Submitted:
26 September 2023
Posted:
29 September 2023
You are already at the latest version
Abstract
The present study was to investigate effects of total flavonoids from Taraxacum mongolicum Hand.-Mazz. (FT) on fermentation quality, antioxidant status and microbial community of Caragana korshinskii Kom. (CK) silage. CK was ensiled with no additive (CON), 1% FT and 2% FT on a fresh weight (FW) basis for 60 days. The results showed that 1% FT and 2% FT groups displayed higher DM content than CON group, and 2% FT group had the best effect on nutrient preservation. Compared with CON and 1% FT groups, 2% FT group exhibited the best silage fermentation quality and the highest antioxidant activity, including increased lactic acid, acetic acid concentrations and the activities of antioxidant enzymes, as well as decreased pH and the ammonia nitrogen (NH3-N) concentration. Moreover, 2% FT addition significantly affected the microbial community, such as increased abundance of Lactobacillus and decreased abundances of Pseudomonas and unidentified Cyanobacteria. The abundances of Lactobacillus parafarraginis and Lactobacillus brevis were negatively correlated to pH, while they were positively correlated with T-AOC, GSH-Px and CAT activities. In conclusion, 2% FT may be used as additives to inhibit the growth of undesirable microorganisms and promote fermentation quality and antioxidant activity of CK silage.
Keywords:
Caragana korshinskii Kom.
; flavonoids
; Taraxacum mongolicum Hand.-Mazz.
; fermentation quality
; antioxidant status
; microbial community
1. Introduction
Caragana korshinskii Kom. (CK), a woody legume shrub, is grown in arid and semi-arid regions to help stop desertification and provide environmental protection [1]. However, overgrowth of CK can result in reduced plant species diversity and regeneration of fresh CK branches [2]. Consequently, routine pruning of older branches is essential, a practice that generates approximately 4 million tons of stubble annually in China [3]. In addition, the expansion of livestock production and the shortage of traditional forage resources necessitate the development of new feed sources. CK, characterized by its high nutritional value, presents a promising option to support livestock production [4]. Cai [5] reported that the preservation of nutrients in woody forage through silage could help address the scarcity of forage resources in arid regions. In general, converting woody plants into naturally high-quality silage proves challenging due to the limited presence of fermentation substrates in legumes [5]. Several studies have indicated that the incorporation of rice bran, Lactobacillus plantarum, and molasses to facilitate lactic acid fermentation in Caragana silage, thereby creating a valuable high-quality forage resource for livestock production [6,7].
In the context of food safety, one promising dietary approach involves incorporating natural compounds extracted from plants into the diets of ruminant [8]. The usage of metabolites derived from medicinal plants in domestic animals has gained traction in animal research and applications throughout the food production chain [9,10]. As plant secondary metabolites, flavonoids are known for their antioxidant properties and other bioactivities, such as antimicrobial and anti-inflammatory effects, which contribute to reducing the risk of diseases [11]. Despite their potential benefits, there has been limited research on the utilization of flavonoids from plants in forage silage. Flavonoids and polysaccharides exist in astragalus and hawthorn residues have been shown to inhibit the growth of various spoilage microorganisms, including Enterobacter, and enhance the quality of alfalfa silage [12,13,14]. Apple pomace and grape pomace are rich in flavonoids and other polyphenols, which inhibited proteolysis and altered the fatty acid composition of ensiled alfalfa [15]. Sea buckthorn pomace has relatively high flavonoids and other bioactive substances [16], and Lactobacillus plantarum and Lactobacillus brevis could be enriched during alfalfa silage [17]. In conclusion, flavonoids derived from medicinal plants have the potential to serve as additives for enhancing the quality of forage silage.
Taraxacum mongolicum Hand.-Mazz., widely distributed in the warmer temperate zones of the northern hemisphere [18], has a long history of traditional medicinal and dietary use in China [19]. This plant is recognized for its anti-inflammatory, choleretic, antibacterial, anti-allergic, and antioxidant properties, attributed to its bioactive metabolites such as phenolic compounds, sesquiterpene lactones, polysaccharides, and flavonoids [20]. Flavonoids from Taraxacum mongolicum Hand.-Mazz. (FT) have antibacterial and antioxidant capabilities [21], and results have showed that FT after fermentation exhibited stronger DPPH free radical scavenging ability and reducing power than that before fermentation [22]. Antioxidants can effectively neutralize excessive free radical production, mitigating oxidative stress when the animal’s antioxidant system balance is disrupted due to metabolic disturbances [23,24]. Given this background, we hypothesized that FT could serve as a valuable silage additive to enhance the fermentation quality and antioxidant capacity of CK silage.
To the best of our knowledge, there is limited information available on the application of FT in the ensiling process of CK. Therefore, the objective of this study was to investigate the effects of FT on the fermentation quality, antioxidant status, and microbial community of CK silage.
2. Materials and Methods
2.1. Materials and Silage Preparation
The branch of CK (no inflorescence and leaf) was harvested from Yanchi county, Ningxia, China in May 2022. After the cutting, the CK branches were broken down and processed into slices less than 1 cm in size using a kneading machine (RC-400, QuFuZhiZao Conveyor Co., Ltd., Shandong, China). The freshly prepared CK material was adjusted to attain a dry matter (DM) content of approximately 40%. Three treatments were applied based on fresh weight (FW): no additive (CON), 1% FT and 2% FT. The FT (98% total flavonoids; Shaanxi Guanchen Biotech Co., Ltd.) used in this experiment was extracted from Taraxacum mongolicum Hand.-Mazz. and kept as a fine dry power.
Subsequently, 500 g of the CK material was uniformly mixed with the respective additives and manually packed into polyethylene bags measuring 27 cm × 30 cm (Embossed Food Saver Bag Co., Ltd., Chengdu, China). These bags were vacuum-sealed tightly. A total of 9 bags (comprising 3 treatments with 3 replicates each) were prepared and stored at ambient temperature (24–26°C) for 60 days.
2.2. Analysis of chemical composition, fermentation characteristics and microbial composition
To determine fermentation parameters, 10 g samples of silage were diluted with 90 ml of distilled water, filtered through four layers of cheesecloth, and refrigerated at 4°C for 24 h. The resulting supernatant was used for pH measurement, conducted using a calibrated pH meter (PHS-3G, Mettler Toledo, Zurich, Switzerland). Before analyzing ammonia nitrogen (NH3-N) and organic acids, a subsample of the supernatant was subjected to centrifugation at 2,500 rpm for 10 minutes and then filtered through a 0.22-µm microporous filter. NH3-N concentration was determined using the phenol-sodium hypochlorite colorimetric method. Organic acids were quantified using high-performance liquid chromatography (HPLC) with a KC-811 column (Shodex; Shimadzu, Japan) at an oven temperature of 50°C, a flow rate of 1 ml/min, and detection at 210 nm, following the method outlined by [25].
For dry matter (DM) determination, samples were dried in a forced-air oven at 65°C for 72 hours and ground to pass through a 1.0 mm screen for subsequent chemical analysis. Total nitrogen (TN) content was assessed using a Kjeldahl apparatus (K-360, BUCHI Laboratory Equipment Trade Co., Ltd., Shanghai, China), and crude protein (CP) was calculated by multiplying the TN content by 6.25. Neutral detergent fiber (NDF) was determined using heat-stable α-amylase as the method of Van Soest [26], while acid detergent fiber (ADF) was measured in accordance with the method described by Robertson and Van Soest (1981) using an ANKOM A2000i fiber analyzer (A2000i, ANKOM Technology, New York, USA). Water-soluble carbohydrates (WSC) were quantified via anthrone-sulfuric acid colorimetry [27], and ether extract (EE) was analyzed using the procedures outlined by the Association of Official Analytical Chemists (AOAC) [28].
The plate count method was employed to enumerate the number of lactic acid bacteria (LAB), yeasts, and molds in both the fresh CK materials and silage samples. To achieve this, 5 g samples were homogenized in 50 mL of sterilized physiological saline (8.5 g/L) for 45 minutes in an incubator shaker (25 ± 2°C; shaker rate: 120 rpm). The resulting solution was then serially diluted (10-fold). LAB were enumerated by inoculating MRS broth and incubating at 37°C in an anaerobic incubator for 48–72 h. Yeasts and molds were enumerated by inoculating potato medium (PDA, containing 0.1% chloramphenicol) and incubating at 30°C for 7 d. Microbial numbers were expressed in colony-forming units (cfu), converted into logarithmic form, and reported on a fresh matter (FM) basis.
2.3. Antioxidant capacity analysis
Following the thawing of the sample filtrates, which had been stored at -20°C for antioxidant analysis, the filtrates were subsequently subjected to centrifugation at 12,000 ×g for 10 min at 4°C. The resulting liquid supernatant was employed for the assessment of various antioxidant parameters, including total antioxidant capacity (T-AOC), superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), and catalase (CAT). These analyses were conducted spectrophotometrically using commercially available assay kits sourced from Nanjing Jiancheng Bioengineering Institute, China, in accordance with the manufacturer’s instructions.
2.4. Microbial community analysis
For the DNA extraction of silage samples, a DNA isolation kit (Tiangen, DP302-02, Tiangen, China) was employed in accordance with the manufacturer’s instructions. Subsequently, full-length 16S rRNA gene amplification for SMRT sequencing was carried out through PCR.
The analysis of full-length 16S rRNA amplicon sequencing data, including the construction of the 16S rRNA library, quality control for PCR amplifications, sequence pre-processing, species annotation, and assessment of alpha diversity, followed the procedures outlined in Xu [29]. Furthermore, to assign taxonomy to the sequences, a comparison was made with the NCBI 16S ribosomal RNA database, with classification achieved at a bootstrap threshold of 0.8, using RDP classifier software. The sample ordination, based on beta diversity, was explored using principal coordinate analysis (PCoA).
2.5. Statistical Analysis
To assess the effects of FT, a one-way analysis of variance (ANOVA) was conducted utilizing the general linear model procedure within the Statistical Package for Social Science (SPSS 21.0, SPSS, Inc., Chicago, IL, United States). To discern differences among treatment means, Duncan’s tests were employed. The significance was declared at p < 0.05. An online platform (http://www.omicshare.com/tools) was used to analyze the sequencing data of Microbial community.
3. Results
3.1. Chemical and microbial compositions of CK before ensiled
Table 1 shows the chemical and microbial compositions of CK before ensiled. The DM content was 426.21 g/kg FM, and the contents of CP, EE, WSC, NDF and ADF were 64.79, 25.20, 10.12, 801.81 and 664.81 g/kg DM, respectively. The number of LAB, yeasts, and molds in fresh CK was 3.19, 0.00 and 3.86 log10 cfu/g FM, respectively.
3.2. Effect of FT on the fermentation quality of CK silage
Table 2 illustrates the fermentation quality of CK silage after 60 days of ensiling. The pH and the NH3-N concentration of 2% FT group were lower compared with CON and 1% FT groups (p < 0.01). The lactic acid and acetic acid concentrations of 2% FT group were higher compared with CON and 1% FT groups (p < 0.01). Meanwhile, the highest and lowest LAB number were presented in 2% FT and CON groups (p < 0.01), respectively. Propanoic acid, butyric acid, yeasts and molds were not detected in all groups.
3.3. Effect of FT on the chemical composition of CK silage
Table 3 shows the chemical composition of CK silage after 60 days of ensiling. The DM content of CON group was the lowest compared with 1% FT and 2% FT groups (p < 0.01). The CP contents in CON and 1% FT groups were lower compared with 2% FT group (p < 0.01). The contents of NDF and ADF in 2% FT group were lower compared with CON and 1% FT groups (p < 0.01). Furthermore, there were no significant differences in WSC and EE contents among three groups (p > 0.05).
3.4. Effect of FT on the antioxidant capacity of CK silage
Table 4 illustrates the antioxidant capacity of CK silage after 60 days of ensiling. The T-AOC and CAT activity of 2% FT group were higher than that of CON and 1% FT groups (p < 0.01), and the T-AOC and CAT activity of 1% FT groups were higher than that of CON group (p < 0.01). The SOD activity of 2% FT group was higher than that of CON and 1% FT groups (p < 0.01). The GSH-Px activities of 1% FT and 2% FT groups were higher than that of CON group (p < 0.01).
3.5. Effect of FT on the microbial communities of CK silage
Table 5 shows the alpha-diversity of the microbial communities of silages. The coverage depth for all treatments was above 99%. Compared with CON group, 1% FT and 2% FT groups decreased the richness indices (Ace and Chao 1), however, there were no significant differences among three groups (p > 0.05). And, there were no significant differences in Simpson and Shannon indexs between three groups (p > 0.05).
The principal coordinate analysis was used to study the correlations of microbial community structure in CK silage. There were clear separation and differences in bacterial communities among the three groups (Figure 1).
Figure 2A describes the microbial communities at the phylum level. Firmicutes and Proteobacteria were the dominant phyla in three groups. The abundance of Firmicutes was highest in 2% FT group (84.7%), followed by 1% FT group (62.3%), and CON group (17.8%). The FT-treatments obviously decreased the abundance of Proteobacteria, which was the highest in CON group (56.1%), followed by that in 1% FT group, and 2% FT group (30.7%, and 8.1%, respectively). In addition, 2% FT and 1% FT groups also decreased the abundance of Cyanobacteria and Bacteroidetes compared with CON group (0.1%, 5.6% and 14.7%; 0.3%, 0.8% and 4.3%).
Figure 2B describes the microbial communities at the genus level. Lactobacillus, the main microbes in 2% FT group (83.2%) and 1% FT group (50.1%), and that in CON group was 4.5%. The relative abundance of Pseudomonas and unidentified Cyanobacteria in 2% FT and 1% FT groups were higher than that in CON group (3.9%, 13.5% and 35.2%; 0.0%, 3.5% and 14.7%).
Figure 2C describes the microbial communities at the species level. Lactobacillus parafarraginis was the most dominant in 2% FT group (77.1%) and Lactobacillus plantarum was the most dominant in 1% FT group (45.0%). The relative abundance of Pardosa pseudoannulata in 2% FT and 1% FT groups were lower than that in CON group (0.0%, 0.0% and 14.2%). Moreover, the relative abundance of Lactobacillus brevis in 2% FT was higher than that in 1% FT and CON groups (4.5%, 2.7% and 0.5%).
The taxonomic differences between the different treatments are shown in the clade representation generated by LEfSe (Figure 3). 1% FT and 2% FT additions significantly affected the microbial composition of CK silage (LAD > 4). 1% FT group increased the abundance of Pigmentiphaga. 2% FT group increased the abundance of Lactobacillaceae, Lactobacillales, Lactobacillus, Oceanobacillus profundus, Oceanobacillus, Lactobacillus paralimentarius.
3.6. The correlation between microorganisms and fermentation parameters, antioxidant capacity of CK silage
Figure 4 illustrates the correlation between microorganisms (species level) and fermentation parameters, antioxidant capacity of CK silage. The abundance of Lactobacillus parafarraginis exerted positive correlations with lactic acid and acetic acid concentrations, LAB number (p < 0.01), T-AOC, GSH-Px and CAT activities (p < 0.05), while it showed negative correlations with pH and NH3-N concentration (p < 0.05). The abundance of Lactobacillus brevis showed positive correlations with acetic acid concentration, LAB number (p < 0.01), T-AOC, GSH-Px and CAT activities (p < 0.05), while it showed negative correlations with pH (p < 0.05). The abundances of Weissella cibaria and Pararhizobium giardinii showed negative correlations with SOD activity (p < 0.05). The abundance of Pardosa pseudoannulata showed negative correlations with acetic acid concentration and GSH-Px activity (p < 0.05). The abundance of Caragana microphylla showed positive correlations with NH3-N concentration (p < 0.05).
4. Discussion
4.1. Effect of FT on the chemical composition of fresh CK and Silage
The WSC content plays a pivotal role in determining the quality of silage fermentation. Ensuring satisfactory fermentation quality requires WSC values exceeding 5% DM [30]. Additionally, the presence of LAB at levels surpassing 5.0 log 10 cfu/g during ensiling is crucial for achieving well-preserved silage [31]. Instances of lower WSC content and LAB number, coupled with elevated mold counts suggested a requirement for additives in the preparation of CK silage. In this study, it was observed that FT-treated silage exhibited a higher DM content compared to CON group. This outcome suggested that FT effectively mitigated DM loss during the ensiling process. Furthermore, in comparison to freshly harvested CK, silage treated with FT preserved protein content remarkably well. Notably, 2% FT group displayed significantly higher CP levels than both 1% FT and CON groups. This preservation of protein content could be attributed to two potential mechanisms: Firstly, the active inhibition of proteases in the low-pH environment directly reduced proteolysis. Secondly, the formation of protein-polyphenol complexes in the silage treated with 2% FT, facilitated by the presence of flavonoids, which are a subgroup of polyphenols [32]. Fresh CK exhibited substantial levels of NDF and ADF, while 2% FT group exhibited lower fiber contents compared to the CON group in silage. This reduction in NDF and ADF contents within 2% FT group could be attributed to processes such as acidic hydrolysis and potential fibrinolytic enzyme production by microorganisms during the fermentation of silage [17]. The findings of this study collectively suggest that the application of 2% FT yields the most favorable results in terms of nutrient preservation in CK silage, highlighting FT’s potential as a valuable silage additive.
4.2. Effect of FT on the fermentation quality of CK silage
The organic acids (mainly lactic acid and acetic acid) and NH3-N concentrations, and pH are vital indicators of well-preserved forage silage quality [33]. The accumulation of NH3-N during ensiling is a recognized marker of protein degradation [34]. 2% FT group exhibited lower pH and NH3-N concentration than CON, meanwhile, the highest lactic acid concentration and LAB number was found in 2% FT group. Flavonoids are part of polyphenols, and the decreased NH3-N content in 2% FT-treated silage might be due to the formation of complexes of polyphenols and proteins, or inhibition of plant and/or bacterial proteolytic enzymes [35]. These results indicated that FT could promote the proliferation of LAB, which increased the lactic acid concentration. Similarly, apple pomace and grape pomace contains polyphenol, such as phenolic acids, flavonoids, and so on [36,37], and the additions resulted in an elevated lactic acid concentration coupled with a decrease in pH and nonprotein nitrogen concentrations in the ensiled alfalfa [15]. In the present study, the acetic acid concentration of 2% FT group was the highest among three groups. The reason might be due to Lactobacillus parafarraginis was the most dominant in 2% FT group (77.1%). Lactobacillus parafarraginis belonging to Lactobacillus buchneri group [38], is known for its capability to enhance acetic acid concentration and bolster the aerobic stability of silage across a wide spectrum of temperatures [39].
It is worth noting that propanoic acid, butyric acid, yeasts, and molds were not detected in any of the groups, possibly due to the inhibitory effects of the low-pH environment on undesirable bacteria and fungi, including Clostridia. Prior research has illuminated that Lactobacillus spp. showcase a higher degree of resistance to phenolic compounds when compared to other bacterial groups such as Clostridium spp., Bacteroides spp., Escherichia coli, and Bacillus subtilis [32].
Hence, the incorporation of 2% FT may foster a microenvironment characterized by low pH value and elevated levels of bioactive components, thereby facilitating the rapid proliferation of desirable microorganisms such as LAB under anaerobic conditions. To delve deeper into these findings, microbial community analysis was conducted.
4.3. Effect of FT on the antioxidant capacity of CK silage
Oxidative stress has been associated with various health disorders in cattle [40], and it can significantly impair animal production outcomes [41]. Numerous studies have highlighted the role of dietary antioxidants in enhancing both the antioxidant capacity of animals and the antioxidant properties of animal-derived products [42,43,44]. Therefore, augmenting the antioxidant capacity of silage is of paramount importance and highly desirable in the context of animal nutrition. Prior investigations by Zhang [45,46] have underscored that alfalfa silage inoculated with LAB exhibiting robust antioxidant activity displayed the highest T-AOC and CAT activity. Generally, endogenous antioxidants like SOD, CAT and GSH-Px serve as the initial line of defense against oxidative stress within cells. These enzymes perform functions such as dismutating superoxide radicals (O2−), breaking down hydrogen peroxide (H2O2), and converting hydroperoxides into harmless molecules (H2O / alcohol and O2), respectively [47]. Compared with CON group, 1% FT and 2% FT groups showed better antioxidant capacity. And 2% FT-treated CK silage exhibited the highest T-AOC, SOD, GSH-Px and CAT activities, which was probably due to the substantial antioxidant capacity inherent in FT. As an important subgroup of plant phenols, flavonoids endow high antioxidant activity, attributable to their polyhydroxy structure [48,49]. In vitro assays have revealed that flavonoids extracted from fermented dandelion exhibit superior antioxidant activity compared to those from unfermented dandelion, as evidenced by their effectiveness in scavenging DPPH radicals and their reducing power [22]. Consequently, these findings lend support to the hypothesis that FT functions analogously to chemical antioxidants, bolstering the antioxidant capacity of silage.
4.4. Effect of FT on the microbial communities of CK silage
In this study, the high coverage value of approximately 0.99 for each sample indicated the comprehensive sequencing coverage achieved, signifying that the majority of microbial species were successfully detected. The beta-diversity analysis unveiled distinct separation and variations in bacterial communities within the CK silage, strongly suggesting that the microbial composition underwent significant changes due to FT-treatment during the ensiling process.
Specific alterations in bacterial composition were observed at various taxonomic levels among the three experimental groups. Firmicutes and Proteobacteria emerged as the top two predominant phyla in CK silage, consistent with prior research findings [50,51,52]. Lactobacillus, recognized for its pivotal role and beneficial attributes in ensiled forage, is typically the dominant bacterial strain responsible for driving lactic fermentation during ensiling [30]. Intriguingly, FT-treatment led to a notable increase in the abundance of Lactobacillus, particularly in 2% FT group, where it constituted as much as 83% of the total microbial population. Within them, Lactobacillus parafarraginis emerged as the most dominant species in 2% FT group, comprising a substantial 77.1% of the population. This might be due to the addition of 2% FT could provide substrates and energy supplementation for the propagation of Lactobacillus parafarraginis, and/or to the fact that Lactobacillus parafarraginis is highly resistant to phenolic compounds. Previous studies have demonstrated that Lactobacillus parafarraginis ZH1 contributes to the aerobic stability and safety of silage by producing anti-yeast compounds such as acetic acid, hexadecanoic acid, benzoic acid, tetradecanoic acid, and octadecanoic acid [53]. Lactobacillus parafarraginis is classified as a heterofermentative LAB, endowed with the ability to convert lactic acid into acetic acid and 1, 2-propylene glycol [50]. This capability might elucidate the observed high acetic acid content in 2% FT group. These findings collectively suggest that the addition of 2% FT might enhance the aerobic stability and safety of CK silage by fostering the proliferation of Lactobacillus parafarraginis. However, it is essential to note that this hypothesis warrants further experimental validation. Lactobacillus plantarum is a common dominant bacterium in silage, and that was the most dominant in 1% FT group (45.0%). However, the abundance of Lactobacillus plantarum in 2% FT group was considerably lower, and this might be due to the replacement of Lactobacillus plantarum by Lactobacillus parafarraginis in 2% FT group during later silage. As photosynthetic bacterial phylum, Cyanobacteria exist in a variety of growing environments and a wide variety of generated products [54]. Li [55] reported that Cyanobacteria was the dominant phylum in four fresh tropical forages, and Cyanobacteria decreased to marginal levels in silage of paspalum and stylo. Another research also reported that Cyanobacteria was the dominant phylum in the control Neolamarckia cadamba leaf silage, and intrinsic tannins significantly decreased the abundance of Cyanobacteria. [56]. Similar to above studies, FT-treatment markedly decreased abundance of Pseudomonas and unidentified Cyanobacteria (Pardosa pseudoannulata), especially for 2% FT group. Pseudomonas, considered an undesirable bacterial strain in silage due to its potential for biogenic amine production and the reduction of protein content and nutritional value [57], experienced a marked reduction in abundance following FT treatment, especially in 2% FT group. These observations align with findings from previous studies involving various forage types, including corn stover, red clover, alfalfa, and Moringa oleifera leaves [50,51,58,59]. Therefore, the incorporation of 2% FT appears to be an effective strategy for improving silage conditions, leading to alterations in both the silage environment and microbial dynamics.
4.5. The correlation between microorganisms and fermentation parameters, antioxidant capacity of CK silage
Silage fermentation is a highly intricate biological process in which Lactobacillus assumes a central role. Lactobacillus has been observed to exert a positive influence on lactic acid production, while concurrently affecting pH and NH3-N levels [17,60]. Similar to the above research, there existed a negative correlation between pH and the abundance of Lactobacillus, particularly Lactobacillus parafarraginis and Lactobacillus brevis. Furthermore, it was evident that the abundance of Lactobacillus parafarraginis exhibited a negative correlation with NH3-N concentration in the present study. Research by Cai [31] has elucidated that lactic acid-producing cocci initiate lactic fermentation during the early stages of ensiling; however, their viability diminishes under acidic conditions. Consequently, lactic acid-producing rods, exemplified by Lactobacillus, assume a pivotal role in pH reduction during the latter stages of ensiling. Lactobacillus’s sensitivity to reduced pH levels underscores the notable correlation coefficients between NH3-N and Lactobacillus [52]. It was worth noting that the abundances of Lactobacillus parafarraginis and Lactobacillus brevis were positively correlated with acetic acid concentration. It was probably because of both species were heterofermentative LAB that could convert lactic acid to acetic acid [61]. The abundance of Caragana microphylla (belonged to Cyanobacteria) was positively correlated with NH3-N concentration. The explanation was that Caragana microphylla could be led to proteolysis.
The abundances of Lactobacillus parafarraginis and Lactobacillus brevis exhibited positive correlations with T-AOC, GSH-Px and CAT activities, and results showed Lactobacillus parafarraginis and Lactobacillus brevis might promote the improvement of antioxidant capacity. The precise mechanisms underlying this phenomenon warrant further investigation to elucidate the specific pathways or compounds through which Lactobacillus parafarraginis and Lactobacillus brevis exert their influence on antioxidant activity. The abundances of Weissella cibaria and Pararhizobium giardinii were negatively correlated with SOD activity, Weissella cibaria and Pararhizobium giardinii potentially reduced antioxidant capacity in CK silage. To better understand this phenomenon, it is essential to delve into the mechanisms by which these microorganisms impact SOD activity and explore whether they produce metabolites or compounds that interfere with the antioxidant defense systems in silage.
5. Conclusions
This study revealed that 2% FT addition had the best effect on nutrient preservation in CK silage, and 2% FT addition could improve the silage quality and antioxidant activity. In particular, 2% FT addition increased the proportion of LAB and inhibited the growth of undesirable microorganisms in CK silage. The results of this research contributes to the application of FT as a potential functional feed additive in CK silage. In the future, it could be possible to reveal the functional microbial with FT through metagenomics and develop other similar medicinal herbs in forage silage.
Author Contributions
Conceptualization, S.H.; methodology, S.H. and L.G.; software, L.G.; validation, L.G.; formal analysis, W.K.; investigation, L.G., X.Z. and C.M.; resources, S.H.; data curation, L.G.; writing—original draft preparation, S.H.; writing—review and editing, S.H. and W.K.; visualization, Q.L.; supervision, S.H.; project administration, S.H.; funding acquisition, S.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Science Foundation of Ningxia, grant number 2023AAC03094.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We sincerely thank the Key Laboratory of High-efficiency Production Model Innovation of Forage of the Ministry of Agriculture for their technical support for this research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, S.J.; Chaudhry, A.S.; Osman, A.; Shi, C.Q.; Edwards, G.R.; Dewhurst, R.J.; Cheng, L. Associative effects of ensiling mixtures of sweet sorghum and alfalfa on nutritive value, fermentation and methane characteristics. Animal. Feed. Sci. Technol. 2015, 206, 29–38. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.R. Effects of planting Caragana shrubs on soil nutrients and stoichiometries in desert steppe of Northwest China. Catena. 2019, 183, 104213. [Google Scholar] [CrossRef]
- Li, W.C.; Zhang, S.J.; Zhang, T.Z.; Shen, Y.Q.; Han, L.J.; Peng, Z.J.; Xie, Z.X.; Zhong, C.; Jia, S.R. Bacterial cellulose production from ethylenedi-amine pretreated Caragana korshinskii Kom. Ind. Crop. Prod. 2021, 164, 113340. [Google Scholar] [CrossRef]
- Wang, G.H.; Chen, Z.X.; Shen, Y.Y.; Yang, X.L. Efficient prediction of profile mean soil water content for hillslope-scale Caragana korshinskii plantation using temporal stability analysis. Catena, 2021, 206, 105491. [Google Scholar] [CrossRef]
- Cai, Y.M.; Du, Z.M.; Yamasaki, S.; Nguluve, D.; Tinga, B.; Macome, F.; Oya, T. Community of natural lactic acid bacteria and silage fermentation of cornstover and sugarcane tops in Africa. Asian-Australasian. J. Animal. Sci. 2020, 33, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- You, J.T.; Zhang, H.; Zhu, H.F.; Xue, Y.L.; Cai, Y.M.; Zhang, G.J. Microbial community, fermentation quality, and in vitro degradability of ensiling Caragana with lactic acid bacteria and rice bran. Front. Microbiol. 2022, 13, 804429. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.C.; Wang, Y.; Rinne, M.; Franco, O.M.; Li, F.H.; Lin, Y.F.; Zhang, Q.; Cai, Y.M.; Zhang, G.J. Effects of lactic acid bacteria and molasses on the fermentation quality, in vitro dry matter digestibility, and microbial community of Korshinsk peashrub (Caragana korshinskii Kom.) silages harvested at two growth stages. Grass. Forage. Sci 2023, 1–13. [Google Scholar] [CrossRef]
- Chen, L.; Bao, X.Y.; Guo, G.; Huo, W.J.; Xu, Q.F.; Wang, C.; Liu, Q. Treatment of alfalfa silage with tannin acid at different levels modulates ensiling characteristics, methane mitigation, ruminal fermentation patterns and microbiota. Animal. Feed. Sci. Technol. 2021, 278, 114997. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Firkins, J.L.; Dijkstra, J.; Kebreab, E.; Waghorn, G.; Makkar, H.P.S.; Adesogan, A.T.; Yang, W.; Lee, C.; Gerber, P.J.; Henderson, B.; Tricarico, J.M. Special topics-Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. J. Anim. Sci. 2013, 91, 5045–5069. [Google Scholar] [CrossRef] [PubMed]
- Mertenat, D.; Cero, M.D.; Vogl, C.R.; Ivemeyer, S.; Meier, B.; Maeschli, A.; Hamburger, M.; Walkenhorst, M. Ethnoveterinary knowledge of farmers in bilingual regions of Switzerland—Is there potential to extend veterinary options to reduce antimicrobial use? J. Ethnopharmacol. 2019, 246, 112184. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.F.; Gan, Q.; Liu, S.A.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food. Chem. 2022, 383, 132531–132531. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, S.; Davoodi, H. Herbal plants and their derivatives as growth and health promoters in animal nutrition. Vet. Res. Commun. 2011, 35, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Bouaziz, F.; Helbert, C.B.; Romdhane, M.B.; Koubaa, M.; Bhiri, F.; Kallel, F.; Chaari, F.; Driss, D.; Buon, L.; Chaabouni, S.E. Structural data and biological properties of almond gum oligosaccharide: Application to beef meat preservation. Int. J. Biol. Macromol. 2015, 72, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.K.; Wang, X.K.; Lu, Y.; Guo, L.N.; Li, X.M.; Yang, F.Y. Exploring the silage quality of alfalfa ensiled with the residues of astragalus and hawthorn. Bioresource. Technol. 2020, 297, 122249. [Google Scholar] [CrossRef]
- Ke, W.C.; Yang, F.Y.; Undersander, D.J.; Guo, X.S. Fermentation characteristics, aerobic stability, proteolysis and lipid composition of alfalfa silage ensiled with apple or grape pomace. Animal. Feed. Sci. Technol. 2015, 202, 12–19. [Google Scholar] [CrossRef]
- Kitrytė, V.; Povilaitis, D.; Kraujalienė, V.; Šulniūtė, V.; Pukalskas, A.; Venskutonis, P.R. Fractionation of sea buckthorn pomace and seeds into valuable components by using high pressure and enzyme-assisted extraction methods. LWT-Food. Sci. Technol. 2017, 85, 534–538. [Google Scholar] [CrossRef]
- Chen, L.Y.; Qu, H.; Bai, S.Q.; Yan, L.J.; You, M.H.; Gou, W.L.; Li, P.; Gao, F.Q. Effect of wet sea buckthorn pomace utilized as an additive on silage fermentation profile and bacterial community composition of alfalfa. Bioresource. Technol. 2020, 314, 123773. [Google Scholar] [CrossRef]
- González-Castejón, M.; Visioli, F.; Rodriguez-Casado, A. Diverse biological activities of dandelion. Nutr. Rev. 2012, 70, 534–547. [Google Scholar] [CrossRef]
- Duan, L.; Zhang, C.M.; Zhao, Y.; Chang, Y.Z.; Guo, L.; Donato, P.D.; Silvestri, B. Comparison of bioactive phenolic compounds and antioxidant activities of different parts of Taraxacum mongolicum. Molecules. 2020, 25, 3260–3272. [Google Scholar] [CrossRef]
- Schütz, K.; Carle, R.; Schieber, A. Taraxacum-A review on its phytochemical and pharmacological profile. J. Ethnopharmacol. 2006, 107, 313–323. [Google Scholar] [CrossRef]
- Park, S.; Kim, D. S.; Wu, X.G. , Yi, Q.J. Mulberry and dandelion water extracts prevent alcohol-induced steatosis with alleviating gut microbiome dysbiosis. Exp. Biol. Med. 2018, 243, 882–894. [Google Scholar] [CrossRef]
- Liu, N.; Song, M.; Wang, N.F.; Wang, Y.; Wang, R.F.; An, X.P.; Qi, J.W. The effects of solid-state fermentation on the content, composition and in vitro antioxidant activity of flavonoids from dandelion. Plos. One. 2020, 15, e0239076. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.R.; Wang, L.N.; Zhang, J.; Ke, W.C.; Zhou, J.W.; Zhu, J.X.; Guo, X.S.; Long, R.J. Characterization of antioxidant properties of lactic acid bacteria isolated from spontaneously fermented yak milk in the Tibetan Plateau. J. Funct. Foods. 2017, 35, 481–488. [Google Scholar] [CrossRef]
- Tian, X.Z.; Paengkoum, P.; Paengkoum, S.; Chumpawadee, S.; Ban, C.; Thongpea, S. Short communication: purple corn (Zea mays L.) stover silage with abundant anthocyanins transferring anthocyanin composition to the milk and increasing antioxidant status of lactating dairy goats. J. Dairy. Sci. 2018, 102, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.P.; Yu, Y.D.; Yu, Z.; Shao, T.; Na, R.S.; Zhao, M.M. Effects of lactic acid bacteria inoculants and cellulase on fermentation quality and in vitro digestibility of Leymus chinensis silage. Grassland. Sci. 2014, 60, 199–205. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy. Sci. 1991, 74, 3583–97. [Google Scholar] [CrossRef]
- Murphy, R. P. A method for the extraction of plant samples and the determination of total soluble carbohydrates. J. Sci. Food. Agr. 1958, 9, 714–717. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International; Association of Analytical Communities: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Xu, D.M.; Ding, W.R.; Ke, W.C.; Li, F.H.; Zhang, P.; Guo, X.S. Modulation of metabolome and bacterial community in whole crop corn silage by inoculating homofermentative Lactobacillus plantarum and heterofermentative Lactobacillus buchneri. Front. Microbiol. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Ni, K.K.; Wang, F.F.; Zhu, B.G.; Yang, J.X.; Zhou, G.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresoure. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef]
- Cai, Y.M.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Kumai, S.; Nakase, T. Influence of Lactobacillus spp. from an inoculant and of Weissella and Leuconostoc spp. from forage crops on silage fermentation. Appl. Environ. Microbiol. 1998, 64, 2982–2987. [Google Scholar] [CrossRef]
- Lee, M.R.F.; Scott, M.B.; Tweed, J.K.S.; Minchin, F.R.; Davies, D.R. Effects of polyphenol oxidase on lipolysis and proteolysis of red clover silage with and without a silage inoculant (Lactobacillus plantarum L54). Animal. Feed. Sci. Technol. 2008, 144, 125–136. [Google Scholar] [CrossRef]
- Hashemzadeh-Cigari, F.; Khorvash, M.; Ghorbani, G.R.; Ghasemi, E.; Taghizadeh, A.; Kargar, S.; Yang, W.Z. Interactive effects of molasses by homofermentative and heterofermentative inoculants on fermentation quality, nitrogen fractionation, nutritive value and aerobic stability of wilted alfalfa (Medicago sativa L) silage. J. Anim. Physiol. Anim. Nutr. 2014, 98, 290–299. [Google Scholar] [CrossRef]
- Nkosi, B.D.; Meeske, R.; van der Merwe, H.J.; Groenewald, I.B. Effects of homofermentative and heterofermentative bacterial silage inoculants on potato hash silage fermentation and digestibility in rams. Animal. Feed. Sci. Technol. 2010, 157, 195–200. [Google Scholar] [CrossRef]
- Albrecht, K.A.; Muck, R.E. Proteolysis in ensiled forage legumes that vary in tannin concentration. Crop. Sci. 1991, 31, 464–469. [Google Scholar] [CrossRef]
- Lu. Y.R.; Foo L.Y. The polyphenol constituents of grape pomace. Food. Chem. 1999, 65, 1–8. [Google Scholar] [CrossRef]
- Garcia, Y.D.; Valles, B.S.; Lobo, A.P. Phenolic and antioxidant composition of by-products from the cider industry: apple pomace. Food. Chem. 2009, 117, 731–738. [Google Scholar] [CrossRef]
- Endo, A.; Okada, S. Lactobacillus farraginis sp. nov. and Lactobacillus parafarraginis sp. nov., heterofermentative lactobacilli isolated from a compost of distilled shochu residue. Int. J. Syst. Evol. Microbiol. 2007, 57, 708–712. [Google Scholar] [CrossRef]
- Liu, Q.H.; Yang, F.Y.; Zhang, J.G.; Shao, T. Characteristics of Lactobacillus parafarraginis ZH1 and its role in improving the aerobic stability of silages. J. Appl. Microbiol. 2014, 117, 405–416. [Google Scholar] [CrossRef]
- Miller, J.K.; Brzezinska-Slebodzinska, E.; Madsen, F.C. Oxidative stress, antioxidants, and animal function. J. Dairy. Sci. 1993, 76, 2812–2823. [Google Scholar] [CrossRef]
- Liu, Q.H.; Shao, T.; Bai, Y.F. The effect of fibrolytic enzyme, Lactobacillus plantarum and two food antioxidants on the fermentation quality, alpha-tocopherol and beta-carotene of high moisture napier grass silage ensiled at different temperatures. Animal. Feed. Sci. Technol. 2016, 221, 1–11. [Google Scholar] [CrossRef]
- Kotsampasi, B.; Christodoulou, C.; Tsiplakou, E.; Mavrommatis, A.; Mitsiopoulou, C.; Karaiskou, C.; Dotas, V.; Robinson, P.H.; Bampidis, V.A.; Christodoulou, V.; Zervas, G. Effects of dietary pomegranate pulp silage supplementation on milk yield and composition, milk fatty acid profile and blood plasma antioxidant status of lactating dairy cows. Animal. Feed. Sci. Technol. 2017, 234, 228–236. [Google Scholar] [CrossRef]
- Khosravi, M.; Rouzbehan, Y.; Rezaei, M.; Rezaei, J. Total replacement of corn silage with sorghum silage improves milk fatty acid profile and antioxidant capacity of Holstein dairy cows. J. Dairy. Sci. 2018, 101, 10953–10961. [Google Scholar] [CrossRef]
- Tian, X.Z.; Paengkoum, P.; Paengkoum, S.; Chumpawadee, S.; Ban, C.; Thongpea, S. Purple corn (Zea mays L.) stover silage with abundant anthocyanins transferring anthocyanin composition to the milk and increasing antioxidant status of lactating dairy goats. J. Dairy. Sci. 2019, 102, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.X.; Ke, W.C.; Bai, J.; Li, F.H.; Xu, D.M.; Ding, Z.T.; Guo, X.S. The effect of Pediococcus acidilactici J17 with high-antioxidant activity on antioxidant, α-tocopherol, β-carotene, fatty acids, and fermentation profiles of alfalfa silage ensiled at two different dry matter contents. Animal. Feed. Sci. Technol. 2020, 268, 114614. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Huang, Z.P.; Vyas, D.; Adesogan, A.T.; Franco, M.; Ke, W.C.; Li, F.H.; Bai, J.; Ding, Z.T.; Guo, X.S. Antioxidant status, chemical composition and fermentation profile of alfalfa silage ensiled at two dry matter contents with a novel Lactobacillus plantarum strain with high-antioxidant activity. Animal. Feed. Sci. Technol. 2021, 272, 114751. [Google Scholar] [CrossRef]
- Ighodaro, O.M. , Akinloye, O.A.. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Bao, T.; Xu, Y.; Gowd, V.; Zhao, J.C.; Xie, J.H.; Liang, W.K.; Chen, W. Systematic study on phytochemicals and antioxidant activity of some new and common mulberry cultivars in China. J. Funct. Foods. 2016, 25, 537–547. [Google Scholar] [CrossRef]
- Jakub, T. , Karel, Š. Flavonoids as potent scavengers of hydroxyl radicals. Compr. Rev. Food. Sci. F. 2016, 15, 720–738. [Google Scholar] [CrossRef]
- Xu, Z.S.; He, H.Y.; Zhang, S.S.; Kong, J. Effects of inoculants Lactobacillus brevis and Lactobacillus parafarraginis on the fermentation characteristics and microbial communities of corn stover silage. Sci. Rep. 2017, 7, 13614. [Google Scholar] [CrossRef]
- Dong, Z.H.; Li, J.F.; Chen, L.; Wang, S.R.; Shao, T. Effects of freeze-thaw event on microbial community dynamics during red clover ensiling. Front. Microbiol. 2019, 10, 1559. [Google Scholar] [CrossRef]
- Zi, X.J.; Li, M.; Chen, Y.Y.; Lv, R.L.; Zhou, H.L.; Tang, J. Effects of citric acid and Lactobacillus plantarum on silage quality and bacterial diversity of king grass silage. Front. Microbiol. 2021, 12, 631096. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.H.; Lindow, S.E.; Zhang, J.G. Lactobacillus parafarraginis ZH1 producing anti-yeast substances to improve the aerobic stability of silage. Anim. Sci. J. 2018, 89, 1302–1309. [Google Scholar] [CrossRef]
- Heberline, J.A. Cyanobacteria: omics and manipulation. Environ. Prog. Sustain. 2017, 36, 980. [Google Scholar] [CrossRef]
- Li, D.X.; Ni, K.K.; Zhang, Y.C.; Lin, Y.L.; Yang, F.Y. Fermentation characteristics, chemical composition and microbial community of tropical forage silage under different temperatures. Asian-Australas. J. Anim. Sci. 2019, 32, 665–674. [Google Scholar] [CrossRef]
- He, L.W.; Lv, H.J.; Xing, Y.Q.; Chen, X.Y.; Zhang, Q. Intrinsic tannins affect ensiling characteristics and proteolysis of Neolamarckia cadamba leaf silage by largely altering bacterial community. Bioresource. Technol. 2020, 311, 123496. [Google Scholar] [CrossRef]
- Dunière, L.; Sindou, J.; Chaucheyras-Durand, F.; Chevallier, I.; Sergentet, D. Silage processing and strategies to prevent persistence of undesirable microorganisms. Animal. Feed. Sci. Technol. 2013, 182, 1–15. [Google Scholar] [CrossRef]
- Ogunade, I.M.; Jiang, Y.; Pech Cervantes, A.A.; Kim, D.H.; Oliveira, A.S.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: effects of Escherichia coli. O157:H7 and silage additives. J. Dairy Sci. 2018, 101, 2048–2059. [Google Scholar] [CrossRef]
- Wang, Y.; He, L.W.; Xing, Y.Q.; Zhou, W.; Pian, R.Q.; Yang, F.Y.; Chen, X.Y.; Zhang, Q. Bacterial diversity and fermentation quality of Moringa oleifera leaves silage prepared with lactic acid bacteria inoculants and stored at different temperatures. Bioresource. Technol. 2019, 284, 349–358. [Google Scholar] [CrossRef]
- Yang, L.L.; Yuan, X.J.; Li, J.F.; Dong, Z.H.; Shao, T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresource. Technol. 2019, 275, 280–7. [Google Scholar] [CrossRef] [PubMed]
- Danner, H.; Holzer, M.; Mayrhuber, E.; Braun, R. Acetic acid increases stability of silage under aerobic conditions. Appl. Environ. Microb. 2003, 69, 562–567. [Google Scholar] [CrossRef]
Figure 1.
Principal coordinate analysis of the microbial communities of silage. CON, control group; FT1, silage treated with 1% FT; FT2, silage treated with 2% FT.
Figure 1.
Principal coordinate analysis of the microbial communities of silage. CON, control group; FT1, silage treated with 1% FT; FT2, silage treated with 2% FT.
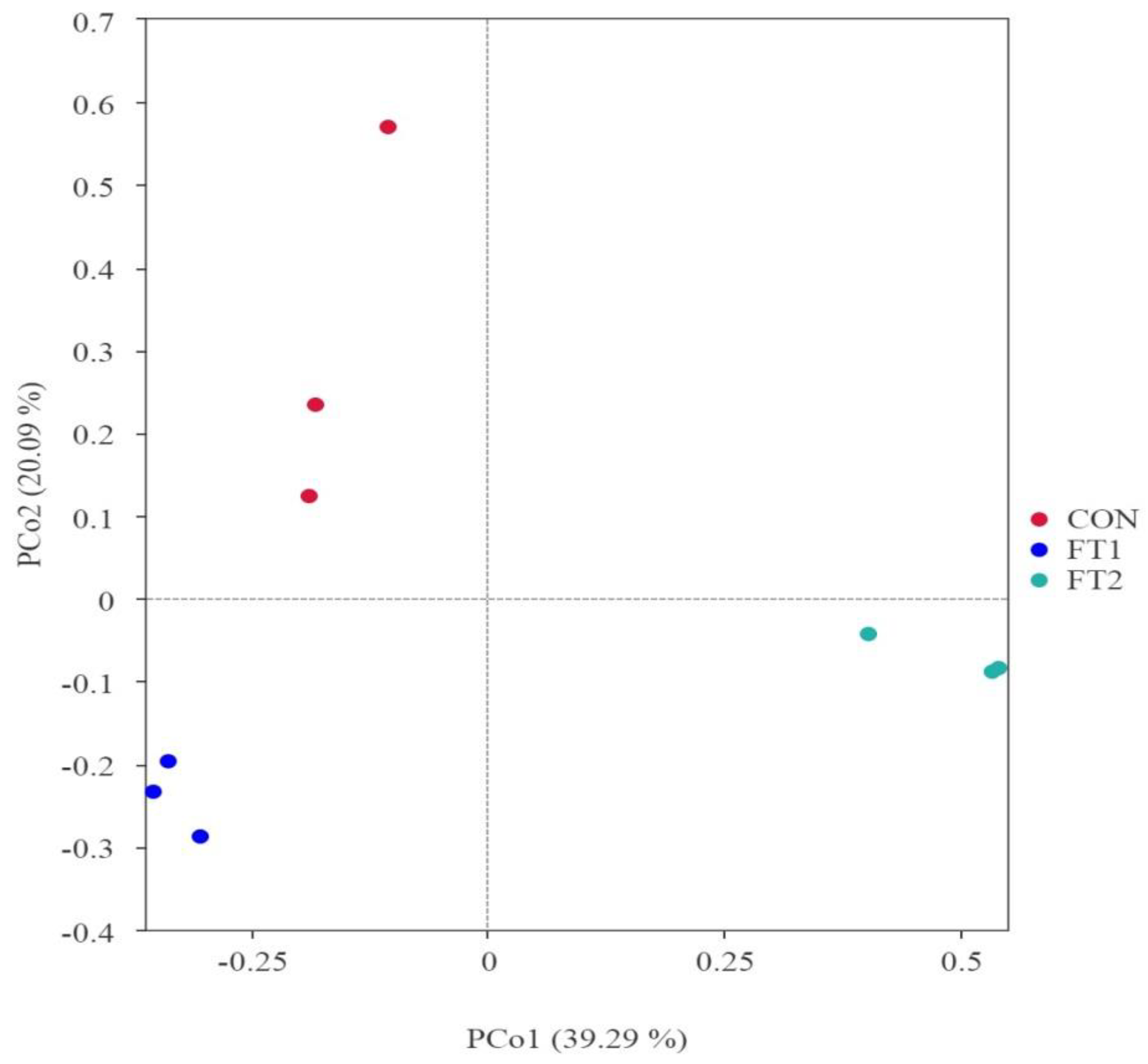
Figure 2.
The microbial abundance at the phylum (A), genus (B) and species (C) levels in silage. CON, control group; FT1, silage treated with 1% FT; FT2, silage treated with 2% FT.
Figure 2.
The microbial abundance at the phylum (A), genus (B) and species (C) levels in silage. CON, control group; FT1, silage treated with 1% FT; FT2, silage treated with 2% FT.
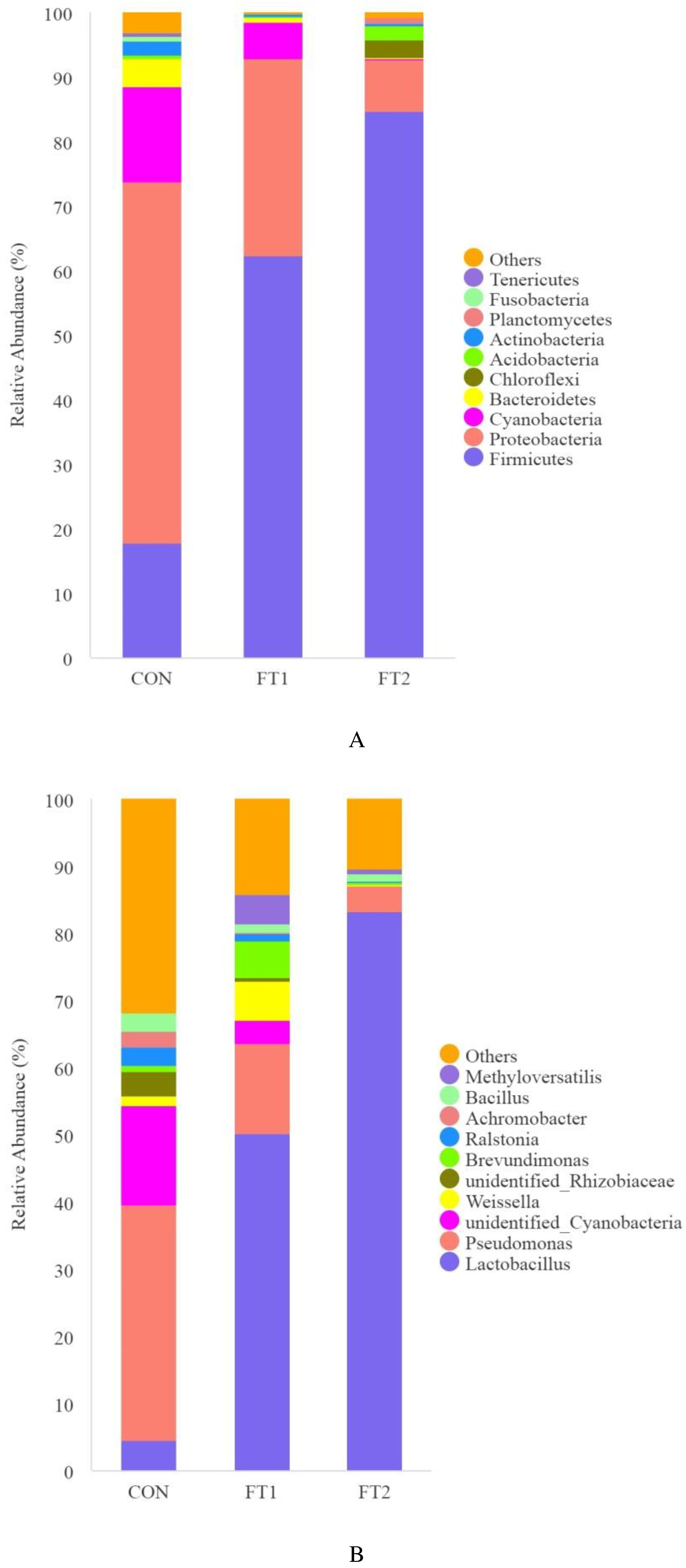

Figure 3.
LDA value distribution and evolutionary branch of different species in silage treated with different additives. (A) LDA value distribution of different species. (B) Evolutionary branch diagram of various species. FT1, silage treated with 1% FT; FT2, silage treated with 2% FT.
Figure 3.
LDA value distribution and evolutionary branch of different species in silage treated with different additives. (A) LDA value distribution of different species. (B) Evolutionary branch diagram of various species. FT1, silage treated with 1% FT; FT2, silage treated with 2% FT.
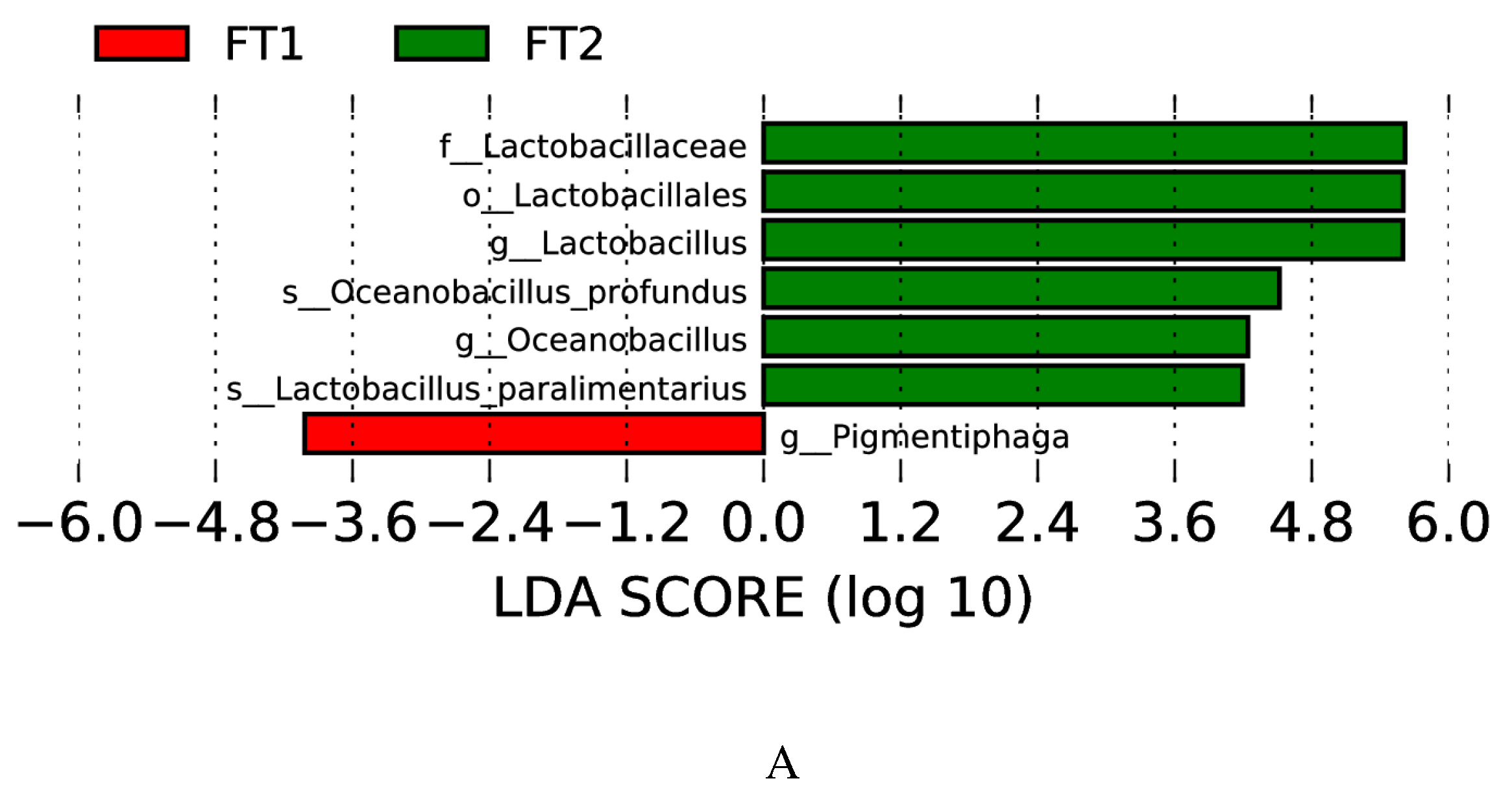
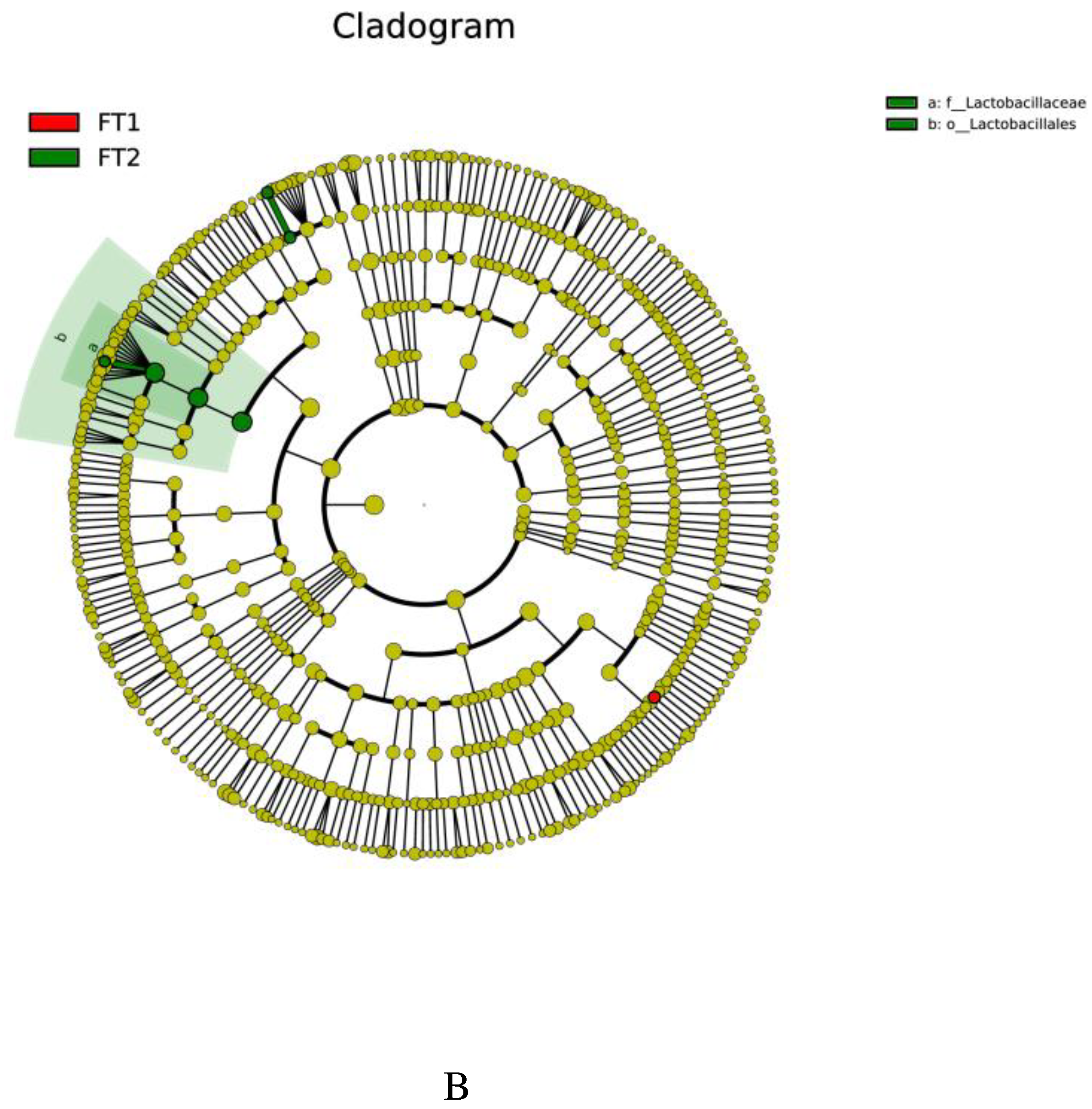
Figure 4.
The correlation between microorganisms and fermentation parameters, antioxidant capacity at the species level using Spearman’s correlation analysis. LA, lactic acid; AA, acetic acid; AN, NH3-N; LAB, lactic acid bacteria; TAOC, total anti-oxidation competence; SOD, superoxide dismutase; GPX, glutathione peroxidase; CAT, catalase. * represents p < 0.05 and ** represent p < 0.01.
Figure 4.
The correlation between microorganisms and fermentation parameters, antioxidant capacity at the species level using Spearman’s correlation analysis. LA, lactic acid; AA, acetic acid; AN, NH3-N; LAB, lactic acid bacteria; TAOC, total anti-oxidation competence; SOD, superoxide dismutase; GPX, glutathione peroxidase; CAT, catalase. * represents p < 0.05 and ** represent p < 0.01.
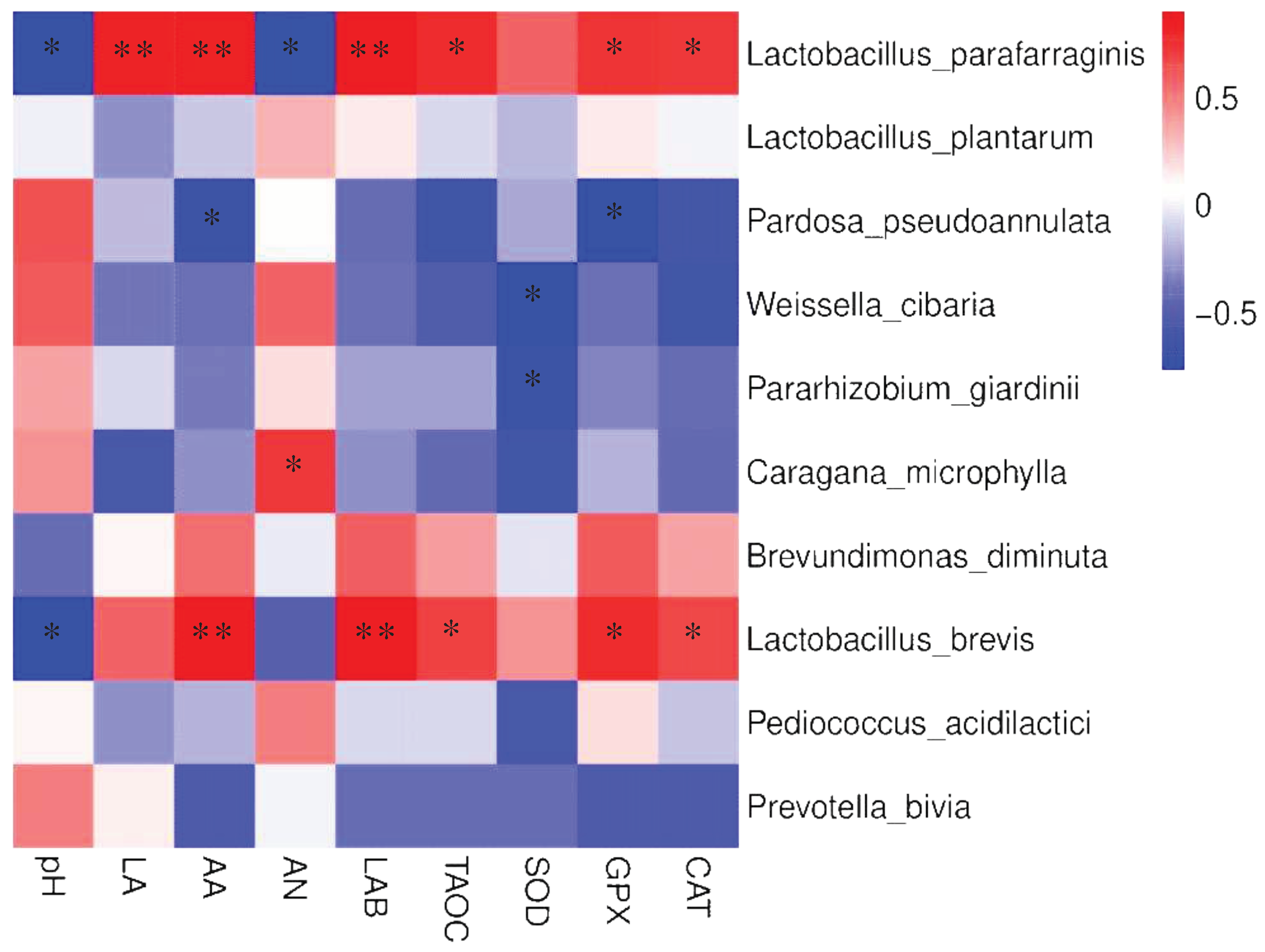

Table 1.
Chemical and microbial compositions of CK before ensiled.
| Items1 | CK |
|---|---|
| DM (g/kg FM) | 426.21 |
| CP (g/kg DM) | 64.79 |
| EE (g/kg DM) | 25.20 |
| WSC (g/kg DM) | 10.12 |
| NDF (g/kg DM) | 801.81 |
| ADF (g/kg DM) | 664.81 |
| LAB (log10 cfu/g FM) | 3.19 |
| Yeasts (log10 cfu/g FM) | 0.00 |
| Molds (log10 cfu/g FM) | 3.86 |
1 DM, dry matter; CP, crude protein; EE, ether extract; WSC, water-soluble carbohydrates; NDF, neutral detergent fiber; ADF, acid detergent fiber; cfu, colony forming unit; LAB, lactic acid bacteria; FM, fresh matter.
Table 2.
Effects of FT on the fermentation quality of CK silage.
| Items1 | Treatments2 | SEM3 | p-Value | ||
|---|---|---|---|---|---|
| CON | 1% FT | 2% FT | |||
| pH | 3.78 a | 3.77 a | 3.72 b | 0.01 | <0.01 |
| Lactic acid (g/kg DM) | 37.02 b | 37.32 b | 44.69 a | 1.23 | <0.01 |
| Acetic acid (g/kg DM) | 2.90 b | 4.02 b | 12.28 a | 1.05 | <0.01 |
| Propanoic acid (g/kg DM) | ND | ND | ND | ND | - |
| Butyrate (g/kg DM) | ND | ND | ND | ND | - |
| NH3-N (g/kg TN−1) | 14.65 a | 14.40 a | 11.65 b | 0.48 | <0.01 |
| LAB (log 10 cfu/g FM) | 2.49 c | 3.72 b | 4.50 a | 0.22 | <0.01 |
| Yeasts (log 10 cfu/g FM) | ND | ND | ND | ND | - |
| Molds (log 10 cfu/g FM) | ND | ND | ND | ND | - |
Different letters indicate significant differences in the same row (p < 0.05). 1 DM, dry matter; NH3-N, ammonia nitrogen; TN, total nitrogen; LAB, lactic acid bacteria; cfu, colony forming unit; FM, fresh matter. ND, not detected. 2 CON, control group; 1% FT, silage treated with 1% FT; 2% FT, silage treated with 2% FT. 3 SEM, standard error of the mean.
Table 3.
Effects of FT on the chemical composition of CK silage.
| Items1 | Treatments2 | SEM3 | p-Value | ||
|---|---|---|---|---|---|
| CON | 1% FT | 2% FT | |||
| DM (g/kg FM) | 417.14 c | 423.53 b | 428.10 a | 1.20 | <0.01 |
| CP (g/kg DM) | 65.85 b | 66.13 b | 67.40 a | 0.18 | <0.01 |
| WSC (g/kg DM) | 3.50 | 3.62 | 3.57 | 0.06 | 0.73 |
| EE (g/kg DM) | 44.39 | 43.58 | 42.28 | 0.54 | 0.28 |
| NDF (g/kg DM) | 760.98 a | 758.31 a | 737.85 b | 3.71 | <0.05 |
| ADF (g/kg DM) | 647.40 a | 643.68 a | 603.86 b | 5.27 | <0.01 |
Different letters indicate significant differences in the same row (p < 0.05). 1 DM, dry matter; FM, fresh matter; CP, crude protein; WSC, water-soluble carbohydrate; EE, ether extract; NDF, neutral detergent fiber; ADF, acid detergent fiber. 2 CON, control group; 1% FT, silage treated with 1% FT; 2% FT, silage treated with 2% FT. 3 SEM, standard error of the mean.
Table 4.
Effects of FT on the antioxidant capacity of CK silage.
| Items1 | Treatments2 | SEM3 | p-Value | ||
|---|---|---|---|---|---|
| CON | 1% FT | 2% FT | |||
| T-AOC (mmol/g Prot) | 72.74 c | 84.51 b | 90.45 a | 2.66 | <0.01 |
| SOD (U/g FW) | 173.19 b | 172.67 b | 215.11 a | 7.80 | <0.01 |
| GSH-Px (U/g FW) | 210.41 b | 320.27 a | 334.14 a | 19.94 | <0.01 |
| CAT (U/g FW) | 8.08 c | 16.22 b | 21.73 a | 1.99 | <0.01 |
Different letters indicate significant differences in the same row (p < 0.05). 1 T-AOC, total anti-oxidation competence; SOD, superoxide dismutase; GSH-Px, glutathione peroxidase; CAT, catalase; Prot, protein of the homogenate; FW, fresh weight. 2 CON, control group; 1% FT, silage treated with 1% FT; 2% FT, silage treated with 2% FT. 3 SEM, standard error of the mean.
Table 5.
Effects of FT on the alpha-diversity of microbial communities of CK silage.
| Items | Treatments1 | SEM2 | p-Value | ||
|---|---|---|---|---|---|
| CON | 1% FT | 2% FT | |||
| Ace | 378.95 | 215.13 | 199.38 | 80.02 | 0.66 |
| Chao1 | 364.49 | 197.63 | 188.53 | 77.34 | 0.64 |
| Simpson | 0.61 | 0.72 | 0.37 | 0.12 | 0.52 |
| Shannon | 3.89 | 3.26 | 1.85 | 0.75 | 0.59 |
| Coverage | 0.99 | 0.99 | 0.99 | 0.00 | 0.66 |
1 CON, control group; 1% FT, silage treated with 1% FT; 2% FT, silage treated with 2% FT. 2 SEM, standard error of the mean.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



