
Submitted:
18 October 2023
Posted:
19 October 2023
You are already at the latest version
Abstract
The goal of our study was to identify signatures of selection in the Turopolje pigs and other commercial pig breeds. We conducted a comprehensive analysis of five datasets, including one local pig breed (Turopolje) and four commercial pig breeds (Large White, Landrace, Pietrain, and Duroc), using strict quality control measures. Our final dataset consisted of 485 individuals and 54,075 single nucleotide polymorphisms (SNPs). To detect selection signatures within these pig breeds, we utilized the XP-EHH and XP-nSL methodologies, which allowed us to identify candidate genes that have been subject to positive selection. Our analysis consistently highlighted the PTBP2 and DPYD genes as commonly targeted by selection across all studied breeds. Both of these genes are associated with muscular development in pigs and other species. Furthermore, in the Large White breed a number of genes were detected by the two methods, such as ATP1A1, CASQ2, CD2, IGSF3, MAB21L3, NHLH2, SLC22A15, VANGL1. In Duroc breed a different set of genes was detected, such as ARSB, BHMT, BHMT2, DMGDH, JMY. The function of these genes was related to body weight, production efficiency and meat quality, average daily gain and other similar traits. Overall, our results have identified a number of genomic regionsthat are under selective pressure between local and commercial pig breeds. This information can help to improve our understanding of the mechanisms underlying pig breeding, and ultimately contribute to the development of more efficient and sustainable pig production practices. Our study highlights the power of using multiple genomic methodologies to detect genetic signatures of selection, and provides important insights into the genetic diversity of pig breeds.
Keywords:
Selection Signatures
; Genomic Analysis
; Muscular Development Genes
1. Introduction
Studying selective sweeps in domesticated animals is a valuable tool for identifying genomic regions that have been influenced by natural and artificial selection. Such studies can uncover important mutations, genes, and pathways associated with economically significant traits [1]. Over the years, commercial pig breeding programs have been developed to improve desirable traits such as growth rate, meat quality, and disease resistance, in order to meet the increasing demand for pig products. Selection strategies within breeds and also crossbreeding are commonly used to achieve these goals [2]. These adaptations can result from both hard and soft selective pressures. Hard selective pressures involve the complete removal of deleterious alleles or fixation of advantageous alleles, leading to a rapid and strong selective sweep. On the other hand, soft selective pressures involve selection on multiple alleles with smaller effects, resulting in a more gradual and less pronounced selective sweep [3]. However, there are concerns that the intense selection pressure in commercial pig breeding programs may also lead to unintended consequences, such as a decrease in genetic diversity and increased susceptibility to disease [4]. Positive selection tends to favor a small number of highly productive animals, which can result in the loss of rare genetic variants that may be important for adaptation to changing environments or resistance to diseases.
Understanding the genetic mechanisms underlying these selective pressures can provide valuable insights into the evolutionary history of pigs and inform breeding strategies for improving pig health and productivity. Selective sweeps are important evolutionary events that contribute to the adaptation of populations to changing environments, and many methods are available to study the genetic mechanisms underlying these events. One such method is cross-population extended haplotype homozygosity (XP-EHH), the extension of the EHH method. This is a haplotype-based approach that measures the difference in haplotype length between two populations and identifies genomic regions that have undergone recent positive selection [5]. XP-EHH has been successfully used to detect selective sweeps in a variety of species, including humans [6], sheep [7], and pigs [8]. Another method that can be used to capture genomic patterns resulting from local selective sweeps is cross-population number of segregating sites by length (XP-NSL) is an extension of haplotype-based statistic (nSL). This method is useful for performing a genomic scan to identify regions of local adaptation by comparing haplotype patterns between two populations. XP-nSL has been shown to have good power in detecting both ongoing and recently completed hard and soft sweeps, making it a valuable tool for studying the genetic mechanisms underlying adaptation to different environments [9]. The ability to focus on recent selection pressures could be especially valuable in the analysis of the Turopolje breed, as the Austrian population was established only in the 1990s.
Integrating these two methods can provide us with a greater confidence in identifying genomic regions that have undergone positive selection. Overall, understanding the genetic basis of desirable traits in domesticated animals is essential for improving breeding programs, and the detection of selection signatures can play an important role in achieving this goal.
The aim of the study was to identify genomic regions and genes, that markedly differ between the Austrian Turopolje breed, and the commercial breeds. find selection signatures commercial breed
2. Materials and Methods
2.1. Data
The present study leveraged genomic data from Austrian Turopolje and commercial pig populations. The commercial dataset comprised Large White (82 individuals), Landrace (76 individuals), Pietrain (69 individuals), and Duroc (74 individuals), while the local population (Turopolje) consisted of 184 individuals.We conducted quality control using PLINK with the following parameters; only autosomal variants were retained, variants with less than 10% missing data (--mind 0.1) and individuals with less than 10 % missing genotypic data were retained (--geno 0.1). After conducting quality control measures total of 54,075 single nucleotide polymorphisms (SNPs) were retained for analysis, with 485 individuals included in the final dataset. The genotyping rate for the dataset was 0.996.
2.2. Methods
In this study, two methods were used to identify regions under selection in the genomes of commercial and local pigs, cross-population extended haplotype homozygosity (XP-EHH) and cross-population number of segregating sites by length (XP-nSL). These methods were implemented in the software Selscan v.2.080 [10].
To perform haplotype phasing, Beagle v.5.4 [11] was used. Two approaches were used to determine the selection signatures with both methods: (1) Each commercial population was compared with the Turopolje breed separately; (2) and then all populations were combined into one group, and jointly compared to Turopolje.
Normalization of both methods scores across all chromosomes was performed using the "norm" program distributed along with Selscan. Normalized XP-EHH scores were considered as the signature of selection.
XP-nSL statistics were used to detect local adaptation by comparing haplotype patterns between two populations around the same allele of interest. XP-nSL, designed to identify genomic regions involved in local adaptation between two populations. Building upon the nSL method used for within-population analysis, XP-nSL compares haplotype sets from different populations, calculating statistics based on ancestral and derived alleles. Computation involves assessing the similarity of consecutive sites within haplotypes, followed by normalization across the genome [9]. This method has the power to detect both ongoing and recently completed hard and soft sweeps.
In total, 68 windows were analyzed using these methods to identify regions under selection in the genomes of commercial and local pigs.
3. Results
Our research revealed that commercial breeds had multiple regions under selection, as shown in Figure 1, Figure 2 and Figure 3 for Large White, Duroc, and the entire population, respectively. The Manhattan plot for Pietrain and Landrace is available in the supplementary section.
In our study, xp-EHH and xp-nSL detected 133 and 53 genes, respectively, as candidates for signatures of selection. The selected candidate regions are distributed across different chromosomes. There were 15 common genomic regions between two methods, indicating both positive and divergent selection acted on these regions. These candidate regions were located on chromosomes 2 and 4 (Table 1)
The XP-EHH and XP-nSL approaches consistently highlighted PTBP2 and DPYD genes as commonly targeted by selection across all studied breeds (Table 2).
In Large White breed a number of genes were detected by both selection signature methods used in this study, such as: ATP1A1, CASQ2, CD2, IGSF3, MAB21L3, NHLH2, SLC22A15, VANGL1 (Table 2).
In the Duroc breed, a distinct set of genes showed similarity in our study between the two methods, including ARSB, BHMT, BHMT2, DMGDH, and JMY (Table 2).
4. Discussion
The candidate regions we have identified are spread out across various chromosomes. This distribution suggests that both natural selection and random mating may have played a partial role in shaping the genomes of these breeds [12]. In our study, we identified 15 common genomic regions that exhibited evidence of both positive and divergent selection when analyzed with two distinct methods. These regions were specifically localized on chromosomes 2 and 4, as detailed in Table 2. Notably, our analysis using the XP-EHH and XP-nSL approaches consistently highlighted the PTBP2 and DPYD genes as recurrently favored targets of selection across all breeds studied, underscoring the robustness and reliability of these findings.
A study by Santana et al. [13] suggested that genetic variation in the DPYD gene could influence muscle growth and development in Nellore cattle, and might be associated with rib eye area as a measure of muscle quantity. The authors found significant associations between a single nucleotide polymorphism (SNP) in the DPYD gene and rib eye area in Nellore cattle, providing further support for the potential relevance of this gene in pig breeding. Studies showed that the DPYD is one of the candidate genes in Duroc pig population that were detected within or nearby significant SNPs related to intramuscular fat (IMF) metabolism. Also, the DPYD is related to muscle contraction, muscle system process, developmental process, and sphingolipid signalling pathway [14]. Furthermore, the studies showed that PTBP2 may play a role in regulating alternative splicing of specific muscle-specific genes during development and regeneration and the potential involvement of PTBP2 in various muscular diseases in human, including myotonic dystrophy and spinal muscular atrophy. The dysregulation of PTBP2-mediated splicing could contribute to the pathology of these diseases [15].
In Large White breed a number of genes were detected by both selection signature methods used in this study, such as: ATP1A1, CASQ2, CD2, IGSF3, MAB21L3, NHLH2, SLC22A15, VANGL1 (Table 2).
Study by Shanshan et al [16] suggested that the ATP1A1 gene is a protein-coding gene that encodes the alpha-1 subunit of the Na+/K+-ATPase enzyme, which is involved in the active transport of sodium and potassium ions across cell membranes. Also, there is a disrupted potassium ion homeostasis in the ciliary muscle in negative lens-induced myopia, and that this may be related to changes in the expression and activity of Na+/K+-ATPase, which is encoded by the ATP1A1 gene. Specifically, they noted that there is decreased expression of ATP1A1 at both the mRNA and protein levels in the ciliary muscles with negative lens induced myopia, as well as reduced Na+/K+-ATPase activity. These findings suggest that a disruption of potassium ion homeostasis may be contributing to the development of myopia in these animals, and that changes in the expression and activity of ATP1A1 may be involved in this process [16]. CASQ2 gene same with DPYD revealed significant enrichment in KEGG pathways in relation to muscle contraction, muscle system process, developmental process, and sphingolipid signalling pathway [14]. In investigating intramuscular fat (IMF) and pork quality, researchers identified sex-specific biomarkers related to IMF in pigs. Among these, the CD2 molecule (CD2) emerged as a notable biomarker for high-IMF pigs, confirmed via qRT-PCR and shedding light on IMF deposition mechanisms for enhanced meat quality [17]. A genomic study by Fontanesi et al. [18] delved into the Average Daily Gain of Italian Large White pigs, suggesting a connection between the IGSF3 gene and variations in average daily gain.
The results underscored that the identified candidate genes primarily revolve around pig production and meat quality, indicating targeted selective pressures in commercial pig breeds. These genes appear to have undergone significant selection to enhance production efficiency and meat quality.
Our result also showed selection pressure on NHLH2, SLC22A15 and VANGL1 in Large White pigs, which located on chromosome 4. The NHLH2 gene encodes a basic helix-loop-helix transcription factor implicated in regulating adult body weight and fertility, based on mouse knockout studies. Notably, two SNPs in the NHLH2 gene have functional implications. One SNP within the 3' untranslated region (3'UTR) results in increased mRNA instability and reduced protein levels when linked to luciferase mRNA. The other SNP in the coding region leads to an amino acid change affecting protein migration and secondary structure. One of these SNPs is associated with obesity in humans, while the other is prevalent among individuals. These findings suggest a potential role for NHLH2 in body weight regulation and warrant further investigation into their influence on human weight and fertility phenotypes [19]. In the pig population studied, the gene SLC22A15 was examined as part of the validation process using qRT-PCR to confirm the RNA-Seq results. SLC22A15 was identified as one of the down-regulated genes. The qRT-PCR analysis of SLC22A15's expression in pigs showed consistent results with the transcriptomic data, indicating that changes in SLC22A15 expression due to different feeding patterns were reliably captured using both RNA-Seq and qRT-PCR methods. This highlights the role of SLC22A15 in responding to varying feeding regimens within the pig population [20].
In other study by Bergamaschi et al. [21] results showed that the composition of the gut microbiome was influenced by the host's genetics, and several SNPs were found to be associated with specific microbial taxa at different time points during the pigs' growth. These SNPs were located within genomic regions that contained a total of 68 genes. One of these genes, VANGL1, located in a genomic region on SSC4, was identified as part of the Wnt signaling pathway. VANGL1 was found to be highly expressed in gut tissues and has been linked to processes such as cell proliferation and turnover. Interestingly, the study highlighted that VANGL1 was associated with specific microbial Operational Taxonomic Units (OTUs), particularly those classified as Anaerostipes. This finding suggested a connection between VANGL1 and the abundance of certain gut microbes.
In the Duroc breed, a distinct set of genes showed similarity in our study between the two methods, including ARSB, BHMT, BHMT2, DMGDH, and JMY. Similar to our results, the study explored genetic diversity and selection signals in pig populations, particularly highlighting the genes ARSB, DMGDH, and BHMT within the supergene region. Using ROH analysis on data from American Duroc (AD) and Canadian Duroc (CD) pig populations were studied. AD pigs exhibited higher genetic diversity and lower inbreeding compared to CD pigs, likely due to distinct selective pressures. In the overlapping ROH hotspots of both populations, a significant missense mutation (rs81216249) was identified in the supergene region containing ARSB, DMGDH, and BHMT genes. This variant allele, originating from European pigs, was nearly fixed in Duroc pigs. Strong selection on this supergene was indicated, possibly contributing to improvements in body weight and length. These findings suggest the potential for genetic enhancement of pigs using these genes as trait-related markers. They identified a significant genetic variant situated within the overlapping section of three genes: ARSB, DMGDH, and BHMT. Specifically, the variant manifested as a missense mutation within the DMGDH gene. Analysis of various genetic indicators pointed towards the positive selection acting upon this genetic region encompassing the three genes (ARSB-DMGDH-BHMT). This observation suggested that this region had undergone a process of favorable selection.
This observation strongly implied a history of favorable selection shaping this genomic region. Notably, these findings mirror our own outcomes, where both XP-EHH and XP-nSL methodologies demonstrated a similar pattern of positive selection on these genes, as showcased in Figure 3. The results showed a significant genetic variant within the ARSB-DMGDH-BHMT gene cluster that underwent positive selection, as evidenced by various genetic indicators and the uniform genetic haplotype among Duroc pigs. This gene cluster's influence on growth and fat deposition traits adds to its biological significance [22].
The gene JMY also has potential involvement in muscle development, along with other genes like HOMER1, as well as ITGA1 and RAB32. These genes are associated with functions such as glycogen metabolism and mitochondrial dynamics, which are relevant to muscle health and development [23].
BHMT2, functioning as a methyl transferase, facilitates the transfer of a methyl group from betaine to homocysteine, resulting in the production of methionine. This biochemical process intricately interweaves with essential metabolic pathways involving dimethylglycine, betaine, and methionine. BHMT2's influence extends to growth and metabolism, as its capability to catalyze methyl transfers contributes depth and intricacy to these metabolic interactions. Notably, the genetic correlation between betaine, dimethylglycine, and methionine levels and growth rates suggests that BHMT2 plays a role in influencing growth potential [24].
The results of this study indicate that the candidate genes identified were primarily associated with pig production and meat quality, suggesting that selection pressure in commercial pig breeds has been largely focused on these traits. Specifically, these genes appear to have been under strong selective pressure in order to improve the production efficiency and meat quality of commercial pig breeds.
5. Conclusions
Our study has successfully identified numerous candidate genes that are under selective pressure in commercial pig breeds, particularly in muscle development. Two key genes, PBTP2 and DYPD, were found in all commercial breeds and are located in chromosome 4. This information is crucial in understanding the genetic processes that contribute to phenotypic differentiation in pig breeds. With a better understanding of natural and artificial selection processes, we can improve pig breeding efficiency and drive pig production forward.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: title; Table S1: title; Video S1: title.
Author Contributions
Conceptualization, F.A., G.M. and J.S.; methodology, F.A., G.M. and J.S.; validation, G.M. and J.S.; formal analysis, F.A., G.M. and J.S; data curation, J.S., B.B and C.D.; writing—original draft preparation, F.A. and G.M.; writing—review and editing, G.M., J.S. visualization, F.A., G.M., J.S., B.B. and C.D.; supervision, G.M. and J.S;. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The genomic data resulted from a collaboration involving Boku University, the Institut für Biologische Landwirtschaft und Biodiversität der Nutztiere at HBLFA Raumberg-Gumpenstein, and Österreichische Schweineprüfanstalt GmbH.
Acknowledgments
The author extends sincere thanks to Dr. Siavash Salek Ardestani for his technical assistance in the preparation of this paper. His expertise greatly improved the quality of the work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhao, F.; McParland, S.; Kearney, F. et al. Detection of selection signatures in dairy and beef cattle using high-density genomic information. Genet Sel Evol 47, 49 (2015).
- Gonzalez-Dieguez, D.; Tusell, L.; Bouquet, A.; Legarra, A.; Vitezica, Z.G. Purebred and Crossbred Genomic Evaluation and Mate Allocation Strategies To Exploit Dominance in Pig Crossbreeding Schemes. G3 Genes|Genomes|Genetics 10, 2829-41 (2020).
- Hermisson, J.; Pennings, P.S. Soft sweeps: molecular population genetics of adaptation from standing genetic variation. Genetics 169, 2335-52 (2005).
- . Zanella, R.; Peixoto, J.O.; Cardoso, F.F.; Cardoso, L.L.; Biegelmeyer, P.; Cantão, M.E.; Otaviano, A.; Freitas, M.S.; Caetano, A.R.; Ledur, M.C. Genetic diversity analysis of two commercial breeds of pigs using genomic and pedigree data. Genetics Selection Evolution 48, 1-10 (2016).
- Sabeti, P.C.; Varilly, P.; Fry, B.; Lohmueller, J.; Hostetter, E.; Cotsapas, L.; Brooks, L.D. Genome-wide detection and characterization of positive selection in human populations. Nature 449, 913-8 (2007).
- Grossman, S.R.; Shylakhter, I.; Karlsson, E.K.; Byrne, E.H.; Morales, S.; Frieden, G.; Hostetter, E.; Angelino, E.; Garber, M.; Zuk, O. A composite of multiple signals distinguishes causal variants in regions of positive selection. Science 327, 883-6 (2010).
- Eydivandi, S.; Roudbar, M.A.; Ardestani, S.S.; Momen, M.; Sahana, G. A selection signatures study among Middle Eastern and European sheep breeds. Journal of Animal Breeding and Genetics 138, 574-88 (2021).
- Yang, W.; Liu, Z.; Zhao, Q.; Du, H.; Yu, J.; Wang, H.; Liu, X.; Liu, H.; Jing, X.; Yang, H. Population Genetic Structure and Selection Signature Analysis of Beijing Black Pig. Frontiers in genetics 13 2022. [CrossRef] [PubMed]
- Szpiech, Z.A.; Novak, T.E.; Bailey, N.P.; Stevison, L.S. Application of a novel haplotype-based scan for local adaptation to study high-altitude adaptation in rhesus macaques. Evolution letters 5, 408-21 (2021).
- Szpiech, Z.A.; Hernandez, R.D. selscan: an efficient multithreaded program to perform EHH-based scans for positive selection. Molecular Biology and Evolution, 31, 2824–2827 (2014).
- Browning, B.L.; Tian, X.; Zhou, Y.; Browning, S.R. Fast two-stage phasing of large-scale sequence data. The American Journal of Human Genetics, 108(10), 1880-90 (2021).
- Kim, E.S.; Elbeltagy, A.R.; Aboul-Naga, A.M.; Rischkowsky, B.; Sayre, B.; Mwacharo, J.M.; Rothschild, M.F. Multiple genomic signatures of selection in goats and sheep indigenous to a hot arid environment. Heredity 116(3), 255-64 (2016).
- Santana, M.H.d.A.; Ventura, R.V.; Utsunomiya, Y.T.; Neves, H.H.d.R.; Alexandre, P.A.; Oliveira Junior, G.A.; Gomes, R.d.C.; Bonin, M.d.N.; Coutinho, L.L.; Garcia, J.F. A genomewide association mapping study using ultrasound-scanned information identifies potential genomic regions and candidate genes affecting carcass traits in Nellore cattle. Journal of Animal Breeding and Genetics 132, 420-7 (2015).
- Ding, Rongrong, et al. "Single-locus and multi-locus genome-wide association studies for intramuscular fat in Duroc pigs." Frontiers in genetics 10 (2019): 619.
- Hinkle, Emma R., et al. "RNA processing in skeletal muscle biology and disease." Transcription 10.1 (2019): 1-20.
- Wu, Shanshan, et al. "Disrupted potassium ion homeostasis in ciliary muscle in negative lens-induced myopia in Guinea pigs." Archives of Biochemistry and Biophysics, Volume 688, 2020,108403.
- Yang, Y.; Wang, X.; Wang, S.; Chen, Q.; Li, M.; Lu, S. Identification of Potential Sex-Specific Biomarkers in Pigs with Low and High Intramuscular Fat Content Using Integrated Bioinformatics and Machine Learning. Genes 14(9), 1695 (2023).
- Fontanesi, L. et al. A genomewide association study for average daily gain in Italian Large White pigs. Journal of Animal Science, Volume 92, Issue 4, April 2014, Pages 1385–1394.
- AL_Rayyan, N.; Wankhade, UD.; Bush, K.; Good, DJ. Two single nucleotide polymorphisms in the human nescient helix-loop-helix 2 (NHLH2) gene reduce mRNA stability and DNA binding. Gene 512(1), 134-42 (2013).
- Zhang, H.; Xia, P.; Feng, L.; Jia, M.; Su, Y. Feeding frequency modulates the intestinal Transcriptome without affecting the gut microbiota in pigs with the same daily feed intake. Frontiers in Nutrition 2021. [CrossRef] [PubMed]
- Bergamaschi, M.; Maltecca, C.; Schillebeeckx, C.; McNulty, NP.; Schwab, C.; Shull, C.; Fix, J.; Tiezzi, F. Heritability and genome-wide association of swine gut microbiome features with growth and fatness parameters. Scientific reports 10(1), 10134 (2020).
- Wang, X.; Li, G.; Ruan, D.; Zhuang, Z.; Ding, R.; Quan, J.; Wang, S.; Jiang, Y.; Huang, J.; Gu, T.; Hong, L. Runs of homozygosity uncover potential functional-altering mutation associated with body weight and length in two Duroc pig lines. Frontiers in Veterinary Science 2022. [CrossRef] [PubMed]
- Hao, X.; Plastow, G.; Zhang, C.; Xu, S.; Hu, Z.; Yang, T.; Wang, K.; Yang, H.; Yin, X.; Liu, S.; Wang, Z. Genome-wide association study identifies candidate genes for piglet splay leg syndrome in different populations. BMC genetics 2017. [CrossRef] [PubMed]
- Dervishi, E.; Bai, X.; Dyck, MK.; Harding, JC.; Fortin, F.; Dekkers, JC.; Plastow, G. GWAS and genetic and phenotypic correlations of plasma metabolites with complete blood count traits in healthy young pigs reveal implications for pig immune response. Frontiers in Molecular Biosciences 2023. [CrossRef] [PubMed]
Figure 1.
Manhattan plot in Large White population compare with local population in XP-ehh and XP-nSL methods.
Figure 1.
Manhattan plot in Large White population compare with local population in XP-ehh and XP-nSL methods.
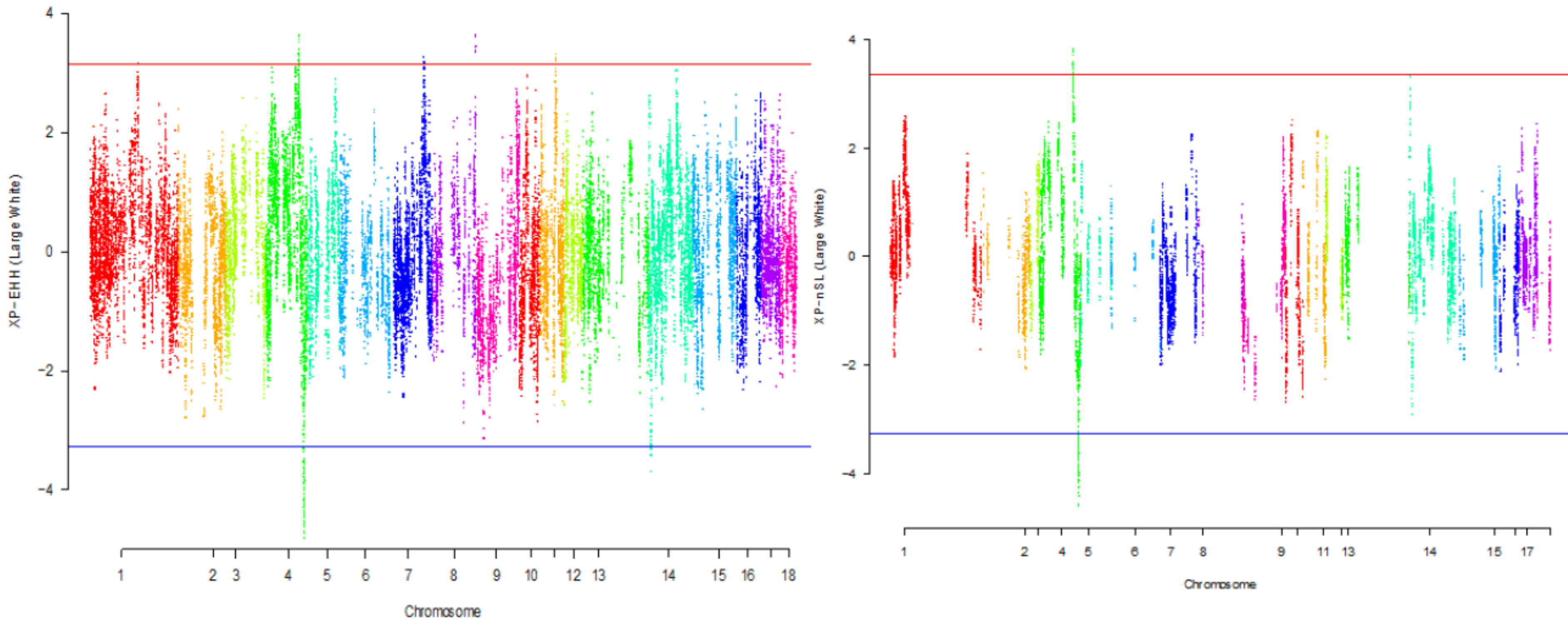
Figure 2.
Manhattan plot in Duroc population compare with local population in XP-ehh and XP-nSL methods.
Figure 2.
Manhattan plot in Duroc population compare with local population in XP-ehh and XP-nSL methods.
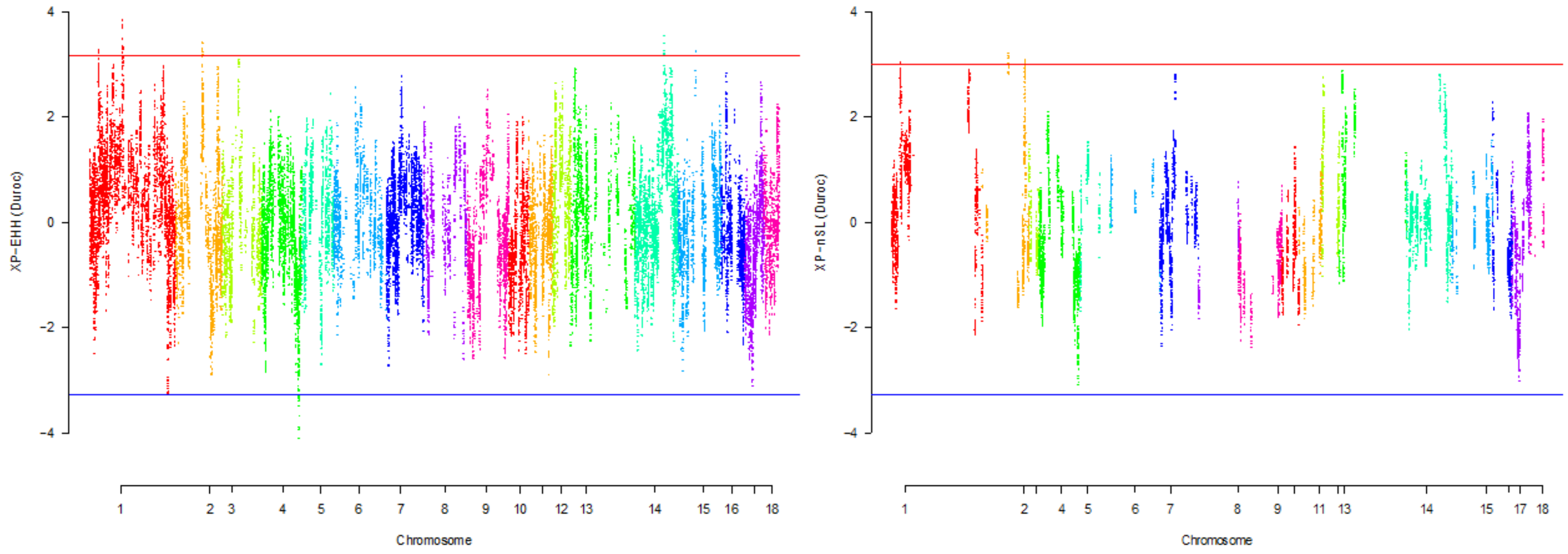
Figure 3.
Manhattan plot in All population compare with local population in XP-ehh and XP-nSL methods.
Figure 3.
Manhattan plot in All population compare with local population in XP-ehh and XP-nSL methods.
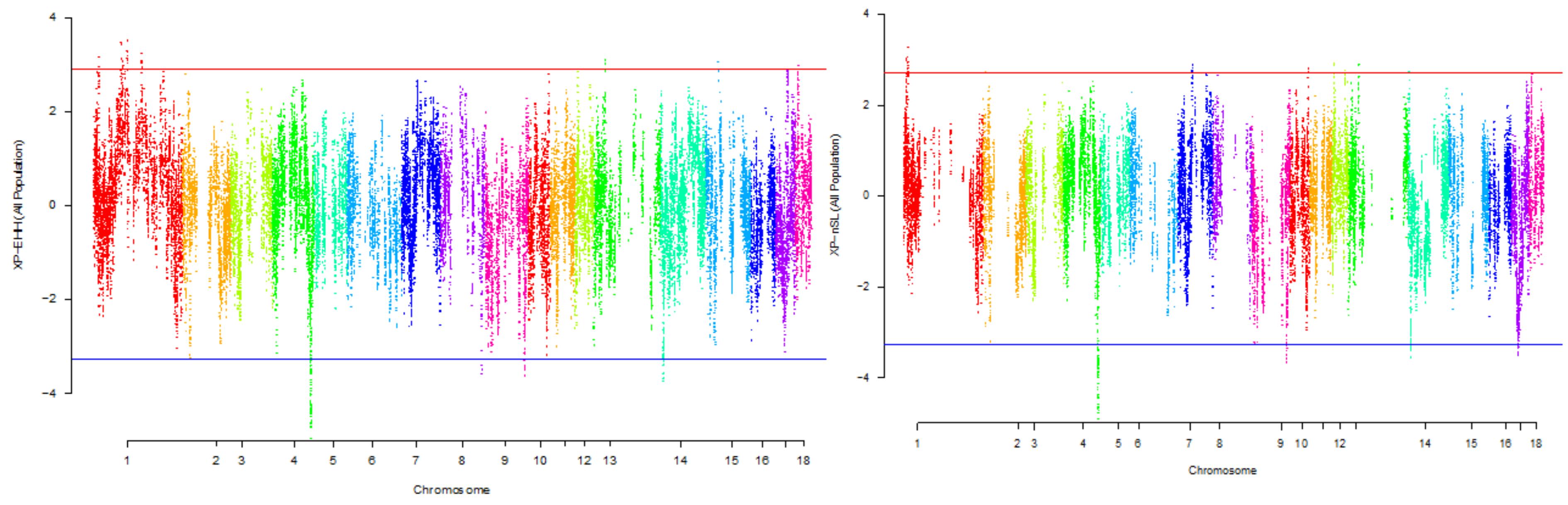

Table 1.
This is a figure. Schemes follow the same formatting.
| Gene name | Chromosome number | Position |
| DPYD PTBP2 ATP1A1 |
4 | 119931414:120712459 |
| 4 | 120961381:121041802 | |
| 4 | 104353508:104384547 | |
| CASQ2 CD2 |
4 | 104918501:104983607 |
| 4 | 103963345:103977292 | |
| IGSF3 MAB21L3 NHLH2 SLC22A15 |
4 | 103994390:104166988 |
| 4 | 104599818:104625234 | |
| 4 | 104857635:104858042 | |
| 4 | 104664015:104755672 | |
| VANGL1 ARSB |
4 | 104996073:105048775 |
| 2 | 87614963:87778447 |
Table 2.
common gene between different methods and pig species.
| Comparing Breed and methods | Common Genes | ||
| Xpehh | Xp-Nsl | Between Xpehh and XpNsl | |
| Large White(Between two methods) | - | - | DPYD, PTBP2, ATP1A1, CASQ2, CD2, IGSF3, MAB21L3, NHLH2 SLC22A15,VANGL1 |
| Large White and Duroc | DPYD, PTBP2 | PTBP2 | PTBP2 |
| Large White and Pietrain | DPYD, PTBP2 | DPYD, PTBP2 | DPYD, PTBP2 |
| Large White and Landrace | DPYD, PTBP2 | PTBP2 | DPYD, PTBP2 |
| Duroc(Between two methods) | - | - | ARSB, BHMT, BHMT2, DMGDH, JMY |
| Duroc and Pietrain | DPYD, PTBP2 | PTBP2 | PTBP2 |
| Duroc and Landrace | DPYD, PTBP2 | PTBP2 | PTBP2 |
| Pietrain(Between two methods) | - | - | DPYD, PTBP2 |
| Pietrain and Landrace | DPYD, PTBP2 | PTBP2 | DPYD, PTBP2 |
| Landrace(Between two methods) | - | - | PTBP2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



