
Submitted:
03 March 2024
Posted:
05 March 2024
You are already at the latest version
Abstract
Abstract: More than 40 hypotheses predict associations between features of nest architecture and at least one measure of reproductive success. However, quantitative studies of reproductive consequences of nest characteristics remain scarce. In addition, most studies were conducted on model species of which nests can be easily monitored with artificial nest-boxes. Here we review the replicability of research protocols and findings in model species with many repeat studies focusing mainly on nest-size components, animal-derived nest material, or fresh greenery in blue tits (Cyanistes caeruleus), great tits (Parus major), tree swallows (Tachycineta bicolor), and starlings (Sturnus spp.). The studies looked for correlations between nest traits and aspects of breeding performance that can be easily quantified in the field, such clutch size, brood size at hatching or fledging, the % of eggs that hatch or fledge, or nestling characteristics assumed to reflect qualities associated with survival probabilities (e.g., morphometry, body condition, blood profiles). We discuss the consequences of poor replicability of research methodologies and provide explanations for why many of these studies reported poor associations between nest design and breeding success at different spatiotemporal scales. We also make suggestions for future research.
Keywords: Nest design, Breeding success, Nest-box, Cavity-nesting birds, Replicability
Keywords:
Nest design
; Breeding success
; Nest-box
; Cavity-nesting birds
; Replicability
1. Introduction
Eggs and offspring of many taxa regularly develop in constructions, like nests, containers, burrows, or holes [1,2,3,4,5,6,7,8,9,10]. Animal constructions are amongst the first examples mentioned in discussions about extended phenotypes or ‘niche construction’, which also implies that nest design should have important fitness-associated implications [3,5,11,12].
A bird nest typically consists of a non-random combination of various vegetation-derived, animal-derived, and/or mineral-based materials [5]. Whereas the harder nest components (e.g., sticks, roots, mud) are used to strengthen the nest base, soft vegetation fragments (e.g., moss, grass, and leaves) form the basis of the nest walls, and animal-derived materials (e.g., feathers, hair, fur) are most often placed in the vicinity of vulnerable eggs and chicks [13,14]. The primary role of an avian nest is to provide a place where eggs are laid and incubated. In many species it is also the place where the chicks are reared to fledging [5]. An essential design element of a nest is to protect its occupants against environmental dynamics [reviewed in 5]. Nests have insulation properties that reduce cooling or accelerate warming so that energy-based expenditure during incubation or brooding is minimized [15]. In addition, nests have physical properties that help control the dynamics of nest humidity [16] or minimize exposure to heavy rain or wind [17]. However, large nests with abundant animal-derived material may hamper cooling and increase risks associated with hyperthermia when environmental temperatures are elevated [18].
Nests also provide signals exploited in conspecific or heterospecific interactions [19,20]. Although many nest structures may have been designed to improve crypsis, abundant nest components might increase nest predation risks when they increase nest detectability [21,22]. Bird nests cannot only be breeding environments for the nest builders and their offspring, but also for other non-avian species, often invertebrates that use bird nests for reproduction [23,24]. Therefore, some nest components act as a physical barrier to protect eggs or nestlings against nest parasites or disease. Finally, nest architectures will be determined by constraints, which include nest builder characteristics (e.g., body size and condition, biased perception characteristics), nest-site design (e.g., cavities imposing nest shapes) or availability of resources used for nest construction. Therefore, components of nest design should be the outcome of a combination of benefits, costs, and constraints, which have been discussed in more detail in books or review papers (e.g., [5,20,25,26]).
Bird nests have become a more popular research topic in recent years when research became interested in reproductive consequences of human-induced environmental change. Broad nest types have been described for more than 5,000 of the more than 10,000 identified bird species, but the size of nest structures is known for less than 400 species, and the descriptions and measurements of the masses of nest components in dismantled nests have been done for less than 50 species [5]. The plant or animal species used as materials to construct or decorate bird nests have been rarely mentioned in scientific publications (e.g., [5,13,27,28,29,30,31,32]). Empirical studies of associations between avian nest design and breeding success remain scarce [33], even though costs and benefits of nest design are always expected to influence at least one aspect of reproductive success. This is because a quantitative study of reproductive consequences of nest design is a time-consuming activity requiring adequate sample sizes of differing nest architectures and their associated breeding performances.
Bird nests have two crucial and different roles in successful breeding. The first is that the nest is where the eggs are laid and incubated. Therefore, variation in nest architecture is likely to have some form of impact upon the incubating bird and the likelihood that eggs will hatch. All bird nests fulfil this function and reproductive success is measured by percentage hatchability. The second role involves the nest as a location for rearing of offspring. In many species, hatchlings leave the nest soon after hatching and the nest plays no role in their subsequent development. However, in many, especially altricial species, the nest has a key part of play in ensuring survival and successful fledging of the offspring. However, the functional aspects of the nest need to deal with the dynamic pattern of ontogeny as the small ectothermic hatchlings grow and mature into large endothermic fledglings. A well-insulated nest may be crucial for keeping small, naked hatchlings warm, but this may adversely affect thermoregulation in older, mature birds. Reproductive success in this case is typically the percentage of the eggs in a clutch that become hatchlings or fledglings, often alongside fledgling size, and condition. Nest architecture may influence the first half of post-hatching development more than the second half.
Field studies that looked for associations between aspects of nest design and reproductive success have been conducted most in nest-box breeding passerine birds that also belong to the best-studied non-domesticated vertebrates during the last century (e.g., [33,34,35,36,37]). Secondary cavity-exploiting birds build nests within cavities they do not excavate themselves and breeding opportunities are limited by the availability of tree holes in secondary forest or human-transformed habitat. In the absence of natural holes, these species of birds readily accept and nest in human-made boxes attached to different substrates, like buildings, artificial posts, or trees. Human-made boxes and their surroundings can have properties that substantially differ from that of natural cavities or tree holes (e.g., [38,39,40,41]). However, artificial boxes improve accessibility for humans to routinely monitor bird populations and reproductive performance. Use of nest-boxes also facilitate experiments in human-controlled environments to reveal more easily the underlying proximate mechanisms that link the individual responses to environmental change. Costs and benefits of nest-box use have been reviewed recently by Thompson et al. [42], Zhang et al. [43] and Marcus et al. [44].
An important scientific challenge of all wildlife research on complex ecological interactions is to improve ‘replicability’ of repeat studies that investigate the same problem with different data samples and in different settings. When replicability is high, repeat studies are expected to obtain similar results. However, lack of replicability has been highlighted in different research fields, including research on behaviour and ecology [45,46]. Nature’s dynamics and constraints impose on science investigations, and so each empirical study will most probably be unique in its cocktail of research process components so that true or exact replication (e.g., [45]) cannot be realized in practice. For instance, poor replicability can be caused by spatiotemporal bias in at least one of the research process components, such as protocols, observers, study design, logistics (e.g., tools), sampling efforts, hypothesis-driven data analysis, or study system characteristics. However, repeat studies that apply the same research protocols do not necessarily have to produce the same results when individual-specific, population-specific, or species-specific responses to an external environment express spatiotemporal variation (e.g., [47,48,49]). Therefore, both the biology-specific characteristics of model systems, and the laboratory-specific research protocols applied to understand these model systems, might influence the level of replicability of research findings in repeat studies. Identifying their relative importance requires research collaboration standardizing or randomizing characteristics of research protocols at different spatiotemporal scales (e.g., [4,50]), ideally defined before the initiation of collaborative studies (e.g., [37,51]). However, most field studies have been initiated independently from international collaboration networks, which was also the case for investigations of reproductive consequences of nest characteristics.
Here we discuss the replicability of methodologies and research findings in intraspecific repeat studies that investigated breeding consequences of avian nest design. We focus our review on the avian model species that provided most of the repeat studies, which are non-domesticated secondary-cavity nesters that will regularly nest in artificial nest-boxes. We compare methods and research findings mainly in great tits (Parus major, 13 studies), blue tits (Cyanistes caeruleus, 22 studies), tree sparrows (Tachycineta bicolor, 7 studies), and starlings (Sturnus spp., 9 studies). The studies examined nest-size components, animal-derived nest material, and fresh aromatic plant fragments placed on the nest rim. It also involved monitoring of breeding traits expressed after the nest building phase, such as the size of the incubated clutch, the size of the brood at hatching or fledging, success of hatching or fledging, and/or nestling phenotypes that reflect qualities linked to physical condition and future survival probabilities (e.g., morphometry, blood profiles). Many of these studies have been guided by hypotheses that serve to explain why nest characteristics might be associated with at least one aspect of breeding success. These hypotheses, the environmental conditions required to test them, and the publications in which at least one hypothesis has been proposed or tested, are summarized in Table 1, many of which are discussed in the different sections below.
2. Methods
Published information on the architecture and breeding success of bird nests has been extracted from articles and their references or citations that were exposed on the electronic platforms HAL, ResearchGate, Google Scholar, JSTOR, journal websites and species-devoted websites (e.g., treeswallows.com). The verified publications dealing with breeding consequences of nest architecture regularly cited the same publications indicating that the bibliographic searches were exhaustive. Authors were contacted to obtain additional information about recent findings.
3. Replicability of Methods and Research Findings in Studies of Nest-Box Breeders
3.1. Replicability of Methodologies
3.1.1. Nest-Size Components
Nest-size components include all the nest characteristics associated with length, volume, thickness, or mass. This can involve measurements of the whole nest (e.g., nest depth, nest volume, nest mass) or measurements of nest components (e.g., moss mass, stick mass, nest cup thickness), which are also used to calculate proportions of the whole nest (e.g., mass proportion, stick proportion) (e.g., [52,53]). The simplest field measure is a measurement of the external vertical distance between the nest bottom and the top of the nest rim [1,54], and variously named nest thickness [55], nest depth [1,56] or nest height [54,57,58,59]. In long-term studies, most measurements were taken at the external side of the nest that was closest to the entrance in boxes of which the front door could be removed (e.g., [57,60,61]). However, in some years or in some study sites, measurements were also taken at the external side of the nest that was farthest from the entrance, especially in study sites where nests expressed important asymmetry in the height of the nest rim [62] or in study sites with nest-boxes of which the front panel could not be removed (e.g., [63]). In symmetrical nests and in studies that used more than one box type, nest height was used to calculate the nest volume by multiplying the nest height with the internal surface of the nest-box floor [64].
Some studies measured the mass of fresh nests during the periods of egg laying or incubation whereas other studies measured the dry mass of deserted nests collected after the breeding season (e.g., [65,66,67,68,69]). Nest-size components of non-deconstructed nests, such as the thickness of the nest material under the cup or the wall surrounding the cup, or the nest cup’s depth, diameter, or volume, have occasionally been quantified (e.g., [52,53,54,66,70,71]). However, debris produced by parents or nestlings (e.g., feather fragments) can accumulate inside the nest significantly increasing the mass of successful nests (e.g., [66,68,69]). In addition, the physical and associated insulation properties of a nest might change throughout the breeding event when the nest becomes compressed because of the fast-growing brood [72]. Nest compression is probably more important in populations where nest bases are mainly built from moss and brood mass is higher, for example in large clutches of great tits or blue tits. Most recent studies of the insulative properties of nest walls have focused on species that nest in open environments [15]. Laboratory studies with collected or artificial nests also show that dried hair, feathers, and moss have higher insulative properties than dried leaves, grass, or roots and that the position of the nest materials relative to the position of the eggs might also be important (e.g., reviewed in [5,15]). Studies of thermal properties of nests for species breeding in nest-boxes in combination with measures of breeding success are rare and involve small sample sizes (e.g., [67,72]). There are also few studies of breeding success that investigated nest-size components as niches for non-flying nest parasites (e.g., [23,24]) or as odour sources for flying nest parasites (e.g., [73,74]). Obviously, studies interested in reproductive consequences of the composition of entire nests cannot dismantle nests before the breeding cycle is finished and breeding performance quantified.
Most studies compared nests from one box type. However, different research teams did not use the same nest-box size, even though positive associations between nest-size components and nest-box size have been repeatedly reported (e.g., [4,64,70,75,76,77,78,79,80,81]). Therefore, some researchers monitored larger nests whereas other researchers monitored smaller nests without providing scientific arguments for nest-box type choice. Most studies did not use the same nest-box design, so the relative importance of nest-box characteristics and nest characteristics in the expression of breeding success could not be established with rigorous research approaches. However, nest-box size was used as a proxy of nest size to allow multi-factor analyses at a macro-geographic scale [4,50]. Such a comparative approach is useful when nest size varies less within box types than across box types, which has been validated by case studies (see above).
The complexity of the statistical approaches is associated with the number of nest traits, study years, explanatory variables and/or characteristics of study design (e.g., [4,50,52,54,61,65,66,76,82,83]). For instance, Álvarez and Barba [54] measured nine nest-size components that were reduced to three principal components that reflected the size of the nest, the nest cup, and the basal area of the nest. The composition of research teams and their statistical tools have also evolved over time, perhaps influencing the outcome of analyses and associated interpretations. Reanalyzing older data with new statistical approaches might therefore be useful. For instance, multi-factor analyses of associations between breeding performance and nest size remain rare often ignoring key drivers of breeding performance other than nest traits (e.g., identity and characteristics of nest-builders, environmental factors) (e.g., [57,60,61,83]). Some investigations examined associations between nest size as the explanatory variable and clutch size as the dependent variable [4,52,53,57,62,65,78,81], whereas other studies considered nest size and clutch size as proxies of nest-builder quality potentially influencing breeding success (e.g., [60,61].
Big nests might provide thermal benefits for incubation when ambient temperatures are low, whereas small nests might be beneficial when ambient temperatures are high (e.g., [18,62,82,84,85]). However, multi-factor analyses of associations between breeding success and nest size rarely consider the dynamics in meteorological conditions during the ectothermic phase of breeding, i.e., when altricial chicks lack insulating plumage and rely on their parents for thermoregulation [63]. In multi-site analyses, the first-egg date or geographic location could be used as thermal proxies when the average meteorological conditions change with the progress of the breeding season and latitude or altitude (e.g., [4,50,63,84,86]).
Nest-builder age has been included as a potential driver of breeding success in studies of nests with older endothermic nestlings with full plumage (e.g., [57,60,61]). However, the functional properties of nests are expected to be expressed mainly during the earlier phases of breeding (e.g., [5,84,85]), that is before the parents are captured and identified. Therefore, studies of nest desertion during the ectothermic phases of breeding ignored the age or identity of the nest builders as explanatory variables. In addition, studies that looked at characteristics of nests that did not produce fledglings did not make a distinction between the ectothermic and endothermic phase of the nestling stage (e.g., [57,60,61,63]).
Research protocols (e.g., nest-chamber design) cannot be truly replicated when experimental design is hypothesis-dependent. For instance, Mazgajski and Rykowska [76] and Kaliński et al. [87] changed the design of large nest-boxes to alter the distance between the nest-box entrance hole and the nest rim to examine associations between nest-box design and nest size on the one hand and nest predation risks on the other hand. By contrast, Järvinen et al. [82] examined the thermal benefits of nest size by adding or removing nest material without changing nest-box design and without quantifying the composition of entire nests. However, Järvinen et al. [82] worked with small boxes and an experimental increase of the height of the nest cup might have increased a risk of nest surface cooling because of a higher exposure to wind blowing into the nest cavity via the entrance hole. Thus, studies manipulating nest size independent from nest-chamber size should also consider influences of nest-chamber characteristics (e.g., chamber size, chamber depth, entrance orientation, box age) potentially influencing exposure to wind inside the nest chamber (e.g., [62]). Small passerine birds cannot remove eggs from nest-boxes (e.g., [88]), and so making a distinction between effects of nest predation and insulation also requires a study of ectothermic chick fitness components, including quantification of the disappearance versus desertion of eggs.
Although nest-size components can be easily manipulated from a logistic point of view, only two blue tit studies have done this in natural conditions, and research protocols were only partially replicated [82,89,90]. Moreover, there are few field experiments that used artificial cavity nests [91,92] or exchanged cavity nests between breeding pairs or species (e.g., [93,94,95]). Switching nests between study populations or species can examine reproductive consequences of significant changes in nest-size components in the absence of the time-consuming research process of nest deconstruction given that species consistently differ in nest architecture. The thermal properties of different types of nest materials have been investigated in laboratory conditions, but the fitness consequences of experimental changes in nest-size components remain unknown in natural conditions. For instance, a higher mass of wood-based material that has lower insulation properties might reduce breeding success in study sites where a nesting material preferred at other sites is lacking (e.g., [53,64]). However, there are no European field experiments that examined reproductive consequences of significant changes in the proportion of wood-based material inside cavity nests, probably because most field studies were conducted in non-urbanized areas where wood-based material is rarely used or absent (e.g., [52]).
3.1.2. Animal-Derived Nest Material
Animal-derived nest material [ADNM] usually include heterospecific feathers, mammalian hair and fur, and arthropod silk [1] although human-made materials are also used for nesting (e.g., [96,109]). The types of materials used to line nests differ between study populations or species (e.g., [13,14,49,66,97,98]), which explains why studies conducted on different species do not focus on the same nest materials and therefore cannot be fully replicated. There are few published measurements of the average mass of animal-derived nest material in secondary nest-cavity species (e.g., [53,66,97]) because the ability to measure its mass will also depend on the ability to separate it from other nest material. It is not too complicated to separate heterospecific feathers from other nest material, which can explain why many studies of the reproductive consequences of lining material focused on feathers (e.g., [83,99,100,101,102,103]). However, published reports of the mass of mammal-derived nest material in association with measures of breeding success remain an exception (e.g., [51,53,66,68,104,105]), even though more than 70% of the European passerine bird species regularly use hair or fur for nesting [30,106]. Mammal-derived material is often tightly mixed up with other nest material (e.g., moss), especially in nests that produce fledglings, so that it takes much time to extract it. Animal-derived material has been quantified more rapidly with estimates not requiring nest deconstruction (e.g., scores, photographs) assumed to reliably reflect relative differences in nest composition (e.g., [70,83]). Some studies only investigated the visible components of animal-derived nest material to test specific hypotheses associated with signaling, crypsis, or thermoregulation (e.g., [15,51,83,101]). A few experiments have used artificial eggs and dummy nests covered with hair to examine associations between the level of egg covering on the one hand and the probability of nest destruction [107] or nest usurpation by other species [108] on the other hand. The mass of lining material is sometimes difficult to obtain when nestlings contaminate nests.
Birds frequently incorporate human-derived nest material (anthropogenic) into the nest, perhaps because anthropogenic and non-anthropogenic nest materials have similar physical properties. According to Reynolds et al. [109], human-derived nest material always undergoes human-based processing and include plastics or dyed tissues used to produce cloths. Hanmer et al. [110] referred to dyed, treated cotton and artificial stuffing materials, whereas other studies mentioned use of strings, papers, textiles, and pieces of plastic [111,112]. McCabe [27] also referring to H. C. Battell reports that house wren (Troglodytes aedon) nests could contain nails, hairpins, safety pins, paper fasteners, pieces of wire, cellophane cigar wrappers, or thin transparent plastic. However, defining anthropogenic nest material with precision is not an easy task given that fragments of domesticated plant or animal species might also be considered as anthropogenic via the human-guided process of domestication. What then exactly is anthropogenic-based nest material when heterospecific species (e.g., fur from wildlife species versus domesticated dogs; twigs from native versus introduced tree species) exploited for avian nest building are rarely identified with precision? Only recently, ornithologists became interested in breeding consequences of artificial anthropogenic materials exploited by birds to line their nests (e.g., [110,111,112]). Anthropogenic nest materials have been weighed with high precision balances (e.g., errors of 0.01 g, 0.0005 g, or 0.0001 g), which implies that they often represent only a tiny fraction of the entire nest. Presence of human-made material inside bird nests have been used as bioindicators of human presence or waste pollution (e.g., [112]). Consequently, its presence might covary with other environmental drivers of avian breeding success. Experimental studies involving human-derived nest material remain rare [92,96].
3.1.3. Greenery
Greenery is represented by fresh plant fragments other than moss (which often remains green even in old nests) that are used to decorate or cover the nest surface without having functions to strengthen the nest or improve nest insulation properties. Often such materials are brought to the nest during incubation or nestling rearing rather than being incorporated into the original nest. At least 10 hypotheses have been proposed to explain why birds include fresh greenery into the nest [113,114]. For instance, volatile chemicals emitted from greenery might hide avian-generated odours as a nest-protection strategy, stimulate immune functions in adults or nestlings, or modulate conspecific sex-associated interactions (e.g., [74,114,115,116,117,118,119]). In secondary-cavity nesters, greenery is considered to be a signal aimed to attract mates or repel conspecifics or other species to reduce competition or parasitism, although they may have other functions in open-nesting species (reviewed in [114]).
Use of greenery, and how it is delivered to the nest, is a species-specific trait. For instance, European starling (Sturnus vulgaris) males deliver greenery in the presence of a female and mostly before the onset of egg laying [118,120], whereas blue tits mostly deliver greenery after the onset of egg laying (e.g., [121,122]). Most studies investigated impacts of greenery on aspects of breeding success independent from other nest traits [89]. For instance, the functions and reproductive consequences of fresh greenery delivered by males or females have been frequently investigated in nest-box breeding European starlings and spotless starlings (Sturnus unicolor) rarely taking other aspects of nest composition into account (e.g., [123,124,125,126,127,128,129]).
Fresh greenery incorporated into the nest before the initiation of hatching might rapidly deteriorate [28,122,130] and therefore not be found during the phase of dismantling of dried nests after the breeding cycle is finished. Herb-fragment deterioration could be verified when a known amount of greenery is added to the nest at the time of egg laying and their presence checked in nests collected after the fledging stage. However, parents might remove or add herb-fragments after greenery loads are experimentally changed [122,131]. Therefore, experimentally added fresh herb fragments stored inside small cloth sacs or containers could be incorporated inside the nest without bird parents having access to it. Sampling fresh plant fragments at the time they are incorporated into the nest might have implications for the study of associations between greenery and breeding success. Photographs of nest surfaces could be taken with the drawback that the greenery or other nest material inserted inside the nest might be ignored [132]. Alternatively, delivery of fresh plant fragments could also be recorded with camera devices and their frequency of delivery quantified as is done with nestling food cf. [133].
In an interspecific comparative analysis of starlings, the use of greenery is more common in cavity-nesting than in open-nesting species [134]. This result suggests that the actions of the greenery volatiles might be more efficient inside cavities or the requirement to hide nestling odours with plant volatiles might be less important in open nests. For instance, one might speculate that volatiles emitted by fresh greenery will most probably be influenced by the level of wind exposure. Therefore, volatile concentrations are expected to be higher inside cavity nests not exposed to an air flow than in open nests exposed to wind. However, the frequency of fresh greenery delivery aimed to maintain a minimum level of active volatiles should depend on the temporal dynamics of the volatiles expressed inside the nest chamber. For instance, Petit et al. [28] showed that the quantities of volatiles of fresh aromatic plant fragments placed inside an empty nest box will decrease rapidly within a 48h time-window. Therefore, we cannot exclude that active volatile dynamics will also be influenced by the frequency and duration that nest boxes are opened, especially during windy days. However, to our best knowledge, there are currently no published field studies that measured dynamics in volatile concentrations emitted by fresh greenery inside active nests. Consequently, we do not know the associations between nest-box visitation protocols and their consequences for volatile dynamics and associated avian breeding success. Although nest-boxes could be checked with mirrors without opening them following the research protocols applied in studies of nests built inside tree holes [135,136], we are not aware that this was done to improve the understanding of the reproductive consequences of bird nest chemistry.
It is not easy to quantify greenery loads incorporated into nests without penalizing breeding success (e.g., [137]). Therefore, most studies used an experimental approach adding or removing fresh greenery (e.g., [131,137,138]). Grass was sometimes used as a control treatment to compare the reproductive performances of nests with versus without fresh greenery (e.g., [116,119]). The species of greenery that have been used for the experimental treatments were not replicated across studies [114]. Although a cocktail of different greenery species might be more efficient in fighting parasites or disease than the action of a single greenery species (e.g., [122]), field experiments have never compared the effects of a cocktail of greenery species resembling multi-therapy treatments with the effects of a single greenery species. Medical-oriented experiments might test herbs that are not exploited by the birds, which has also been done in species that do not incorporate greenery in the nest [139,140]. In addition, some populations might be experimentally exposed to novel greenery not locally available when an experimental design involving tests with the same greenery species is replicated at a wide spatial scale, (e.g., [141]).
A simple experimental design could be expected to be replicated most often across studies (e.g., [51,114,141]). However, the design of research protocols cannot always be truly replicated when the protocol must be adjusted to the characteristics of the local model system and different potential drivers of breeding success must also be considered. For instance, in Corsica, blue tits continuously deliver aromatic fragments from different plant species (e.g., Achillea lugistica All., Lavandula stoechas L., Helichrysum italicum Roth., Mentha suaveolens Ehrh.) throughout the breeding cycle [28,122] and blow flies continuously contaminate the nest after hatching (e.g., [131]). Therefore, an experiment that examines the direct effect of aromatic herbs on breeding performance must control for nest-builder behaviour and the dynamics of nest parasitism also influencing breeding success. In addition, associations between the mass of aromatic plant fragments and breeding performance might become stronger under more severe environmental constraints. Therefore, Mennerat et al. [131] combined aromatic plant manipulations with brood-size manipulations in nests where aromatic plants and blowflies had to be removed every second day and nest compositions significantly altered from 5-days up to 15-days post-hatch. Some field experiments might be logistically so complex that application of true replication procedures becomes complicated from a logistic point of view.
2.2. Replicability of Research Findings
Population-specific or species-specific characteristics might lower the replicability of research findings across studies. In great tits and blue tits, the female builds the cavity nest, and takes care of nest sanitation, the production and incubation of the eggs, brooding of the hatchlings and rearing of older nestlings. By contrast, the male partner significantly contributes to territorial defence, courtship feeding, offspring feeding and helps with removal of faecal sacs produced by nestlings (e.g., [88]). In Spanish blue tits, males deliver feathers (e.g., [102,103]) in a sexual selection framework, although this behaviour has not been observed in Norwegian blue tits (T. Slagsvold, pers. comm.). The sex-specific roles in reproduction in these two cavity-nesting species mean that one might expect that (1) the female-built nest characteristics are expected to have the strongest impact during the ectothermic phase that is under better female control, and (2) the reproductive efforts of the male partner that do not directly contribute to nest building can modify the strength of the association between nest design and fledgling productivity. In tree swallows, however, the males cover the female-built nests with feathers, which might strengthen the associations between nest components and aspects of breeding success (e.g., [70,99,100,142]).
3.2.1. Nest-Size Components
Many nest-box studies predicted higher breeding success in larger than in smaller boxes when increased nest-building efforts associated with the occupation of larger nest chambers would better reflect aspects of the quality of the nest-builders, their mates, or their territories [4,44,63,82,89,143,144,145,146,147]. Previous studies in contrasting habitats throughout Europe also showed that great tits or other cavity-nesting passerines occupy more often the larger chamber when there is an option to choose between at least two chamber sizes. In addition, the preference for larger chambers in choice experiments with nest-box breeding great tits is maintained in contrasting habitats (e.g., Scandinavian versus Mediterranean; urban versus rural) (e.g., [4,50,143,148,149,150,151,152,153,154,155,156,157]). Maziarz et al. [158] also noticed that there is considerable overlap in the tree cavity characteristics exploited by great tits from different study areas. Therefore, the naturally selected forest birds might have evolved genetic-based preferences for larger chambers where chamber size would signal aspects of future survival probabilities for breeders and their offspring (e.g., [37,157]). In addition, nest-box breeders, like great tits, produce larger nests with more eggs inside larger cavities (e.g., [4,148,149,150,151,152,157], and references therein). This preference suggests that the birds prefer larger cavity nests for reproduction, perhaps because larger and thicker nests have a higher insulative value increasing incubation efficiency [84,85,86,97,150,159] or reduce risks associated with hyperthermia, sibling competition for space, nestling crowding complicating nest cleaning, nest-cup humidity, nest-cup rigidity, flooding, nest predation, nestling-induced gas accumulation (e.g., NH3), or nest parasitism [4,91,143,148,149,150,152,155,158,160,161,162]. Alternatively, in contrast to eggs or nestlings, parents might escape more easily from nest predators when nest chambers are substantially larger. However, in the short-term field studies, associations between breeding success and nest-size components differed between study years, study sites and/or model species (e.g., [53,54,66,81]). A few studies reported that bigger or heavier nests were associated with higher hatching success or more hatchlings (e.g., [52,53,61,163]), but others have not (e.g., [78], although the strength of the associations differed between study sites and species [53,163] and how the data were analyzed (e.g., [70]). Studies that lasted at least four years never reported strong positive associations between the height of fresh nests and brood size at fledging or fledging success (e.g., [57,60,61,83]).
Lombardo [70] found an impressive number of associations between nest-size components (nest-cup volume, nest volume, thickness of the nest under the cup, nest depth, cup index) and breeding success in tree swallows. To obtain these results, he exploited sub-samples to calculate simple correlations dealing with a single year, a single female age-class, or a biased breeding time-window (either early or late breeding). Results were not easy to interpret because the strength and direction of the associations depended on how the data samples were selected.
In one study plot in mainland southern France, two repeat studies investigated associations between the height or volume of fresh nests and different aspects of breeding success in blue tits [57,60]. The multi-factor studies did not consider the same drivers of breeding success and did not use the same statistical methods, but the two studies revealed that breeding success was weakly associated with nest size after controlling for other drivers of breeding success. These studies also considered nest-builder age as a potential driver of reproductive performance, so data only involved nests from captured parents when endothermic nestlings were older than 8 days post-hatching. The analyses repeatedly revealed that clutch size is a significant driver of hatchling and fledgling productivity, a result confirmed with multi-site investigations in Corsican blue tits [61].
Several studies reported positive associations between brood size at fledging and the mass of non-deconstructed nests collected after fledging [65,69], whereas such an association was not found for fresh nests measured during egg laying or incubation [69]. This can be explained by the fact that larger broods produce more nestling skin dust that makes nests with more fledglings significantly heavier than nests with less fledglings [66,68]. In other words, higher breeding success caused heavier non-deconstructed nests at the end of the breeding season, whereas heavier fresh nests did not cause higher breeding success.
Several studies simultaneously investigated fresh nests or dismantled nests in great tits and blue tits that occupied the same study sites during the same breeding seasons [66,110,163]. These studies showed that the associations between breeding success and nest size were species-specific, probably because the two species differ in body size or do not exploit the same resources for nesting and breeding (e.g., [49,58,164]).
Field studies that experimentally changed nest size by adding or removing nest material without changing nest-box design did not find significant associations between nest-size treatment on the one hand and measures of breeding success on the other hand [82,89,90]. In house wrens (Troglodytes aedon), stick removal did not change breeding success compared to controls where sticks were not removed [90].
Perhaps the best evidence that breeding performance is weakly associated with nest size is that secondary-cavity nesting passerines successfully breed in nest-boxes that vary in nest-chamber size assumed to be closely correlated with nest size (e.g., [4,37,50]). In three field studies of Paridae, nesting in larger boxes resulted in larger nests without significantly increasing the average hatchling or fledgling productivity per nest compared to nesting in smaller boxes (e.g., [64,75,80,157]). In urban great tits, Demeyrier et al. [157] reported a lower average breeding success per nest in larger than in smaller nest-boxes, attributed to the effects of an ecological trap mechanism. On the other hand, in Algerian blue tits, the average ratio of the number of fledglings to clutch size per nest was higher in larger than in smaller boxes [80]. In the UK, three different nest-box sizes showed no relationship with any measure of reproductive success in blue tits [78]. The absence of a significant association between nest-box size and reproductive performance was also reported in other cavity-nesting species (e.g., [81,104,152]). However, Van Balen [152] argued that nestling survival might be lowered in very small nest-boxes when a small bottom area would prevent nestlings from escaping from physical contact therefore increasing hyperthermia risks. This would imply that the strength of the association between nest size and breeding success might also depend on which nest-chamber sizes are considered in studies involving more than one box type.
Overall, the simplest conclusion is that in the best-studied non-domesticated nest-box breeding model species the number of fledglings per nest or the ratio of the number of fledglings to clutch size per nest are poorly associated with nest-size components. There are several potential mechanisms that can explain why nest design and breeding success are poorly associated, as illustrated with an example in Box 1. For instance, one scenario is that nest-size components, that also influence the nest-insulation properties and associated thermal conditions of breeding, are not adjusted to the dynamics of the ambient temperature. Consequently, in many instances, nest design might become suboptimal as expressed in increased hypothermia or hyperthermia potentially reducing breeding success. Also note that, for birds, nest-box position and orientation might become important when nest chambers exposed to direct sunlight can increase the surface temperature of non-incubated eggs up to 5°C above the ambient temperature [165]. A significant rise in egg temperature because of direct sun exposure might initiate maladaptive egg development before the onset of incubation, perhaps forcing females to initiate incubation before the clutch is completed which would increase the occurrence of asynchronous hatching [165,166].
Box 1. A hypothetical model for associations between nest insulation, environmental temperature, and breeding success.
In some birds, nest insulation has an inverse relationship with environmental temperature during the period of nest construction (Figure 1) [84,86,211]. Moreover, Crossman et al. [212] showed that in Canada prevailing climate in two different locations affected size and composition of songbird nests. These results suggest that birds seem to be building nests that reflect the prevailing environmental conditions and which may provide an optimum nesting environment. For example, if the prevailing temperature is cold during construction the bird needs to build a well-insulated nest to minimise its own heat loss (and that of its eggs), and so minimise energy expenditure during incubation.
Let us postulate that nest insulation is constant between years but environmental temperature varies from year to year (Figure 2). For most years there will be a mismatch between environmental temperature at the time of nest construction and nest insulation. A constant nest insulation will only be optimum for the prevailing environmental temperature in a few years (e.g., see arrows in Figure 2). Under such conditions it is predicted that reproductive performance would be suboptimal during most years and there would be a curvilinear relationship with environmental temperature (Figure 2). This is because inadequate nest insulation would lead in cool years to hypothermia of the incubating bird, eggs, or chicks. Moreover, too much nest insulation would lead to hyperthermia in warmer years. Both conditions would reduce reproductive performance (Figure 2).
By contrast, if nest insulation varies from year to year because the birds probably respond to the environment by inversely matching nest insulation to prevailing environmental temperature conditions (Figure 3). As a result, nest insulation will mirror environmental temperature from year to year (Figure 3). Nest insulation would produce environmental conditions in the nest that would be close to optimum for most years and as a result there would be no relationship between reproductive performance and environmental temperature (Figure 3).
In support of this idea, Deeming & Merrils-Brown [213] reported that for song thrush (Turdus philoemlos) nests built in Devon, UK built within 2 km of each other, but over a period of six years thermal conductance of the nest wall varied among years. Morover, neither year nor thermal conductance had no any significant effect on either date of clutch initiation, or any measure of breeding success. Therefore, observational or experimental studies may not reveal effects of variation in nest insulation on reproductive performance of birds because of the behavioural plasticity of the birds leads to nest construction that matches the prevailing environmental conditions, and optimises the incubation environment.
Alternatively, the nest-size components and their associated insulation properties might be adjusted adaptively to the spatiotemporal dynamics of the thermal conditions of breeding so that different nest designs can have the same breeding success [Box 1]. This adjustment in nest design might be done before or after the onset of egg laying. At European latitudes, warmer breeding conditions will probably require smaller nests combined with smaller broods to prevent hyperthermia (e.g., [18]). This might also explain why the size of the nest components and associated brood size are larger in first than in replacement or second broods without altering the ratio of the number of fledglings to clutch size per nest (e.g., [57], Lambrechts et al., unpubl. Data). Consequently, nest builders might indeed be able to anticipate the future thermal conditions of breeding, like egg layers do when they adjust their clutch size to clutch type (first clutch versus replacement or second clutch) or local habitat characteristics (e.g., food availability, latitude, ambient temperature) without substantially changing the proportion of eggs that produce fledglings [160,167]. In a similar vein, in painted turtles (Chrysemys picta) that nest at different locations in North America, nest temperatures are optimized for embryonic development by females selecting nest sites that offer the best environmental conditions [10]. In other words, the turtles can anticipate future breeding conditions via the selection of an appropriate nesting site.
However, we currently do not know whether variation in the associations between breeding performance and the different measures of nest size result from a combination of spatiotemporal bias in research protocols and/or study system characteristics. Therefore, it is recommended to standardize or randomize research protocols at different spatiotemporal scales or to conduct local field experiments altering nest-size components to explore causal effects of nest morphology on breeding success (e.g., [65,82,87]). For instance, the macro-geographic analysis of Møller et al. [4,50] assumed that there exist a tight association between nest-box size and nest size, which has been confirmed by different local studies [64,76,78,79]. Other long-term field studies involving measures of the size of fresh nests that produced endothermic nestlings revealed strong positive associations between clutch size on the one hand and the brood size at hatching or fledging on the other hand (e.g., [57,60,61]). Therefore, the macro-geographic patterns involving clutch size and nest-box size as reported in Møller et al. [4,50] might also be found in similar analyses that focus on brood size at hatching and fledging.
3.2.2. Animal-Derived Nest Material
Long-term studies of associations between animal-derived nest material and proxies of breeding success are rare, and findings of short-term studies varied across local populations or model species, probably also because of spatiotemporal variation in study site characteristics or year effects. The correlative or experimental field studies that examined associations between feather use and breeding success were mainly conducted on tree swallows and blue tits that cover female-built nests with heterospecific feathers (e.g., [70,83,99,100,102,168,169]), whereas the associations between mammal-derived nest material and breeding success were mainly conducted in great tits that rarely adorn nests with feathers [51,53,66,98].
Studies in tree swallows examined the benefits of feather use as insulation material minimizing heat loss improving the energy budgets of the nestlings or as a physical barrier minimizing exposure to parasites hidden inside the nest. Although the associations between a measure of feather presence on the one hand and hatchling or fledgling productivity on the other hand were weak in several studies [70,99,100,168,169], there were positive correlations between nestling development patterns and different measures of feather abundance [99,100,168,169]. For instance, Stephenson et al. [168] showed that nestlings were larger in control nests than in nests where feathers were experimentally removed. In addition, nestling growth was positively associated with the number of feathers in the nest. However, the number of fledglings were not associated with feather abundance or feather treatment, perhaps because heavy rainfall at the time of fledging modified the strength of the association between feather counts and fledgling productivity. In addition, feather treatments combined with insecticide treatments did not have a significant impact on nest parasites attributed to the fact that the number of nest parasites counted in this study population were relatively low compared to that of other study populations in North America. There were no counts of the number of parasites attached to nestlings so that the physical barrier effect of the feathers was not examined. In some tree swallow studies, males competed for feathers and delivered a significantly larger number of feathers to the nest following experimental feather removal (e.g., [168]), which might have penalized hatchling or fledgling productivity.
In Spanish blue tits, feather carrying in males can impact on sexual interactions altering extrapair copulation patterns, intraspecific competition, or parental investment in males [102,103]. For instance, correlative and experimental approaches revealed that feather carrying in males induced females to lay more eggs, improved nestling quality and fledgling productivity [102]. However, experimentally adding feathers might also have negative effects when it would become signals of male-male competition prior to incubation [103,126,170]. In a two-year blue tit study conducted by Britt and Deeming [66], there were no significant correlations between the mass of the lining material (feathers, fur, hair) on the one hand and the proportion of eggs hatched or the proportion of hatchlings fledging on the other hand. In an 8-year blue tit study in Finish blue tits, Järvinen and Brommer [83] reported for the first time a significant positive association between feather use and recruitment probabilities of locally born offspring. However, the proportion of feathers in the nest lining and the presence/absence of feather nest ornaments were not correlated with fledging probabilities. Thus, the reproductive consequences of nest feathering in blue tits remain poorly understood also because of lack of experimental repeat studies. In addition, the outcome of such a study might depend on which sex is feathering the nest, which seems to be a population-specific trait.
In spotless starlings, females adorn nests with feathers in response to the carrying of green plants by males, probably in the framework of sexual selection [101]. The experimental addition of feathers to nests increased clutch size and reduced nestling mortality in polygynous males, not in monogamous males, whereas the morphometric measurements of nestlings did not differ between experimental and control nests [101].
Britt and Deeming [66] could not explain why there was a significant positive correlation between the mass of the animal-derived nest material and the proportion of eggs hatching in one of two study years in great tits. Although Álvarez et al. [53] measured the mass of mammal-derived materials and several measures of breeding success (clutch size, hatching success, fledging success) and nestling phenotypes (size, mass, condition), they did not report that hair mass was statistically significantly correlated with breeding success or nestling phenotypes. Surprisingly, however, Álvarez et al. [53] found in different habitats that the condition of great tit nestlings declined when there were more feathers in the nest, perhaps because the amount or proportion of feathers covaried with other nest components that might have lowered breeding success (e.g., sticks) or the great tit nests were visited by competing blue tits. In the macro-geographic study of Loukola et al. [51], the reproductive success of great tits was not associated with hair mass or the level of clutch coverage that reflected the amount of animal-derived nest material incorporated into the nest, perhaps because the study was conducted during a very late year. In addition, Loukola et al. [51] assumed that the replacement of the natural lining material by the same amount of sheep hair during the egg-laying stage did not significantly alter the breeding conditions after nest-material exchange, which obviously should imply that different types of animal-derived nest materials have similar fitness consequences. Finally, Glądalski et al. [92] replaced great tits nests by artificial nests that also included cotton as lining material and showed an impact on nestling blood profiles compared to controls. However, this study did not report the consequences of artificial lining material for hatching or fledgling success.
The overall picture is that animal-derived lining material seems to have the strongest impact on nestling phenotypes via improved nestling growth or condition. However, its association with reproductive success seems to be less straightforward, also because of the impact of unexpected external interventions during the nestling stage, which can be nest predation, extreme weather conditions, or other year effects potentially influencing whole study populations. Improved nestling development might have long-term consequences expressed in measures of recruitment of locally born offspring, but field studies that linked aspects of animal-based nest materials to recruitment probabilities are very rare [83].
3.2.3. Greenery
Studies of associations between fresh greenery loads and different measures of breeding performance have been replicated most in nest-box breeding blue tits and starlings. The conclusions from the older studies [reviewed in 114, 171] did not differ much from the conclusions of the more recent studies [119,172]. For instance, in blue tits from the Mediterranean region, it has been repeatedly shown that higher greenery loads or experimental treatments with fresh greenery were not translated into more hatchlings or more fledglings [119,130,137,138,172]. Some blue tit studies reported that an experimental change in the amounts of aromatic plant fragments significantly changed the morphometry, growth patterns, or blood profiles of nestlings compared to control treatments (e.g., [131,141]), whereas other field experiments found little or no effects on nestlings (e.g., [119,137,172]). For instance, Glądalski et al. [141] reported an effect of greenery on nestling hemoglobin levels in Polish blue tit broods treated with a cocktail of lavender Lavandula angustifolia (ca. 0.33 g), common yarrow Achillea millefolium (ca. 0.33 g), and Immortelle Helichrysum italicum (ca. 0.33 g) compared to controls treated with fresh grass Poaceae spp. (ca. 1 g). By contrast, Garrido-Bautista et al. [119] did not find such an effect in Mediterranean blue tit broods treated with 0.5 g of Mentha sp. compared to controls treated with 0.5 g of grass. Garrido-Bautista et al. [119] also exposed broods to cross-fostering without altering the original brood size. Mennerat et al. [131] and Pires et al. [172] reported effects of aromatic fragment treatments on some nestling characteristics, but only in larger or enlarged broods, which indicates that the greenery-associated effects on nestlings might be modified by environmental constraints.
In European starlings, Fauth et al. [173] did not find a difference in the breeding performance (pre-fledging survival, post-fledging survival, and morphometry of nestlings) between nests with and without green nest material, which was confirmed by the study of Brouwer and Komdeur [124]. Gwinner et al. [116] also found that European starling nests treated with a cocktail of local herbs was not translated into a higher fledgling success compared to nests experimentally treated with grass. However, compared to controls, nestling body mass and blood profiles (basophils, lymphocytes, haematocrit) changed after the herb treatment without changing the nest parasite loads. In addition, the return rate of the birds to the colony the year after hatching was higher for those from herb than grass nests implying long-term reproductive effects of greenery via an improved fledgling condition. Gwinner et al. [125] and Gwinner et al. [129] also showed that European starlings adjust their yolk testosterone levels and incubation behaviour to nest type assumed to have positive effects on nestling phenotypes. By contrast, experimentally increasing breeding densities in spotless starlings resulted in higher loads of greenery and more feather use, but lowered breeding success probably because of increased intraspecific competition [127].
Thus, in the best-studied model species, fresh greenery seems to have a stronger effect on nestling phenotypes assumed to be associated with post-fledging survival, less on fledgling productivity per se. However, research protocols involving greenery treatments and their reproductive consequences were poorly replicated, so we currently do not know the relative importance of research protocols and study system characteristics (population, environment) influencing nestling phenotypes in experiments involving greenery manipulations.
4. Concluding Remarks and Perspectives
Despite the impressive number of published hypotheses that explained for decades why nest design should be correlated with reproductive success at an ecological scale (Table 1), there is no robust evidence for direct effects of nest characteristics on breeding performance in natural conditions. In addition, studies that reported a significant association between nest characteristics and breeding success proposed more than one explanation for its existence. The relative importance of nest architectures and other potential drivers of breeding success will most probably differ between study years, environments, individuals, populations, or species. For instance, it remains to be established whether qualities of the parents and/or their territories are the common cause for the reported associations between nest characteristics and reproductive success, or whether it is the nest architecture per se that directly influences breeding performance (e.g., [174]). In addition, the combination of environmental pressures that select nest shapes at an evolutionary scale might express important spatiotemporal variation at a local short-term ecological scale. For instance, nest predation pressures that select for smaller cavity nests, or cold spells that select for cavity nests with more lining material, are not expressed every year in a local study plot. In addition, strengths of associations between nest design and breeding performance can be modified by external factors that are not under complete control of the nest builders, which include interspecific interventions or impacts of extreme weather sometimes impacting whole study populations. Thus, one of the reasons why an association between nest design and breeding success might be weak is that a hypothesis has not been investigated in the right environmental conditions or not with an adequate model species. It is evident that it makes little sense to examine the functional benefit of fresh greenery in species that rarely use greenery to finish the nest, although this has been done in field studies that tested the drug or nest protection hypothesis (e.g., [139,140]). Knowing the basic biology of a model species is essential to propose and adequately test biologically relevant hypotheses from an evolutionary point of view. In addition, it would make no sense to test the nest-soaking hypothesis [162] in dry breeding environments or in nest-chambers where nests will never be exposed to flooding. However, the external influences modifying the associations between nest design and breeding success (e.g., year effects, [51,168]) cannot be forecasted by researchers that initiate a short-term field study on breeding success, and therefore researchers will often not be able to predict with precision the outcome of such a study. When long-term data sets are available, one option might be to select only the adequate environmental conditions in which a specific hypothesis can be examined. For instance, if the goal would be to test whether better insulated nests buffer best the effects of extreme weather conditions, only nests exposed to heavy rain or cold days might be selected for statistical analyses, eventually to be compared with findings obtained in less extreme weather conditions. Thus, one of the biggest challenges will be to determine the environmental conditions in which a specific hypothesis can be adequately tested in natural conditions. Laboratory experiments that alter availability of nest material or create different environmental conditions of nesting and breeding could also be used to identify proximate underlying mechanisms that link nest design to reproductive success.
Future Research Directions
This review has highlighted inadequacies in our understanding of how nest architecture can impact upon reproductive success in birds. This may reflect our inability to recognize such effects or perhaps, like painted turtles [10], the birds are outwitting us by building optimal nests every time? To answer such questions, we would like to suggest some potential future research direction of intraspecific studies.
Firstly, individual optimization of reproduction predicts that there is a tight positive correlation between clutch size on the one hand and brood size at hatching and fledging on the other hand [175,176]. The question then is to what extent these correlations can be altered by nest characteristics independent from individual-specific abilities to rear nestlings. One approach is to conduct multi-factor analyses that consider clutch size together with nest architectures as potential drivers of breeding success (cf. [60,61]). Most intraspecific field studies of the reproductive consequences of avian nest design did not last more than four years and most experimental studies lasted one or two years. Therefore, the long-term reproductive consequences of nest architectures for next generations remain understudied. The simplest scenario is that certain nest architectures improve the phenotypic qualities of the offspring that also impact on the probability to be recruited into the next breeding population. It is well established that phenotypic qualities of offspring are associated with local recruitment probabilities (e.g., [177,178,179,180,181,182]). However, additional long-term studies will be required to better investigate the long-term effects of intraspecific variation in nest design, which can be a time-consuming task.
Quantitative intraspecific studies of the composition of entire nests in association with environmental factors and breeding success remain rare, even in the best-studied model species. Different types of nest material might improve breeding success in the same way, but because of different reasons or depending on the environmental conditions of breeding (cf. Table 1). Experimental investigations involving animal-derived material or greenery should ideally quantify the composition of entire nests to better control for the potential effects of nest components other than those that have been experimentally manipulated. These studies could be combined with measurements of the environmental factors assumed to have evolutionary shaped the diversity in the species-specific nest architectures. Moreover, few research studies have extracted mammal-derived nest material from bird nests, even though the importance of this nest component has been highlighted for decades. Cavity nests differ in the types of mammal-derived nest material (e.g., thick hair from boar or horses versus wool from sheep versus fur from rabbits versus anthropogenic materials) but the reproductive consequences of this diversity in the properties of mammal-derived nest material remain poorly understood.
Does nest design result from constraints or restraints? Birds may reduce investment in nest building because of constraints (e.g., shortage of nest material, inabilities to carry materials) or evolutionary adaptive decision-making restraining costly reproductive investments [183]. The restraints hypothesis can be tested in environments where building material is superabundant. For instance, Corsican blue tits use small quantities of strongly smelling aromatic Achillea ligustica fragments to adorn the nest [28,122], even in territories where there are hundreds of Achillea plants and only a couple of plants are enough to cover the whole nest or fill up the nest chamber [M. Lambrechts pers. obs.]. One potential reason to restrain delivery of fresh Achillea fragments is that very high volatile concentrations inside the cavity nest might be costly for parents or nestlings. Moreover, in many forest habitats, the superabundance of moss might allow the birds to fill up more than one nest cavity and rapidly construct replacement nests (e.g., [57]). Despite this overabundance of moss, and the fact that the nest-building phase can last several weeks, the nest rim remains at a certain distance below the entrance hole, probably to reduce predation risks or minimize direct wind exposure. Quantifying the availability of nest materials in breeding territories can contribute to a better understanding of the underlying mechanisms responsible for individual-specific or species-specific nest designs (e.g., [49,184]).
Can nest builders and their nest designs predict the future conditions of incubation and nestling rearing? Birds rarely have a reproductive success of 100% given that in many nests not all the eggs hatch or not all the nestlings fledge [160,181,182,185,186,187,188,189]. In addition, the breeders produce an excessive number of eggs per individual lifetime given that in passerines, like great tits, only one out six fledglings will become a breeder that makes contributions to the next generation of breeders [187]. This situation suggests that the nest builders cannot adequately predict and anticipate the future conditions of breeding. However, despite this breeding inefficiency, egg layers use cues (e.g., vernal changes in daylength, ambient temperature, rainfall, food used to form eggs, aspects associated with body condition) to predict and anticipate the optimal breeding time often reflected in the amount of food required to rear the nestlings [190,191]. Therefore, one might ask what cues nest builders can use to predict the future breeding conditions so that the breeding success per nest can be maximized. One of the obvious candidates are aspects associated with ambient temperature (e.g., temperature change) that will influence the thermal conditions of breeding, and of which we assume that it consistently differs across latitudes, altitudes, and time-windows within and between breeding seasons (e.g., [84,86]). Nager and van Noordwijk [192] used ice packs externally placed against one of the walls of nest-boxes to lower the nest-chamber temperatures during the nesting period of Swiss great tits. They reported that the position of the nest cups avoided the coldest nest-chamber wall, which would mean that the nest builders indeed perceived changes in nest-chamber temperatures. However, sample sizes were small, and few repeat studies in natural conditions examined how nest builders adjusted nest shape in responses to changes in nest-chamber temperatures. For instance, Holland and Shutler [142] showed that nest-chamber warming did not change the feathering behaviour of tree swallows, therefore not supporting the hypothesis that nest feathering is used to improve nest insulation. Other studies reported that the height or the shape of the nest are adjusted to the size of the entrance hole, nest-box damage, or nest-chamber illumination, perhaps as a plastic response to nest predation risks or exposure to air flow inside the cavity [62,79,87,193,194,195]. However, we currently do not truly know the physiological mechanisms that translate environmental cues into adaptive breeding decisions [160,191,196,197,198] and their citations. Perhaps body-associated cues expressed via feelings, like experiences of fatigue or cold, might also influence nest-building decisions and associated breeding performances, but we currently do not possess the field methodologies to adequately examine body feelings as cues to predict the future.
Most natural studies of reproductive consequences of nest architectures have been conducted with species that use nest-boxes for breeding. These species have a strongly biased nesting-associated biology compared to species that construct other nest types. Nest-box design can modify the associations between the design of cavity nests and breeding success, for instance when nest-box design influences the size and composition of nest parasites or the insulation properties of nests (e.g., [37,199]). Therefore, it has been recommended for quite a while to take the importance of nest-box design into account during data analysis. However, most research teams continue to use one box type, box types differ between research teams without explaining why, and most of the box types are not those that are preferred by the model species, often not allowing the full expression of the nest. The relative importance of nest-box design and the architecture of cavity nests in the expression of breeding success therefore remains to be clarified.
Finally, there are many multi-factorial influences that affect nest architecture in natural conditions, and so controlled laboratory studies might better identify proximate mechanisms that link nest characteristics to breeding success. For instance, Breen et al. [200] recently used laboratory zebra finches (Taeniopygia guttata) building nests with stiff or flexible string. Higher fledging was associated with use of fewer stiff string pieces and more flexible string pieces in the nest. Although they claimed that they had the first experimental evidence of a link between a physical property of nest material and breeding performance, Breen et al. [200] offered no functional explanation as to why flexibility of string would be important. Quantitative investigations of the breeding consequences of nest types other than cavity nests are more than welcome. Many hypotheses about costs and benefits of nest design make the same predictions of associations between nest characteristics and breeding success in natural conditions (Table 1). An experimental approach combined with a better identification of the relevant environmental conditions of testing will be required to examine more efficiently the causes of costs and benefits of nest design. In addition, field experiments allow to better identify maladaptive nest designs lowering fitness. Urbanization can provide unique research opportunities when nests are exposed to novel environmental conditions, as highlighted in recent review papers (e.g., [201]).
To conclude, we can expect that the level of difficulty needed to understand the underlying mechanisms that link nest design to environmental dynamics and breeding success to be at least as high as that for the best-studied life-history traits, such as the timing of reproduction or clutch size. This review has only dealt with those species that nest within secondary cavities and has not considered the many studies of species that produce their own cavities or nest in the open, and whether these conditions affect reproductive success. We are only just beginning to appreciate the importance of nests in avian reproductive biology and how, and if, these crucial structures can help birds adapt to our rapidly changing world.
Author Contributions
Writing – Original draft preparation M.M.L.; Writing - Review and Editing D.C.D. and M.M.L.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
There are no data associated with this review article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hansell, M. Bird nests and construction behaviour. Cambridge University Press, 2000.
- Healy, S.; Walsh, P.; Hansell, M. Nest building by birds. Current Biology 2008, 18, R271–R273. [Google Scholar] [CrossRef]
- Schaedelin, F.C.; Taborsky, M. Extended phenotypes as signals. Biological Reviews 2009, 84, 293–313. [Google Scholar] [CrossRef]
- Møller, A.P.; Adriaensen, F.; Artemyev, A.V.; Bańbura, J.; Barba, E.; Biard, C.; Blondel, J.; Bouslama, Z.; Bouvier, J.-C.; Camprodon, J.; et al. Variation in nest size in relation to clutch size in birds. Ecol. Evol. 2014, 4, 3583–3595. [Google Scholar] [CrossRef] [PubMed]
- Deeming, D.C. A review of the roles materials play in determining functional properties of bird nests. Acta Ornithol. 2023, 58, 1–28. [Google Scholar] [CrossRef]
- Deeming, D. C. Nest construction in mammals: a review of the patterns of construction and functional roles. Phil. Trans. R. Soc. B 2023, 378, 20220138. [Google Scholar] [CrossRef] [PubMed]
- Svensson, O.; Kvarnemo, C. How sexual and natural selection interact and shape the evolution of nests and nesting behaviour in fishes. Phil. Trans R. Soc. B 2023, 378, 20220139. [Google Scholar] [CrossRef]
- Fischer, E.K. Form, function, foam: evolutionary ecology of anuran nests and nesting behaviour. Phil. Trans R. Soc. B 2023, 378, 20220141. [Google Scholar] [CrossRef] [PubMed]
- O’Fallon, S.; Drager, K.; Zhao, A.; Suarez, A.; Pinter-Wollman, N. Foraging behaviour affects nest architecture in a cross-species comparison of ant nests. Phil. Trans R. Soc. B 2023, 378, 20220146. [Google Scholar] [CrossRef]
- Bodensteiner, B.L.; Iverson, J.B.; Lea, C.A.; Milne-Zelman, C.L.; Michell, T.S.; Refsnider, J.H.; Voves, K.; Warner, D.A.; Janzen, F.J. Mother knows best: nest-site choice homogenises embryo thermal environments among populations in a widespread ectotherm. Phil. Trans R. Soc. B 2023, 378, 20220155. [Google Scholar]
- Dawkins, R. The Extended Phenotype. The Gene as the Unit of Selection. Oxford University Press, 1982.
- Laland, K.N.; Sterelny, K. Perspective: seven reasons (not) to neglect niche construction. Evolution 2006, 60, 1751–1762. [Google Scholar]
- Biddle, L.E.; Broughton, R.E.; Deeming, D.C.; Goodman, A.M. Composition of bird nests is a species-specific characteristic. Avian Biol. Res. 2018, 11, 1–22. [Google Scholar] [CrossRef]
- Dickinson, A.M.; Locke, E.; Gray, L.A.; Bennett, S.L.; Biddle, L.E.; Goodman, A.M.; Deeming, D.C. Composition of nests constructed by species in the Motacillidae, Sylviidae and Prunellidae. Avian Biol. Res. 2022, 15, 21–33. [Google Scholar] [CrossRef]
- Deeming, D.C.; Dickinson, A.M.; Broughton, R.E.; Locke, E.; Gray, L.A.; Bennett, S.L.; Gilchrist, R.; Muniz, S.; Goodman, A.M.; Biddle, L.E. Factors affecting thermal insulation of songbird nests as measured using temperature loggers. Physiol. Biochem. Zool. 2020, 93, 488–504. [Google Scholar] [CrossRef] [PubMed]
- Deeming, D.C.; Gilchrist, R.; Szafraniec, M.; Pollins, J.M. Water vapour conductance of passerine nest walls. Acta Ornithol. 2020, 55, 13–21. [Google Scholar] [CrossRef]
- Biddle, L.E.; Dickinson, A.M.; Broughton, R.E.; Gray, L.A.; Bennett, S.L.; Goodman, A.M.; Deeming, D.C. Nest materials affect the hydrological properties of bird nests. J. Zool. 2019, 309, 161–171. [Google Scholar] [CrossRef]
- Mertens, J.A.L. Thermal conditions for successful breeding in Great Tits (Parus major L.). Oecologia 1977, 28, 1–56. [Google Scholar] [CrossRef]
- Moreno, J. Avian nests and nest-building as signals. Avian Biol. Res. 2012, 5, 238–251. [Google Scholar] [CrossRef]
- Mainwaring, M.C.; Hartley, I.R.; Lambrechts, M.M.; Deeming, D.C. The design and function of birds’ nests. Ecol. Evol. 2014, 4, 3909–3928. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.P. Nest predation selects for small nest size in the blackbird. Oikos 1990, 57, 237–240. [Google Scholar] [CrossRef]
- Grégoire, A.; Garnier, S.; Dréano, N.; Faivre, B. Nest predation in blackbirds and the influence of nest characteristics. Ornis Fenn. 2003, 80, 1–10. [Google Scholar]
- Heeb, P.; Werner, I.; Richner, H.; Kölliker, M. Horizontal Transmission and Reproductive Rates of Hen Fleas in Great 432 Tit Nests. J. Anim. Ecol. 1996, 65, 474–484. [Google Scholar] [CrossRef]
- Mennerat, A.; Charmantier, A.; Perret, P.; Hurtrez-Boussès, S.; Lambrechts, M.M. Parasite intensity is driven by temperature in a wild bird. Peer Community Journal 2021, 1, e60. [Google Scholar] [CrossRef]
- Deeming, D.C.; Reynolds, S.J. Nests, Eggs, and Incubation: New Ideas about Avian Reproduction. Oxford University Press, Oxford, 2015.
- Perez, D.M.; Gardner, J.L.; Medina, I. Climate as an evolutionary driver of nest morphology in birds: A Review. Front Ecol. Evol. 2020, 8, 566018. [Google Scholar] [CrossRef]
- McCabe, R.A. Nest construction by house wrens. Condor 1965, 67, 229–234. [Google Scholar] [CrossRef]
- Petit, C.; Hossaert-McKey, M.; Perret, P.; Blondel, J.; Lambrechts, M.M. Blue tits use selected plants and olfaction to maintain an aromatic environment for nestlings. Ecol. Lett. 2002, 5, 585–589. [Google Scholar] [CrossRef]
- Mennerat, A.; Perret, P.; Lambrechts, M.M. Local individual preferences for nest materials in a passerine bird. PLoS ONE 2009, 4, e5104. [Google Scholar] [CrossRef]
- Harničárová, K.; Adamík, P. Mammal hair in nests of four cavity-nesting songbirds: occurrence, diversity and seasonality. Bird Study 2016, 63, 181–186. [Google Scholar] [CrossRef]
- Glądalski, M.; Wolski, G.J.; Bańbura, M.; Kaliński, A.; Markowski, M.; Skwarska, J.; Wawrzyniak, J.; Bańbura, J. Differences in use of bryophyte species in tit nests between two contrasting habitats: an urban park and a forest. Eur. Zoo. J. 2021, 88, 807–815. [Google Scholar] [CrossRef]
- Hauber, M.E.; Nagy, J.; Sheard, C.; Antonson, N.D.; Street, S.E.; Healy, S.D.; Lala, K.N.; Mainwaring, M.C. Nest architecture influences host use by avian brood parasites and is shaped by coevolutionary dynamics. Proc. R. Soc. Lond. Biol. Sci. 2024, 291, 20231734. [Google Scholar] [CrossRef]
- Mainwaring, M.C. Causes and consequences of intraspecific variation in nesting behaviors: insights from blue tits and great tits. Front Ecol. Evol. 2017, 5, 39. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H. (Ed.) Reproductive success. Studies of individual variation in contrasting breeding systems. The University of Chicago Press, 1988.
- Otter, K.A. (Ed.) Ecology and behavior of chickadees and titmice. An integrated approach. Oxford University Press, 2007.
- Clutton-Brock, T.; Sheldon, B.C. Individuals and populations: the role of long-term, individual-based studies of animals in ecology and evolutionary biology. Trends Ecol. Evol. 2010, 25, 562–573. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Adriaensen, F.; Ardia, D.R.; Artemyev, A.V.; Atienzar, F.; Banbura, J.; Barba, E.; Bouvier, J.-C.; Camprodon, J.; Cooper, C.B.; et al. The design of artificial nestboxes for the study of secondary hole-nesting birds: a review of methodological inconsistencies and potential biases. Acta Ornithol. 2010, 45, 1–26. [Google Scholar] [CrossRef]
- Wesołowski, T. Nest-sites of hole-nesters in a primaeval temperate forest (Białowieża National Park, Poland). Acta Ornithol. 1989, 25, 321–351. [Google Scholar]
- Czeszczewik, D.; Walankiewicz, W.; Mitrus, C.; Nowakowski, W.K. Nest-box data of pied flycatcher Ficedula hypoleuca may lead to erroneous generalizations. Vogelwelt 1999, 120, 361–365. [Google Scholar]
- Maziarz, M.; Broughton, R.K.; Wesołowski, T. Microclimate in tree cavities and nest-boxes: Implications for hole-nesting birds. Forest Ecol. Manage. 2017, 389, 306–313. [Google Scholar] [CrossRef]
- Sudyka, J.; Di Lecce, I.; Szulkin, M. Microclimate shifts in nest-boxes and natural cavities throughout reproduction. J. Avian Biol. 2023, e03000. [Google Scholar] [CrossRef]
- Thompson, E.K.; Keenan, R.J.; Kelly, L.T. The use of nest boxes to support bird conservation in commercially managed forests: a systematic review. For. Ecol. Manage. 2023, 550, 121504. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, X.; Chen, Z. et al. Negative effects of artificial nest boxes on birds: a review. Avian Research 2023, 14, 100101. [CrossRef]
- Markus, J.M.; Hart, A.G.; Goodenough, A.E. Predator-proofing avian nest boxes: a review of interventions, opportunities, and challenges. Birds 2024, 5, 1–23. [Google Scholar] [CrossRef]
- Kelly, C.D. Rate and success of study replication in ecology and evolution. PeerJ 2019, 7, e7654. [Google Scholar] [CrossRef]
- Fraser, H.; Barnett, A.; Parker, T.H.; Fidler, F. The role of replication studies in ecology. Ecol. Evol. 2020, 10, 5197–5207. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Perret, P.; Maistre, M.; Blondel, J. Do experiments with captive non-domesticated animals make sense without population field studies? A case study with Blue Tits’ breeding time. Proc. R. Soc. Lond. B Biol. Sci. 1999, 266, 1311–1315. [Google Scholar] [CrossRef]
- Stroebe, W.; Strack, F. The alleged crisis and the illusion of exact replication. Pers. Psychol. Sci. 2014, 9, 59–71. [Google Scholar] [CrossRef]
- Alambiaga, I.; Álvarez, E.; Diez-Méndez, D.; Verdejo, J.; Barba, E. “The tale of the three little tits”: different nest building solutions under the same environmental pressures. Avian Biol. Res. 2020, 13, 49–56. [Google Scholar] [CrossRef]
- Møller, A.P.; Adriaensen, F.; Artemyev, A.V.; Bańbura, J.; Barba, E.; Biard, C.; Blondel, J.; Bouslama, Z.; Bouvier, J.-C.; Camprodon, J.; et al. Clutch-size variation in Western Palaearctic secondary hole-nesting passerine birds in relation to nest box design. Methods Ecol. Evol. 2014, 5, 353–362. [Google Scholar] [CrossRef]
- Loukola, O.J.; Adamik, P.; Adriaensen, F.; Barba, E.; Doligez, B.; Flensted-Jensen, E.; Eeva, T.; Kivelä, S.M.; Laaksonen, T.; Morosinotto, C.; et al. The roles of temperature, nest predators and information parasites for geographical variation in egg covering behaviour of tits (Paridae). J. Biogeogr. 2020, 47, 1482–1493. [Google Scholar] [CrossRef]
- Alabrudzińska, J.; Kaliński, A.; Słomczyński, R.; Wawrzyniak, J.; Zieliński, P.; Bańbura, J. Effects of nest characteristics on breeding success of Great Tits Parus major. Acta. Ornithol. 2003, 38, 151–154. [Google Scholar] [CrossRef]
- Álvarez, E.; Belda, E.J.; Verdejo, J.; Barba, E. Variation in great tit nest mass and composition and its breeding consequences: a comparative study in four Mediterranean habitats. Avian Biol. Res. 2013, 6, 39–46. [Google Scholar] [CrossRef]
- Álvarez, E.; Barba, E. Nest quality in relation to adult bird condition and its impact on reproduction in great tits. Acta Ornithol. 2008, 43, 3–9. [Google Scholar] [CrossRef]
- Hurtrez-Boussès, S.; de Garine-Wichatitsky, M.; Perret, P.; Blondel, J.; Renaud, F. Variations in prevalence and intensity of blow fly infestations in an insular Mediterranean population of blue tits. Can. J. Zool. 1999, 77, 337–341. [Google Scholar] [CrossRef]
- Glądalski, M.; Kaliński, A.; Markowski, M.; Skwarska, J.; Wawrzyniak, J.; Bańbura, J. Nest size parameter of great tits and blue tits: a long-term study. J. Ornithol. 2023. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Aimé, C.; Midamegbe, A.; Galan, M.-J.; Perret, P.; Grégoire, A.; Doutrelant, C. Nest size and breeding success in first and replacement clutches: an experimental study in blue tits Cyanistes caeruleus. J. Ornithol. 2012, 153, 173–179. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Blondel, J.; Bernard, C.; Caro, S.P.; Charmantier, A.; Demeyrier, V.; Doutrelant, C.; Dubuc-Messier, G.; Fargevieille, A.; de Franceschi, C.; et al. Exploring biotic and abiotic determinants of nest size in Mediterranean great tits (Parus major) and blue tits (Cyanistes caeruleus). Ethology 2016, 122, 492–501. [Google Scholar] [CrossRef]
- Weduwen, D. der; Keogan, K.; Samplonius, J.M.; Phillimore, A.B.; Shut, J.D. The correlates of intraspecific variation in nest height and nest building duration in the Eurasian blue tit Cyanistes caeruleus. J. Avian Biol. 2021, 52, e02528. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Marrot, P.; Fargevieille, A.; Giovannini, P.; Lucas, A.; Demeyrier, V.; Midamegbe, A.; Perret, P.; Grégoire, A.; Charmantier, A.; Doutrelant, C. Nest size is not closely related to breeding success in blue tits: a long-term nest-box study in a Mediterranean oak habitat. Auk 2016, 133, 198–204. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Blondel, J.; de Franceschi, C.; Doutrelant, C. Nest size is positively correlated with fledging success in Corsican blue tits (Cyanistes caeruleus) in an insular oak-dominated habitat mosaic. J. Ornithol. 2017, 158, 125–132. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Haurez, J.; Bodineau, G.; Gagliardi, G.; Maistre, M.; Perret, P.; Pihan, P.; Wilhelm, B.; Wilhelm, J.; Bernard, C.; Blondel, J. Coal tits Periparus ater build larger nests than blue tits Cyanistes caeruleus and great tits Parus major living in the same Mediterranean coniferous woodland habitat. Acta Ornithol. 2016, 51, 123–129. [Google Scholar] [CrossRef]
- Sonnenberg, B.R.; Branch, C.L.; Benedict, L.M.; Pitera, A.M.; Pravosudov, V.V. Nest construction, ambient temperature and reproductive success in a cavity-nesting bird. Anim. Behav. 2020, 165, 43–58. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Charmantier, A.; Demeyrier, V.; Lucas, A.; Perret, S.; Abouladzé, M.; Bonnet, M.; Canonne, C.; Faucon, V.; Grosset, S.; et al. Nest design in a changing world: great tit Parus major nests from a Mediterranean city environment as a case study. Urban Ecosyst. 2017, 20, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, J.G. Aspects of the breeding biology of great tits Parus major L. and blue tits Parus caeruleus L. in Belfast. Ir. Nat. J. 1998, 26, 99–103. [Google Scholar]
- Britt, J.; Deeming, D.C. First-egg date and air temperature affect nest construction in blue tits (Cyanistes caeruleus), but not in great tits (Parus major). Bird Study 2011, 58, 78–89. [Google Scholar] [CrossRef]
- Cruz, Á.; Álvarez, E.; Barba, E. Nest insulation capacity during incubation and after fledging are related. Avian Biol. Res. 2016, 9, 22–27. [Google Scholar] [CrossRef]
- Briggs, K.B. , Deeming D.C. Use of materials in nest construction by pied flycatchers Ficedula hypoleuca reflects localized habitat and geographical location. Bird Study 2016, 63, 516–524. [Google Scholar] [CrossRef]
- Harnist, I.; Dubiec, A.; Mazgajski, T.D. Changes in nest mass in relation to nesting stages in the great tit Parus major. Bird Study 2020, 67, 292–299. [Google Scholar] [CrossRef]
- Lombardo, M.P. Nest architecture and reproductive performance in tree swallows (Tachycineta bicolor). Auk 1994, 111, 814–824. [Google Scholar] [CrossRef]
- Soler, J.J.; Møller, A.P.; Soler, M. 1998. Nest building, sexual selection and parental investment. Evol. Ecol. 1998, 12, 427–441. [Google Scholar] [CrossRef]
- Deeming, D.C.; Pike, T.W. Nest surface temperature predicts fledging success of blue tits Cyanistes caeruleus but not great tits Parus major. Acta Ornithol. 2015, 50, 247–251. [Google Scholar] [CrossRef]
- Tomás, G.; Merino, S.; Martínez-de la Puente, J.; Moreno, J.; Morales, J.; Lobato, E. Determinants of abundance and effects of blood-sucking flying insects in the nest of a hole-nesting bird. Oecologia 2008, 156, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Castaño-Vázquez, F.; Merino, S.; Cuezva, S.; Sánchez-Moral, S. Nest gasses as a potential attraction cue for biting flying insects and other ectoparasites of cavity nesting birds. Front. Ecol. Evol. 2020, 8, 258. [Google Scholar] [CrossRef]
- Slagsvold, T.; Amundsen, T. Do great tits adjust hatching spread, egg size and offspring sex ratio to changes in clutch size? J. Anim. Ecol. 1992, 61, 249–258. [Google Scholar] [CrossRef]
- Mazgajski, T.D.; Rykowska, Z. Dependence of nest mass on nest hole depth in the great tit Parus major. Acta Ornithol. 2008, 43, 49–55. [Google Scholar] [CrossRef]
- Stanback, M.T.; Mercadante, A.N.; Cline, E.L.; Burke, T.H.; Roth, J.E. Cavity depth, not experience, determines nest height in Easthern bluebirds. Wilson J. Ornithol. 2013, 125, 301–306. [Google Scholar] [CrossRef]
- Deeming, D.C.; Morton, F.E.M.; Laverack, K.L. Nest box size affects mass and proportions of materials used in blue tit Cyanistes caeruleus nests. Bird Study 2019, 66, 130–135. [Google Scholar] [CrossRef]
- Holveck, M.-J.; Grégoire, A.; Doutrelant, C.; Lambrechts, M.M. Nest height is affected by lamppost lighting proximity in addition to nestbox size in urban great tits. J Avian. Biol. 2019, 50, e01798.
- Sakraoui, W.; Bouslama, Z.; Belabed, A.I. Does increasing nestboxes size affects breeding success of blue tits (Cyanistes caeruleus)? Study in a locality of Seraidi (Northeast, Algeria). Eco. Env. Cons. 2019, 25, 609–613. [Google Scholar]
- Briggs, K.B.; Deeming, D.C. Effects of year and box size on construction of Eurasian Nuthatch Sitta europaea nests. Ardea, 2022; 110, 61–74. [Google Scholar]
- Järvinen, P.H.; Kluen, E.; Tiiri, M.; Brommer, J.E. Experimental manipulation of blue tit nest height does not support the thermoregulation hypothesis. Ornis Fenn. 2017, 94, 82–91. [Google Scholar] [CrossRef]
- Järvinen, P.H.; Brommer, J.E. Lining the nest with more feathers increases offspring recruitment probability: selection on an extended phenotype in the blue tit. Ecol. Evol. 2020, 10, 13327–13333. [Google Scholar] [CrossRef] [PubMed]
- Deeming, D.C.; Mainwaring, M.C.; Hartley, I.R.; Reynolds, S.J. Local temperature and not latitude determines the design of blue tit and great tit nests. Avian Biol. Res. 2012, 5, 203–205. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Caro, S.P. Egg cooling associated with nest size in a passerine bird. J. Therm. Biol. 2022, 110, 103383. [Google Scholar] [CrossRef]
- Mainwaring, M.C.; Hartley, I.R.; Bearhop, S.; Brulez, K.; du Feu, C.R.; Murphy, G.; Plummer, K.E.; Webber, S.L.; Reynolds, S.J.; Deeming, C.D. Latitudinal variation in blue tit and great tit nest characteristics indicates environmental adjustment. J. Biogeogr. 2012, 39, 1669–1677. [Google Scholar] [CrossRef]
- Kaliński, A.; Wawrzyniak, J.; Bańbura, M.; Skwarska, J.; Zielinski, P.; Gladalski, M.; Bańbura, J. Does the threat of European pine marten (Martes martes) predation influence the height of nests built by blue tits (Cyanistes caeruleus) and great tits (Parus major)? Avian Biol. Res. 2014, 7, 83–90. [Google Scholar] [CrossRef]
- Perrins, C.M. British Tits. Collins, 1979.
- Tomás, G.; Merino, S.; Martínez-de la Puente, J.; Moreno, J.; Morales, J.; Rivero-de Aguilar, J. Nest size and aromatic plants in the nest as sexually selected female traits in blue tits. Behav. Ecol. 2013, 24, 926–934. [Google Scholar] [CrossRef]
- Alworth, T. An experimental test of the function of sticks in the nests of house wrens. Condor 1996, 98, 841–844. [Google Scholar] [CrossRef]
- Slagsvold, T. Experiments on clutch size and nest size in passerine birds. Oecologia 1989, 80, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Glądalski, M.; Kaliński, A.; Wawrzyniak, J.; Bańbura, M.; Markowski, M.; Skwarska, J.; Bańbura, J. Physiological condition of nestling great tits Parus major in response to experimental reduction in nest micro- and macro-parasites. Conserv. Physiol. 2018, 6, coy062. [Google Scholar] [CrossRef] [PubMed]
- Remeš, V.; Krist, M. Nest design and the abundance of parasitic Protocalliphora blow flies in two hole-nesting passerines. Écoscience 2005, 12, 549–553. [Google Scholar] [CrossRef]
- Moreno, J.; Merino, S.; Lobato, E.; et al. Nest-dwelling ectoparasites of two sympatric hole-nesting passerines in relation to nest composition. An experimental study. Écoscience 2009, 16, 418–427. [Google Scholar] [CrossRef]
- Cantarero, A.; López-Arrabé, J.; Saavedra-Garcés, I.; Rodríguez-García, V.; Palma, A.; Moreno, J. The significance of nest structure and nesting material for hole-nesting passerines: an experimental study with Nuthatches Sitta europaea. Acta Ornithol. 2014, 49, 143–155. [Google Scholar] [CrossRef]
- Surgey, J.; du Feu, C.R.; Deeming, D.C. Opportunistic use of a wool-like artificial material as lining of tit (Paridae) nests. Condor 2012, 114, 385–392. [Google Scholar]
- Deeming, D.C.; Mainwaring, M.C. Functional properties of nests. In: Deeming, D.C. and Reynolds, S.J. (Eds.), Nests, eggs, and incubation: new ideas about avian reproduction, pp. 29–49. Oxford University Press, Oxford, 2015.
- Aasen, M.; Slagsvold, T. No cultural transmission of use of nest materials in titmice Paridae. Anim. Behav. 2020, 170, 27–32. [Google Scholar] [CrossRef]
- Winkler, D.W. Use and importance of feathers as nest lining in tree swallows (Tachycineta bicolor). Auk 1993, 110, 29–36. [Google Scholar]
- Lombardo, M.P.; Bosman, R.M.; Faro, C.A.; Houtteman, S.G.; Kluisza, T.S. Effect of feathers as nest insulation on incubation behavior and reproductive performance of tree swallows (Tachycineta bicolor). Auk 1995, 112, 973–981. [Google Scholar] [CrossRef]
- Veiga, J.P.; Polo, V. Feathers in the spotless starlig nests: a sexually selected trait? Behaviour 2011, 148, 1359–1375. [Google Scholar] [CrossRef]
- Sanz, J.J.; García-Navas, V. Nest ornamentation in blue tits: is feather carrying ability a male status symbol? Behav. Ecol. 2011, 22, 240–247. [Google Scholar] [CrossRef]
- García-Navas, V.; Ortega, J.; Ferrer, E.S.; Sanz, J.J. Feathers, suspicions, and infidelities: an experimental study of parental care and certainty of paternity in the blue tit. Biol. J. Linn. Soc. 2013, 109, 552–561. [Google Scholar] [CrossRef]
- Briggs, K.B.; Biddle, L.E.; Deeming, D.C. Geographical location affects size and materials used in the construction of European pied flycatcher (Ficedula hypoleuca) nests. Avian Res. 2019, 10, 17. [Google Scholar] [CrossRef]
- Briggs, K.B.; Deeming, D.C. Localized habitat affects size and materials used in the construction of Common Redstart Phoenicurus phoenicurus nests. Bird Study 2021, 68, 9–20. [Google Scholar] [CrossRef]
- Møller, A.P. On the use of feathers in birds’ nests: Predictions and tests. Ornis Scand. 1984, 15, 38–42. [Google Scholar] [CrossRef]
- White, D.W.; Kennedy, E.D. Effect of egg covering and habitat on nest destruction by house wrens. Condor 1997, 99, 873–879. [Google Scholar] [CrossRef]
- Slagsvold, T. , Wiebe, K.L. Egg covering in cavity nesting birds may prevent nest usurpation by other species. Behav. Ecol. Sociobiol. 2021, 75, 116. [Google Scholar] [CrossRef]
- Reynolds, S.J.; Davies, C.S.; Elwell, E.; Tasker, P.J.; Williams, A.; Sadler, J.P.; Hunt, D. Does the urban gradient influence the composition and ectoparasite load of nests of an urban bird species? Avian Biol. Res. 2016, 9, 224–234. [Google Scholar] [CrossRef]
- Hanmer, H.J.; Thomas, R.L.; Beswick, G.J.F.; Collins, B.P.; Fellowes, M.D.E. Use of anthropogenic material affects bird nest arthropod community structure: influence of urbanisation, and consequences for ectoparasites and fledging success. J. Ornithol. 2017, 158, 1045–1059. [Google Scholar] [CrossRef]
- Girão, J.; Bessa, F.; Garrido-Bautista, J.; Ferreira, B.; Santos-Baena, C.; Marques, M.P.M.; Batista de Carvalho, L.A.E.; Ramos, J.A.; Norte, A.C. Variation in the use of anthropogenic materials in tit nests: influence of human activities and pandemic restrictions. Urban Ecosyst. 2024. [Google Scholar] [CrossRef]
- Jagiello, Z.; Corsini, M.; Dylewski, L.; Ibáñez-Álamo, J.D.; Szulkin, M. The extended avian urban phenotype: anthropogenic solid waste pollution, nest design, and fitness. Science Total Environ. 2022, 838, 156034. [Google Scholar] [CrossRef] [PubMed]
- Clark, L. The nest protection hypothesis: the adaptive use of plant secondary compounds by European starlings. In: Loye, J. E. and Zuk, M. (Eds.). Bird-parasite interactions: ecology, evolution, and behaviour. Oxford, Oxford University Press, pp. 205-221, 1991.
- Dubiec, A.; Gózdz, I.; Mazgajski, T.D. Green plant material in avian nests. Avian Biol. Res. 2013, 6, 133–146. [Google Scholar] [CrossRef]
- Wimberger, P.H. The use of green plant material in bird nests to avoid ectoparasites. Auk 1984, 101, 615–618. [Google Scholar] [CrossRef]
- Gwinner, H.; Oltrogge, M.; Trost, L.; Nienaber, U. Green plants in starling nests: effects on nestlings. Anim. Behav. 2000, 59, 301–309. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Hossaert-McKey, M. Olfaction, volatile compounds and reproduction in birds. Acta Zool. Sinica 2006, 52, 284–287. [Google Scholar]
- Veiga, J.P.; Polo, V.; Vinuela, J. Nest green plants as a male status signal and courtship display in the spotless starling. Ethology 2006, 112, 196–204. [Google Scholar] [CrossRef]
- Garrido-Bautista, J.; Ramos, J.A.; Arce, S.I.; Melero-Romero, P.; Ferreira, R.; Santos-Baena, C.; Guímaro, H.R.; Martín-Villegas, C.; Moreno-Rueda, G.; Norte, A.C. Is there a role for aromatic plants in blue tits (Cyanistes caeruleus) nests? Behav. Ecol. Sociobiol. 2023, 77, 118. [Google Scholar] [CrossRef]
- Eens, M.; Pinxten, R.; Verheyen, R.F. On the function of singing and wing-waving in the European starling Sturnus vulgaris. Bird Study 1990, 37, 48–52. [Google Scholar] [CrossRef]
- Cowie, R.J.; Hinsley, S.A. Timing and return with green vegetation by nesting blue tits Parus caeruleus. Ibis 1988, 130, 553–555. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Dos Santos, A. Aromatic herbs in Corsican blue tit nests: the ‘Potpourri’ hypothesis. Acta Oecol. 2000, 21, 175–178. [Google Scholar] [CrossRef]
- Gwinner, H. The function of green plants in nests of European starlings. Behaviour 1997, 134, 337–351. [Google Scholar] [CrossRef]
- Brouwer, L.; Komdeur, J. Green nest material has a function in mate attraction in the European starling. Anim. Behav. 2004, 67, 539–548. [Google Scholar] [CrossRef]
- Gwinner, H.; Yohannes, E.; Schwabl, H. Nest composition and yolk hormones: do female European starlings adjust yolk androgens to nest quality? Avian Biol. Res. 2013, 6, 307–312. [Google Scholar] [CrossRef]
- Polo, V.; Rubalcaba, J.G.; Veiga, J.P. Green plants in nests reduce offspring recruitment rates in the spotless starling. Behav. Ecol. 2015, 26, 1131–1137. [Google Scholar] [CrossRef]
- Rubalcaba, J.G.; Fuentes, D.; Veiga, J.P.; Polo, V. Nest decoration as social signals by males and females: greenery and feathers in starling colonies. Behav. Ecol. 2017, 28, 1369–1375. [Google Scholar] [CrossRef]
- Soler, J.J.; Ruiz-Castellano, C.; Figuerola, J.; Martín-Vivaldi, M.; Martínez-de la Puente, J.; Ruiz-Rodríguez, M.; Tomás, G. Telomere length and dynamics of spotless starling nestlings depend on nest-building materials used by parents. Anim. Behav. 2017, 126, 89–100. [Google Scholar] [CrossRef]
- Gwinner, H.; Capilla-Lasheras, P.; Cooper, C.; Helm, B. ‘Green incubation’: avian offspring benefit from aromatic nest herbs through improved parental incubation behaviour. Proc. R. Soc. Lond. Biol. Sci. 2018, 285, 20180376. [Google Scholar] [CrossRef]
- Tomás, G.; Merino, S.; Martínez-de la Puente, J.; Moreno, J.; Morales, J.; Lobato, E.; Rivero-de Aguilar, J.; del Cerro, S. Interacting effects of aromatic plants and female age on nest-dwelling ectoparasites and blood-sucking flies in avian nests. Behav. Proc. 2012, 90, 246–253. [Google Scholar] [CrossRef]
- Mennerat, A.; Perret, P.; Bourgault, P.; Blondel, J.; Gimenez, O.; Thomas, D.W.; Heeb, P.; Lambrechts, M.M. Aromatic plants in nests of blue tits: positive effects on nestlings. Anim. Behav. 2009, 77, 569–574. [Google Scholar] [CrossRef]
- Sugasawa, S.; Edwards, S.C.; Stanforth, R.; Bruton, E.; Hansell, M.; Reilly, M.; Healy, S.D. A non-destructive approach to collect nest material data using photographs. Ibis 2021, 163, 1457–1462. [Google Scholar] [CrossRef]
- Bańbura, J.; Blondel, J.; de Wilde-Lambrechts, H.; Perret, P. Why do female blue tits (Parus caeruleus bring fresh plants to their nests? J. Ornithol. 1994, 136, 217–221. [Google Scholar] [CrossRef]
- Rubalcaba, J.G.; Polo, V.; Maia, R.; Rubenstain, D.R.; Veiga, J.P. Sexual and natural selection in the evolution of extended phenotypes: the use of green nesting material in starlings. J. Evol. Biol. 2016, 29, 1585–1592. [Google Scholar] [CrossRef]
- Rendell, W.B.; Robertson, R.J. Nest-site characteristics, reproductive success and cavity availability for tree swallows breeding in natural cavities. Condor 1989, 91, 875–885. [Google Scholar] [CrossRef]
- Wesołowski, T. Clutch size and breeding performance of marsh tits Parus palustris in relation to hole size in a primeval forest. Acta Ornithol. 2003, 38, 65–72. [Google Scholar] [CrossRef]
- Mennerat, A.; Perret, P.; Caro, S.P.; Heeb, P.; Lambrechts, M.M. Aromatic plants in blue tit Cyanistes caeruleus nests: no negative effect on blood-sucking Protocalliphora blow fly larvae. J. Avian Biol. 2008, 39, 127–132. [Google Scholar] [CrossRef]
- Pires, B.A.; Belo, A.D.F.; Rabaça, J.E. Aromatic plants in Eurasian blue tit nests: the ‘nest Protection Hypothesis’ revisited. Wilson J. Ornithol. 2012, 124, 162–165. [Google Scholar] [CrossRef]
- Dawson, R.D. Does fresh vegetation protect avian nests from ectoparasites? An experiment with tree swallows. Can. J. Zool. 2004, 82, 1005–1010. [Google Scholar] [CrossRef]
- Shutler, D.; Campbell, A.A. Experimental addition of greenery reduces flea loads in nests of a non-greenery using species, the tree swallow Tachycineta bicolor. J. Avian Biol. 2007, 38, 7–12. [Google Scholar] [CrossRef]
- Glądalski, M.; Bańbura, M.; Kaliński, A.; Markowski, M.; Skwarska, J.; Wawrzyniak, J.; Zieliński, P.; Bańbura, J. Consequences of experimental addition of fresh, aromatic plants into nests of blue tits (Cyanistes caeruleus) on the physiological condition of nestlings. Behav. Ecol. Sociobiol. 2020, 74, 29. [Google Scholar] [CrossRef]
- Holland, E.R.; Shutler, D. Nest feathering responses by tree swallows (Tachycineta bicolor). J. Ornithol. 2018, 159, 991–998. [Google Scholar] [CrossRef]
- Slagsvold, T. Nest site preference and clutch size in the pied flycatcher Ficedula hypoleuca. Ornis Scand. 1987, 18, 189–197. [Google Scholar] [CrossRef]
- Lens, L.; Wauters, L.A.; Dhondt, A.A. Nest-building by crested tit Parus cristatus males: an analysis of costs and benefits. Behav. Ecol. Sociobiol. 1994, 35, 431–436. [Google Scholar] [CrossRef]
- Mainwaring, M.C.; Benskin, C.; Mc, W.H.; Hartley, I.R. The weight of female-built nests correlates with female but not male quality in the blue tit Cyanistes caeruleus. Acta Ornithol. 2008, 43, 43–48. [Google Scholar] [CrossRef]
- Broggi, J.; Senar, J.C. Brighter great tit parents build bigger nests. Ibis 2009, 151, 588–591. [Google Scholar] [CrossRef]
- Mainwaring, M.C.; Hartley, I. R The energetic costs of nest building in birds. Avian Biol. Res. 2013, 6, 12–17. [Google Scholar] [CrossRef]
- Löhrl, H. Einfluβ des Brutraumfläche auf die Gelegegröβe des Kohlmeise (Parus major). J. Ornithol. 1973, 114, 339–347. [Google Scholar] [CrossRef]
- Löhrl, H. Weitere Versuche zur Frage „Brutraum und Gelegegröβe“ bei des Kohlmeise, Parus major. J. Ornithol. 1980, 121, 403–405. [Google Scholar] [CrossRef]
- Löhrl, H. Experimente zur Bruthöhlenwahl der Kohlmeise (Parus major). J. Ornithol. 1986, 127, S51–S59. [Google Scholar] [CrossRef]
- Karlsson, J.; Nilsson, S.G. The influence of nest-box area on clutch size in some hole-nesting passerines. Ibis 1977, 119, 207–211. [Google Scholar] [CrossRef]
- Van Balen, J.H. The relationship between nest-box size, occupation and breeding parameters of the great tit Parus major and some other hole-nesting species. Ardea 1984, 72, 163–175. [Google Scholar]
- Nilsson, S.G. The evolution of nest-site selection among hole-nesting birds: the importance of nest predation and competition. Ornis Scand. 1984, 15, 167–175. [Google Scholar] [CrossRef]
- Gustafsson, L.; Nilsson, S.G. Clutch size and breeding success of pied and collared flycatchers Ficedula spp. in nest-boxes of different sizes. Ibis 1985, 127, 380–385. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Abouladzé, M.; Bonnet, M.; Demeyrier, V.; Doutrelant, C.; Faucon, V.; le Prado, G.; Lidon, F.; Noell, T.; Pagano, P.; et al. Nest-box size influences where secondary-cavity exploiters roost and nest: a choice experiment. J. Ornithol. 2013, 154, 563–566. [Google Scholar] [CrossRef]
- Slagsvold, T.; Kleiven, K.W.; Eriksen, A.; Johannessen, L.E. Vertical and horizontal transmission of nest-site preferences in titmice. Anim. Behav. 2013, 85, 323–328. [Google Scholar] [CrossRef]
- Demeyrier, V.; Lambrechts, M.M.; Perret, P.; Grégoire, A. Experimental demonstration of an ecological trap for a wild bird in a man-transformed environment. Anim. Behav. 2016, 118, 181–190. [Google Scholar] [CrossRef]
- Maziarz, M.; Wesołowski, T.; Hebda, G.; Cholewa, M. Natural nest sites of Great Tits (Parus major) in a primeval temperate forest (Białowiez_a National Park, Poland). J. Ornithol. 2015, 156, 613–623. [Google Scholar] [CrossRef]
- Deeming, D.C. How does the bird-nest incubation unit work? Avian Biol. Res. 2016, 9, 103–113. [Google Scholar] [CrossRef]
- Kluijver, H.N. The population ecology of the great tit, Parus m. major L. Ardea 1951, 39, 1–135. [Google Scholar]
- Erbelding-Denk, C.; Trillmich, F. Das Mikroklima im Nistkasten und seine Auswikungen auf die Nestlinge beim Star (Sturnus vulgaris). J. Ornithol. 1990, 131, 73–84. [Google Scholar] [CrossRef]
- Wesołowski, T.; Czeszczewik, D.; Rowinski, P.; Walankiewicz, W. Nest soaking in natural holes – a serious cause of breeding failure? Ornis Fenn. 2002, 79, 132–138. [Google Scholar]
- Glądalski, M.; Bańbura, M.; Kaliński, A.; Markowski, M.; Skwarska, J.; Wawrzyniak, J.; Zieliński, P.; Cyżewska, I.; Bańbura, J. Effects of nest characteristics on reproductive performance in Blue Tits Cyanistes caeruleus and Great Tits Parus major. Avian Biol. Res. 2016, 9, 37–43. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Demeyrier, V.; Fargevieille, A.; Giovannini, P.; Lucas, A.; Marrot, P.; Midamegbe, A.; Perret, P.; Charmantier, A.; Doutrelant, C.; Grégoire, A. Great tits build shallower nests than blue tits. Avian Biol. Res. 2014, 7, 251–254. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Caizergues, A.E.; Perrier, C.; Charmantier, A.; Caro, S. Surface temperatures of non-incubated eggs in great tits (Parus major) are strongly associated with ambient temperature. Int. J. Biometeorol. 2020, 64, 1767–1775. [Google Scholar] [CrossRef]
- Mainwaring, M.C. Nest construction and incubation in a changing climate. In: Deeming DC, Reynolds SJ (Eds.) Nests, eggs, and incubation: new ideas about avian reproduction. Oxford University Press, Oxford, pp. 65–74, 2015.
- Lambrechts, M.M.; Rieux, A.; Galan, M.-J.; Cartan-Son, M.; Perret, P.; Blondel, J. Double-brooded great tits (Parus major) in Mediterranean oak habitats: do first broods always perform better than second broods? Russ. J. Ecol. 2008, 39, 516–522. [Google Scholar] [CrossRef]
- Stephenson, S.; Hannon, S.; Proctor, H. The function of feathers in tree swallow nests: insulation or ectoparasite barrier? Condor 2009, 111, 479–487. [Google Scholar] [CrossRef]
- Dawson, R.D.; O’Brien, E.L.; Mlynowski, T.J. The price of insulation: costs and benefits of feather delivery to nests for male tree swallows Tachycineta bicolor. J. Avian Biol. 2011, 42, 93–102. [Google Scholar] [CrossRef]
- Mainwaring, M.C.; Wolfenden, A.; Read, J.E.; Robson, J.M.A.; Tomlinson, C.J.; Hartley, I.R. Feathering the nest: the effects of feather supplementation to blue tit nests. Avian Biol. Res. 2016, 9, 89–95. [Google Scholar] [CrossRef]
- Scott-Baumann, J.F.; Morgan, E.R. A review of the nest protection hypothesis: does inclusion of fresh green plant material in birds’ nests reduce parasite infestation? Parasitology 2015, 142, 1016–1023. [Google Scholar] [CrossRef]
- Pires, B.A.; Belo, A.D.F.; Diamantino, F.; Rabaça, J.E.; Merino, S. Development of nestling blue tits (Cyanistes caeruleus) is affected by experimental addition of aromatic plants. Avian Biol. Res. 2020, 13, 44–48. [Google Scholar] [CrossRef]
- Fauth, P.T.; Krementz, D.G.; Hines, J.E. Ectoparasitism and the role of green nesting material in the Europena starling. Oecologia 1991, 88, 22–29. [Google Scholar] [CrossRef]
- McCarty, J.P.; Secord, A.L. Nest-building behavior in PCB-contaminated tree swallows. Auk 1999, 116, 55–63. [Google Scholar] [CrossRef]
- Pettifor, R.A.; Perrins, C.M.; McCleery, R.H. Individual optimization of clutch size in great tits. Nature 1988, 336, 160–162. [Google Scholar] [CrossRef]
- Tinbergen, J.M.; Both, C. Is clutch size individually optimized? Behav. Ecol. 1999, 10, 504–509. [Google Scholar] [CrossRef]
- Tinbergen, J.M.; Boerlijst, M.C. Nestling weight and survival in individual great tits (Parus major). J. Anim. Ecol. 1990, 59, 1113–1127. [Google Scholar] [CrossRef]
- Both, C.; Visser, M.E.; Verboven, N. Density-dependent recruitment rates in great tits: the importance of being heavier. Proc. R. Soc. Lond. Biol. Sci. 1999, 266, 465–469. [Google Scholar] [CrossRef]
- Naef-Daenzer, B.; Widmer, F.; Nuber, M. Differential post-fledging survival of great and coal tits in relation to their condition and fledging date. J. Anim. Ecol. 2001, 70, 730–738. [Google Scholar] [CrossRef]
- Perrins, C.M.; McCleery, R.H. The effect of fledgling mass on the lives of great tits Parus major. Ardea 2001, 89, 135–142. [Google Scholar]
- Rodríguez, S.; Álvarez, E.; Barba, E. Factors affecting fledgling output of great tits, Parus major, in the long term. Anim. Biodiv. Cons. 2016, 39, 147–154. [Google Scholar] [CrossRef]
- Rodríguez, S.; van Noordwijk, A.J.; Álvarez, E.; Barba, E. A recipe for post fledgling survival in great tits Parus major: be large and be early (but not too much). Ecol. Evol. 2016, 6, 4458–4467. [Google Scholar] [CrossRef]
- Curio, E. Why do young birds reproduce less well? Ibis 1982, 125, 400–404. [Google Scholar] [CrossRef]
- O’Neill, L.G.; Parker, T.H.; Griffith, S.C. Nest size is predicted by female identity and the local environment in the blue tit (Cyanistes caeruleus), but is not related to the nest size of the genetic or foster mother. R. Soc. open sci. 2018, 5, 172036. [Google Scholar] [CrossRef]
- Lack, D. Family-size in titmice of the genus Parus. Evolution 1950, 4, 279–290. [Google Scholar] [CrossRef]
- Blondel, J.; Gosler, A.; Lebreton, J.D.; McCleery, R. (Eds.) Population biology of passerine birds: an integrated approach. NATO ASI Series G, volume 24. Springer-Verlag, Berlin and Heidelberg, 1990.
- Payevsky, V.A. Mortality rate and population density regulation in the great tit, Parus major L.: a review. Russ. J. Ecol. 2006, 37, 180–187. [Google Scholar] [CrossRef]
- Chamberlain, D.E.; Cannon, A.R.; Toms, M.P.; Leech, D.I.; Hatchwell, B.J.; Gaston, K.J. Avian productivity in urban landscapes: a review and meta-analysis. Ibis 2009, 151, 1–18. [Google Scholar] [CrossRef]
- Mägi, M.; Mänd, R.; Tamm, H.; Sisask, E.; Kilgas, P.; Tilgar, V. Low reproductive success of great tits in the preferred habitat: a role of food availability. Écoscience 2009, 16, 145–157. [Google Scholar] [CrossRef]
- Dawson, A. Control of the annual cycle in birds: endocrine constraints and plasticity in response to ecological variability. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 12, 363, 1621–1633. [Google Scholar] [CrossRef]
- Williams, T.D. Physiological adaptations for breeding in birds; Princeton University Press: Princeton, New Jersey, USA, 2012. [Google Scholar]
- Nager, R.G.; van Noordwijk, A.J. Energetic limitations in the egg-laying period of great tits. Proc. R. Soc. Lond. B Biol. Sci. 1992, 249, 259–263. [Google Scholar]
- Stanback, M.T.; Bartholomew, J.E.; Bergner, L.M.; Cline, E.L.; Helms, P.I.; McGovern, P.G.; Millican, D.M.; Roth, J.E. House wrens alter nest architecture to compensate for cavity vulnerability. Wilson J. Ornithol. 2013, 125, 174–178. [Google Scholar] [CrossRef]
- Maziarz, M.; Wesołowski, T. Does darkness limit the use of tree cavities for nesting birds? J. Ornithol. 2014, 155, 793–799. [Google Scholar] [CrossRef]
- Podkowa, P.; Surmacki, A. The importance of illumination in nest site choice and nest characteristics of cavity nesting birds. Scientific Reports 2017, 7, 1329. [Google Scholar] [CrossRef] [PubMed]
- Tollenaar, D. Legperioden en eierproductie bij eenige wilde vogelsoorten, vergeleken met die bij hoenderrassen. Mededeelingen Landbouwhoogeschool Wageningen 1922, 23, 1–46. [Google Scholar]
- Caro, S.P.; Schaper, S.V.; Hut, R.A.; Ball, G.F.; Visser, M.E. The case of the missing mechanism: how does temperature influence seasonal timing in endotherms? PLoS Biol. 2013, 11, e1001517. [Google Scholar] [CrossRef] [PubMed]
- Bonamour, S.; Chevin, L.-M.; Charmantier, A.; Teplitsky, C. Phenotypic plasticity in response to climate change: the importance of cue variation. Phil. Trans. R. Soc. Lond. B 2019, 374, 20180178. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.P. Parasites, predators and nest boxes: facts and artefacts in nest box studies of birds? Oikos 1989, 56, 421–423. [Google Scholar] [CrossRef]
- Breen, A.J.; Healy, S.D.; Guillette, L.M. Reproductive consequences of material use in avian nest construction. Behav. Proc. 2021, 193, 104507. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, S.J.; Ibáñez-Álamo, J.D.; Sumasgutner, P.; Mainwaring, M.C. Urbanisation and nest building in birds: a review of threats and opportunities. J. Ornithol. 2019, 160, 841–860. [Google Scholar] [CrossRef]
- Slagsvold, T. On the evolution of clutch size and nest size in passerine birds. Oecologia 1989, 79, 300–305. [Google Scholar] [CrossRef]
- Tomás, G.; Merino, S.; Moreno, J.; Sanz, J.J.; Morales, J.; Garcia-Fraílle, S. Nest weight and female health in the Blue Tit (Cyanistes caeruleus). Auk 2006, 123, 1013–1021. [Google Scholar] [CrossRef]
- Smith, J.A.; Harrison, T.J.E.; Martin, G.R.; Reynolds, S.J. Feathering the nest: food supplementation influences nest construction by blue (Cyanistes caeruleus) and great tits (Parus major). Avian Biol. Res. 2013, 6, 18–25. [Google Scholar] [CrossRef]
- Martínez-de la Puente, J.; Merino, S.; Lobato, E.; Moreno, J.; Tomás, G.; Morales, J. Male nest-building activity influences clutch mass in pied flycatchers Ficedula hypoleuca. Bird Study 2009, 56, 264–267. [Google Scholar] [CrossRef]
- Hilton, G.M.; Hansell, M.H.; Ruxton, G.D.; Reid, J.; Monaghan, P. Using artificial nests to test importance of nesting material and nest shelter for incubation energetics. Auk 2004, 121, 777–787. [Google Scholar] [CrossRef]
- Polo, V.; Veiga, J.P. Nest ornamentation by female spotless starlings in response to a male display: an experimental study. J. Anim. Ecol. 2006, 75, 942–947. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.D.; Doherty, P.F.; Piaggio, A.J.; Huyvaert, K.P. Sex and nest type influence avian blood parasite prevalence in a high-elevation bird community. Parasites Vectors 2021, 14, 145. [Google Scholar] [CrossRef]
- Clark, L.; Mason, J.R. Effect of biologically active plants used as nest material and the derived benefits to starling nestlings. Oecologia 1988, 77, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Soler, J.J.; Cuervo, J.J.; Møller, A.P.; de Lope, F. Nest building is a sexually selected behaviour in the Barn Swallow. Anim. Behav. 1998, 56, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Mainwaring, M.C.; Deeming, D.C.; Jones, C.I.; Hartley, I.R. Adaptive latitudinal variation in common blackbird Turdus merula nest characteristics. Ecol. Evol. 2014b, 4, 841–851. [Google Scholar] [CrossRef]
- Crossman, C.A.; Rohwer, V.G.; Martin, P.R. Variation in the structure of bird nests between northern Manitoba and southeastern Ontario. PLoS ONE 2011, 6, e19086. [Google Scholar] [CrossRef]
- Deeming, D.C.; Merrils-Brown, M. 2024 Architecture differentially affects thermal properties of Song Thrush Turdus philomelos nest walls. Bird Study 2024, in press. [Google Scholar]
Figure 1.
A hypothetical relationship between nest insulation and environmental temperature.
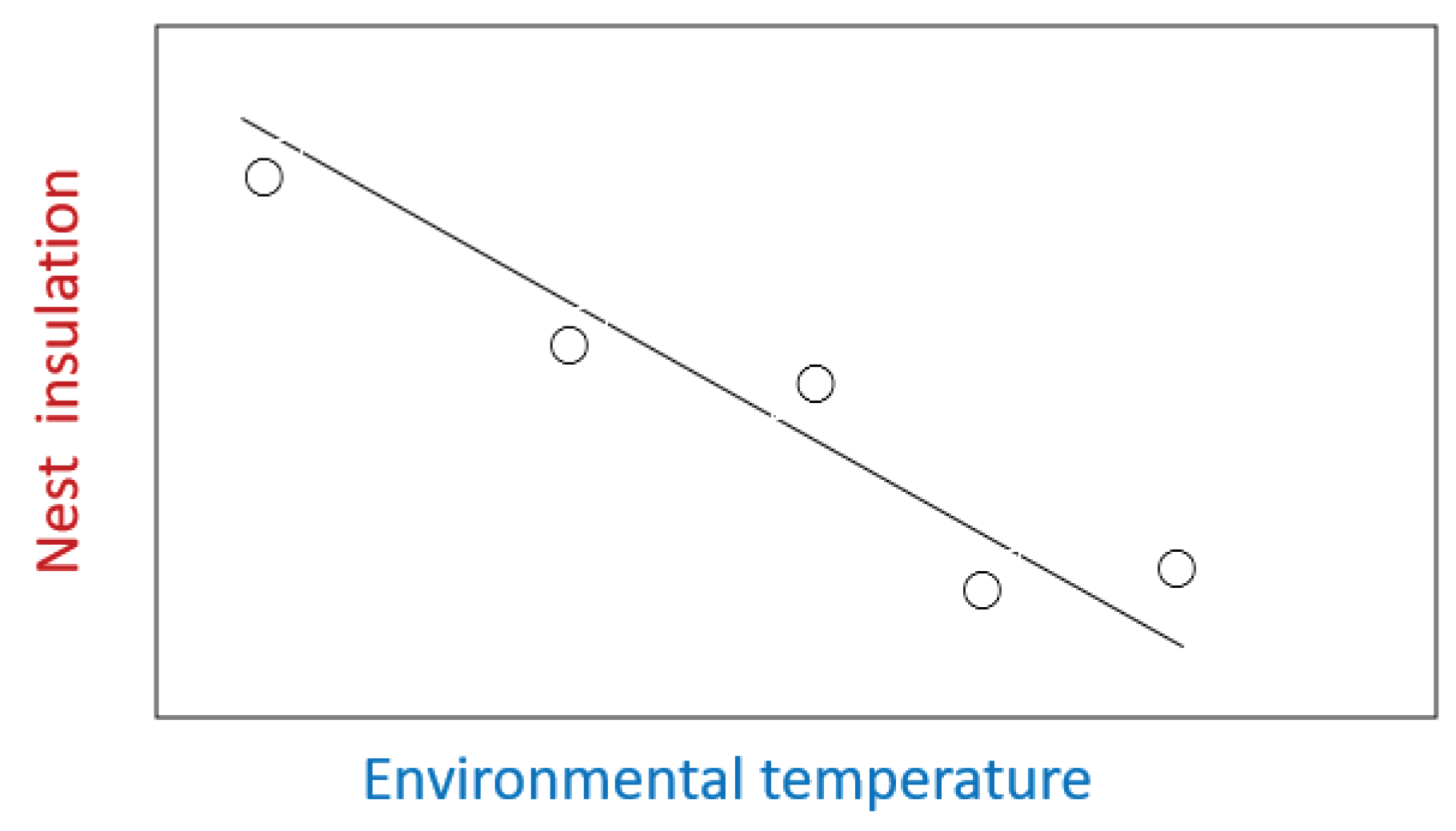
Figure 2.
A. Hypothetical indication of a constant nest insulation over many years (red line on left panel) with the blue line indicating the environmental temperature. Nest insulation only matches the environmental temperature in certain years (arrows). B. the right panel shows the reproductive performance of the birds building the nests with optimal conditions producing the highest reproductive performance at some environmental temperatures. For cool temperatures the nest insulation is sub-optimal and reproductive performance is reduced. At high temperatures there is too much insulation and reproductive performance also suffers.
Figure 2.
A. Hypothetical indication of a constant nest insulation over many years (red line on left panel) with the blue line indicating the environmental temperature. Nest insulation only matches the environmental temperature in certain years (arrows). B. the right panel shows the reproductive performance of the birds building the nests with optimal conditions producing the highest reproductive performance at some environmental temperatures. For cool temperatures the nest insulation is sub-optimal and reproductive performance is reduced. At high temperatures there is too much insulation and reproductive performance also suffers.
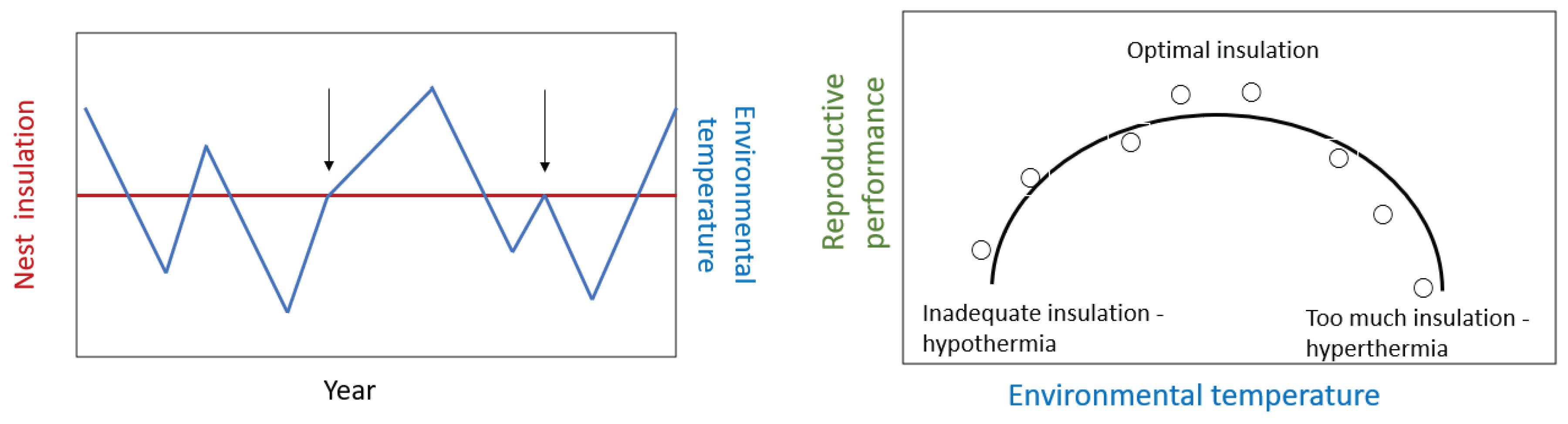
Figure 3.
A. Hypothetical indication of changes in nest insulation that inversely match environmental temperature over many years (red line and blue lines, respectively, on left panel). Nest insulation matches the environmental temperature and so is optimal most years. B. Reproductive performance is unaffected by prevailing environmental temperatures because nest insulation is always close to optimum allowing, all other things being equal, reproductive performance to be maximised every year (right panel).
Figure 3.
A. Hypothetical indication of changes in nest insulation that inversely match environmental temperature over many years (red line and blue lines, respectively, on left panel). Nest insulation matches the environmental temperature and so is optimal most years. B. Reproductive performance is unaffected by prevailing environmental temperatures because nest insulation is always close to optimum allowing, all other things being equal, reproductive performance to be maximised every year (right panel).
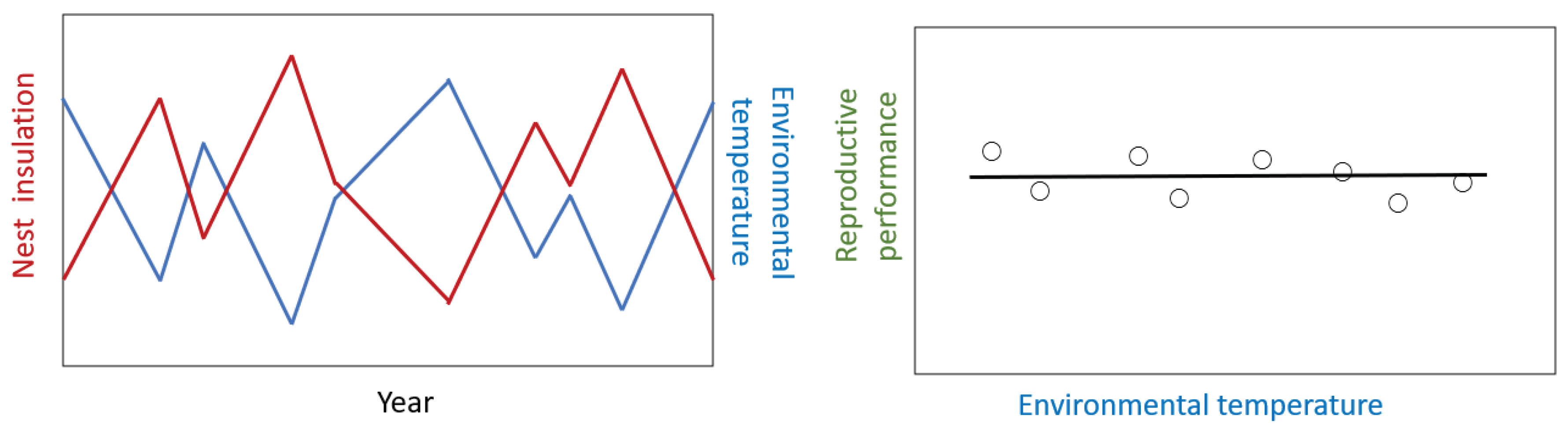

Table 1.
Hypotheses and associated predictions about associations between nest characteristics and aspects of breeding performance in secondary-cavity nesters. Species key: GT = great tit (Parus major); BT = blue tit (Cyanistes caeruleus); TS = tree swallow (Tachycineta bicolor); ES = European starling (Sturnus vulgaris); SS = spotless starling (Sturnus unicolor); CT = coal tit (Poecile ater); MT = marsh tit, (Periparus palustris); PF = pied flycatcher (Ficedula hypoleuca); CF = collared flycatcher (Ficedula albicollis); EN = European nuthatch (Sitta europaea); HW = house wren (Troglodytes aedon).
Table 1.
Hypotheses and associated predictions about associations between nest characteristics and aspects of breeding performance in secondary-cavity nesters. Species key: GT = great tit (Parus major); BT = blue tit (Cyanistes caeruleus); TS = tree swallow (Tachycineta bicolor); ES = European starling (Sturnus vulgaris); SS = spotless starling (Sturnus unicolor); CT = coal tit (Poecile ater); MT = marsh tit, (Periparus palustris); PF = pied flycatcher (Ficedula hypoleuca); CF = collared flycatcher (Ficedula albicollis); EN = European nuthatch (Sitta europaea); HW = house wren (Troglodytes aedon).
| Hypothesis | Prediction | Species studied | Analysis and conditions tested | Conditions of expression or selective factors | Reference |
|---|---|---|---|---|---|
| Nest-size components | |||||
| Direct effects | |||||
| Bigger cavity nests reduce egg breaking risks | Less nest desertion or more hatchlings in bigger nests | GT | Not tested | Cavities with irregular floor and wall surfaces | 54 |
| Bigger nests can physically support heavier or larger broods | Larger clutches and larger broods in larger nests | Cavity-nesting passerines GT, BT, PF, CF |
Correlative Experimental |
Cavity size (depth, floor area) | 4, 143, 149, 202 |
| Bigger nests allow nest cup expansion reducing competition for space or overheating in crowded nest cups | More fledglings in nest cups that expand more | BT, PF, GT, TS | Experiments with artificial nests in BT and PF or very small boxes in GT | Bigger nestlings and larger broods in larger cavities, also depending on ambient temperatures | 18, 70, 91, 152, 202 |
| Thicker nests reduce negative effects of nest compression due to growing nestlings | Bigger or heavier broods in thicker nests | GT, BT | Not tested | Cavities with irregular floor and wall surfaces, and nest bases mainly built from soft nest material, like moss | 4, 72 |
| Bigger nests provide better insulation improving incubation or brooding when the incubator is on the eggs | More ectotherm nestlings, or better growing embryos or nestlings, in bigger nests | GT, BT | Correlative Experimental |
Cold weather | 72, 82, 84, 86 |
| Bigger or asymmetric nests can block air flows in damaged cavities containing cracks or slits | Improved incubation efficiency increasing survival probabilities of ectotherm embryos or nestlings | GT, BT, CT | Not tested | Cold and windy weather | 62 |
| Bigger nests hamper cooling of species-specific eggs or ectotherm nestlings when the incubator or brooder is off the nest | Better egg survival or improved embryo growth in bigger nests | GT | Laboratory | Cold weather | 85 |
| Smaller nest cups reduce heat loss during incubation | Higher hatching success in smaller nest cups | BT, PF | Experiments with artificial nests, but not tested | Cold weather | 91 |
| Bigger nests reduce nest soaking risks | Less nest desertion, and higher breeding success in bigger and thicker nests | PF, CF, MT, EN | Natural cavities Anecdotal |
Cavities exposed to heavy rain | 162 |
| Nest size determines exposure to nest parasites, depending on the types of parasites. Larger nests might harbor more non-flying nest parasites or produce more gasses that attract more flying nest parasites |
Breeding success associated with nest parasitism but depending on the types of parasites infesting the nest. Some nest parasites might vaccinate bird hosts potentially improving contributions to next generations. |
GT, BT | Correlative | Cavities exposed to nest parasites. Climate-dependent parasite activity and growth |
23, 24, 73 |
| Bigger nests reduce contact with cavity walls increasing sanitary conditions | More hatchlings or fledglings, and better growing embryos or nestlings, in bigger nests | GT | Not tested | Wetter cavities promoting micro-organism development | 52 |
| Nest size controls cavity illumination influencing predation risks or abilities to perceive nestling phenotypes | Darker nests suffer less from nest predation, and cavity illumination is associated with nestling phenotypes and brood characteristics | GT | Correlative Experimental, but not tested |
Light intensity combined with structure of predator community | 79, 194, 195 |
| Smaller nests with lower insulation properties prevent overheating of eggs prior to incubation | Higher egg hatching success in smaller, less insulated, nests | Cavity-nesting passerines TS |
Not tested | Hot conditions and direct sunlight exposure combined with larger clutches favour asynchronous hatching | 166 M. Lambrechts (Idea) |
| Smaller nests with lower insulation properties prevent overheating of nestlings | Higher fledging success in smaller, less insulated, nests | GT | Not tested | Hot conditions combined with larger broods | 18 |
| Smaller or thinner nests reduce nest predation risks | More eggs, hatchlings, or fledglings in smaller or thinner nests | GT, BT | Correlative, Experimental | High nest predation pressures | 60, 61, 76, 87 |
| Dry nests lower cooling of eggs or ectotherm nestlings. Smaller moss-based nests dry out more quickly and therefore built in conditions with more rainfall | More hatchlings and fledglings in smaller moss-based nests | GT, BT | Correlative Not tested |
Cavities frequently exposed to rainfall | 58, 91 |
| Indirect effects | |||||
| Better parents produce bigger nests | Less nest desertion, larger clutches, and more hatchlings or fledglings, in bigger nests | BT, GT | Correlative | Individual-specific physiology | 54, 144, 145, 146, 203 |
| Territories providing more resources contain bigger nests | Less nest desertion, larger clutches, and more hatchlings or fledglings, in bigger nests | BT | Food supplementation | Individual-specific resource availability | 54, 147, 204 |
| Bigger nests attract better mates or stimulate reproductive investment | Larger clutches, more hatchlings, and more fledglings in bigger nests | PF, BT | Correlative Experimental |
Individual-specific resource availability and post-mating investments in nest building | 71, 89, 205, 210 |
| Animal-derived nest material (ADNM) | |||||
| Direct effects | |||||
| Nests with more ADNM better hide clutches reducing interspecific competition or predation risks | Less nest desertion or more hatchlings in nests with more ADNM | HW, GT, BT | Correlative, Experimental |
Environments with more competitors | 51, 107, 108 |
| ADNM creates a physical barrier to keep incubators, eggs and nestlings dry in wet moss-based nests. Wet nests accelerate cooling | More hatchlings or ectotherm nestlings, or better growing embryos or nestlings in wet nests with more ADNM | Cavity-nesters mainly using moss GT, BT |
Not tested | Wet environments combined with use moss to build the foundation of the nest given that moss-based materials rapidly absorb rainwater and slowly dry out, whereas hair or fur dry out faster | 91 M. Lambrechts & D.C. Deeming (Idea) |
| Nests with more ADNM provide better insulation when the incubator is on the eggs or ectotherm nestlings | More hatchlings or ectotherm nestlings, or better growing embryos or nestlings, in nests with more ADNM | Small-bodied passerines, TS, BT, GT | Correlative Experiments |
Colder climates and smaller bird species | 51, 66, 100, 106, 142, 147, 168, 170 |
| Nests with more ADNM hamper cooling of eggs or ectotherm nestlings when the incubator or brooder is off the eggs | More hatchlings or ectotherm nestlings, or better growing embryos or nestlings, in nests with more ADNM | GT, TS | Laboratory Not tested in the field |
Colder climates and smaller bird species | 99, 100, 106, 165, 206 |
| ADNM creates a physical barrier against nest parasites | Ectotherm and endotherm nestlings grow better in nests with more ADNM | TS, GT, BT | Correlative, Experimental |
Cavity nests with nest parasites | 92, 99, 100, 168, 170 |
| ADNM provides comfort during incubation or parental care stages | More hatchlings or fledglings in nests with more ADNM | SS | Not tested | Nest foundations built from hard nest material | 207 |
| Nests with less visible ADNM components attract less predators | Less nest desertion, and more hatchlings or fledglings, in nests with less ADNM | Passerines with nest rims close to the entrance hole | Not tested | Environments with nest predators. More likely in open nesters. | 106 |
| Nests with more mammal-derived material reduce risks of nest take overs or brood parasitism | Less nest desertion, and more hatchlings or nestlings, in nests with more mammal-derived material | GT | Correlative Experimental |
Environments with nest predators or competitors | 51, 108 |
| Nests with less ADNM decrease risks associated with premature egg heating or hyperthermia in nestlings | More hatchlings or fledglings in nests with less ADNM | Passerines with abundant ADNM | Not tested | Large broods combined with hot weather | 106 M. Lambrechts & D.C. Deeming (Idea) |
| Indirect effects | |||||
| Better parents deliver more ADNM used in status signaling | More hatchlings and fledglings in nests with more ADNM | SS, BT | Correlative Experimental |
Spatiotemporal variation in availability combined with increased intraspecific competition | 83, 102, 207 |
| Better territories result in more ADNM | More hatchlings and fledglings in nests with more ADNM | GT, BT | Food supplementation | Spatiotemporal variation in availability | 204 |
| Nests with more ADNM attract better mates or stimulate reproductive investment | More hatchlings and fledglings in nests with more ADNM | SS, BT | Experimental | Spatiotemporal variation in availability combined with increased intraspecific competition | 102, 207 |
| ADNM induces costly intraspecific competition | More nest desertion and lower breeding success or lower-quality nestlings in nests with more ADNM | BT | Experimental | High population densities promoting intraspecific competition | 103, 170 |
| Greenery | |||||
| Direct effects | |||||
| Greenery protects the roosting or incubating parents against pathogens or blood parasites | Breeders, and consequently nestlings, in better physical condition in nests with more greenery | BT | Not tested | Cavities increasing risks associated with pathogens or invertebrate vector exposure | 121, 208 |
| Greenery reduces nest parasitism reducing deleterious effects on breeders or nestlings | Breeders or nestlings in better physical condition in nests with more greenery | BT, ES, SS, TS | Correlative Experimental |
Cavities increasing risks associated with nest parasite exposure | 124, 131, 140, 172, 209 |
| Greenery produces more volatile compounds improving health status or immunocompetence | Breeders or nestlings grow better and have a higher body condition in nests with more greenery | ES | Experimental | All cavity types | 116, 129 |
| Greenery provides comfort during incubation | More hatchlings in nests with more greenery | ES | Correlative | All cavity types | 129 |
| Indirect effects | |||||
| Better parents deliver more greenery used in status signaling | Less nest desertion, and higher breeding success or higher-quality nestlings in nests with more greenery | SS, B | Experimental | Spatiotemporal variation in availability combined with increased intraspecific competition | 89, 126, 207 |
| Better territories result in more greenery delivered to the nest | Less nest desertion, and higher breeding success or higher-quality nestlings in nests with more greenery | Passerines from dry or heavily urbanized regions BT, ES, SS |
Not tested | Climate change reduces availability of nest material. Spatiotemporal variation in greenery availability perhaps covarying with other essential humidity-associated resources required for reproduction |
166 M. Lambrechts & D.C. Deeming (Idea) |
| More greenery improves mate attraction or stimulates reproductive investment in partners or offspring | Less nest desertion, and higher breeding success or higher-quality nestlings in nests with more greenery | ES, SS, BT | Experimental | Spatiotemporal variation in availability combined with increased intraspecific competition | 89, 118, 124, 207 |
| Greenery induces costly intraspecific competition | More nest desertion and lower breeding success or lower-quality nestlings in nests with more greenery | SS | Experimental | High population densities promoting intraspecific competition | 126 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



