
Submitted:
01 September 2023
Posted:
06 September 2023
You are already at the latest version
Abstract
Cryopreserved semen is widely used in Assisted Reproductive Technologies (ART) enabling the conservation and broad use of genetically superior semen. The quality of semen post-thawing determines the success of ART and despite the great advances in cryopreservation methods, the quality of frozen/thawed semen is still sub-optimal. Post-thawing spermatozoa endure oxidative stress (OS) due to the high levels of reactive oxygen and nitrogen species, which are produced during the freezing/thawing process, and the depletion of antioxidants. To counteract this depletion, supplementation of sperm preparation medium with antioxidants has been widely applied. Melatonin is a hormone with diverse biological roles and a potent antioxidant, with an ameliorative effect on spermatozoa. In the present study, the effect of melatonin on bovine spermatozoa was evaluated during in vitro sperm handling and under oxidative conditions in terms of sperm quality parameters and antioxidant capacity. Melatonin (100 μΜ) improved the kinematic parameters of spermatozoa, enhanced viability and reinforced the antioxidant status of spermatozoa by increasing cellular GSH levels and inhibiting iNOS protein expression.
Keywords:
antioxidant capacity
; glutathione
; hydrogen peroxide
; nitric oxide synthase
; oxidative stress
; su-peroxide ion
; sperm physiology
; reactive oxygen species
1. Introduction
The use of cryopreserved semen is widely applied in assisted reproductive technologies (ART) for the apparent reason that it enables broad use and conservation of genetically superior semen. Undoubtedly, the quality of semen post-thawing determines the success of all processes applied in ART [1]. Despite the advances in cryopreservation of bull semen, the viability of spermatozoa after thawing is still relatively low and highly variable [2], mainly because of an imbalance in the redox status of spermatozoa. The high content of polyunsaturated fatty acids (PUFA) in the cell membranes of spermatozoa render them particularly vulnerable to oxidation by reactive oxygen species (ROS), which are over-produced by spermatozoa during freezing procedures [3]. Moreover, freezing induces alterations in membrane fluidity, which damage mitochondrial membranes, thus increasing mitochondrial membrane potential and producing excessive amounts of ROS [4,5]. During their preparation for ART, spermatozoa are deprived of the antioxidants that are present in the seminal plasma, their intracellular antioxidant defenses are depleted by the overload of ROS produced during freeze/thawing, and thus sustain OS manifested as lipid peroxidation (LPO), DNA damage and apoptosis [3,6,7]. To prevent this, researchers have employed supplementation of antioxidants in the sperm preparation media [8,9,10,11,12]. Nevertheless, preparation of spermatozoa for artificial insemination (AI) or in vitro embryo production (IVEP) protocols, apart from the freeze-thawing procedure, requires several additional steps of handling. Antioxidant supplementation has been considered as a potential approach to preserve the fertilizing ability of spermatozoa during these processes and to improve male reproductive performance. Thus, the role of antioxidants in male reproductive physiology has been investigated in vivo through oral supplementation or in vitro as supplements in culture media. [13,14]). Currently, the consensus is that the inclusion of antioxidants in the sperm preparation media may significantly improve the quality parameters of spermatozoa (for a review see [9]).
Melatonin is an amphiphilic hormone produced in several tissues but mainly in the pineal gland [15]. In addition to its apparently diverse biological roles, such as regulation of the circadian cycle, immune response, and seasonal reproduction [16], melatonin is also a potent antioxidant [17]. Its antioxidant properties are manifested either directly by neutralization of free radicals or indirectly by inducing the activity and expression of key antioxidant enzymes, such as glutathione peroxidase, catalase, and superoxide dismutase (for a review see [15]). Melatonin is also present in seminal plasma [18]. The ameliorative effect of melatonin as an additive in sperm preparation media has been assessed in many species, such as human [19,20], red deer [21], sheep [22], swine [23] and cattle [24,25,26,27]. In addition to scavenging ROS, melatonin may also exert its biological effect through its receptors MT1 and MT2, which are classic G protein-coupled receptors (GPCRs) that stimulate downstream signaling cascades by activating G proteins [20,28,29]. It appears that melatonin may mediate its biological effects in a dose-dependent manner [30,31,32]. To date the consensus is that at micromolar concentrations melatonin reduces ROS and cAMP levels thus modulating sperm capacitation, acrosome reaction and fertilization rate [21,27,33,34]. At nanomolar concentrations melatonin interacts with intracellular ligands and modulates the activity of antioxidant enzymes, whereas at picomolar concentrations it binds to cell membrane receptors, thus triggering downstream signaling pathways [35,36]. Melatonin action has also been correlated with the production of nitric oxide (NO) and the expression of the inducible isoform of nitric oxide synthase (iNOS) in macrophages and other cell types [37]. The NO synthases (NOSs) comprise a family of enzymes which utilize oxygen and the amino acid L-arginine to produce the highly reactive molecule NO [38]. NO and the nitrogen species which are derived from NO mediate important roles in reproductive processes.
The addition of natural or synthetic antioxidants in the medium of spermatozoa is an effective approach to prevent OS and thus counteract the negative effects of ROS on spermatozoa during ART [39,40,41]. We have previously shown the ameliorative effects of crocin and crocetin on post-thawing spermatozoa [11,42,43], while melatonin has a proven biological role in reproductive processes by acting either as an antioxidant or as a ligand for the melatonin receptors [16,18,28]. The aim of the present study was to investigate if supplementation of the in vitro sperm preparation media with melatonin can preserve the quality characteristics of post-thawing spermatozoa over time after thawing, anticipating its use to prevent sperm from oxidative attack in procedures of AI or IVEP protocols.
2. Materials and Methods
2.1. Reagents
All reagents were purchased from Sigma Aldrich Co. (Darmstadt, Germany), unless otherwise specified.
Sperm TALP (Tyrode's albumin lactate pyruvate) containing 100 mm NaCl, 3.1 mm KCl, 25 mm NaHCO3, 0.29 mm NaH2PO4, 21.6 mm Na lactate, 2 mm CaCl2, 1.5 mm MgCl2, 10 mm Hepes, 1 mm sodium pyruvate and 50 μg/mL gentamycin in water for embryo transfer [44] was equilibrated overnight at 37 C in a 5% CO2 humidified atmosphere before use.
A fresh stock 10 mM solution of melatonin (Cayman, Clinical, Ann Arbor, Michigan, USA) was prepared before each experiment. Melatonin was diluted in dimethylosulfoxide (DMSO), which reached a final concentration of 1% (v/v) to all samples, including the controls (vehicle).
RIPA buffer consisted of 50 mM Tris-HCl, pH 7,2, 150 mM NaCl, 1% NP-40, 1mM EDTA, 1 mM EGTA and 1% (v/v) of protease inhibitor cocktail (Sigma-Aldrich, Darmstadt, Germany).
2.2. Semen collection, sample preparation and experimental design
Six mature, healthy bulls (3 Simmental, 2 Holstein and 1 Brown Swiss) of proven fertility (housed at the Department of Artificial Insemination, Directorate of Veterinary Centre of Thessaloniki, National Ministry of Rural Development and Food, Ionia, Greece) were used as sperm donors throughout the study. The age of the animals ranged from 48 to 60 months at the beginning of the experiment. Four different ejaculates were collected from each bull with the use of an artificial vagina. Samples were collected from all animals at the same period of the year (November 2022 - February 2023). Ejaculates were evaluated by the same person; only semen characterized with >70% initial motility, >75% viability, and a total concentration of at least 4 × 109 spermatozoa/ejaculate was cryopreserved. Semen was diluted in a Tris-citric acid home-made extender containing glycerol and egg yolk (20% Tris-egg yolk, 7% glycerol, 78 mM citric acid, 69 mM fructose, 50 μg tylosin, 250 μg gentamycin, 150 μg lincomycin, 300 μg spectinomycin in each ml of extended frozen semen) and packed into 0.5 ml plastic straws with 50 × 106 spermatozoa/ml. The straws were labeled with the name and registration code of the bull and the date of freezing prior to freezing. In brief, straws were allowed to equilibrate at 5οC for 4 h after which they were placed on a horizontal rack, 3 cm above the surface of liquid nitrogen in order to be bathed in liquid nitrogen vapors and thus reach -100οC within 7–10 min. Once this temperature was reached, the straws were plunged and stored in liquid nitrogen (-196°C). The storage period of semen in liquid nitrogen ranged from 2 to 24 months.
One straw (from a single ejaculate) was used every time for each analysis. The straws were thawed by immersion in a 37°C water-bath for 40 sec and pooled together into a sterile plastic tube (CellstarTubes, Greiner Bio One, Frickenhausen, Germany), washed twice with 3 × volumes of Sperm TALP and centrifuged at 1500 rpm for 10 min at room temperature (RT). Sperm concentration was determined by cell count on a Neubauer chamber (OptikLabor, Grale HDS, New South Wales, Australia). Finally, the pellet was re-suspended in Sperm TALP to a final concentration of 50 × 106 cells/ml and the samples were divided in four groups, with a total volume of 100 μl each. Two served as Control and were incubated in TALP with 1% DMSO (vehicle), while the other two were supplemented with 100 μΜ melatonin. All samples were incubated in a 37°C water-bath for 60 min and aliquots were removed to be analyzed for each assay at the beginning of the incubation (time point t0) and after 60 min, at which time point (t60) 100 μΜ οf H2O2 were added to the appropriate vials and aliquots were also analyzed. All assays were repeated 6 times, unless otherwise indicated.
2.3. Motility Assessment
Motility assessment was performed by a Computer Assisted Sperm Analyzer (CASA), using Integrated Semen Analysis System Software (ISAS MvCo, Valencia, Spain). The CASA system consisted of a triocular optical phase microscope (Nicon Eclipse C1, Nikon, Tokyo, Japan), equipped with a heated plate (Tokai, Tokyo, Japan) and a Baler Scout CCD digital camera (Basler Vision Technologies, Ahrensburg, Germany), connected to a computer. Each time a 5-μl-aliquot of sperm suspension was placed on a pre-warmed slide on the heated plate at 37°C and 9 different parameters were measured: spermatozoa with rapid, medium, slow movement, static spermatozoa, progressive motility (PM), curvilinear velocity (VCL), straight line velocity (VSL), average path velocity, (VAP), amplitude lateral head, (ALH). CASA default settings: 60 frames/second, total of 25 frames captured; minimum contrast of 80 and medium cell size of 5 pixels; a cutoff value for progressive cells of 50 μm/sec for VAP and 70% for medium threshold straightness; static cells: VAP cutoff 25 μm/sec and VSL cutoff 10 μm/sec.).
2.4. Viability Assessment
At the indicated time points (0, 60 min), 10 μl of each sample were mixed with an equal volume of trypan blue solution and cells were smeared on a slide [45]. Two hundred spermatozoa in each slide were evaluated microscopically (×1000) as alive (unstained) or dead (stained).
2.5. Preparation of whole cell lysates
To measure intracellular content, spermatozoa were resuspended in RIPA buffer and subjected to two cycles of freeze-thawing followed by two cycles of sonication at 28 kHz for 30 sec. Whole cell lysates were centrifuged at 5000 × g for 10 min to remove debris and the supernatant was used for further analysis.
2.6. Determination of Intracellular Glutathione
To determine intracellular GSH levels, spermatozoa lysates were incubated with 0.33 mM DTNB [5,5’-dithiobis (2-nitrobenzoic acid)] [46]. The thiol groups of GSH in the lysates cleave the disulfide bond in DTNB to yield 2-nitro-5-thiobenzoic acid, which ionizes to the TNB2− dianion that has a yellow color and can be quantified at 412 nm, using a spectrophotometer (Pharmacia LKB-Novaspec II, U.K.). Results were expressed as percentage (%) of Control.
2.7. Determination of the Intracellular superoxide anion content
The nitroblue-tetrazolium (NBT) assay was performed to evaluate the intracellular generation of the superoxide anion (O2−) [47]. The NBT test is based on the production of blue-purple NBT formazan crystals, as a result of the reduction of yellow NBT chloride (2,2'-bis(4-nitrophenyl)-5,5'-diphenyl-3,3'-(3,3'-dimethoxy-4,4'diphenylene) to ditetrazolium chloride by O2−. The NBT was dissolved in Dulbecco’s PBS at 5 mg/ml and administered to the whole cell suspension. Intact spermatozoa were incubated with NBT (final concentration 0.5 mg/ml) for 60 min and subsequently, were washed twice with PBS and centrifuged (300 × g) for 10 min. Cells and the formed formazan crystals were dissolved in 120 μl of 2 M ΝaOH and 140 μl of DMSO. Optical density was assessed at 630 nm with a ΒioTek EL800 microplate reader (Thomas Scientific, Swedesboro, NJ, U.S.A.).
2.8. Western Blot Analysis
The intracellular levels of inducible nitric oxide synthase (iNOS) were determined by Western blotting as previously described [48]. A total of 50 μg of total protein were analyzed on a 10% SDS polyacrylamide gel and then transferred on nitrocellulose membrane. iNOS protein was detected with the iNOS antibody (1:1000) and a goat anti-Rabbit IgG alkaline phosphatase conjugated antibody (1:10,000) (Proteintech Group, Manchester, UK). Nitroblue tetrazolium (NBT), 5-bromo-4-chloro-3- indolyl phosphate (BCIP) and p-nitrophenyl phosphate were used for the alkaline phosphatase reaction. iNOS protein quantitation was performed by densitometry of the appropriate molecular weight band using the ImageJ 1.53a software. The analysis was performed 4 times with samples from independent experiments.
2.9. Statistical analysis
Statistical tests exploited in the study were chosen after performing analyses for the normality of residuals. Non-parametric tests have been employed when values did not follow the Gaussian distribution when verified by the normality tests run (Anderson-Darling, D'Agostino-Pearson omnibus, Shapiro-Wilk, and Kolmogorov-Smirnov tests). A standard one-way ANOVA was performed for the statistical examination of rapid motility, viability and NBT test. Kruskal-Wallis test, followed by Dunn’s multiple comparison for progressive motility, total motility and viability. GSH data were processed with the Kruskal–Wallis test and pairwise comparisons were performed with the Mann–Whitney U test.
The statistical significance level was set at p≤0.05. All experimental data were analyzed with the SPSS version 25.0 (IBM Corp., Armonk, New York, USA) and are presented as mean ± SEM.
3. Results
3.1. Melatonin inhibits the loss of motility over time during the handling of spermatozoa
Frozen-thawed spermatozoa undergo OS-induced damage during their preparation for ART. In this experimental set up, incubation of bovine spermatozoa at 37°C for 60 min negatively affected their motility parameters (Table 1). The % of motile spermatozoa (total motile in Table 1) was reduced during the 60 min from 77.05% at t0 to 64.98% (p=0.0016) at t60 (Figure 1A) and concomitantly the % of static spermatozoa was increased (Table 1). Rapidly moving spermatozoa (rapid) were reduced from 49.59% at t0 to 27.67% (p<0.0001) at t60 (Figure 1B), while the % of spermatozoa with slow motility increased from 5.12% at t0 to 8.87% at t60 (Table 1). Although progressive motility was not affected, VCL (p=0.0053) and ALH (p=0.0152) were also reduced, whereas VSL and VAP showed a tendency to decline.
When 100 μΜ melatonin was included in the incubation medium, the motility was not affected during the 60 min incubation period, as no change was observed in any of the motility parameters between the Control or the Melatonin groups at t0 and the Melatonin group at t60 (Table 1). The group that was incubated in the presence of melatonin presented higher % of motile spermatozoa (72.68%, p=0.0655) and higher % of rapid spermatozoa (43.22%, p=0.0002) compared to the Control group at the same time point (Figure 1). Moreover, VCL (p=0.007), VAP (p=0.0494) and ALH (p=0.01) at the end of the 60 min incubation, were higher in the melatonin treated spermatozoa compared to the control cells (Table 1).
3.2. In the presence of melatonin, spermatozoa retain their motility during an insult by hydrogen peroxide
The deterioration of spermatozoa over time during their handling has been attributed to the generation of ROS [49]. To augment this effect, by the end of the 60-min incubation spermatozoa were exposed to 100 μM H2O2. As anticipated, hydrogen peroxide had overall a negative effect on the motility of spermatozoa and mediated a further reduction of the motility parameters at t60 (Table 1). In particular, hydrogen peroxide induced an additional reduction in total motility from 64.68% (Control group at t60) to 56.03% (p=0.0471, Figure 2A). Melatonin alleviated this effect and retained the % of motile spermatozoa to levels (70.32%) higher than the HP group (p=0.0015) and similar to those of the Melatonin group (72.68%) (Table 1).
In the presence of melatonin, the percentage of rapid spermatozoa exposed to hydrogen peroxide remained at levels (32.2%) that were even higher than the % of Control cells at t60 (27.67%), but with no statistical significance. In any case, melatonin inhibited the loss of rapid spermatozoa in the presence of HP (32.2% in the Melatonin-HP group versus 20.48% in the HP group, p=0.0043) (Table 1, Figure 2B). Melatonin had similar effect on other kinematic parameters, as spermatozoa incubated in the presence of 100 μM melatonin retained higher VCL, VAP and ALH values after exposure to hydrogen peroxide compared to the control H2O2-treated cells (Table 1). These data demonstrate that pre-incubation of spermatozoa with melatonin preserves their movement characteristics even under conditions of acute increase of hydrogen peroxide.
3.3. Melatonin protects viability of spermatozoa against oxidative stress
After thawing, 62.5% of the spermatozoa were alive but during the 60-min incubation at 37°C this % was reduced to 40.63% (p<0.001) after 60 min (Figure 3). Supplementation with melatonin effectively inhibited this reduction in viability, increasing the % of alive spermatozoa both in the presence (56.22%, p=0.0107) or absence (59.56%, p=0.0018) of hydrogen peroxide (Figure 3 and Figure 4). The insult by hydrogen peroxide at t60 had no effect on viability compared to the untreated cells but in any case, both melatonin treated groups had higher % of alive cells (p<0.01) compared to the control spermatozoa treated with hydrogen peroxide (Figure 4).
3.4. Melatonin increases the intracellular GSH content of spermatozoa
Hydrogen peroxide depleted spermatozoa of their GSH reserves, as determined by the colorimetric GSH assay and demonstrated in Figure 5. In particular, spermatozoa that were pre-incubated at 37°C for 60 min and then exposed to 100 μΜ H2O2 had 22% lower GSH levels compared to the Control group (p=0.007). Melatonin supplementation increased GSH levels both in the presence (p=0.063) and absence of H2O2 (p=0.015). No statistically significant differences were observed between the Melatonin and the Melatonin+HP group.
3.5. Melatonin does not affect the intracellular levels of superoxide ion)
Because superoxide ion (O2−) is the major ROS of human spermatozoa [50], we determined the effect of hydrogen peroxide and melatonin on the levels of O2− in spermatozoa in this experimental system. Surprisingly, exposure of spermatozoa to 100 μM H2O2 after a 60 min-incubation slightly decreased O2− levels (p>0.1) compared to the Control at t60 (Figure 6). Melatonin also reduced intracellular O2− to equal levels, irrespective to the presence of H2O2 (Figure 6, black bars), but this effect also had no statistical significance.
3.6. Melatonin reduces the inducible nitric oxide synthase (iNOS) protein levels
The expression of the inducible isoform of NOS (iNOS) by spermatozoa was determined by Western blot analysis. Hydrogen peroxide had no effect on iNOS levels (Figure 7, white bars). Preincubation of spermatozoa for 60 min at 37°C in the presence of 100 μM melatonin slightly reduced iNOS (88.71%) compared to the Control group (arbitrarily set at 100%), but this effect was not statistically significant. Interestingly, the addition of hydrogen peroxide in the melatonin-treated spermatozoa further decreased iNOS levels to 50.53% (p=0.0489) of Control values (Figure 7).
4. Discussion
Crucial reproductive processes, both in male and female, are affected by ROS. Regarding spermatozoa, low and moderate concentrations of ROS are required for processes such as capacitation and acrosome reaction [3,51,52,53,54]. On the other hand, elevated amounts of ROS reduce sperm motility [55]. The excessive amounts of ROS produced during preparation of spermatozoa for ART are considered a major contributor to the relatively poor outcome of these processes, despite the developments achieved in ART [55,56]. In bovine the blastocyst rate that is achieved during IVF rarely exceeds 40–50% [57], which is similar to that obtained the previous decades [58]. The last decade various antioxidants have been tested as supplements in bovine ART to improve different steps of these processes, but their effects are still under discussion [8,9,59,60].
Melatonin is a ubiquitously expressed hormone with well documented antioxidant and free radical scavenging activities [61,62]. However, its exact molecular mechanism(s) of action is (are) yet to be described. Nevertheless, melatonin plays a rather important role in the physiology of reproduction [63,64,65,66] and has been considered as a supplement for media in ART [9,67].
In the present study, we assessed the effect of melatonin on frozen/thawed bovine spermatozoa during their incubation at 37°C and after their challenge with hydrogen peroxide. We also attempted to shed some light on the mechanisms by which melatonin elicits its action. The concentration of hydrogen peroxide was determined based on previous data [20,68,69,70] and pilot experiments. Griveau et al have demonstrated that 50 μΜ H2O2 is required to induce capacitation of bovine spermatozoa [68], so higher concentrations probably induce OS.
Despite the accumulating data in many species, such as human, ram and bovine [20,22,24,28,29,35,71], the role of melatonin in sperm physiology remains controversial. Regarding bovine spermatozoa, few studies focus on the antioxidant properties of melatonin [24,27,33,72]. There is a wide range in the concentrations of melatonin used in these experiments from nanomolar to millimolar, that could correspond to the different physiological concentrations of melatonin related to sex (male or female genital tract), different species [73], or experimental conditions [27].
In the present study, melatonin was supplemented in the medium at a 100 μM concentration, at which the molecule mediates an antioxidant role [20,70]. The spermatozoa that were incubated for 60 min at 37°C in the presence of 100 μΜ melatonin were more motile than the corresponding Control group, as determined by different motility parameters (Figure 1 and Figure 2, Table 1). In addition, melatonin increased viability attested by the higher percentage of alive spermatozoa at the end of the 60-min period (Figure 3). Melatonin not only increased the motility and viability of spermatozoa, but it also rendered them more capable in sustaining the insult by hydrogen peroxide in terms of motility (Table 1, Figure 2) compared to the Control group. Fernandez-Alegre et al. reported that melatonin differentially affects spermatozoa depending on the species and the experimental conditions [27]. In red deer, 0.1 mM or 1 mM melatonin did not improve sperm motility after a 4-h incubation [21]. On the other hand, the same concentration of melatonin (1 mM) improved the motility of human spermatozoa after a 30-minute incubation [72] and the fertilizing capacity of normozoospermic and oligozoospermic samples [19]. The work by Li et al established that experimental conditions have a great impact on the outcome of melatonin supplementation [24].
The ameliorative effect on bovine spermatozoa appears to be attributed to the antioxidant properties of melatonin. Relatively low concentrations of melatonin (10 μM) decreased the levels of ROS and improved the antioxidant status of sex-sorted bovine spermatozoa by increasing the activities of key antioxidant enzymes, such as glutathione peroxidase, superoxide ion dismutase and catalase [24]. In the current study, the ameliorative effect of melatonin on the motility and viability of spermatozoa was accompanied by a concomitant increase in intracellular GSH levels (Figure 5), although it had no effect on superoxide ion levels (Figure 6). This could be attributed to the reaction of superoxide ions with NO to form peroxinitrate ions (ONOO-) [74], an interaction of highly reactive molecules characterized by Korkmaz et al as the “devil’s triangle” [75]. Melatonin is known to directly scavenge ROS and NO and directly detoxify ONOO- [76], thus reducing overall the reactive molecules in the cell milieu. On the other hand, melatonin may inhibit the production of NO from L-arginine. The enzymes that regulate this reaction are called NO synthases (NOS), and one particular isoform, the inducible NOS (iNOS) is regulated at the transcriptional level [77]. Therefore, we assessed the modulation of iNOS protein levels in bovine spermatozoa by melatonin to demonstrate that melatonin decreases the expression of iNOS in cells exposed to hydrogen peroxide (Figure 7). This is in agreement with the work of Gilad et al, who demonstrated that melatonin, at concentrations between 1 μM and 1 mM inhibits the expression of iNOS in cultured murine macrophages stimulated with lipopolysaccharide [37]. Thus, melatonin may reduce not only ROS but also reactive nitrogen species (RNS) in spermatozoa. Alternatively, the beneficial effect of melatonin on the motility of spermatozoa could be attributed to changes induced in cytoskeletal elements of spermatozoa mediated by the interaction of melatonin with calmodulin, and preservation of ATP reserves, which are required to maintain flagellum activity [78]. Korkmaz et al have stated that melatonin is the only currently available molecule which is known to block all aspects of the “devil’s triangle” as well as modulate the expression of antioxidant proteins, thereby preventing a redox imbalance and thus oxidative stress.
Nevertheless, the differential effects of melatonin on different types of cells appear to be cell type- and concentration-dependent. As mentioned above, melatonin at micromolar concentrations modulates sperm capacitation in spermatozoa, whereas at nano or picomolar concentrations does not affect sperm motility [35]. These reports, along with the data presented here, clearly impose the need for further studies where a broad range of melatonin concentrations under different conditions (capacitating or not) will be tested in order to elucidate the mechanisms that mediate the effects of melatonin on spermatozoa.
5. Conclusions
the results of the present study demonstrate that during the first hour after thawing of cryopreserved spermatozoa, melatonin improved the motility and viability of spermatozoa, even under induced OS (exposure to hydrogen peroxide) either through its antioxidant activities or by inhibiting iNOS. We anticipate that these results will set a starting point for future studies, in order to optimize the applications of melatonin in ART.
Author Contributions
Conceptualization, V.S., M.P.T; Methodology, S.N.L., V.S., M.P.T; Experimental analysis, S.N.L., V.S., E.T., I.M.; Data validation and analysis, S.N.L., V.S., E.T, M.P.T.; Statistical analysis , E.T., I.M.; Draft Preparation, S.N.L., V.S., M.P.T.
Funding
This research did not receive any specific grants from funding agencies in public, commercial, or non-profit organizations.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon request (S.N.L., slavrent@vet.auth.gr).
Acknowledgments
The authors are grateful to Dr. Ioannis Panagiotidis, DVM and the Department of Artificial Insemination, Directorate of Veterinary Centre of Thessaloniki, National Ministry of Rural Development and Food, Ionia, Greece for providing the semen samples and Mr. Ioannis Drosos for providing technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Martinez, M.; Majzoub, A. Best Laboratory Practices and Therapeutic Interventions to Reduce Sperm DNA Damage. Andrologia 2021, 53. [CrossRef]
- Ugur, M.R.; Saber Abdelrahman, A.; Evans, H.C.; Gilmore, A.A.; Hitit, M.; Arifiantini, R.I.; Purwantara, B.; Kaya, A.; Memili, E. Advances in Cryopreservation of Bull Sperm. Front Vet Sci 2019, 6, 268. [CrossRef]
- Lenzi, A.; Picardo, M.; Gandini, L.; Dondero, F. Lipids of the Sperm Plasma Membrane: From Polyunsaturated Fatty Acids Considered as Markers of Sperm Function to Possible Scavenger Therapy. Hum Reprod Update 1996, 2, 246–256. [CrossRef]
- Wang, A.W.; Zhang, H.; Ikemoto, I.; Anderson, D.J.; Loughlin, K.R. Reactive Oxygen Species Generation by Seminal Cells during Cryopreservation. Urology 1997, 49, 921–925. [CrossRef]
- Said, T.M.; Gaglani, A.; Agarwal, A. Implication of Apoptosis in Sperm Cryoinjury. Reprod Biomed Online 2010, 21, 456–462. [CrossRef]
- Safarinejad, M.R.; Hosseini, S.Y.; Dadkhah, F.; Asgari, M.A. Relationship of Omega-3 and Omega-6 Fatty Acids with Semen Characteristics, and Anti-Oxidant Status of Seminal Plasma: A Comparison between Fertile and Infertile Men. Clinical Nutrition 2010, 29, 100–105. [CrossRef]
- Baldi, E.; Tamburrino, L.; Muratori, M.; Degl’Innocenti, S.; Marchiani, S. Adverse Effects of in Vitro Manipulation of Spermatozoa. Anim Reprod Sci 2020, 220, 106314. [CrossRef]
- Tsantarliotou, M.P.; Sapanidou, V.G. The Importance of Antioxidants in Sperm Quality and in Vitro Embryo Production. Journal of Veterinary Andrology 2018, 3, 1–12.
- Sapanidou, V.; Tsantarliotou, M.P.; Lavrentiadou, S.N. A Review of the Use of Antioxidants in Bovine Sperm Preparation Protocols. Anim Reprod Sci 2023, 251, 107215. [CrossRef]
- Sapanidou, V.; Taitzoglou, I.; Tsakmakidis, I.; Kourtzelis, I.; Fletouris, D.; Theodoridis, A.; Lavrentiadou, S.; Tsantarliotou, M. Protective Effect of Crocetin on Bovine Spermatozoa against Oxidative Stress during in Vitro Fertilization. Andrology 2016, 4, 1138–1149. [CrossRef]
- Sapanidou, V.; Lavrentiadou, S.N.; Errico, M.; Panagiotidis, I.; Fletouris, D.; Efraimidis, I.; Zervos, I.; Taitzoglou, I.; Gasparrini, B.; Tsantarliotou, M. The Addition of Crocin in the Freezing Medium Extender Improves Post-Thaw Semen Quality. Reproduction in Domestic Animals 2022, 57, 269–276. [CrossRef]
- Pintus, E.; Ros-Santaella, J.L. Impact of Oxidative Stress on Male Reproduction in Domestic and Wild Animals. Antioxidants 2021, 10, 1154. [CrossRef]
- Pintus, E.; Luis, J.; Santaella, R.-; Drevet, J.R.; Sorrentino, R. Impact of Oxidative Stress on Male Reproduction in Domestic and Wild Animals. 2021. [CrossRef]
- Bathgate, R. Antioxidant Mechanisms and Their Benefit on Post-Thaw Boar Sperm Quality. Reproduction in Domestic Animals 2011, 46, 23–25. [CrossRef]
- Hacışevki, Aysun, and B.B. An Overview of Melatonin as an Antioxidant Molecule: A Biochemical Approach. In Melatonin - Molecular Biology, Clinical and Pharmaceutical Approaches; Drăgoi, C.M., Nicolae, A.C., Eds.; IntechOpen, 2018; pp. 59–85 ISBN 978-1-78984-504-4.
- Tsantarliotou, M.P.; Kokolis, N.A.; Smokovitis, A. Melatonin Administration Increased Plasminogen Activator Activity in Ram Spermatozoa. Theriogenology 2008, 69, 458–465. [CrossRef]
- Lialiaris, T.S. Melatonin in Prevention of Mutagenesis, Oxidation and Other Damage to Cells.; 2011.
- Casao, A.; Cebrián, I.; Asumpção, M.E.; Pérez-Pé, R.; Abecia, J.A.; Forcada, F.; Cebrián-Pérez, J.A.; Muiño-Blanco, T. Seasonal Variations of Melatonin in Ram Seminal Plasma Are Correlated to Those of Testosterone and Antioxidant Enzymes. Reproductive Biology and Endocrinology 2010, 8, 59. [CrossRef]
- Monllor, F.; Espino, J.; Marchena, A.M.; Ortiz, Á.; Lozano, G.; García, J.F.; Pariente, J.A.; Rodríguez, A.B.; Bejarano, I. Melatonin Diminishes Oxidative Damage in Sperm Cells, Improving Assisted Reproductive Techniques. Turkish Journal of Biology 2017, 41, 881–889. [CrossRef]
- Espino, J.; Ortiz, Á.; Bejarano, I.; Lozano, G.M.; Monllor, F.; García, J.F.; Rodríguez, A.B.; Pariente, J.A. Melatonin Protects Human Spermatozoa from Apoptosis via Melatonin Receptor- and Extracellular Signal-Regulated Kinase-Mediated Pathways. Fertil Steril 2011, 95, 2290–2296. [CrossRef]
- Domínguez-Rebolledo, A.E.; Fernández-Santos, M.R.; Bisbal, A.; Ros-Santaella, J.L.; Ramón, M.; Carmona, M.; Martínez-Pastor, F.; Garde, J.J. Improving the Effect of Incubation and Oxidative Stress on Thawed Spermatozoa from Red Deer by Using Different Antioxidant Treatments. Reprod Fertil Dev 2010, 22, 856–870. [CrossRef]
- Casao, A.; Abecia, J.A.; Cebrián-Pérez, J.A.; Muiño-Blanco, T.; Vázquez, M.I.; Forcada, F. The Effects of Melatonin on in Vitro Oocyte Competence and Embryo Development in Sheep | Efecto de La Melatonina En La Competencia Del Oocito y El Desarrollo Embrionario Ovino in Vitro. Spanish Journal of Agricultural Research 2010, 8, 35–41. [CrossRef]
- Jang, H.Y.; Kim, Y.H.; Kim, B.W.; Park, I.C.; Cheong, H.T.; Kim, J.T.; Park, C.K.; Kong, H.S.; Lee, H.K.; Yang, B.K. Ameliorative Effects of Melatonin against Hydrogen Peroxide-Induced Oxidative Stress on Boar Sperm Characteristics and Subsequent in Vitro Embryo Development. Reproduction in Domestic Animals 2010, 45, 943–950. [CrossRef]
- Li, C.-Y.; Hao, H.-S.; Zhao, Y.-H.; Zhang, P.-P.; Wang, H.-Y.; Pang, Y.-W.; Du, W.-H.; Zhao, S.-J.; Liu, Y.; Huang, J.-M.; et al. Melatonin Improves the Fertilization Capacity of Sex-Sorted Bull Sperm by Inhibiting Apoptosis and Increasing Fertilization Capacitation via MT1. Int J Mol Sci 2019, 20. [CrossRef]
- Pang, Y.-W.; Sun, Y.-Q.; Jiang, X.-L.; Huang, Z.-Q.; Zhao, S.-J.; Du, W.-H.; Hao, H.-S.; Zhao, X.-M.; Zhu, H.-B. Protective Effects of Melatonin on Bovine Sperm Characteristics and Subsequent in Vitro Embryo Development. Mol Reprod Dev 2016, 83, 993–1002. [CrossRef]
- Carlos Gutié Rrez-Añez Id, J.; Henning, H.; Lucas-Hahn, A.; Baulain, U.; Aldag, P.; Sieg, B.; Hensel, V.; Herrmann, D.; Niemann, H. Melatonin Improves Rate of Monospermic Fertilization and Early Embryo Development in a Bovine IVF System. 2021. [CrossRef]
- Fernández-Alegre, E.; Álvarez-Fernández, I.; Domínguez, J.C.; Casao, A.; Martínez-Pastor, F. Melatonin Non-Linearly Modulates Bull Spermatozoa Motility and Physiology in Capacitating and Non-Capacitating Conditions. Int J Mol Sci 2020, 21. [CrossRef]
- Casao, A.; Gallego, M.; Abecia, J.A.; Forcada, F.; Pérez-Pé, R.; Muiño-Blanco, T.; Cebrián-Pérez, J.Á. Identification and Immunolocalisation of Melatonin MT(1) and MT(2) Receptors in Rasa Aragonesa Ram Spermatozoa. Reprod Fertil Dev 2012, 24, 953–961. [CrossRef]
- Gonzalez-Arto, M.; Luna, C.; Pérez-Pé, R.; Muiño-Blanco, T.; Cebrián-Pérez, J.A.; Casao, A. New Evidence of Melatonin Receptor Contribution to Ram Sperm Functionality. Reprod Fertil Dev 2016, 28, 924–935. [CrossRef]
- Succu, S.; Berlinguer, F.; Pasciu, V.; Satta, V.; Leoni, G.G.; Naitana, S. Melatonin Protects Ram Spermatozoa from Cryopreservation Injuries in a Dose-Dependent Manner. J Pineal Res 2011, 50, 310–318. [CrossRef]
- Fujinoki, M. Melatonin-Enhanced Hyperactivation of Hamster Sperm. REPRODUCTION 2008, 136, 533–541. [CrossRef]
- Gwayi, N.; Bernard, R.T.F. The Effects of Melatonin on Sperm Motility in Vitro in Wistar Rats. Andrologia 2002, 34, 391–396. [CrossRef]
- Ashrafi, I.; Kohram, H.; Ardabili, F.F. Antioxidative Effects of Melatonin on Kinetics, Microscopic and Oxidative Parameters of Cryopreserved Bull Spermatozoa. Anim Reprod Sci 2013, 139, 25–30. [CrossRef]
- Cheuquemán, C.; Arias, M.E.; Risopatrón, J.; Felmer, R.; Álvarez, J.; Mogas, T.; Sánchez, R. Supplementation of IVF Medium with Melatonin: Effect on Sperm Functionality and in Vitro Produced Bovine Embryos. Andrologia 2015, 47, 604–615. [CrossRef]
- Gimeno-Martos, S.; Casao, A.; Yeste, M.; Cebrián-Pérez, J.A.; Muiño-Blanco, T.; Pérez-Pé, R. Melatonin Reduces CAMP-Stimulated Capacitation of Ram Spermatozoa. Reprod Fertil Dev 2019, 31, 420–431. [CrossRef]
- Tomás-Zapico, C.; Coto-Montes, A. Melatonin as Antioxidant under Pathological Processes. Recent Patents on Endocrine, Metabolic & Immune Drug Discov- ery 2007, 1, 63–82. [CrossRef]
- Gilad, E.; Wong, H.R.; Zingarelli, B.; Virág, L.; O’connor, M.; Salzman, A.L.; Szabó, C. Melatonin Inhibits Expression of the Inducible Isoform of Nitric Oxide Synthase in Murine Macrophages: Role of Inhibition of NFκB Activation. The FASEB Journal 1998, 12, 685–693. [CrossRef]
- Luo, Y.; Zhu, Y.; Basang, W.; Wang, X.; Li, C.; Zhou, X. Roles of Nitric Oxide in the Regulation of Reproduction: A Review. Front Endocrinol (Lausanne) 2021, 12. [CrossRef]
- Catt, J.W.; Henman, M. Toxic Effects of Oxygen on Human Embryo Development. Hum Reprod 2000, 15 Suppl 2, 199–206. [CrossRef]
- Gadea, J.; García-Vazquez, F.; Matás, C.; Gardón, J.C.; Cánovas, S.; Gumbao, D. Cooling and Freezing of Boar Spermatozoa: Supplementation of the Freezing Media with Reduced Glutathione Preserves Sperm Function. J Androl 2005, 26, 396–404. [CrossRef]
- Gadea, J.; Gumbao, D.; Cá novas, S.; Alberto García-Vá zquez, F.; Alberto Grulló, L.; Carlos Gardó, J. Supplementation of the Dilution Medium after Thawing with Reduced Glutathione Improves Function and the in Vitro Fertilizing Ability of Frozen-Thawed Bull Spermatozoa. [CrossRef]
- Sapanidou, V.; Taitzoglou, I.; Tsakmakidis, I.; Kourtzelis, I.; Fletouris, D.; Theodoridis, A.; Zervos, I.; Tsantarliotou, M. Antioxidant Effect of Crocin on Bovine Sperm Quality and in Vitro Fertilization. Theriogenology 2015, 84, 1273–1282. [CrossRef]
- Sapanidou, V.; Taitzoglou, I.; Tsakmakidis, I.; Kourtzelis, I.; Fletouris, D.; Theodoridis, A.; Lavrentiadou, S.; Tsantarliotou, M. Protective Effect of Crocetin on Bovine Spermatozoa against Oxidative Stress during in Vitro Fertilization. Andrology 2016. [CrossRef]
- Vajta, G.; Holm, P.; Greve, T.; Callesen, H. Factors Affecting Survival Rates of in Vitro Produced Bovine Embryos after Vitrification and Direct In-Straw Rehydration. Anim Reprod Sci 1996, 45, 191–200. [CrossRef]
- Mayer, D.T.; Squiers, C.D.; Bogart, R.; Oloufa, M.M. The Technique for Characterizing Mammalian Spermatozoa as Dead or Living by Differential Staining. J Anim Sci 1951, 10, 226–235. [CrossRef]
- Papadopoulou, A.; Petrotos, K.; Stagos, D.; Gerasopoulos, K.; Maimaris, A.; Makris, H.; Kafantaris, I.; Makri, S.; Kerasioti, E.; Halabalaki, M.; et al. Enhancement of Antioxidant Mechanisms and Reduction of Oxidative Stress in Chickens after the Administration of Drinking Water Enriched with Polyphenolic Powder from Olive Mill Waste Waters. Oxid Med Cell Longev 2017, 2017.
- Becerra, M.C.; Eraso, A.J.; Albesa, I. Comparison of Oxidative Stress Induced by Ciprofloxacin and Pyoverdin in Bacteria and in Leukocytes to Evaluate Toxicity. Luminescence 2003, 18, 334–340. [CrossRef]
- Katsipis, G.; Tzekaki, E.E.; Tsolaki, M.; Pantazaki, A.A. Salivary GFAP as a Potential Biomarker for Diagnosis of Mild Cognitive Impairment and Alzheimer’s Disease and Its Correlation with Neuroinflammation and Apoptosis. J Neuroimmunol 2021, 361, 577744. [CrossRef]
- Gualtieri, R.; Kalthur, G.; Barbato, V.; Longobardi, S.; Rella, F. Di; Adiga, S.K.; Talevi, R.; Moretti, E. Sperm Oxidative Stress during In Vitro Manipulation and Its Effects on Sperm Function and Embryo Development. Antioxidants 2021, 10. [CrossRef]
- Toor, J.S.; Sikka, S.C. Chapter 1.6 - Human Spermatozoa and Interactions With Oxidative Stress. In Oxidants, Antioxidants and Impact of the Oxidative Status in Male Reproduction; Henkel, R., Samanta, L., Agarwal, A., Eds.; Academic Press, 2019; pp. 45–53 ISBN 978-0-12-812501-4.
- O’Flaherty, C.; de Lamirande, E.; Gagnon, C. Positive Role of Reactive Oxygen Species in Mammalian Sperm Capacitation: Triggering and Modulation of Phosphorylation Events. Free Radic Biol Med 2006, 41, 528–540. [CrossRef]
- de Lamirande, E.; O’Flaherty, C. Sperm Activation: Role of Reactive Oxygen Species and Kinases. Biochim Biophys Acta Proteins Proteom 2008, 1784, 106–115. [CrossRef]
- Bansal, A.K.; Bilaspuri, G.S. Impacts of Oxidative Stress and Antioxidants on Semen Functions. Vet Med Int 2011, 2011. [CrossRef]
- Bansal, A.K.; Bilaspuri, G.S. Oxidative Stress Alters Membrane Sulfhydryl Status, Lipid and Phospholipid Contents of Crossbred Cattle Bull Spermatozoa. Anim Reprod Sci 2008, 104, 398–404. [CrossRef]
- Agarwal, A.; Said, T.M.; Bedaiwy, M.A.; Banerjee, J.; Alvarez, J.G. Oxidative Stress in an Assisted Reproductive Techniques Setting. Fertil Steril 2006, 86, 503–512. [CrossRef]
- Nicole McPherson, and; Agarwal, A.; Maldonado Rosas, I.; Anagnostopoulou, C.; Cannarella, R.; Boitrelle, F.; Villar Munoz, L.; Finelli, R.; Durairajanayagam, D.; Henkel, R.; et al. Oxidative Stress and Assisted Reproduction: A Comprehensive Review of Its Pathophysiological Role and Strategies for Optimizing Embryo Culture Environment. Mexico; imaldonado@citmer.mx (I.M.R 1152, 11, 477. [CrossRef]
- Lonergan, P.; Fair, T. Maturation of Oocytes in Vitro. Annu Rev Anim Biosci 2016, 4, 255–268. [CrossRef]
- Rizos, D.; Clemente, M.; Bermejo-Alvarez, P.; De La Fuente, J.; Lonergan, P.; Gutiérrez-Adán, A. Consequences of in Vitro Culture Conditions on Embryo Development and Quality. Reproduction in Domestic Animals 2008, 43, 44–50. [CrossRef]
- Martins da Silva, S.J. DNA Fragmentation, Antioxidants and ART. In Assisted Reproduction Techniques; 2021; pp. 606–611 ISBN 9781119622215.
- Agarwal, A.; Durairajanayagam, D.; Du Plessis, S.S. Utility of Antioxidants during Assisted Reproductive Techniques: An Evidence Based Review; 2014;
- Reiter, RusselJ.; Tan, D.-X.; Cabrera, J.; D’Arpa, D.; Sainz, RosaM.; Mayo, J.; Ramos, S. The Oxidant/Antioxidant Network: Role of Melatonin. Biol Signals Recept 1999, 8, 56–63. [CrossRef]
- Ahmad, B.; Ali, A.; Bilal, M.; Rashid, S.M.; Amir, ·; Wani, B.; Rahil, ·; Bhat, R.; Muneeb, ·; Rehman, U.; et al. Melatonin and Health: Insights of Melatonin Action, Biological Functions, and Associated Disorders. Mol Neurobiol 2023, 43, 2437–2458. [CrossRef]
- Feng, X.; Zhang, Y.; Li, N.; Zhang, Y.; Zheng, Q.; Sun, M.; Tang, J.; Xu, Z. Melatonin in Reproductive Medicine: A Promising Therapeutic Target? Curr Med Chem 2023, 30, 3090–3118. [CrossRef]
- Dehdari Ebrahimi, N.; Sadeghi, A.; Ala, M.; Ebrahimi, F.; Pakbaz, S.; Azarpira, N. Protective Effects of Melatonin against Oxidative Stress Induced by Metabolic Disorders in the Male Reproductive System: A Systematic Review and Meta-Analysis of Rodent Models. Front Endocrinol (Lausanne) 2023, 14. [CrossRef]
- Dehdari Ebrahimi, N.; Shojaei-Zarghani, S.; Taherifard, E.; Dastghaib, S.; Parsa, S.; Mohammadi, N.; Sabet Sarvestani, F.; Moayedfard, Z.; Hosseini, N.; Safarpour, H.; et al. Protective Effects of Melatonin against Physical Injuries to Testicular Tissue: A Systematic Review and Meta-Analysis of Animal Models. Front Endocrinol (Lausanne) 2023, 14. [CrossRef]
- Silva, B.R.; Silva, J.R.V. Mechanisms of Action of Non-Enzymatic Antioxidants to Control Oxidative Stress during in Vitro Follicle Growth, Oocyte Maturation, and Embryo Development. Anim Reprod Sci 2023, 249. [CrossRef]
- Role, W.; S Silva, A.M.; Kim, Y.-B.; Li, Z.; Zhang, K.; Zhou, Y.; Zhao, J.; Wang, J.; Lu, W. Citation: Molecules Role of Melatonin in Bovine Reproductive Biotechnology. 2023, 28. [CrossRef]
- Griveau, J.E.; Renard, P.; Lannou, D.L. An in Vitro Promoting Role for Hydrogen Peroxide in Human Sperm Capacitation. Int J Androl 1994, 17, 300–307. [CrossRef]
- Rivlin, J.; Mendel, J.; Rubinstein, S.; Etkovitz, N.; Breitbart, H. Role of Hydrogen Peroxide in Sperm Capacitation and Acrosome Reaction1. Biol Reprod 2004, 70, 518–522. [CrossRef]
- ChaithraShree, A.R.; Ingole, S.D.; Dighe, V.D.; Nagvekar, A.S.; Bharucha, S. V; Dagli, N.R.; Kekan, P.M.; Kharde, S.D. Effect of Melatonin on Bovine Sperm Characteristics and Ultrastructure Changes Following Cryopreservation. Vet Med Sci 2020, 6, 177–186. [CrossRef]
- Li, C.; Zhou, X. Melatonin and Male Reproduction. Clinica Chimica Acta 2015, 446, 175–180. [CrossRef]
- Ortiz, A.; Espino, J.; Bejarano, I.; Lozano, G.M.; Monllor, F.; García, J.F.; Pariente, J.A.; Rodríguez, A.B. High Endogenous Melatonin Concentrations Enhance Sperm Quality and Short-Term in Vitro Exposure to Melatonin Improves Aspects of Sperm Motility. J Pineal Res 2010, 50, 132–139. [CrossRef]
- Cebrián-Pérez, J.A.; Casao, A.; González-Arto, M.; dos Santos Hamilton, T.R.; Pérez-Pé, R.; Muiño-Blanco, T. Melatonin in Sperm Biology: Breaking Paradigms. Reproduction in Domestic Animals 2014, 49, 11–21. [CrossRef]
- Szabó, C.; Ischiropoulos, H.; Radi, R. Peroxynitrite: Biochemistry, Pathophysiology and Development of Therapeutics. Nat Rev Drug Discov 2007, 6, 662–680. [CrossRef]
- Korkmaz, A.; Reiter, R.J.; Topal, T.; Manchester, L.C.; Oter, S.; Tan, D.-X. Melatonin: An Established Antioxidant Worthy of Use in Clinical Trials. Molecular Medicine 2009, 15, 43–50. [CrossRef]
- Du Plessis, S.S.; Hagenaar, K.; Lampiao, F. The in Vitro Effects of Melatonin on Human Sperm Function and Its Scavenging Activities on NO and ROS. Andrologia 2010, 42, 112–116. [CrossRef]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric Oxide Synthases: Structure, Function and Inhibition. Biochemical Journal 2001, 357, 593–615. [CrossRef]
- Pandi-Perumal, S.R.; Trakht, I.; Srinivasan, V.; Spence, D.W.; Maestroni, G.J.M.; Zisapel, N.; Cardinali, D.P. Physiological Effects of Melatonin: Role of Melatonin Receptors and Signal Transduction Pathways. Prog Neurobiol 2008, 85, 335–353. [CrossRef]
Figure 1.
The effect of melatonin on the motility of spermatozoa. Frozen-thawed spermatozoa were incubated at 37°C for 60 min in the presence (Melatonin) or absence (Vehicle) of 100 μM melatonin. The % of motile spermatozoa (A) and the % of spermatozoa with rapid motility (B) were determined by CASA and the results are presented as mean values (% of total spermatozoa) ± SEM. The asterisk (*) indicates statistically significant differences (p<0.001, n=6).
Figure 1.
The effect of melatonin on the motility of spermatozoa. Frozen-thawed spermatozoa were incubated at 37°C for 60 min in the presence (Melatonin) or absence (Vehicle) of 100 μM melatonin. The % of motile spermatozoa (A) and the % of spermatozoa with rapid motility (B) were determined by CASA and the results are presented as mean values (% of total spermatozoa) ± SEM. The asterisk (*) indicates statistically significant differences (p<0.001, n=6).
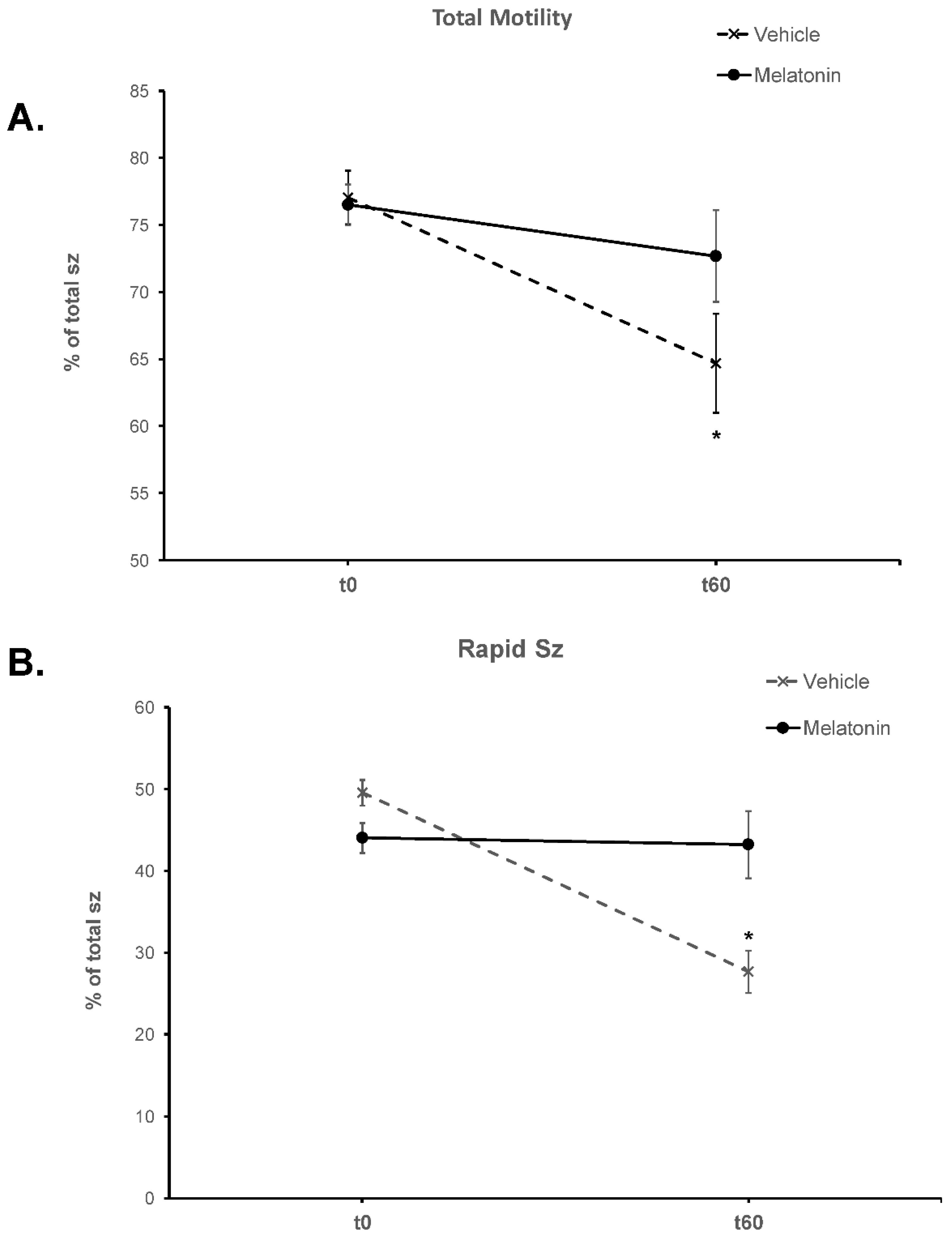
Figure 2.
The effect of melatonin on the percentage of total motility (A) and rapid spermatozoa (B) after exposure to 100 μΜ hydrogen peroxide. Cells were incubated for 60 min in the presence (black bars) or absence (white bars) of 100 μΜ melatonin, at which point the indicated samples were exposed to 100 μΜ hydrogen peroxide (HP). The % of spermatozoa with total (A) or rapid (B) motility were determined by CASA. Results are presented as mean values (% of total spermatozoa) ± SEM and different letters indicate statistically significant differences (p<0.05, n=6).
Figure 2.
The effect of melatonin on the percentage of total motility (A) and rapid spermatozoa (B) after exposure to 100 μΜ hydrogen peroxide. Cells were incubated for 60 min in the presence (black bars) or absence (white bars) of 100 μΜ melatonin, at which point the indicated samples were exposed to 100 μΜ hydrogen peroxide (HP). The % of spermatozoa with total (A) or rapid (B) motility were determined by CASA. Results are presented as mean values (% of total spermatozoa) ± SEM and different letters indicate statistically significant differences (p<0.05, n=6).
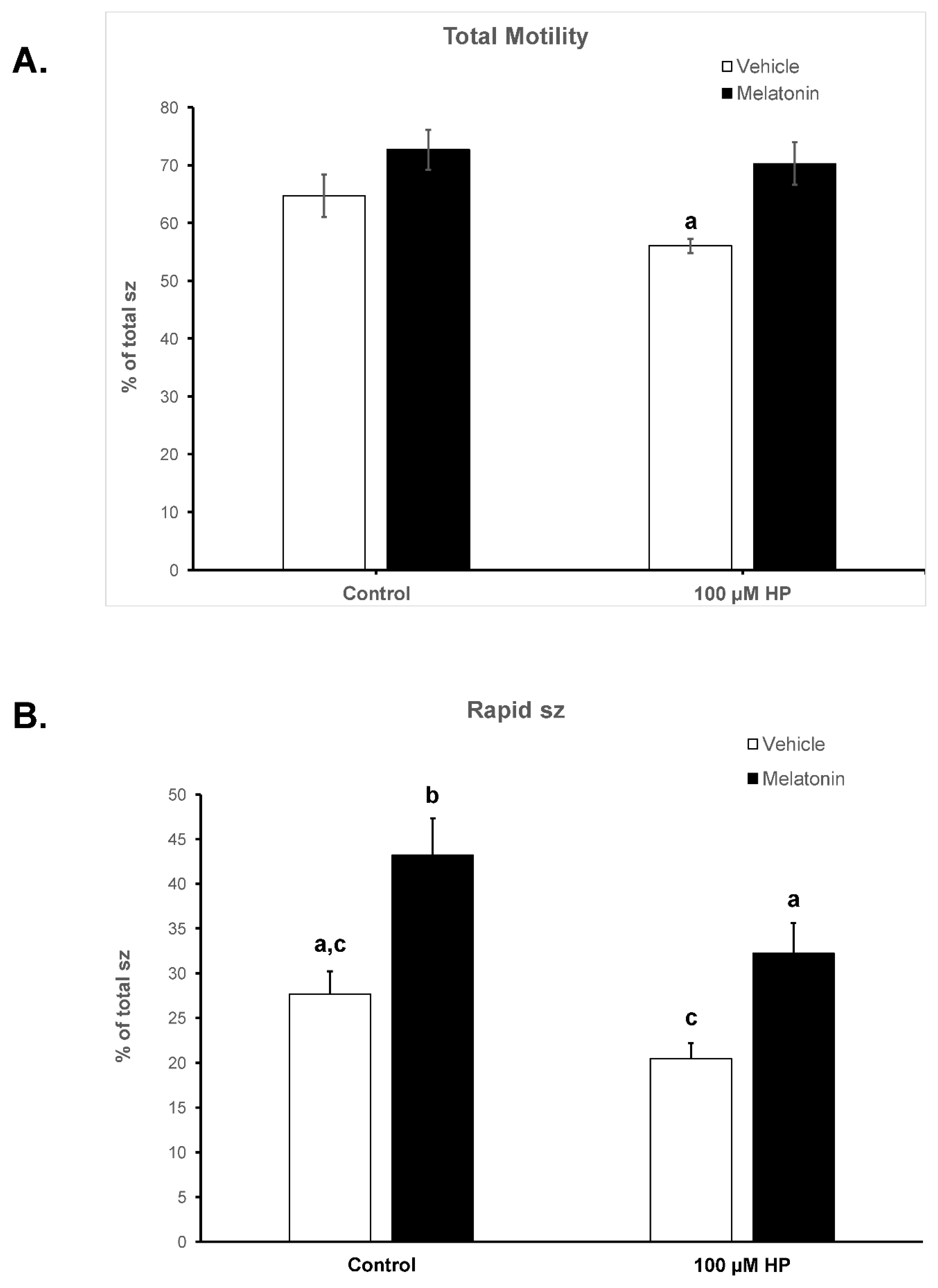
Figure 3.
The effect of melatonin on the viability of spermatozoa. Frozen-thawed spermatozoa were incubated at 37°C for 60 min in the presence (Melatonin) or absence (Vehicle) of 100 μM melatonin. The % of alive spermatozoa was determined by optical microscopy after staining of spermatozoa with trypan blue solution. The results are presented as mean values (% of total spermatozoa) ±SEM (n=6). The asterisk (*) indicates statistically significant different values from the t0 or the t60-Melatonin group (p≤0.01).
Figure 3.
The effect of melatonin on the viability of spermatozoa. Frozen-thawed spermatozoa were incubated at 37°C for 60 min in the presence (Melatonin) or absence (Vehicle) of 100 μM melatonin. The % of alive spermatozoa was determined by optical microscopy after staining of spermatozoa with trypan blue solution. The results are presented as mean values (% of total spermatozoa) ±SEM (n=6). The asterisk (*) indicates statistically significant different values from the t0 or the t60-Melatonin group (p≤0.01).
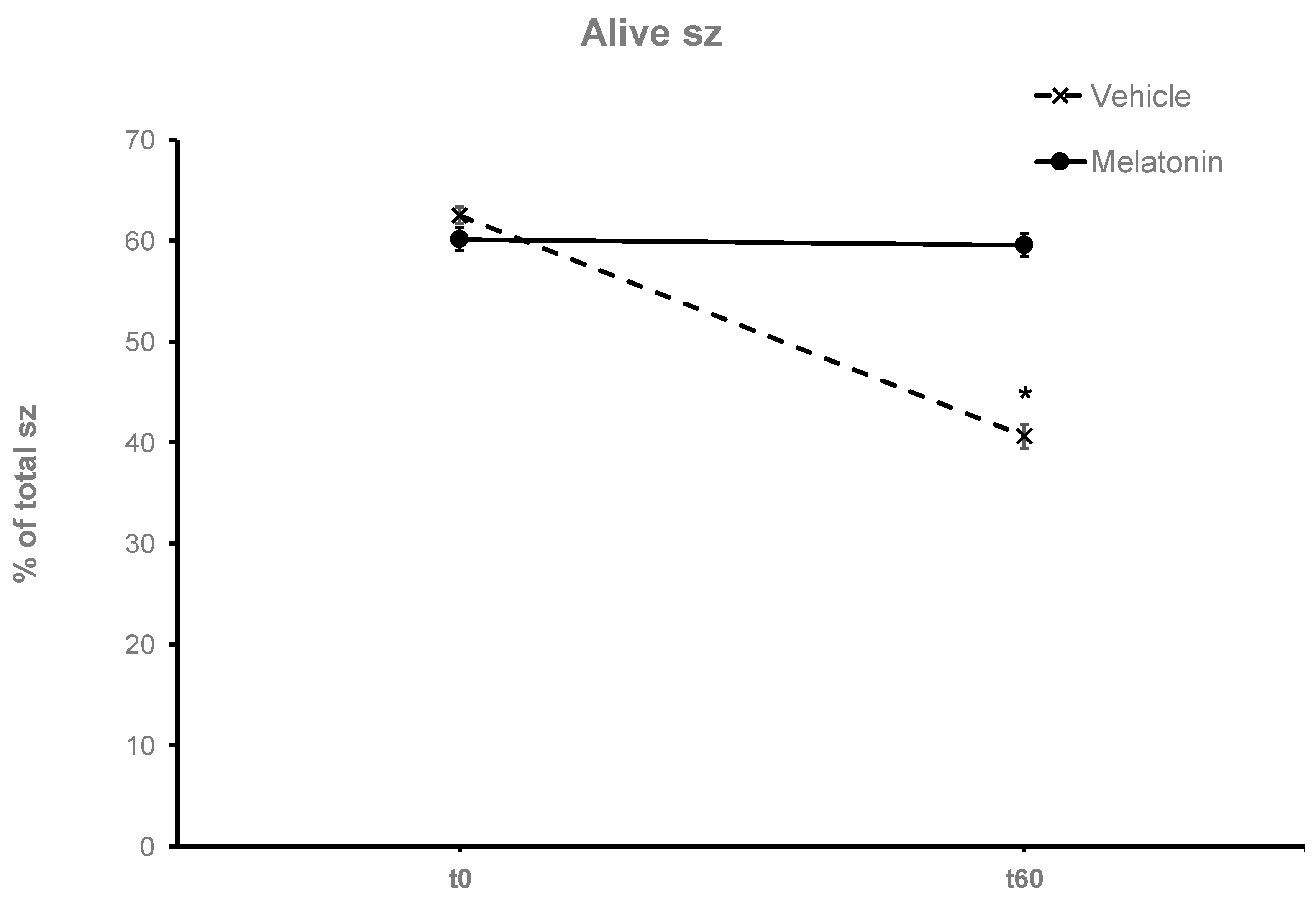
Figure 4.
The effect of melatonin on the viability of spermatozoa after exposure to 100 μΜ hydrogen peroxide. Cells were incubated for 60 min in the presence (black bars) or absence (white bars) of 100 μΜ melatonin, at which point the indicated samples were exposed to 100 μΜ H2O2 (HP), washed and stained with trypan blue solution. Spermatozoa were evaluated as alive (unstained) or dead (stained) cells by observation on an optical microscope. Results are presented as mean values ± SEM (n=6). The asterisk indicates statistically significant difference compared to the Control group or HP group. (P<0.01).
Figure 4.
The effect of melatonin on the viability of spermatozoa after exposure to 100 μΜ hydrogen peroxide. Cells were incubated for 60 min in the presence (black bars) or absence (white bars) of 100 μΜ melatonin, at which point the indicated samples were exposed to 100 μΜ H2O2 (HP), washed and stained with trypan blue solution. Spermatozoa were evaluated as alive (unstained) or dead (stained) cells by observation on an optical microscope. Results are presented as mean values ± SEM (n=6). The asterisk indicates statistically significant difference compared to the Control group or HP group. (P<0.01).
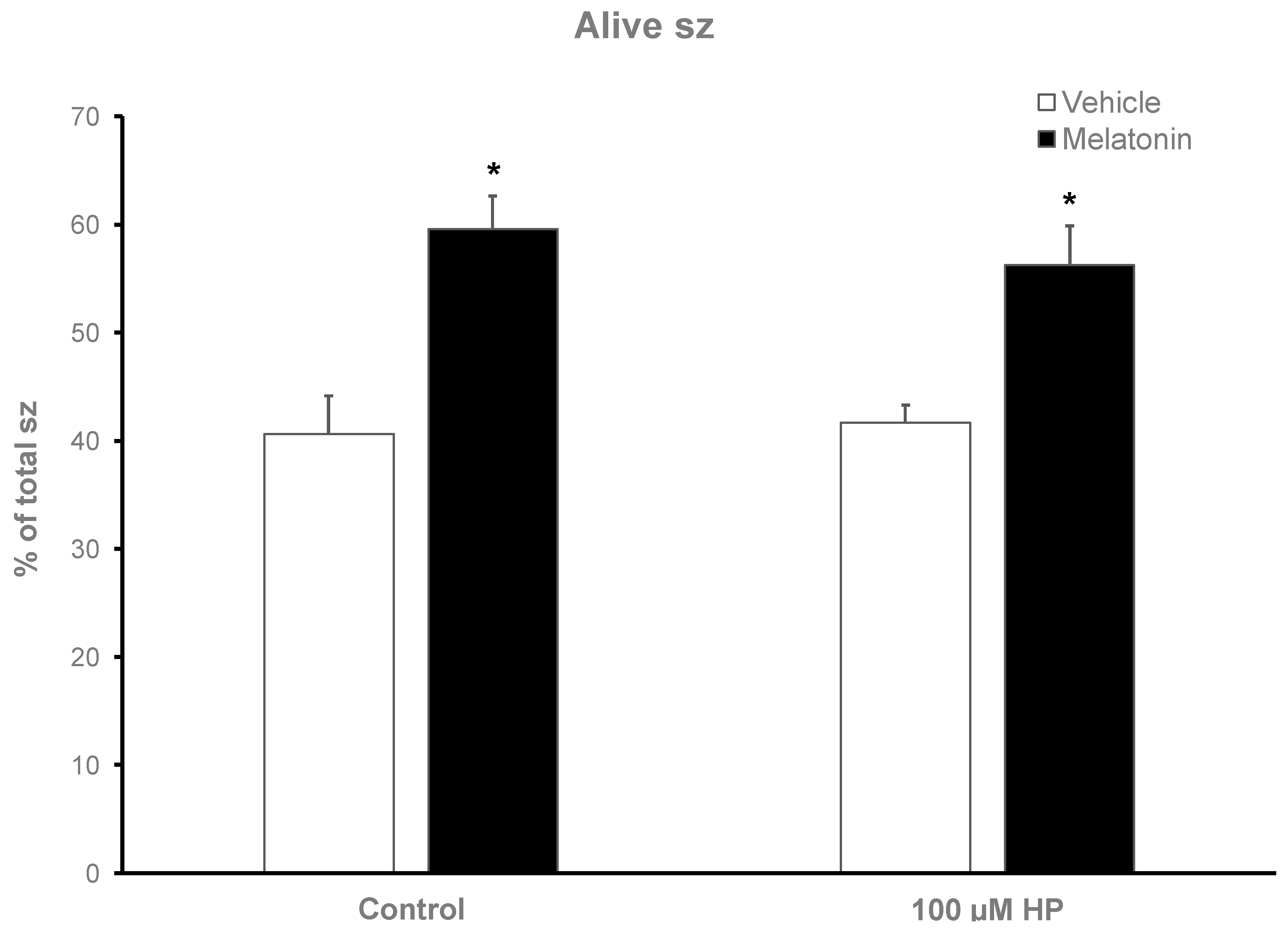
Figure 5.
The effect of melatonin on intracellular GSH levels after exposure to 100 μΜ H2O2. Frozen-thawed bovine spermatozoa were incubated in the presence (black bars) or absence (white bars) of 100 μΜ melatonin for 60 minutes, at which point the indicated samples were exposed to 100 μΜ H2O2.(HP). Results are presented as mean values ± SEM (n=6) and the asterisk indicates statistically significant difference compared to the Control group (p< 0.05).
Figure 5.
The effect of melatonin on intracellular GSH levels after exposure to 100 μΜ H2O2. Frozen-thawed bovine spermatozoa were incubated in the presence (black bars) or absence (white bars) of 100 μΜ melatonin for 60 minutes, at which point the indicated samples were exposed to 100 μΜ H2O2.(HP). Results are presented as mean values ± SEM (n=6) and the asterisk indicates statistically significant difference compared to the Control group (p< 0.05).
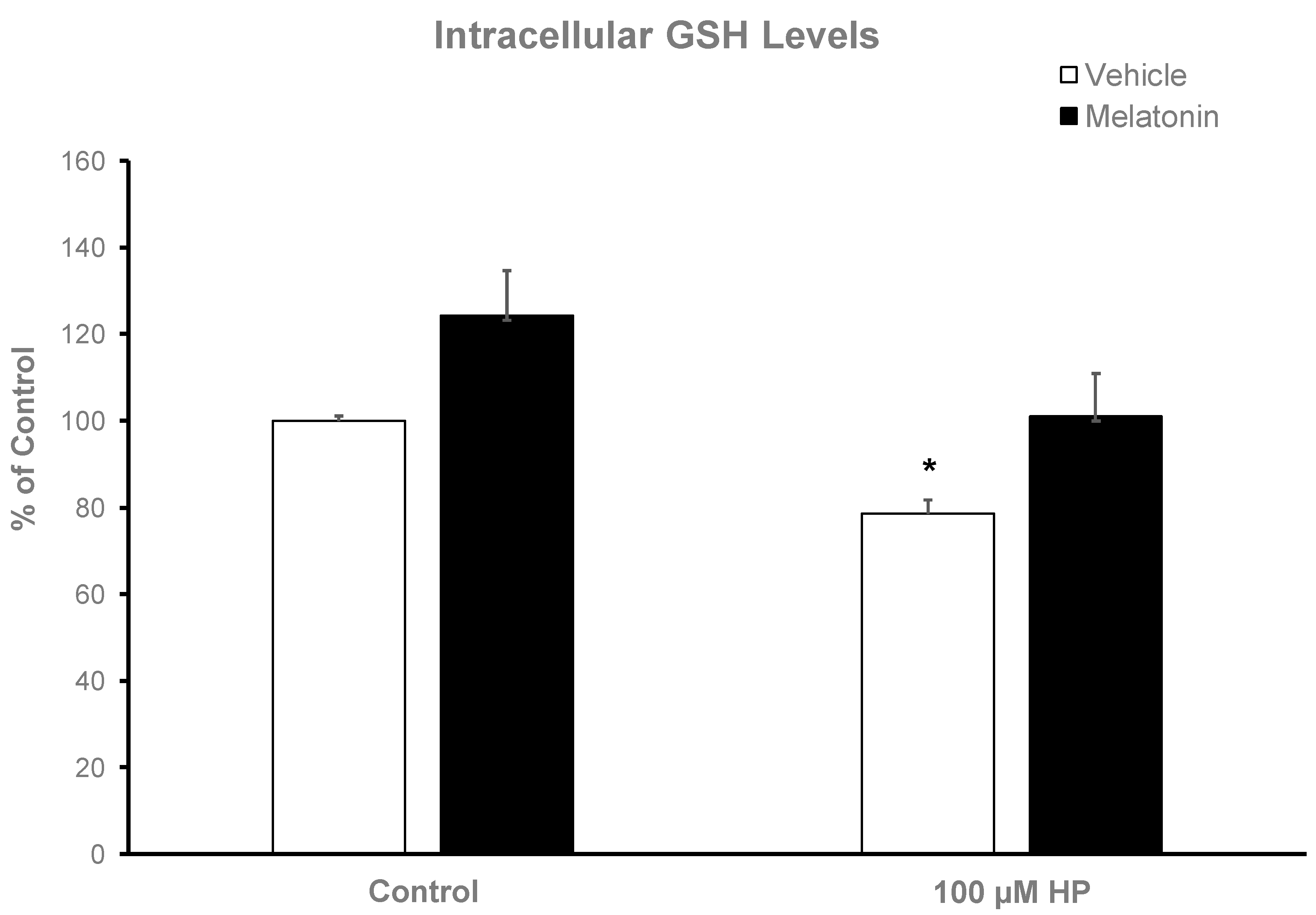
Figure 6.
Cellular O2− levels in bovine spermatozoa incubated in the presence (black bars) or absence (white bars) of 100 μΜ melatonin for 60 minutes, at which point 100 μΜ hydrogen peroxide (HP) was added to the respective groups. Cells were washed and O2− levels were determined by the NBT assay. Data are presented as mean ± SEM (n=6).
Figure 6.
Cellular O2− levels in bovine spermatozoa incubated in the presence (black bars) or absence (white bars) of 100 μΜ melatonin for 60 minutes, at which point 100 μΜ hydrogen peroxide (HP) was added to the respective groups. Cells were washed and O2− levels were determined by the NBT assay. Data are presented as mean ± SEM (n=6).
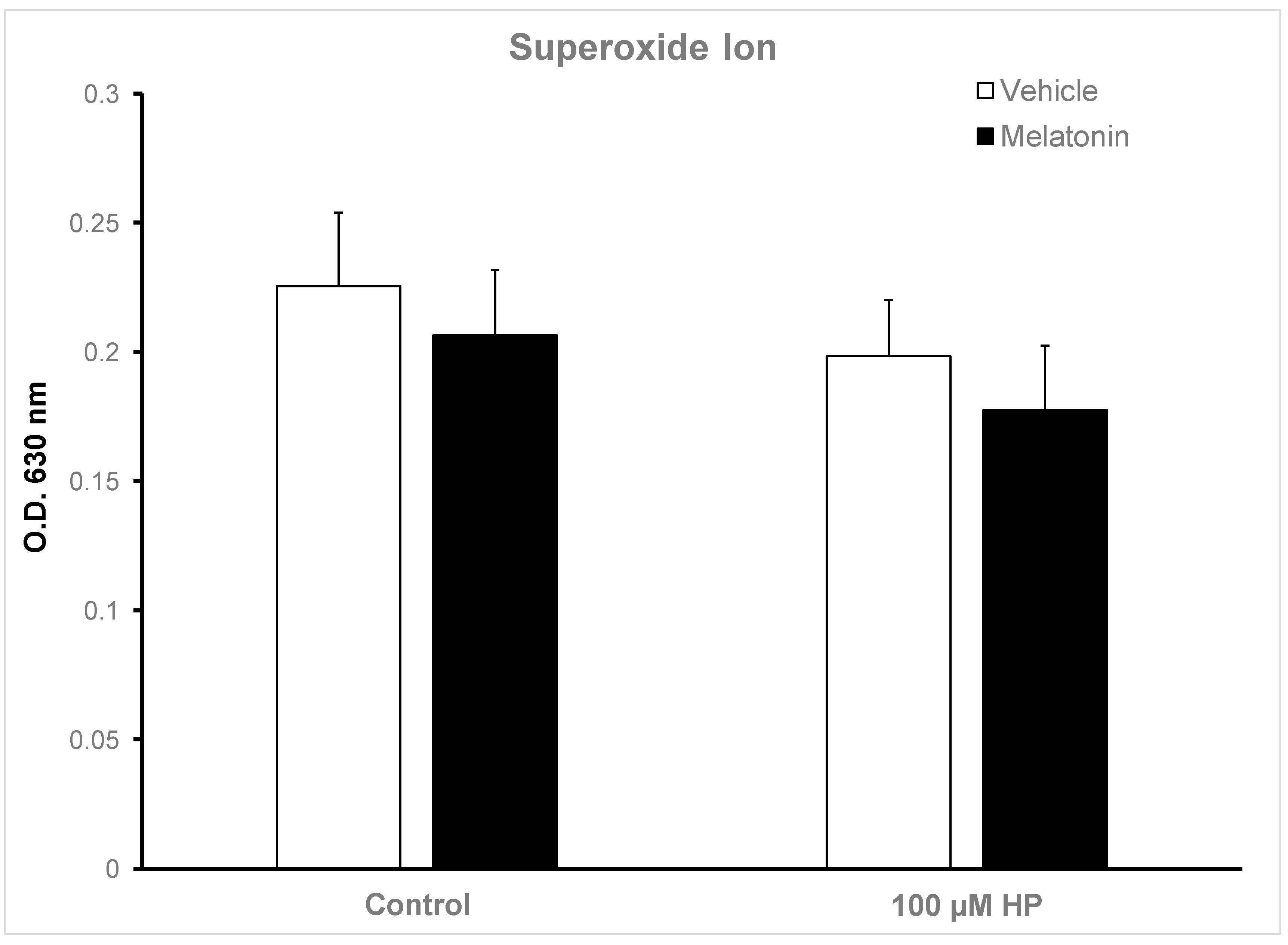
Figure 7.
A. Protein levels of iNOS in bovine spermatozoa incubated for 60 min with (black bars) or without (white bars) 100 μΜ melatonin and exposed to 100 μΜ H2O2 (HP), as determined by Western blot analysis. Results are expressed as percentage of the Control. Each bar represents mean values +/-SEM (n=4). The asterisk (*) indicates statistical significance (p<0.05). B. Representative membrane after blotting with the anti-iNOS antibody. MWM: molecular weight markers, C: Control, HP: 100 μM H2O2, M-HP: 100 μM melatonin + 100 μM H2O2, M: 100 μM melatonin.
Figure 7.
A. Protein levels of iNOS in bovine spermatozoa incubated for 60 min with (black bars) or without (white bars) 100 μΜ melatonin and exposed to 100 μΜ H2O2 (HP), as determined by Western blot analysis. Results are expressed as percentage of the Control. Each bar represents mean values +/-SEM (n=4). The asterisk (*) indicates statistical significance (p<0.05). B. Representative membrane after blotting with the anti-iNOS antibody. MWM: molecular weight markers, C: Control, HP: 100 μM H2O2, M-HP: 100 μM melatonin + 100 μM H2O2, M: 100 μM melatonin.
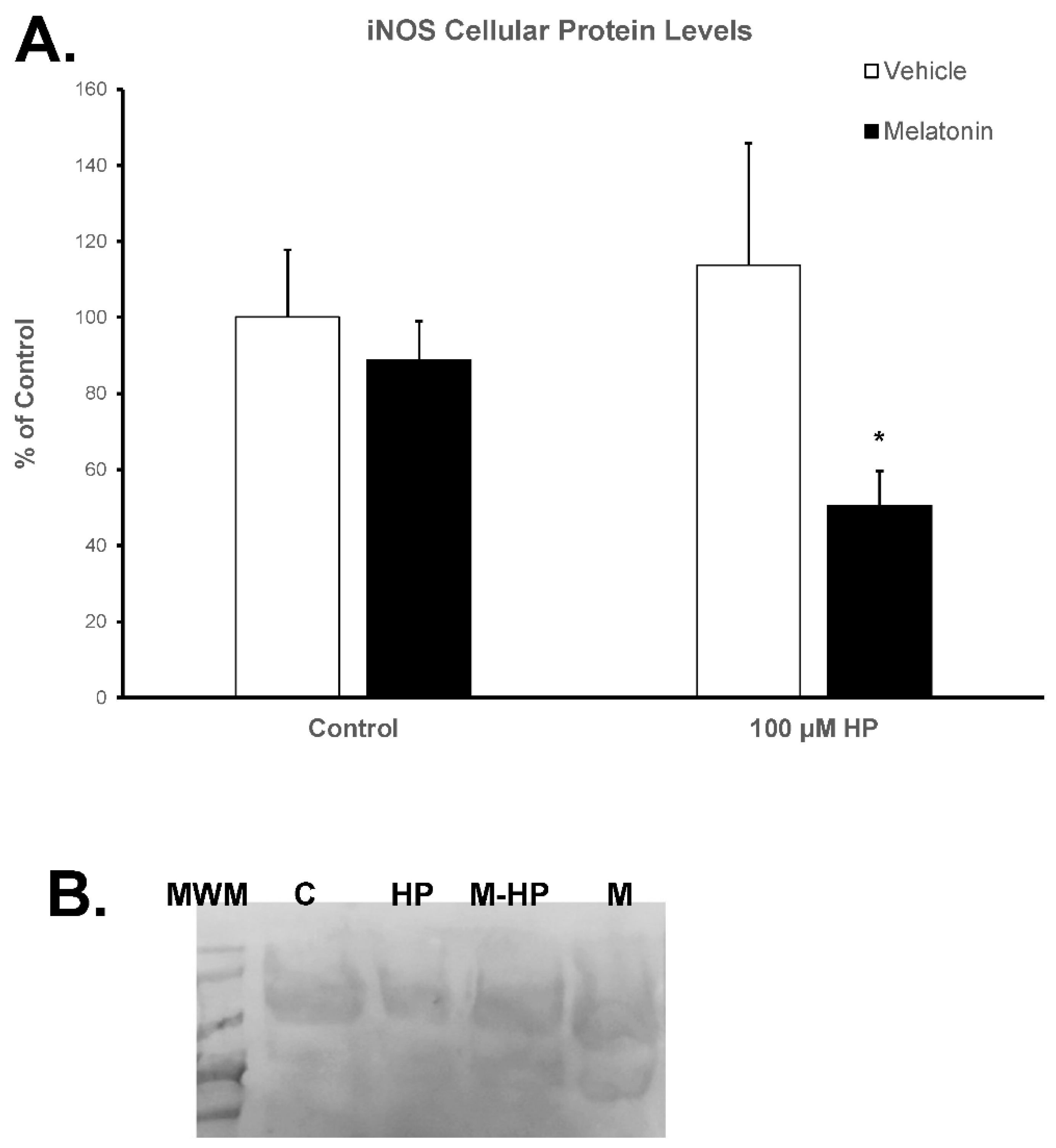

Table 1.
Kinematic parameters of spermatozoa as determined by CASA at the beginning (t0) and after 60 min of incubation at 37°C (t60) in the presence or absence of 100 μM melatonin and 100 μM H2O2. The data are presented as mean ± SEM (n=6). Different letters indicate statistically different values (p<0.05). The * indicates statistically different values from the Control t60-HP.
Table 1.
Kinematic parameters of spermatozoa as determined by CASA at the beginning (t0) and after 60 min of incubation at 37°C (t60) in the presence or absence of 100 μM melatonin and 100 μM H2O2. The data are presented as mean ± SEM (n=6). Different letters indicate statistically different values (p<0.05). The * indicates statistically different values from the Control t60-HP.
| Group | Total motile (%) | Static (%) | Rapid (%) |
Medium (%) | Slow (%) |
PM | VCL | VSL | VAP | ALH |
|---|---|---|---|---|---|---|---|---|---|---|
| Control t0 | 77.05±1.99 | 22.95±1.99 | 49.57±1.52 | 22.38±1.14 | 5.12±1.17 | 21.16±1.74 | 60.89±3.17 | 19.43±1.19* | 34.52±1.87b,d,e | 3.26±0.16* |
| Melatonin t0 | 76.5±1.53 | 24.33±1.93 | 44.03±1.84 | 25.94±1.37 | 6.53±0.9 | 23.77±1.55 | 59.58±1.22 | 20.93±0.57* | 35.83±0.46b,d,e | 3.09±0.08 |
| Control t60 | 61.9±3.69a,b,d | 35.32±3.69 | 27.67±2.58a,b,c | 28.15±1.27 | 8.87±0.74 | 23.58±2.23 | 49.82±1.68a,b | 18.55±0.96 | 30.47±1.09b,c,e | 2.72±0.2 |
| Control t60-HP | 54.96±1.2a,b | 43.98±1.27 | 20.48±1.73a,b,c | 25.53±1.19 | 10.02±0.85 | 20.25±4.62 | 43.72±5.6a,b,c | 17.83±1.89 | 23.85±4.17a,b,c | 2.93±0.05 |
| Melatonin t60 | 72.68±3.42b,c | 23.98±4 | 43.22±4.13 | 23.58±1.79 | 5.88±0.63 | 24.3±4.38 | 62.15±1.84 | 21.55±1.95* | 36.67±1.49d, | 3.38±0.18* |
| Melatonin t60-HP | 70.32±4.51a,c,d | 34.43±3.36 | 32.2±3.39a,b | 25.77±2.29 | 9.27±1.46 | 19.12±2.81 | 54.38±4.31b,c | 18.4±1.08 | 32±1.93b,c,d | 3.18±0.19 |
VCL: Curvilinear Velocity, VSL: Straight Line Velocity, VAP: Average Path Velocity, ALH: Amplitude Lateral Head movement.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



