
Submitted:
29 May 2023
Posted:
30 May 2023
You are already at the latest version
Abstract
Glutathione (GSH) and melatonin (MLT) are two antioxidants that have been described as having a protective role against oxidative stress in several animal species, improving sperm quality and survival after cryopreservation. Since there is no knowledge on the simultaneous use of these two substances concerning its effects on ram semen cryopreservation, this study aimed to evaluate the effects of MLT, GSH, or their association, on in vitro parameters of ram semen after thawing. During eight weekly sessions, the semen from three Merino rams was pooled, divided into four aliquots, and diluted with an egg yolk-based semen extender without (control) and with antioxidants (MLT 1mM, GSH 5mM, and MLT 1mM + GSH 5mM). Diluted semen was evaluated before and after freezing process. Supplementation of fresh semen with GSH at 5 mM had a deleterious effect on total motility progressive (TPM), curvilinear velocity (VCL), straight-line velocity (VSL), average-path velocity (VAP), linearity (LIN), straightness (STR) and increased slow spermatozoon (%). MLT at 1 mM also has a negative effect on TPM, VSL, and STR in fresh semen. In thawed semen, 1 mM of melatonin increased the TPM regarding the group control. Also, the combination of both antioxidants attenuates the negative effect of GSH 5mM on TPM, VSL and BCF. These results indicate that 5mM GSH impairs or does not improve sperm kinetic parameters in fresh semen and suggest that melatonin when combined with GSH plays a protective role on these effects.
Keywords:
Sperm cryopreservation
; semen quality
; antioxidants
; sperm motility
; rams
1. Introduction
The goal of semen cryopreservation is to safeguard semen quality, allowing its use in the future [1]. Current available cryopreservation methodology requires the preparation of semen extender and semen processing, and usually semen is stored in liquid nitrogen at −196 °C [2]. However, 40–50% loss in sperm viability after thawing occurs [3] and optimization protocols are required. Spermatozoa (SPZ) are exposed to many adverse conditions, such as ice crystal formation, thermal shock, chemical toxicity, osmotic and/or oxidative stress during the freezing and thawing procedures [4].
Oxidative stress is caused by the excessive formation of free radicals, particularly reactive oxygen species (ROS) [5]. At physiological levels, ROS are important for the maintenance of normal spermatozoa function; however, in situations of oxidative stress, high levels of these compounds compromise sperm viability, causing morphological and functional damage through the oxidation of lipids, proteins, sugars, and DNA present in these cells [5,6,7]. It is believed that hydrogen peroxide (H2O2) is the main ROS responsible for these damages at the cellular level, which results in decreased motility and viability of SPZ, as well as changes that hinder sperm fusion with the oocyte at fertilisation [7].
In order to protect spermatozoa from oxidative stress, there is an intrinsic antioxidant system, present in the seminal plasma [8] and sperm membranes [9]. This system contains both enzymatic and non-enzymatic antioxidants that eliminate free radicals [8]. Enzymatic antioxidants include the enzymes catalase, superoxide dismutase, glutathione reductase, and glutathione peroxidase [10]. The latter is considered to be the main responsible for eliminating the H2O2 [7]. The non-enzymatic antioxidants include substances such as urate, ascorbic acid, vitamin E, taurine, and hypotaurine, carotenoids, ubiquinones, and reduced glutathione (GSH) [10]. The effectiveness of this system is compromised by the cryopreservation process, since it leads to changes in the activity and distribution of the enzymes that compose it [9]. To overcome this condition, the addition of antioxidant substances to the freezing semen extenders seems to be a beneficial strategy for long-term storage of semen to reduce the negative effects of ROS [11,12,13].
Reduced glutathione (GSH) is part of the largest category of antioxidants present in the body, the thiols [14]. It is the main endogenous non-enzymatic antioxidant, and is widely distributed in mammalian cells [15,16]. It plays an important role in defending cells against oxidative stress; besides being able to react with ROS, it is also used by glutathione peroxidase to reduce H2O2 to H20 and lipoperoxides to alkyl alcohols [12,14]. These defence functions are also present in sperm and seminal plasma [17]. In this regard, several studies have described the protective role of GSH during storage and sperm cryopreservation, both in humans and horses, cats, pigs, cattle, goats, and sheep [18].
Endogenous melatonin (MLT), which participates in the control of the circadian rhythm and the regulation of reproductive seasons in seasonal species, also has anti-inflammatory and antioxidant properties [19]. Several studies, in different species, such as humans [20], bulls [21], chickens [22], rams [11], rabbits [23], dogs [24] and rats [25] have reported that the addition of melatonin as an antioxidant to the extenders used for semen cryopreservation, has a protective effect against oxidative stress, improving sperm quality, survival, and fertility [5]. This protective effect results from its direct action in eliminating free radicals, and from its indirect action in stimulating the activity and regulation of the synthesis of antioxidant enzymes such as glutathione peroxidase, glutathione reductase, catalase, and superoxide dismutase [19,26,27]. Melatonin’s direct action on ROS during cryopreservation safeguards membrane integrity and improves mitochondrial function; increases endogenous antioxidant defences, through its influence on enzyme activity; inhibits apoptosis and protects sperm DNA [28].
To our best knowledge, there are no previous studies on the concomitant use of GSH and melatonin as antioxidant supplements added to extenders for ram sperm cryopreservation. Considering that both substances participate in the antioxidant system previously described, our goal is to evaluate whether the addition of these substances to the egg yolk-based semen extender will have a beneficial effect on in vitro parameters in thawed ram sperm.
2. Materials and Methods
2.1. Local and Animals
The experimental work took place at the Estação Zootécnica Nacional (EZN), at Fonte Boa farm (39°11′57.008″ N 8°44′22.495″ W) during the breeding season from 18th October to 6th December 2022. These trial used semen samples from three Portuguese Merinos rams housed on the EZN facilities, in the same management conditions reported by Barbas et al. [29].
The animals were aged between 2 and 3 years and were kept confined, with water and hay ad libitum, and provided with 1 kg of commercial concentrate per day to keep a body condition score of 3.25 (5-points scale) [30]. Rams were previously dewormed and vaccinated against clostridial and pasteurellosis diseases. The flock was classified as brucellosis-free.
2.2. Semen extender, antioxidants supplementation and experimental design
Semen extender was formulated in the EZN laboratory according to Fernandes et al. [31], and consisted of 15% of EY (egg yolk), 6% of glycerol, 2.1805 g of TRIS, 0.3 g of glucose, 1.1194 g of citric acid, 0.005 g of penicillin and 38 ml of ultrapure water (Milli Q). The EY used were obtained from daily fresh chicken eggs and were manually separated from the egg white using filter paper and a sterile syringe to remove the chalaza.
GSH and melatonin USP powders, purchased from Sigma-Aldrich, Co. (Sintra, Portugal), were weighed and added to the previously formulated semen extender in the corresponding quantities to obtain concentrations of 5 mMol of GSH and 1 mMol of Melatonin. Four groups were made to assess in vitro sperm parameters: G1—semen extender without antioxidant (control group); G2—supplementation of melatonin (1 mMol), G3—supplementation of GSH (5 mMol), and G4—supplementation of GSH (5 mMol) + melatonin (1 mMol). After supplementing the antioxidant(s) in the respective groups, the tubes were shaken using a vortex shaker and then kept in a water bath at 30 °C until semen dilution.
The three Merino rams were used, and 1 ejaculate per ram was collected at each session. Eight semen collection sessions were carried out, totalizing 24 ejaculates. The sessions were carried out weekly on the 18th and 25th of October, the 3rd to 29th of November, and on the 6th of December.
2.3. Semen collection and processing
The ejaculates were collected by the artificial vagina method. After collection, they were immediately sent to the laboratory and placed in a 30 °C water bath and processed according to Fernandes et al. [31]. Each collected ejaculate was individually evaluated and if they fulfilled the minimum quality requirements, i.e., volume > 0.4 mL and > 55% of individual motility (IM), the ejaculates from the three rams were mixed to form a pool of each collection session. When these requisites were not fulfilled, they were rejected, and new semen collection was repeated from the respective rams. The evaluation of the IM (%) of the ejaculates was performed under a microscope (200 x) with platinum heated at 38 °C.
After the formation of the pool, total volume was determined by reading the graduated collecting tube and sperm concentration were determined by spectrophotometric analysis with the equipment (WPA-S106) calibrated for the sheep species. Previously, we diluted (1:400) ram pool, placing in a plastic cuvette 10 μl of pure semen and 3990 μl of bi-distilled water. Sperm concentration was expressed in x 106 total sperm/mL. Total sperm (SPZ) in the ejaculate was counted by the product of volume and concentration. Next, semen pool was divided into four aliquots of equal volume and the corresponding semen extender was added to each of them according to the experimental design. In all groups, the semen was diluted to obtain a concentration of 800 million SPZ/mL. Afterwards, fresh diluted semen of the four groups was immediately evaluated concerning subjective IM (%), sperm motility kinetics by CASA, and smears of the different aliquots were performed with eosin/nigrosine dyes.
In the next step, semen from each of the groups was aspirated separately into 0.25 mL straws by negative pressure and sealed with polyvinyl alcohol (PVA). A colour code was used to identify the four study groups, recording the colour of the straw and the PVA used in each group. They were placed in a water bath at 28 °C in a dry beaker and cooled to 4 °C in a refrigerated chamber during 4 hours. Then, the straws were placed horizontally in a floating freezing rack® (Minitübe GmbH, Tiefenbach, Germany), 4 cm above liquid nitrogen and frozen in liquid nitrogen vapours (−120 °C) during 20 minutes. After this time, the straws were submerged in liquid nitrogen (−196 °C) and stored in liquid nitrogen containers.
For thawing, the straws were immersed in a glass beaker with water at 38 °C for 1 minute. After drying with absorbent paper, the straws were placed in a glass tube containing 1 ml of isotonic saline solution at 38 °C and both tips of the straws were cut to empty their contents. Subsequently, these were briefly homogenized using a vortex (3 seconds), and seminal evaluation was initiated two minutes later. For each study group, one-two semen straws were thawed.
2.4. Seminal assessment
Subjectively, individual motility, viability, and sperm morphology parameters were determined [29].
For the determination of subjective individual motility (IM; %) semen was previously diluted at 1/100 dilution (10 μL of semen in 1 ml isotonic saline solution at 30 °C). Subsequently 10 μl of semen was placed in a glass plate covered with a coverslip and observed under a phase contrast microscope with platinum heated at 37 °C using a magnification of 200x. The IM value (%) of the ejaculate was estimated by the % of SPZ with good quality of movements (progressive, rectilinear, and uniform) observed in several microscopic fields.
By performing an eosin-nigrosine smear, it was possible to determine sperm viability (% live SPZ) and sperm morphology. For this, we used an optic microscope (Olympus BX40 microscopic®, Tokyo, Japan) selecting the immersion objective with a 1000x magnification and counting at least 100 SPZ to determine sperm viability (% of live SPZ). The live SPZ had a pink or white colouration, since the dye cannot cross the plasma membrane. In dead SPZ, which have alterations in plasma membrane allow dyes cross it and sperm cells turn to a purple colour. In the same observation, the % of normal and abnormal SPZ was accounted and its origin namely head, midpiece, and tail.
The CASA system (Computer Assisted Semen Analysis; ISASv1®, Valencia, Spain) was used to obtain semen motility parameters, namely kinetics and sperm trajectory movement, using standard settings for sheep species, namely head size (15–70 µm2), and progressive motility (80% of rectilinearity (STR)). For this purpose, a drop of 10 μL of diluted semen was put on a glass slide, covered with a coverslip and over the heated platinum (37 °C) of the microscope, the following motility parameters were estimated: total motility (TM; %), total progressive motility (TPM; %), total static (TS; %), motility subpopulations (slow, medium and fast; %), curvilinear velocity (VCL; μm/s), linear velocity (VSL; μm/s), mean path velocity (VAP; μm/s), linearity (LIN = [VSL/VCL] x 100; %), amplitude of lateral head displacement (ALH; μm), straightness (STR = [VSL/VAP] x 100; %), head oscillation (WOB = [VAP/VCL] x 100; %) and the cross frequency of the SPZ tail beat (BCF; Hz). The results are expressed as the average obtained by video capture (50 images/sec) of 5 microscopic fields with about 200 SPZ per field for each group under study at each session.
All parameters evaluated were determined in fresh semen and thawed semen of the four groups.
2.5. Statistical analysis
After Shapiro–Wilk W test evaluation, an arcsine square root transformation (Sqrt) of SPZ data was made to reach or approach normal distribution.
A multivariable mixed linear model for repeated measures, using the restricted maximum likelihood (REML) method, was built for each SPZ parameter according to the following equation:
where,
Yijm = Hi + Lj + (HxL)ij + tmi + eijm
Yijm is a vector of all observations and represented by the least square value;
Hi is the fixed effect for group (4 levels: G1, G2, G3, and G4);
Lj is the fixed effect for processing semen (2 levels: fresh sperm, thawed sperm);
(HxL)ij, is the two-way interaction;
Tmi is the random effect for session (m); and,
eijm is a vector of residuals.
Differences of fixed factors and estimate variance component of the mixed factor were evaluated by using the Tukey test and Wald, respectively.
The software JMP® 16 for Windows (SAS Institute, Cary, NC, USA) was used to build the models. All the results were presented as least squares means ± (Sqrt)SEM for a 0.05 level of significance.
3. Results
According to Table 1, all seminal parameters were negatively affected by the cryopreservation process except for intermediate piece (IP) defects and straightness (STR). Regarding CASA sperm evaluation, an effect of the group (G) was observed for TS, TPM, Slow, Medium, VCL, VSL, VAP, WOB and BCF semen parameters. Interactions semen processing x group (SP x G) were observed for TPM, Medium, VCL, VSL, VAP, LIN and STR semen parameters.
Regarding subjective evaluation of SPZ parameters, only sperm viability (death and alive SPZ) and sperm IM were consistently affected by the freezing-thawing cycle throughout groups (Table 2). Overall, abnormal SPZ remains similar between groups and freezing-thawing cycle, according to the Tukey test. Only a decrease of tail defects % and an increase of head defects % after thawing was observed in G3 and G4, respectively.
A great impact of semen processing was observed in TS, from 2.7% in fresh semen to 38.9% in thawed semen, without differences between groups (Table 3). Inversely, TM decreased from 95.4% (fresh semen) to 58.2% (thawed semen), and no group effect was observed. In fresh semen, the supplementation of 5 mM GSH (G3) had a deleterious effect in several SPZ kinetics parameters (TPM, Slow, VCL, VSL, VAP, LIN, and STR) relatively to control group (G1). The same was observed in G2 for three SPZ parameters (TPM, VSL, and STR) relatively to control group. In G4, this adverse effect was observed for TPM, VSL, LIN and STR compared to the control group.
In frozen semen, highest values of TPM were observed in G2 with no differences among the other groups. VSL was lower in G3 relatively to G2 group. No other differences were detected among groups for the remaining parameters. All the results are reported in Table 3.
Groups x semen processing interactions of TPM, VSL, STR, VAP, and LIN are reported in Figure 1.
4. Discussion
Several studies in different species have been conducted to prevent oxidative stress during semen storage in both chilled and frozen semen by using antioxidants such as melatonin and GSH, with contradictory results. Some authors reported that addition of melatonin [11,32] and GSH [12,17,18] improves semen quality and motility, while others didn’t noticed these effects [33,34].
In the present study there was a highly significant decrease in IM, live sperm and a significant increase in TS (CASA) in frozen semen. Also, in frozen semen, we observed that isolated addition of melatonin at 1 mM (G2) and GSH at 5 mM (G3), or together (G4) did not improved SPZ parameters being 40% and 60% for IM and TM (CASA), respectively. During cryopreservation several cellular structures are damaged, including plasma membrane, acrosome, axoneme, and mitochondria [35,36]. This damage will influence motility, which is dependent on available energy that is produced via oxidative phosphorylation in the mitochondria, and glycolysis in the fibrous sheath [37]. Thus, if during cryopreservation these structures are damaged, the capacity for ATP production is reduced, and sperm motility is impaired [38].
Sperm motility is one of the most important characteristics used in the evaluation of semen quality, since it is fundamental that spermatozoa can migrate through the female reproductive tract and reach the oocyte for fertilization [38]. Studies indicate that the addition of 100 and 200 mM GSH to fresh semen improves SPZ kinetic parameters during refrigerated storage at 5 °C in rams [18]. The same was found for frozen semen of goat [12] and boar [39] when 1 and 5 mM GSH were added. In our study, the addition of 5 mM GSH to fresh semen did not improve sperm motility. On the contrary, on fresh semen, the kinetic parameters TPM, VCL, VSL, VAP, LIN, and STR were negatively affected (p < 0.05) in comparison to control group. Our results are contradictory to Shi et al. [18] who found no differences in motility parameters of ram semen immediately after GSH addition (at 0 h). However, at 72 h of storage the groups with 100 and 200 mM of GSH increased TPM, VCL, VSL, and VAP. Nevertheless, Ansari et al. [40], Salmani et al. [34] and Banday et al. [33] reported that the addition of GSH at concentrations of 3, 5, and 10 mM had no effect or was detrimental to sperm quality parameters. These authors refer that the addition of 3 mM did not affect the progressive motility of buffalo semen [40], nor that 5 mM affected the motility and viability of ram semen [33]. Also, using 5 mM GSH in goat semen did not influence total motility and progressive motility, however, when a concentration of 10 mM was used, there was an adverse effect on the parameters of TM, VAP, and VCL [34]. Considering that sperm motility patterns can be altered by the physicochemical properties of the extender, the addition of GSH before cryopreservation may have altered its inherent characteristics, impairing motility parameters [40]. The adverse effects may be associated with the negative effect caused by osmotic stress due to excessive GSH concentrations which was observed in ram semen [41] and equine semen [42]. In our study, osmolarity was not controlled, so negative results may have been assigned to added (5 mM) concentrations of GSH. On the other hand, the detrimental effects on motility and other characteristics of sperm kinetics may also be a consequence of mitochondrial and axoneme damage of spermatozoa induced by high concentrations of GSH [34].
In frozen semen, except for TPM, there were no differences (p > 0.05) between control and the other groups. The addition of 1 mM melatonin to the extender increased the TPM in the frozen semen. This finding agrees with the results obtained in rams by Succu et al. [43] and Ashrafi et al. [44]. Also, Fadl et al. [23] studied the effect of melatonin addition on the quality and DNA integrity of frozen rabbit semen and found that supplementation with 1 mM melatonin significantly increased TM and TPM compared to the control group. In addition, Shahandeh et al. [25] also demonstrated that the addition of 1 mM melatonin to the semen extender not only increased TM, but corresponded to the group with the highest value of progressive motility in frozen rat semen.
In the remaining parameters, no differences were recorded between groups. This corroborates what was reported by ChaithraShree et al. [32], who studied the melatonin effects on sperm motility and structural changes in bovine SPZ after 48 h cryopreservation and found that its addition had no significant effects on motility parameters, which was also verified in rat semen [25]. It should be noted that results comparison with other authors becomes difficult, since most studies do not describe CASA system configurations applied. This fact is essential, since semen parameters estimated by CASA are conditionated by the selected configurations, as well as by the type of slide/camera used, semen processing method and the operator, because he choose the microscopic fields to be analysed [45].
Luo et al. [46] investigated the effects of MLT, GSH, Vit. E and their combinations on Mediterranean buffalo semen cryopreservation. In this study, the single 0.2 mM GSH or combined 1 mM MLT + 0.1 mM GSH supplementation significantly increased the viability and kinetic parameters of spermatozoa compared to the control group. In our study, the antioxidants association had no beneficial effect compared to the control group. These contradictory results may be assigned by the differences among species, semen extender, semen processing, cryopreservation techniques, as well as by the joined concentrations of antioxidants and their associations. Even though they are similar for MLT, yet GSH concentration was 25 (0.2 mM) to 50 (mM) times lower than in the present study. Interestingly, in our study, the addition of MLT to GSH (G4) revealed a protective effect (p < 0.05) because a positive effect was observed in TPM, VSL, and BCF parameters relatively to GSH group. Additionally, the group x freezing interactions confirm these observations. This effect was similar to the one reported by Alagbonsi and Olayaki [47] who investigated the melatonin effects on Δ9-tetrahydrocannabinol (THC)-induced reduction in rat sperm motility. They found that, like in our study, melatonin alone increased progressive motility and sperm kinetic parameters. However, when combined with THC, melatonin attenuated the reduction in progressive motility (by 42%) and kinetic parameters caused by THC.
As previously proposed, osmotic stress may be at the origin of the adverse effects observed in this study when using a high concentration of GSH (5 mM). Burnaugh et al. [48] found an interaction between osmotic stress and oxidative stress, noting that exposure to hypo- and hyperosmotic stress in equine semen can increase the synthesis of superoxide anions by viable spermatozoa and consequently increase ROS formation, amplifying oxidative stress. Melatonin protects spermatozoa against oxidative stress, not only by eliminating excess ROS but also by stimulating endogenous antioxidant enzymes [28]. Several studies have described the strong antioxidant capacity of melatonin. Luo et al. [46] suggest that 1 mM MLT maintains the antioxidant capacity of thawed Mediterranean buffalo sperm. Bhalothia et al. [49] showed that the addition of 1 mM MLT to ram semen extender protects spermatozoa by preventing free radical formation during storage at 4 °C. These facts may thus explain the protective role of melatonin observed when combined with damaging GSH concentrations.
5. Conclusions
We conclude that GSH, at doses considered beneficial in the literature (5 mM), impaired or did not improve some kinetic parameters in fresh semen, although they were similar to the control group in thawed semen. It is further suggested that MLT had a protective role in these effects. Prospective studies should be conducted to optimise the doses and associations of these antioxidants, as well as to establish the mechanisms by which they act to improve ram semen freezability.
Author Contributions
Conceptualization and methodology, C.C., J.S. and J.P.B. Formal analysis, data curation, original draft preparation, and writing—review and editing were equally performed by C.C., J.P.., J.S. and J.P.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by projects UIDP/CVT/00772/2020 (CECAV) and LA/P/0059/2020 (AL4AnimalS), funded by the Portuguese Foundation for Science and Technology (FCT).
Institutional Review Board Statement
This research was approved by the Institutional Review Board of INIAV protocol code 1/UEBRG/INIAV/2023, and it is in accordance with Portuguese and European legislation on animal welfare issues.
Data Availability Statement
The data present in this study are available on request from the corresponding author.
Acknowledgments
We thank EZN-INIAV (Santarem, Portugal) for providing the necessary facilities for the execution this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Allai, L.; Benmoula, A.; Marciane da Silva, M.; Nasser, B.; El Amiri, B. Supplementation of Ram Semen Extender to Improve Seminal Quality and Fertility Rate. Anim. Reprod. Sci. 2018, 192, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Binyameen, M.; Saleem, M.; Riaz, A. Recent Trends in Bovine Reproductive Biotechnologies. CAB Rev. 2019, 14. [Google Scholar] [CrossRef]
- Watson, P.F. The Causes of Reduced Fertility with Cryopreserved Semen. Anim. Reprod. Sci. 2000, 60–61, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Wu, G.; Hong, Q.; Quan, G. Spermatozoa Cryopreservation: State of Art and Future in Small Ruminants. Biopreserv. Biobanking 2019, 17, 171–182. [Google Scholar] [CrossRef]
- Medrano, A.; Contreras, C.Bs.; Herrera, F.; Alcantar-Rodriguez, A. Melatonin as an Antioxidant Preserving Sperm from Domestic Animals. Asian Pac. J. Reprod. 2017, 6, 241–246. [Google Scholar] [CrossRef]
- Baskaran, S.; Finelli, R.; Agarwal, A.; Henkel, R. Reactive Oxygen Species in Male Reproduction: A Boon or a Bane? Andrologia 2021, 53, e13577. [Google Scholar] [CrossRef]
- Meyers, S.A. Cryostorage and Oxidative Stress in Mammalian Spermatozoa. In Studies on Men’s Health and Fertility; Agarwal, A., Aitken, R.J., Alvarez, J.G., Eds.; Oxidative Stress in Applied Basic Research and Clinical Practice; Humana Press: Totowa, NJ, 2012; pp. 41–56. ISBN 978-1-61779-775-0. [Google Scholar]
- Soren, S.; Singh, S.V.; Singh, P. Influence of Season on Seminal Antioxidant Enzymes in Karan Friesbulls under Tropical Climatic Conditions. Turk. J. Vet. Anim. Sci. 2016, 40, 797–802. [Google Scholar] [CrossRef]
- Marti, E.; Marti, J.I.; Muino-Blanco, T.; Cebrian-Perez, J.A. Effect of the Cryopreservation Process on the Activity and Immunolocalization of Antioxidant Enzymes in Ram Spermatozoa. J. Androl. 2008, 29, 459–467. [Google Scholar] [CrossRef]
- Gadea, J.; Sellés, E.; Marco, M.A.; Coy, P.; Matás, C.; Romar, R.; Ruiz, S. Decrease in Glutathione Content in Boar Sperm after Cryopreservation. Theriogenology 2004, 62, 690–701. [Google Scholar] [CrossRef]
- Dai, G.; Meng, Y.; Zhang, L.; Du, Y.; Wen, F.; Feng, T.; Hu, J. Effect of Addition of Melatonin on Liquid Storage of Ram Semen at 4 °C. Andrologia 2019, 51, e13236. [Google Scholar] [CrossRef]
- Gadea, J.; Gumbao, D.; Gómez-Giménez, B.; Gardón, J.C. Supplementation of the Thawing Medium with Reduced Glutathione Improves Function of Frozen-Thawed Goat Spermatozoa. Reprod. Biol. 2013, 13, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Pezo, F.; Yeste, M.; Zambrano, F.; Uribe, P.; Risopatrón, J.; Sánchez, R. Antioxidants and Their Effect on the Oxidative/Nitrosative Stress of Frozen-Thawed Boar Sperm. Cryobiology 2021, 98, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Bucak, M.N.; Ateşşahin, A.; Yüce, A. Effect of Anti-Oxidants and Oxidative Stress Parameters on Ram Semen after the Freeze–Thawing Process. Small Ruminant Research 2008, 75, 128–134. [Google Scholar] [CrossRef]
- Ali, S.S.; Ahsan, H.; Zia, M.K.; Siddiqui, T.; Khan, F.H. Understanding Oxidants and Antioxidants: Classical Team with New Players. J. Food Biochem. 2020, 44, e13145. [Google Scholar] [CrossRef] [PubMed]
- Gadea, J.; Molla, M.; Selles, E.; Marco, M.A.; Garcia-Vazquez, F.A.; Gardon, J.C. Reduced Glutathione Content in Human Sperm Is Decreased after Cryopreservation: Effect of the Addition of Reduced Glutathione to the Freezing and Thawing Extenders. Cryobiology 2011, 62, 40–46. [Google Scholar] [CrossRef]
- Zou, J.; Wei, L.; Li, D.; Zhang, Y.; Wang, G.; Zhang, L.; Cao, P.; Yang, S.; Li, G. Effect of Glutathione on Sperm Quality in Guanzhong Dairy Goat Sperm During Cryopreservation. Front. Vet. Sci. 2021, 8, 771440. [Google Scholar] [CrossRef]
- Shi, L.; Jin, T.; Hu, Y.; Ma, Z.; Niu, H.; Ren, Y. Effects of Reduced Glutathione on Ram Sperm Parameters, Antioxidant Status, Mitochondrial Activity and the Abundance of Hexose Transporters during Liquid Storage at 5 ℃. Small Ruminant Research 2020, 189, 106139. [Google Scholar] [CrossRef]
- Reiter, R.J.; Mayo, J.C.; Tan, D.-X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an Antioxidant: Under Promises but over Delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef]
- Minucci, S.; Venditti, M. New Insight on the In Vitro Effects of Melatonin in Preserving Human Sperm Quality. Int. J. Mol. Sci. 2022, 23, 5128. [Google Scholar] [CrossRef]
- Inyawilert, W.; Rungruangsak, J.; Liao, Y.; Tang, P.; Paungsukpaibool, V. Melatonin Supplementation Improved Cryopreserved Thai Swamp Buffalo Semen. Reprod. Domest. Anim. 2021, 56, 83–88. [Google Scholar] [CrossRef]
- Appiah, M.O.; He, B.; Lu, W.; Wang, J. Antioxidative Effect of Melatonin on Cryopreserved Chicken Semen. Cryobiology 2019, 89, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Fadl, A.M.; Ghallab, A.R.M.; Abou-Ahmed, M.M.; Moawad, A.R. Melatonin Can Improve Viability and Functional Integrity of Cooled and Frozen/Thawed Rabbit Spermatozoa. Reprod. Domest. Anim. 2021, 56, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Rodriguez, J.A.; Carbajal, F.J.; Martinez-De-Anda, R.; Alcantar-Rodriguez, A.; Medrano, A. Melatonin Added to Freezing Diluent Improves Canine (Bulldog) Sperm Cryosurvival. Reproduction and Fertility 2020, 1, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Shahandeh, E.; Ghorbani, M.; Mokhlesabadifarahani, T.; Bardestani, F. Melatonin and Selenium Supplementation in Extenders Improves the Post-Thaw Quality Parameters of Rat Sperm. Clin. Exp. Reprod. Med. 2022, 49, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Cipolla-Neto, J.; Amaral, F.G. Melatonin as a Hormone: New Physiological and Clinical Insights. Endocr. Rev. 2018, 39, 990–1028. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolin, I.; Herrera, F.; Martin, V.; Reiter, R.J. Regulation of Antioxidant Enzymes: A Significant Role for Melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef]
- Ofosu, J.; Qazi, I.H.; Fang, Y.; Zhou, G. Use of Melatonin in Sperm Cryopreservation of Farm Animals: A Brief Review. Anim. Reprod. Sci. 2021, 233, 106850. [Google Scholar] [CrossRef]
- Barbas, J.P.; Pimenta, J.; Baptista, M.C.; Marques, C.C.; Pereira, R.M.L.N.; Carolino, N.; Simões, J. Ram Semen Cryopreservation for Portuguese Native Breeds: Season and Breed Effects on Semen Quality Variation. Animals 2023, 13, 579. [Google Scholar] [CrossRef]
- Russel, A.J.F.; Doney, J.M.; Gunn, R.G. Subjective Assessment of Body Fat in Live Sheep. J. Agric. Sci. 1969, 72, 451–454. [Google Scholar] [CrossRef]
- Fernandes, M.; Hernández, P.R.; Simões, J.; Barbas, J.P. Effects of Three Semen Extenders, Breeding Season Month and Freezing–Thawing Cycle on Spermatozoa Preservation of Portuguese Merino Sheep. Animals 2021, 11, 2619. [Google Scholar] [CrossRef]
- ChaithraShree, A.R.; Ingole, S.D.; Dighe, V.D.; Nagvekar, A.S.; Bharucha, S.V.; Dagli, N.R.; Kekan, P.M.; Kharde, S.D. Effect of Melatonin on Bovine Sperm Characteristics and Ultrastructure Changes Following Cryopreservation. Vet. Med. Sci. 2020, 6, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Banday, M.N.; Lone, F.A.; Rasool, F.; Rashid, M.; Shikari, A. Use of Antioxidants Reduce Lipid Peroxidation and Improve Quality of Crossbred Ram Sperm during Its Cryopreservation. Cryobiology 2017, 74, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Salmani, H.; Nabi, M.M.; Vaseghi-Dodaran, H.; Rahman, M.B.; Mohammadi-Sangcheshmeh, A.; Shakeri, M.; Towhidi, A.; Shahneh, A.Z.; Zhandi, M. Effect of Glutathione in Soybean Lecithin-Based Semen Extender on Goat Semen Quality after Freeze-Thawing. Small Ruminant Research 2013, 112, 123–127. [Google Scholar] [CrossRef]
- Alcay, S.; Ustuner, B.; Nur, Z. Effects of Low Molecular Weight Cryoprotectants on the Post-Thaw Ram Sperm Quality and Fertilizing Ability. Small Ruminant Research 2016, 136, 59–64. [Google Scholar] [CrossRef]
- Keskin, N.; Erdogan, C.; Bucak, M.N.; Ozturk, A.E.; Bodu, M.; Ili, P.; Baspinar, N.; Dursun, S. Cryopreservation Effects on Ram Sperm Ultrastructure. Biopreserv. Biobanking 2020, 18, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A. Energy Metabolism in Mammalian Sperm Motility. WIREs Mech. Dis. 2022, 14, e1569. [Google Scholar] [CrossRef] [PubMed]
- Van De Hoek, M.; Rickard, J.P.; De Graaf, S.P. Motility Assessment of Ram Spermatozoa. Biology 2022, 11, 1715. [Google Scholar] [CrossRef]
- Giaretta, E.; Estrada, E.; Bucci, D.; Spinaci, M.; Rodríguez-Gil, J.E.; Yeste, M. Combining Reduced Glutathione and Ascorbic Acid Has Supplementary Beneficial Effects on Boar Sperm Cryotolerance. Theriogenology 2015, 83, 399–407. [Google Scholar] [CrossRef]
- Ansari, M.S.; Rakha, B.A.; Ullah, N.; Andrabi, S.M.H.; Iqbal, S.; Khalid, M.; Akhter, S. Effect of Exogenous Glutathione in Extender on the Freezability of Nili-Ravi Buffalo (Bubalus Bubalis) Bull Spermatozoa. Anim. Sci. Pap. Rep. 2010, 28, 235–244. [Google Scholar]
- Silva, S.; Soares, A.; Batista, A.; Almeida, F.; Nunes, J.; Peixoto, C.; Guerra, M. In Vitro and In Vivo Evaluation of Ram Sperm Frozen in Tris Egg-Yolk and Supplemented with Superoxide Dismutase and Reduced Glutathione: In Vitro and In Vivo Effect of the Addition of SOD and GSH. Reprod. Dom. Anim. 2011, 46, 874–881. [Google Scholar] [CrossRef]
- De Oliveira, R.A.; Wolf, C.A.; De Oliveira Viu, M.A.; Gambarini, M.L. Addition of Glutathione to an Extender for Frozen Equine Semen. Journal of Equine Veterinary Science 2013, 33, 1148–1152. [Google Scholar] [CrossRef]
- Succu, S.; Berlinguer, F.; Pasciu, V.; Satta, V.; Leoni, G.G.; Naitana, S. Melatonin Protects Ram Spermatozoa from Cryopreservation Injuries in a Dose-Dependent Manner: Melatonin Mitigates Cryopreservation Injuries of Ram Spermatozoa. J. Pineal Res. 2011, 50, 310–318. [Google Scholar] [CrossRef]
- Ashrafi, I.; Kohram, H.; Naijian, H.; Bahreini, M.; Poorhamdollah, M. Protective Effect of Melatonin on Sperm Motility Parameters on Liquid Storage of Ram Semen at 5 °C. Afr. J. Biotechnol. 2011, 10, 6670–6674. [Google Scholar]
- Peter, A.T.; Brito, L.; Althouse, G.; Aurich, C.; Chenoweth, P.; Fraser, N.; Lopate, C.; Love, C.; Luvoni, G.; Waberski, D. Andrology Laboratory Review: Evaluation of Sperm Motility. Clinical Theriogenology 2021, 13, 24–36. [Google Scholar] [CrossRef]
- Luo, X.; Wu, D.; Liang, M.; Huang, S.; Shi, D.; Li, X. The Effects of Melatonin, Glutathione and Vitamin E on Semen Cryopreservation of Mediterranean Buffalo. Theriogenology 2023, 197, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Alagbonsi, I.A.; Olayaki, L.A. Melatonin Attenuates Δ 9 -Tetrahydrocannabinol-Induced Reduction in Rat Sperm Motility and Kinematics in-Vitro. Reprod. Toxicol. 2018, 77, 62–69. [Google Scholar] [CrossRef]
- Burnaugh, L.; Ball, B.A.; Sabeur, K.; Thomas, A.D.; Meyers, S.A. Osmotic Stress Stimulates Generation of Superoxide Anion by Spermatozoa in Horses. Animal Reproduction Science 2010, 117, 249–260. [Google Scholar] [CrossRef]
- Bhalothia, S.K.; Mehta, J.S.; Kumar, T.; Prakash, C.; Talluri, T.R.; Pal, R.S.; Kumar, A. Melatonin and Canthaxanthin Enhances Sperm Viability and Protect Ram Spermatozoa from Oxidative Stress during Liquid Storage at 4 °C. Andrologia 2022, 54, e14304. [Google Scholar] [CrossRef]
Figure 1.
Interaction of group x semen processing on the parameters that showed the highest significance (p < 0.001 and p < 0.01), on the overall model. (a) In fresh semen, total progressive motility (TPM; %), was lower in the treated groups (MLT, GSH and GSH + MLT) when compared to the control group. In frozen semen, MLT group was higher than GSH and control group, however both were similar to GSH + MLT group. (b) In fresh semen the mean values of the straight-line velocity (VSL; µm/s) were lower in the treated groups (MLT, GSH and GSH + MLT) compared to the control group. In frozen semen the MLT group was superior to the GSH group, with no other differences among groups. (c) In fresh semen the straightness (STR; %) was lower in the treated groups (MLT, GSH and GSH + MLT) compared to the control group. In frozen semen there were no differences between the studied groups. (d) The mean average-path velocity (VAP; µm/s) in fresh semen was lower in the GSH group and in frozen semen no differences were detected among groups. (e) In fresh semen Linearity (LIN; %) was lower in the GSH group. In frozen semen there were no differences among groups.
Figure 1.
Interaction of group x semen processing on the parameters that showed the highest significance (p < 0.001 and p < 0.01), on the overall model. (a) In fresh semen, total progressive motility (TPM; %), was lower in the treated groups (MLT, GSH and GSH + MLT) when compared to the control group. In frozen semen, MLT group was higher than GSH and control group, however both were similar to GSH + MLT group. (b) In fresh semen the mean values of the straight-line velocity (VSL; µm/s) were lower in the treated groups (MLT, GSH and GSH + MLT) compared to the control group. In frozen semen the MLT group was superior to the GSH group, with no other differences among groups. (c) In fresh semen the straightness (STR; %) was lower in the treated groups (MLT, GSH and GSH + MLT) compared to the control group. In frozen semen there were no differences between the studied groups. (d) The mean average-path velocity (VAP; µm/s) in fresh semen was lower in the GSH group and in frozen semen no differences were detected among groups. (e) In fresh semen Linearity (LIN; %) was lower in the GSH group. In frozen semen there were no differences among groups.
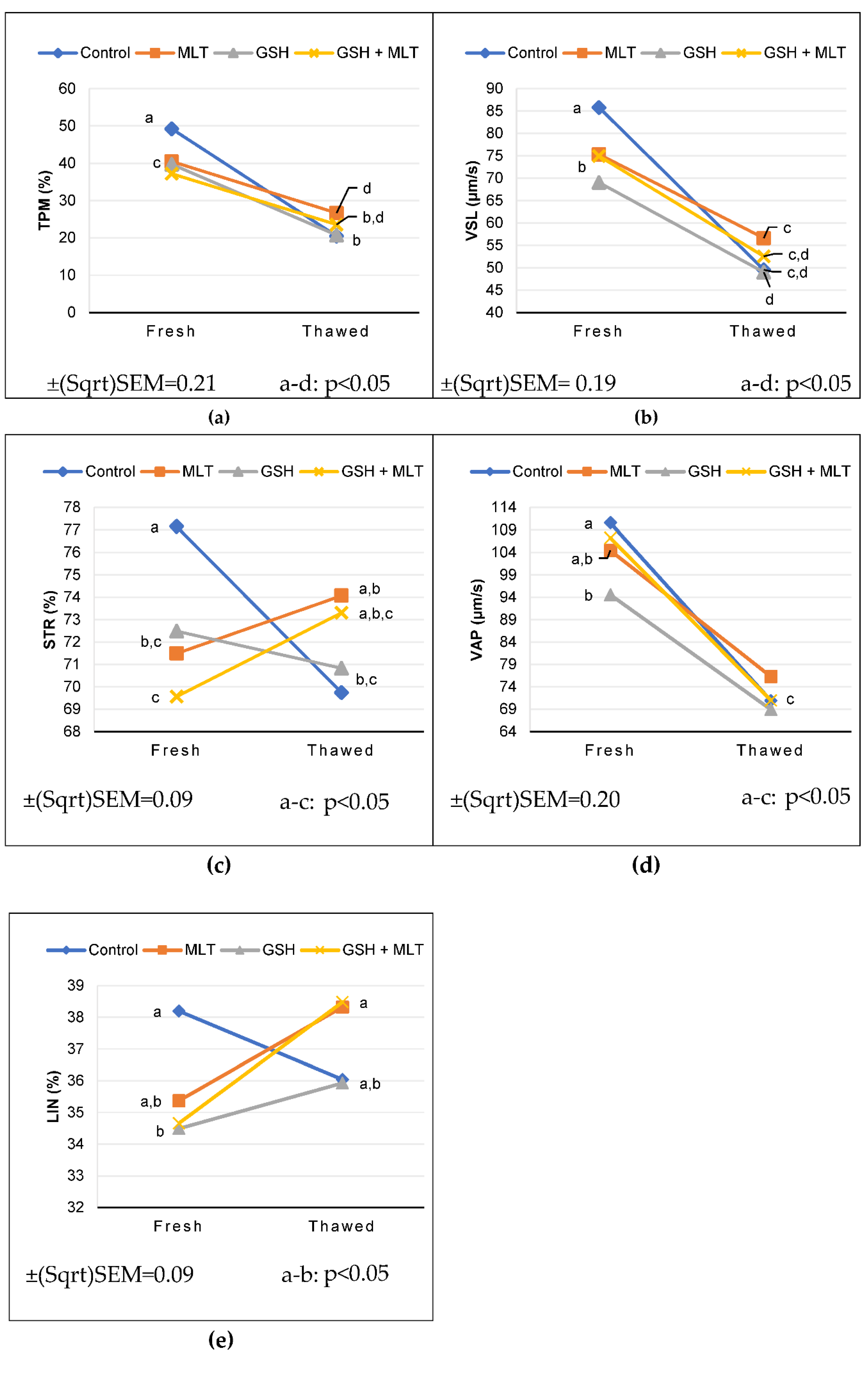

Table 1.
Effect of fixed variables (semen processing and group) and their one-way interactions.
| Parameters | Fixed Variables | Interactions | |
|---|---|---|---|
| Semen processing (SP) | Group (G) | SP x G | |
| IM | *** | NS | NS |
| Alive | *** | NS | NS |
| Death | *** | NS | NS |
| Abnormal | ** | NS | NS |
| Head defects | *** | NS | NS |
| IP defects | NS | NS | NS |
| Tail defects | *** | NS | NS |
| TS | *** | * | NS |
| TM | *** | NS | NS |
| TPM | *** | ** | *** |
| Slow | *** | * | NS |
| Medium | *** | * | * |
| Rapid | *** | NS | NS |
| VCL | *** | * | * |
| VSL | *** | *** | *** |
| VAP | *** | *** | ** |
| ALH | *** | NS | NS |
| LIN | ** | NS | ** |
| STR | NS | NS | *** |
| WOB | *** | ** | NS |
| BCF | *** | * | * |
*—p < 0.05; **—p < 0.01, ***—p < 0.001; NS—not significant (p > 0.05). IM—individual motility; IP—intermediate piece; TS—total static; TM—total motility; TPM—total progressive motility; VCL—curvilinear velocity; VSL—straight-line velocity; VAP—average-path velocity; ALH—amplitude of lateral head displacement; LIN—linearity; STR—straightness; WOB—wobble; BCF—beat cross frequency.
Table 2.
Effects of the antioxidants Melatonin (1 mM) and GSH (5 mM) on the percentage of individual motility (IM), viability (alive/death SPZ) and morphology of spermatozoa according to subjective evaluation.
Table 2.
Effects of the antioxidants Melatonin (1 mM) and GSH (5 mM) on the percentage of individual motility (IM), viability (alive/death SPZ) and morphology of spermatozoa according to subjective evaluation.
| Parameters | G1 (Control) | G2 (Melatonin) | G3 (GSH) | G4 (GSH + Melatonin) | ±(Sqrt)SEM | ||||
|---|---|---|---|---|---|---|---|---|---|
| Fresh | Thawed | Fresh | Thawed | Fresh | Thawed | Fresh | Thawed | ||
| Alive (%) | 78.7 a | 40.1 b | 79.1 a | 38.1 b | 80.9 a | 40.9 b | 82.9 a | 41.1 b | 0.17 |
| Death (%) | 20.9 a | 58.8 b | 20.0 a | 61.7 b | 18.6 a | 58.8 b | 16.6 a | 58.4 b | 0.22 |
| IM (%) | 60.0 a | 39.7 b | 58.7 a | 37.2 b | 58.7 a | 40.3 b | 59.3 a | 38.0 b | 0.08 |
| Abnormal (%) | 15.3 a | 11.6 a | 15.5 a | 11.4 a | 20.3 a | 10.8 a | 15.0 a | 12.9 a | 0.35 |
| Head D (%) | 0.8 a,b | 2.6 a,b | 1.2 a,b | 1.7 a,b | 1.1 a,b | 1.7 a,b | 0.7 b | 3.0 a | 0.37 |
| IP D (%) | 3.1 a | 3.8 a | 1.9 a | 2.7 a | 2.2 a | 2.2 a | 1.5 a | 2.1 a | 0.24 |
| Tail D (%) | 9.6 a,b,c | 3.8 c | 10.9 a,b | 5.3 b,c | 15.3 a | 5.5 b,c | 11.0 a,b | 6.3 b,c | 0.40 |
a–c different superscript letters in the same row: p < 0.05. IM—individual motility; IP—intermediate piece; D—defects; GSH—reduced glutathione.
Table 3.
Effects of antioxidants Melatonin (1 mM) and GSH (5 mM) on the percentage of total static (TS), total motility (TM), total progressive motility (TPM) and its subpopulations of spermatozoa evaluated by CASA.
Table 3.
Effects of antioxidants Melatonin (1 mM) and GSH (5 mM) on the percentage of total static (TS), total motility (TM), total progressive motility (TPM) and its subpopulations of spermatozoa evaluated by CASA.
| Parameters | G1 (Control) | G2 (Melatonin) | G3 (GSH) | G4 (GSH + Melatonin) | ±(Sqrt)SEM | ||||
|---|---|---|---|---|---|---|---|---|---|
| Fresh | Thawed | Fresh | Thawed | Fresh | Thawed | Fresh | Thawed | ||
| TS | 4.1 a | 37.6 b | 2.7 a | 35.3 b | 4.4 a | 38.9 b | 2.9 a | 36.8 b | 0.17 |
| TM | 93.4 a | 59.5 b | 95.4 a | 62.3 b | 94.0 a | 58.2 b | 95.4 a | 61.1 b | 0.11 |
| TPM | 49.2 a | 20.5 b | 40.5 c | 26.7 d | 39.7 c | 20.8 b | 37.2 c | 23.6 b,d | 0.21 |
| Slow | 0.5 a | 3.4 b | 0.4 a | 3.2 b | 2.3 b | 3.7 b | 0.4 a | 4.5 b | 0.23 |
| Medium | 3.5 b,c | 6.1 a | 3.5 b,c | 5.5 a,b | 5.8 a,b | 5.9 a,b | 2.6 c | 5.8 a,b | 0.20 |
| Rapid | 87.1 a | 46.9 b | 89.0 a | 50.3 b | 83.0 a | 44.0 b | 89.5 a,b | 44.8 b | 0.21 |
| VCL | 222.0 a | 137.2 c | 211.3 a,b | 147.5 c | 197.8 b | 136.3 c | 216.6 a,b | 135.8 c | 0.29 |
| VSL | 85.8 a | 49.5 c,d | 75.3 b | 56.6 c | 69.0 b | 49.0 d | 75.0 b | 52.5 c,d | 0.19 |
| VAP | 110.6 a | 70.9 c | 104.5 a,b | 76.3 c | 94.6 b | 68.9 c | 107.3 a | 71.0 c | 0.20 |
| ALH | 3.6 a | 2.9 b | 3.6 a | 2.9 b | 3.5 a | 2.9 b | 3.6 a | 2.8 b | 0.03 |
| LIN | 38.2 a | 36.0 a,b | 35.4 a,b | 38.3 a | 34.5 b | 35.9 a,b | 34.7 b | 38.5 a | 0.09 |
| STR | 77.2 a | 69.7 b,c | 71.5 b,c | 74.1 a,b | 72.5 b,c | 70.8 b,c | 69.6 c | 73.3 a,b,c | 0.09 |
| WOB | 49.4 b,c,d | 51.3 a,b,c | 49.2 c,d | 51.5 a,b | 47.5 d | 50.4 a,b,c | 49.4 b,c,d | 52.2 a | 0.05 |
| BCF | 22.8 a | 18.5 c,d | 20.8 a,b | 19.7 b,c | 20.8 a,b | 17.6 d | 21.4 a,b | 19.2 b,c,d | 0.09 |
a–d different superscript letters in the same row: p < 0.05. GSH—reduced glutathione; TS—total static (%); TM—total motility (%); TPM—total progressive motility (%); Slow (%); Medium (%); Rapid (%); VCL—curvilinear velocity (μm/s); VSL—straight-line velocity (μm/s); VAP—average-path velocity (μm/s); ALH—amplitude of lateral head displacement (μm); LIN—linearity (%); STR—straightness (%); WOB—wobble (%); BCF—beat cross frequency (Hz).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



