Submitted:
08 February 2024
Posted:
12 February 2024
You are already at the latest version
Abstract
Arbuscular mycorrhizal fungi (AMF) and rhizobium play a significant role in plant symbiosis. However, their influence on the soil microbiome associated with plant growth and soil health is not well defined in the Northern Great Plains. This study investigated the effect of microbial inoculants as seed treatment on plant yield, nutrient uptake, potential microbial functions, and rhizosphere soil microbial communities using high-throughput sequencing of 16S and ITS rRNA genes. The experiment was conducted under two contrasting dryland conditions with four treatments: control, single inoculation with AMF or Rhizobium, and dual inoculations of AMF and Rhizobium (AMF+Rhizobium). Our findings revealed that microbial inoculation efficacy was site-specific. In a non-P nutrient limiting condition, AMF+Rhizobium exhibited a synergistic effect on grain yield in Sidney, Montana (MT), dryland field site (DFS) 2. While AMF conferred plant resilience but marginal yield in low soil organic matter and acidic soil conditions in Froid, MT (DFS 1). Noteworthy, variation in microbial community structure and composition indicates site-specific response to AMF and AMF+Rhizobium microbial inoculants. Overall, site-specific factors had a greater influence on plant nutrient uptake, microbial community dynamics and functional potential. It underscores the need for tailored management strategies that consider site-specific characteristics to optimize benefits from microbial inoculation.
Keywords:
field pea
; arbuscular mycorrhizal fungi (AMF)
; Rhizobium
; microbial inoculants
; symbiosis
; microbiome
; agroecosystem
; drylands
1. Introduction
Dryland farming accounts for 60% of the total crop production, which plays a critical role in feeding the world’s growing population [1,2]. Over 40% of the global dryland area is utilized as cropland, often characterized by limited water availability [1,2]. Other major challenges are poor soil quality due to low organic matter content, nutrients, and biological activity [3]. In resource-limited semi-arid regions like the Northern Great Plains, it is imperative to adopt crop management that promotes soil health, which has distinct effects on microbial populations involved in water use efficiency, nutrient cycling, and other critical soil ecosystem functions.
Peas establish symbiotic associations with beneficial soil microorganisms and provide 3-20% photosynthetically assimilated carbon (C) to AMF and Rhizobium partners [4]. As a leguminous plant, it could obtain over 80% of the nitrogen (N) requirement from biological N fixation [5]. Further, mycorrhizal associations enhance water and nutrient uptake, including fixed N by bacteria and phosphorus (P) at significantly higher rates than nonmycorrhizal plants [6,7,8]. The synergistic interaction of AMF and Rhizobia enhances efficient nutrient uptake. These microbial symbioses impose a high demand for carbon from plants that necessitates increased CO2 assimilation from the atmosphere [8], which in turn increases plant exudates, residue, and microbial biomass that enhances soil organic C storage [9].
Agricultural practices heavily relying on fertilizers and pesticides can deplete soil carbon and fertility, and decrease pH, reducing soil microbial diversity and plant vigor [10,11]. The utilization of beneficial microbes is considered an emerging tool to circumvent agricultural intensification [12,13]. Microorganisms have been utilized as biofertilizers, biocontrol agents, and biostimulants over a decades. Several studies have reported the successful use of microbial inoculants, increasing crop yield and plant fitness [13,14] while others have reported nil to varying efficacy in field conditions [15,16]. In particular, arbuscular mycorrhizal fungi (AMF) have been recognized for their C, N, and P cycling, soil structure improvement, water and nutrient uptake, and plant tolerance to abiotic and biotic stresses [10]. Beneficial soil microorganisms enhance plant tolerance to biotic stress through Induced Systemic Resistance (ISR) [17], and environmental stresses through the production of the enzyme 1-amino-cyclopropane carboxylate deaminase (ACCD), which breaks down stress ethylene precursor ACC into 2- oxobutanoate and NH3 that decrease ethylene levels and facilitate plant growth [18]. These soil microbes’ beneficial traits have the potential to offset plant abiotic and biotic stresses and reduce reliance on agrochemical input. However, the influence of AMF and Rhizobium as microbial inoculants on crop agronomic performance is not well resolved in dryland field conditions.
The leguminous and mycorrhizal plant symbioses with Rhizobium and AMF had been known as tripartite interaction. However, the plant-soil microbiome interaction is considered highly complex, involving the plant host, microbial communities, soil, and associated environment contributing to soil health and productivity [12,13]. Studies of natural populations have shown that groups of microbes with distinct functions are crucial in nutrient mineralization, degradation of organic residues, and nutrient availability for plant utilization [19,20]. Recent studies have reported that AMF, phosphate-solubilizing bacteria, and the plant microbiome contribute to nutrient transformation and improved yield in other economically important crops such as barley [19] and corn [20]. Further, the rhizosphere soil microbial communities are largely influenced by plant exudates, soil-physicochemical properties, and management practices [21,22,23]. The microbial inoculants were applied as seed treatments, which may have a 'priority effect' in the microbiome assembly and function in the early plant developmental stage [24,25]. Understanding the rhizosphere soil microbial communities in response to microbial inoculants, dryland crop management practices, and site-specific conditions can provide insights into the essential functions contributing to plant fitness and resilience. Currently, there is a limited understanding of the influence of microbial inoculants on both bacterial and fungal communities relative to soil health in the drylands.
To better understand the influence of AMF and Rhizobium inoculation on plant growth and soil health at contrasting dryland sites, we used high-throughput sequencing of the 16S and ITS rRNA gene amplicons to elucidate the bacterial and fungal communities and their potential ecophysiological functions. The specific objectives of this research were to determine the influence of microbial inoculants on plant growth and nutrient dynamics; microbial diversity, structure, and composition; and potential microbial metabolic and ecophysiological functions. We hypothesize that microbial inoculation and dryland site-specific conditions significantly affect the pea rhizosphere microbial community, affecting crop productivity and soil health.
2. Materials and Methods
Site and soil characterization
The study was conducted in contrasting dryland field sites, in Froid (48C15'18.972 N, 104°29'39.843" W) and Sidney (48°15'18.972 N, 104°29'39.843" W), eastern Montana (Figure S1). The dryland field site (DFS) 1 in Froid field soil is characterized as Dooley sandy loam, while DFS 2 in Sidney is Williams loam soil, as previously described by Sainju et al. 2022 [26]. Both sites have a mean temperature of 21°C. The precipitation during the growing season was 55 mm and 25 mm for DFS 1 and DFS 2, respectively (Table S1). The DFS 1 has an available water supply of 0.17 cm of water per cm of soil, while DFS 2 has an available water supply of 0.18 cm per cm of soil (Table S2, USDA Soil Survey).
Soil samples were collected to characterize each site’s basal soil physicochemical properties. Soil analysis was performed at the Ward laboratories, Kearney, NE, following the prescribed protocol [27,28,29]. The available soil nutrients were compared to the pea nutrient requirements (Table S3). DFS 1 had a strongly acidic soil (pH = 4.7), low organic matter content (1.5% LOI), medium nitrogen (28.33 ppm N), very high phosphorus (73.00 ppm P), and potassium (214.5 ppm). Meanwhile, DFS 2 had a slightly neutral pH (pH= 6.33), moderate organic matter content (2.5% LOI), low nitrogen (7.60 ppm N), high phosphorus (44.75 ppm P), and very high potassium (277.87 ppm K).
Microbial inoculants
The microbial inoculants specific for field peas were arbuscular mycorrhizal fungi (AMF) Glomus intraradices (syn. Rhizophagus irregularis), Rhizobium leguminosarum bv. viceae and a dual inoculant of endomycorrhizal (AMF) and rhizobial products (AMF+Rhizobium) commercially produced by Premier Tech (Riviere-du-Loup, Quebec, Canada). The active ingredients were 2,750 viable spores of G. intraradices/g and 1.6 x 109 viable cells of R. leguminosarum bv. viceae per gram of the product. These inoculants were applied as seed treatment following the recommended 300g/ha application rate for pea [30].
Dryland field experiment
Seed inoculated peas (variety 4010) with microbial inoculants were planted in the dryland field sites described above. The study was set up in 1,858 m2, subdivided into five blocks of equal size, and each block had four plots. Each plot had a 6.10 m width x 15.24 m length. There were four treatments: control, AMF, Rhizobium, and dual inoculants (AMF+Rhizobium) arranged in a randomized complete block design (RCBD) with five replications in each site. All field sites were managed under no-till and regular cultural practices for field pea with no N fertilizer application (Table S4). P and K basal fertilizer was applied at seeding at a rate of 56 kg/ha and 45 kg/ha, respectively. Basagran 5l herbicide was applied at an 897g/ha rate to control weeds at the vegetative stage. Field pea plots were planted on 15 May (DFS 1) and 17 May 2022 (DFS 2). Before harvesting, biomass and plant yield components were gathered. The pea grains were harvested at maturity on 19 August (DFS 1) and 9 August 2022 (DFS 2) using a combine harvester.
Plant nutrient uptake, yield, and microbial dependency
The plant biomass and grain were ground separately to determine the nutrient concentration. A weight of 0.29 to 0.30g per sample was placed into a ceramic boat. C and N concentrations were measured using the LECO FP-2000 C-N analyzer (LECO Trumac Series, LECO Corporation, RI, USA). P content in grain was measured with an inductively coupled plasma (ICP) after hot block digestion following the methods previously described [31]. Nutrient uptake and carbon sequestration were calculated with the formula: Nutrient uptake or Carbon sequestration= (grain yield kg/ha x nutrient concentration) + (biomass yield kg/ha x nutrient concentration ) [32]. Total residual nutrient (nitrate, phosphorus) in the 0-60 cm depth soil was calculated with the formula: Total residual = nutrient concentration in ppm (mg/kg) x mass of soil (kg/ha), where mass of soil (k/ha) =100 (m) x 100 (m) x soil depth (m) x soil bulk density x 1000 (1000 kg=1 ton conversion factor). Microbial dependency (MD) of inoculation treatment was calculated according to Van Der Heijden 2002 [33] as follows: MD %= [(T - C)/C] x100, where T is the mean plant growth parameters (biomass or yield) in the given replicates of the microbial inoculated treatment and C is the mean in the corresponding noninoculated treatment or control group. Microbial dependency with positive values indicates plant growth promotion and negative values indicates plant growth suppresion by microbial inoculants.
Microbial community analysis
We explored bacterial and fungal populations by MiSeq sequencing, targeting the 16S and ITS1 rRNA gene amplicons, respectively. Forty-two days after seeding at the pea vegetative stage, rhizosphere soil samples from three plants in each treatment per plot in each site were collected on June 29 for DFS 1 and June 30, 2022 for DFS 2. Genomic DNA of rhizosphere soil fraction was extracted using a PowerSoil kit (Qiagen, Valencia, CA, USA) following the manufacturer’s instructions. The extracted DNA samples were analyzed for quality and quantity using NanoDrop 1000 (Thermo Scientific, Wilmington, DE, USA). The DNA samples were sent to the University of Minnesota Genomics Center Microbiome Services for sequencing following the 16S and ITS Illumina Amplicon Protocol. The V4 – V5 region of the 16S rRNA gene was amplified with the 515F/926R primer pair [34,35], while the fungal ITS1 region was amplified using ITS1F/ITS2_Nextera primer pairs [36,37]. The PCR product was diluted 1:100 with molecular-grade water. The diluted amplicons were indexed using barcoded PCR primers [37]. Barcoded amplicons were normalized using SequalPrep kits (Invitrogen). The normalized libraries were cleaned with AMPure XP mag beads (Beckman Coulter), quantified by Qubit (Invitrogen), and sequenced using the MiSeq v3 600-cycle kit on the MiSeq platform (Illumina).
Quantitative Insights into Microbial Ecology (QIIME 2 2023.3) bioinformatic pipeline was used to analyze the raw sequences [38]. 16S rRNA demultiplexed paired-end fastq files have been demultiplexed with adapters removed in the process, while the primer and adapter sequences of ITS demultiplexed paired-end fastq files were trimmed off at the 5’ and 3’ end, respectively, using cutadapt [39]. DADA2 was used for denoising, sequence correction, and removal of chimeras (qiime dada2 denoise-paired with the following parameters: --p-trim-left-f 0 paired --p-trim-left-r 0 --p-trunc-len-f 290 --p-trunc-len-r 290 for 16S rRNA sequences, while --p-trim-left-f 0 paired --p-trim-left-r 0 --p-trunc-len-f 170 --p-trunc-len-r 170 for the ITS sequences) [40]. The forward and reverse reads of all the sequences were truncated based on the quality scores (phred ≥ 30).
The BIOM table was summarized with the Qiime2 ‘feature table summarize’ command. A phylogenetic tree was constructed using qiime phylogeny align-to-tree-mafft-fasttree [41,42]. The amplicon sequence variant (ASV) taxonomic identification was performed by using the q2-feature-classifier [43] against the SILVA 132 [44] database and BLAST+ consensus taxonomy classifier [45] for prokaryote 16S rRNA gene, while the UNITE v8 database [46] and classify sklearn for fungal ITS. We obtained 1,474,948 total 16S rRNA sequences, of which 162,466 were prokaryotes with high-quality reads clustered into 2,862 amplicon sequence variants (ASVs). Of the 1,264,985 ITS sequences, 750,118 were high-quality fungal reads clustered into 2,190 ASVs. Data were filtered for low counts of 20% prevalence of a minimum of four counts in each sample and a 10% low variance filter based on the interquartile range. All samples were rarified to even sequencing depth based on the lowest sampling depth. Principal coordinate analysis (PcoA) was used to visualize the effect of microbial inoculants on the microbial community composition and structure [47]. Microbiome datasets with metadata were further visualized using microbiomeAnalyst [48] and RAWgraphs [49].
The Tax4Fun2 R package was used to estimate the metabolic functional features of bacterial communities, which integrates data from 16S rRNA genes from the Kyoto Encyclopedia of Genes and Genomes [50]. We focused on predicted genes involved in C fixation, N metabolism, and P. Fungal ecological functions were predicted using the database FungalTraits [51]. The database FungalTraits was utilized to forecast the ecological activities of fungi [51]. We concentrated on the dynamics of the potential functions associated with soil health, plant-microbe dynamics, and nutrient cycling.
The influence of microbial inoculants on rhizosphere microbial communities in the dryland sites was examined using permutational multivariate analysis of variance (PERMANOVA) [52]. The heat tree analysis was used to plot the differential abundance of bacterial and fungal ASVs between the microbial inoculated and the control based on median abundance and non-parametric Wilcoxon test [53]. Two-way ANOVA was conducted using JMP Pro Statistics, version 17 (SAS Institute, Cary, NC) to test the effects of microbial inoculants and dryland site conditions on nutrient content, nutrient uptake, and yield. The normal distribution and the variance homoscedasticity were analyzed using Shapiro-Wilk and Levene’s tests. Non-parametric Kruskal-Wallis test analysis was performed on the variables that failed the test. Post hoc mean comparisons were completed with Protected Fisher’s Least Significant Difference (LSD).
3. Results
Influence of microbial inoculants on plant agronomic performance and nutrient dynamics
Pea grain yield was significantly affected by AMF+Rhizobium in the dryland field site 2 (DFS 2) with moderate soil organic matter (SOM) and neutral pH (Figure 1a). Plants inoculated with AMF+Rhizobium had a significantly higher yield than the control (z=2.506, P=0.012). Based on microbial dependency analysis, a positive contribution of AMF+Rhizobium (16%) and a negative contribution of AMF treatment (-8%) were linked to yield and overall plant growth under DFS 2 (Figure 1b). The effect of microbial inoculation was insignificant in DFS 1 with low SOM and soil pH, although AMF treatment had an increased positive contribution to plant growth and yield by 25% (Figure 1b). DFS 2 had a significantly higher grain yield than DFS 1. The high initial soil nitrogen did not translate to a higher grain yield, although vegetative plant growth was more profuse in DFS 1 (Figure S2). Across dryland sites with high soil P fertility levels, microbial inoculants’ contribution was 12% by AMF+Rhizobium, 8% by AMF, and 4 % by Rhizobium to the overall plant growth (Figure 1b and Table S5). No significant variations among the treatments were observed in plant stand, nodulation, and plant biomass (Figure S3).
Meanwhile, nutrient contents in the grains were less influenced by microbial inoculants but significantly differed between dryland sites. Although, plants treated with AMF+Rhizobium and AMF inoculants had a significantly higher percentage of carbon (% C) in the grains than the control in DFS 1 (P=0.04, Figure 2a and Table S6). Carbon content (kg/ha) in plant biomass and grains was significantly higher in DFS 2 than in DFS 1(Table S7). There was no significant difference among treatments on N and P uptake within each site. Nonetheless, trend showed an increased nitrogen and phosphorus content in pea grains of AMF+Rhizobium and AMF treatments, respectively (Table S6). The nitrogen content in the grains was significantly higher in DFS 1 with greater initial soil N than in DFS 2. Phosphorus content in the grains was significantly higher in DFS 2, although it had a lesser initial soil P than in DFS 1 with strongly acidic soil (Table S2). The effect of site-specific conditions was greater than that of microbial inoculants on plant nutrient uptake (Tables S7 and S8). DFS 2 had a significantly higher total plant carbon and P uptake between sites, while DFS 1 had a significantly higher N uptake irrespective of the microbial inoculants ( P ≤0.001) (Tables S7 and S8). The initial soil nutrients were significantly high particularly in DFS 1, corresponding to higher nutrient residuals than DFS 2 (Figure 2c–f and Table S2). After the pea cropping, the N and P nutrient residuals were significantly higher than the nutrients utilized by the plants in DFS 1(Figure 2c–f, and Table S9). In DFS 2, plants utilized more nutrients efficiently, thus, there was lesser residuals. The difference in nutrient dynamics indicates a greater influence of site-specific variation than microbial inoculation influencing nutrient availability and plant uptake beyond initial soil nutrients.
Variation in the soil microbial communities associated with increase plant yield in two contrasting dryland sites
Microbial alpha-diversity analysis in dryland field sites displayed various levels of richness, as indicated by the number of species observed (Figure 3a,b). AMF+Rhizobium and the control exhibited the highest bacterial diversity, while AMF treated in the DFS 1 showed the lowest mean number of bacterial species (Figure 3a). No significant difference was observed in bacterial species richness between sites (Table S10 and Figure S4). On the other hand, fungal species richness significantly increased in the rhizosphere of plants treated with microbial inoculants (Figure 3b). DFS 2 had significantly higher fungal species richness than DFS 1 (P =1.02e-07). The Bray-Curtis index was used to calculate beta-diversity values, which were visualized in PCoA plots and compared using the PERMANOVA test of significance to determine the variations in microbial community structures. Clustering of samples based on the site and site-specific response to microbial inoculants significantly explained (P < 0.001) the highest percentage of variation in community structure for the bacterial fraction (R2 = 0.31 and R2=0.43) (Figure 3c,d) and fungal (R2 = 0.48 and R2=0.55) fractions of the population (Figure 3e,f). This data showed that specific site favors distinct soil microbial communities and a more prevalent site-specific response to microbial inoculants. However, the fraction of variance in microbial community structure attributed to microbial inoculants was minimal. This indicates that site-specific conditions strongly influenced microbial community structures, and the response varies between sites.
The compositional differences of rhizosphere soil microbiomes as influenced by site-specific response to microbial inoculants were shown in the distribution of the dominant phyla and taxonomic orders (Figure 4, Table S11). Of note is the significant (P < 0.01) enrichment of Actinobacteria (relative abundance ~35-38%) in the bacterial communities in both dryland sites (Figure4a). It was significantly high in Rhizobium-treated microbial communities. By comparison, DFS 2 soils contained the highest percentage of Bacteroidetes, Firmicutes, and Nitrospirae (Table S11), while WPS2 and Planctomycetes were enriched in DFS 1. These dominant phyla were represented by 11 abundant orders (>1% of all sequences). Of these, seven taxonomic orders, which include Propionibacteriales (relative abundance ~5 – 19%), Rhizobiales (relative abundance ~5 – 7%), Chitinophagales (relative abundance ~5 – 7%), Frankiales (relative abundance ~3 – 7%), Elsterales (relative abundance ~2 – 6%), Thermomicrobiales (relative abundance ~0.3 – 6%), and Bacillales (relative abundance ~0.8 – 5%) were significantly enriched in DFS 2 (Figure 4c and Table S11). While Acetobacterales, Acidobacteriales (relative abundance ~0.5 – 6%), Gaiellales (relative abundance ~6%), and uncultured bacterium (relative abundance ~9%) were enriched in DFS 1 (Figure 4c). Rhizobiales was more abundant in the microbial treatments than in the control in DFS 2 (Figure 4c). AMF+Rhizobium had significant enrichment of Elsterales. AMF treated microbial communities were enriched in Acidobacteriales (Figure 4c and Table S11). Elsterales, Nitrospirales, Nitrososphaerales, Thermomicrobiales, Frankiales, Betaproteobacteriales, Chitinophagales, and Bacillales were abundant in DFS 2 than DFS 1 (Figure 4c).
Moreover, fungal community patterns were more delineated by sites. Mortierellomycota, Chytridiomycota, and unassigned phyla were enriched in DFS 2, while Mucoromycota was enriched in DFS 1 (Figure 4b and Table S12). Significant enrichment of taxonomic orders of Mortierellales, Pleosporales, Coniochatales, Helotiales, and Chaetosphaeriales was observed in DFS 2, while Eurotiales, Filobasidiales, Holtermanniales, Rhizophlyctidales, Sordariales and Tremellales in DFS 1 (Figure 4d and Table S12). The fungal composition data showed that site-specific factors strongly influence the fungal communities in the rhizosphere, wherein each site harbors a unique set of fungal taxa.
Taxonomic hierarchical data comparison based on the heat tree analysis of the microbial communities associated with high grain yield in the AMF treated in DFS 1 revealed a lesser abundance of Chujaibacter, and Segetibacter bacteria than in the untreated control (Figure 5a and Table S13). While fungal communities in the AMF treated had a significantly higher Selenophoma under the order Dothideales, Keissleriella, and Parastagonospora under the family Lentitheciaceae within the fungal order Pleosporales than the untreated control (Figure 5b, and Table S14). The decreased bacterial and increased abundance of fungal groups indicate a community shift in response to the AMF treatment, favoring some fungal over bacterial groups in challenging conditions. Meanwhile, microbial communities in the AMF+Rhizobium-treated plants associated with significantly high yield in DFS 2 revealed an enrichment of several uncultured bacteria under Elsterales (Figure 5c and Table S13). Further, Exophiala, Mortierella, Fusicolla, and other fungal species under the Sordariomycetes were enriched in the AMF+Rhizobium treated than the untreated control (Figure 5d and Table S14). The differentially abundant taxa indicate the influence of AMF and AMF+Rhizobium on the microbial communities, contributing to increased plant yield in unfavorable and non-P limiting conditions, respectively.
Effect of microbial inoculants on the potential functions of the microbial community relative to nutrient cycling and soil health
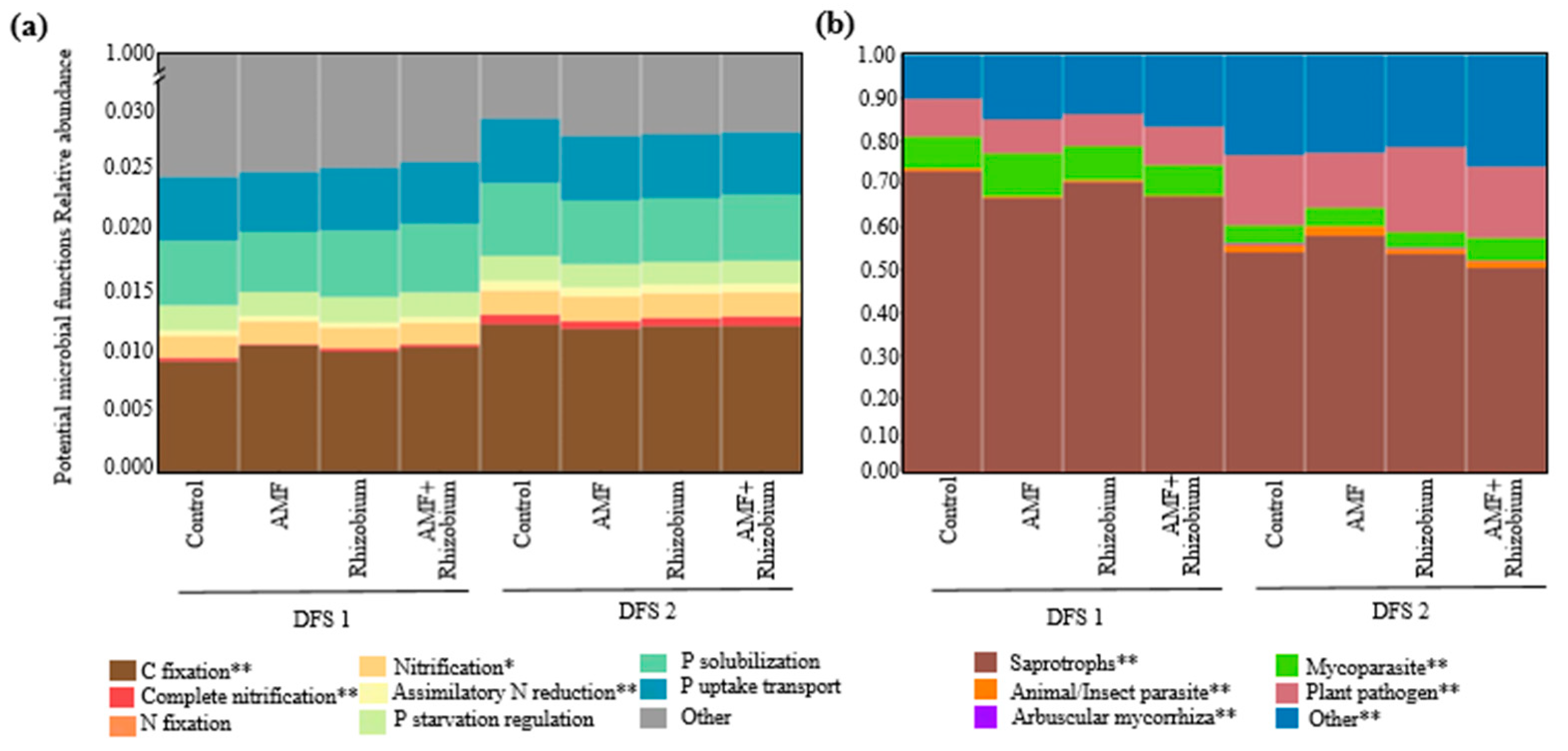
The predicted metabolic functional profile of bacterial communities was obtained using Tax4Fun2 based on the 16S rRNA genes and the KEGG Ortholog groups (KOs). We narrowed down the predicted genes to C, N, and P nutrient cycling (Figure 6a and Table S15). The C and N genes relative abundance were significantly higher in DFS 2 than in DFS 1(P ≤ 0.001). Meanwhile P uptake, P solubilization, and P starvation regulation did not vary between sites with very high P. Investigating fungal ecophysiological functions revealed significant differences between dryland sites, indicating that each site's fungal community has distinct ecological roles and functional adaptations. DFS 2 had a higher relative abundance of arbuscular mycorrhizal fungi (AMF), biocontrol agents such as animal/insect parasites, plant pathogens, and others. While DFS 1 had a higher relative abundance of saprotrophs and mycoparasites (Figure 6b and Table S16). In non-P nutrient or other limiting conditions, distinct bacterial nutrient cycling and fungal ecological functions were more strongly influenced by site-specific factors rather than microbial inoculations.
4. Discussion
In this study, the effectiveness of microbial inoculants exhibited variation across contrasting dryland sites. AMF+Rhizobium contributed positively to plant growth and yield in non-P nutrient-limited but low N conditions. The synergy of AMF and Rhizobium had been reported in previous studies in other legumes such as soybean (Glycine max) [54] and alfalfa (Medicago spp.) [4]. Conversely, AMF treatment had a negative impact on plant growth and yield in high P fertility levels. These findings corroborate with previous studies that AMF does not further improve the host plant’s P budget at high P levels in the rhizosphere [4,55,56]. Across dryland sites, the marginal effect of microbial inoculants on grain yield suggests that the net costs of the symbiosis exceed the benefits for the plants when nutrients are abundant. Plants tend to rely on mineral uptake independent from beneficial microbes, which requires less energy than the demanding process of microbial-dependent uptake [57]. It has long been known that inorganic fertilizers alter AMF and Rhizobium symbioses [58, 59]. In our study, the high initial soil fertility corresponds to high N and P soil nutrient residuals.
Under unfavorable soil conditions, a positively high microbial dependency on AMF contributed to plant growth and yield. Nonetheless, the yield in DFS 1 was significantly lesser than in DFS 2, which follows the principles of limiting factors that dictate the level of crop production, which cannot exceed what is allowed by th e maximum limiting factor [60]. In this study, the soil acidity resulted in nodulation failure irrespective of any microbial inoculants. The acidic soil conditions affect plants and microbial symbionts and restrict nutrient availability [61]. Rhizobium symbiosis, which demands high phosphorus (P), faces challenges in acidic soils where P fixation or immobilization occurs, leading to reduced P bioavailability [62]. The high initial soil nutrients in DFS 1 did not translate to a higher yield, indicating that soil pH, soil organic matter (SOM), and other factors substantially influenced yield.
Noteworthy, microbial inoculation of AMF and AMF+Rhizobium significantly increased carbon content in the grains in DFS1, suggesting that microbial inoculants positively accumulated more carbon in the grains. AMF+Rhizobium had the highest C sequestered in plants, which indicates C sink stimulation from the symbioses [8]. However, single inoculations (AMF or Rhizobium) showed a slightly lower C sequestered than the dual inoculated plants, which points to the plant cost of C allocation being higher than the nutrients gained from AMF or Rhizobium symbiosis alone [63]. Our results showed plants had a higher microbial dependency to acquire water, P, and N resources from AMF+Rhizobium than single inoculations. These findings point to the significance of dual microbial inoculants in promoting carbon sequestration and efficient resource acquisition by the plants in dryland conditions.
Crop productivity and soil health are associated with belowground microbial diversity [12,13,64]. Microbial diversity would likely be low in DFS 1 with low soil organic matter, which has less energy source for the microbes, and the low pH would select few microbial species tolerant to acidic soil conditions. However, our 16S rRNA microbiome data revealed no significant variation in bacterial species richness between the dryland sites, which suggests that bacterial species have a broad adaptation to varying soil conditions and distinct bacterial groups thrive in each site. The fungal communities showed a more pronounced site-specific response to soil conditions and microbial inoculants. Fungal diversity was higher in the site with favorable soil conditions (DFS 2) and responded more strongly to AMF and AMF+Rhizobium inoculations. Of important note, AMF and AMF+Rhizobium treated in DFS 1 were comparable to the fungal species richness in DFS 2, which points to possible stress alleviation by microbial inoculants. The site-specific response to microbial inoculants was more pronounced for the fungal communities than the bacterial communities, which support earlier reports that fungal communities tend to be more sensitive to different agronomic practices and environmental changes due to their complex life cycles and niche requirements than bacterial communities [65,66]. AMF+Rhizobium enriched fungal species at different dryland gradients.
In our study, the microbial community structure and composition variation indicates that specific site favors distinct soil microbial communities and a more prevalent site-specific response to AMF and AMF+Rhizobium microbial inoculants. The bacterial phylum Actinobacteria was enriched in both dryland sites, consistent with another study that this phylum is well adapted to water-limited or drought conditions [67]. At DFS 2, the bacterial communities were dominated by Bacteroidetes [68] and Nitrospirae [69] phyla, which are involved in nutrient cycling. Firmicutes was associated in high P soils [70], in plant growth promotion and pathogen suppression [71]. Microbial inoculations with AMF, Rhizobium, and AMF+Rhizobium were enriched in Rhizobiales. This taxonomic order is comprised of mostly nitrogen-fixing bacteria [72]. AMF+Rhizobium treatment significantly enriched Esterales and Frankiales, which suggests a synergistic influence on the abundance of these copiotrophic bacteria thriving in nutrient-rich substrates [73]. AMF, particularly Rhizophagus irregularis (syn. Glomus intraradices) has been reported to harbor bacterial communities in the hyphosphere of the fungus and increase bacterial resilience in water-limited conditions [74]. While at DFS 1, the bacterial communities were dominated by Acidobacteria, a phylum comprised of oligotrophic bacteria that breaks down dissolved organic matter [75]; persists in low pH [76]; and is associated with low nutrient soil conditions [70]. Planctomycetes enrichment correlated with soil NO3 levels [77]; and the other less studied phylum, WPS2. The taxonomic order Acidobacteriales, under Acidobacteria, was significantly enriched in the AMF treated. These findings suggest that soil conditions and microbial inoculants substantially influence the composition of bacterial communities in the rhizosphere.
In the fungal community, Ascomycota and Basidiomycota were the dominant fungal phyla across sites, but only the composition of other major phyla differed significantly between sites. At DFS 2, the fungal communities were dominated by saprophytic phyla, Chytridiomycota, and Mortierellomycota [75], and unassigned phylum. In contrast, at DFS 1, the fungal communities were dominated by Mucoromycota phylum with a wide range of beneficial and pathogenic members [78]. There was a significant enrichment of saprophytic and mycoparasitic fungal orders in DFS 2 compared to DFS 1. Some patterns of more saprophytic and mycoparasitic fungal orders in the microbial inoculated, such as Orbillales, Cystobasidiales, and Myrmecridiales in DFS 1, while Mortieralles and Chaetosphaeriales in DFS 2 [51]. Unlike in the untreated control with more fungal groups that have both saprotrophic and pathogenic life strategies, such as Agaricales, Pezizales, Xylariales, Rhizophlyctidales, and Dothideales [51].
Microbial community dynamics linked with improved crop performance in challenging conditions showed a shift in composition in response to the AMF treatment. AMF promoted the growth of saprophytic fungal genera Selenophoma and Keissleriella [51], which may compensate for the absence of putative C-cycling bacteria Chujaibacter [79], and other nutrient cycling bacteria such as Elsterales (Alphaproteobacteria), Segetibacter (Bacteroidetes) [68], and uncultured Chloroflexi bacterium in DFS 1. On the other hand, the microbial communities in the AMF+Rhizobium-treated plants associated with significantly higher yield in DFS 2 had a higher abundance of several uncultured bacteria under Esterales. These bacteria have not been characterized, but they may be beneficial in promoting plant growth and yield. The fungal communities in the AMF+Rhizobium-treated plants had a higher abundance of fungal genera Exophiala, Mortierella, and Fusicolla. These fungi are soil saprotrophs, fungal decomposers, and mycoparasite [51], which may positively affect soil microbes and plants. The enrichment of these beneficial bacteria and fungi in the AMF and AMF+Rhizobium-treated plants may represent positive interactions on possible nutrient exchanges or protection from environmental stress that contribute to increased plant yield.
Potential functions of the microbial community based on the relative abundance of the predicted genes involved in nutrient cycling (C fixation, N metabolism, and P) and ecophysiological traits related to overall soil health revealed a stronger influence of site-specific factors than the microbial inoculants under a non-P limiting and unfavorable soil conditions. This indicates that the site conditions’ soil pH, SOM, and nutrient availability greatly influence plant-microbe interactions [25,80], including the microbial inoculants. Meanwhile, fungal communities in each site have distinct ecological roles and functional adaptations. Fungal communities in DFS 2 were more involved in nutrient cycling and plant-microbe interaction due to a higher relative abundance of AMF, biocontrol agents, and plant pathogens. On the other hand, fungal communities in DFS 1 were more involved in decomposition and nutrient cycling. These findings suggest that the soil conditions in each site play a major role in shaping the functional potential of the fungal communities. These findings suggest managing soil pH, SOM, and nutrient availability as a prerequisite for optimizing the benefits of microbial inoculants in improving the functional potential of microbial communities in dryland soils.
This study focused on the influence of microbial inoculants on the pea rhizosphere soil microbial communities under contrasting dryland field conditions. Mycorrhizal colonization was not assessed due to variability in root collection and constraints in delineating the inoculants from native beneficial soil microbes. The inoculants, both pea symbionts applied as seed treatment, have a niche advantage that could influence the microbial community directly or indirectly [25,26]. Although, in an optimal environments, it is likely that established plant-microbiome equilibrium may resist the influence of external inoculants due to low competitive pressure [81]. Noteworthy, we found a site-specific microbiome variation in response to microbial inoculation which resulted to either shift in microbial community composition or enrichment of beneficial soil microbes in sub-optimal dryland conditions. The extent of the microbial inoculants’ influence to cause significant impact on microbial community functions relative to various agricultural management practices, other abiotic and biotic stresses in the drylands necessitates further studies. Our in situ field study captured a snapshot of the bacterial and fungal communities influenced by the inoculants in contrasting dryland field sites.
5. Conclusions
Our study provides valuable insights into the influence of AMF and Rhizobium inoculants on plant yield, C sequestration, nutrient dynamics, microbial diversity, and the potential microbial community functions between contrasting dryland field sites. We used a high throughput sequencing approach to investigate the influence of microbial inoculants on the rhizosphere bacterial and fungal communities. Our findings revealed that the efficacy of microbial inoculants is highly site-dependent. AMF and AMF+Rhizobium significantly increased grain yield and improved nutrient acquisition under specific dryland conditions. While microbial inoculants offer benefits in favorable conditions, their effectiveness is limited in high soil fertility, low soil organic matter, and acidic soil conditions. Microbial community dynamics exhibited site-specific variation, which showed a distinct microbial community pattern and a shift in bacterial and fungal species abundance between sites. The soil conditions showed a profound impact on potential microbial functions relative to nutrient cycling and ecophysiological functions. Additional follow-up studies will need to examine the influence of microbial inoculants on low-input field management, qPCR quantification of microbial inoculants, validation of microbial community functions, and disease build up in a continous pulse rotation. Future efforts to innovate crop and soil management tailored to specific drylands' unique characteristics and challenges to enrich the plant-soil microbiome for overall soil health and sustainable dryland crop production.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Monthly mean temperature and precipitation from May to August during 2022 growing seasons at the two-dryland sites in Froid (DFS 1) and Sidney, Montana (DFS 2); Table S2: Characterization of the soil physicochemical analysis of the two contrasting dryland sites; Table S3: Pea nutrient requirement and baseline soil physicochemical analysis of the two contrasting dryland sites; Table S4: Field management practices and chemical applications; Table S5: Microbial dependency (%) data showing the microbial inoculants’ contribution to aboveground biomass, grain yield and overall plant growth at two dryland sites; Table S6: Effect of microbial inoculations on plant biomass and grain nutrient content (% by mass) at two dryland sites; Table S7: Effect of microbial inoculants on carbon sequestered/stored in the harvested plant biomass and grains at two dryland sites; Table S8: Effect of microbial inoculants on plant N and P nutrient uptake at two dryland sites; Table S9: Effect of microbial inoculation on residual NPK (kg/ha) after pea cropping; Table S10: Alpha and beta diversity of pea rhizosphere soils as influenced by microbial inoculants at two dryland conditions, MT 2022; Table S11: Influence of microbial inoculants at two dryland sites on the most abundant (relative sequences abundance > 1% of all bacterial sequences) bacterial taxonomic group level phylum, order and genera on pea rhizosphere soil; Table S12: Influence of microbial inoculants at two dryland sites on the most abundant (relative sequences abundance > 1% of all fungal sequences) fungal taxonomic group level phylum, order and genera on pea rhizosphere soil; Table S13: Bacterial taxonomic difference between inoculated microbial communities and the control obtained from heat tree analysis, using the median abundance and non-parametric Wilcoxon Rank Sum test (Foster et al., 2017); Table S14: Fungal taxonomic difference between inoculated microbial communities and the control obtained from heat tree analysis, using the median abundance and non-parametric Wilcoxon Rank Sum test [53]; Table S15: Influence of the microbial inoculants on the relative abundance of potential gene category involved in plant nutrient uptake at two dryland sites. The functional profiles of bacterial communities were predicted based on the 16S rRNA genes of retrieved bacterial taxa using Tax4Fun2 according to the KEGG Ortholog groups (KOs); Table S16: The relative abundance of potential fungal traits in dryland condition. The ecological functional profiles of fungal communities were predicted based on the FungalTraits database. Figure S1: The field sites at (a) DFS 1(Froid) and (b) DFS 2 (Sidney) were managed under no-till practices. Each site was set up in 20,000 sq.ft. (USDA Soil Survey; Figure S2: The yield components across dryland field sites. Two-way ANOVA analysis, means followed by different letters are significantly different according to Tukey’s Test HSD at P <0.05 (n=400 plants); Figure S3: Boxplots of the (a) plant stand, (b) nodulation rating scale using Yates et. al., 2017 [82], and (c) plant biomass across dryland field sites. Two-way ANOVA analysis, means followed by different letters are significantly different according to LSD at P <0.05; Figure S4: Influence of microbial inoculants on microbial species richness comparison among treatments and between sites on (a, b) bacterial, and (c, d) fungal communities. Boxplot and LS mean lines with common letter are not significantly different based on LSD tests at 0.05% probability level.
Author Contributions
Conceptualization, S.R.D; experiment implementation and investigation, R.B.C. and S.R.D; data processing, visualization, and writing—original draft preparation, R.B.C; writing—review and editing, S.R.D; supervision, project administration and funding acquisition, S.R.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Specialty Crop Block Grant of Montana Department of Agriculture (Grant: 22SC002314 - Influence of Arbuscular Mycorrhizal Fungi and Rhizobium on Soil Health and Plant Growth).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequence data analyzed in this study were deposited in the NCBI database, BioProject PRJNA1020395 (https://www.ncbi.nlm.nih.gov).
Acknowledgments
The authors gratefully acknowledge Brett Allen, William Bart Stevens, Upendra Sainju, Jay Jabro, Bill Iversen, and Josh Campbell for their expert recommendation. Dora Alvarez, Michael Johnson, Nichole Davidson, Margaret Duffy, Chloe Turner-Meservy, Nancy Webb, Julia Seiller, Rachel Cross, Lacie Selting and Jessie Selting for their assistance with field and laboratory work. Portions of this research were conducted with high performance computing resources through the USDA SCINet Scientific Computing.
Conflicts of Interest
The authors declare no conflict of interest. USDA Disclaimer: Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply a recommendation or endorsement by the U.S. Department of Agriculture. USDA is an equal opportunity provider and employer.
References
- FAO of the, UN. Food and Agriculture Organization of the United Nation. 2018 [cited 2019 Jan 5]. CropWat | Land & Water | Food and Agriculture Organization of the United Nations. Available from: http://www.fao.org/land-water/databases-and-software/crop-information/soybean/en/.
- Liebig, M.A.; Acosta-Martinez, V.; Archer, D.W.; Halvorson, J.J.; Hendrickson, J.R.; Kronberg, S.L.; Samson-Liebig, S.E.; Vetter, J.M. Conservation practices induce tradeoffs in soil function: Observations from the northern Great Plains. Soil Sci. Soc. Am. J. 2022, 86, 1413–1430. [Google Scholar] [CrossRef]
- Peterson, GA. Dryland Farming. In: Reference Module in Earth Systems and Environmental Sciences. Elsevier; 2018.
- Püschel, D.; Janoušková, M.; Voříšková, A.; Gryndlerová, H.; Vosátka, M.; Jansa, J. Arbuscular Mycorrhiza Stimulates Biological Nitrogen Fixation in Two Medicago spp. through Improved Phosphorus Acquisition. Front. Plant Sci. 2017, 8, 390. [Google Scholar] [CrossRef]
- Ruisi, P.; Giambalvo, D.; Di Miceli, G.; Frenda, A.S.; Saia, S.; Amato, G. Tillage Effects on Yield and Nitrogen Fixation of Legumes in Mediterranean Conditions. Agron. J. 2012, 104, 1459–1466. [Google Scholar] [CrossRef]
- Jones MD, Durall DM, Tinker PB. A comparison of arbuscular and ectomycorrhizal Eucalyptus coccifera : Growth response, phosphorus uptake efficiency and external hyphal production. New Phytol. 1998, 140, 125–134.
- Goltapeh EM, Danesh YR, Prasad R, Varma A. Mycorrhizal fungi: what we know and what should we know? In: Mycorrhiza. Springer; 2008. p. 3–27.
- ezáčová V, Konvalinková T, Jansa J. Carbon fluxes in mycorrhizal plants. Mycorrhiza-eco-physiology, Second Metab Nanomater. 2017, 1–21.
- Yang, L.; Luo, Y.; Lu, B.; Zhou, G.; Chang, D.; Gao, S.; Zhang, J.; Che, Z.; Cao, W. Long-term maize and pea intercropping improved subsoil carbon storage while reduced greenhouse gas emissions. Agric. Ecosyst. Environ. 2023, 349. [Google Scholar] [CrossRef]
- Khangura, R.; Ferris, D.; Wagg, C.; Bowyer, J. Regenerative Agriculture—A Literature Review on the Practices and Mechanisms Used to Improve Soil Health. Sustainability 2023, 15, 2338. [Google Scholar] [CrossRef]
- Xavier, G.R.; Jesus, E.d.C.; Dias, A.; Coelho, M.R.R.; Molina, Y.C.; Rumjanek, N.G. Contribution of Biofertilizers to Pulse Crops: From Single-Strain Inoculants to New Technologies Based on Microbiomes Strategies. Plants 2023, 12, 954. [Google Scholar] [CrossRef]
- Chouhan, G.K.; Verma, J.P.; Jaiswal, D.K.; Mukherjee, A.; Singh, S.; Pereira, A.P.d.A.; Liu, H.; Abd_Allah, E.F.; Singh, B.K. Phytomicrobiome for promoting sustainable agriculture and food security: Opportunities, challenges, and solutions. Microbiol. Res. 2021, 248, 126763. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Li, Y.; Laterrière, M.; Lay, C.-Y.; Klabi, R.; Masse, J.; St-Arnaud, M.; Yergeau, É.; Lupwayi, N.Z.; Gan, Y.; Hamel, C. Effects of arbuscular mycorrhizal fungi inoculation and crop sequence on root-associated microbiome, crop productivity and nutrient uptake in wheat-based and flax-based cropping systems. Appl. Soil Ecol. 2021, 168, 104136. [Google Scholar] [CrossRef]
- Ryan, M.H.; Graham, J.H. Little evidence that farmers should consider abundance or diversity of arbuscular mycorrhizal fungi when managing crops. New Phytol. 2018, 220, 1092–1107. [Google Scholar] [CrossRef]
- Qiu, Z.; Egidi, E.; Liu, H.; Kaur, S.; Singh, B.K. New frontiers in agriculture productivity: Optimised microbial inoculants and in situ microbiome engineering. Biotechnol. Adv. 2019, 37, 107371. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Lugtenberg B, Kamilova F. Plant-Growth-Promoting Rhizobacteria. 2009; Annual Reviews Microbiol. 2009:63:541.
- Masrahi, A.S.; Alasmari, A.; Shahin, M.G.; Qumsani, A.T.; Oraby, H.F.; Awad-Allah, M.M.A. Role of Arbuscular Mycorrhizal Fungi and Phosphate Solubilizing Bacteria in Improving Yield, Yield Components, and Nutrients Uptake of Barley under Salinity Soil. Agriculture 2023, 13, 537. [Google Scholar] [CrossRef]
- Cozzolino V, Monda H, Savy D, Di Meo V, Vinci G, Smalla K. Cooperation among phosphate-solubilizing bacteria, humic acids and arbuscular mycorrhizal fungi induces soil microbiome shifts and enhances plant nutrient uptake. Chem Biol Technol Agric. 2021, 8, 1–18.
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2013, 8, 790–803. [Google Scholar] [CrossRef]
- Calderon, R.B.; Jeong, C.; Ku, H.-H.; Coghill, L.M.; Ju, Y.J.; Kim, N.; Ham, J.H. Changes in the Microbial Community in Soybean Plots Treated with Biochar and Poultry Litter. Agronomy 2021, 11, 1428. [Google Scholar] [CrossRef]
- Dastogeer, K.M.; Tumpa, F.H.; Sultana, A.; Akter, M.A.; Chakraborty, A. Plant microbiome–an account of the factors that shape community composition and diversity. Curr. Plant Biol. 2020, 23, 100161. [Google Scholar] [CrossRef]
- Fitzpatrick, C.R.; Schneider, A.C. Unique bacterial assembly, composition, and interactions in a parasitic plant and its host. J. Exp. Bot. 2020, 71, 2198–2209. [Google Scholar] [CrossRef]
- Wei, Z.; Gu, Y.; Friman, V.-P.; Kowalchuk, G.A.; Xu, Y.; Shen, Q.; Jousset, A. Initial soil microbiome composition and functioning predetermine future plant health. Sci. Adv. 2019, 5, eaaw0759. [Google Scholar] [CrossRef]
- Sainju UM, Liptzin D, Stevens WB. How soil carbon fractions relate to soil properties and crop yields in dryland cropping systems? Soil Sci Soc Am J. 2022, 86, 795–809.
- Gelderman RH, Beegle D. Nitrate-nitrogen. Recomm Chem soil test Proced North Cent Reg North Cent Reg Res Publ. 1998, 221:17–20.
- Warncke D, Brown JR. Potassium and other basic cations. Recomm Chem soil test Proced North Cent Reg. 1998, 1001:31.
- Combs SM, Nathan M V. Soil organic matter. Recomm Chem soil test Proced North Cent Reg. 1998, 221:53–8.
- AGTIV. Active ingredients [Internet]. 2023 [cited 2023 Nov 8]. Available from: https://www.ptagtiv.com/en/active-ingredients/#mycorrhizae.
- Campbell CR and PCO. Reference plant analysis procedures for the southern region of the United States. South Co-op Ser Bull. 1992, 368:71–3.
- Paye WS, Szogi AA, Shumaker PD, Billman ED. Annual Ryegrass (Lolium multiflorum Lam.) Growth Response to Nitrogen in a Sandy Soil Amended with Acidified Manure and Municipal Sludge after “Quick Wash” Treatment. Agronomy. 2023, 13.
- Van Der Heijden MGA. Arbuscular mycorrhizal fungi as a determinant of plant diversity: in search of underlying mechanisms and general principles. In: Mycorrhizal ecology. Springer; 2002. p. 243–65.
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef]
- Quince, C.; Lanzen, A.; Davenport, R.J.; Turnbaugh, P.J. Removing Noise From Pyrosequenced Amplicons. BMC Bioinform. 2011, 12, 38. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc a Guid to methods Appl 1990, 18, 315–322. [Google Scholar]
- Gohl, D.M.; Vangay, P.; Garbe, J.; MacLean, A.; Hauge, A.; Becker, A.; Gould, T.J.; Clayton, J.B.; Johnson, T.J.; Hunter, R.; et al. Systematic improvement of amplicon marker gene methods for increased accuracy in microbiome studies. Nat. Biotechnol. 2016, 34, 942–949. [Google Scholar] [CrossRef]
- Estaki, M.; Jiang, L.; Bokulich, N.A.; McDonald, D.; González, A.; Kosciolek, T.; Martino, C.; Zhu, Q.; Birmingham, A.; Vázquez-Baeza, Y.; et al. QIIME 2 Enables Comprehensive End-to-End Analysis of Diverse Microbiome Data and Comparative Studies with Publicly Available Data. Curr. Protoc. Bioinform. 2020, 70, e100. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Katoh K, Misawa K, Kuma KI, Miyata T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066.
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Bokulich NA, Kaehler BD, Rideout JR, Dillon M, Bolyen E, Knight R, et al. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome. 2018, In Press.
- Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, et al. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–6.
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef]
- Gower, JC. Principal coordinates analysis. Wiley StatsRef Stat Ref online. 2014, 1–7. [Google Scholar]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Mauri M, Elli T, Caviglia G, Uboldi G, Azzi M. RAWGraphs: a visualisation platform to create open outputs. In: Proceedings of the 12th biannual conference on Italian SIGCHI chapter. 2017. p. 1–5.
- Wemheuer, F.; Taylor, J.A.; Daniel, R.; Johnston, E.; Meinicke, P.; Thomas, T.; Wemheuer, B. Tax4Fun2: prediction of habitat-specific functional profiles and functional redundancy based on 16S rRNA gene sequences. Environ. Microbiome 2020, 15, 11. [Google Scholar] [CrossRef]
- Põlme, S.; Abarenkov, K.; Nilsson, R.H.; Lindahl, B.D.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; Baldrian, P.; et al. FungalTraits: a user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA); Wiley statsref Stat Ref online. 2014;1–15. [CrossRef]
- Foster ZSL, Weiland JE, Scagel CF, Grünwald NJ. The Composition of the Fungal and Oomycete Microbiome of Rhododendron Roots under Varying Growth Conditions, Nurseries, and Cultivars. Phytobiomes. 2020, 4, 156–164.
- Qin, Y.; Yan, Y.; Cheng, L.; Lu, Y.; Chen, J.; Liu, F.; Tan, J. Arbuscular Mycorrhizal Fungi and Rhizobium Facilitate Nitrogen and Phosphate Availability in Soybean/Maize Intercropping Systems. J. Soil Sci. Plant Nutr. 2023, 23, 2723–2731. [Google Scholar] [CrossRef]
- Liu, A.; Ku, Y.-S.; Contador, C.A.; Lam, H.-M. The Impacts of Domestication and Agricultural Practices on Legume Nutrient Acquisition Through Symbiosis With Rhizobia and Arbuscular Mycorrhizal Fungi. Front. Genet. 2020, 11, 583954. [Google Scholar] [CrossRef]
- Smith SE, Read DJ. Mycorrhizal symbiosis. Academic press; 2010.
- Zhao, B.; Jia, X.; Yu, N.; Murray, J.D.; Yi, K.; Wang, E. Microbe-dependent and independent nitrogen and phosphate acquisition and regulation in plants. New Phytol. 2023. [Google Scholar] [CrossRef]
- Akiyama, K.; Matsuzaki, K.-I.; Hayashi, H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 2005, 435, 824–827. [Google Scholar] [CrossRef]
- Bender SF, Schlaeppi K, Held A, Heijden MGA Van Der. Agriculture, Ecosystems and Environment Establishment success and crop growth e ff ects of an arbuscular mycorrhizal fungus inoculated into Swiss corn fields. Agric Ecosyst Environ. 2019, 273, (2018):13–24.
- Zahran, H.H. Rhizobium -Legume Symbiosis and Nitrogen Fixation under Severe Conditions and in an Arid Climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [CrossRef]
- Pinto AP, Faria JMS, Dordio A V, Carvalho AJP. Organic Farming–a Sustainable Option to Reduce Soil Degradation. Agroecol Approaches Sustain Soil Manag. 2023, 83–143.
- Jindo, K.; Audette, Y.; Olivares, F.L.; Canellas, L.P.; Smith, D.S.; Voroney, R.P. Biotic and abiotic effects of soil organic matter on the phytoavailable phosphorus in soils: a review. Chem. Biol. Technol. Agric. 2023, 10, 1–12. [Google Scholar] [CrossRef]
- Kiers, E.T.; Duhamel, M.; Beesetty, Y.; Mensah, J.A.; Franken, O.; Verbruggen, E.; Fellbaum, C.R.; Kowalchuk, G.A.; Hart, M.M.; Bago, A.; et al. Reciprocal Rewards Stabilize Cooperation in the Mycorrhizal Symbiosis. Science 2011, 333, 880–882. [Google Scholar] [CrossRef]
- Busby, P.E.; Soman, C.; Wagner, M.R.; Friesen, M.L.; Kremer, J.; Bennett, A.; Morsy, M.; Eisen, J.A.; Leach, J.E.; Dangl, J.L. Research priorities for harnessing plant microbiomes in sustainable agriculture. PLOS Biol. 2017, 15, e2001793. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2014, 9, 1177–1194. [Google Scholar] [CrossRef]
- Dangi SR, Allen BL, Jabro JD, Rand TA, Campbell JW, Calderon RB. The Effect of Alternative Dryland Crops on Soil Microbial Communities. Soil Systems 2023, 8, 4.
- Hu, J.; Cyle, K.T.; Miller, G.; Shi, W. Water deficits shape the microbiome of Bermudagrass roots to be Actinobacteria rich. FEMS Microbiol. Ecol. 2023, 99, fiad036. [Google Scholar] [CrossRef]
- Pan, X.; Raaijmakers, J.M.; Carrión, V.J. Importance of Bacteroidetes in host–microbe interactions and ecosystem functioning. Trends Microbiol. 2023, 31, 959–971. [Google Scholar] [CrossRef]
- Xu, S.; Wang, B.; Li, Y.; Jiang, D.; Zhou, Y.; Ding, A.; Zong, Y.; Ling, X.; Zhang, S.; Lu, H. Ubiquity, diversity, and activity of comammox Nitrospira in agricultural soils. Sci. Total. Environ. 2019, 706, 135684. [Google Scholar] [CrossRef]
- Oliverio, A.M.; Bissett, A.; McGuire, K.; Saltonstall, K.; Turner, B.L.; Fierer, N. The Role of Phosphorus Limitation in Shaping Soil Bacterial Communities and Their Metabolic Capabilities. mBio 2020, 11. [Google Scholar] [CrossRef]
- Hashmi I, Bindschedler S, Junier P. Firmicutes. In: Beneficial microbes in agro-ecology. Elsevier; 2020. p. 363–96.
- Wang, S.; Meade, A.; Lam, H.-M.; Luo, H. Evolutionary Timeline and Genomic Plasticity Underlying the Lifestyle Diversity in Rhizobiales. mSystems 2020, 5. [Google Scholar] [CrossRef]
- Idbella, M.; Bonanomi, G. Uncovering the dark side of agriculture: How land use intensity shapes soil microbiome and increases potential plant pathogens. Appl. Soil Ecol. 2023, 192, 105090. [Google Scholar] [CrossRef]
- Hestrin, R.; Kan, M.; Lafler, M.; Wollard, J.; A Kimbrel, J.; Ray, P.; Blazewicz, S.J.; Stuart, R.; Craven, K.; Firestone, M.; et al. Plant-associated fungi support bacterial resilience following water limitation. ISME J. 2022, 16, 2752–2762. [Google Scholar] [CrossRef]
- Shen, J.; Liang, Z.; Kuzyakov, Y.; Li, W.; He, Y.; Wang, C.; Xiao, Y.; Chen, K.; Sun, G.; Lei, Y. Dissolved organic matter defines microbial communities during initial soil formation after deglaciation. Sci. Total. Environ. 2023, 878, 163171. [Google Scholar] [CrossRef]
- Conradie, T.; Jacobs, K. Seasonal and Agricultural Response of Acidobacteria Present in Two Fynbos Rhizosphere Soils. Diversity 2020, 12, 277. [Google Scholar] [CrossRef]
- Buckley, D.H.; Huangyutitham, V.; Nelson, T.A.; Rumberger, A.; Thies, J.E. Diversity of Planctomycetes in Soil in Relation to Soil History and Environmental Heterogeneity. Appl. Environ. Microbiol. 2006, 72, 4522–4531. [Google Scholar] [CrossRef]
- Bonfante, P.; Venice, F. Mucoromycota: going to the roots of plant-interacting fungi. Fungal Biol. Rev. 2020, 34, 100–113. [Google Scholar] [CrossRef]
- Zhang, Q.; Guo, T.; Sheng, K.; Shi, W.; Han, Y.; Wang, Y.; Li, H. Continuous straw return for 8 years mitigates the negative effects of inorganic fertilisers on C-cycling soil bacteria. Eur. J. Soil Sci. 2022, 73, e13322. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Anand, G.; Gaur, R.; Yadav, D. Plant–microbiome interactions for sustainable agriculture: a review. Physiol. Mol. Biol. Plants 2021, 27, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Yates RJ, Abaidoo R, Howieson JG. Field experiments with rhizobia. In Australian Centre for International Agricultural Research; 2016.
Figure 1.
The influence of microbial inoculants on (a) plant grain yield and (b) microbial dependency (%) based on plant biomass and grain yield at contrasting dryland site conditions. Error bars indicate standard error of the mean from five replications. Bars with common letters are not significantly different based on Wilcoxon tests at 0.05% probability level.
Figure 1.
The influence of microbial inoculants on (a) plant grain yield and (b) microbial dependency (%) based on plant biomass and grain yield at contrasting dryland site conditions. Error bars indicate standard error of the mean from five replications. Bars with common letters are not significantly different based on Wilcoxon tests at 0.05% probability level.
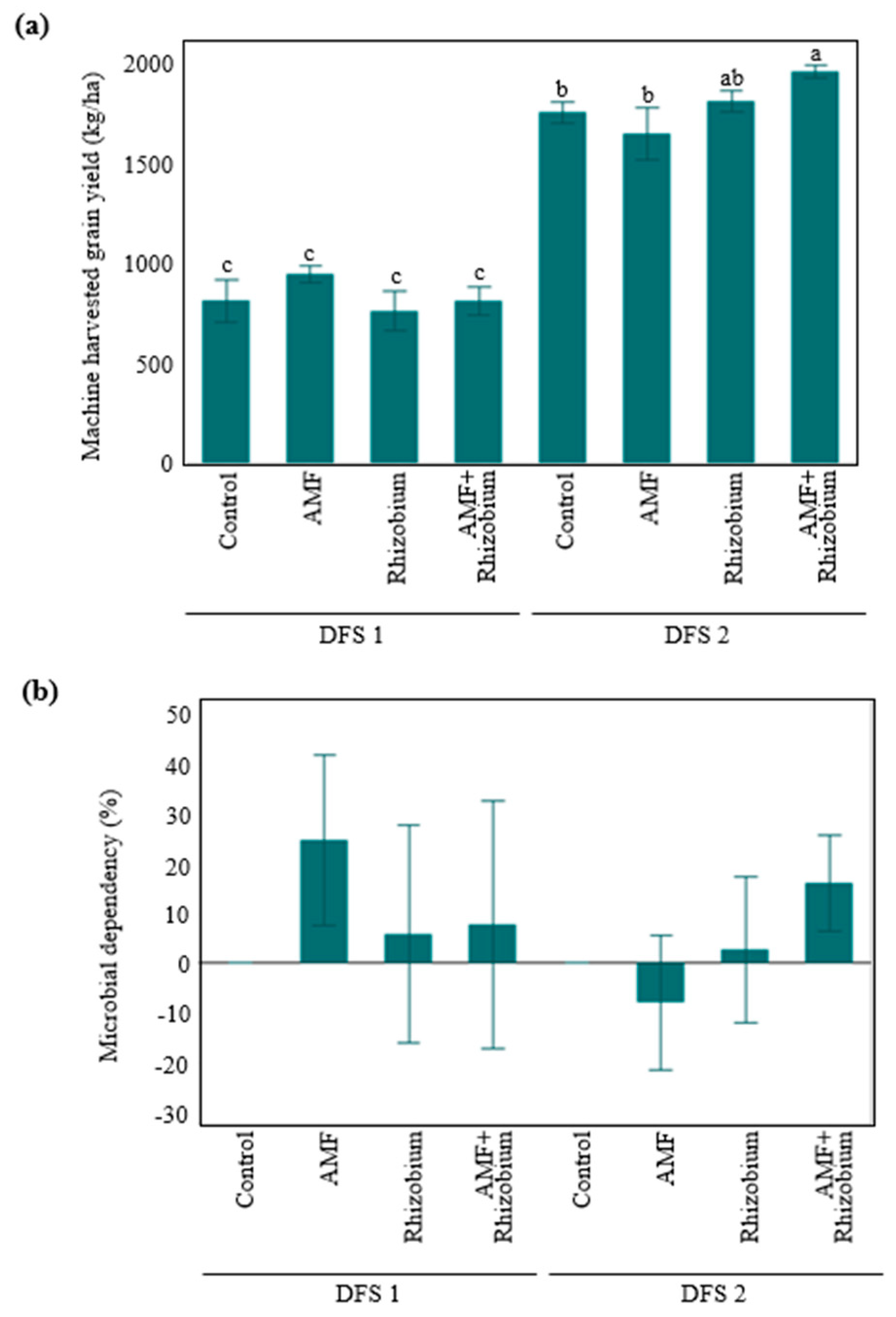
Figure 2.
The influence of microbial inoculants on grain nutrient content and plant nutrient uptake (a,b) carbon (% C). The mean plot shows the influence of microbial inoculants on soil nutrient residuals after pea cropping: (c) nitrogen residual (kg/ha), (d) plant nitrogen uptake vs soil N residual (kg/ha), (e) phosphorus residual , and (f) grain phosphorus uptake (kg/ha) vs soil P residual at contrasting dryland sites. The vertical bars in the least square means denote confidence intervals. Bars with common letter are not significantly different based on LSD tests at 0.05% probability level.
Figure 2.
The influence of microbial inoculants on grain nutrient content and plant nutrient uptake (a,b) carbon (% C). The mean plot shows the influence of microbial inoculants on soil nutrient residuals after pea cropping: (c) nitrogen residual (kg/ha), (d) plant nitrogen uptake vs soil N residual (kg/ha), (e) phosphorus residual , and (f) grain phosphorus uptake (kg/ha) vs soil P residual at contrasting dryland sites. The vertical bars in the least square means denote confidence intervals. Bars with common letter are not significantly different based on LSD tests at 0.05% probability level.
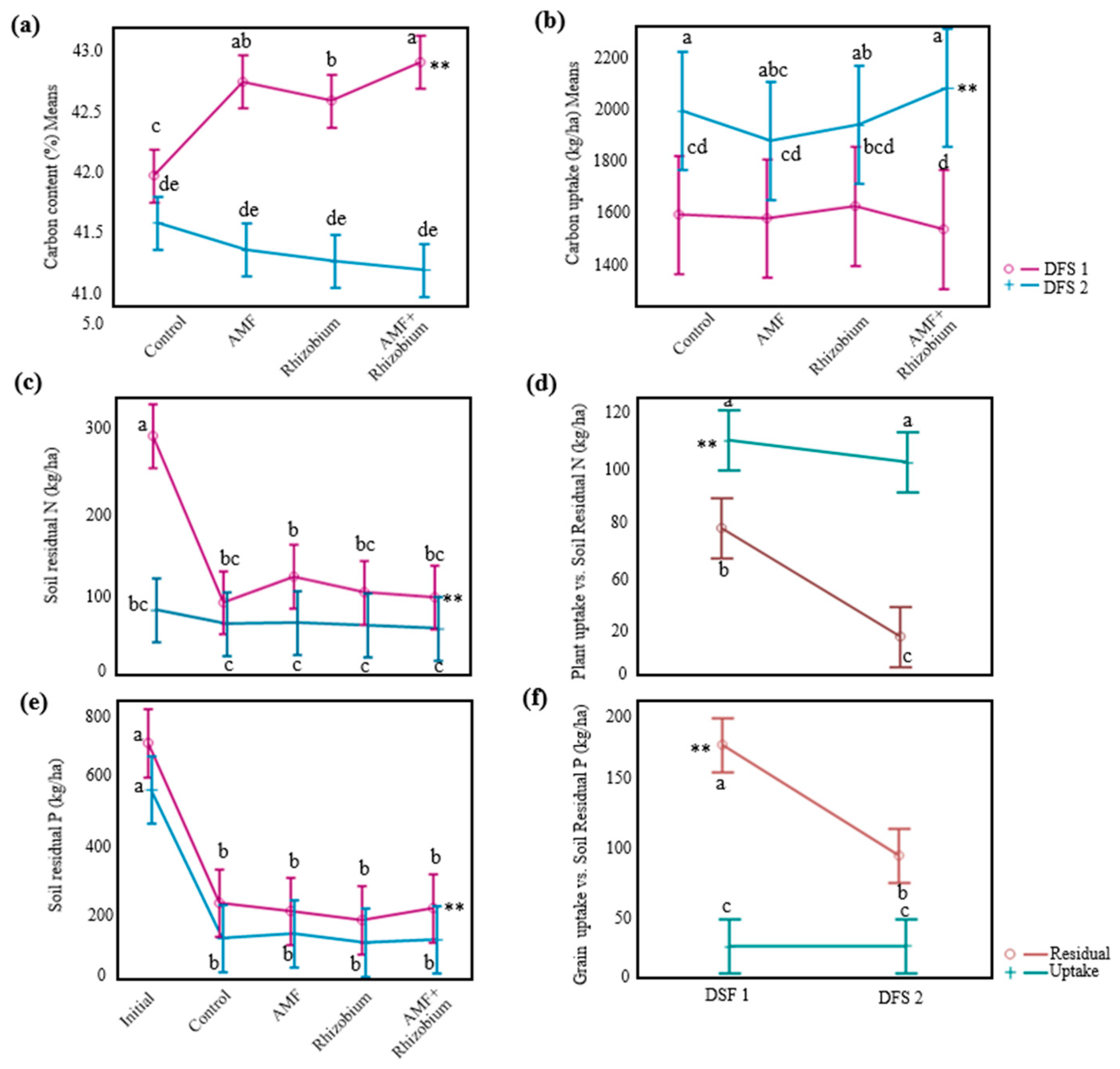
Figure 3.
Site-specific effect response to microbial inoculation on microbial diversity. Alpha-diversity of (a) bacterial and (b) fungal communities was calculated as observed number of species per sample and visualized using box-plots. Beta-diversity of microbial inoculant treatments (c,e) across sites and (d,f) between sites for bacterial and fungal communities. Beta diversity was calculated using the Bray–Curtis index and visualized using principal coordinate analysis (PCoA) ordination plots. The different groups are highlighted by ellipses showing a 95% confidence range for the variation within each group.
Figure 3.
Site-specific effect response to microbial inoculation on microbial diversity. Alpha-diversity of (a) bacterial and (b) fungal communities was calculated as observed number of species per sample and visualized using box-plots. Beta-diversity of microbial inoculant treatments (c,e) across sites and (d,f) between sites for bacterial and fungal communities. Beta diversity was calculated using the Bray–Curtis index and visualized using principal coordinate analysis (PCoA) ordination plots. The different groups are highlighted by ellipses showing a 95% confidence range for the variation within each group.
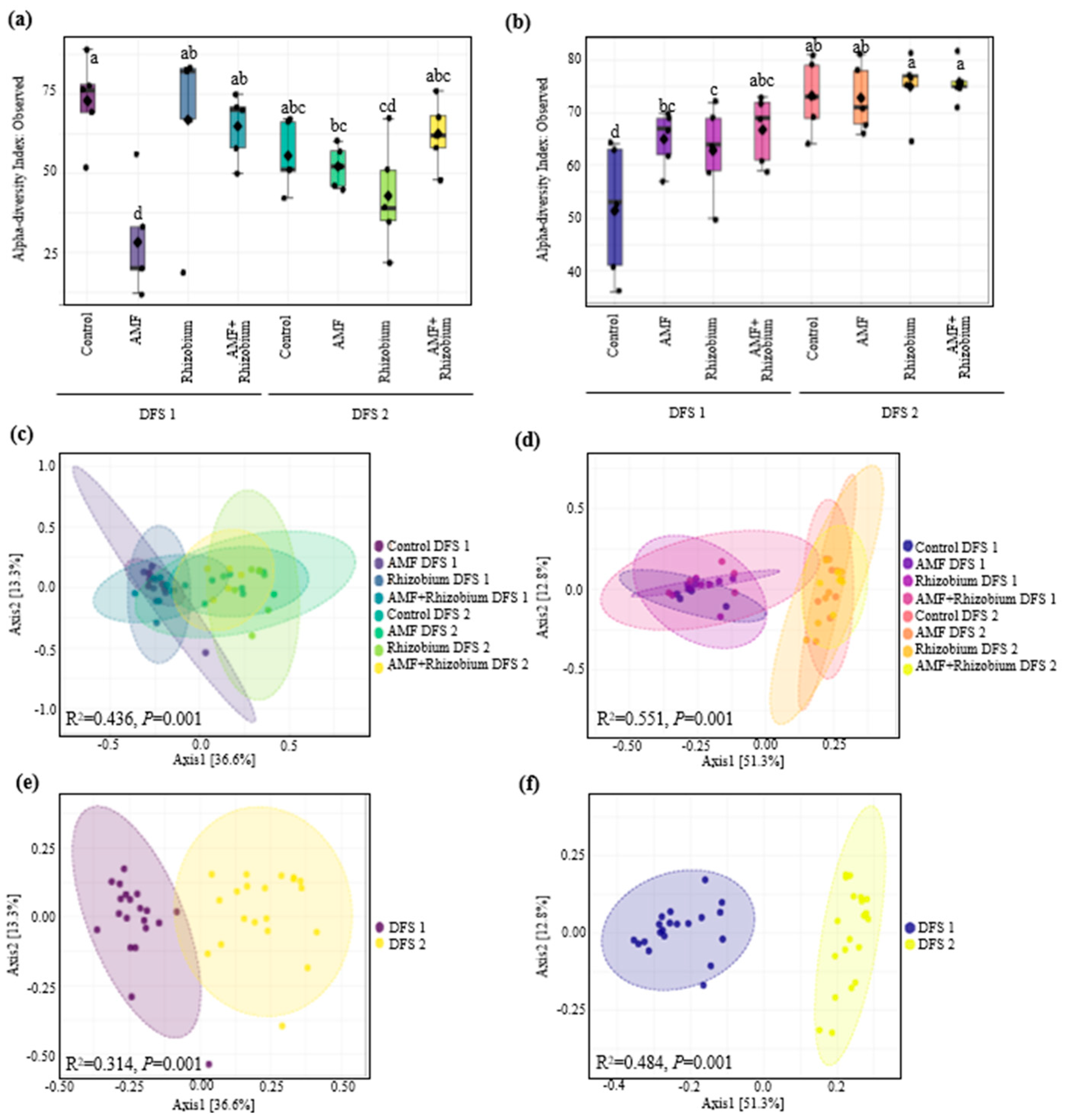
Figure 4.
Taxabar plots showing the microbial community profiles of (a) bacterial and (b) fungal communities at the phylum level. Heatmap showing the taxonomic order level composition of the (c) bacterial and (d) fungal communities in contrasting dryland field conditions.
Figure 4.
Taxabar plots showing the microbial community profiles of (a) bacterial and (b) fungal communities at the phylum level. Heatmap showing the taxonomic order level composition of the (c) bacterial and (d) fungal communities in contrasting dryland field conditions.
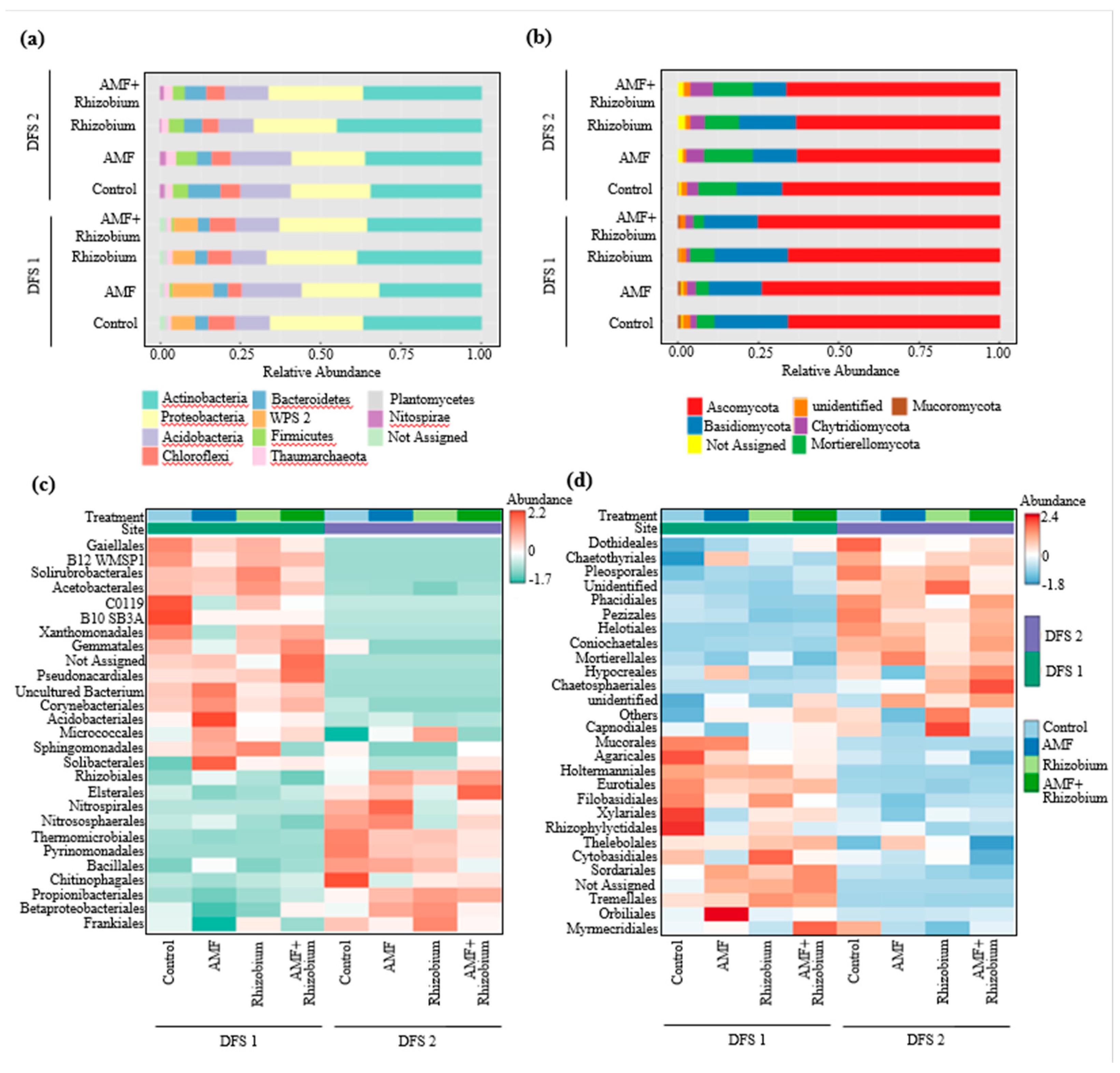
Figure 5.
The microbial communities of AMF and AMF+Rhizobium associated with increase crop performance in DFS 1 and DFS 2, respectively. The taxonomic differences between (a) AMF treated bacterial and (b) fungal communities vs the control in dryland field site 1 (DFS 1); and (c) AMF+Rhizobium treated bacterial and (d) fungal communities vs the control in DFS 2. The heat tree analysis leverages the hierarchical structure of taxonomic classifications quantitatively using the median abundance and statistically using the non-parametric Wilcoxon Rank Sum test (Foster et al., 2017).
Figure 5.
The microbial communities of AMF and AMF+Rhizobium associated with increase crop performance in DFS 1 and DFS 2, respectively. The taxonomic differences between (a) AMF treated bacterial and (b) fungal communities vs the control in dryland field site 1 (DFS 1); and (c) AMF+Rhizobium treated bacterial and (d) fungal communities vs the control in DFS 2. The heat tree analysis leverages the hierarchical structure of taxonomic classifications quantitatively using the median abundance and statistically using the non-parametric Wilcoxon Rank Sum test (Foster et al., 2017).

Figure 6.
Impact of microbial inoculants on the relative abundance of potential microbial functions (a) Functional profile of bacterial communities relative to C, N, and P nutrient cycling genes, predicted using Tax4Fun2 based on the 16S rRNA genes according to the KEGG Ortholog groups (KOs). (b) Ecophysiological functions of fungal communities relative to nutrient cycling, plant-microbe interaction, and soil health based on FungalTraits database.
Figure 6.
Impact of microbial inoculants on the relative abundance of potential microbial functions (a) Functional profile of bacterial communities relative to C, N, and P nutrient cycling genes, predicted using Tax4Fun2 based on the 16S rRNA genes according to the KEGG Ortholog groups (KOs). (b) Ecophysiological functions of fungal communities relative to nutrient cycling, plant-microbe interaction, and soil health based on FungalTraits database.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



