
Submitted:
25 July 2023
Posted:
26 July 2023
You are already at the latest version
Abstract
Mesenchymal stem cells (MSCs) are an attractive therapeutic tool for tissue engineering and regenerative medicine owing to their regenerative and trophic properties. The best-known and most widely used are bone marrow MSCs which are currently being harvested and developed from a wide range of adult and perinatal tissues. MSCs from different sources are believed to have different secretion potentials and production which may influence their therapeutic effects. To confirm it, we performed a quantitative proteomic analysis based on the TMT technique of MSCs from three different sources: Wharton’s jelly (WJ), dental pulp (DP) and bone marrow (BM). Our analysis focused on MSC biological properties of interest for tissue engineering. We identified a total of 611 differentially expressed human proteins. WJ-MSCs showed the greatest variation compared with the other sources. WJ produced more extracellular matrix (ECM) proteins and ECM-affiliated proteins and appeared more able to modulate the inflammatory and immune response. BM-MSCs displayed enhanced differentiation and paracrine communication capabilities. DP-MSC appeared to promote exosome production. The results obtained confirm the existence of differences between WJ, DP and BM-MSC and the need to select the MSC origin according to the therapeutic objective sought.
Keywords:
mesenchymal stem cells
; proteomics
; tissue engineering
; regenerative medicine
1. Introduction
MSCs were initially isolated from bone marrow (BM) based on their ability to adhere to plastic culture dishes and to form colony-forming unit-fibroblast (CFU-F) [1]. They are capable of self-renewal, produce extracellular matrix (ECM) and are able to differentiate into multiple cell types [2]. BM-MSC has been shown to have immunomodulatory properties, controlling inflammation and modifying nearby immune cells [3,4]. All of these properties have made MSC a prime candidate for tissue engineering and regenerative medicine. BM-MSC transplants have therefore been shown to be beneficial for cartilage regeneration [5], bone tissue regeneration [6] and in acute and chronic models of muscle degeneration [7].
Tissue engineering aims to replace, restore, maintain or improve the function of human tissues through the laboratory production of biological substitutes for transplantation. In order to create these biological substitutes, the cell part is composed of stem cells which are used primarily for their ability to differentiate into a desired cell type. MSCs, for example, are widely used for their ability to differentiate into osteoblasts to generate a bone substitute [8]. In this domain, an approach consisting of using only one MSC capacity (e.g., BMP2 synthesis, osteo-differentiation) has provided only limited results [9,10] for good reason: mimicking the in vivo environment (the aim of tissue engineering) means reproducing a complex micro-environment. MSCs are good candidates since they produce ECM which participates in the establishment and maintenance of tissues and secrete many factors that promote, among other things, homing [11].
Since the use of bone marrow derived MSCs, it is recognized that MSC populations can be isolated from a variety of tissues including adipose tissue, muscle, tendon, peripheral blood, umbilical cord, skin, dental tissue… [12,13,14,15]. MSCs isolated from umbilical cord (UC-MSC), adipose tissue (AT-MSC) or dental pulp (DP-MSC) have significant advantages over BM-MSCs, such as a painless collection, easy extraction or a high proliferation capacity. For these reasons, these MSCs now tend to replace BM-MSCs in tissue engineering and are beginning to be used in regenerative medicine [16,17,18,19].
But do MSCs from different tissues have exactly the same capabilities? Studies first compared different sources of MSCs (mainly UC-MSC, AT-MSC and BM-MSC) on their ability to form colonies (CFU-F), their expansion potential, their differentiation capacity and their cell surface marker expressions [20,21,22,23,24]. Apart from their proliferative potential, no major differences were found between these different MSC sources. Other studies then took the analysis further using omics-based methods: comparisons of the transcriptome, proteome, secretome and even exosomes of these different MSC sources were performed. [25,26,27,28,29].
In the present study, we made a proteomic comparison of three different sources of MSC using the TMT-based quantitative technique. Our analysis focused on the MSC biological properties of interest for tissue engineering and regenerative medicine. Indeed, MSCs used in innovative therapy and mainly in tissue engineering must be able to recreate (alone or by paracrine effect) the injured tissue: (i) by differentiating into cells of interest, (ii) by producing ECM and/or (iii) by homing thanks to secreted proteins and finally, (iv) they must limit inflammation and modulate the immune response.
We compared three sources of MSC: umbilical cord-derived Wharton’s jelly MSC (WJ-MSC), dental pulp MSC (DP-MSC) and bone marrow MSC (BM-MSC), cultivated under proliferative conditions. The objective was to identify whether there are sufficient proteomic differences to make an MSC source more attractive for a given cell therapy application.
2. Results
The TMT-based quantitative proteomics technique we used enabled us to analyze the proteins present in all of the samples.
2.1. Qualitative Analyses of the 3 Sources of hMSC
2.1.1. Differentially Expressed Proteins and Biological Processes Involved
We identified a total of 611 differentially expressed human proteins. The biological processes affected by these variations of expression between cell sources are shown in Figure 1. The biological process with the greatest variation in protein expression (outside of the "other metabolic or biological" groups) was cellular organization and biogenesis at 12%.
2.1.2. Group Comparison
Pairwise comparison: BM vs. DP, BM vs. WJ and DP vs. WJ (Figure 2), showed a stronger difference in protein expression (either up- and downexpressed) between BM-MSC and WJ-MSC (Figure 2a). In contrast, BM vs. DP had the fewest variant proteins (Supplemental Figure S1). These results can be explained by the fact that BM-MSC and DP-MSC are both adult MSCs.
The GO classification of differentially expressed proteins with comparisons: BM vs. DP, BM vs. Wj and DP vs. WJ (Figure 2b) highlight several features: camparisons BM vs. DP and BM vs. WJ revealed many underexpressed proteins in the majority of biological processes. This suggests that BM-MSC expressed fewer proteins involved in different biological processes than other cell sources with the exception of signal transduction. The pairwise comparison DP vs. WJ showed overexpression of proteins involved in nine of the 13 biological processes listed In Figure 2b. DP-MSC overexpress more proteins than other sources for the following biological processes: cell cycle or cell proliferation, cell organization and biogenesis, developmental processes, stress response, transport and DNA metabolism. WJ-MSC overexpressed proteins in the major processes: cell adhesion, protein metabolism, RNA metabolism and transcription, and cell-cell signaling (Supplemental Figures S2).
2.2. Analyses Focused on the Functions of Interest of MSC in Cell Therapy
2.2.1. MSC Characteristics
The International Society for Cellular Therapy has defined MSCs as cells with a specific immunophenotype, ex vivo plastic-adherent growth, and multilineage differentiation [30]. We therefore looked for differences in protein expression from these features.
- CD markers of MSC
Among the CD marker profiles, four membrane proteins had differences in expression between MSC sources with three in the CAM family (Table 1). BM-MSCs expressed the most Endoglin: CD105, VCAM1: CD106 and MCAM: CD146 compared with other sources. WJ-MSC down expressed VCAM1: CD106 and ALCAM: CD166 compared with other sources. More than just cell markers, these proteins are involved in important processes such as angiogenesis and immune response.
- Differentiation capacities
Some proteins have been identified in those differentially expressed between the different sources of MSC as being involved in osteoblastic, adipocyte, neuronal or muscle differentiation (Table 2). Alkaline phosphatase, an important enzyme in bone mineralization, was underexpressed by WJ-MSC compared with DP-MSC and BM-MSC. Compared with the others, WJ-MSC produced more Protaglandin G/H synthase 2, while BM-MSC produced more Adipogenesis regulatory factor. BM-MSCs seemed to be the most capable of ensuring adipocyte differentiation as opposed to WJ-MSCs. Regarding other types of differentiation, it seems that BM-MSC, which overexpressed more proteins, was more capable of neuronal or muscle differentiation.
2.2.2. ECM Production
ECM is a major component of the cellular microenvironment. It is composed of structural components (collagens, ECM glycoproteins and proteoglycans) and ECM-associated proteins. The production of ECM by MSCs is one of the criteria for selecting them for tissue engineering.
- collagens, ECM glycoproteins and proteoglycans
The ECM structural components differentially expressed in the pairwise comparisons BM/DP, BM/WJ and DP/WJ as shown in Figure 3.
Collagen is the most abundant fibrous protein in ECM. There are 28 different types in humans and in our study 11 collagens were identified as being differentially expressed in the three MSC sources. Col14A1 was most highly expressed by BM-MSC. WJ-MSC most expressed Col3A1, Col4A1, Col4a2, Col5A1 and Col16A1. DP-MSCs underexpressed Col4A1, Col8A1 and Col11A1 compared with other sources.
Some ECM glycoproteins such as fibronectin are ubiquitous, while others have more specific localizations such as the laminins of basal membranes. They contain several structural and functional domains, several sites of cell attachment via integrins or other receptors, the most frequent of which contain the Arg-Gly-Asp (RGD) sequence and several sites of interaction with other extracellular macromolecules. Since they are capable of numerous interactions with the microenvironment, a strong variation in their expression can have a real impact on MSC capacities. In our study, among the proteins identified as statistically variant, members of insulin-like growth factor binding protein (IGFBP) family: IGFBP-4, IGFBP-5 and IGFBP-7 were found to be highly expressed by BM-MSCs compared with DP-MSC. BM-MSC overexpressed MFGE8 protein compared with the other sources of MSC. DP-MSC strongly expressed fibrilin-2 known to regulate the early process of elastic fiber assembly. This protein also regulates osteoblast maturation by controlling TGF-beta bioavailability and calibrating TGF-beta and BMP levels. WJ-MSC overexpressed Fibronectin, EMILIN-1, Fibulin-2, IGFBP7 and Nidogen-2 compared with the other sources.
Proteoglycans make up the interstitial matter. They form a hydrophilic gel with a wide range of functions such as tissue hydration, modulation of signaling pathways and resistance to tensile forces. Our analysis revealed a strong variation in the expression of a number of proteoglycans in WJ-MSC. WJ-MSC strongly underexpressed three of the four members of the Syndecan family. Conversely, they overexpressed the other identified proteoglycans: Biglycan, Tenascin and VCAN. DP-MSC overexpressed Syndecan-1 and Syndecan-4 compared with BM- and WJ-MSC.
- ECM-associated proteins
All differentially expressed proteins known to be associated with, interact with or regulate ECM are listed in Figure 4 (and Supplemental Figures S3).
Most of the ECM regulators presented here are enzymes or proteins involved in ECM remodeling through bond formation or degradation. In the BM/DP comparison, among the nine proteins differentially expressed, all proteins are overexpressed by BM-MSCs with the exception of Neprilysin. DP-MSC was found to overexpressed just one protein compared with the other two sources: Neprilysin, a protein involved in elastin degradation. WJ-MSC overexpressed a number of proteins compared with other sources such as: ADAMTSL, Dipeptidyl peptidase 4, ITIH1, ITIH2, SERPINE1 or TGM2.
The group of ECM-affiliated proteins was largely made up of the integrin family. The expression of the various integrins changed from source to source of MSC, with no particular trend. WJ underexpressed Integrin beta-like protein 1: a protein that promotes cell migration.
2.2.3. Cell-Cell Signaling
Proteins belonging to the biological process "cell-cell signaling" (GO term) identified as significantly variant between the different MSC sources are shown in Figure 5.
The ratios with WJ-MSC showed the most significant differences in protein expression. Compared with BM and DP, WJ-MSC overexpressed CXCL6, GDF15, GPC6, HMGA2, INHBA, NAMPT, PLAT and PTGS2, and for some of them very strongly. WJ-MSC underexpressed only three proteins: CAV1, DAB2 and PLPP3 compared with other cell sources. DP-MSC overexpressed three proteins: AMPH, NXN and SDC1 and underexpressed JUP. BM-MSC overexpressed GPC4 but underexpressed more proteins: CTHRC1, GREM1 and LTBP4 (Supplemental Figures S3).
2.2.4. Inflammation and Immune Response
MSCs have been shown to secrete a broad spectrum of bioactive molecules that induce a variety of responses including inhibition of inflammatory and/or immune responses. We identified seven proteins involved in the inflammatory response and 11 proteins in the immune response (Figure 6).
BM-MSC overexpressed PTGES3 when DP-MSC overexpressed Follistatin and Galectin-3 compared with the others. WJ-MSC overexpressed PTGS2 and PTX3 compared with BM and DP-MSC. The differences between cell sources were most marked in the immune response with the presence of important proteins such as ADGRE5, ICAM1, CD200, VCAM1, or DPP4. WJ-MSC overexpressed ICAM1, CD200, CXCL6 and DPP4 compared with BM and DP-MSC but underexpressed ALCAM, CAMK1D, VCAM1 and SERPINB1. BM-MSC only overexpressed VCAM1 compared with the two others. DP-MSC overexpressed Nectin-3 and underexpressed CD200 and DDP4 compared with BM and WJ-MSC (Supplemental Figures S3).
3. Discussion
TMT-based quantitative proteomic analysis was performed on MSCs grown under conventional culture conditions. We did not generate any particular condition such as hypoxia treatment that could generate a response to the stimuli. The aim of this study was to highlight differences in level protein expression and only the proteins produced by all of the samples were analyzed.
The comparison of our study with other articles is not obvious since no published study to date has performed the same proteomic analysis of BM-MSC, WJ-MSC and DP-MSC. One team compared human mesenchymal stem cells derived from dental pulp, bone marrow, adipose tissue, and umbilical cord tissue by expression of 15 pluripotent stem cell genes [23]. There are articles on comparative proteomic analysis but the cell sources are not the same. Our comparison with the literature can only be partial.
Qualitative analyses of the three MSC sources revealed a number of differentially expressed proteins of the same order of magnitude as those found in other articles [27,28]. Comparing these articles, biological processes impacted by these differences in expression are not always the same (it depends in part on the cell sources), but we have in common among the most variant: "cell organization and biogenesis" and "secretion by cell" which for us was included in the group "other biological process".
In group comparison, the Venn diagram shows a greater difference between WJ-MSC and other sources. This result can be explained by the fact that BM and DP are both adult MSC whereas WJ-MSC has a fetal origin. Shin et al. showed that the secretome of fetal-derived MSCs, such as PL and WJ, had a more diverse composition than that of AD and BM-derived MSCs [28].
In analyzing the quantitative results, we chose to focus on the elements that make MSCs a tool of choice for cell therapy.
CD markers are classically analyzed by cytometry. This type of analysis highlights the positivity or otherwise of these cells to the CD markers sought. Proteomic analysis gives a much more precise idea of the expression of these markers. We were therefore able to show that BMs are the source of cells that express the most CD105, CD106 and CD146, while WJs express CD106 and CD166 the least. This difference in expression could reflect a preference of interaction. The function of these markers on the MSC surface is still poorly understood. We hypothesize that the difference in expression observed in BM-MSCs compared with other sources could be because they belong to the hematopoietic niche [31].
Another important feature of MSCs is their ability to differentiate into various cellular types. This is the point most studied during MSC source comparisons. It is important to couple differentiation capacity with proliferation capacity. Indeed, it is known that there is a balance between proliferation and differentiation. Thus, WJ-MSCs, which proliferate faster than BM-MSCs, appear to have little or no capacity to differentiate into osteoblasts or adipocytes. [21,22,32]. This is also what we found in our study with WJ-MSCs underexpressing ALPL, a protein that is essential for osteoblastic mineralization, and overexpressing PTGS2, a protein that suppresses adipocyte differentiation. Our results suggested that BM-MSCs are superior to other sources in osteogenic, adipogenic, neuronal or muscular differentiation. By comparing gene expression profiles, Hsieh and his team found that BM-MSCs were more capable of osteogenic and adipogenic differentiation, while WJ-MSCs proliferated more [33]. In a study comparing fetal MSC sources (WJ, fetal and maternal side of placenta) and adult MSCs (BM, AT), BM-MSCs showed the greatest capacity for differentiation [34]. But not all articles go in the same direction. For example, Donders and his team showed that WJ overexpressed genes involved in differentiation, maturation and neuronal support, compared with BM-MSC [35].
In tissue engineering, one of the most important points is the ability of the cells used to generate extracellular matrix. In addition to providing physical support to cells, ECM actively participates in the establishment and maintenance of differentiated tissues and organs by regulating growth factors, hydration levels and the pH of the local environment [36]. Among ECMs, collagens are well represented in our study. WJ-MSCs overexpressed the most collagen types compared to other sources, and collagens of different classes: fibrillar, fibrillar-associated collagen with interrupted triple helics, basement membrane, filamentous, short chain and multiplexins. Compared with the other two groups, BM-MSC overexpressed only Col14A1 which is often present in areas of high mechanical stress, indicating that it potentially has a role in maintaining mechanical tissues.
In the proteoglycans identified as differentially expressed, again it is WJ-MSCs that overexpress the most proteins. However, we noted the presence of three members of the IGFBP family. These proteins are more highly expressed by BM-MSCs than by DP-MSCs. This IGFBP protein family serves as a transport protein for insulin-like growth factor-1 (IGF-1), influences the bioavailability of IGFs and therefore reduces their signaling with cell receptors. IGFs stimulate the proliferation of differentiated chondrocytes, leading to enlargement of the conjugation cartilage and elongation of the bone. In addition, IGFs promote the growth of all tissues, stimulate protein synthesis and enhance Ca2+ uptake. IGF-1 therefore enables growth or at least limits age-related bone loss [37].
If we look at all of the components of all ECM, we can see that WJ-MSCs overexpress most of the players in the basal lamina, such as collagen 4, laminins, nidogen and integrins. The basal lamina enables epithelial cells to adhere to the underlying connective tissue, and constitutes a major interface between epithelial tissue cells and the body's interior for the regulation and diffusion of nutrients. It also plays a role in the survival, proliferation and differentiation of cells in the various epithelial tissues [36].
Among the ECM regulators, BM and WJ-MSC shared the overexpression of the various proteins identified. ECM-affiliated proteins were mainly represented by the integrin family. To be functional, integrins must form heterodimers (composed of an alpha and beta chain). Here, no functional heterodimer appears to be over- or underexpressed. In fact, the alpha integrins identified dimerized with beta 1 integrin, which showed no difference in expression. Similarly, integrin beta 3 normally associates with alphaIIb or alpha V, which were not present in our analysis. Apart the integrins, we can see the overexpression of Galectin-3 by the DP-MSC compared with BM and WJ. Galectin-3 modulates important interactions between epithelial cells and extracellular matrix, which promotes tissue vascularization [38].
Proteins belonging to the biological process "cell-cell signaling" are largely composed of transcriptional regulators, growth factor or hormone regulators. A number of Wnt pathway players are presented: FZD7, NXN, RECK, WNT5A. The Wnt signaling pathway is very important in MSC (as Notch). It is involved in osteoblastic, adipocytic and chondrocytic differentiation [39,40]. Above all, it plays a role in MSC tissue regeneration [41].
Two proteins involved in exocytosis are also differentially expressed: SDC1 and AMPH. They are overexpressed by DP-MSC compared with BM and WJ. Exosomes secreted by MSCs have been the subject of recent studies. They are proving to be one of the main mechanisms of the therapeutic action of MSCs which has so far been neglected [42,43,44]. Comparative proteomic analysis of exosomes from three MSC sources revealed differences in capacity. This would justify the choice of an MSC source based on potential applications [27].
Our study did not reveal differences in the expression of important factors secreted by MSCs such as proteins involved in angiogenesis: HGF, IGF-1, MCP-1, Angiogenin or VEGF, or proteins involved in hematopoiesis: TGFB1, TGFB2, GDF6, VEGF-C, M-CSF, CSF or interleukins.
In contrast, expression differences were observed for proteins involved in inflammatory and/or immune responses. Within these two categories, WJ-MSCs showed the greatest variation in protein expression. Like Donders et al., we found the overexpression of CD200, ICAM-1 by WJ-MSCs [35]. Although the main secreted factors responsible for the immunological and anti-inflammatory competence of MSCs described in the literature did not emerge in our study [45,46], the proteins identified here suggest that WJ-MSCs are the most potent for modulating the inflammatory and immune response.
4. Materials and Methods
Proteins were extracted using EasyPrep sample preparation kit (Pierce, ThermoFisher Scientific) using manufacturer recommendations. The proteins were quantified using the Micro BCA kit method (Pierce, San Jose, CA, USA). Twenty-five micrograms of proteins of each sample were digested and labeled with TMTpro™ 16-plex reagents (ThermoFisher Scientific, San Jose, CA, USA), mixed in equimolar amounts. A fractionation was purchased using the High pH Reversed-Phase Peptide Fractionation Kit (Pierce, San Jose, CA, USA) according to manufacturer recommendations. The tryptic peptide solutions were dried under vacuum and reconstituted in 20 µL water/1% formic acid (v/v) each. The LC–MS/MS platform consisted of an Ultimate 3000 RSLC UPLC system coupled to an Orbitrap Fusion mass spectrometer (MS) (ThermoFisher Scientific) with a nano-trap column (Acclaim PepMap 100 Å C18, 5 µm, 100 µm i.d. x 2 cm length, ThermoFisher Scientific) and an Easy-Spray column (Acclaim PepMap 100 Å C18, 2 µm, 75 µm i.d. x 50 cm length, ThermoFisher Scientific). Ten fractions of the TMT-labeled digest were separated by on-line nanoLC and analyzed by nano-electrospray tandem mass spectrometry. The overall workflow of the analysis is presented in Figure 1. The peptide mixtures were injected onto a nano-trap column with a flow of 5 µL/min and subsequently gradient eluted with a flow of 300 nL/min, from 4% to 30% acetonitrile (v/v) for 140 min. Each fraction was analyzed on an Orbitrap Fusion MS using synchronous precursor selection (SPS) MS3 quantitation. The full scan was performed in the range of 375–2000 m/z at nominal resolution of 120,000 at 200 m/z and AGC set to 4.105, followed by selection of the most intense ions above an intensity threshold of 5000 for collision-induced dissociation (CID)-MS2 fragmentation in the linear ion trap with 35% normalized collision energy. The isolation width for the frontal cortex samples was set to 0.7 m/z with no offset. The top 10 fragment ions for each peptide MS2 were notched out with an isolation width of 2 m/z and co-fragmented to produce MS3 scans analyzed in the MS at a nominal resolution of 50,000 after higher-energy collision dissociation (HCD) fragmentation at a normalized collision energy of 65%. Data were processed using Proteome Discoverer 2.5 (ThermoFisher Scientific, Bremen, Germany) before being run against Homo sapiens Uniprot database (release 2022_12). Parameters were specified as follows: trypsin enzyme, two miscleavages allowed, minimum peptide length of six amino acids, TMT tags on lysine residues and peptide N-termini (+304.207 Da), carbamidomethylation of cysteine residues (+57.021 Da) as fixed modifications, oxidation of methionine residues (+15.995 Da) and acetylation of protein N-termini (+42.011 Da) as variable modifications, precursor mass tolerance of 10 ppm, and a fragment mass tolerance of 0.6 Da. Peptide spectral match (PSM) error rates were determined using the target-decoy strategy coupled to Percolator modeling of true and false matches [47]. Reporter ions were quantified from MS3 scans using an integration tolerance of 20 ppm with the most confident centroid setting. An MS2 spectral assignment false discovery rate (FDR) of less than 1% was achieved by applying the target-decoy strategy. Following spectral assignment, peptides were assembled into proteins and were further filtered based on the combined probabilities of their constituent peptides to a final FDR of 1%. In addition, we only validated the proteins that were present in the five biological replicates. In cases of redundancy, shared peptides were assigned to the protein sequence with the most matching peptides, thereby adhering to the principles of parsimony. The DAPs were identified based on a t-test with p-value less than 0.05 and with a fold change >2 or <0.5 (Supplemental Figures S1). The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium (http://www.proteomexchange.org (accessed on July 19, 2023) via the PRIDE partner repository with dataset identifier PXD043912 an [48].
5. Conclusions
In this study, we succeeded in highlighting the differences in proteomic expression of three MSC sources and linking these differences with their applicative interest for regenerative medicine and tissue engineering. WJ-MSCs showed the greatest variation compared with the other sources. WJ produced more ECM proteins or ECM-affiliated proteins and appeared to be better able to modulate the inflammatory and immune response. BM-MSCs displayed enhanced differentiation and paracrine communication capabilities. DP-MSC appeared to promote exosome production. None of the MSC sources is without interest. The results obtained confirm the need to select the origin of MSCs according to the desired therapeutic objective.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, M.N., J.L.B. and P.M.; methodology, J.L.B, P.M.; software, P.M.; validation, M.N.,J.L.B. and P.M.; formal analysis, P.M.; investigation, M.N., J.L.B.; resources, M.N., P.M.; data curation, P.M.; writing-original draft preparation, M.N.; writing-review and editing, M.N., P.M.; visualization, M.N., P.M.; supervision, M.N., P.M.; project administration, M.N., P.M.; funding acquisition, P.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors would like to thank Emma Catel-Dobel for her preliminary study, which helped conceptualize this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Friedenstein, A.J.; Chailakhyan, R.K.; Gerasimov, U.V. Bone marrow osteogenic stem cells: In vitro cultivation and transplantation in diffusion chambers. Cell Prolif. 1987, 20, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Bianco, P. ; Robey; P. G., Simmons, P.J. Mesenchymal stem cells: revisiting history, concepts, and assays. Cell Stem Cell. 2008, 2, 313–319. [Google Scholar] [CrossRef]
- Ryan, J.M.; Barry, F.P.; Murphy, J.M.; Mahon, B.P. , Mesenchymal stem cells avoid allogeneic rejection. J. of Inflammation 2005, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Chiu, S.M.; Motan, D.A.L.; Zhang, Z.; Chen, L.; Ji, H.-L.; Tse, H.-F.; Fu, Q.-L.; Lian, Q. Mesenchymal stem cells and immunomodulation: current status and future prospects. Cell Death & Disease 2016, 7, e2062. [Google Scholar] [CrossRef]
- Nam, H.Y.; Karunanithi, P.; Loo, W.C.; Naveen, S.; Chen, H.; Hussin, P.; Chan, L.; Kamarul, T. The effects of staged intra-articular injection of cultured autologous mesenchymal stromal cells on the repair of damaged cartilage: a pilot study in caprine model. Arthritis Research & Therapy 2013, 15, R129. [Google Scholar] [CrossRef]
- Bruder, S.P.; Kurth, A.A.; Shea, M.; Hayes, W.C.; Jaiswal, N.; Kadiyala, S. Bone regeneration by implantation of purified, culture-expanded human mesenchymal stem cells. J Orthop Res. 1998, 16, 155–162. [Google Scholar] [CrossRef]
- Salah-Mohellibi, N.; Millet, G.; André-Schmutz, I.; Desforges, B.; Olaso, R.; Roblot, N.; Courageot, S.; Bensimon, G.; Cavazzana-Calvo, M.; Melki, J. Bone marrow transplantation attenuates the myopathic phenotype of a muscular mouse model of spinal muscular atrophy. Stem Cells. 2006, 12, 2723–2732. [Google Scholar] [CrossRef] [PubMed]
- Oryan, A.; Kamali, A.; Moshiri, A.; Eslaminejad, M.B. Role of mesenchymal stem cells in bone regenerative medicine: what is the evidence? Cells Tissues Organs 2017, 204, 59–83. [Google Scholar] [CrossRef] [PubMed]
- Decambron, A.; Fournet, A.; Bensidhoum, M.; Manassero, M.; Sailhan, F.; Petite, H.; Logeart-Avramoglou, D.; Viateau, V. Low-dose BMP-2 and MSC dual delivery onto coral scaffold for critical-size bone defect regeneration in sheep. J Orthop Res. 2017, 12, 2637–2645. [Google Scholar] [CrossRef] [PubMed]
- Zong, C.; Xue, D.; Yuan, W.; Wang, W.; Shen, D.; Tong, X.; Shi, D.; Liu, L.; Zheng, Q.; Gao, C.; Wang, J. Reconstruction of rat calvarial defects with human mesenchymal stem cells and osteoblast-like cells in poly-lactic-co-glycolic acid scaffolds. Eur Cell Mater. 2010, 20, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.; Liu, D.; Thakor, A.S. Mesenchymal Stromal Cell Homing: Mechanisms and Strategies for Improvement. iScience 2019, 15, 421–438. [Google Scholar] [CrossRef]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.-W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; Norotte, C.; Teng, P.-N.; Traas, J.; Schugar, R.; Deasy, B.M.; Badylak, S.; Buhring, H.-J.; Giacobino, J.-P.; Lazzari, L.; Huard, J.; Péault, B. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell. 2008, 3, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Mankani, M.; Brahim, J.; Gehron Robey, P.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc Natl Acad Sci USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed]
- Seo, B.-M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.-Y.; Shi, S. Investigation of Multipotent Postnatal Stem Cells from Human Periodontal Ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Riekstina, U.; Muceniece, R.; Cakstina, I.; Muiznieks, I.; Ancans, J. Characterization of human skin-derived mesenchymal stem cell proliferation rate in different growth conditions. Cytotechnology 2008, 58, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Qu, Z.; Yin, X.; Shang, C.; Ao, Q.; Gu, Y.; Liu, Y. Efficacy of umbilical cord-derived mesenchymal stem cell-based therapy for osteonecrosis of the femoral head: A three-year follow-up study. Molecular Medecine Reports 2016, 14, 4209–4215. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.S.; Yen, H.H.; Chang, C.Y.; Lien, W.C.; Huang, S.H.; Lee, S.S.; Wang, L.; Wang, H.D. Adipose-Derived Stem Cell-Incubated HA-Rich Sponge Matrix Implant Modulates Oxidative Stress to Enhance VEGF and TGF-β Secretions for Extracellular Matrix Reconstruction In Vivo. Oxid Med Cell Longev. 2022, 9355692. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Yin, Z.; Wu, T.; Li, Y.; Luo, X.; Xu, M.; Duan, L.; Li, J. Cell Transplantation 2018, 27, 1634–1643. [CrossRef]
- Suda, S.; Nito, C.; Ihara, M.; Iguchi, Y.; Urabe, T.; Matsumaru, Y.; Sakai, N.; Kimura, K. Randomised placebo-controlled multicentre trial to evaluate the efficacy and safety of JTR-161, allogeneic human dental pulp stem cells, in patients with Acute Ischaemic stRoke (J-REPAIR). BMJ Open 2022, 12, e054269. [Google Scholar] [CrossRef]
- Prasanna, J.S.; Gopalakrishnan, D.; Shankar, S.R.R.; Vasandan, A.B. Pro-Inflammatory Cytokines, IFNc and TNFa, Influence Immune Properties of Human Bone Marrow and Wharton Jelly Mesenchymal Stem Cells Differentially. PLoS ONE 2010, 5, e9016. [Google Scholar] [CrossRef]
- Bosch, J.; Houben, A.P.; Radke, T.F.; Stapelkamp, D.; Bünemann, E.; Balan, P.; Buchheiser, A.; Liedtke, S.; Kögler, G. Distinct Differentiation Potential of ‘‘MSC’’ Derived from Cord Blood and Umbilical Cord: Are Cord-Derived Cells True Mesenchymal Stromal Cells? Stem Cells and Development 2012, 21, 1977–1988. [Google Scholar] [CrossRef]
- Raicevic, G.; Najar, M.; Pieters, K.; De Bruyn, C.; Meuleman, N.; Bron, D.; Toungouz, M.; Lagneaux, L. Inflammation and Toll-Like Receptor Ligation Differentially Affect the Osteogenic Potential of Human Mesenchymal Stromal Cells Depending on Their Tissue Origin. Tissue Eng Part A 2012, 18, 1410–1418. [Google Scholar] [CrossRef]
- Stanko, P.; Kaiserovab, K.; Altanerovab, V.; Altanerb, C. Comparison of human mesenchymal stem cells derived from dental pulp, bone marrow, adipose tissue, and umbilical cord tissue by gene expression. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2014, 158, 373–377. [Google Scholar] [CrossRef]
- Trivanović, D.; Jauković, A.; Popović, B.; Krstić, J.; Mojsilović, S.; Okić-Djordjević, I.; Kukolj, T.; Obradović, H.; Santibanez, J.F.; Bugarski, D. Mesenchymal stem cells of different origin: Comparative evaluation of proliferative capacity, telomere length and pluripotency marker expression. Life Sciences 2015, 141, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Pires, A.O.; Mendes-Pinheiro, B.; Teixeira, F.G.; Anjo, S.I.; Ribeiro-Samy, S.; Gomes, E.D.; Serra, S.C.; Silva, N.A.; Manadas, B. , Sousa, N.; Salgado, A.J. Unveiling the Differences of Secretome of Human Bone Marrow Mesenchymal Stem Cells, Adipose Tissue derived Stem Cells and Human Umbilical Cord Perivascular Cells: A Proteomic Analysis. Stem Cells Dev. 2016, 25, 1073–1083. [Google Scholar] [CrossRef]
- Barrett, A.N.; Fong, C.Y.; Subramanian, A.; Liu, W. , Feng, Y.; Choolani, M.; Biswas, A.; Rajapakse, J.; Bongso, A. Human Wharton’s Jelly Mesenchymal Stem Cells Show Unique Gene Expression Compared to Bone Marrow Mesenchymal Stem Cells Using Single-Cell RNA-Sequencing. Stem Cells Dev. 2019, 28, 196–211. [Google Scholar] [CrossRef]
- Wang, Z.G.; He, Z.Y.; Liang, S.; Yang, Q.; Cheng, P.; Chen, A.M. Comprehensive proteomic analysis of exosomes derived from human bone marrow, adipose tissue, and umbilical cord mesenchymal stem cells. Stem Cell Research & Therapy, 2020, 11, 511. [Google Scholar] [CrossRef]
- Shin, S.; Lee, J.; Kwon, Y.; Park, K.-S.; Jeong, J.-H.; Choi, S.-J.; Bang, S.I.; Chang, J.W.; Lee, C. Comparative Proteomic Analysis of the Mesenchymal Stem Cells Secretome from Adipose, Bone Marrow, Placenta and Wharton’s Jelly. Int. J. Mol. Sci. 2021, 22, 845. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Swaminathan, G.; Garcia-Marques, F.; Regmi, S.; Yarani, R.; Primavera, R.; Chetty, S.; Bermudez, A.; Pitteri, S.J.; Thakor, A.S. Integrated transcriptome-proteome analyses of human stem cells reveal source-dependent differences in their regenerative signature. Stem Cell Reports 2022, 18, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; DJ Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Crippa, S.; Bernardo, M.E. Mesenchymal Stromal Cells: Role in the BM Niche and in the Support of Hematopoietic Stem Cell Transplantation. HemaSphere 2018, 2, 6. [Google Scholar] [CrossRef]
- Naudot, M.; Barre, A.; Caula, A.; Sevestre, H.; Dakpé, S.; Mueller, A.A.; Devauchelle, B.; Testelin, S.; Marolleau, J.P.; Le Ricousse, S. Co-transplantation of Wharton's jelly mesenchymal stem cellderived osteoblasts with differentiated endothelial cells does not stimulate blood vessel and osteoid formation in nude mice models. J Tissue Eng Regen Med. 2020, 14, 257–271. [Google Scholar] [CrossRef]
- Hsieh, J.Y.; Fu, Y.S.; Chang, S.J.; Tsuang, Y.H.; Wang, H.W. Functional module analysis reveals differential osteogenic and stemness potentials in human mesenchymal stem cells from bone marrow and Wharton’s Jelly of umbilical cord. Stem Cell and Development, 2010, 19, 1895–1910. [Google Scholar] [CrossRef]
- Sangeetha, K.N.; Vennila, R.; Secunda, R.; Sakthivel, S.; Pathak, S.; Jeswanth, S.; Surendran, R. Functional variations between Mesenchymal Stem Cells of different tissue origins: A comparative gene expression profiling. Biotechnol Lett. 2020, 42, 1287–1304. [Google Scholar] [CrossRef]
- Donders, R.; Bogie, J.F.J.; Ravanidis, S.; Gervois, P.; Vanheusden, M.; Marée, R.; Schrynemackers, M.; Smeets, H.J.M.; Pinxteren, J.; Gijbels, K.; Walbers, S.; Mays, R.W.; Deans, R.; Van Den Bosch, L.; Stinissen, P.; Lambrichts, I.; Gyselaers, W.; Hellings, N. Human Wharton's Jelly-Derived Stem Cells Display a Distinct Immunomodulatory and Proregenerative Transcriptional Signature Compared to Bone Marrow-Derived Stem Cells. Stem Cells Dev. 2018, 27, 65–84. [Google Scholar] [CrossRef] [PubMed]
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular matrix assembly: a multiscale deconstruction. Nat Rev Mol Cell Biol. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Tahimic, C.G.T.; Wang, Y.; Bikle, D.D. Anabolic effects of IGF-1signaling on the skeleton. Frontiers in Endocrinology 2013, 4, 6. [Google Scholar] [CrossRef]
- Ochieng, J.; Furtak, V.; Lukyanov, P. Extracellular functions of galectin-3. Glycoconj J. 2002, 19, 527–535. [Google Scholar] [CrossRef]
- James, A.W. Review of signaling pathways governing MSC osteogenic and adipogenic differentiation. Scientifica 2013, 684736. [Google Scholar] [CrossRef]
- Shizas, N.P.; Zafeiris, C.; Neri, A.-A.; Anastasopoulos, P.P.; Papaioannou, N.A.; Dontas, I.A. Inhibition versus activation of canonical Wnt-signaling, to promote chondrogenic differentiation of Mesenchymal Stem Cells. A review. Orthopedic Reviews 2021, 13, 1. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, J.; Chen, Q.; Yan, H.; Du, H.; Luo, W. Regulation of pathophysiological and tissue regenerative functions of MSCs mediated via the WNT signaling pathway (Review). Molecular Medecine Reports 2021, 24, 648. [Google Scholar] [CrossRef]
- Toh, W.S.; Lai, R.C.; Zhang, B.; Lim, S.K. MSC exosome works through a protein-based mechanism of action. Biochemical Society Transactions 2018, 46, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Teo, K.Y.W.; Chuah, S.J.; Lai, R.C.; Lim, S.K.; Toh, W.S. MSC exosomes alleviate temporomandibular joint osteoarthritis by attenuating inflammation and restoring matrix homeostasis. Biomaterials 2019, 200, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Liu, Y.; Yang, Y.; Wang, H.; Xu, Y.; Zhang, Z. MSC-Derived Exosomes-Based Therapy for Peripheral Nerve Injury: A Novel Therapeutic Strategy. BioMed Research International 2019, 6458237. [Google Scholar] [CrossRef] [PubMed]
- De Miguel, M.P.; Fuentes-Julián, S.; Blázquez-Martínez, A.; Pascual, C.Y.; Aller, M.A.; Arias, J.; Arnalich-Montiel, F. Immunosuppressive Properties of Mesenchymal Stem Cells: Advances and Application. Current Molecular Medicine 2012, 12, 574–591. [Google Scholar] [CrossRef]
- Konala, V.B.R.; Bhonde, R.; Pal, R. Secretome studies of mesenchymal stromal cells (MSCs) isolated from three tissue sources reveal subtle differences in potency. In Vitro Cellular & Developmental Biology - Animal 2020, 56, 689–700. [Google Scholar] [CrossRef]
- The, M.; Maccoss, M.; Noble, W.; Käll, L. Fast and AccurateProtein False Discovery Rates on Large-Scale Proteomics Data Sets with Percolator 3.0. Journal of the American Society for Mass Spectrometry 2016, 27, 1719–1727. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D. J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; Pérez, E.; Uszkoreit, J.; Pfeuffer, J.; Sachsenberg, T.; Yılmaz, ¸ S.; Tiwary, S.; Cox, J.; Audain, E.; Walzer, M.
- Jarnuczak, A.F.; Ternent, T.; Brazma, A.; Vizcaíno, J.A. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar]
Figure 1.
Biological process classification of differentially expressed proteins.
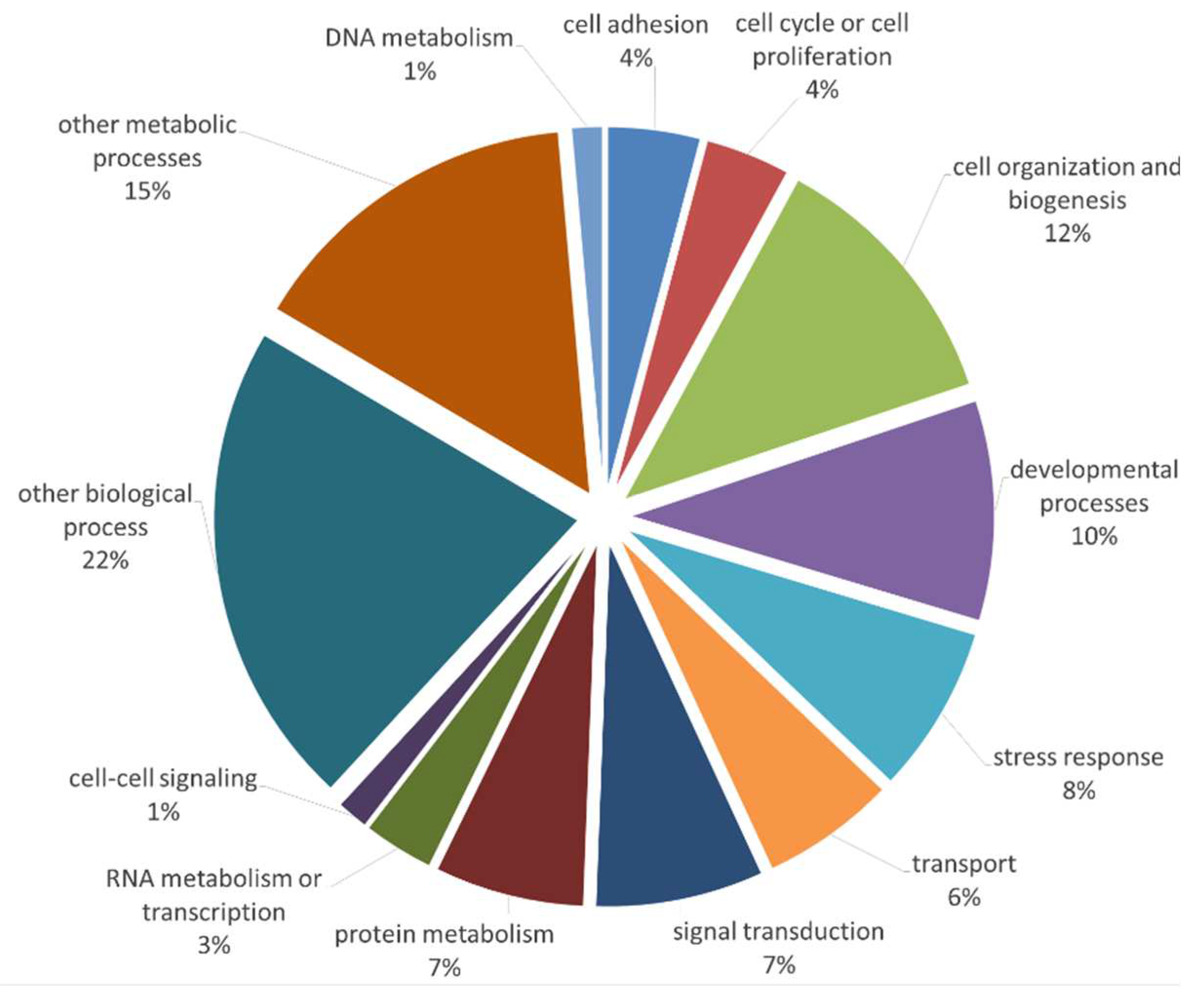
Figure 2.
(a) Venn diagram of differentially expressed proteins, (b) : GO classification of differentially expressed proteins using pairwise comparison: BM vs. DP, BM vs. WJ and DP vs. WJ.
Figure 2.
(a) Venn diagram of differentially expressed proteins, (b) : GO classification of differentially expressed proteins using pairwise comparison: BM vs. DP, BM vs. WJ and DP vs. WJ.
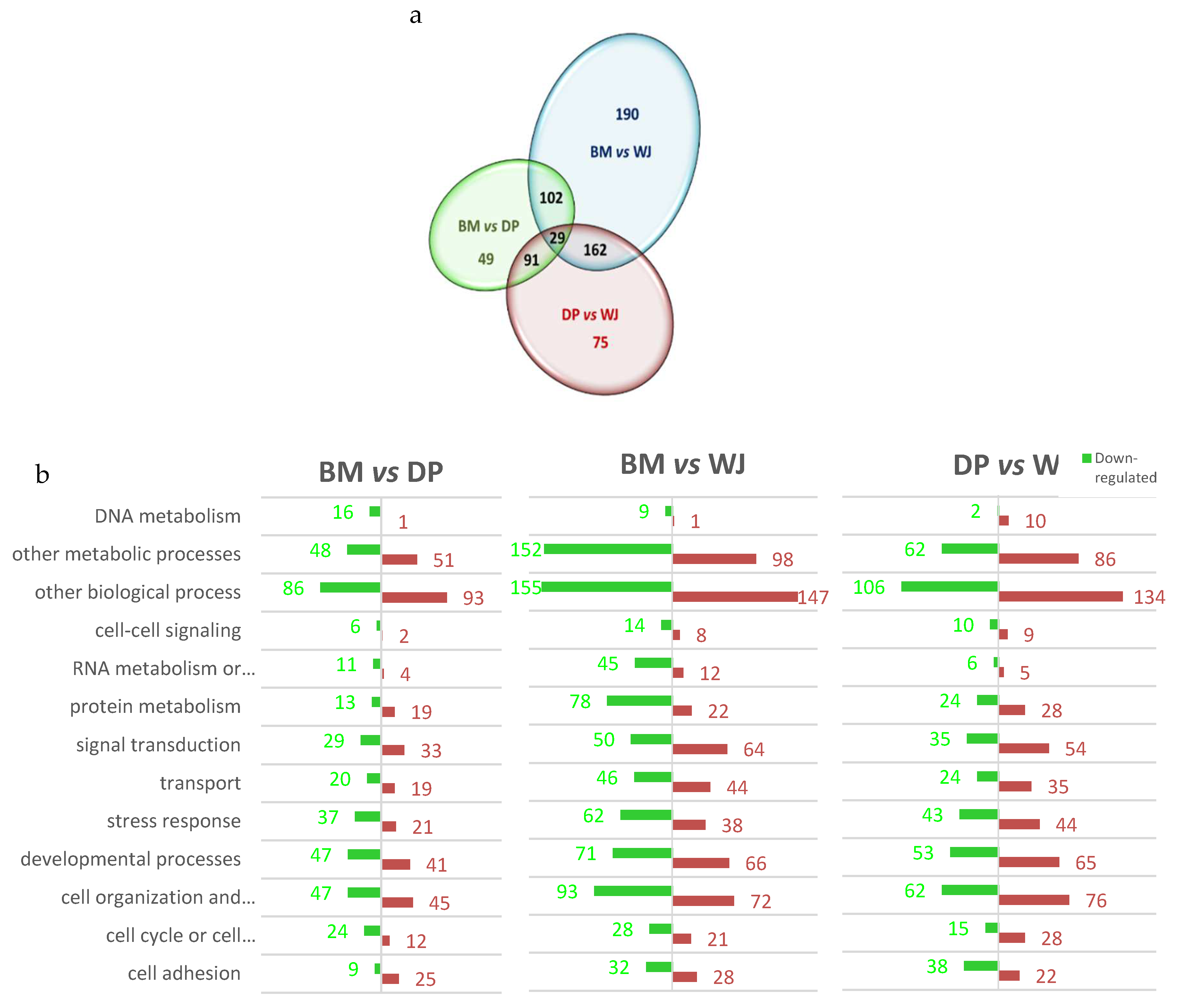
Figure 3.
Heatmaps of differentially expressed proteins in the following categories: collagens, proteoglycans or extracellular matrix glycoproteins, using pairwise comparison: BM/DP, BM/WJ and DP/WJ. Color coding in the heatmap depicts the variation between the maximum (coded in red tones) to minimum (coded in green tones) observed of Log10FC and statistically insignificant results are color-coded white.
Figure 3.
Heatmaps of differentially expressed proteins in the following categories: collagens, proteoglycans or extracellular matrix glycoproteins, using pairwise comparison: BM/DP, BM/WJ and DP/WJ. Color coding in the heatmap depicts the variation between the maximum (coded in red tones) to minimum (coded in green tones) observed of Log10FC and statistically insignificant results are color-coded white.

Figure 4.
Heatmaps of differentially expressed proteins in the following categories: ECM regulators and ECM affiliated proteins, using pairwise comparison: BM/DP, BM/WJ and DP/WJ. Color coding in the heatmap depicts the variation between the maximum (coded in red tones) to minimum (coded in green tones) observed of Log10FC and statistically insignificant results are color-coded white.
Figure 4.
Heatmaps of differentially expressed proteins in the following categories: ECM regulators and ECM affiliated proteins, using pairwise comparison: BM/DP, BM/WJ and DP/WJ. Color coding in the heatmap depicts the variation between the maximum (coded in red tones) to minimum (coded in green tones) observed of Log10FC and statistically insignificant results are color-coded white.
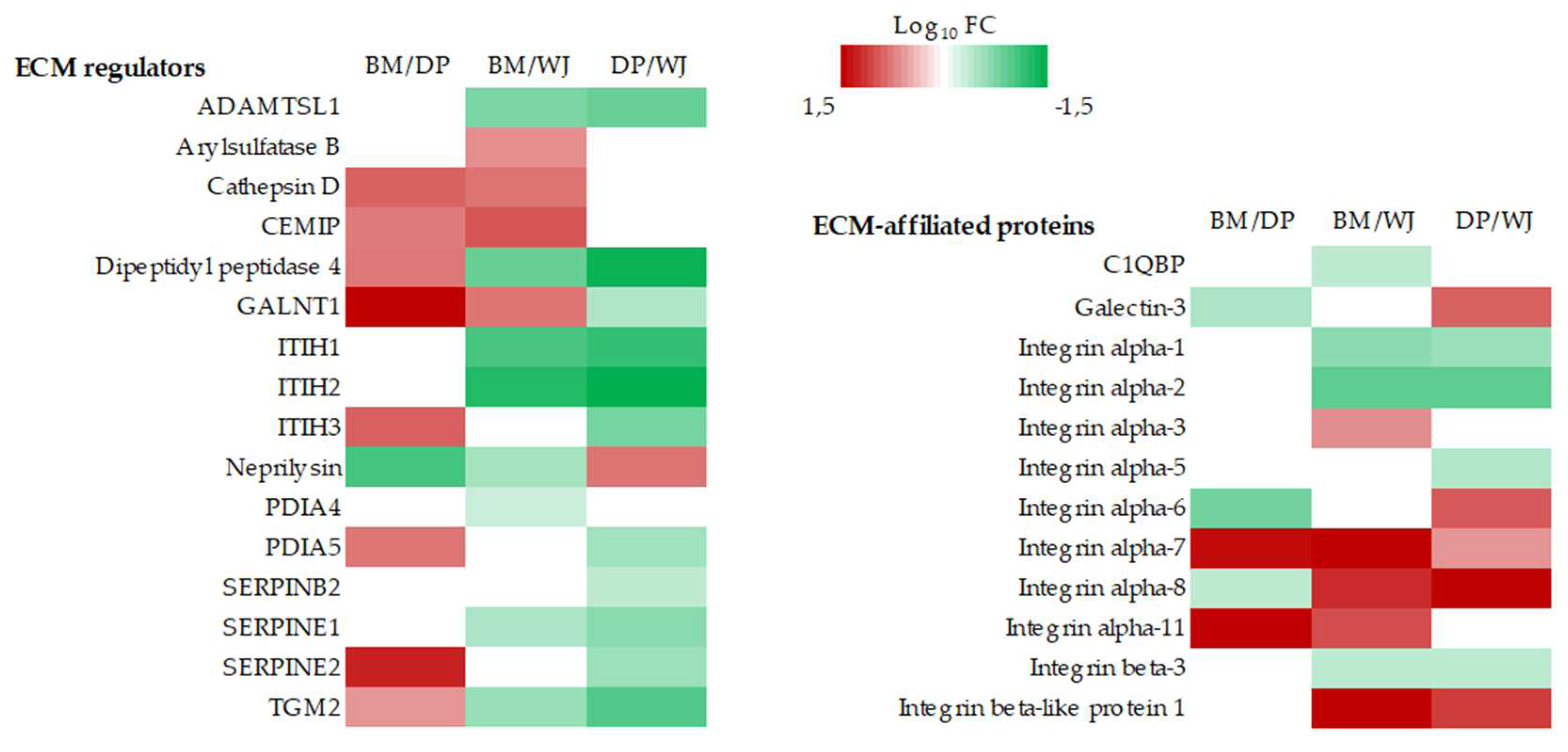
Figure 5.
Proteins involved in cell-cell signaling significantly underexpressed (Log10FC negative) or overexpressed (Log10FC positive) according to BM/DP, BM/WJ and DP/WJ ratios. Only statistically significant values are shown.
Figure 5.
Proteins involved in cell-cell signaling significantly underexpressed (Log10FC negative) or overexpressed (Log10FC positive) according to BM/DP, BM/WJ and DP/WJ ratios. Only statistically significant values are shown.
Figure 6.
Heatmaps of differentially expressed proteins: Inflammatory Response and Immune Response using pairwise comparison: BM/DP, BM/WJ and DP/WJ. Color coding in the heatmap depicts the variation between the maximum (coded in red tones) to minimum (coded in green tones) observed of Log10FC and statistically insignificant results are color-coded white.
Figure 6.
Heatmaps of differentially expressed proteins: Inflammatory Response and Immune Response using pairwise comparison: BM/DP, BM/WJ and DP/WJ. Color coding in the heatmap depicts the variation between the maximum (coded in red tones) to minimum (coded in green tones) observed of Log10FC and statistically insignificant results are color-coded white.
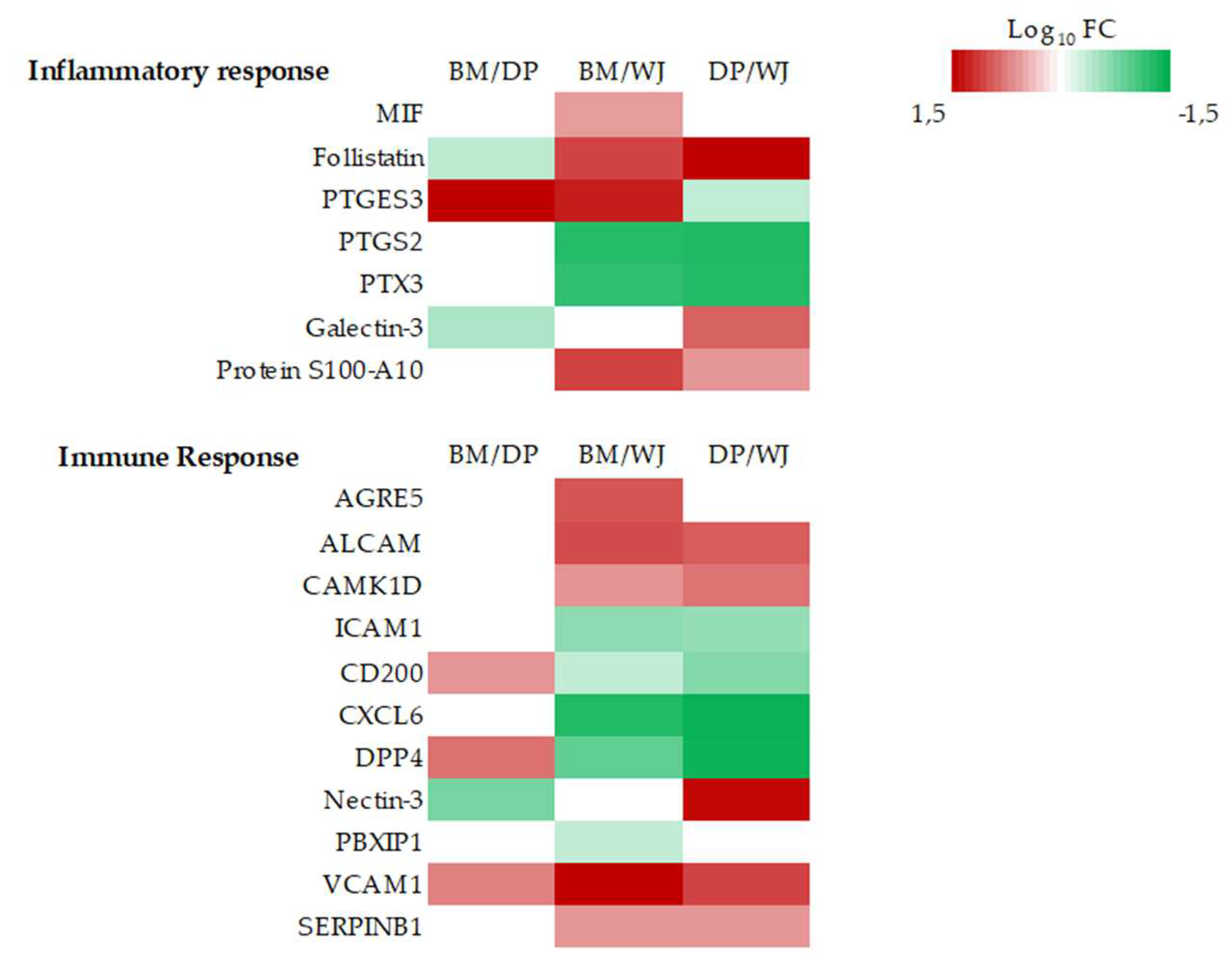

Table 1.
CD proteins differentially expressed in MSCs sources. Only statistically significant values are shown.
Table 1.
CD proteins differentially expressed in MSCs sources. Only statistically significant values are shown.
| Protein Name | Ratio | Most expressed by | Least expressed by | ||
| BM/DP | BM/WJ | DP/WJ | |||
| CD105: Endoglin | 2.842 | 2.542 | - | BM | |
| CD106: VCAM1 | 2.419 | 9.136 | 3.776 | BM | WJ |
| CD146: MCAM | 2.689 | 2.477 | - | BM | |
| CD166: ALCAM | - | 3.567 | 3.145 | WJ | |
Table 2.
Differentially expressed proteins in MSCs playing a role in cell differentiation. Only statistically significant values are shown.
Table 2.
Differentially expressed proteins in MSCs playing a role in cell differentiation. Only statistically significant values are shown.
| Protein Name | Gene Symbol | Ratio BM/DP | Ratio BM/WJ | Ration DP/WJ | Role of Protein |
|---|---|---|---|---|---|
| Osteoblast differentiations | |||||
| Alkaline phosphatase, tissue-nonspecific isozyme | ALPL | - | 5.508 | 4.441 | Promotes calcification |
| Transforming growth factor beta-1 proprotein | TGFB1 | - | - | 0.452 | TGF-β/BMP pathway controls the differentiation of mesenchymal precursor cells |
| Fibronectin | FN1 | - | - | 0.206 | Marker of osteoblast maturation |
| Adipocyte differentiations | |||||
| Prostaglandin G/H synthase 2 | PTGS2 | - | 0.077 | 0.073 | Suppressor of adipocytic differentiation |
| Adipogenesis regulatory factor | ADIRF | 3.25 | 5.483 | - | transcriptional regulator of white adipocyte differentiation |
| Neuronal differentiations | |||||
| Glia-derived nexin | SERPINE2 | 6.058 | - | 0.269 | Promotes neurite extension by inhibiting thrombin |
| Dihydropyrimidinase-related protein 2 | DPYSL2 | - | 2.706 | - | Involved in the regulation of axon formation during neuronal polarization as well as in axon growth and guidance |
| Echinoderm microtubule-associated protein-like 1 | EML1 | - | 2.598 | 4.279 | Required for normal proliferation of neuronal progenitor cells |
| Neuronal growth regulator 1 | NEGR1 | 2.263 | - | - | May function as a trans-neural growth-promoting factor in regenerative axon sprouting |
| Other differentiations | |||||
| Actin2, aortic smooth muscle | ACTA2 | 2.068 | - | - | Involved in vascular contractility and blood pressure homeostasis |
| Transgelin | TAGLN | 3.061 | 2.052 | - | Ubiquitously expressed in vascular and visceral smooth muscle and is an early marker of smooth muscle differentiation |
| Caldesmon | CALD1 | 2.139 | 2.48 | - | Regulated actomyosin interactions in smooth muscle and non-muscle cells Involved in Schwann cell migration during peripheral nerve regeneration |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



