
Submitted:
26 January 2024
Posted:
29 January 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Advances in biological sciences have been applied in medicine, agriculture and industry with great benefit. Some recent trends in society, governance structures and communications, however, appear to be hampering the use of new scientific findings for controlling infectious diseases. These limitations are illustrated with examples related to (i) the adaptation of fresh water mosquito vectors of major arboviral diseases to salinity in coastal areas with consequences for disease transmission, (ii) understanding the implications of reduced dengue transmission during the COVID-19 lockdown for dengue control, (iii) causes underlying the rapid spread of the malaria vector Anopheles stephensi in South Asia and Africa, (iv) the application of serodiagnostic techniques for Lyme disease and tick-borne relapsing fever caused by tick-borne bacteria of the genus Borrelia, and (v) COVID-19 prevention.
Keywords:
Aedes aegypti
; Aedes albopictus
; Anopheles culicifacies
; Anopheles stephensi
; Borreliosis
; Dengue
; Climate change
; COVID-19
; Lyme disease
; Malaria
; Salinity adaptation in mosquito vectors
; Serodiagnosis
; Tick-borne relapsing fever
; Vaccines
1. Introduction
The use of other living organisms and their products for human benefit has a very ancient origin. Identification of herbal medicines, selection of wild cereal crops to form cultivated varieties, domestication of goats, sheep and cattle, cultivation of cotton, and fermentation of plant products to form alcoholic beverages are some examples [1]. However, elucidation of the structure of DNA and proteins in the middle of the 20th century spawned the science of molecular biology which has been applied in innovative ways in medicine, agriculture, and industrial production for human benefit. For example, biotechnology, defined by the European Federation of Biotechnology in 1981 as ‘the integrated use of biochemistry, microbiology and chemical engineering in order to achieve the technological application of the capacities of microbes and cultured cells’ [1], has created useful products such as monoclonal antibodies for clinical therapy of COVID-19, reagents for the laboratory diagnosis of infectious diseases [2,3,4], and the CRISPR-Cas9 gene editing technology for advancing human medicine, agriculture and animal husbandry [5].
Some societal limitations that presently have an impact on human well-being are well known. Two prominent examples are the (i) rapid expansion of human populations and improvements in their quality of life, largely attributable to science-based improvements in healthcare and food production, but leading also to the unsustainable use of the planet’s biotic and abiotic resources, and (ii) continuing high levels of greenhouse gas emissions and environmental pollution, causing many undesirable effects, including global warming. While the two factors also influence the prevalence of infectious diseases, less-well understood societal factors that limit application of recent advances in biological sciences for controlling infectious diseases have emerged. These constraints are discussed with specific examples from (i) the transmission of dengue by Aedes mosquitoes, (ii) the transmission of malaria by the urban malaria vector Anopheles stephensi, (iii) the application of serological techniques for the laboratory diagnosis of Lyme disease and tick-borne relapsing fever borreliosis, (iv) understanding immunity to SARS-CoV-2 and the delivery of COVID-19 vaccines.
2. Development of Aedes mosquito vectors in coastal brackish water and its impact on the control of dengue and other arboviral diseases
Aedes aegypti mosquitoes are the principal vectors of human arboviral diseases, including dengue, chikungunya, yellow fever and Zika, in tropical and sub-tropical countries [6,7,8]. Aedes albopictus is a secondary arboviral vector that has recently expanded its range to several temperate zone countries, partly through developing diapausing eggs able to survive winters [6,7,8]. Both Aedes species are widely regarded to lay eggs and undergo preimaginal development only in fresh water (FW) habitats containing <0.5 gL-1 salt, so that the present World Health Organization (WHO) [6,7] and US Centres for Disease Control and Prevention (CDC) [8] guidelines for larval source reduction through applying larvicides and elimination of preimaginal habitats are only directed towards fresh water habitats. Larval source reduction measures are important worldwide for controlling Aedes vectors, compared to the spraying of adulticides indoors and the use of insecticide impregnated bed nets which are effective against Anopheles mosquito vectors of malaria. This is because the two Aedes species, in contrast to the Anopheles vectors, tend to blood feed on humans outdoors during the day. Most countries follow WHO recommendations for controlling arboviral diseases, including dengue which is the most prevalent arboviral disease worldwide with 5.2 million annual cases reported to the WHO in 2019 but with a higher estimated incidence of 100 – 400 million cases worldwide [6,7,8]. The WHO recommends measures to minimize or eliminate fresh water collections where the Aedes vectors oviposit and undergo preimaginal development, the application of the organophosphate larvicide Temephos at concentrations up to 1 mgL-1 to fresh water storage containers which constitute important Aedes larval habitats in the tropics, and space spraying with adulticides to control dengue epidemics [7,9].
Aedes aegypti and Ae. albopictus were first reported to be capable of ovipositing and undergoing preimaginal development to adulthood in brackish water (BW) habitats of up to 15 gL-1 salt in the northern coastal peninsula of Jaffna in the island of Sri Lanka (Figure 1) with FW, BW and saline water defined as containing < 0.5, 0.5–30 and > 30 gL-1 salt, respectively [10,11,12,13,14,15].
The preimaginal development of Ae. aegypti and Ae. albopictus in BW has since been reported from the coastal areas of other countries including Brunei Darussalam [17], the US [18], Brazil [19], Mexico [20], Indonesia [21], and India [22]. Typical BW habitats of Ae. aegypti are BW accumulations in beach debris, fishing boats, and coastal wells, as well as discarded containers in swamps, stagnant surface drains, surface ground water and household containers in coastal areas [10,11,12,13,14,15,17,18,19,20,21,22]. Some BW collections where Ae. aegypti larvae were found in Jaffna city and the Jaffna peninsula are illustrated in Figure 2.
BW-derived Ae. aegypti and Ae. albopictus larvae in the Jaffna peninsula were more salinity-tolerant, possessing higher LC50 for salt than the corresponding FW-derived larvae from the Sri Lankan mainland as shown in Figure 3 [10,15,23]. Aedes albopictus was observed to be more salinity tolerant than Ae. aegypti at both locations (Figure 3).
Further investigations on laboratory-maintained colonies demonstrated that, although they remained reproductively compatible, BW-adapted or salinity-tolerant Ae. aegypti differed from FW Ae. aegypti significantly in several important features that are summarized in Table 1. These structural and physiological differences have been suggested to be related to the development of salinity tolerance in BW Ae. aegypti.
All the salinity-adaptive structural and molecular changes seen in Ae. aegypti are expected to be paralleled in the secondary arboviral vector Ae. albopictus, as well as other FW mosquito vectors that have adapted to salinity, e.g., the malaria vector Anopheles culicifacies discussed in section 4.
The euryhaline nature of salinity-tolerant Aedes vectors in coastal areas and their neglect in vector control programs, allows them to serve as arboviral reservoirs in coastal areas and function as bridging vectors for adjoining inland areas, thereby enhancing arboviral disease transmission and facilitating epidemics [26]. As a consequence, it is predicted that small islands and countries with long coastlines in relation to their total land area, e.g. Indonesia, Sri Lanka and the Caribbean islands, will be particularly prone to the enhanced transmission of arboviral diseases like dengue, chikungunya and Zika [27,28]. Moreover, it is expected that global warming leading to a rise in sea level will increase ground water salinisation in coastal areas, and further exacerbate arboviral disease transmission in this manner [29,30]. It is therefore important that in coastal zones throughout the world there is a need to (i) extend larval source reduction efforts also to the BW habitats of Aedes vectors, and (ii) monitor the efficacy of widely-used larvicides in coastal areas [15].
Dengue is the most prevalent arboviral disease worldwide and causes >100,000 cases annually in Sri Lanka. Despite overwhelming published evidence for more than a decade following the first demonstration of salinity tolerant Aedes vectors in Sri Lanka in 2011 [10], the country’s Ministry of Health has only recently begun to target BW collections in coastal areas in its dengue control program. However, the WHO and CDC have yet to modify their guidelines for controlling dengue and other arboviral diseases, which continue to focus solely on FW habitats of the Aedes vectors [6,7,8,9].
3. Understanding the implications for dengue control of the reduced Aedes vector densities and dengue incidence observed during the COVID-19 lockdown
Public health measures that severely curtailed the movement of people to reduce COVID-19 transmission during the period from 2020 to 2022, termed lockdown, were widely expected to adversely affect mosquito control programs and increase the incidence of diseases like malaria and dengue in endemic countries [31]. A significant increase in the incidence and mortality from malaria during the COVID-19 lockdown was indeed reported in Zimbabwe [32]. A small increase in dengue incidence was also reported from Singapore [33] which is a country that employs stringent larval source reduction measures in both public places and residences to control dengue transmission.
However, the incidence of dengue incidence markedly decreased in all districts of Sri Lanka during the COVID-19 lockdown [34,35,36,37]. An 89% reduction in the number of predicted dengue cases during the most drastic lockdown period from March 2020 to April 2021 in the northern Jaffna district (Figure 4) was also accompanied by an 89% drop in Aedes larvae collected from ovitraps in Jaffna city [34,35].
Dynamics of disease transmission by vector mosquitoes for a non-immune population is described by the Ross–MacDonald equation [30,38] which can be usefully applied to dengue transmission in the present context. This equation relates the number of secondary infections generated from a single infected person (R0) to vector parameters as follows:
Ro = ma2αβpn/ r[−loge (p)]
m = ratio of the number of vector mosquitoes to the number of humans;
a = average number of human blood meals taken by a mosquito in a day;
α = probability of transmission of pathogen from an infected human to a biting mosquito;
β = probability of transmission of a pathogen from an infected mosquito to a non-immune human during feeding;
p = daily probability of survival of the mosquito vector;
n = duration in days from infection of a biting mosquito until the mosquito becomes capable of infecting humans after the pathogen undergoes obligatory development in the mosquito, also termed the extrinsic incubation period;
r = recovery rate in humans (inverse of the average duration of infectiousness in days).
The anthropophagic Ae. aegypti and partly anthropophagic Ae. albopictus vectors are daytime feeders [6,7,8,9,10,39] that are highly prevalent in premises of schools, hospitals, government offices, transport hubs, and factories in Sri Lanka [14,40]. The closure of schools and offices, as well other forms of restrictions on the movement of people outside of residences, can therefore be expected to reduce blood feeding by the two vectors, particularly Ae. aegypti, leading to a reduction in ‘a’ in the equation. Any reduction in blood meals will result in reduced oviposition and decreased vector densities, manifesting as reduced values of ‘m’ and ‘p’. Lower values for ‘a’, ‘m’, and ‘p’ will diminish R0, which will be particularly impacted by its exponential relationship with ‘a’ and ‘p’. R0 is directly related to the rate of dengue transmission and therefore the number of dengue cases in the population. Reduced oviposition as a result of decreased blood feeding is consistent with diminished larval collections from ovitraps in Jaffna city [34,35], which was also independently observed in the western Colombo district [36], during the lockdown in Sri Lanka.
In particular, diminished access of Aedes vectors to blood meals in public places like schools, transport hubs, shops, and temples during the COVID-19 lockdown were postulated to reduce vector densities and vector survival and therefore dengue transmission [34,35]. Additionally, infection of uninfected Aedes vectors through feeding on infected persons, and infection of uninfected persons by infected Aedes vectors in new locations are both promoted by population mobility, and could have contributed to reduced dengue incidence during the COVID-19 lockdown. Dengue and malaria incidences fell by 75% and 46% respectively during the 2020 lockdown in India [41], a country with similar dengue epidemiology and mosquito vector control programmes to Sri Lanka. Statistics for malaria were not available for Sri Lanka because malaria had been eliminated from the island in 2013 [42]. The early expectation of epidemiologists, held also by the WHO, that the incidence of dengue and malaria will increase due to a collapse of vector control programmes during the COVID-19 pandemic [31], was therefore not borne out in India and Sri Lanka. From the perspective of more effective dengue control in Sri Lanka, India and other countries with similar dengue epidemiology, it would appear that this early prevalent view [31] detracts from experimental findings [34,35] which strongly suggest that dengue control measures involving Aedes vector larval source reduction need to be more stringently applied in public places where people congregate.
4. Adaptation of fresh water Anopheles malaria vectors to salinity and its consequences for malaria control
Some salinity-tolerant Anopheles that transmit malaria parasites in coastal areas have evolved into distinct species as a result of their reproductive isolation. These include An, farauti in Australia and neighbouring countries, An. sundaicus in Asia, An. melas and An. merus in Africa, as well as An. aquasalis and An. albimanus in the Americas [28,29,30]. It was recently observed for the first time that An. culicifacies, the principal rural FW vector of malaria in South Asia, can also lay eggs and undergo preimaginal development in BW of up to 4 gL-1 salt in coastal swamps of eastern Sri Lanka [43] and the Jaffna peninsula [44,45]. It is possible that An. culicifacies may similarly develop in coastal BW habitats of other South and Southeast Asian countries. Larval source reduction was an important component of efforts to control or eradicate malaria in the past no longer considered to be the case by the WHO [46]. However, this position needs to be reconsidered, and the performance of further vector surveys supported, because global warming leading to a rise in sea levels can increase brackish water habitats for anopheline malaria vectors in coastal areas [28,29,30].
Anopheles stephensi, the principal urban vector of malaria in South Asia, was first detected in Sri Lanka as having arrived from India to Mannar island (Figure 1) in 2017 [47]. Anopheles stephensi was later observed to develop in BW of up to 3.5 gL-1 in cement water storage tanks and domestic wells in different locations within Jaffna city (Figure 1) as well as Mannar island [16,47,48]. It postulated that its adaptation to similar BW habitats in in neighbouring coast of Tamil Nadu state in South India, and the movement of refugees and combatants in boats during the 1983-2009 civil war in Sri Lanka, as well as monsoonal winds across the 64 -137 km-wide separating Palk strait (Figure 1) may have facilitated this expansion in the range of An. stephensi [45,48]. Anopheles stephensi also recently expanded its range westwards to the Arabian Peninsula and after that to Northeast Africa with coastal zones playing a prominent role in this process [46,48]. Further expansion of the range of An. stephensi to West and East Africa was confirmed in 2023 [46]. It seems reasonable to postulate that, as in Sri Lanka [48], civil wars in Yemen, Eritrea, Ethiopia, Sudan and Somalia that were accompanied by the collapse of societal infrastructure, governments and vector control programs, as well as the movement of refugees, had a major role in the spread of An. stephensi and the consequent increased malaria transmission in the Arabian Peninsula and Africa [46].
5. Laboratory tests supporting clinical diagnosis of Lyme disease and tick-borne relapsing fever
The pathogen causing Lyme disease (LD) was first identified in ticks in 1982, and then in human patients in the US in 1983, to be a spirochete bacterium later named Borrelia burgdorferi. Borrelia burgdorferi sensu stricto (Bbss) is the principal species responsible for LD in the US [4,49]. Other species of the genus Borrelia termed Lyme Disease Borreliae (LDB), or alternatively Borrelia burgdorferi sensu lato, are now known to cause LD in many temperate zone countries [4,49]. Some clinical manifestations of LD are shared with tick-borne relapsing fever (TBRF) caused by a different group of Borrelia species termed relapsing fever Borreliae (RFB) [4,50], and indeed non-borrelial tick-borne diseases [51]. TBRF is prevalent in both tropical and temperate climates in many parts of the world [4,50].
In the absence of distinct pathognomonic features, other than erythema migrans at the site of a tick bite which often goes unnoticed, a diagnosis of LD relies heavily on laboratory confirmation of infection. Because LDB are often present at low concentrations in blood, the detection of antibodies in patient sera has become the preferred laboratory diagnostic method [4,52]. The recommendation from the US National Conference on Serologic Diagnosis of Lyme Disease meeting in 1994 of a standard two-tier test (STTT) for the serological diagnosis of LD was an important diagnostic advance [52]. The first tier of the STTT is an enzyme immunoassay (EIA) or immunofluorescence assay (IFA) on whole Bbss cell antigens, followed by a second-tier confirmatory western blot (WB) on whole Bbss cell lysates for sera that give positive or equivocal results in the first-tier test [52]. The rationale for the STTT was that the first tier EIA or IFA was highly sensitive but inadequately specific, while the second tier WB was highly specific for detecting serum antibodies to LDB [52]. Drawbacks to STTTs include the need for a cumbersome WB procedure and variable sensitivity for detecting infections with LDB species other than Bbss, both of which can be overcome by using relevant recombinant proteins as target antigens [4]. As a result, the CDC has been approving modified two-tier tests (MTTTs) amenable to machine reading of results in the US since 2019 [53].
Line immunoblot tests (IBs) based on purified recombinant proteins applied as lines on blotting media strips have been used to support the diagnosis of LD in European laboratories from the beginning of the 21st century [4,54]. LD IB tests used potential single tier tests had comparable sensitivity and specificity to STTTs and were also able to detect antibodies against common European LDB species [54]. LD IBs tests have recently been developed together with analogous TBRF IB tests that detect RFB [4,55]. The use of TBRF IBs to test patients identified with LD-like symptoms in Ukraine, Australia and the US strongly suggested the many of them had TBRF and not LD [4,55]. The parallel use of LD and TBRF IBs, as illustrated in Figure 5, can therefore be expected to be more reliable for diagnosing tick-borne borreliosis worldwide [4,55].
The CDC estimated, based on commercial insurance claims data, that approximately 476,000 persons were treated every year for LD in the US during the period 2010-2018 [56]. The number LD cases in western Europe was estimated to be >200,000 per year and increasing every year [57]. The CDC reported 483 cases of TBRF during the period 1990-2011 in the US mainly confined to the western states [50], contrasting with other findings that suggest a greater incidence and wider distribution within the US [55]. Limited discourse within the relevant scientific communities and many myths about LD propagated by the print, broadcast and internet media [58], appear to be shortcomings that need to be overcome in the field of human borreliosis.
6. SARS-CoV-2 infectivity and COVID-19 vaccines
The WHO estimated that 774 million cases of COVID-19 had caused 7 million deaths worldwide up to the end of 2023 [59]. Vaccines against COVID-19 were rapidly developed after COVID-19 was first identified to provide the urgently needed initial immunity in the global population to combat the pandemic. One of the first to complete clinical trials was the University of Oxford-Astra Zeneca ChAdOx1 nCoV-19 vaccine utilising a recombinant chimpanzee adenovirus vector expressing the spike protein of SARS-CoV-2 as an immunogen [60]. This vaccine was offered for use on a not-for-profit basis for the duration of the pandemic, and also licensed to be manufactured inexpensively by the Serum Institute of India as a product named Covishield.
During the first year of COVID-19 vaccination from December 2020 to December 2021, 55.9% of the global population was estimated to have received one dose of a vaccine, 45.5% two doses, and 4.3% a third booster immunisation [61]. Based on officially reported COVID-19 deaths, 14·4 million (95% confidence interval 13·7–15·9), and alternatively the more reliable excess mortality data, 19·8 million (95% interval 19·1–20·4) deaths are estimated to have been averted by vaccination in 185 countries over this one-year period [61]. Similar findings were reported at the country level from the US and Israel [62,63]. Vaccination ameliorated the serious effects of the spread SARS-CoV2 infections among persons who had not previously had COVID-19, serving to enhance population immunity that minimized severe disease and mortality. Thus, COVID vaccines had a major beneficial impact on the early course of the pandemic and they continue to be used to boost immunity in elderly and immunocompromised persons at the present time. More lives would have been saved if resource-constrained countries had better early access to vaccines [61], some governments had not delayed beginning vaccinations or been more transparent about the local COVID-19 situation, and more people had not resisted receiving the vaccine due to cultural perceptions and misconceptions spread through the print, broadcast and internet media [64,65].
Increased susceptibility to infection by SARS-CoV-2 and consequent higher mortality during the temperate zone winter in persons with an ancestry in warm tropical countries, was widely attributed to socio-economic factors but not a likely weaker early immune response in the upper respiratory tract for which there is now increasing evidence [66,67,68,69,70]. Timely recognition of this could have led to the implementation of appropriate public health and clinical measures that might have reduced infections, morbidity and mortality during the COVID-19 pandemic among vulnerable immigrant communities living in temperate zone countries.
7. Conclusions
Some of the shortcomings in society that appear to negatively impact the control of COVID-19 and the vector-borne infectious diseases Lyme disease, tick-borne relapsing fever, malaria and dengue have been highlighted in this article. The identified drawbacks affect morbidity and mortality due to these diseases. Societal factors may similarly impact other infectious diseases as well. It is possible that more extensive investigations are needed to fully evaluate the relevant scientific findings cited here, but there appears to be little support for this. Governments as well as national and international organizations with relevant responsibilities need to recognize the likely limitations identified here and then address them. It is encouraging in this context that the issue of misleading information in the print, broadcast and internet media, the potential dangers of advanced computer technology in facilitating such misinformation, and assigning liability to the source of misinformation, have recently achieved prominence [65].
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data supporting the conclusions of this article are included within the article.
Conflicts of Interest
The author is affiliated to IDFISH and IGeneX which develop and apply diagnostic techniques for tick-borne and other infectious diseases.
Abbreviations
CDC – US Centres for Disease Control and Prevention; COVID-19—coronavirus disease 2019; EIA- enzyme immunoassay; IFA- immunofluorescence assay; LD - Lyme disease; LDB - Lyme disease Borreliae; MTTT- modified two-tier test; RFB - relapsing fever Borreliae; TBRF - tick-borne relapsing fever; SARS-CoV-2—severe acute respiratory syndrome coronavirus 2; STTT- standard two-tier test; WB - western blot; WHO—World Health Organization.
References
- Ramasamy, R. The scope of modern biotechnology. J. Natl. Sci. Found. Sri Lanka 1994, 22A, S1-S5. [CrossRef]
- Turgeon, M.L. Immunology and Serology in Laboratory Medicine, 4th ed.; Mosby Elsevier: St. Louis, MO, USA, 2009; pp.1–525.
- Shah, J.S.; Ramasamy, R. Fluorescence in situ hybridization (FISH) tests for identifying protozoan and bacterial pathogens in infectious diseases. Diagnostics 2022; 12(5):1286. [CrossRef]
- Shah, J.S.; Burrascano, J.J.; Ramasamy, R. Recombinant protein immunoblots for differential diagnosis of tick-borne relapsing fever and Lyme disease. J. Vector Borne Dis. 2023, 60(4), 353-364. [CrossRef]
- Adashi, E.Y.; Gruppuso, P.A; Cohen, I.G. CRISPR therapy of sickle cell disease: The dawning of the gene editing era. Am. J. Med. 2024, S0002-9343(23)00798-2. doi: 10.1016/j.amjmed.2023.12.018. Epub ahead of print. [CrossRef]
- World Health Organization. Fact sheet on dengue and severe dengue. Available online: https:// www. who. int/ news- room/ fact- sheets/ detail/dengue- and-severe- dengue (accessed 14 January 2024).
- World Health Organization. Dengue guidelines for diagnosis, treatment, prevention and control. Available online: https:// whqli bdoc. who. int/ publications/ 2009/ 97892 41547 871_ eng. pdf (2009a) (accessed 17 August 2023).
- Centers for Disease Control and Prevention. Dengue. Available online: https:// www. cdc. gov/ dengue/ index. Html (accessed 15 August 2023).
- World Health Organization. Temephos in drinking-water: use for vector control in drinking-water sources and containers. Available online: https://www. who. int/ docs/ defau lt- source/ wash- documents/ wash- chemicals/ temephos- background- document. pdf? sfvrsn= c34fd a71_4 (accessed 17 August 2023).
- Ramasamy, R.; Surendran, S.N.; Jude, P.J.; Dharshini, S.; Vinobaba, M. Larval development of Aedes aegypti and Aedes albopictus in peri-urban brackish water and its implications for transmission of arboviral diseases. PLoS. Negl. Trop. Dis. 2011, 5, e1369. [CrossRef]
- Surendran, S.N.; Jude, P.J.; Thabothiny, V.; Raveendran, S.; Ramasamy, R. Preimaginal development of Aedes aegypti in brackish and fresh water urban domestic wells in Sri Lanka. J. Vector Ecol. 2012, 37(2), 471–473. [CrossRef]
- Jude, P.J.; Tharmasegaram, T.; Sivasubramaniyam, G.; Senthilnanthanan, M.; Kannathasan, S.; Raveendran, S.; Ramasamy, R.; Surendran, S.N. Salinity-tolerant larvae of mosquito vectors in the tropical coast of Jaffna, Sri Lanka and the effect of salinity on the toxicity of Bacillus thuringiensis to Aedes aegypti larvae. Parasit. Vectors 2012, 5, 269. [CrossRef]
- Surendran, S.N.; Jayadas, T.T.P.; Sivabalakrishnan, K.; Santhirasegaram, S.; Karvannan, K.; Weerarathne, T.C.; Karunaratne, S.H.P.; Ramasamy, R. Development of the major arboviral vector Aedes aegypti in urban drain-water and associated pyrethroid insecticide resistance is a potential global health challenge. Parasit. Vectors 2019, 12(1), 337. [CrossRef]
- Surendran, S.N.; Jayadas, T.T.P.; Thiruchenthooran, V.; Raveendran, S.; Tharsan, A.; Santhirasegaram, S.; Sivabalakrishnan, K.; Karunakaran, S.; Ponnaiah, B.; Gomes, L.; et al. Aedes larval bionomics and implications for dengue control in the paradigmatic Jaffna peninsula, northern Sri Lanka. Parasit. Vectors 2021, 14(1), 162. Erratum in: Parasit. Vectors 2021, 14(1), 218. [CrossRef]
- Sivabalakrishnan, K.; Thanihaichelvan, M.; Tharsan, A.; Eswaramohan, T.; Ravirajan, P.; Hemphill, A.; Ramasamy, R.; Surendran, S.N. Resistance to the larvicide temephos and altered egg and larval surfaces characterize salinity-tolerant Aedes aegypti. Sci. Rep. 2023, 13(1), 8160. [CrossRef]
- Surendran, S.N.; Kesavan, L.; Jayadas, T.T.P.; Sivabalakrishnan, K.; Tharsan, A.; Liyanagedara, N.; Eswaramohan, T.; Raveendran, S.; Singh,. OP.; Ramasamy, R. Morphological and odorant-binding protein 1 gene intron 1 sequence variations in Anopheles stephensi from Jaffna city in northern Sri Lanka. Med. Vet. Entomol. 2022, 36(4), 496-502. [CrossRef]
- Idris, F.H.; Usman, A.; Surendran, S.N.; Ramasamy, R. Detection of Aedes albopictus pre-imaginal stages in brackish water habitats in Brunei Darussalam. J. Vector Ecol. 2013, 38(1), 197-199. [CrossRef]
- Yee, D.A.; Himel, E.; Reiskind, M.H.; Vamosi, S.M. Implications of saline concentrations for the performance and competitive interactions of the mosquitoes Aedes aegypti (Stegomyia aegypti) and Aedes albopictus (Stegomyia albopictus). Med. Vet. Entomol. 2014, 28(1), 60-69. [CrossRef]
- de Brito Arduino, M.; Mucci, L.F.; Serpa, L.L.; Rodrigues, Mde. M. Effect of salinity on the behavior of Aedes aegypti populations from the coast and plateau of southeastern Brazil. J. Vector Borne Dis. 2015, 52(1),79-87.
- Galavíz-Parada, J.D.; Vega-Villasante, F.; Marquetta, M. del C.; Guerrero-Galvan, S.R.; Chong, O.; Navarrete-Heredia, J.L.; Cupul-Magaña, F.G. Effect of Temperature and Salinity on the Eclosion and Survival of Aedes aegypti (L) (Diptera: Culicidae) from Western Mexico. Rev. Cuba Med. Trop. 2019, 71(2), e353.
- Ratnasari, A.; Jabal, A.R.; Syahribulan; Idris, I.; Rahma, N.; Rustam, S.N.R.N.; Karmila, M.; Hasan, H.; Wahid, I. Salinity tolerance of larvae Aedes aegypti inland and coastal habitats in Pasangkayu, West Sulawesi, Indonesia. Biodiversitas 2021, 22, 1203-1210.
- Shamna, A.K.; Vipinya, C.; Sumodan, P.K. Detection of Aedes albopictus (Diptera: Culicidae) breeding in brackish water habitats in coastal Kerala, India and its Implications for dengue scenario in the state. Int. J. Entomol. Res. 2022, 7, 214–216.
- Ramasamy, R.; Jude, P.J.; Veluppillai, T.; Eswaramohan, T.; Surendran, S.N. Biological differences between brackish and fresh water-derived Aedes aegypti from two locations in the Jaffna peninsula of Sri Lanka and the implications for arboviral disease transmission. PLoS. One 2014, 9, e104977. [CrossRef]
- Surendran, S.N.; Sivabalakrishnan, K.; Jayadas, T.T.P.; Santhirasegaram, S.; Laheetharan, A.; Senthilnanthanan, M.; Ramasamy, R. Adaptation of Aedes aegypti to salinity: Characterized by larger anal papillae in larvae. J. Vector Borne Dis. 2018. 55(3), 235-238. [CrossRef]
- Ramasamy, R.; Thiruchenthooran, V.; Jayadas, T.T.P.; Eswaramohan,T.; Santhirasegaram, S.; Sivabalakrishnan, K.; Naguleswaran, A.; Uzest, M.; Cayrol, B.; Voisin, S.N.; et al. Transcriptomic, proteomic and ultrastructural studies on salinity-tolerant Aedes aegypti in the context of rising sea levels and arboviral disease epidemiology. BMC. Genomics 2021, 22(1), 253. [CrossRef]
- Ramasamy, R.; Surendran, S.N. Mosquito vectors developing in atypical anthropogenic habitats: Global overview of recent observations, mechanisms and impact on disease transmission. J. Vector Borne Dis. 2016, 53(2), 91-98.
- Ramasamy, R. Adaptation of freshwater mosquito vectors to salinity increases arboviral disease transmission risk in the context of anthropogenic environmental changes. In: Shapshak, P.; Sinnott, J.T.; Somboonwit, C.; Kuhn, J.H. editors. Global Virology I - Identifying and Investigating Viral Diseases. New York City (NY): Springer; 2015. pp. 45–54.
- Ramasamy, R.; Surendran, S.N.; Jude, P.J.; Dharshini, S.; Vinobaba, M. Adaptation of mosquito vectors to salinity and its impact on mosquito-borne disease transmission in the South and Southeast Asian tropics. In: Morand, S.; Dujardin, J.P.; Lefait, R.; Apiwathnasorn, C. editors. Socio-Ecological Dimensions of Infectious Diseases in Southeast Asia. Singapore: Springer; 2015. pp 107–122.
- Ramasamy, R.; Surendran, S.N. Possible impact of rising sea levels on vector-borne infectious diseases. BMC. Infect. Dis. 2011, 11, 18. [CrossRef]
- Ramasamy, R.; Surendran, S. Global climate change and its potential impact on disease transmission by salinity-tolerant mosquito vectors in coastal zones. Front. Physiol. 2012, 3, 198. [CrossRef]
- Wilder-Smith, A.; Tissera, H.; Ooi, E.E.; Coloma, J.; Scott, T.W.; Gubler,. D.J. Preventing dengue epidemics during the COVID-19 pandemic. Am. J. Trop. Med. Hyg. 2020, 103(2), 570-571.
- Gavi, S.; Tapera, O.; Mberikunashe, J.; Kanyangarara, M. Malaria incidence and mortality in Zimbabwe during the COVID-19 pandemic: analysis of routine surveillance data. Malaria J. 2021, 20(1), 233. [CrossRef]
- Lim, J.T.; Dickens, B..S.L.; Chew, L.Z.X.; Choo, E..LW.; Koo,. J.R.; Aik, J.; Ng, L.C.; Cook, A.R. Impact of sars-cov-2 interventions on dengue transmission. PLoS. Negl. Trop. Dis. 2020, 14(10), e0008719.
- Surendran, S.N.; Nagulan, R.; Sivabalakrishnan, K.; Arthiyan, S.; Tharsan, A.; Jayadas, T.T.P.; Raveendran, S.; Kumanan, T.; Ramasamy, R. Reduced dengue incidence during the COVID-19 movement restrictions in Sri Lanka from March 2020 to April 2021. BMC. Public Health 2022, 22(1), 388. [CrossRef]
- Surendran, S.N.; Nagulan, R.; Tharsan, A.; Sivabalakrishnan, K.; Ramasamy, R. Dengue Incidence and Aedes Vector collections in relation to COVID-19 population mobility restrictions. Trop. Med. Infect. Dis. 2022, 7(10), 287. [CrossRef]
- Niriella, M.A.; Ediriweera, D.S.; De Silva, A.P.; Premarathna, B..H.R.; Jayasinghe, S.; de Silva, H.J. Dengue and leptospirosis infection during the coronavirus 2019 outbreak in Sri Lanka. Trans. R. Soc. Trop. Med. Hyg. 2021, 115(9), 944-946. [CrossRef]
- Ariyaratne, D.; Gomes, L.; Jayadas, T.T.P.; Kuruppu, H.; Kodituwakku, L.; Jeewandara, C.; Pannila Hetti, N.; Dheerasinghe, A.; Samaraweera, S.; Ogg, G.S.; et al. Epidemiological and virological factors determining dengue transmission in Sri Lanka during the COVID-19 pandemic. PLoS. Glob. Public Health 2022, 2(8), e0000399. [CrossRef]
- MacDonald, G. The Epidemiology and Control of Malaria. Oxford University Press: London, UK, 1957.
- Paramasivan, R.; Philip Samuel, P.; Pandian, R.S. Biting rhythm of vector mosquitoes in a rural ecosystem of South India. Int. J. Mosq. Res. 2015, 2, 106–113.
- Kusumawathie, P.H.D. Larval infestation of Aedes aegypti and Ae. albopictus in six types of institutions in a dengue transmission area in Kandy, Sri Lanka. Dengue Bull. 2005, 29, 165–168.
- Mayilsamy, M.; Vijayakumar, A.; Veeramanoharan, R.; Rajaiah, P.; Balakrishnan, V.; Kumar, A. Impact of COVID-19 lockdown during 2020 on the occurrence of vector-borne diseases in India. J. Vector Borne Dis. 2023, 60(2), 207-210. [CrossRef]
- Wijesundere, D.A.; Ramasamy, R. Analysis of historical trends and recent elimination of malaria from Sri Lanka and its applicability for malaria control in other countries. Front. Public Health 2017, 5, 212. [CrossRef]
- Jude, P.J.; Dharshini, S.; Vinobaba, M.; Surendran, S.N.; Ramasamy, R. Anopheles culicifacies breeding in brackish waters in Sri Lanka and implications for malaria control. Malar. J. 2010, 9, 106. [CrossRef]
- Jude, P.J.; Tharmasegaram, T.; Sivasubramaniyam, G.; Senthilnanthanan, M.; Kannathasan, S.; Raveendran, S.; Ramasamy, R.; Surendran, S.N. Salinity-tolerant larvae of mosquito vectors in the tropical coast of Jaffna, Sri Lanka and the effect of salinity on the toxicity of Bacillus thuringiensis to Aedes aegypti larvae. Parasit. Vectors 2012, 5, 269. [CrossRef]
- Surendran, S.N.; Jayadas, T.T.P., Tharsan, A.; Thiruchenthooran, V.; Santhirasegaram, S.; Sivabalakrishnan, K.; Raveendran, S.; Ramasamy, R. Anopheline bionomics, insecticide resistance and transnational dispersion in the context of controlling a possible recurrence of malaria transmission in Jaffna city in northern Sri Lanka. Parasit. Vectors 2020, 13(1), 156. [CrossRef]
- World Health Organization. World Malaria Report 2023. Available online: https://www.who.int/publications/i/item/9789240086173 (accessed 15 January 2024).
- Surendran, S.N.; Sivabalakrishnan, K.; Gajapathy, K.; Arthiyan, S.; Jayadas, T.T..P; Karvannan, K.; Raveendran, S.; Karunaratne S.H.P.P.; Ramasamy, R. Genotype and biotype of invasive Anopheles stephensi in Mannar Island of Sri Lanka. Parasit. Vectors 2018, 11(1), 3. [CrossRef]
- Surendran, S.N.; Sivabalakrishnan, K.; Sivasingham, A.; Jayadas, T.T.P.; Karvannan, K.; Santhirasegaram, S.; Gajapathy, K.; Senthilnanthanan, M.; Karunaratne, S.H.P.P.; Ramasamy, R. Anthropogenic factors driving recent range expansion of the malaria vector Anopheles stephensi. Front. Public Health 2019, 7, 53. [CrossRef]
- Centers for Disease Control and Prevention. Lyme Disease. Available online: https://www.cdc.gov/lyme/index.html (accessed 15 January 2024).
- Centers for Disease Control and Prevention. Tick- and Louse-Borne Relapsing Fever. Available online: https://www.cdc.gov/relapsing-fever/index.html (accessed 15 January 2024).
- Boulanger, N.; Boyer, P.; Talagrand-Reboul, E.; Hansmann ,Y. Ticks and tick-borne diseases. Med. Mal. Infect.. 2019, 49(2), 87-97.
- Centers for Disease Control and Prevention. Recommendations for test performance and interpretation from the second national conference on serologic diagnosis of Lyme disease. MMWR. 1995, 44(31), 590–591.
- Mead, P.; Petersen, J.;, Hinckley, A. Updated CDC recommendation for serologic diagnosis of Lyme disease. MMWR. 2019, 68(32), 703. [CrossRef]
- Liu, S.; Cruz, I.D.; Ramos, C.C.; Taleon, P.; Ramasamy, R.; Shah, J. Pilot study of immunoblots with recombinant Borrelia burgdorferi antigens for laboratory diagnosis of Lyme disease. Healthcare (Basel) 2018, 6(3), 99. [CrossRef]
- Shah, J.S.; Liu, S.; Du Cruz, I.; Poruri, A.; Maynard, R.; Shkilna, M.; Korda, M.; Klishch, I.; Zaporozhan, S.; Shtokailo, K.; et al. Line immunoblot assay for tick-borne relapsing fever and findings in patient sera from Australia, Ukraine and the USA. Healthcare (Basel) 2019, 7(4), 121. [CrossRef]
- Kugeler, K.J.; Schwartz, A.M.; Delorey, M.J.; Mead, P.S.; Hinckley, A.F. Estimating the frequency of Lyme disease diagnoses, United States, 2010-2018. Emerg. Infect. Dis. 2021, 27(2), 616-619. [CrossRef]
- Marques, A.R.; Strle, F.; Wormser, G.P. Comparison of Lyme disease in the United States and Europe. Emerg. Infect. Dis. 2021, 27(8), 2017-2024. [CrossRef]
- Glauser, W. Combatting Lyme disease myths and the "chronic Lyme industry". CMAJ 2019, 191(40), E1111-E1112. Erratum in: CMAJ 2019, 191(50), E1389.
- World Health Organization. Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed 17 January 2024).
- Ramasamy, M.N.; Minassian, A.M.; Ewer, K.J.; Flaxman, A.L.; Folegatti,. PM.; Owens, D.R.; Voysey, M.; Aley, P.K.; Angus, B.; Babbage, G.; et al. Safety and immunogenicity of ChAdOx1 nCoV-19 vaccine administered in a prime-boost regimen in young and old adults (COV002): a single-blind, randomised, controlled, phase 2/3 trial. Lancet 2021, 396(10267), 1979-1993.
- Watson, O.J.; Barnsley, G.; Toor, J.; Hogan, A.B.; Winskill, P.; Ghani, A.C. Global impact of the first year of COVID-19 vaccination: a mathematical modelling study. Lancet Infect. Dis. 2022, 22(9), 1293-1302. [CrossRef]
- Yamana, T.K.; Galanti, M.; Pei, S.; Di Fusco, M.; Angulo, F.J.; Moran, M.M.; Khan, F.; Swerdlow, D.L.; Shaman, J. The impact of.
- COVID-19 vaccination in the US: Averted burden of SARS-CoV-2-related cases, hospitalizations and deaths. PLoS ONE 2023, 18, e0275699.
- Haas, E.J.; McLaughlin, J.M.; Khan, F.; Angulo, F.J.; Anis, E.; Lipsitch, M.; et al. Infections, hospitalisations, and deaths averted via a nationwide vaccination campaign using the Pfizer–BioNTech BNT162b2 mRNA COVID-19 vaccine in Israel: A retrospective surveillance study. Lancet Infect. Dis. 2022, 22, 357–366.
- Bonnevie E, Goldbarg J, Gallegos-Jeffrey AK, Rosenberg SD, Wartella E, Smyser J. Content themes and influential voices within vaccine opposition on Twitter, 2019. Am. J. Public Health 2020, 110(S3), S326-S330. [CrossRef]
- Nature Editorial. How online misinformation exploits 'information voids' - and what to do about it. Nature 2024, 625(7994), 215-216. [CrossRef]
- Ramasamy, R. Nasal conditioning of inspired air, innate immunity in the respiratory tract and SARS-CoV-2 infectivity. Available online: https://doi.org/10.31219/osf.io/4j95b (accessed 14 January 2024).
- Ramasamy, R. Perspective of the relationship between the susceptibility to initial SARS-CoV-2 infectivity and optimal nasal conditioning of inhaled air. Int. J. Mol .Sci. 2021, 22(15), 7919.
- Ramasamy R. Innate and adaptive immune responses in the upper respiratory tract and the infectivity of SARS-CoV-2. Viruses 2022, 14(5), 933.
- Aquino, Y.; Bisiaux, A.; Li, Z.; O'Neill, M.; Mendoza-Revilla, J.; Merkling, S.H.; Kerner, G.; Hasan, M.; Libri, V.; Bondet, V.; et al. Dissecting human population variation in single-cell responses to SARS-CoV-2. Nature 2023, 621(7977), 120-128.
- Ramasamy R. COVID-19 vaccines for optimizing immunity in the upper respiratory tract. Viruses 2023, 15(11), 2203. [CrossRef]
Figure 1.
(a) Map showing the location of Sri Lanka in relation to South India; (b) the relative locations of the Jaffna peninsula, Mannar island and Jaffna city within Sri Lanka; (c) sites within Jaffna city where Anopheles stephensi (discussed in section 4) larvae are found. Reproduced with permission under the creative commons licence from [16].
Figure 1.
(a) Map showing the location of Sri Lanka in relation to South India; (b) the relative locations of the Jaffna peninsula, Mannar island and Jaffna city within Sri Lanka; (c) sites within Jaffna city where Anopheles stephensi (discussed in section 4) larvae are found. Reproduced with permission under the creative commons licence from [16].
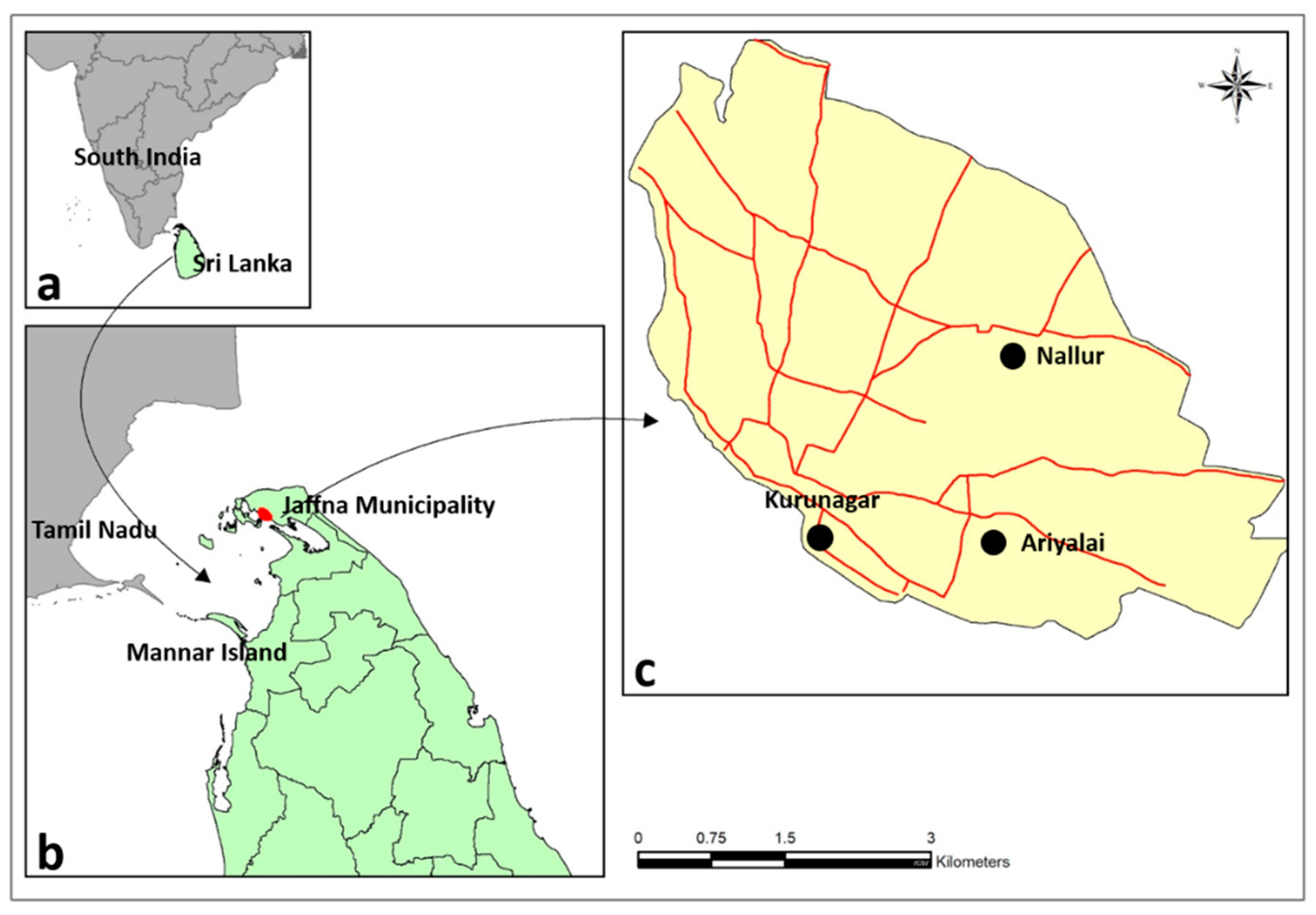
Figure 2.
BW habitats of Ae. aegypti in the Jaffna peninsula. Photographs show the brackish water collections containing larvae in: A & B - disused boats; C & E: abandoned wells; D & F: discarded food and beverage containers. Reproduced with permission under the creative commons licence from [10].
Figure 2.
BW habitats of Ae. aegypti in the Jaffna peninsula. Photographs show the brackish water collections containing larvae in: A & B - disused boats; C & E: abandoned wells; D & F: discarded food and beverage containers. Reproduced with permission under the creative commons licence from [10].

Figure 3.
Effect salinity on Ae. aegypti and Ae. albopictus from (A) coastal Jaffna peninsula and (B) mainland Sri Lanka on the L1and L3 larvae to adult transformation. L1 and L3 refer to first and third instar larvae respectively. Ppt – parts per thousand or gL-1. Reproduced with permission under the creative commons licence from [10].
Figure 3.
Effect salinity on Ae. aegypti and Ae. albopictus from (A) coastal Jaffna peninsula and (B) mainland Sri Lanka on the L1and L3 larvae to adult transformation. L1 and L3 refer to first and third instar larvae respectively. Ppt – parts per thousand or gL-1. Reproduced with permission under the creative commons licence from [10].
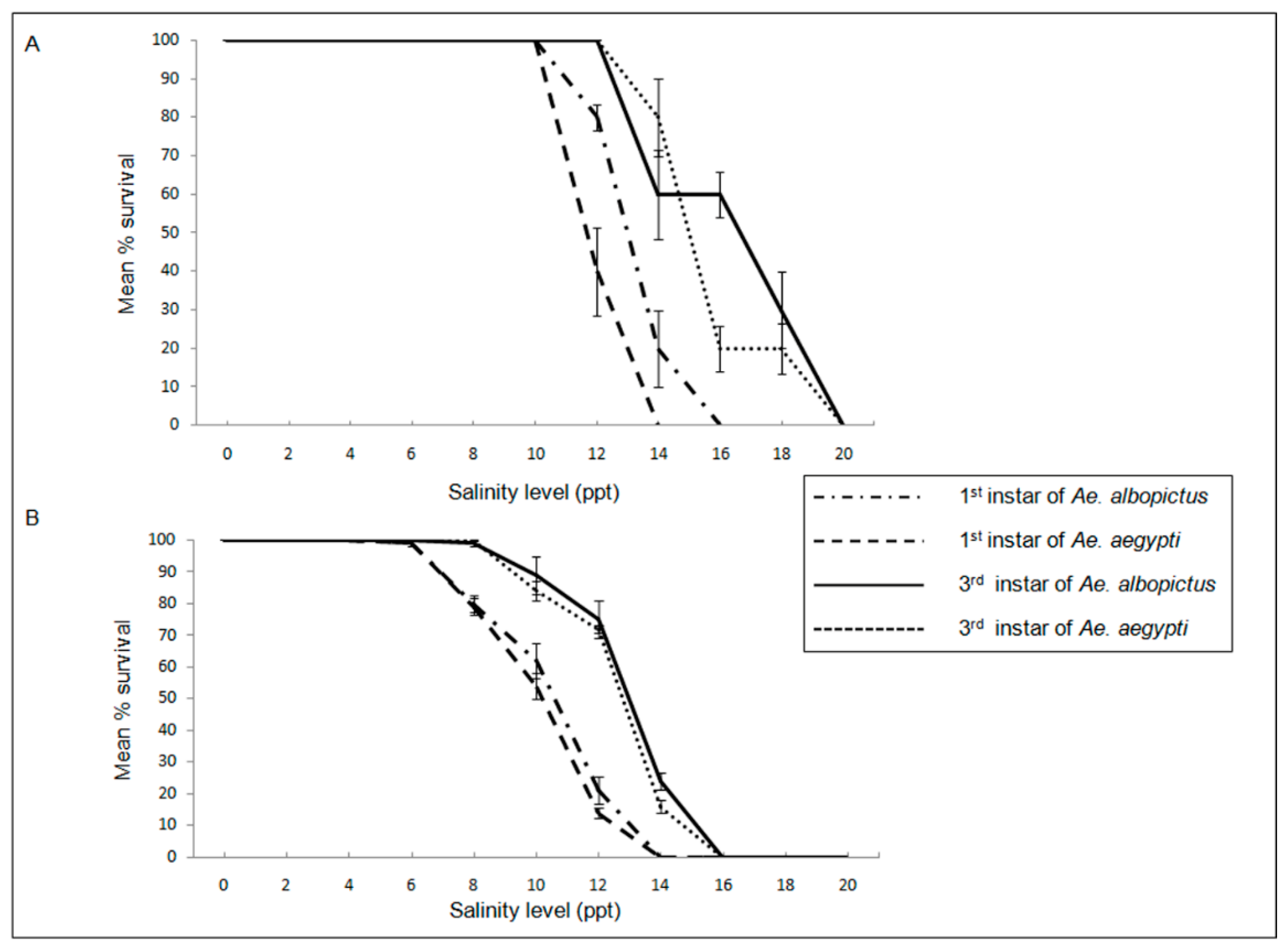
Figure 4.
Numbers of actual and predicted dengue cases in the Jaffna district during COVID-19 movement restrictions. Severe restrictions on movement of people were in place from 1 March 2020 to 30 April 2021 (period A) to lower COVID-19 transmission. These were variably eased and reimposed from 1 May 2021 until 22 November 2021 (period B), but removed altogether with the full opening of all schools from 22 November 2021 onward (period C). Reproduced with permission under the creative commons licence from [35].
Figure 4.
Numbers of actual and predicted dengue cases in the Jaffna district during COVID-19 movement restrictions. Severe restrictions on movement of people were in place from 1 March 2020 to 30 April 2021 (period A) to lower COVID-19 transmission. These were variably eased and reimposed from 1 May 2021 until 22 November 2021 (period B), but removed altogether with the full opening of all schools from 22 November 2021 onward (period C). Reproduced with permission under the creative commons licence from [35].
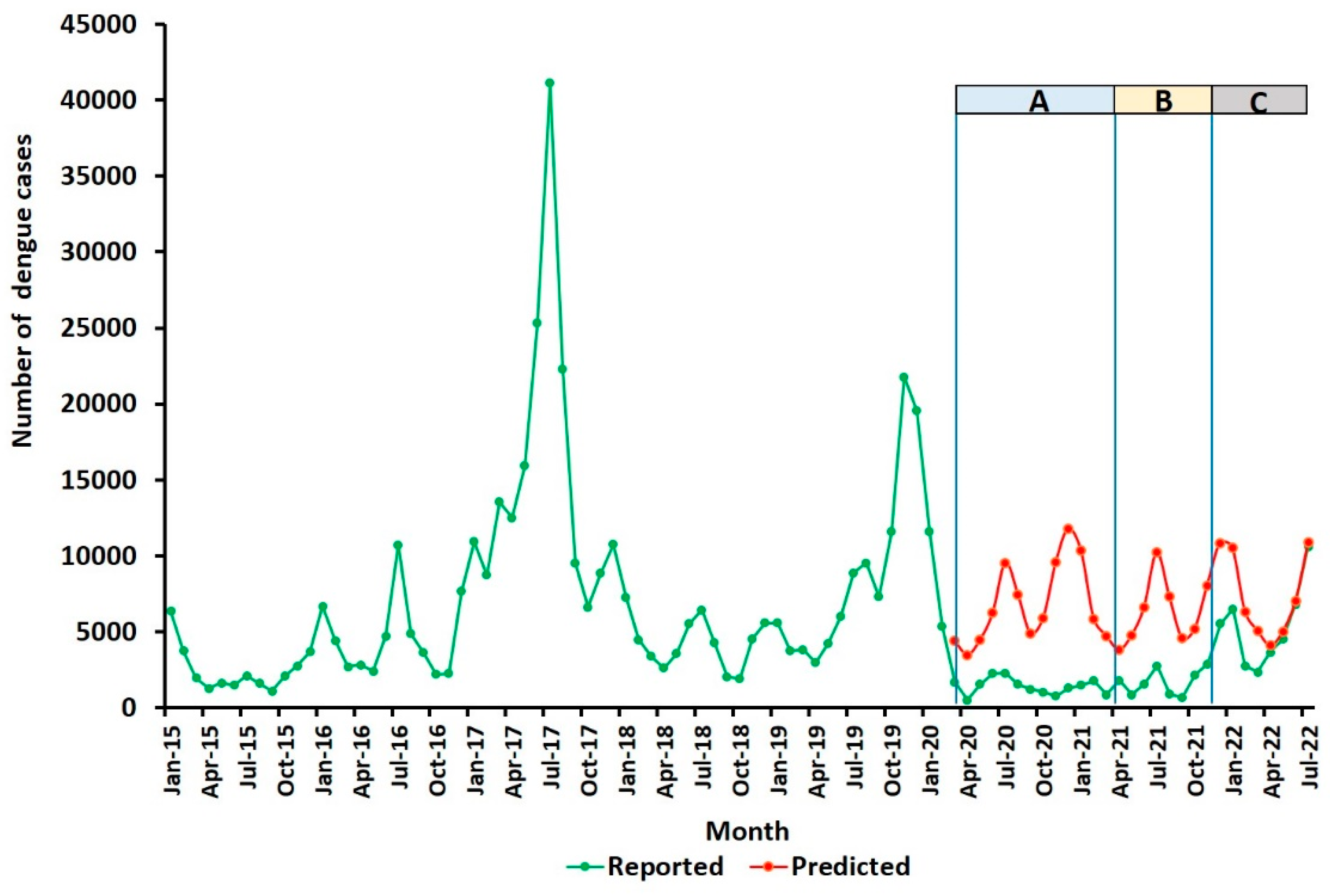
Figure 5.
Representative TBRF and LD IgM and IgG IBs with five sera from patients with LD-like symptoms. Sera of both patients in lanes 1 and 2 were only positive in TBRF IBs. Serum in lane 3 was positive in both TBRF and LD IBs. Serum in lane 4 was negative in both IBs. Serum in lane 5 was only positive in LD IBs. The detection of either IgM or IgG antibodies as shown in Figure 5 provides greater sensitivity than detecting either IgM or IgG alone. The different LDB and RFB antigens used in the LD and TBRF IBs are explained elsewhere [54,55]. Reproduced with permission under the creative commons licence from [55].
Figure 5.
Representative TBRF and LD IgM and IgG IBs with five sera from patients with LD-like symptoms. Sera of both patients in lanes 1 and 2 were only positive in TBRF IBs. Serum in lane 3 was positive in both TBRF and LD IBs. Serum in lane 4 was negative in both IBs. Serum in lane 5 was only positive in LD IBs. The detection of either IgM or IgG antibodies as shown in Figure 5 provides greater sensitivity than detecting either IgM or IgG alone. The different LDB and RFB antigens used in the LD and TBRF IBs are explained elsewhere [54,55]. Reproduced with permission under the creative commons licence from [55].
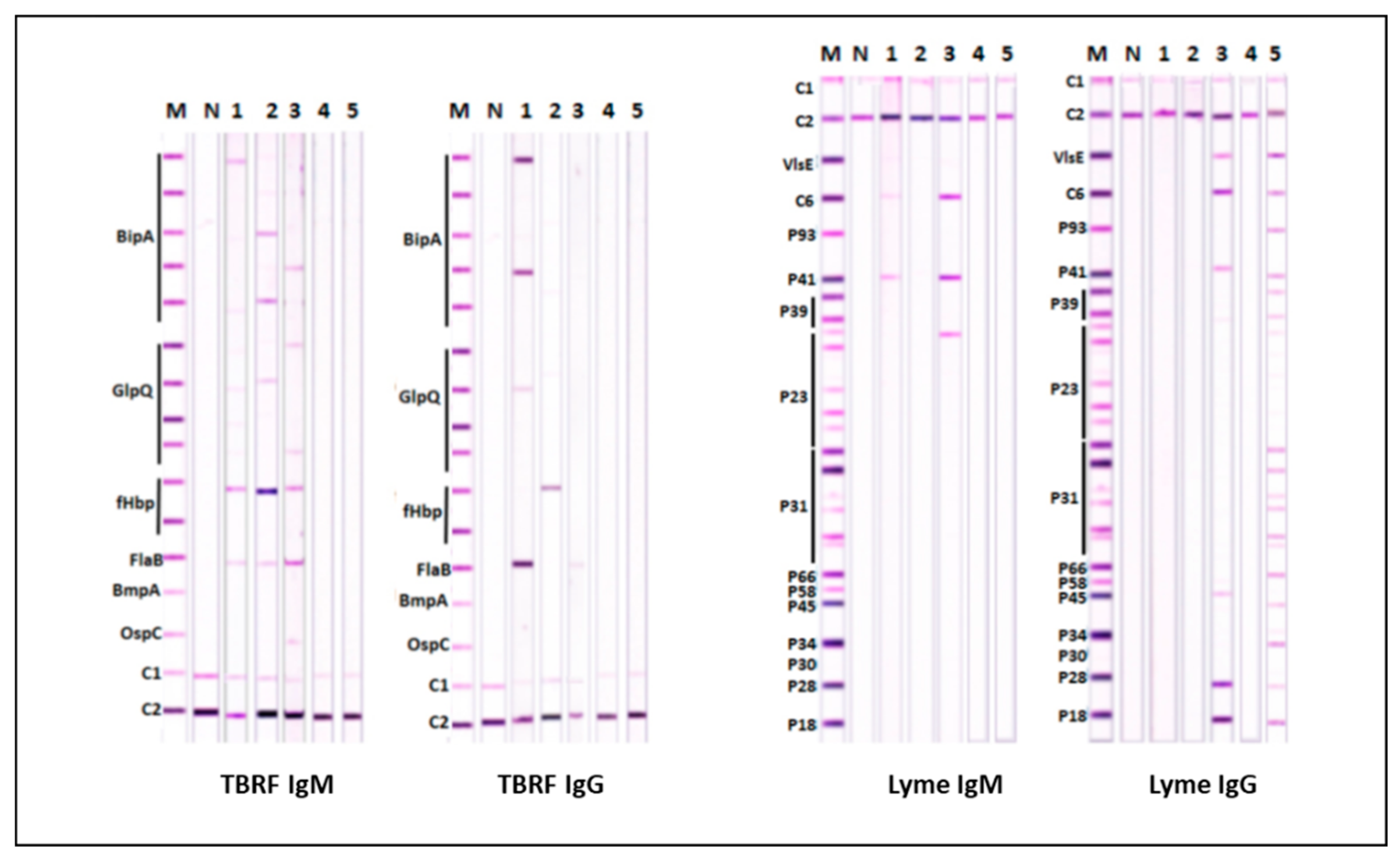

Table 1.
Differences between salinity-tolerant (BW) Ae. aegypti and FW Ae. aegypti.
| Characteristic | Differences | Cited References |
|---|---|---|
| LC50 for salt | Significantly higher LC50 in BW Ae. aegypti for the L1 and L3 to adult transition. An inheritable characteristic | [10,15,23] |
| Osmoregulatory anal papillae in L3 larvae |
Significantly larger anal papillae in BW Ae. aegypti. An inheritable characteristic | [24] |
| Gene expression in mid-L4 larvae |
Marked differences, particularly in genes for cuticle proteins, and others associated with cuticle synthesis | [25] |
| Protein composition of L4 cuticles | Marked differences compatible with the gene expression data | [25] |
| Cuticle structure by TEM | Thicker cuticles in L4 larvae and adult abdomen with more prominent endocuticles and exocuticles in BW Ae. aegypti | [25] |
| Surfaces of shed L3 and L4 cuticles | More pronounced surface undulations in BW Ae. aegypti cuticles by AFM and SEM | [15] |
| Egg sizes | Significantly smaller eggs in BW Ae. aegypti | [15] |
| Surfaces of eggs by AFM and SEM | BW Ae. aegypti egg surfaces were significantly less elastic by AFM, with more undulating surfaces seen by AFM and SEM | [15] |
| Hatchability of eggs and preimaginal development to adults | Hatchability of eggs laid and preimaginal development to adults by FW Ae. aegypti is decreased in 10 gL-1 salt BW. These properties were maternally inherited in genetic crosses | [15] |
| Susceptibility of L3 and L4 to the common larvicide Temephos | BW Ae. aegypti were significantly more resistant than FW Ae. aegypti in a 24h assay |
[15] |
| L1-L4: first to fourth instar larval stages, AFM: atomic force microscopy, SEM: scanning electron microscopy, TEM: transmission electron microscopy, LC50: concentration producing 50% lethality | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



