
Submitted:
16 November 2023
Posted:
17 November 2023
You are already at the latest version
Abstract
Anti-Müllerian hormone (AMH) has been proposed as biomarker for fertility in cattle, yet this associative relationship appears to be influenced by heat stress. Objective was to test serum AMH and AMH-related single nucleotide polymorphisms (SNPs) as markers potentially predictive of reproductive traits in heat-stressed dairy cows. The study included 300 Holstein cows that were genotyped using BovineSNP50 (54,000 SNP) after blood samples were collected and DNA extracted. A genome wide association study was then executed. Nine intragenic SNPs within the pathways that influence the AMH gene were found important with multiple comparisons adjustment test (p<1.09x10-6). A further validation study was performed in an independent Holstein cattle population, which was divided into moderate (MH; n=152) and severe heat-stressed (SH; n=128) groups, and subjected to a summer reproductive management program. Serum AMH was confirmed as a predictor of fertility measures (p<0.05) in MH but not in SH group. Cows were genotyped using the TaqMan allelic discrimination method and qPCR, which revealed four SNPs as predictive markers for serum AMH (p<0.01), reproductive traits (p<0.01) and physiological variables (p<0.05). These SNPs were in the genes AMH, IGFBP1, LGR5 and TLR4. In conclusion, both serum AMH concentrations and AMH-related SNPs appear to be associated with fertility traits in dairy cows managed during summer.
Keywords:
AMH
; fertility
; Holstein cows
; markers
; SNPs
1. Introduction
Fertility is important to the efficiency of dairy herds, as it accounts for one of the major costs of production. In the last century, selection programs in dairy cattle systems were focused primarily on improving milk yield traits [1], which caused a reduction in the genetic merit for fertility [2]. The high-production dairy cows that comprise the current population were produced with great success using this selection strategy [3]. However, the progressive decline in dairy cow fertility stimulated remarkable change in dairy cow selection indices in the past 25 years. Therefore, modern indices include functional traits such as those that are specifically designated to enhance reproduction and health, in addition to traits related to milk production [1,3,4].
Currently, the combination of reproductive management techniques and breeding selection programs have improved on-farm reproduction and accelerated the rate of fertility genetic progress in dairy cows [5,6,7]. To incorporate reproductive features into breeding programs, one of the biggest challenges has been measuring phenotypes with a reasonable heritability. For instance, heritability estimates for mostly fertility traits are usually low. Another important issue for many fertility traits is to identify easy-to-measure phenotypic variables or genetic markers that correlate to relevant fertility variables [8].
Evaluations of new measurable traits, such as endocrine or molecular markers for reproductive phenotypes, offer potential new tools to help genetic improvement of fertility in cattle [9]. Then, a phenotypic trait with high variability, heritability, and repeatability that is also associated with fertility could be a candidate to include in selection strategies [10]. The Anti-Müllerian hormone (AMH) has emerged as potential biomarker for fertility in dairy cows [11]. Concentrations of AMH in cattle have been reported to have high variability among animals and high repeatability within individual animals [12], and it is considered as a moderately heritable trait (0.36 to 0.46) [13]. Therefore, circulating AMH is proposed as reproductive management tool. The success of this tool has been reported in breeding and superovulation protocols in dairy heifers and lactating cows [14].
Novel elements that should be investigated to improve fertility in dairy cows include the range of circulating AMH and its threshold values, as well as genome-wide association studies (GWAS) that identify single nucleotide polymorphisms (SNPs) linked to phenotypic variations in AMH concentrations. The establishment of an AMH threshold, and an test executable on a dairy farm, would enable farmers to selectively breed cows with elevated AMH levels, in order to increase the reproductive efficiency in their herds [15]. Similarly, GWAS technologies make possible to identify functional genes associated with serum AMH that are directly or indirectly involved in reproductive processes, as well as to recognize specific molecular markers that could play a role enhancing fertility-related physiological events [16].
Efficacy of endocrine and molecular markers as predictors appear to be highly influenced by climate, especially in semi-desert regions that can induce heat-stressed climatic conditions [17,18]. Heat stress (HS) during summer is a major factor responsible to disrupt several reproductive events. High environmental temperatures affect heat dissipation in lactating dairy cows leading to an increase in body temperature, which is strongly associated with impaired reproduction [19]. HS affects a number of reproductive processes, including the formation of the corpus luteum, ovarian follicular growth, gonadotropin production, oocyte competence, embryonic growth, and uterine endometrial responses [20,21].
Cellular and molecular impairments appear to be part of the response of lactating diary cows to heat stress. When oocytes are exposed to heat shock during maturation, it hinders their ability to reorganize their microtubules and microfilaments and damaged spindle apparatus [22]. Oocytes that were exposed to HS showed an increase in both apoptotic gene expression and production of reactive oxygen species (ROS), which decreased the oocyte's capacity to split and develop into a blastocyst [23,24]. Taken together, heat stress-induced alterations in oocytes might be associated with fertilization failure, embryo development defects, and consequently low conception rates and (or) high embryo mortality rates [19].
Therefore, our objective was to evaluate associations of serum AMH concentrations and AMH-related single nucleotide polymorphisms (SNPs) with reproductive performance in Holstein dairy cows exposed to heat-stressed environmental conditions.
2. Materials and Methods
The Institutional Animal Care and Use Committee of the Insituto Tecnologico de Sonora approved all procedures performed in animals (Approval code 2017-0079).
2.1. Location and experimental population
Three neighboring dairy farms in the Yaqui Valley Sonora, Mexico collaborated in the current study, and they were managed as intensive milk production systems. The geographic coordinates are 27°21’ North Latitude and 109°54’ West Longitude. The site is located 46 meters above sea level. Local climate ranges from dry to semi-humid, with summer rains, an average of 371.6 mm of precipitation annually, and an a yearly mean temperature of 23 °C. During the summer, this zone can record ambient temperatures above 48 °C, and a temperature-humidity index (THI) that often exceeds 85 units.
Three hundred lactating and spring-calved Holstein cows with average body weight of 635.1 ± 32.5 kg, body condition score from 2.5 to 3.5, and 4 to 6 yrs-old were used in this study. Cows received a similar management and were kept in shaded stables with free access to a water source and a commercial trace mineral supplement. Floor and shade areas were provided according to the Holstein cows requirements. A mixed diet was supplied twice daily, which was designed to meet requirements for dairy cows with average body weight of 650 kg and daily milk production of 30 kg. This ration's average composition was 3.5% fat and 3.2% crude protein.
2.2. Blood sampling for AMH
Blood samples were collected early in the summer by puncturing the coccygeal vein using serum tubes, which were promptly placed on ice. After clotting, samples were centrifuged for 30 min at room temperature (18 to 22 °C). Subsequently, serum samples were stored at -20 °C until analysis. Serum AMH concentrations were measured using the Minitube of America AMH-bovine specific immunoassay (AMH Fertility AssayTM), which has a sensitivity of 0.04 pg/mL and an intra-assay coefficient of variation of 2.2%.
2.3. SNP data and quality control
An additional blood sample of 3 ml was drawn from each cow through venipuncture of the coccygeal vein using disposable sterile syringes. Five drops of the collected whole blood were spotted on Fast Technology for Analysis of Nucleic Acids cards (FTA®), which were sent to Neogen AgriGenomics (Lincoln, NE) for DNA extraction and genotyping. The SNP panel BovineSNP50, which contains 53,218 highly informative SNPs evenly dispersed throughout the whole bovine genome, was used to acquire genotypes of each cow.
PLINK v1.07 software was used to implement quality control for the SNPs that were used in genotype to phenotype association analyses. Only SNPs that met the following criteria were included in analyses: A call rate of greater than 95% or a false discovery rate of less than 5%, a missing genotype frequency of less than 5%, a minor allele frequency (MAF) of more than 5%, and no-deviation from Hardy-Weinberg equilibrium (p-value of Chi-square goodness-of-fit test greater than 0.05, X2 > 0.05). The analyses also excluded SNPs with uncertain loci or those on the sex chromosomes. After quality control, a total of 45,832 SNPs were retained for further analyses.
2.4. Genome-wide association study (GWAS)
Batch effects/stratification of the test input data were corrected by using principal component analysis (PCA) option. The genomic analysis was then performed using a single-locus mixed model to study associations between genotypes of each SNP marker with serum AMH level as a phenotypic observation (i.e., single-marker SNP GWAS). The software SNP & Variation Suite v8 (SVSv8; Golden Helix, Inc., Bozeman, MT, www.goldenhelix.com) was used for this purpose by fitting all SNP simultaneously.
The additive mixed model used was: y = Xβ + Za + e, where y was the vector of phenotypic observations (AMH), X was the incidence matrix of fixed effects, β was the vector of fixed effects, Z was the design matrix of random additive genetic effects, a was the vector of random additive genetic effects, and e was the vector of residual effects. This model assumed that a~N (0, Gσ2a) and e~N (0, Iσ2e), where σ2a was the additive genetic variance, σ2e was the residual variance component, G was the genomic relationship matrix and I was the identity matrix.
2.5. Multiple testing adjustment
The p-values obtained from the genomic analyses were subjected to the Bonferroni correction test (b=α/n), which assumed independence between SNPs to adjust for multiple comparisons. The experiment-wise error was α=0.05 and the number of tests (n) was taken to be the number of the useful SNPs (n=45,832). Therefore, the 5% genome-wide threshold was equivalent to a nominal p-value of 1.09 × 10−6, corresponding to 5.96 on a −log10(P-value) scale.
2.6. Validation population
An independent Holstein cattle population (n=280) was selected to perform the validation study for the candidate SNP markers previously identified as associated with serum AMH. Cows were selected from two neighboring dairy herds in the Yaqui Valley; they were spring-calved Holstein, with average body weight of 651.3 ± 34.7 kg, mean body condition score of 3.0 ± 0.5, and aged 4 to 7. Cows were randomly assigned to one of two experimental periods. The moderate heat-stressed (MH; n=152) group included cows managed during late spring and early summer (AT= 28.1 ºC, RH= 55.8% and THI= 75.7 units), whereas the severe heat-stressed (SH; n=128) group included cows managed during middle and late summer (AT= 30.6 ºC, RH= 71.4% and THI= 82.2 units). Level of heat stress was established according to the classification described by Collier et al. [25]: (1) absence of heat stress (< 68 units), (2) heat tress threshold (68–72 units), (3) moderate heat stress (73–79 units), and (4) severe heat stress (> 79 units).
2.7. Reproductive management
Cows were subjected to an ovulation synchronization protocol that included the insertion of an intravaginal device for delayed progesterone release (CIDR®, Pfizer, Mexico) at day 0 plus an intramuscular (im) administration of 0.01 mg of GnRH (Fertagyl®, Intervet, Mexico). The CIDR was removed 7 days later and then 25 mg im of prostaglandin F2a (PGF2a, Lutalyse®, Pfizer, Mexico) was applied. The cows were given a second dose of 0.01 mg of GnRH at day 9, and 24 hours later they were artificially inseminated (FTAI; day 10). At 30 days following FTAI, ultrasound scanning was used to diagnose pregnancy. The number of pregnant cows divided by the total number of FTAI services at day 30 was used to compute the conception rate (CR).
A trans-rectal ultrasonography inspection was performed in cows utilizing a 7.5 MHz transducer (Sonosite MicroMaxx TM®, Bothell, WA, USA) to assess the health of their reproductive system, mainly uterine body and horns. Ovarian activity was also evaluated through measuring dominant follicle diameter (FOL) on day 7 of the synchronization protocol, as well as Corpus Luteum diameter (CL) on day 14 after AI.
2.8. Physiological traits and climatic data
During the course of the study, both rectal temperature (RT; °C) and respiratory rate (RR; breaths/min) were measured at 1200 h two times per week. The RT was obtained using a digital thermometer (TES- 1310®) with a contact sounding line (type K; 9-cm large), which was inserted into the animal to touch the rectal mucosa, while the RR was determined by visually counting the intercostal movements. A blood sample was collected at the beginning of the study to measure serum AMH as described above.
Data on the ambient temperature (AT; °C) and relative humidity (RH; %) were obtained from the REMAS website (http://www.siafeson.com/remas) by a local meteorological station (Network of Automatic Meteorological Stations of Sonora). Daily records from both AT and RH were used to calculate the temperature-humidity index (THI), by processing the following equation: THI = (0.8 AT) + [(RH/100) (AT 14.4)] + 46.4 [26].
2.9. SNP marker genotyping
An additional blood sample was collected using Vacutainer tubes with EDTA (EDTA-Na2; Venoject®, Terumo, Lakewood, CA, USA), by puncturing the jugular vein from each cow at the beginning of the study. The samples were centrifuged at 3500 RMP for 15 min. Then, 200 μl of leukoplatelet layer was removed by pipet and refrigerated at −20 °C. Next, the DNA was extracted using a commercial kit (DNeasy Blood & Tissue Kits; QIAGEN, Hilden, Germany) and following the manufacturer’s instructions. The concentration and purity of the DNA were determined using a NanoDrop automated spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
The nine significant SNPs reported in this study were genotyped using the TaqMan method for allelic discrimination and RT-qPCR (StepOneTM, Applied Biosystems, Foster City CA, USA), in accordance with the methods described by Castillo-Salas et al. [27]. The PCR was carried out using the StepOne Real-Time PCR System from Thermo Fisher Scientific (Massachusetts, USA). Finally, genotyping and analysis of PCR data were performed using StepOne software (Life Technologies Corporation, version 2.3).
2.10. Statistics for the validation analysis
Mean values of the analyzed traits were calculated using the MEANS and FREQ procedures for continuous and categorical variables, respectively. The assumptions of normality and homogeneity of variances were tested through the UNIVARIATE procedure. Allele and genotype frequencies for each polymorphism and deviations from Hardy-Weinberg equilibrium were obtained using the ALLELE program.
A mixed effects statistical model was used to perform an associative study using the MIXED and GLIMMIX procedures for continuous (AMH, FOL, CL, RT and RR) and categorical variables (CR), respectively. For the statistical processing of the information, the SAS (Statistical Analysis System) program version 9.4 was used, which included the procedures for genetic studies [28]. The statistical model included the fixed effects of the SNP genotype, herd and lactation, as well as the covariant of days in milk, and the random effect of sire.
If genotype term was confirmed as significant source of variation (p < 0.05) in the associative statistical analysis, then LSMEANS procedure was performed to make comparisons between average genotypic values, considering the Bonferroni adjustment [29]. The allele substitution effects (i.e., effect of substituting one allele for another within the population) was determined using a regression model that included the allelic effect as a covariate [29]. Additive genetic effects were obtained using the procedures described by Luna-Nevárez et al. [30].
3. Results
3.1. Climate and AMH sampling
During the validation study, moderate HS characterized the climatic conditions in late spring and early summer (i.e., THI ranged from 72 to 79 units), whereas severe heat stressed conditions prevailed in mid- and late summer (i.e., THI ranged from 79 to 84), as observed in Figure 1.
Average values for serum AMH, reproductive traits and physiological variables differed (p < 0.05) in cows managed during moderate heat stress compared with those cows exposed to severe heat stress (Table 1). A decrease in serum AMH, FOL, CL and CR was observed when the THI surpassed the threshold of 79 units, whereas an slight increase was detected in the physiological traits RT and RR.
3.2. AMH as endocrine marker during summer
In late spring and early summer, high correlations (r = 0.71 to 0.75, p < 0.05) were observed between serum AMH and reproductive traits (i.e., FOL and CL; Table 2). However, this relationship diminished during summer when the THI surpassed 79 units, as correlations dropped (r < 0.30) due to the severe heat stress. A similar associative relationship trend was observed between serum AMH and physiological traits (i.e., RT and RR) through the study, although correlations were mostly not significant.
The simple regression analysis detected serum AMH as predictor for the reproductive traits FOL (R2 = 0.56; p < 0.01) and CL (R2 = 0.52; p < 0.01) but only for cows managed under moderate heat stress (THI = 72 to 79 units; Figure 2a,b). When the THI increased above 79 units, the severe heat stress appeared to affect the ability of serum AMH to predict the ovarian traits FOL and CL (Figure 2c,d).
3.1. Genome-wide association study
After applying quality control criteria, 45,832 SNPs were interpreted useful and were used in GWAS analysis. Then, the single-locus mixed model identified 81 genome- wide SNPs associated with serum concentrations of AMH (p < 0.00001). However, only 16 of these SNPs surpassed multiple testing correction because they were significant at lower p-value than the Bonferroni adjusted threshold (i.e., 1.09 × 10−6; Figure 3). From these 16 SNPs, nine were located within genes (i.e., intronic), whereas the remaining seven SNPs showed an intergenic position as observed in Table 3.
3.2. SNPs selection
From the 16 genome-wide SNPs significantly associated with AMH serum levels, the 9 intragenic SNPs were selected for use in a validation study because they are located within genes. However, only 4 intragenic SNPs did not deviate from the Hardy–Weinberg equilibrium (HWE, X2 > 0.05) and met the criteria for minor allele frequency higher than 10% (MAF > 0.10). These 4 SNPs were rs876084180, rs445674221, rs42849475 and rs8193046, which were in the genes AMH, IGFBP1, LGR5 and TLR4 (Table 4). Therefore, only these 4 SNPs were further tested for validation as molecular markers through a genotype to phenotype association study.
3.3. SNP markers validation
The least square means for AMH and reproductive traits according to SNP genotypes are reported in Table 5. The SNP rs876084180 was associated with serum AMH, FOL and CL (p < 0.01), whereas the SNP rs42849475 was associated with ovarian traits and CR (p < 0.05). Interestingly, the SNPs rs445674221 and rs8193046 were predictors of serum AMH (p < 0.0001) and all reproductive traits (p < 0.01). The SNPs rs445674221 and rs8193046 were also associated with the physiological traits RT and RR (p < 0.01), whereas the SNPs rs876084180 and rs42849475 were associated only with RT (p < 0.05) as observed in Table 6.
From the SNPs rs876084180, rs445674221, rs42849475 and rs8193046, the most favorable genotypes were AA, GG, GG and AA, respectively, because they were associated with a better reproductive and physiological response. According to the allele substitution effects, the SNP rs876084180 had the highest allele contribution (p < 0.0001) on serum AMH levels. However, the SNPs rs445674221 and rs8193046 were the highest contributors for the reproductive traits FOL, CL and CR (p < 0.1). Interestingly, these two SNPs were also the most beneficial contributors for the physiological traits RT and RR (p < 0.01), as they were associated with a slight reduction in both variables.
A significant additive fixed effect was confirmed for the SNPs rs876084180, rs445674221, rs42849475 and rs8193046, suggesting that the sum of their individual effects was equal to their combined allele effects.
3.4. SNP effects on reproductive phenotypes
A significant improvement in serum AMH and reproductive traits (p < 0.05) was observed as the number of favorable genotypes of the four genome-wide significant SNP markers increased from 0 to 4. Serum AMH differed in cows having 0 to 1, 2 to 3 and 4 favorable genotypes; FOL differed among all favorable genotype categories; CL differed between 0, 1, 2 to 3 and 4 favorable genotypes; and CR differed in cows having 0, 1 to 3 and 4 favorable genotypes (Figure 4a–d).
4. Discussion
Anti-Müllerian hormone (AMH) has been suggested as potential biomarker associated with ovarian function and fertility traits in dairy cows [13,31]. AMH has been positively associated with the total number of healthy oocytes and follicles in the cow, and this relationship is influenced by climate [17]. An increase in temperature and humidity lead to heat stress (HS) in lactating dairy cows, which affect oocyte quality and development, follicular growth and embryo viability [32,33]. Severe heat-stressed conditions appear to compromise the predictive ability of AMH as physiological marker. However, the reliability of AMH as molecular marker appears to be unaffected by adverse environmental conditions. In the current study, we detected that serum AMH appeared to be an endocrine marker for summer reproductive response in Holstein cows, but only when HS did not exceed a moderate threshold. Furthermore, we identified four SNPs associated with serum AMH as molecular markers for reproductive traits, which also appeared to be predictors of physiological traits indicative of HS tolerance.
In northwestern Mexico, the Yaqui Valley experiences extremely hot weather conditions characterized by an average THI that varies from 75.7 units in late spring to 82.2 units at the end of summer. These THI values indicate heat-stressed climatic conditions progressing from moderate to severe during summer, and creating a unique hot weather favorable to study heat stress influence on physiological and molecular markers in dairy cattle [18,34]. Therefore, further studies are proposed to elucidate the ability of circulating AMH concentrations to predict fertility in Holstein dairy cows exposed to HS. In addition, the study of AMH polymorphisms as predictors for reproductive performance and thermotolerance, appears to be a viable strategy for genetic improvement of Holstein cows managed under warm environments.
Endocrine and molecular technologies have been proposed to identify practical and reliable markers for fertility in cattle. The circulating AMH concentration is proposed as potential candidate marker due to this trait having high variability, repeatability and heritability [35,36,37]. In the current study, we first tested serum AMH as endocrine marker in Holstein cows managed from late spring until the end of the summer. Serum AMH collected at the beginning of a reproductive management program was associated with higher FOL, CL and CR, and resulted as predictor for ovarian response (p < 0.05) under moderate HS. However, this predictive ability was not detected in Holstein cows when HS progressed from moderate to severe during summer.
We observed a significant reduction in average serum AMH in Holstein cows after HS increased from moderate to severe (417.26 ± 4.51 to 136.94 ± 4.03 pg/mL). According to Gobikrushanth et al. [15] AMH concentrations in dairy cows are classified as low (≤140 pg/mL), intermediate (>140 to ≤450 pg/mL) or high (>450 pg/mL). In our study, serum AMH dropped from intermediate to low levels as the summer season progressed. A significant decrease in follicular growth, ovulation rate and embryo quality was observed in cows averaging serum AMH below 400 pg/ml [17]. In another study, cows with high and intermediate plasma AMH concentrations were 1.42 and 1.51 times more likely to be associated with pregnant than cows with low plasma AMH concentrations within 84 days following the start of the mating season [38].
High temperatures and humidity in cattle affect ovarian hormone concentrations (i.e., AMH, FSH, LH, etc.), uterine environment, and follicular development, which compromises oocyte competence and early embryonic development [39,40,41]. Fertilization process and embryonic development were disrupted by heat stress leading to a 30-35% decrease in conception rates in dairy cows [6]. Reductions in hematocrit and modifications in water balance during heat stress may attribute to the decline in hormone concentrations within peripheral blood [42].
After executing GWAS, we detected 16 SNPs associated with serum AMH that were significant at the Bonferroni adjusted p-value. Only 9 of these SNPs were selected as candidate marker because they were located within a gene. From these SNPs, 4 of them surpassed the quality control criteria and were tested through a genotype to phenotype association study. Such SNPs were rs876084180, rs445674221, rs42849475 and rs8193046, which were within the genes AMH, IGFBP1, LRG5 and TLR4. These SNP were then validated as molecular markers for fertility traits and thermotolerance in Holstein cows subjected to a reproductive management program during the summer.
Nawaz et al. [13] reported candidate genome-wide SNPs on BTA11 and BTA20 associated with AMH concentrations and located in the genes DENND1A, NR5A1, NR6A1, PTGS1, NDUFA8 and FST, which have plausible roles on fertility and AMH regulation. The DENND1A gene was also described as a candidate gene associated with total number of collected and viable embryos in superovulated Canadian Holstein cows [43]. Gobikrushanth et al. [15] reported genome-wide SNPs associated with variation in serum AMH in Holstein cows, one within the AMH gene (BTA7) and four in the genes SCAI and PPP6C (BTA11), and FGF18 and EEF2K (BTA20). A similar GWAS study for AMH concentration in cows from several dairy breeds detected a single quantitative traits locus (QTL) on BTA11. This QTL included candidate genes linked to fertility-related phenotypes, some of them previously reported (NR5A1, HSPA5, CRB2, DENND1A and NDUFA8) [38].
The four genome-wide SNPs validated in the current study as markers associated with fertility traits and thermotolerance in Holstein cows are located in the genes AMH, IGFBP1, LRG5 and TLR4. The SNPs rs876084180 is located within the AMH (Anti-Müllerian hormone) gene. AMH is a dimeric glycoprotein synthesized by granulosa cells within growing preantral and antral follicles [44]. This protein is considered a potential marker of the ovarian follicular reserve and healthy follicular population in cows [31,35], because it is highly variable, repeatable and heritable [35,36,37].
In our study, results suggested that the SNPs rs876084180 in the gene AMH was a predictor for reproductive traits (i.e., FOL, CL and CR), because AMH regulates follicular growth and oocyte competence, as well as fertilization and embryo development. Such reproductive functions of AMH appear to be due to its close relationship with the FSH signaling pathway, which regulate and promote ovarian follicle development [45]. In addition, the ability of AMH to predict RT is because the direct participation of this protein within TGFβ pathway, which is involved in architecture remodeling of cells exposed to heat shock [46].
The SNP rs445674221 is within the gene IGFBP1. This gene encodes a protein primarily produced in hepatocytes and renal cells, which is a key modulator of the IGF1. The IGFBP1 is released into the bloodstream by the liver and binds to IGF1 more strongly than its receptor IGFR1 to control the bioavailability and activity of IGF1 [29]. The IGF1 peptide is essential in the regulation of reproductive functions by acting synergistically with gonadotropins [47]. This peptide plays an important role in oocyte development, follicular growth, oocyte maturation, ovulation, luteal function, and also prevent cellular apoptosis and enhance proliferation in the ovary [48,49,50].
Under heat stress, IGFBP1 transports IGF1 to the membrane of the oocyte allowing the interaction with its receptor (IGF1R), which induces activation of PI3K/AKT signal pathway [51]. Then, AKT inactivates caspases (CASP-3 and CASP-9) [52], and regulates the levels of the BCL-2 family proteins, which reduce cellular alterations caused by heat stress (i.e., increase in ROS and H2O2) [53]. As per activation of the receptor by the IGF1 ligand, the functionality of the mitochondrial membrane is stabilized, the continuation of apoptosis in the oocyte is deferred, and the cell survival pathways are activated [54,55]. Addition of IGF1 (12.5 to 25 ng/ml) to oocytes cultured at 41 °C tended to minimize the reduction in both mitochondrial activity and developmental competence, two processes affected by heat stress [56,57].
The SNP rs42849475 is within the gen LGR5. This gene is expressed in the epithelial surface of the ovarian stem cells [58], as well as in the surface of the Corpus Luteum [59]. It could explain the association of LGR5 gene with the diameter of FOL and CL in the current study. Similarly, this gene is also expressed in the uterine epithelia, where appears to collaborate in uterine functions needed to support implantation and pregnancy establishment [60]. It seems to explain the relationship between LGR5 gene and CR.
LGR5 is a potential target of the Wnt signaling pathway [61], which promotes cell proliferation and cell survival during heat stress. The proliferative mechanism has been involved in the growth of the mammary gland in dairy cows [62]. This pathway is also associated with energy metabolism, a critical function required to maintain a thermal balance within the body once the animal is exposed to heat stress [63]. It appeared to support effects of LGR5 gene on thermotolerance-related traits.
The SNP rs8193046 is in the gene TLR4. This gene is a crucial mediator of the inflammation-like response in the uterus triggering the production of regulatory T (Treg) cells, which are required to facilitate and promote embryo implantation [64]. Treg cells suppress anti-fetal effector responses and support the process of uterine vascular remodeling, facilitating strong placental development that supports growth and survival of the fetus [65,66]. It provides evidence of the mechanisms through that TLR4 is helping to improve conception rate as observed in the current study.
As part of the immune response against pathogens and external stressors, TLR4 induces the expression of pro-inflammatory cytokines [67], which are regulated by nuclear factor kappa-B (NF-κB) expression [68,69]. NF-κB is an important intracellular signaling protein that has a critical role in preventing heat stress-induced early apoptosis, and instead promotes growth and survival in cells exposed to heat-stressed conditions [70]. TLR4 has been associated with a reduction in both lypolitic action and adypocite mobilization in Holstein cows exposed to heat stress as a body response to minimize heat load [71,72]. It appeared to explain the associative relationship between TLR4 gene and thermotolerance traits observed in the current study.
The allele substitution effects detected in the current study supported the concept of the favorable contribution of the genes AMH, IGFBP1, LGR5 and TLR4 to reproductive and physiological traits evaluated in the current study. This positive effect of the four validated genes was confirmed by the improvement in serum AMH, FOL diameter, CL diameter and conception rate as the number of SNPs with favorable genotypes increased from 0 to 4. A previous report also involving GWAS and a validation study identified 3 genes (i.e., TLR4, GRM8 and SMAD3) as favorable markers for milk yield and thermotolerance in Holstein cows managed in a heat stressed environment [18].
The identification of genetic makers associated with variations in circulating AMH concentrations in Holstein cows exposed to HS would be potentially helpful to identify and select future elite genetic merit cows with greater potential to respond successfully in summer reproductive programs [15]. The discovery and validation of novel molecular markers associated with serum AMH appeared to be a promissory strategy to identify dairy cows with superior fertility during summer. Selection of these cows within a genetic improvement program could lead to faster genetic gain in reproductive efficiency in dairy production systems within in semiarid regions [13,31]. As stated by Hansen et al. [73], selecting for genetic thermotolerance makes possible to reduce the impact of HS on reproduction. Genomic studies in Australian Holstein cows reported higher breeding values for fertility in cows that were genetically more thermotolerant [74].
A limitation of the current study is the low number of cows included in the GWAS analysis, which combined with the multiple-testing statistical adjustment may reduce the number of significant SNPs detected [75]. In addition, due to the polygenic effect of AMH a reduced variation explained by the markers is expected. Fertility is a highly polygenic trait that is influenced by thousands of SNP effects [15,76], and the candidate genes we detected appeared to be a small number of the many that slightly contribute to phenotypic variance of serum AMH concentrations. Interestingly, our validation study in an independent Holstein population confirmed the ability of these genes as predictors for fertility and thermotolerance traits. Because the likelihood of an SNP being significant in two different populations is low, validating the effects of SNPs in independent animal populations seems to be the most reliable method for determining the significance of candidate SNPs [77,78].
5. Conclusions
Identification of physiological and genetic markers for fertility in Holstein cows exposed to warm climate conditions during summer could be a favorable strategy for dairy producers. Anti-Müllerian hormone (AMH) has been recognized as potential blood marker associated with reproductive performance in Holstein cows; however, there is limited information on its effectiveness as a marker under extreme climatic conditions of heat. In the current study, we provided evidence that serum AMH concentration was an endocrine biomarker for summer reproductive response but only when heat stress did not exceed a moderate threshold. Interestingly, molecular analysis revealed four SNPs associated with serum AMH concentrations as genetic markers to improve fertility traits and thermotolerance in dairy cows managed under severe summer weather conditions. However, further GWAS analyses including extensive dairy populations are recommended in order to identify and validate more candidates SNPs associated with serum AMH. Also, additional studies using these markers in the processes to estimate breeding values for fertility in Holstein cows exposed to different heat stress intensities could be helpful in dairy cattle genetic improvement programs.
Author Contributions
Conceptualization, J.F.M., M.G.T., R.M.E., S.E.S. and P.L.-N.; methodology, L.A.C.-M., P.A.L.-C., and F.R.-A.; software, G.L.-N. and P.L.-N.; validation, P.A.L.-C. and P.L.-N..; formal analysis, L.A.C.-M.., G.L.-N. and P.L.-N.; investigation, L.A.C.-M., F.R.-A. and P.L.-N..; resources, J.F.M., M.G.T. and P.L.-N.; data curation, G.L.-N.; writing—original draft preparation, M.G.T. and P.L.-N.; writing—review and editing, J.F.M., M.G.T., R.M.E. and S.E.S.; visualization, L.A.C.-M. and P.L.-N.; supervision, J.F.M., M.G.T. and P.L.-N.; project administration, J.F.M. and P.L.-N.; funding acquisition, J.F.M., M.G.T. and P.L.-N. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by UCMEXUS-CONACYT Grant Program 2016 through the project titled “Genomic analyses of thermotolerance in Holstein dairy cattle managed during summer in southern Sonora, Mexico” (Project Number CN-16-123). This project was also funded by PROFAPI- ITSON Grant Program 2017 through the project titled “Validation of genetic markers associated with thermotolerance and milk production in Holstein dairy cattle managed under heat-stressed environmental conditions” (Project Number 2017-0079).
Institutional Review Board Statement
The Institutional Animal Care and Use Committee of the Instituto Tecnologico de Sonora approved all procedures performed on animals (Protocol code 2017-02 approved on 1 March 2017).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, P.L.-N., upon reasonable request.
Acknowledgments
We thank the respective staff of the cooperative dairy herds, dairy producers, and students involved during the project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cole, J.B.; VanRaden, P.M. Symposium review: Possibilities in an age of genomics: The future of selection indices. J. Dairy Sci. 2018, 101, 3686–3701. [Google Scholar] [CrossRef] [PubMed]
- Butler, S.T. Genetic control of reproduction in dairy cows. Reprod. Fertil. Dev. 2013, 26, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Miglior, F.; Fleming, A.; Malchiodi, F.; Brito, L.F.; Martin, P.; Baes, C.F. A 100-Year Review: Identification and genetic selection of economically important traits in dairy cattle. J. Dairy Sci. 2017, 100, 10251–10271. [Google Scholar] [CrossRef] [PubMed]
- Pryce, J.E.; Nguyen, T.T.T.; Axford, M.; Nieuwhof, G.; Shaffer, M. Symposium review: Building a better cow-The Australian experience and future perspectives. J. Dairy Sci. 2018, 101, 3702–3713. [Google Scholar] [CrossRef] [PubMed]
- Thomasen, J.R.; Willam, A.; Egger-Danner, C.; Sørensen, A.C. Reproductive technologies combine well with genomic selection in dairy breeding programs. J. Dairy Sci. 2016, 99, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, P.D.; Santos, V.G.; Giordano, J.O.; Wiltbank, M.C.; Fricke, P.M. Development of fertility programs to achieve high 21-day pregnancy rates in high-producing dairy cows. Theriogenology 2018, 1, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Lucy, M.C. Symposium review: Selection for fertility in the modern dairy cow-Current status and future direction for genetic selection. J. Dairy Sci. 2019, 102, 3706–3721. [Google Scholar] [CrossRef] [PubMed]
- Häggman, J.; Christensen, J.M.; Mäntysaari, E.A.; Juga, J. Genetic parameters for endocrine and traditional fertility traits, hyperketonemia and milk yield in dairy cattle. Animal 2019, 13, 248–255. [Google Scholar] [CrossRef]
- Crowe, M.A.; Hostens, M.; Opsomer, G. Reproductive management in dairy cows—the future. Ir. Vet. J. 2018, 8, 1. [Google Scholar] [CrossRef]
- Jimenez-Krassel, F.; Scheetz, D.M.; Neuder, L.M.; Ireland, J.L.; Pursley, J.R.; Smith, G.W.; Tempelman, R.J.; Ferris, T.; Roudebush, W.E.; Mossa, F.; et al. Concentration of anti-Müllerian hormone in dairy heifers is positively associated with productive herd life. J. Dairy Sci. 2015, 98, 3036–3045. [Google Scholar] [CrossRef]
- Mossa, F.; Ireland, J.J. Physiology and endocrinology symposium: Anti-Müllerian hormone: A biomarker for the ovarian reserve, ovarian function, and fertility in dairy cows. J. Anim. Sci. 2019, 97, 1446–1455. [Google Scholar] [CrossRef]
- Gobikrushanth, M.; Dutra, P.A.; Bruinjé, T.C.; Colazo, M.G.; Butler, S.T.; Ambrose, D.J. Repeatability of antral follicle counts and anti-Müllerian hormone and their associations determined at an unknown stage of follicular growth and an expected day of follicular wave emergence in dairy cows. Theriogenology 2017, 1, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.Y.; Jimenez-Krassel, F.; Steibel, J.P.; Lu, Y.; Baktula, A.; Vukasinovic, N.; Neuder, L.; Ireland, J.L.H.; Ireland, J.J.; Tempelman, R.J. Genomic heritability and genome-wide association analysis of anti-Müllerian hormone in Holstein dairy heifers. J. Dairy Sci. 2018, 101, 8063–8075. [Google Scholar] [CrossRef] [PubMed]
- Alward, K.J.; Bohlen, J.F. Overview of Anti-Müllerian hormone (AMH) and association with fertility in female cattle. Reprod Domest Anim. 2020, 55, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Gobikrushanth, M.; Purfield, D.C.; Colazo, M.G.; Butler, S.T.; Wang, Z.; Ambrose, D.J. The relationship between serum anti-Müllerian hormone concentrations and fertility, and genome-wide associations for Anti-Müllerian hormone in Holstein cows. J. Dairy Sci. 2018, 101, 7563–7574. [Google Scholar] [CrossRef] [PubMed]
- Martínez, C.A.; Roca, J.; Barranco, I. Editorial: Molecular Biomarkers in Animal Reproduction. Front. Vet. Sci. 2021, 1, 802187. [Google Scholar] [CrossRef] [PubMed]
- Torres-Simental, J.F.; Peña-Calderón, C.; Avendaño-Reyes, L.; Correa-Calderón, A.; Macías-Cruz, U.; Rodríguez-Borbón, A.; Leyva-Corona, J.C.; Rivera-Acuña, F.; Thomas, M.G.; Luna-Nevárez, P. Predictive markers for superovulation response and embryo production in beef cattle managed in northwest Mexico are influenced by climate. Liv, Sci. 1045. [Google Scholar] [CrossRef]
- Zamorano-Algandar, R.; Medrano, J.F.; Thomas, M.G.; Enns, R.M.; Speidel, S.E.; Sánchez-Castro, M.A.; Luna-Nevárez, G.; Leyva-Corona, J.C.; Luna-Nevárez, P. Genetic markers associated with milk production and thermotolerance in Holstein dairy cows managed in a heat-stressed environment. Biology 2023, 12, 679. [Google Scholar] [CrossRef] [PubMed]
- Wolfenson, D.; Roth, Z. Impact of heat stress on cow reproduction and fertility. Anim. Front. 2018, 9, 32–38. [Google Scholar] [CrossRef]
- Hansen, P.J. Exploitation of genetic and physiological determinants of embryonic resistance to elevated temperature to improve embryonic survival in dairy cattle during heat stress. Theriogenology 2007, 68 (Suppl 1), S242–9. [Google Scholar] [CrossRef]
- Gendelman, M.; Aroyo, A.; Yavin, S.; Roth, Z. Seasonal effects on gene expression, cleavage timing, and developmental competence of bovine preimplantation embryos. Reproduction 2010, 140, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Roth, Z.; Hansen, P.J. Disruption of nuclear maturation and rearrangement of cytoskeletal elements in bovine oocytes exposed to heat shock during maturation. Reproduction 2005, 129, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.M.; Chiaratti, M.R.; Macabelli, C.H.; Rodrigues, C.A.; Ferraz, M.L.; Watanabe, Y.W.; Smith, L.C.; Meirelles, F.V.; Baruselli, P.S. The infertility of repeat-breeder cows during summer is associated with decreased mitochondrial DNA and increased expression of mitochondrial and apoptotic genes in oocytes. Biol. Reprod. 2016, 94, 66. [Google Scholar] [CrossRef] [PubMed]
- Roth, Z. Effect of heat stress on reproduction in dairy cows: Insights into the cellular and molecular responses of the oocyte. Annu. Rev. Anim. Biosci. 2017, 5, 151–170. [Google Scholar] [CrossRef] [PubMed]
- Collier, R.J.; Zimbelman, R.B.; Rhoads, R.P.; Rhoads, M.L.; Baumgard, L.H. A re- evaluation of the impact of temperature humidity index (THI) and black globe humidity index (BGHI) on milk production in high producing dairy cows. Proc. 24th Annual South. Nut. Man. Conf., Tempe, Arizona, USA 2009, 113–125. [Google Scholar]
- Mader, T.L.; Johnson, L.J.; Gaughan, J.B. A comprehensive index for assessing environmental stress in animals. J. Anim Sci. 2010, 88, 2153–2165. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Salas, C.A.; Luna-Nevárez, G.; Reyna-Granados, J.R.; Luna-Ramirez, R.I.; Limesand, S.W.; Luna-Nevárez, P. Molecular markers for thermo-tolerance are associated with reproductive and physiological traits in Pelibuey ewes raised in a semiarid environment. J. Therm. Biol. 2023, 112, 103475. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S. Forensics: Handbook of Statistical Genetics; John Wiley and Sons: Hoboken, NJ, USA, 2001. [Google Scholar]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Longman Scientific and Technical: New York, NY, USA, 1996. [Google Scholar]
- Luna-Nevarez, P.; Rincon, G.; Medrano, J.F.; Riley, D.G.; Chase, C.C.Jr.; Coleman, S.W.; VanLeeuwen, D.; DeAtley, K.L.; Islas-Trejo, A.; Silver, G.A.; et al. Single nucleotide polymorphisms in then growth hormone-insulin like growth factor axis in straightbred and crossbred Angus, Brahman, and Romosinuano heifers: Population genetic analyses and association of genotypes with reproductive phenotypes. J. Anim. Sci. 2011, 8, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Rico, C.; Fabre, S.; Médigue, C.; di Clemente, N.; Clément, F.; Bontoux, M.; Touzé, J.L.; Dupont, M.; Briant, E.; Rémy, B.; Beckers, J.F.; et al. Anti-mullerian hormone is an endocrine marker of ovarian gonadotropin-responsive follicles and can help to predict superovulatory responses in the cow. Biol. Reprod. 2009, 80, 50–59. [Google Scholar] [CrossRef]
- De Rensis, F.; Saleri, R.; Garcia-Ispierto, I.; Scaramuzzi, R.; López-Gatius, F. Effects of heat stress on follicular physiology in dairy cows. Animals 2021, 11, 3406. [Google Scholar] [CrossRef]
- Dahl, G.E.; Tao, S.; Laporta, J. Heat stress impacts immune status in cows across the life cycle. Front. Vet. Sci. 2020, 7, 116. [Google Scholar] [CrossRef]
- Leyva-Corona, J.C.; Reyna-Granados, J.R.; Zamorano-Algandar, R.; Sanchez-Castro, M.A.; Thomas, M.G.; Enns, R.M.; Speidel, S.E.; Medrano, J.F.; Rincon, G.; Luna-Nevarez, P. Polymorphisms within the prolactin and growth hormone/insulin-like growth factor-1 functional pathways associated with fertility traits in Holstein cows raised in a hot-humid climate. Trop. Anim. Health Prod. 2018, 50, 1913–1920. [Google Scholar] [CrossRef]
- Monniaux, D.; Drouilhet, L.; Rico, C.; Estienne, A.; Jarrier, P.; Touzé, J.; Sapa, J.; Phocas, F.; Dupont, J.; Dalbiès-Tran, R.; Fabre, S. Regulation of anti-Müllerian hormone production in domestic animals. Reprod. Fertil. Dev. 2012, 25, 1–16. [Google Scholar] [CrossRef]
- Ribeiro, E.S.; Bisinotto, R.S.; Lima, F.S.; Greco, L.F.; Morrison, A.; Kumar, A.; Thatcher, W.W.; Santos, J.E.P. Plasma anti-Müllerian hormone in adult dairy cows and associations with fertility. J. Dairy Sci. 2014, 97, 6888–6900. [Google Scholar] [CrossRef]
- Souza, A.H.; Carvalho, P.D.; Rozner, A.E.; Vieira, L.M.; Hackbart, K.S.; Bender, R.W.; Dresch, A.R.; Verstegen, J.P.; Shaver, R.D.; Wiltbank, M.C. Relationship between circulating anti-Müllerian hormone (AMH) and superovulatory response of high-producing dairy cows. J. Dairy Sci. 2015, 98, 169–178. [Google Scholar] [CrossRef]
- Gobikrushanth, M.; Purfield, D.C.; Canadas, E.R.; Herlihy, M.M.; Kenneally, J.; Murray, M.; Kearney, F.J.; Colazo, M.G.; Ambrose, D.J.; Butler, S.T. Anti-Müllerian hormone in grazing dairy cows: Identification of factors affecting plasma concentration, relationship with phenotypic fertility, and genome-wide associations. J. Dairy Sci. 2019, 102, 11622–11635. [Google Scholar] [CrossRef]
- Dash, S.; Chakravarty, A.K.; Singh, A.; Upadhyay, A.; Singh, M.; Yousuf, S. Effect of heat stress on reproductive performances of dairy cattle and buffaloes: A review. Vet. World. 2016, 9, 235–244. [Google Scholar] [CrossRef]
- Santana, M.L.; Bignardi, A.B.; Pereira, R.J.; Stefani, G.; El Faro, L. Genetics of heat tolerance for milk yield and quality in Holsteins. Animal 2017, 11, 4–14. [Google Scholar] [CrossRef]
- Succu, S.; Sale, S.; Ghirello, G.; Ireland, J.J.; Evans, A.C.O.; Atzori, A.S.; Mossa, F. Exposure of dairy cows to high environmental temperatures and their lactation status impairs establishment of the ovarian reserve in their offspring. J. Dairy Sci. 2020, 103, 11957–11969. [Google Scholar] [CrossRef]
- Lamp, O.; Derno, M.; Otten, W.; Mielenz, M.; Nürnberg, G.; Kuhla, B. Metabolic heat stress adaption in transition cows: Differences in macronutrient oxidation between late-gestating and early-lactating German Holstein dairy cows. PLoS ONE 2015, 10, e0125264. [Google Scholar] [CrossRef]
- Jaton, C.; Schenkel, F.S.; Sargolzaei, M.; Cánovas, *!!! REPLACE !!!*; Malchiodi, F.; Price, C.A.; Baes, C.; Miglior, F. Genome-wide association study and in silico functional analysis of the number of embryos produced by Holstein donors. J. Dairy Sci. 2018, 101, 7248–7257. [Google Scholar] [CrossRef]
- La Marca, A.; Volpe, A. Anti-Müllerian hormone (AMH) in female reproduction: Is measurement of circulating AMH a useful tool? Clin. Endocrinol. (Oxf.) 2006, 64, 603–610. [Google Scholar] [CrossRef]
- Kastora, S.L.; Triantafyllidou, O.; Kolovos, G.; Kastoras, A.; Sigalos, G.; Vlahos, N. Combinational approach of retrospective clinical evidence and transcriptomics highlight AMH superiority to FSH, as successful ICSI outcome predictor. J. Assist. Reprod. Genet. 2020, 37, 1623–1635. [Google Scholar] [CrossRef]
- Luna-Nevárez, G.; Pendleton, A.L.; Luna-Ramirez, R.I.; Limesand, S.W.; Reyna-Granados, J.R.; Luna-Nevárez, P. Genome-wide association study of a thermo-tolerance indicator in pregnant ewes exposed to an artificial heat-stressed environment. J. Therm. Biol. 2021, 101, 103095. [Google Scholar] [CrossRef]
- Gobikrushanth, M.; Purfield, D.C.; Colazo, M.G.; Wang, Z.; Butler, S.T.; Ambrose, D.J. The relationship between serum insulin-like growth factor-1 (IGF-1) concentration and reproductive performance, and genome-wide associations for serum IGF-1 in Holstein cows. J. Dairy Sci. 2018, 101, 9154–9167. [Google Scholar] [CrossRef]
- Monte, A.P.O.; Barros, V.R.P.; Santos, J.M.; Menezes, V.G.; Cavalcante, A.Y.P.; Gouveia, B.B.; Bezerra, M.E.S.; Macedo, T.J.S.; Matos, M.H.T. Immunohistochemical localization of insulin-like growth factor-1 (IGF-1) in the sheep ovary and the synergistic effect of IGF-1 and FSH on follicular development in vitro and LH receptor immunostaining. Theriogenology 2019, 129, 61–69. [Google Scholar] [CrossRef]
- Dai, S.; Zhang, H.; Yang, F.; Shang, W.; Zeng, S. Effects of IGF-1 on the three-dimensional culture of ovarian preantral follicles and superovulation rates in mice. Biology 2022, 11, 833. [Google Scholar] [CrossRef]
- Shimizu, T. Molecular and cellular mechanisms for the regulation of ovarian follicular function in cows. J. Reprod. Dev. 2016, 62, 323–329. [Google Scholar] [CrossRef]
- Dan, H.C.; Antonia, R.J.; Baldwin, A.S. PI3K/Akt promotes feedforward mTORC2 activation through IKKα. Oncotarget 2016, 7, 21064–21075. [Google Scholar] [CrossRef]
- Le Roith, D. The insulin-like growth factor system. Exp. Diabesity Res. 2003, 4, 205–212. [Google Scholar] [CrossRef]
- Poudel, S.B.; Dixit, M.; Neginskaya, M.; Nagaraj, K.; Pavlov, E.; Werner, H.; Yakar, S. Effects of GH/IGF on the aging mitochondria. Cells 2020, 9, 1384. [Google Scholar] [CrossRef]
- Barrera, S.S.; Naranjo-Gomez, J.S.; Rondón-Barragán, I.S. Thermoprotective molecules: Effect of insulin-like growth factor type I (IGF-1) in cattle oocytes exposed to high temperatures. Heliyon 2023, 9, e14375. [Google Scholar] [CrossRef]
- Ascari, I.J.; Alves, N.G.; Jasmin, J.; Lima, R.R.; Quintão, C.C.R.; Oberlender, G.; Moraes, E.A.; Camargo, L.S.A. Addition of insulin-like growth factor I to the maturation medium of bovine oocytes subjected to heat shock: Effects on the production of reactive oxygen species, mitochondrial activity and oocyte competence. Domest. Anim. Endocrinol. 2017, 60, 50–60. [Google Scholar] [CrossRef]
- Lima, R.S.; Risolia, P.H.B.; Ispada, J.; Assumpção, M.E.O.A.; Visintin, J.A.; Orlandi, C.; Paula-Lopes, F.F. Role of insulin-like growth factor 1 on cross-bred Bos indicus cattle germinal vesicle oocytes exposed to heat shock. Reprod. Fertil. Dev. 2017, 29, 1405–1414. [Google Scholar] [CrossRef]
- Yang, S.; Yang, Y.; Hao, H.; Du, W.; Pang, Y.; Zhao, S.; Zou, H.; Zhu, H.; Zhang, P.; Zhao, X. Supplementation of EGF, IGF-1, and connexin 37 in IVM medium significantly improved the maturation of bovine oocytes and vitrification of their IVF blastocysts. Genes (Basel) 2022, 13, 805. [Google Scholar] [CrossRef]
- Flesken-Nikitin, A.; Hwang, C.I.; Cheng, C.Y.; Michurina, T.V.; Enikolopov, G.; Nikitin, A.Y. Ovarian surface epithelium at the junction area contains a cancer-prone stem cell niche. Nature 2013, 495, 241–245. [Google Scholar] [CrossRef]
- Sun, X.; Terakawa, J.; Clevers, H.; Barker, N.; Daikoku, T.; Dey, S.K. Ovarian LGR5 is critical for successful pregnancy. FASEB J. 2014, 28, 2380–2389. [Google Scholar] [CrossRef]
- Sun, X.; Jackson, L.; Dey, S.K.; Daikoku, T. In pursuit of leucine-rich repeat-containing G protein-coupled receptor-5 regulation and function in the uterus. Endocrinology 2009, 150, 5065–5073. [Google Scholar] [CrossRef]
- de Lau, W.; Barker, N.; Low, T.Y.; Koo, B.K.; Li, V.S.; Teunissen, H.; Kujala, P.; Haegebarth, A.; Peters, P.J.; van de Wetering, M.; et al. Lgr5 homologues associate with Wnt receptors and mediate R-spondin signalling. Nature 2011, 476, 293–297. [Google Scholar] [CrossRef]
- Wen, H.; Luo, H.; Yang, M.; Augustino, S.M.A.; Wang, D.; Mi, S.; Guo, Y.; Zhang, Y.; Xiao, W.; Wang, Y.; et al. Genetic parameters and weighted single-step genome-wide association study for supernumerary teats in Holstein cattle. J. Dairy Sci. 2021, 104, 11867–11877. [Google Scholar] [CrossRef]
- Risha, M.A.; Ali, A.; Siengdee, P.; Trakooljul, N.; Haack, F.; Dannenberger, D.; Wimmers, K.; Ponsuksili, S. Wnt signaling related transcripts and their relationship to energy metabolism in C2C12 myoblasts under temperature stress. PeerJ 2021, 14, e11625. [Google Scholar] [CrossRef]
- Erlebacher, A. Mechanisms of T cell tolerance towards the allogeneic fetus. Nat. Rev. Immunol. 2013, 13, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.Y.; Moldenhauer, L.M.; Groome, H.M.; Schjenken, J.E.; Robertson, S.A. Toll-like receptor-4 null mutation causes fetal loss and fetal growth restriction associated with impaired maternal immune tolerance in mice. Sci. Rep. 2021, 11, 16569. [Google Scholar] [CrossRef]
- Robertson, S.A.; Care, A.S.; Moldenhauer, L.M. Regulatory T cells in embryo implantation and the immune response to pregnancy. J. Clin. Investig. 2018, 128, 4224–4235. [Google Scholar] [CrossRef]
- Ramachandran, G. Gram-positive and gram-negative bacterial toxins in sepsis: A brief review. Virulence 2014, 5, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Wittebole, X.; Castanares-Zapatero, D.; Laterre, P.F. Toll-like receptor 4 modulation as a strategy to treat sepsis. Mediators Inflamm. 2010, 2010, 568396. [Google Scholar] [CrossRef]
- Li, W.H.; Liu, Y.L.; Lun, J.C.; He, Y.M.; Tang, L.P. Heat stress inhibits TLR4-NF-κB and TLR4-TBK1 signaling pathways in broilers infected with Salmonella Typhimurium. Int. J. Biometeorol. 2021, 65, 1895–1903. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, G.; Wang, Z.; Guo, X.; Xu, Q.; Huang, Q.; Su, L. NF-κB signaling is essential for resistance to heat stress-induced early stage apoptosis in human umbilical vein endothelial cells. Sci. Rep. 2015, 5, 13547. [Google Scholar] [CrossRef]
- Liu, Z.; Ezernieks, V.; Wang, J.; Arachchillage, N.W.; Garner, J.B.; Wales, W.J.; Cocks, B.G.; Rochfort, S. Heat stress in dairy cattle alters lipid composition of milk. Sci. Rep. 2017, 7, 961. [Google Scholar] [CrossRef]
- Faylon, M.P.; Baumgard, L.H.; Rhoads, R.P.; Spurlock, D.M. Effects of acute heat stress on lipid metabolism of bovine primary adipocytes. J. Dairy Sci. 2015, 98, 8732–8740. [Google Scholar] [CrossRef]
- Hansen, P.J. Reproductive physiology of the heat-stressed dairy cow: Implications for fertility and assisted reproduction. Anim. Reprod. 2019, 16, 497–507. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Bowman, P.J.; Haile-Mariam, M.; Pryce, J.E.; Hayes, B.J. Genomic selection for tolerance to heat stress in Australian dairy cattle. J. Dairy Sci. 2016, 99, 2849–2862. [Google Scholar] [CrossRef] [PubMed]
- Weller, J.I.; Song, J.Z.; Heyen, D.W.; Lewin, H.A.; Ron, M. A new approach to the problem of multiple comparisons in the genetic dissection of complex traits. Genetics 1998, 150, 1699–1706. [Google Scholar] [CrossRef] [PubMed]
- Minozzi, G.; Nicolazzi, E.L.; Stella, A.; Biffani, S.; Negrini, R.; Lazzari, B.; Ajmone-Marsan, P.; Williams, J.L. Genome wide analysis of fertility and production traits in Italian Holstein cattle. PLoS ONE 2013, 8, e80219. [Google Scholar] [CrossRef] [PubMed]
- Visscher, P.M. Sizing up human height variation. Nat. Genet. 2008, 40, 489–490. [Google Scholar] [CrossRef]
- Karlsson, E.K.; Baranowska, I.; Wade, C.M.; Salmon-Hillbertz, N.H.; Zody, M.C.; Anderson, N.; Biagi, T.M.; Patterson, N.; Pielberg, G.R.; Kulbokas, E.J.; et al. Efficient mapping of Mendelian traits in dogs through genome-wide association. Nat. Genet. 2007, 39, 1321–1328. [Google Scholar] [CrossRef]
Figure 1.
Ambient temperature (AT, °C), relative humidity (RH, %), and temperature-humidity index (THI, units) during the study. The straight line indicates the threshol between moderate and severe heat stress.
Figure 1.
Ambient temperature (AT, °C), relative humidity (RH, %), and temperature-humidity index (THI, units) during the study. The straight line indicates the threshol between moderate and severe heat stress.
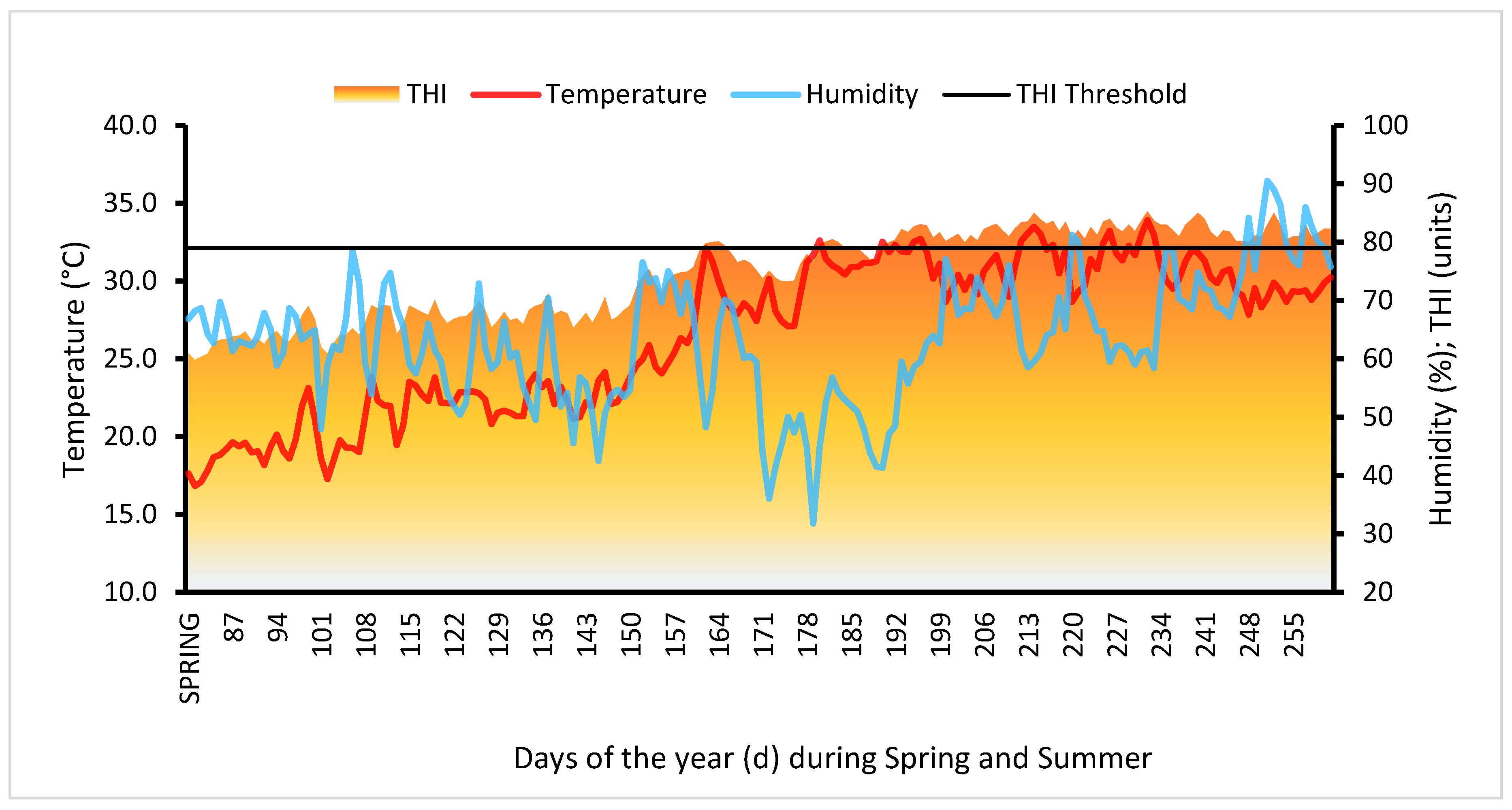
Figure 2.
Regression relationships between Anti-Müllerian hormone (AMH) and ovarian variables according to heat-stressed group. A) Follicle diameter (moderate HS), B) Corpus Luteum diameter (moderate HS), C) Follicle diameter (severe HS), D) Corpus Luteum diameter (severe HS).
Figure 2.
Regression relationships between Anti-Müllerian hormone (AMH) and ovarian variables according to heat-stressed group. A) Follicle diameter (moderate HS), B) Corpus Luteum diameter (moderate HS), C) Follicle diameter (severe HS), D) Corpus Luteum diameter (severe HS).
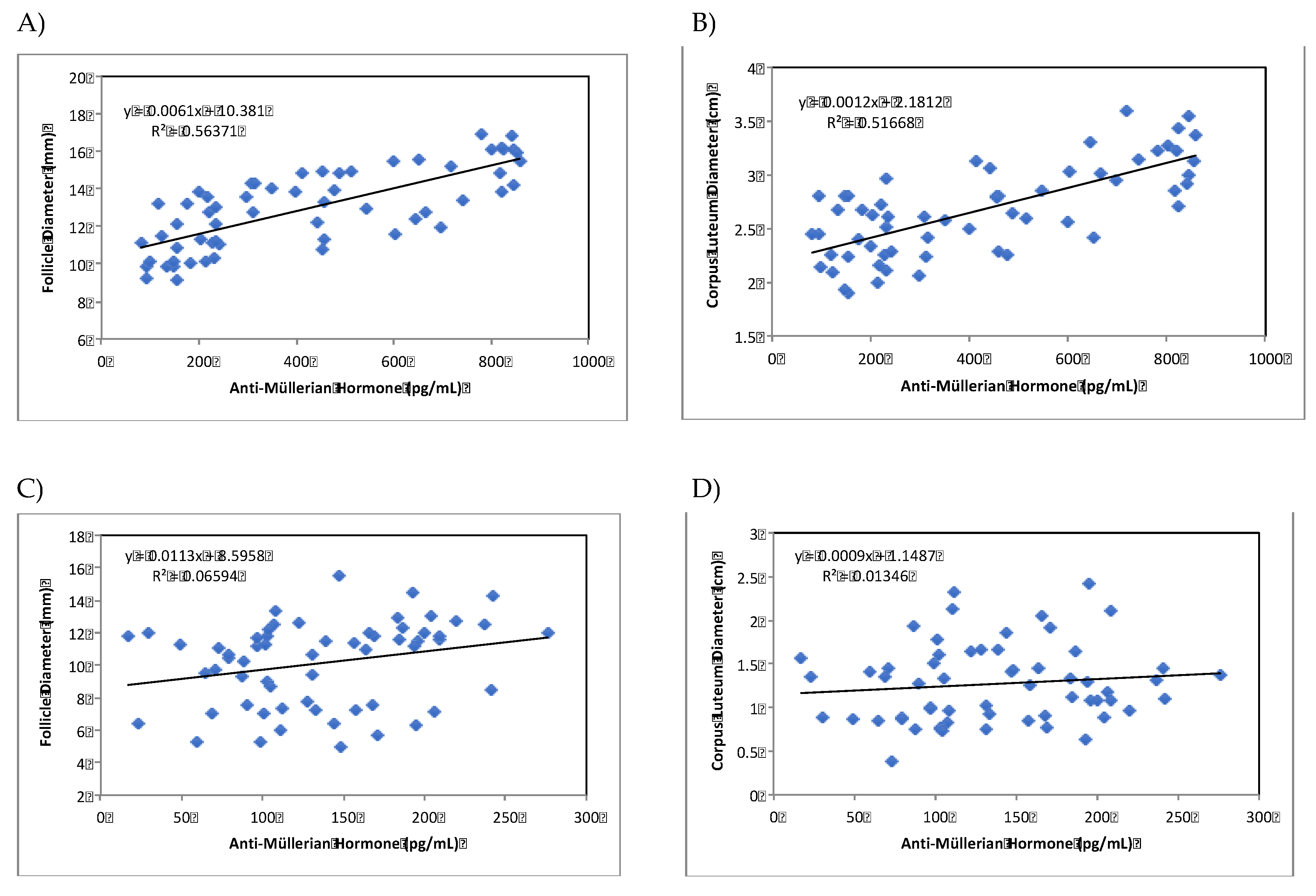
Figure 3.
Manhattan plot from single-marker GWAS for Anti-Mülleriam hormone (AMH) serum concentrations in Holstein cows managed during summer in a semi-desert region.
Figure 3.
Manhattan plot from single-marker GWAS for Anti-Mülleriam hormone (AMH) serum concentrations in Holstein cows managed during summer in a semi-desert region.
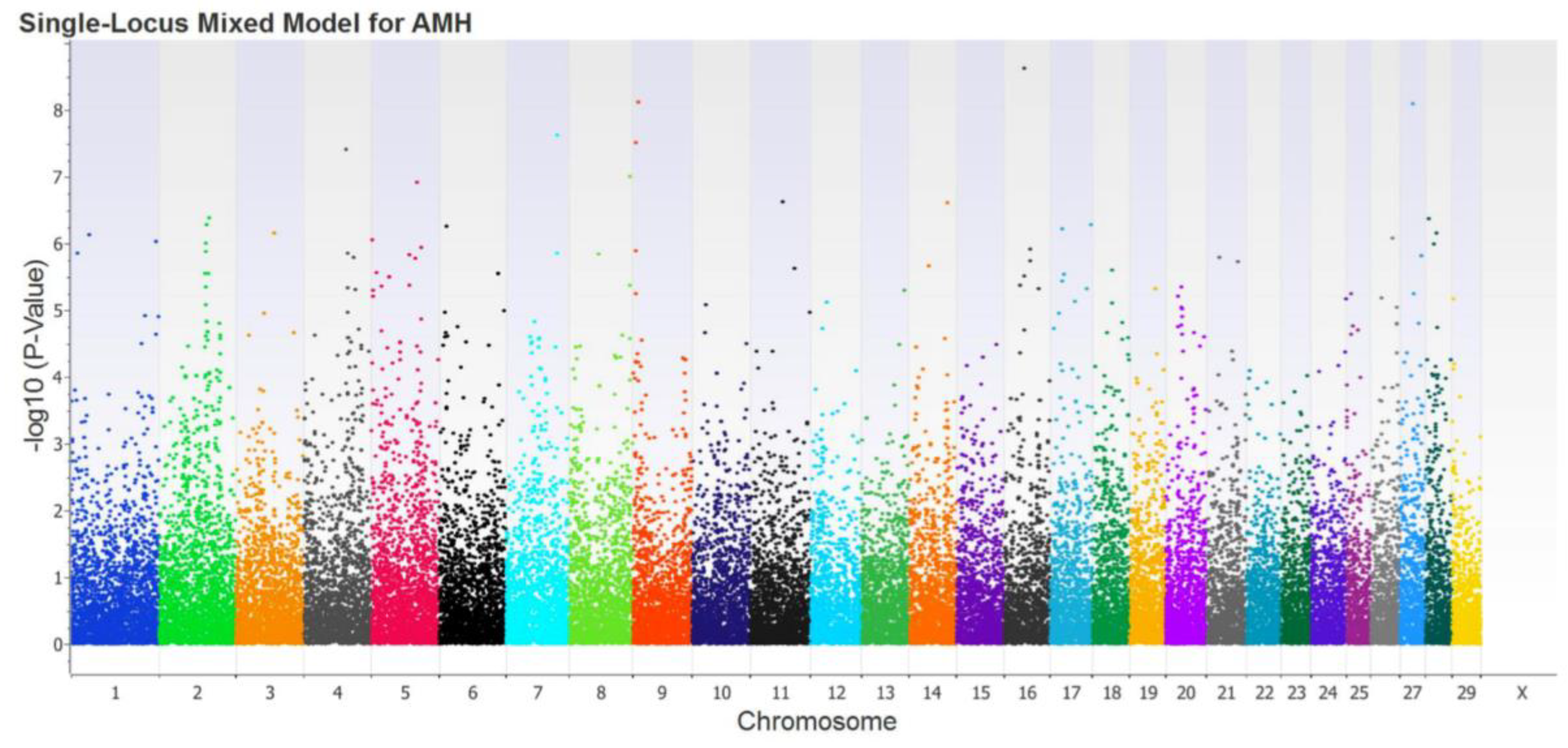
Figure 4.
Average values for serum AMH concentrations and reproductive traits (± SE) during the expeimental period in Holstein cows according to the number of favorable genotype SNP markers (i.e., 0, 1, 2, 3 or 4). A) Serum AMH concentrations (pg/ml); B) Follicle diameter (mm); C) Corpus Luteum diameter (cm); D) Conception rate (%).
Figure 4.
Average values for serum AMH concentrations and reproductive traits (± SE) during the expeimental period in Holstein cows according to the number of favorable genotype SNP markers (i.e., 0, 1, 2, 3 or 4). A) Serum AMH concentrations (pg/ml); B) Follicle diameter (mm); C) Corpus Luteum diameter (cm); D) Conception rate (%).
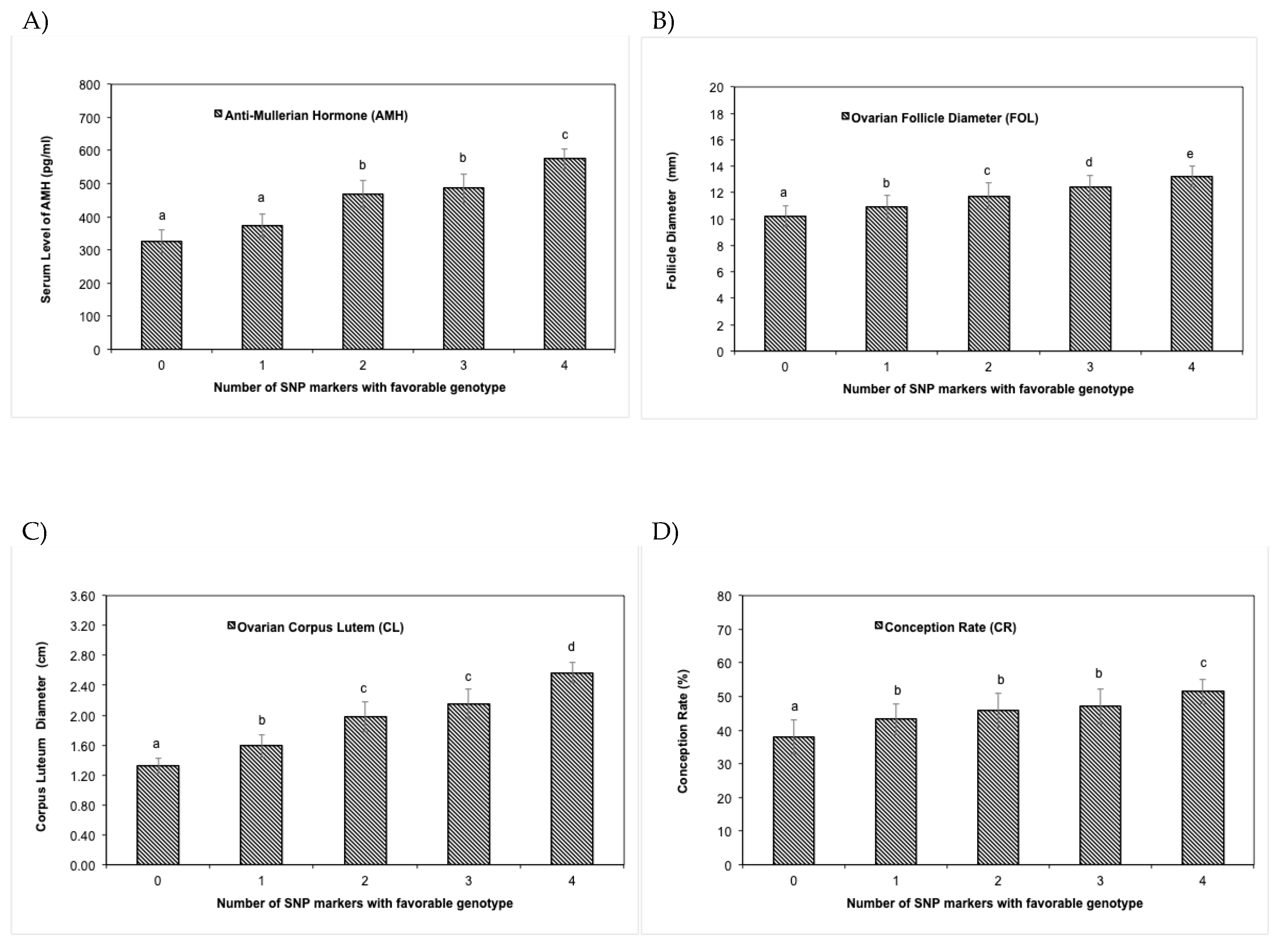

Table 1.
Serum AMH, reproductive traits and physiological variable in Holstein cows according to heat-stressed environmental conditions.
Table 1.
Serum AMH, reproductive traits and physiological variable in Holstein cows according to heat-stressed environmental conditions.
| Variable | Moderate heat stress (n=152; THI = 72-79) |
Severe heat stress (n=128; THI > 79) |
|---|---|---|
| AMH (ng/ml) | 417.26 ± 4.51 a | 136.94 ± 4.03 b |
| FOL (mm) | 12.95 ± 0.93 a | 10.15 ± 1.02 b |
| CL (cm) | 2.68 ± 0.02 a | 1.26 ± 0.04 b |
| CR (%) | 52.63 a | 37.50 b |
| RT (°C) | 38.21 ± 1.25 a | 38.73 ± 3.05 b |
| RR (breaths/min) | 63.35 ± 3.14 a | 67.14 ± 4.96 b |
AMH = Serum Anti-Müllerian hormone; FOL = Follicle diameter; CL = Corpus Luteum diameter; CR = Conception rate; RT = Rectal temperature; RR = Respiratory rate. a, b Different literals between columns indicate a significant statistical difference (p < 0.05).
Table 2.
Pearson correlations between serum Anti-Müllerian hormone (AMH) with ovarian and physiological traits in Holstein cows according to climatic experimental groups (HS = heat stress).
Table 2.
Pearson correlations between serum Anti-Müllerian hormone (AMH) with ovarian and physiological traits in Holstein cows according to climatic experimental groups (HS = heat stress).
| Variable | Moderate HS (p-value) | Severe HS (p-value) |
|---|---|---|
| FOL (mm) CL (cm) RT (°C) RR (breaths/min) |
0.7508 (<0.0001) 0.7188 (<0.0001) 0.3427 (0.0142) 0.2135 (0.1938) |
0.2567 (0.0837) 0.1160 (0.2155) 0.0958 (0.2671) 0.0634 (0.5728) |
FOL = Follicle diameter; CL = Corpus Luteum diameter; RT = Rectal temperature; RR = Respiratory rate.
Table 3.
Significant SNPs (p < 1.09 × 10−6) from a single-marker genome-wide association study (GWAS) with Anti-Müllerian hormone (AMH) in Holstein cows managed in a warm environment.
Table 3.
Significant SNPs (p < 1.09 × 10−6) from a single-marker genome-wide association study (GWAS) with Anti-Müllerian hormone (AMH) in Holstein cows managed in a warm environment.
| SNP ID 1 | Variant 2 | BTA 3 | Position 4 | Gene 5 | Alleles 6 | Var 7 | p-Value 8 |
|---|---|---|---|---|---|---|---|
| rs41807005 | Intergenic | 16 | 34′845050 | -------- | T/C | 0.162 | 2.35 × 10−9 |
| rs109740021 | Intergenic | 9 | 9′329501 | -------- | A/G | 0.157 | 7.53 × 10−9 |
| rs42416336 | Intronic | 27 | 23′354986 | LONRF1 | A/C | 0.152 | 8.16 × 10−9 |
| rs876084180 | Intronic | 7 | 21′401999 | AMH | A/C | 0.143 | 2.38 × 10−8 |
| rs136263395 | Intergenic | 9 | 4′423733 | -------- | T/C | 0.141 | 3.11 × 10−8 |
| rs445674221 | Intronic | 4 | 76′133069 | IGFBP1 | A/G | 0.139 | 3.85 × 10−8 |
| rs8193046 | Intronic | 8 | 108′833985 | TLR4 | T/C | 0.131 | 9.86 × 10−8 |
| rs42849475 | Intronic | 5 | 1′087211 | LGR5 | A/G | 0.129 | 1.20 × 10−7 |
| rs42338999 | Intergenic | 11 | 58′684365 | -------- | T/C | 0.124 | 2.38 × 10−7 |
| rs135450328 | Intergenic | 14 | 68′794329 | -------- | A/C | 0.123 | 2.46 × 10−7 |
| rs478504266 | Intronic | 2 | 89’091847 | SGO2 | G/A | 0.119 | 4.15 × 10−7 |
| rs137194049 | Intronic | 28 | 4′632166 | DISC1 | T/C | 0.118 | 4.24 × 10−7 |
| rs135441773 | Intronic | 2 | 84′744596 | SLC39A10 | T/G | 0.117 | 5.22 × 10−7 |
| rs110893810 | Intronic | 17 | 73′804897 | RSPH14 | T/C | 0.116 | 5.24 × 10−7 |
| rs136745124 | Intergenic | 6 | 12′650752 | -------- | A/G | 0.115 | 5.49 × 10−7 |
| rs43475092 | Intergenic | 3 | 68′078177 | -------- | T/C | 0.114 | 7.04 × 10−7 |
1 SNP reference of the NCBI; 2 SNP chromosome variant; 3 Bos taurus autosomal chromosome number; 4 SNP position within the chromosome; 5 candidate gene symbol (LONRF1 = LON peptidase N-terminal domain and ring finger 1; AMH = Anti-mullerian hormone; IGFBP1 = Insulin-like growth factor binding protein 1; TLR4 = toll-like receptor 4; LGR5 = Leucine rich repeat containing G protein-coupled receptor 5; SGO2 = Shugoshin 2; DISC1 = DISC1 scaffold protein; SLC39A10 = Solute carrier family 39 member 10; RSPH14 = Radial spoke head 14 homolog). 6 alleles from the SNP; 7 percentage of trait variance explained by the SNP; 8 SNP statistical significance.
Table 4.
Identification, gene name, favorable SNP allele, allele frequencies, and Hardy–Weinberg equilibrium analysis for genomic SNPs associated with serum AMH concentrations.
Table 4.
Identification, gene name, favorable SNP allele, allele frequencies, and Hardy–Weinberg equilibrium analysis for genomic SNPs associated with serum AMH concentrations.
| SNP ID 1 | Gene 2 | F. Allele 3 | Allele Frequency 4 | HWE Test 5 | HWE p-Value 6 | |
|---|---|---|---|---|---|---|
| A | C | |||||
| rs876084180 | AMH | T | 0.35 | 0.65 | 0.32 | 0.46 |
| A | G | |||||
| rs445674221 | IGFBP1 | T | 0.53 | 0.47 | 2.63 | 0.18 |
| rs42849475 | LGR5 | T | 0.24 | 0.76 | 1.25 | 0.31 |
| rs8193046 | TLR4 | T | 0.39 | 0.61 | 0.86 | 0.39 |
1 SNP reference of the NCBI; 2 Gene symbol name; 3 F. Allele = allele with the favorable effect on phenotype; 4 Frequency of both alleles within cow population; 5 Hardy–Weinberg equilibrium “χ2” test value; 6 “χ2” test p-value with 1 degree of freedom and α = 0.05.
Table 5.
Least-square means ± SE according to SNP’s marker genotypes for serum AMH and reproductive traits in Holstein cattle from a validation population.
Table 5.
Least-square means ± SE according to SNP’s marker genotypes for serum AMH and reproductive traits in Holstein cattle from a validation population.
| SNP ID 1 | Trait 2 | Least-Square Means by Genotype ± SE 3 | p-Value 4 | AlleleSE | AdditiveFE | ||
|---|---|---|---|---|---|---|---|
| AA | AC | CC | |||||
| rs876084180 | AMH | 501.26 ± 45.3 a | 382.49 ± 37.9 b | 152.94 ± 19.2 c | <0.0001 | 165.12 | 174.16* |
| FOL | 13.01 ± 1.06 a | 11.95 ± 1.04 b | 10.17 ± 0.98 c | 0.0010 | 1.37 | 1.42* | |
| CL | 2.53 ± 0.17 a | 1.92 ± 0.16 ab | 1.47 ± 0.19 b | 0.0086 | 0.50 | 0.53* | |
| CR | 45.20 ± 3.67 a | 36.75 ± 3.32 a | 40.65 ± 4.01 a | 0.6519 | 2.21 | 2.27 | |
| AA | AG | GG | |||||
| rs445674221 | AMH | 141.80 ± 19.6 a | 267.39 ± 23.2 a | 442.65 ± 37.8 b | 0.0009 | 146.01 | 150.42* |
| FOL | 10.63 ± 0.96 a | 11.04 ± 1.05 a | 13.91 ± 1.12 b | 0.0068 | 1.60 | 1.64* | |
| CL | 1.24 ± 0.09 a | 1.82 ± 0.07 b | 2.89 ± 1.18 c | 0.0087 | 0.72 | 0.77* | |
| CR | 34.50 ± 0.47 a | 45.15 ± 0.46 b | 56.25 ± 0.42 c | <0.0001 | 10.55 | 10.87* | |
| rs42849475 | AMH | 291.55 ± 26.7 a | 342.16 ± 37.5 a | 401.29 ± 39.2 a | 0.1506 | 52.76 | 54.87* |
| FOL | 9.86 ± 0.87 a | 11.16 ± 1.09 b | 12.64 ± 1.26 b | 0.0079 | 1.34 | 1.39* | |
| CL | 1.38 ± 0.07 a | 1.92 ± 1.05 b | 2.24 ± 1.90 b | 0.0096 | 0.38 | 0.43* | |
| CR | 39.70 ± 3.07 a | 46.20 ± 3.59 b | 50.25 ± 4.36 b | 0.0188 | 4.91 | 5.27* | |
| rs8193046 | AMH | 474.67 ± 38.7 a | 379.23 ± 37.3 b | 158.34 ± 14.2 c | <0.0001 | 151.78 | 158.16* |
| FOL | 13.28 ± 1.29 a | 11.37 ± 0.94 b | 10.14 ± 0.94 c | 0.0065 | 1.52 | 1.57* | |
| CL | 2.71 ± 2.02 a | 1.86 ± 1.45 b | 1.20 ± 1.08 c | 0.0012 | 0.67 | 0.75* | |
| CR | 54.15 ± 3.67 a | 45.70 ± 3.29 a | 36.75 ± 3.18 b | 0.0034 | 8.45 | 8.70* | |
1 SNP reference of the NCBI; 2 phenotypic traits (AMH = serum Anti-Mullerian hormone, ng/ml; FOL = Follicular diameter, mm; CL = Corpus Luteum diameter, cm; CR = Conception rate, %); 3 least-square means according to SNP genotype ± SE (a,b,c indicate statistical difference among genotypes at); 4 p-value = statistical significance; 5 Allele substitution effect; 4 Additive fixed estimated effect.
Table 6.
Least-square means ± SE according to SNP markers genotypes for physiological traits in Holstein cattle from a validation population.
Table 6.
Least-square means ± SE according to SNP markers genotypes for physiological traits in Holstein cattle from a validation population.
| SNP ID 1 | Trait 2 | Least-Square Means by Genotype ± SE 3 | p-Value 4 | Allele SE | Additive FE | ||
|---|---|---|---|---|---|---|---|
| AA | AC | CC | |||||
| rs876084180 | RT | 38.14 ± 2.98 a | 38.25 ± 2.67 a | 38.79 ± 3.39 b | 0.0316 | -0.28 | 0.32* |
| RR | 63.67 ± 5.22 a | 65.44 ± 5.19 a | 66.35 ± 5.19 a | 0.0928 | -1.63 | 1.69* | |
| AA | AG | GG | |||||
| rs445674221 | RT | 38.89 ± 3.02 a | 38.46 ± 2.96 b | 38.15 ± 2.77 c | 0.0094 | -0.34 | 0.37* |
| RR | 68.32 ± 4.98 a | 65.01 ± 4.67 b | 61.96 ± 4.12 c | 0.0067 | -3.06 | 3.18* | |
| rs42849475 | RT | 38.81 ± 3.54 a | 38.39 ± 2.69 b | 38.26 ± 3.82 b | 0.0235 | -0.24 | 0.27* |
| RR | 65.79 ± 4.28 a | 64.03 ± 5.02 a | 62.99 ± 4.67 a | 0.2138 | -1.35 | 1.40* | |
| rs8193046 | RT | 38.16 ± 2.65 a | 38.49 ± 3.08 b | 38.93 ± 3.19 c | 0.0076 | -0.35 | 0.38* |
| RR | 61.24 ± 5.09 a | 64.96 ± 5.16 b | 68.95 ± 4.87 c | 0.0052 | -3.69 | 3.85* | |
1 SNP reference of the NCBI; 2 phenotypic traits (RT = rectal temperature, °C; RR = respiration rate, breaths/min); 3 least-square means according to SNP genotype ± SE (a,b,c indicate statistical difference among genotypes); 4 p-value = statistical significance; 5 Allele substitution effect; 4 Additive fixed estimated effect.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



