
Submitted:
04 March 2024
Posted:
05 March 2024
You are already at the latest version
Abstract
Spermatogenesis is critical for insect reproduction and is regulated by many different genes. In this study, we found that Forkhead transcription factor Fd59a functions as a key factor in spermatogenesis of Drosophila melanogaster. Fd59a contains a conversed Forkhead domain, and it is clustered to the FoxD subfamily with other FoxD members from some insect and vertebrate species. Mutation in Fd59a caused swelling in the apical region of the testis. More importantly, only a few mature sperm were present in the seminal vesicle of Fd59a mutant flies compared to the control flies, and the fertility of Fd59a2/2 mutant males was significantly lower than that of the control flies. Immunofluorescence staining showed that the homeostasis of testis stem cell niche in Fd59a2/2 mutant and Fd59a RNAi flies was disrupted, and apoptosis of sperm bundles was increased. Furthermore, results from RNA-sequencing and qRT-PCR suggest that Fd59a can regulate expression of genes related to reproductive process and cell death. Taken together, our results indicated that Fd59a plays a key role in the spermatogenesis of Drosophila.
Keywords:
Spermatogenesis
; Forkhead
; FoxD
; Testis
; Apoptosis
1. Introduction
Forkhead box (Fox) transcription factor, which contains a highly conserved DNA binding domain of ~100 amino acids consisting of three α helices, three β folds and two ring connections, plays critical roles in organ development, innate immunity, and other processes [1]. Based on phylogenetic analysis, Fox proteins are assigned to different subclasses and named “Fox, subclass N, member X” [2]. FoxD subfamily members are mainly involved in metabolism and early organ development [3]. In mammals, FoxD1 regulates human early embryonic development and is associated with various diseases. For example, FoxD1 can promote SLC2A1 (Solute carrier family 2 member 1) transcription and inhibit degradation of SLC2A1 to facilitate proliferation, invasion, and metastasis of pancreatic cancer cells [4,5,6]. In planarian, FoxD gene expression was induced by wound signaling, and it was involved in head regeneration [7]. In Drosophila melanogaster, Fd59a/FoxD may regulate development of nervous system and control egg-laying behavior of females [8]. However, the functions of insect FoxD subfamily members are still largely unknown.
Spermatogenesis is a process to produce mature sperm for reproduction. The process of spermatogenesis is conserved from insects to vertebrates; thus, insect testis is an ideal model for studying mechanisms of spermatogenesis [9]. In D. melanogaster, germ stem cells (GSCs) differentiate into goniablasts under the control of stem cell niche, then goniablasts develop into spermatids through mitosis and meiosis. After nuclear elongation and individualization processes, round spermatids finally become mature sperm [10].
Spermatogenesis is regulated by various signaling pathways, such as JAK-STAT (Janus kinase-signal transducer and activator of transcription), BMP (Bone morphogenetic protein), and EGF (Epidermal growth factor) pathways [11], and by many genes [12,13]. Recent studies showed that different genes are involved in spermatogenesis of insects. For example, knockdown expression of ribosomal protein S3 (RpS3) strongly disrupted spermatid elongation and individualization process in D. melanogaster [14]. Knockdown or mutation of cytochrome c1-like (cyt-c1L) gene in early germ cells resulted in male sterility of D. melanogaster [15]. Moreover, BmHen1, a gene in Bombyx mori encoding methyltransferase that modifies piRNAs, was found to regulate eupyrene sperm development [16]. These results indicate that the molecular mechanism of insect spermatogenesis is much more complicated than what we have already known about.
In our previous study, we showed that B. mori FoxA participated in the development of wing disc [17]. Microarray data showed that Fox genes were expressed in B. mori testis and BmFoxD was expressed at a high level [18]. In Drosophila, expression of Fd59a/FoxD was about 2-fold higher in the testis than in the ovary [19], suggesting that Fd59a may play a role in testis development or spermatogenesis. In this study, we showed that mutation and knockdown expression of Fd59a caused swelling in the apical region of testis and decreased male fertility. More importantly, loss of function in Fd59a disrupted the homeostasis of testis stem cell niche and induced apoptosis of sperm bundles, resulting in only a few mature sperm in the seminal vesicle. By analyzing RNA sequencing from the testis of Fd59a2/2 mutants, we found that Fd59a may regulate expression of genes related to reproductive and metabolic processes. Our findings suggest that Fd59a plays a role in Drosophila spermatogenesis.
2. Materials and Methods
2.1. Fly Lines
Wild-type w1118 line was maintained in the laboratory [20]. Nos-Gal4 (TB00040) and UAS-GFP dsRNA (BDSC9331) fly lines were obtained from Tsinghua Fly Center in Beijing, China. Fd59a1/CyO (BDSC56819), Fd59a2/CyO (BDSC56820) and UAS-Fd59a RNAi (BDSC31937) flies were obtained from the Bloomington Drosophila Stock Center in Indiana, USA. Bam-Gal4 fly line was kindly provided by Professor Yufeng Wang at the School of Life Sciences, Central China Normal University, Wuhan, China.
To analyze functions of Fd59a, Fd59a1/CyO males were crossed to Fd59a1/CyO females to generate Fd59a1/1 loss of function flies, while Fd59a2/CyO males were crossed to Fd59a2/CyO females to generate Fd59a2/2 loss of function flies. To knockdown expression of Fd59a, Nos-Gal4 and Bam-Gal4 flies were crossed to UAS-Fd59a RNAi flies, respectively.
All flies were reared on a fresh cornmeal/yeast/brown sugar diet with p-hydroxybenzoic acid methylester as a mold inhibitor at 25°C with a photoperiod of approximately 12 L : 12 D (light : dark) [21].
2.2. Bioinformatics Analysis
Amino acid homology search was performed by BLAST at the National Center for Biotechnology Information (NCBI, https://blast.ncbi.nlm.nih.gov). Sequence alignment was done by Cluster W. Phylogenetic tree was constructed by RAxML (Random Axelerated Maximum Likelikhood) with 1000 bootstrap replications [22]. Functional protein domains were analyzed by the SMART (https://smart.embl.de/). Potential Fox binding sites in the promoter sequences of select genes were predicted using the JASPAR program (https://jaspar.genereg.net/).
2.3. RNA Isolation and Quantitative RT-PCR
To analyze the expression pattern of Fd59a at different developmental stages, around 50 adult flies of w1118 (male : female ratio is ~2 : 1) were collected and mated in cage. Then flies were transferred to a new cage 2 h later. Based on the developmental stages, embryos at 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, and 24 h after egg-laying, and the first, second, third instar larvae, early, mid and late pupae, and 1, 3 and 5-day-old adults were collected. All samples were collected into RNAex Pro reagent (Accurate Biology, Changsha, China) and kept at -80°C.
Total RNA was extracted as described previously [20]. The first-strand cDNA was synthesized from 1 μg of total RNA using HiFiScript gDNA removal cDNA Synthesis Kit (cwbiotech, Taizhou, China). Gene specific primers (Table 1) for the tested genes were designed based on sequences from the Flybase and synthesized by Tsingke Biotechnology (Beijing, China).
Quantitative reverse transcription – polymerase chain reaction (qRT-PCR) was performed using QuantStudio™ 6 Flex (Thermo Fisher Scientific, Waltham, USA) with ChamQ SYBR qPCR Master Mix (Vazyme, Nanjing, China) following the manufacturer’s instructions. The qPCR cycling program was 95°C for 30 s, followed by 40 cycles of 95°C for 10 s, 60°C for 30 s. Relative expression level of genes was calibrated against the reference gene Rp49 by using 2−ΔΔCt calculation method [23].
2.4. Male Fertility Test
For fertility test, 10 1-day-old virgin females and 15 3-day-old males were placed in bottles with egg collection plates as described previously [24]. Egg collection plates were changed every 24 h, and the number of embryos and larvae in the plates were counted.
2.5. Immunofluorescence Staining
Testes from 3-day-old adult flies were dissected in 10 mM phosphate-buffered saline (PBS) (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4, pH 7.4) and fixed in 4% paraformaldehyde for 30 min at room temperature. Testis samples were washed in 3‰ PBT (10 mM PBS containing 0.1% Triton X-100) for 3 or more times (each for 15 min) and blocked with blocking buffer (3‰ PBT containing 5% normal goat serum) for 1 h at room temperature. Next, all samples were incubated with primary antibody prepared in dilution buffer (3‰ PBT containing 3% normal goat serum) at 4°C overnight, washed three times in 3‰ PBT, and then incubated with secondary antibody at room temperature for 3 h in darkness. Finally, samples were washed three times with 3‰ PBT, mounted using VECTASHIELD® antifade Mounting Medium containing 2 μg/ml DAPI (Vector Laboratories, Newark, USA), and observed under confocal microscopy.
Primary antibodies used were anti-Vasa (1:50, Developmental Studies Hybridoma Bank, Iowa, USA), anti-Fasciclin III (Fas III) (1:100, Developmental Studies Hybridoma Bank, 7G10, Iowa, USA), anti-αSpectrin (1:50, Developmental Studies Hybridoma Bank, 3A9, Iowa, USA). The secondary antibodies were goat anti-rabbit 568 and goat anti-mouse 488, which were Alexa Fluor-conjugated (1:500, Invitrogen, Carlsbad, USA). Fluorescent images were captured using an FV3000 confocal microscope (Olympus, Tokyo, Japan).
2.6. TUNEL Assay
To analyze whether loss of function in Fd59a can induce cell death of sperm bundle, testes from 3-day-old adults of Fd59a2/2 and Fd59a RNAi flies were dissected. The collected testes were fixed in 4% paraformaldehyde and washed with 3‰ PBT as described above.
Terminal deoxyribonucleotide transferase (TdT)-mediated dUTP nick end labeling (TUNEL) assay was performed using the one-step TUNEL apoptosis assay kit from Beyotime Biotechnology (Shanghai, China). Testis samples prepared from above were incubated with 50 μL TUNEL reaction mixture containing 5 μL of TdT enzyme and 45 μL of fluorescent labeling solution at 4°C overnight, and then washed three times in 3‰ PBT. DAPI staining was performed as described above.
2.7. RNA Sequencing
Testes from 3-day-old Fd59a2/2 and w1118 flies were dissected in DEPC-treated PBS (10 mM, pH7.4). Total RNA was extracted, and RNA sequencing was performed at Majorbio Bio-Pharm Technology (Shanghai, China) using an Illumina HiSeq 2000 (Illumina, San Diego, CA, USA).
2.8. Bioinformatics Analysis of RNA-seq Data
Transcript abundances in this study were determined using Fragments Per Kilobase per Million mapped reads (FPKM) values. Differentially expressed genes (DEGs) were identified based on a log2 fold-change > 1.5 (or log2 fold-change < −1.5) and a p-adjust < 0.05 with three biological replicates. Gene Ontology (GO) enrichment analysis of DEGs was performed using the GOseq R package.
2.9. Statistical Analysis
The size of apical region from Fd59a2/2 and w1118 testes was quantitated by ImageJ. Three biological replicates and three technical replicates for each biological sample were performed. Statistical significance was analyzed by the student’s t-test (for comparison of two groups) or one-way analysis of variance followed by a least significant difference test (for multiple comparisons among groups). Data were analyzed using GraphPad Prism version 9.0 and presented as mean ± S.E.
3. Results
3.1. Expression Profile of Fd59a in Drosophila
To understand functions of Fd59a, we first determined the expression profile of Fd59a at different developmental stages of Drosophila by qRT-PCR. The results showed that expression of Fd59a mRNA peaked twice during development from embryo to adult, with the first peak around mid-embryonic stage and then a plateau with a relatively high level from late-embryonic stage to 1st instar larval stage, and the second peak just at the metamorphosis period and maintaining at a relative high level till mid pupal stage (Figure 1A), indicating that Fd59a may be involved in Drosophila development.
It has been reported that loss of function in Fd59a affected egg-laying of Drosophila females [8]. Interestingly, microarray data from the Flybase showed that expression level of Fd59a was about 2-fold higher in the testis than in the ovary [19]. We performed qRT-PCR and confirmed that mRNA level of Fd59a was significantly higher in the testis than in the ovary of 3-fay-old w1118 flies (Figure 1B), suggesting that Fd59a may also have a function in the testis of Drosophila.
3.2. Sequence and Phylogenetic Analyses of Fd59a
Fd59a gene is in the 2nd chromosome. The full-length cDNA of Fd59a is 1371 bp long, encoding a protein of 456 amino acid residues, with a calculated molecular weight of 49.1 kDa and pI of 5.19.
Fd59a belongs to the FoxD subfamily. To reveal the evolutionary relationship of Fd59a, Fd59a/FoxD homologous sequences from some insect and vertebrate species were blasted and downloaded from NCBI (Table 2), sequence alignment was performed, and phylogenetic tree was constructed. The results showed that all the select Fd59a/FoxD homologous proteins contain a conserved forkhead box domain (Figure 2A). Meanwhile, Drosophila Fd59a was most close to FoxD3-A of Papilio Xuthus and FoxD3-like of Blattella germanica (Figure 2B) and they contain similar functional domains (Figure 2C). These results suggest that Drosophila Fd59a is evolutionarily close to insect FoxD members.
3.3. Loss of Function in Fd59a Affects Testis Development and Male Fertility
To determine the role of Fd59a in testis development in D. melanogaster, testes from Fd59a1/1 and Fd59a2/2 loss of function mutant flies as well as w1118 flies were dissected. Fd59a1 and Fd59a2 were two mutant types of Fd59a, with Fd59a1 as a hypomorphic mutation and Fd59a2 as a null allele [8]. We found that the apical region of testes from the Fd59a1/1 and Fd59a2/2 mutant flies was swelled (Figure 3A,B), with only a few mature sperm in the seminal vesicle of Fd59a2/2 mutant testis compared to full of mature sperm in the w1118 flies (Figure 3A). As the phenotype of Fa59a2/2 flies was more significant than that of Fa59a1/1 flies, we carried out our following studies in the Fa59a2/2 mutant flies. When Fd59a2/2 males were crossed with w1118 females, the hatching rate of F1 flies was significantly decreased compared to the control (Figure 3C). Together, these results suggest that Fd59a plays a critical role in the development of testis and/or spermatogenesis of D. melanogaster.
3.4. Loss of Function in Fd59a Affects Spermatogenesis
In the apical tip of testis, hub cells are surrounded with GSCs and CySCs to form the stem cell niche, and proliferation and differentiation of testis stem cells are controlled by the niche. Disruption of niche homeostasis can lead to swelling of testis and defect in spermatogenesis [25]. To clarify the role of Fd59a in spermatogenesis, Fas Ⅲ and Vasa antibodies were used to label the hub cells and germ cells, respectively, and αSpectrin antibody was used to label spectrosomes and fusomes for early germ cell development. The results showed that distribution of GSCs and CySCs was scattered in the Fd59a2/2 testis compared to the control testis (Figure 4A, a1-a4, b1-b4, a6, b6), and a strong pattern of fusome and spectrosome formation was displayed in the control testis, while only a few fusomes and spectrosomes were observed in the Fd59a2/2 mutant testis (Figure 4A, a3, a5, b3, b5). These results suggest that Fd59a may play a role in maintaining the homeostasis of testis stem cell niche.
Mammalian FoxD1 is related to apoptosis, as knockdown expression of FoxD1 facilitated apoptosis in HNSCC (head and neck squamous cell carcinoma) cells [26]. To determine whether loss of function in Fd59a can induce apoptosis in the testis, TUNEL assay was performed. TUNEL positive signals were detected in sperm bundles of the Fd59a2/2 mutant flies but not in the w1118 flies (Figure 5A, a1-a4, b1-b4), suggesting that loss of function in Fd59a induced apoptosis of spermatid.
To further confirm the role of Fd59a in testis, expression of Fd59a in GSCs was knocked down by Nos-Gal4 (Figure 4B), and similar phenotypes, such as swelling in the apical region of testis, fewer mature sperm in the seminal vesicle and apoptosis of sperm bundles, were observed in the Nos-Gal4>Fd59a RNAi flies (Figure 4A, c1-c6, d1-d6, and Figure 5A, c1-c4, d1-d4). Meanwhile, knockdown expression of Fd59a in the 4–16 stage of spermatogonia by Bam-Gal4 (Figure 5C) also induced apoptosis of sperm bundles (Figure 5A, e1-e4, f1-f4). Moreover, only a few mature sperms were observed in the seminal vesicles of the Nos-Gal4>Fd59a RNAi and Bam-Gal4>Fd59a RNAi flies, while the control flies were filled with mature sperm (Figure 5B). These combined results suggest that loss of function of Fd59a in the testis resulted in disruption of the stem cell niche during spermatogenesis and increase in apoptosis of sperm bundles, finally leading to fewer mature sperm in the seminal vesicle.
3.5. Fd59a Regulates Gene Expression in the Testis
To better understand how Fd59a plays a role in regulating testis development and spermatogenesis, we carried out RNA-sequencing (RNA-seq) with RNAs isolated from the testes of w1118 (control) and Fd59a2/2 males. Totally 1863 differential expressed genes (DEGs) with at least 1.5-fold change (p-adjust < 0.05) were identified by RNA-seq, with 854 genes upregulated and 1009 genes downregulated in the testis of Fd59a2/2 flies (Figure 6A). This result indicated that Fd59a serves as a transcription factor in the testis.
Gene Ontology (GO) analysis showed that 120 differentially expressed genes (DEGs) are involved in the reproductive process and 475 DEGs participate in the metabolic process (Figure 6B). Among the reproduction related DEGs, many genes have been shown to play a role in gonad development and spermatogenesis, such as Fz2 and Zpg [27,28]. There are also 57 DEGs which are involved in cell death, including Rnrs and Ptp52F [29,30].
To confirm RNA-seq data, totally 20 DEGs associated with reproductive process and cell death were selected for qRT-PCR analysis (Table 3). The results showed that expression profiles of all the select DEGs in the testis detected by qRT-PCR were consistent with those of the RNA-seq data (Figure 6C). Then, 2000 bp promoter sequences upstream the transcriptional start sites of these select genes were downloaded, and the potential Fox binding sites were predicted using JASPAR database (the relative profile score threshold was set to 85%). Except for CG32817 promoter, several Fox binding sites were predicted in the promoter of each select gene, with more than 10 potential Fox binding sites in the promoters of Spd-2, Cal1, Blanks, Ptp52F, Lola and Debcl genes (Table 3). This result further supported that Fd59a is an upstream regulator of these genes. Taken together, our results suggest that Fd59a serves as a transcription factor to regulate expression of genes involved in reproduction, cell growth and death in the testis.
4. Discussion
Drosophila testis is an ideal system to study spermatogenesis. In this study, we found that FoxD transcription factor Fd59a contributes to spermatogenesis of Drosophila. So far, little is known about the functions of Drosophila Fd59a/FoxD and other insect FoxD members. It was reported that Fd59a was expressed in octopaminergic neurons and regulated egg-laying behavior of females [8]. Drosophila CHES-1-like/FoxN inhibited germline stem cell differentiation by upregulating Dpp expression and ectopic expression of CHES-1-like reduced male fertility significantly [31]. In B. mori, Fox family genes were expressed in the testis, with BmFoxL2-2 and BmFoxD at a higher level than other BmFox genes [18]. However, the function of BmFoxD in the testis is still unknown.
In this study, we showed that spermatogenesis was disrupted, and apoptosis of sperm bundles was induced in Fd59a mutant and RNAi flies. Spermatogenesis is a complex process regulated by multiple signaling pathways and many different genes. Over-activation of the JAK-STAT signaling pathway led to overgrowth of testis and disrupted structure of testis stem cell niche in Drosophila [25,32]. Over-activation of the JNK or loss of the Notch signaling caused cell death in the testis of Drosophila [33,34]. In mammals, deletion of Stat3 in Foxd1 cell lineage protected mice from kidney fibrosis [35]. Hypoxia-inducible factors (HIFs) regulated genes related to oxygen homeostasis, and lack of Hif-p4h-2 (HIF prolyl-4-hydroxylases) in FoxD1 lineage led to dysregulation of genes involved in the Notch signaling pathway [36]. These results suggest that there is a genetic interaction between mammalian FoxD subfamily members and the JAK-STAT and Notch pathways. However, mRNA levels of genes related to the above three signaling pathways did not change significantly in the Fd59a2/2 testis (RNA-seq data), indicating that Fd59a is not involved in the JAK-STAT, JNK or Notch signaling pathway. Therefore, insect FoxD members may function differently from mammalian FoxD subfamily members.
Results from RNA-seq and qRT-PCR showed that many genes related to reproduction and cell death were differentially expressed in the testis of Fd59a2/2 flies. Among the reproduction related DEGs, Fz2 and Zpg have been reported to regulate germ stem cell development in Drosophila testis [27,28], while Blanks functioned in sperm individualization [37]. Among cell death related DEGs, Ptp52F enhanced autophagy and apoptosis in Drosophila midgut [30]. In addition, serval potential Fox binding sites were predicted in the promoters of select DEGs. Thus, Fd59a acts as a transcription factor to regulate expression of genes involved in spermatogenesis and in maintaining survival of sperm cells.
It was reported that octopamine was essential for increasing GSCs in mating Drosophila females [38], and β-adrenergic-like octopamine receptor (OctβR) was strongly expressed in adult testis [39]. In Fd59a2/2 adult testis, Octβ2R expression was down-regulated, thus, it is possible that Fd59a regulates spermatogenesis partly through regulating expression of Octβ2R, and Fd59a may be a key factor linking the nervous system to male reproduction system.
Author Contributions
Conceptualization, X.Q.Y., L.W. and Q.H.; methodology, T.T., L.W. and Q.H.; software, M.P., Y.D. and Y.L.; validation, Y.X., Y.D. and L.W.; formal analysis, X.Q.Y. and Q.H.; investigation, T.T., M.P., Y.X., Y.D. and Q.H.; resources, Y.X. and Q.H.; data curation, M.P., Y.L. and Q.H.; writing – original draft preparation, T.T. and Q.H.; writing – review and editing, X.Q.Y., L.W. and Q.H.; visualization, T.T. and Y.L.; supervision, L.W. and Q.H.; project administration, X.Q.Y. and Q.H.; funding acquisition, Q.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (No. 32300389), GuangDong Basic and Applied Basic Research Foundation (No. 2022A1515111055) and China Postdoctoral Science Foundation (No. 2022M721219).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank Professor Yufeng Wang at the School of Life Sciences, Central China Normal University, Wuhan, China, for providing Bam-Gal4 flies.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Akhlaghipour, I.; Fanoodi, A.; Zangouei, A. S.; Taghehchian, N.; Khalili-Tanha, G.; Moghbeli, M. MicroRNAs as the Critical Regulators of Forkhead Box Protein Family in Pancreatic, Thyroid, and Liver Cancers. Biochemical genetics 2023, 61, 1645–1674. [Google Scholar] [CrossRef]
- Kaestner, K.H.; Knochel, W.; Martinez, D.E. Unified nomenclature for the winged helix/forkhead transcription factors. Genes & development 2000, 14, 142–146. [Google Scholar] [CrossRef]
- Quintero-Ronderos, P.; Laissue, P. The multisystemic functions of FOXD1 in development and disease. Journal of molecular medicine (Berlin, Germany) 2018, 96, 725–739. [Google Scholar] [CrossRef]
- Cai, K.; Chen, S.; Zhu, C.; Li, L.; Yu, C.; He, Z.; Sun, C. FOXD1 facilitates pancreatic cancer cell proliferation, invasion, and metastasis by regulating GLUT1-mediated aerobic glycolysis. Cell Death & Disease 2022, 13, 765. [Google Scholar] [CrossRef]
- Jiang, W.; Li, Y.; Li, R.; Chen, W.; Song, M.; Zhang, Q.; Chen, S. The Prognostic Significance of FOXD1 Expression in Head and Neck Squamous Cell Carcinoma. Journal of Personalized Medicine 2023, 13, 530. [Google Scholar] [CrossRef]
- Qiu, S.J.; Li, D.; Shen, Z.S.; Li, Q.; Shen, Y.; Deng, H.X.; Wu, Y.D.; Zhou, C.C. Diagnostic and prognostic value of FOXD1 expression in head and neck squamous cell carcinoma. Journal of Cancer 2021, 12, 693–702. [Google Scholar] [CrossRef]
- Scimone, M.L.; Lapan, S.W.; Reddien, P.W. A forkhead transcription factor is wound-induced at the planarian midline and required for anterior pole regeneration. PLoS genetics 2014, 10, e1003999. [Google Scholar] [CrossRef] [PubMed]
- Lacin, H.; Rusch, J.; Yeh, R. T.; Fujioka, M.; Wilson, B. A.; Zhu, Y.; Robie, A. A.; Mistry, H.; Wang, T.; Jaynes, J. B.; Skeath, J. B. Genome-wide identification of Drosophila Hb9 targets reveals a pivotal role in directing the transcriptome within eight neuronal lineages, including activation of nitric oxide synthase and Fd59a/Fox-D. Developmental biology 2014, 388, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Siddall, N. A.; Hime, G. R. A Drosophila toolkit for defining gene function in spermatogenesis. Reproduction (Cambridge, England) 2017, 153, R121–R132. [Google Scholar] [CrossRef] [PubMed]
- Fabian, L.; Brill, J. A. Drosophila spermiogenesis: Big things come from little packages. Spermatogenesis 2012, 2, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Greenspan, L.J.; de Cuevas, M.; Matunis, E. Genetics of gonadal stem cell renewal. Annual review of cell and developmental biology 2015, 31, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Drummond-Barbosa, D. Local and Physiological Control of Germline Stem Cell Lineages in Drosophila melanogaster. Genetics 2019, 213, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, K.G.; Carthew, R.W. Out of the testis: biological impacts of new genes. Genes & development 2017, 31, 1825–1826. [Google Scholar] [CrossRef]
- Fang, Y.; Zhang, F.; Zhan, Y.; Lu, M.; Xu, D.; Wang, J.; Li, Q.; Zhao, L.; Su, Y. RpS3 Is Required for Spermatogenesis of Drosophila melanogaster. Cells 2023, 12, 573. [Google Scholar] [CrossRef]
- Chen, M. Y.; Duan, X.; Wang, Q.; Ran, M. J.; Ai, H.; Zheng, Y.; Wang, Y. F. Cytochrome c1-like is required for mitochondrial morphogenesis and individualization during spermatogenesis in Drosophila melanogaster. The Journal of experimental biology 2023, 226, jeb245277. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chen, D.; Zheng, S.; Yi, M.; Liu, Z.; Liu, Y.; Yang, D.; Liu, Y.; Tang, L.; Zhu, C.; Huang, Y. BmHen1 is essential for eupyrene sperm development in Bombyx mori but PIWI proteins are not. Insect biochemistry and molecular biology 2022, 151, 103874. [Google Scholar] [CrossRef]
- Hu, Q.; Zhu, Z.; Zhao, D.; Zeng, B.; Zheng, S.; Song, Q.; Deng, H.; Feng, Q. Bombyx mori transcription factors FoxA and SAGE divergently regulate the expression of wing cuticle protein gene 4 during metamorphosis. The Journal of biological chemistry 2019, 294, 632–643. [Google Scholar] [CrossRef]
- Song, J.; Li, Z.; Tong, X.; Chen, C.; Chen, M.; Meng, G.; Chen, P.; Li, C.; Xin, Y.; Gai, T.; Dai, F.; Lu, C. Genome-wide identification and characterization of Fox genes in the silkworm, Bombyx mori. Functional & integrative genomics 2015, 15, 511–522. [Google Scholar] [CrossRef]
- Chintapalli, V.R.; Wang, J.; Dow, J.A. Using FlyAtlas to identify better Drosophila melanogaster models of human disease. Nature genetics 2007, 39, 715–720. [Google Scholar] [CrossRef]
- Hu, Q.; Xiao, Y.; Wei, R.; Tang, T.; Wen, L.; Lu, Y.; Yu, X. Q. Identification and functional analysis of CG3526 in spermatogenesis of Drosophila melanogaster. Insect science 2024, 31, 79–90. [Google Scholar] [CrossRef]
- Biwot, J.C.; Zhang, H.B.; Chen, M.Y.; Wang, Y.F. A new function of immunity-related gene Zn72D in male fertility of Drosophila melanogaster. Archives of Insect Biochemistry and Physiology 2019, 102, e21612. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Xiao, Y.; Huang, B.; Ran, M. J.; Duan, X.; Wang, Y. F.; Lu, Y.; Yu, X. Q. A dual role of lola in Drosophila ovary development: regulating stem cell niche establishment and repressing apoptosis. Cell death & disease 2022, 13, 756. [Google Scholar]
- Loza-Coll, M.A.; Petrossian, C.C.; Boyle, M.L.; Jones, D.L. Heterochromatin Protein 1 (HP1) inhibits stem cell proliferation induced by ectopic activation of the Jak/STAT pathway in the Drosophila testis. Experimental cell research 2019, 377, 1–9. [Google Scholar] [CrossRef]
- Li, J.; Yan, T.; Wu, X.; Ke, X.; Li, X.; Zhu, Y.; Yang, J.; Li, Z. Aberrant overexpression of transcription factor Forkhead box D1 predicts poor prognosis and promotes cancer progression in HNSCC. BMC cancer 2021, 21, 1205. [Google Scholar] [CrossRef]
- Smendziuk, C.M.; Messenberg, A.; Vogl, A.W.; Tanentzapf, G. Bi-directional gap junction-mediated soma-germline communication is essential for spermatogenesis. Development (Cambridge, England) 2015, 142, 2598–2609. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Gao, Y.; Song, X.; Ma, X.; Zhu, X.; Mao, Y.; Yang, Z.; Ni, J.; Li, H.; Malanowski, K. E.; Anoja, P.; Park, J.; Haug, J.; Xie, T. Wnt signaling-mediated redox regulation maintains the germ line stem cell differentiation niche. eLife 2015, 4, e08174. [Google Scholar] [CrossRef] [PubMed]
- Akdemir, F.; Christich, A.; Sogame, N.; Chapo, J.; Abrams, J.M. p53 directs focused genomic responses in Drosophila. Oncogene 2007, 26, 5184–5193. [Google Scholar] [CrossRef]
- Santhanam, A.; Peng, W.H.; Yu, Y.T.; Sang, T.K.; Chen, G.C.; Meng, T.C. Ecdysone-induced receptor tyrosine phosphatase PTP52F regulates Drosophila midgut histolysis by enhancement of autophagy and apoptosis. Molecular and cellular biology 2014, 34, 1594–1606. [Google Scholar] [CrossRef]
- Yu, J.; Liu, Y.; Lan, X.; Wu, H.; Wen, Y.; Zhou, Z.; Hu, Z.; Sha, J.; Guo, X.; Tong, C. CHES-1-like, the ortholog of a non-obstructive azoospermia-associated gene, blocks germline stem cell differentiation by upregulating Dpp expression in Drosophila testis. Oncotarget 2016, 7, 42303–42313. [Google Scholar] [CrossRef] [PubMed]
- Kiger, A.A.; Jones, D.L.; Schulz, C.; Rogers, M.B.; Fuller, M.T. Stem cell self-renewal specified by JAK-STAT activation in response to a support cell cue. Science (New York, N.Y.) 2001, 294, 2542–2545. [Google Scholar] [CrossRef] [PubMed]
- Herrera, S.C.; Bach, E.A. The Emerging Roles of JNK Signaling in Drosophila Stem Cell Homeostasis. International journal of molecular sciences 2021, 22, 5519. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.L.; Qian, Y.; Schulz, C. Notch and Delta are required for survival of the germline stem cell lineage in testes of Drosophila melanogaster. PloS one 2019, 14, e0222471. [Google Scholar] [CrossRef] [PubMed]
- Ajay, A. K.; Zhao, L.; Vig, S.; Fujiwara, M.; Thakurela, S.; Jadhav, S.; Cho, A.; Chiu, I. J.; Ding, Y.; Ramachandran, K.; Mithal, A.; Bhatt, A.; Chaluvadi, P.; Gupta, M. K.; Shah, S. I.; Sabbisetti, V. S.; Waaga-Gasser, A. M.; Frank, D. A.; Murugaiyan, G.; Bonventre, J. V.; Hsiao, L. L. Deletion of STAT3 from Foxd1 cell population protects mice from kidney fibrosis by inhibiting pericytes trans-differentiation and migration. Cell reports 2022, 38, 110473. [Google Scholar] [CrossRef] [PubMed]
- Rosendahl, A. H.; Monnius, M.; Laitala, A.; Railo, A.; Miinalainen, I.; Heljasvaara, R.; Mäki, J. M.; Myllyharju, J. Deletion of hypoxia-inducible factor prolyl 4-hydroxylase 2 in FoxD1-lineage mesenchymal cells leads to congenital truncal alopecia. The Journal of biological chemistry 2022, 298, 101787. [Google Scholar] [CrossRef]
- Sanders, C.; Smith, D.P. LUMP is a putative double-stranded RNA binding protein required for male fertility in Drosophila melanogaster. PloS one 2011, 6, e24151. [Google Scholar] [CrossRef]
- Yoshinari, Y.; Ameku, T.; Kondo, S.; Tanimoto, H.; Kuraishi, T.; Shimada-Niwa, Y.; Niwa, R. Neuronal octopamine signaling regulates mating-induced germline stem cell increase in female Drosophila melanogaster. eLife 2020, 9, e57101. [Google Scholar] [CrossRef]
- Ohhara, Y.; Kayashima, Y.; Hayashi, Y.; Kobayashi, S.; Yamakawa-Kobayashi, K. Expression of β-adrenergic-like octopamine receptors during Drosophila development. Zoological science 2012, 29, 83–89. [Google Scholar] [CrossRef]
Figure 1.
Expression of Fd59a at different developmental stages and in adult ovary and testis. (A) Expression of Fd59a at different developmental stages. Drosophila embryos at 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 and 24 h after egg-laying, first (L1), second (L2), third (L3) instar larvae, early, mid and late pupae, 1, 3 and 5-day-old adult flies were collected to prepare total RNAs for analysis of transcriptional expression of Fd59a by qRT-PCR. (B) Expression of Fd59a in the testis and ovary of 3-day-old adult flies. Data were presented as means ± S.E., and significant difference was determined by the student’s t-test and indicated by asterisks: ** p < 0.01.
Figure 1.
Expression of Fd59a at different developmental stages and in adult ovary and testis. (A) Expression of Fd59a at different developmental stages. Drosophila embryos at 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 and 24 h after egg-laying, first (L1), second (L2), third (L3) instar larvae, early, mid and late pupae, 1, 3 and 5-day-old adult flies were collected to prepare total RNAs for analysis of transcriptional expression of Fd59a by qRT-PCR. (B) Expression of Fd59a in the testis and ovary of 3-day-old adult flies. Data were presented as means ± S.E., and significant difference was determined by the student’s t-test and indicated by asterisks: ** p < 0.01.
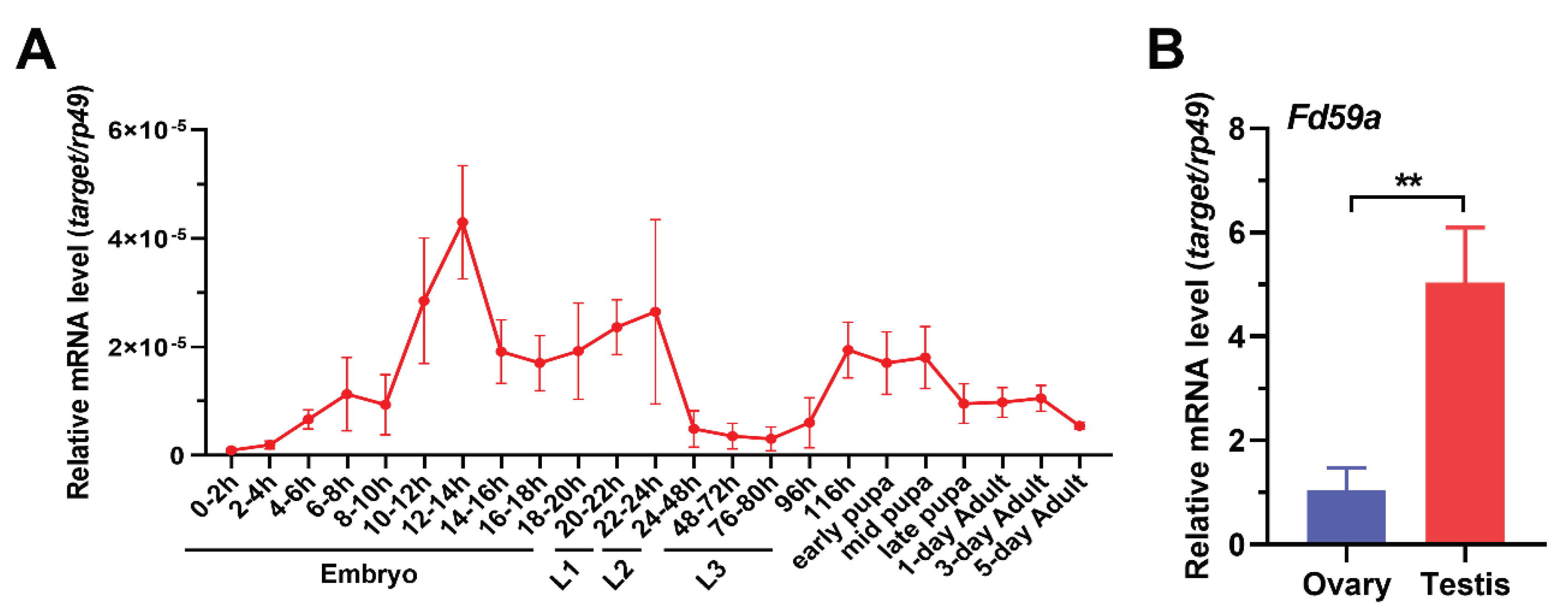
Figure 2.
Sequence analysis of D. melanogaster Fd59a. Sequence alignment (A), Maximum Likelihood phylogenetic tree (B) and functional domains (C) of Fd59a with its homologous proteins from some insect and vertebrate species. For detailed information of the sequences, see Table 2.
Figure 2.
Sequence analysis of D. melanogaster Fd59a. Sequence alignment (A), Maximum Likelihood phylogenetic tree (B) and functional domains (C) of Fd59a with its homologous proteins from some insect and vertebrate species. For detailed information of the sequences, see Table 2.

Figure 3.
Mutation in Fd59a affects testis development and reduces the number of mature sperm in the seminal vesicle. (A) Overall appearance of testes and seminal vesicles from w1118, Fd59a1/1 and Fd59a2/2 flies, nuclei were stained with DAPI. Scale bar in a1, b1, c1 is 100 µm, in a2, b2, c2 is 50 µm, and in a3, b3, c3 is 25 µm. The arrowhead in a3 represents mature sperm. Numbers below the images indicate the pairs of testes with similar phenotypes in the images. (B) Quantitative measurements of the apical region of testis from Fd59a2/2 and w1118 flies. (C) Male fertility test of w1118 and Fd59a2/2 flies. Data were presented as means ± S.E., and significant difference was determined by the student’s t-test and indicated by asterisks: ** p < 0.01, *** p < 0.001.
Figure 3.
Mutation in Fd59a affects testis development and reduces the number of mature sperm in the seminal vesicle. (A) Overall appearance of testes and seminal vesicles from w1118, Fd59a1/1 and Fd59a2/2 flies, nuclei were stained with DAPI. Scale bar in a1, b1, c1 is 100 µm, in a2, b2, c2 is 50 µm, and in a3, b3, c3 is 25 µm. The arrowhead in a3 represents mature sperm. Numbers below the images indicate the pairs of testes with similar phenotypes in the images. (B) Quantitative measurements of the apical region of testis from Fd59a2/2 and w1118 flies. (C) Male fertility test of w1118 and Fd59a2/2 flies. Data were presented as means ± S.E., and significant difference was determined by the student’s t-test and indicated by asterisks: ** p < 0.01, *** p < 0.001.
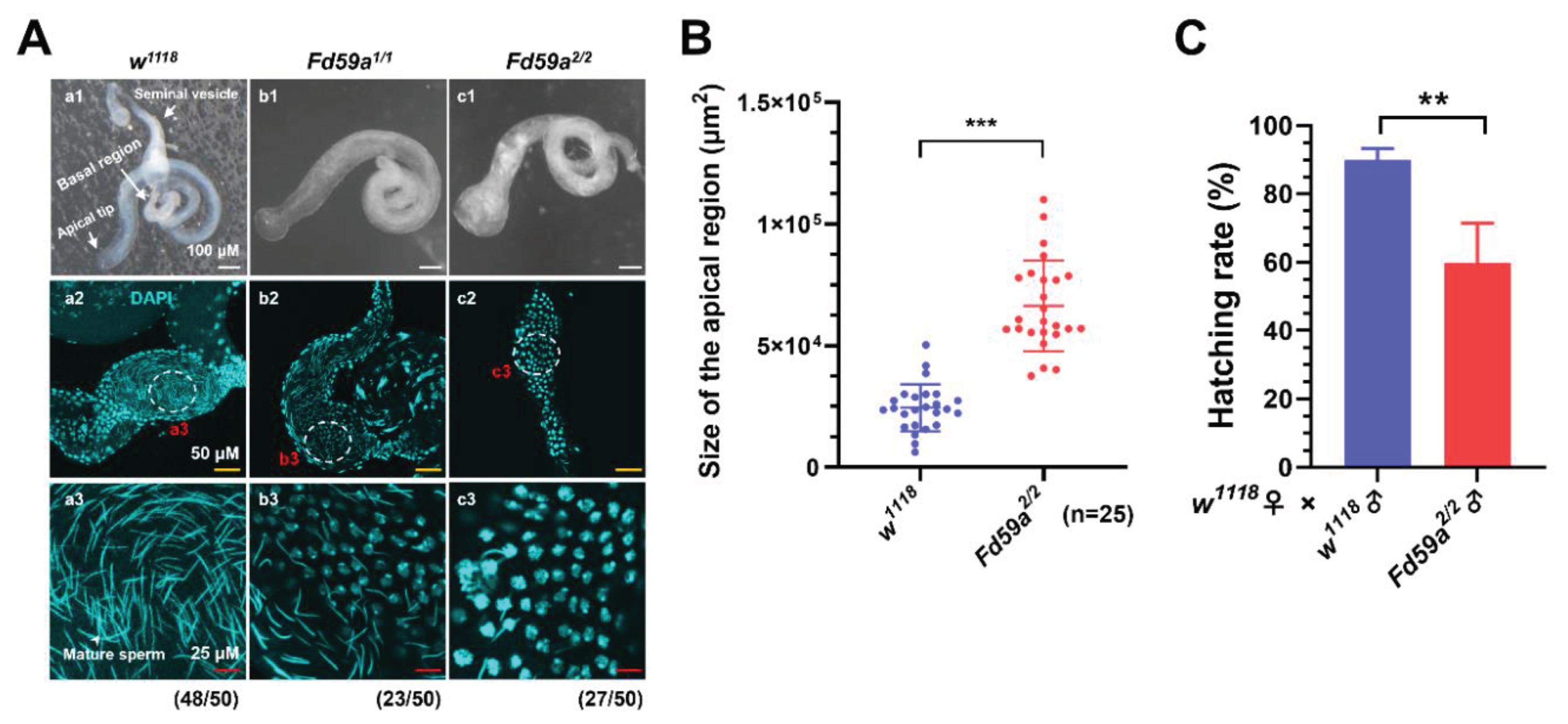
Figure 4.
Loss of function of Fd59a in the testis disrupts the homeostasis of testis stem cell niche. (A) Immunostaining of testis. Testes from the w1118, Fd59a2/2, Nos-Gal4>GFP RNAi and Nos-Gal4>Fd59a RNAi flies were labeled with anti-Vasa (green), anti-FasIII (red) and anti-αSpectrin (red) antibodies, and nuclei were stained with DAPI (blue). (a1-a4, b1-b4, c1-c4, d1-d4) The apical region of testis, (a5, b5, c5, d5) enlarged part of the apical region showing fusomes and spetrosomes, (a6, b6, c6, d6) enlarged part of the apical tip showing the stem cell niche. In the testis of Fd59a2/2 and Nos-Gal4>Fd59a RNAi flies, the distribution of germ cells labeled with anti-Vasa (green) antibody was disrupted, and only a few fusomes and spectrosomes labeled with anti-αSpectrin (red) antibody were observed. Scale bar in a1-a4, b1-b4, c1-c4 and d1-d4 is 50 µm, in a5, b5, c5 and d5 is 25 µm, and in a6, b6, c6 and d6 is 10 µm. The arrow in a3 represents hub cell, and arrowheads in a5 represent fusomes and spectrosomes. (B) Expression of Fd59a in the testis of Nos-Gal4 RNAi flies. Data were presented as means ± S.E., and significant difference was determined by the student’s t-test and indicated by asterisks: ** p < 0.01.
Figure 4.
Loss of function of Fd59a in the testis disrupts the homeostasis of testis stem cell niche. (A) Immunostaining of testis. Testes from the w1118, Fd59a2/2, Nos-Gal4>GFP RNAi and Nos-Gal4>Fd59a RNAi flies were labeled with anti-Vasa (green), anti-FasIII (red) and anti-αSpectrin (red) antibodies, and nuclei were stained with DAPI (blue). (a1-a4, b1-b4, c1-c4, d1-d4) The apical region of testis, (a5, b5, c5, d5) enlarged part of the apical region showing fusomes and spetrosomes, (a6, b6, c6, d6) enlarged part of the apical tip showing the stem cell niche. In the testis of Fd59a2/2 and Nos-Gal4>Fd59a RNAi flies, the distribution of germ cells labeled with anti-Vasa (green) antibody was disrupted, and only a few fusomes and spectrosomes labeled with anti-αSpectrin (red) antibody were observed. Scale bar in a1-a4, b1-b4, c1-c4 and d1-d4 is 50 µm, in a5, b5, c5 and d5 is 25 µm, and in a6, b6, c6 and d6 is 10 µm. The arrow in a3 represents hub cell, and arrowheads in a5 represent fusomes and spectrosomes. (B) Expression of Fd59a in the testis of Nos-Gal4 RNAi flies. Data were presented as means ± S.E., and significant difference was determined by the student’s t-test and indicated by asterisks: ** p < 0.01.
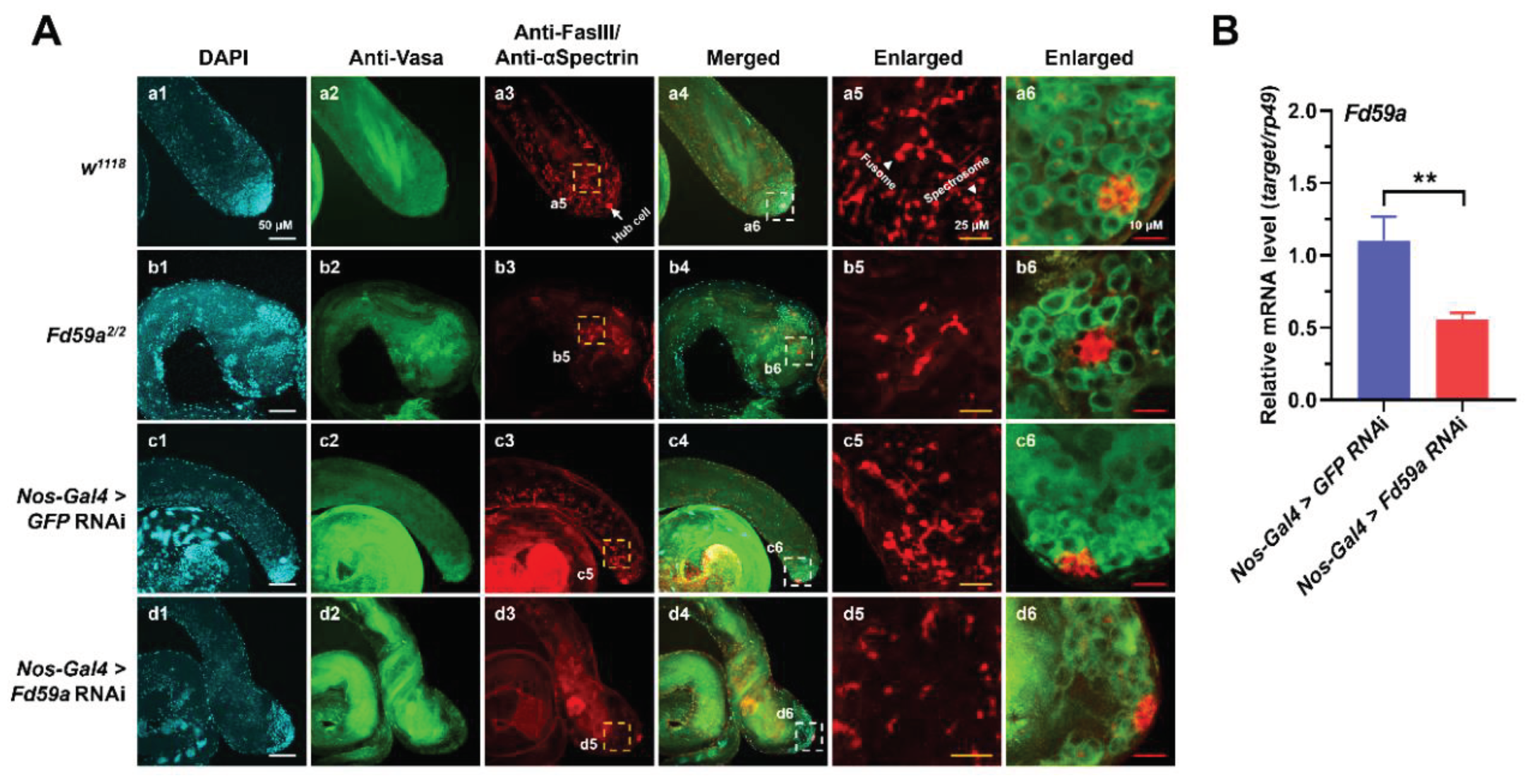
Figure 5.
Loss of function of Fd59a in the testis induces apoptosis of sperm bundles. (A) Detection of apoptotic cells in the testis. Testes from the w1118, Fd59a2/2, Nos-Gal4>GFP RNAi, Nos-Gal4>Fd59a RNAi, Bam-Gal4>GFP RNAi and Bam-Gal4>Fd59a RNAi flies were stained with TUNEL assay for apoptotic cells, and nuclei were stained with DAPI (blue). (a1-a3, b1-b3, c1-c3, d1-d3, e1-e3, f1-f3) The basal region of testis, (a4, b4, c4, d4, e4, f4) enlarged part of the basal region showing the sperm bundles. In the testis of Fd59a2/2, Nos-Gal4>Fd59a RNAi and Bam-Gal4>Fd59a RNAi flies, many TUNEL signals were detected in the sperm bundles, and only a few TUNEL signals were detected in the basal region of w1118, Nos-Gal4>GFP RNAi and Bam-Gal4>GFP RNAi flies, but not in the sperm bundles. Scale bar in a1-a3, b1-b3, c1-c3, d1-d3, e1-e3 and f1-f3 is 50 µm, in a4, b4, e4 and f4 is 10 µm, and in c4, d4 is 20 µm. (B) DAPI staining of seminal vesicle. Seminal vesicles from Nos-Gal4>GFP RNAi, Nos-Gal4>Fd59a RNAi, Bam-Gal4>GFP RNAi and Bam-Gal4>Fd59a RNAi flies were stained with DAPI. Scale bar in a1, b1, c1, d1 is 50 µm, in a2, b2, c2, d2 is 10 µm. Numbers below the images indicate the pairs of testes with similar phenotypes in the images. (C) Expression of Fd59a in the testis of Bam-Gal4 RNAi flies. Data were presented as means ± S.E., and significant difference was determined by the student’s t-test and indicated by asterisks: * p < 0.05.
Figure 5.
Loss of function of Fd59a in the testis induces apoptosis of sperm bundles. (A) Detection of apoptotic cells in the testis. Testes from the w1118, Fd59a2/2, Nos-Gal4>GFP RNAi, Nos-Gal4>Fd59a RNAi, Bam-Gal4>GFP RNAi and Bam-Gal4>Fd59a RNAi flies were stained with TUNEL assay for apoptotic cells, and nuclei were stained with DAPI (blue). (a1-a3, b1-b3, c1-c3, d1-d3, e1-e3, f1-f3) The basal region of testis, (a4, b4, c4, d4, e4, f4) enlarged part of the basal region showing the sperm bundles. In the testis of Fd59a2/2, Nos-Gal4>Fd59a RNAi and Bam-Gal4>Fd59a RNAi flies, many TUNEL signals were detected in the sperm bundles, and only a few TUNEL signals were detected in the basal region of w1118, Nos-Gal4>GFP RNAi and Bam-Gal4>GFP RNAi flies, but not in the sperm bundles. Scale bar in a1-a3, b1-b3, c1-c3, d1-d3, e1-e3 and f1-f3 is 50 µm, in a4, b4, e4 and f4 is 10 µm, and in c4, d4 is 20 µm. (B) DAPI staining of seminal vesicle. Seminal vesicles from Nos-Gal4>GFP RNAi, Nos-Gal4>Fd59a RNAi, Bam-Gal4>GFP RNAi and Bam-Gal4>Fd59a RNAi flies were stained with DAPI. Scale bar in a1, b1, c1, d1 is 50 µm, in a2, b2, c2, d2 is 10 µm. Numbers below the images indicate the pairs of testes with similar phenotypes in the images. (C) Expression of Fd59a in the testis of Bam-Gal4 RNAi flies. Data were presented as means ± S.E., and significant difference was determined by the student’s t-test and indicated by asterisks: * p < 0.05.
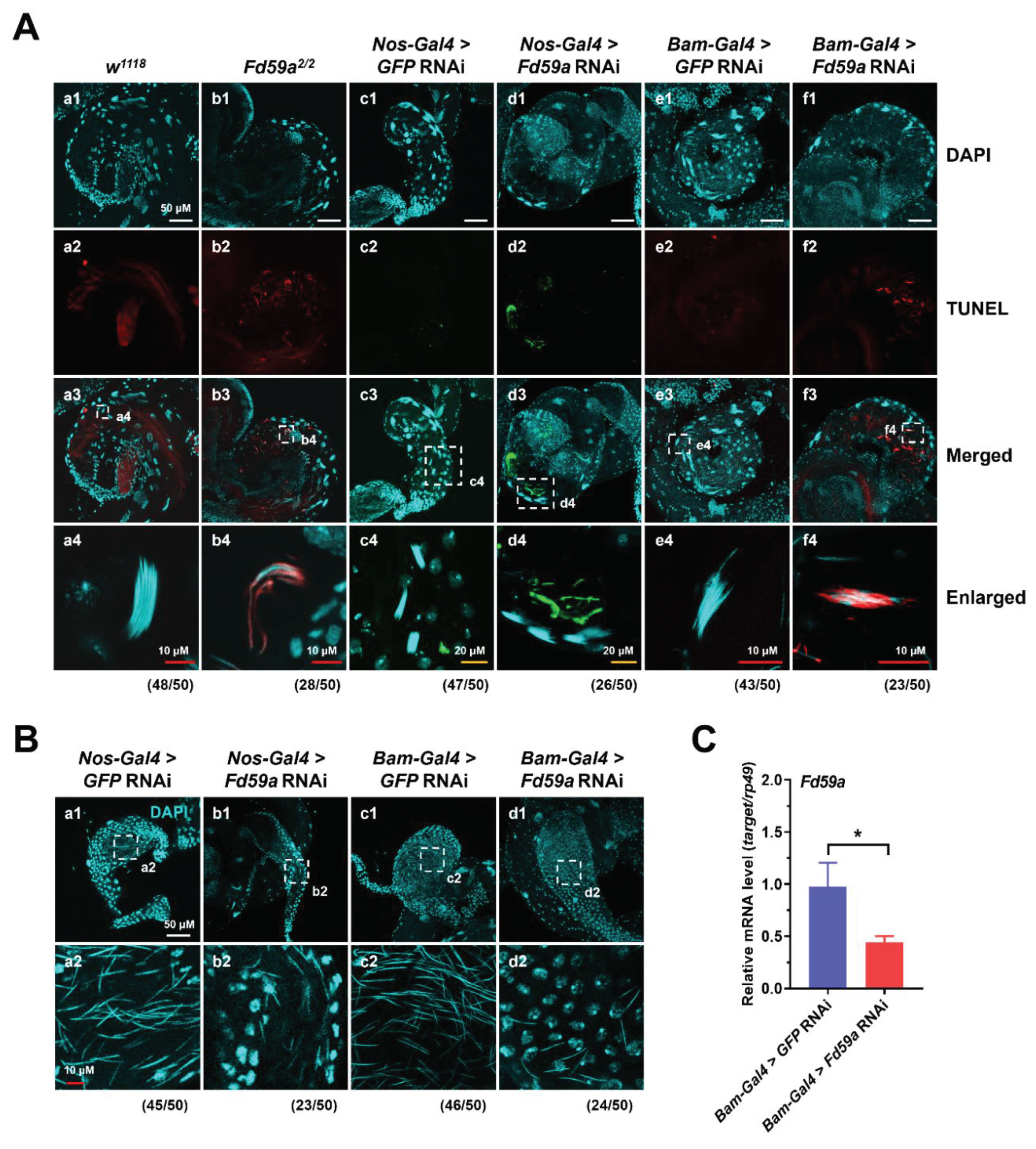
Figure 6.
RNA-seq analysis of RNAs from the testes of Fd59a2/2 and w1118 flies. (A) Heatmap of differentially expressed genes (DEGs) in the testis of Fd59a2/2 flies relative to w1118 flies. (B) GO analysis of DEGs between Fd59a2/2 and w1118 fly testes. (C) qRT-PCR validation of select DEGs from RNA sequencing data. Data were presented as means ± S.E., and significant difference was determined by the student’s t-test and indicated by asterisks: * p < 0.05, ** p < 0.01 and *** p< 0.001.
Figure 6.
RNA-seq analysis of RNAs from the testes of Fd59a2/2 and w1118 flies. (A) Heatmap of differentially expressed genes (DEGs) in the testis of Fd59a2/2 flies relative to w1118 flies. (B) GO analysis of DEGs between Fd59a2/2 and w1118 fly testes. (C) qRT-PCR validation of select DEGs from RNA sequencing data. Data were presented as means ± S.E., and significant difference was determined by the student’s t-test and indicated by asterisks: * p < 0.05, ** p < 0.01 and *** p< 0.001.
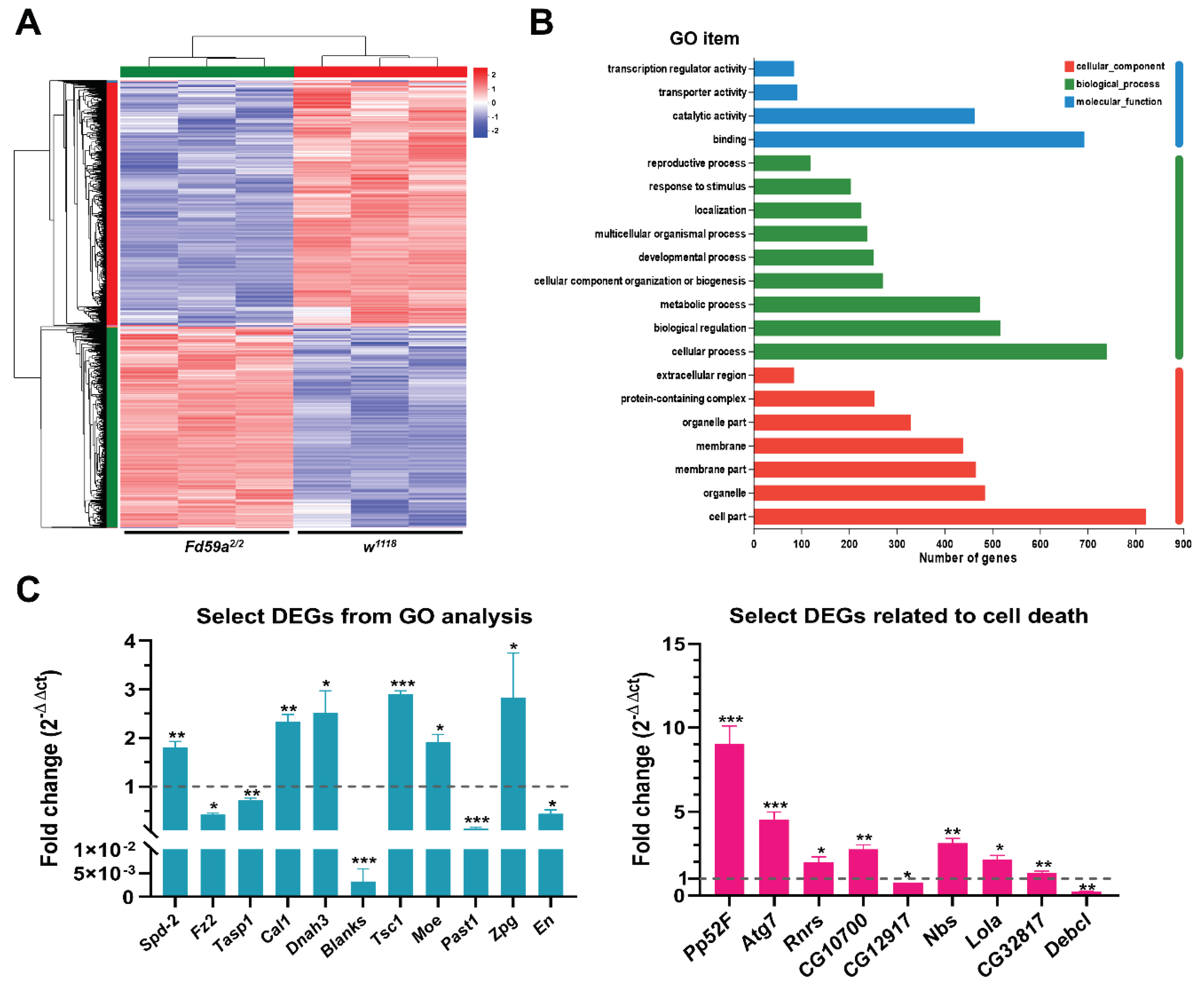

Table 1.
Primers used in this study.
| Name | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| Fd59a-qRT | CAGGGAAGTCAGTCGGGGGA | GTCGCCACATCGAAGGCGTA |
| Rp49-qRT | GCCCAAGGGTATCGACAACA | ACCTCCAGCTCGCGCACGTT |
| Spd-2-qRT | GTGACCCACACGACCCTCTG | GCCGAATGACCAGCCGTTTG |
| Fz2-qRT | TCGCGAGTCACAATTGCACC | GGACGCCACTCTACGGTGTT |
| Tasp1-qRT | CGGCATGCGAGTCTGTTCGG | ACACAAGGCAGCGCAAGTCTA |
| Debcl-qRT | ATCGACAACGGCGGATGGTT | ACGCGATCCCAAGCGAATCT |
| Ptp52F-qRT | TGTCCGACGATCTTTGCGCT | GGCGTAGGGGGAAAGTGGAC |
| Atg7-qRT | TACAACTGCTGGCCGATGAGG | GCACGGAAAGGCGAACCAAT |
| RnrS-qRT | GGACCGTTTGCTCGTGGAGT | GAAATCCGCGTCCAGGGTGA |
| CG10700-qRT | TGTGGAGGCTACGGCCAATC | TCACCACGGCTGTTTCCCAA |
| CG12917-qRT | CAGGGGCTTCCTTCAGTCGG | AAATAGCCAGACACGGGGGC |
| Nbs-qRT | ATTCCCAAAAGCCGCGCAAG | TGGGTCACCTGCCAAATGCT |
| Lola-qRT | CTGCTGAGATATGCGAGCCAGA | GTTCACAATGGCCTCCGCCT |
| Cal1-qRT | GGTGGTGGACGAGGAAACACT | TCCACAGCCTCCTTTGCCAC |
| Dnah3-qRT | AGAGCTGGCAAGAGCGGAAA | ACATTGCGAGACGTGGCACC |
| Blanks-qRT | ACGGGCCAGGAAAGAGCTTG | ACGGCTTCTTTGGCTCGACA |
| En-qRT | CCAACGACGAGAAGCGTCCA | CTCCGCTCGGTCAGATAGCG |
| Tsc1-qRT | GGTTGGCATGACTGGCTCCT | CACGTCCCGGCTGCTTGATA |
| CG32817-qRT | AATCAAGTGTCTAACCCTGAACTGG | GTTGCGCCATCGAAAAGCAT |
| Moe-qRT | GCCTGCGAGAGGTTTGGTTCTT | TCACGTCCTGGTTCATCACCTT |
| Past1-qRT | ACACCCGATCACACAGCCTC | CGCCTGCACTGTGTGGCTAA |
| Zpg-qRT | GGGGCCTATGTGAGCGACAA | CCGCCCTCCCAAATCTTCCA |
Table 2.
Fd59a homologous protein sequences used in the phylogenetic tree.
| Proteins | Species | Accession Number |
|---|---|---|
| Forkhead box protein D5 | Bombyx mori | XP_004922516.1 |
| uncharacterized protein LOC5564025 | Aedes aegypti | XP_001648348.3 |
| Forkhead box protein D3 | Manduca sexta | XP_030032680.1 |
| Forkhead box protein D3-B | Plutella xylostella | XP_037961710.1 |
| Forkhead box protein D3-like | Spodoptera frugiperda | XP_035435321.1 |
| Forkhead box protein D5-like | Spodoptera litura | XP_022815528.1 |
| Forkhead box protein D3-like | Nilaparvata lugens | XP_039277739.1 |
| Forkhead box protein D3-like | Ceratosolen solmsi marchali | XP_011504225.1 |
| Fork head domain-containing protein FD3 | Papilio xuthus | KPJ03207.1 |
| Forkhead box protein D3-like | Bemisia tabac | XP_018914635.1 |
| Forkhead box protein D3-A | Blattella germanica | PSN41724.1 |
| Forkhead box protein D5 | Helicoverpa armigera | XP_021187147.2 |
| Forkhead box protein D3 | Danio rerio | NP_571365.2 |
| Forkhead box protein unc-130 | Caenorhabditis elegans | NP_496411.1 |
| Forkhead box protein D4 | Homo sapiens | NP_997188.2 |
| Forkhead box protein D3 | Mus musculus | NP_034555.3 |
| Forkhead box protein D5-A | Xenopus laevis | NP_001081998.1 |
| Forkhead box protein D3 isoform X1 | Manis javanica | XP_036880296.1 |
| Forkhead box protein D3 | Caretta caretta | XP_048718258.1 |
| Forkhead box protein B1-like | Octopus sinensis | XP_029653697.1 |
| Forkhead box protein D3-like | Branchiostoma floridae | XP_035698942.1 |
| Forkhead box protein D3 | Rattus norvegicus | NP_542952.1 |
Table 3.
Differentially expressed genes selected for qRT-PCR validation and the number of predicted Fox binding sites in the 2 kb promoter regions.
Table 3.
Differentially expressed genes selected for qRT-PCR validation and the number of predicted Fox binding sites in the 2 kb promoter regions.
| Gene Symbol | Log2 Fold Difference | Relative Expression | Biological Functions | Forkhead Binding Sites |
|---|---|---|---|---|
| GO analysis related genes | ||||
| Spd-2 | 1.08 | Up-regulated | Involved in sperm aster formation | 13 |
| Fz2 | -1.66 | Down-regulated | Germ-line stem cell niche homeostasis | 6 |
| Tasp1 | -1.35 | Down-regulated | Involved in spermatogenesis | 7 |
| Cal1 | 0.61 | Up-regulated | Female meiosis chromosome segregation | 10 |
| Dnah3 | 0.70 | Up-regulated | Involved in sperm competition | 6 |
| Blanks | -0.72 | Down-regulated | Involved in sperm individualization | 11 |
| Tsc1 | 0.76 | Up-regulated | Negative regulation of developmental growth | 4 |
| Moe | 0.67 | Up-regulated | Oocyte anterior/posterior axis specification | 10 |
| Past1 | -2.62 | Down-regulated | Involved in sperm individualization | 3 |
| Zpg | 1.33 | Up-regulated | Male germ-line stem cell population maintenance | 5 |
| En | -1.40 | Down-regulated | Involved in gonad development | 8 |
| Cell death related genes | ||||
| Ptp52F | 3.44 | Up-regulated | Involved in larval midgut cell programmed cell death | 15 |
| Atg7 | 1.60 | Up-regulated | Involved in autophagy | 2 |
| RnrS | 0.68 | Up-regulated | Involved in activation of cysteine-type Endopeptidase activity involved in apoptotic process | 9 |
| CG10700 | 0.822 | Up-regulated | Involved in execution phase of apoptosis | 9 |
| CG12917 | -0.62 | Down-regulated | Involved in apoptotic DNA fragmentation | 4 |
| Nbs | 0.78 | Up-regulated | Involved in intrinsic apoptotic signaling pathway in response to DNA damage | 2 |
| Lola | 0.74 | Up-regulated | Involved in nurse cell apoptotic process | 14 |
| CG32817 | 0.78 | Up-regulated | Involved in extrinsic apoptotic signaling pathway | 0 |
| Debcl | -1.92 | Down-regulated | Programmed cell death involved in cell development | 11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



