
Submitted:
18 May 2023
Posted:
19 May 2023
You are already at the latest version
Abstract
Red beetroot, is a well-recognized and established source of bioactives (e.g., betalains and polyphenols) with anti-inflammatory and antimicrobial properties. It is proposed as a potential alternative to zinc oxide, with a focus on gut microbiota modulation and metabolite production. In this study, weaned pigs aged 28-days were fed either a control diet, diet supplemented with zinc oxide (3,000 mg/kg), or 2% and 4% pulverized whole red beetroot (CON, ZNO, RB2 and RB4; respectively) for 14 days. After the pigs were euthanized, blood and digesta samples were collected for microbial composition and metabolite analyses. Results showed, red beetroot supplemented diet at 2% improved the gut microbial richness relative to other diets, but marginally influenced the caecal microbial diversity compared to zinc oxide supplemented diet. Further increase in red beetroot levels (4% -RB4) lead to loss of caecal diversity, decreased short chain fatty acids and secondary bile acid concentrations. An increased Proteobacteria abundance, presumably due to increased lactate/lactic acid producing bacteria was also observed. Summarily, red beetroot contains several components conceived to improve the gut microbiota and metabolite output of weaned pigs. Future studies investigating individual components in red beetroot will better elucidate their contributions to gut microbiota modulation and pig health.
Keywords:
weaned pig
; gut microbiota
; red beetroot
; short chain fatty acids
; bile acids
1. Introduction
Weaning is a stressful phase in pig production, characterized by reduced feed intake, poor growth rate, gut microbiota disruption and diarrhoea [1,2]. It is a transitionary phase of the pig life, associated with compositional and functional alterations of the gut microbiota, resulting in enteric infections. Several measures to prevent dire economic losses at weaning in pig production are currently being explored. Diet provided to weaned pigs have been demonstrated to modulate significant gut microbiota changes leading to increased population of beneficial bacteria species with remarkable changes observed 10 to 14 days post-weaning [3,4].
Pathogenic colonization of the gut, a leading cause of diarrhea and death of young pigs at weaning is thus avoidable via dietary modulation of a healthy gut microbial composition [5,6]. Notably, the composition and diversity of the gut microbiota can be altered by the source and level of protein and fibre, causing increased or depleted gut microbiota metabolite production and corresponding biological responses [7,8,9]. The gut microbiota metabolite levels in return enhance or inhibit the growth of certain bacteria phyla (e.g., Bacteroidetes, Firmicutes and Proteobacteria) in the gastrointestinal tract [10,11].
Similarly, in-feed antibiotics and zinc oxide (ZnO) have been reported to reduce piglet mortality during weaning [12,13]. Their capacity to suppress post weaning diarrhoea, alter host-gut microbiota metabolism, improve feed intake and energy production for growth is well known [14,15,16]. However, despite these advantages, they have been found to destabilize the gut microbial diversity, alter short chain fatty acid (SCFA) levels and support the emergence of harmful and antibiotic-resistant bacteria species [17,18,19].
Evident from past literature, gut microbiota changes to in-feed ZnO have been characterized with increased coliforms [20,21], reduced anaerobic and lactic acid bacteria [22,23] and reduced commensal bacteria population [24,25]. Consequently, the functional potential of the gut microbiota and production of health promoting metabolite (e.g., short chain fatty acids and bile acids) may be compromised.
Additionally, there are concerns about severe environmental pollution from high fecal excretions of zinc [26] linked to in-feed pharmacological doses of zinc oxide, coupled with increasing trends of multidrug-resistant E. coli [27,28]. This signifies a risk to the animal-environment food chain and highlights an urgent need for alternatives to in-feed ZnO. Therefore, there has been an increased research interest in plants containing bioactive compounds e.g., red beetroot with health promoting properties, as possible replacement for in-feed ZnO with emphasis on modulating a healthy gut microbiota thus preventing pathogenic colonization post-weaning [29,30].
Red beetroot (Beta vulgaris subsp. vulgaris conditiva) contains bioactive compounds such as betalains, polyphenols, inorganic nitrate (NO3), fibre and minerals (e.g., potassium, sodium, phosphorus, calcium, magnesium, copper, iron, zinc, and manganese) [31,32,33]. These bioactives (i.e., betalains, polyphenols, nitrate and fibre) have been indicated in the potential prebiotic effect of red beetroot, driving gut microbiota modulation and metabolite production, with impact on host metabolism, physiology and immune functions [34,35,36].
Red beetroot is one of the top ten plants with high antioxidant, anti-inflammatory, antimicrobial, anticarcinogenic and hepatoprotective characteristics [37,38]. It is currently being considered as a therapeutic ingredient, in the treatment of conditions caused by oxidative stress, inflammation and metabolic disorders (e.g., hypertension, diabetes, insulin resistance and kidney dysfunction) [39,40,41]. The health benefits of red beetroot in humans, rodents [42,43] and rainbow trout [44,45] have been widely studied and reported in literature, but studies using pigs have not been considered, and research demonstrating the potential of red beetroot supplementation on the gut microbiota is yet lacking. This study therefore examines the effect of red beetroot on the gut microbiota composition and metabolite output of weaned pigs.
2. Materials and Methods
The animal trial was conducted at the National Pig Centre, UK under an ethical approval granted by the University of Leeds Animal Welfare and Ethical Review Committee (AWERC) with the approval number; 070510HM. All husbandry practices were set by the farm in accordance with the Welfare of Farmed Animals, England Regulation 2007, and all procedures were conducted following the amended Animals (Scientific Procedures) Act 1986. For ethical reasons, the number of piglets per treatment was reduced and determined based on previous studies [46,47,48] focusing on gut microbiota diversity. The basal diet was provided by Primary diets, UK and whole red beetroot powder was purchased from Buy Wholefoods online Ltd (Ramsgate, UK). Reference bile salts for bile acid quantification were purchased from Sigma-Aldrich (Steinheim, Germany) and Cayman (Cambridge, UK). A mixed short chain fatty acid standard solution containing acetate, propionate, butyrate, valerate, isobutyrate and isovalerate was obtained from Supelco - Merck life science Ltd (Dorset, UK). Chemicals, solvents, and other reagents were purchased from Sigma-Aldrich (Germany) and Fischer Scientific (Loughborough, UK) accordingly.
2.1. Experimental animals and experimental design
Forty-eight piglets (Large White X Landrace X Duroc) weaned on day-28 (average body weight: 7.58±0.69 kg) were randomly allocated to one of four diet (n = 12), balancing for body- weight, sex, and litter origin, for a 14-day feeding experiment. The pigs were housed in a temperature-controlled flat deck with open feed troughs and nipple drinkers allowing easy access to feed and water ad libitum. The experimental diets comprised a basal control diet (CON) and diet supplemented with 3000 mg/kg zinc oxide (ZNO), both formulated according to the National Research Council (2012) recommendations (Table 1). Red beetroot supplemented diets (RB2 and RB4) were obtained by adding 2% (20 g/kg) and 4% (40 g/kg) pulverized whole red beetroot to the basal diet respectively, thoroughly mixed with an electric mixer on-site.
2.2. Sample collection
At the end of the experiment period, eight animals per diet (n = 8) were euthanized by captive bolt and exsanguination. Blood samples were collected from the jugular vein into heparinized tubes, from which plasma was obtained after centrifugation at 2000 x g, 4°C for 10 min. Fecal samples were collected from the rectum into designated tubes. The abdominal cavity was immediately opened, each intestinal segment (duodenum, jejunum, ileum, caecum, and colon) was identified, separately cut, and emptied into a sterile beaker. Digesta from each segment was mixed, aliquoted into sterile 2 mL Eppendorf tubes. All samples were snapped frozen in liquid nitrogen and stored at -80℃ for analysis of gut bacterial composition, short chain fatty acids and bile acids.
2.3. Gut microbiota analyses and bioinformatics
Pig gut microbial composition was examined using digesta samples from the jejunum, ileum, and caecum. Genomic DNA was extracted from (approx. 1.0 g) digesta sample with QIAamp Power fecal DNA kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The concentration and purity of DNA samples were measured spectrophotometrically with Nano Drop® ND-1000 (Nano Drop Technologies Inc., Dover, USA), absorbance ratio at 260/280 nm observed were within the range of 1.8 - 2.0. DNA samples were submitted to the University of Leeds, Next Generation Sequencing facility, St. James Hospital Leeds, UK, for quality screening, 16S rRNA gene library preparation and sequencing. According to a previous study [49], the V4 hypervariable region of the 16S rRNA gene was amplified in a two-step polymerase chain reaction (PCR) with specific primers (564F, 806R) and Illumina adaptor overhang. Following the Illumina 16S metagenomics sequencing library preparation protocol, the final libraries were pooled, and pair-end sequenced on the Illumina MiSeq platform (Illumina, San Diego, CA, USA).
Sequence reads were processed in Mothur v.1.43.0 with the MiSeq standard operation procedure developed by the Schloss group [50,51]. Chimera-free and unique sequences were identified and aligned to the SILVA (v.138) database, then sequences with 97% similarity were clustered into operational taxonomic unit. The “Biome” file generated from Mothur was transferred to R (v 3.6.2 and 4.0.0) environment for further analyses including the alpha and beta diversity indices.
The alpha diversity of the gut microbial community was evaluated using Chao1, Shannon and Simpson indices, variables were compared using ANOVA evaluating the effect of diet, gut location, and their interaction with the lmerTest (linear mixed effects). Differences between gut samples (beta diversity) were determined by a permutational multivariate analysis of variance (PERMANOVA) of the non-phylogenetic distance matrix (Bray Curtis) and visualized on a non-metric multidimensional scale (NMDS) plot. The diet effect on each gut location was computed by a paired comparison of distance matrices with pairwise Adonis function (adonis2) in vegan package (v. 2.6.4).
Differentially (distinct) abundant taxa between gut locations per diet was identified in a two-sided Welch’s t-test and Benjamin Hochberg false discovery ratio (FDR) correction in STAMP (Statistical Analysis of Metagenomics and other Profiles) software [52]. Further analysis employed DESeq2 (v. 1.27.32) in R [53] with Wald hypothesis testing for distinct genera in each gut location comparing multiple diet groups. Differences were estimated as fold change (Log 2- fold change) between diet and with FDR-corrected p values.
2.4. Predicted functions of pig gut microbiota
To predict the functional pathways mediated by the gut microbiota, OTU abundance and representative sequences processed in Mothur were submitted to Piphillin (https://piphillin.secondgenome.com/). Gene sequences were matched against KEGG (Kyoto Encyclopedia of Genes and Genomes) database as described in Iwai, et al. [54], using USEARCH version 8.0.1623 with global alignment setting for sequence identification fixed to 90% cut-off (a level significantly associated with PICRUSt - phylogenetic investigation of communities by reconstruction of unobserved states) [55].
Pathways differentially mediated by diet in the different gut locations were computed with DESeq2 in R using Wald test and p values adjusted for multiple inter-diet comparisons.
2.5. Quantification of short chain fatty acids (SCFA) and bile acids
Short chain fatty acids (acetate, propionate, butyrate, valerate, isobutyrate and isovalerate) in plasma, jejunum, ileum, caecum, colon digesta and fecal samples were determined with gas chromatography (Varian 3400; Varian Ltd., Oxford, UK). Method used was as described in Taylor, et al. [56] with slight modification. Briefly, 1.0 g or 1 mL sample was mixed with an equal volume of distilled water in an Eppendorf tube and centrifuged at 12,000 x g, at 4°C for 10 min. Phosphoric acid (50 µL, 85% v/v) was added to the supernatant (500 µL) collected and 150 µL caproic acid (150 mM/L) as internal standard. The mixture was made up to 1 mL with distilled water, centrifuged at 14,000 x g for 20 min, after which the supernatant was collected for SCFA analyses. Individual short chain fatty acid in processed sample was quantified using a standard curve obtained from a mixed volatile fatty acid prepared in the concentration range 0 to 125 mM.
Bile acids in samples were determined as described in Zhang, et al. [57], briefly, 0.3 g digesta was mixed with acetonitrile (final conc. 80% v/w), incubated for 20 min at room temperature and centrifuged at 15,000 x g at 4°C for 20 min. The supernatant collected was passed through Strata-X 33μm polymer based solid phase extraction cartridges (Phenomenex, Torrance USA) after cartridges had been conditioned with methanol and water. Subsequently, bile acids were eluted in 1.5 mL methanol, concentrated, and dried using a solvent evaporator (SP Genevac EZ-2 Series, Pennsylvania, USA), then reconstituted in 150 μL methanol before subjecting to HPLC-MS (Shimadzu, Kyoto, Japan). The mobile phase: A and B was a mixture of 5 mM ammonium acetate in water and methanol respectively, both acidified with 0.012% formic acid. A mixed standard reference (0 - 0.1 mM) containing; taurohyodeoxycholic acid (THCA), glycohyodeoxycholic acid (GHDCA), taurocholic acid (TCA), glycocholic acid (GCA), taurochenodeoxycholic acid (TCDCA), taurodeoxycholic acid (TDCA), glycochenodeoxycholic acid (GCDCA), glycodeoxycholic acid (GDCA), cholic acid (CA), glycolithocholic acid (GLTHCA), chenodeoxycholic acid (CDCA), deoxycholic acid (DCA) and lithocholic acid, was prepared for quantification of bile salts.
2.6. Statistical analyses
SCFA and bile acid concentrations were analyzed in R environment (v. 4.2.2), zero inflated data was analyzed using negative binomial with square root link function, multiple comparison of means was computed using Tukey post hoc test with significance level of p < 0.05. Results were expressed as mean and standard error of mean (SEM) and presented in tables. A Spearman correlation analysis was conducted between SCFA levels, bile acids and the mean relative abundance of top twenty-five genera in each gut location per diet using the “Psych” [58] and “Pheatmap” [59] packages in R (v. 3.31).
3. Results
3.1. Effect of diets on gut microbial diversity and taxonomic composition
In the present study, there was no difference in the growth performance of the pigs, however, the diets (P = 0.01) significantly influenced the species richness and diversity of the gut microbiota with respect to the gut locations (P < 0.001; jejunum, ileum, and caecum) examined. Diet RB2 increased the jejunal species richness (P = 0.02) compared to other diets according to the Chao1 index, but caecal species abundance was comparable for all the diets (Figure 1a). From the Shannon index of alpha diversity, the gut microbial community was diverse, but this was not influenced by the diets (P = 0.07), howbeit a pairwise diet comparison showed, ZNO diet was different from CON and RB4 in the caecum (Figure 1b). No significant species abundance, divergence, or evenness (i.e., dominance) was observed between the diets and in the gut locations for the Simpson index (data not shown).
The beta diversity is as shown in the non-metric multi-dimensional scaling (NMDS) plots (Figure 2a,b). Samples from the caecum clustered distinctively away from the ileum and jejunum, depicting the caecum had a significantly different (P = 0.013) microbial composition from the ileum and jejunum, while the ileum and jejunum microbial community are marginally different (P = 0.051). A subset analysis of the caecal biome with inter-diet comparisons indicated CON pigs had more similar species in the caecum than ZNO (P = 0.03) and RB2 (P = 0.05) pigs but related to RB4 pigs (P = 0.35). Hence, ZNO pigs contained more dissimilar species than RB4 pigs, whereas RB2 and RB4 pigs were not different (P = 0.09).
From the digesta samples analyzed, 15 phyla and 310 genera were observed with approximately 99% of total sequences (17,573,278) assigned. The mean relative phyla and genera abundance is presented in Figure 3a,b. Dominant bacteria phyla with mean relative abundance >1% were Firmicutes, Actinobacteriota, Bacteria unclassified, and Bacteroidota. The mean relative phyla and top genera abundance in the gut were compared across the diet groups and have been presented in Table 2 and S1 respectively. The gut locations mainly influenced (P < 0.05) the relative mean phyla distribution with increase in the caecum compared to other regions examined, however Firmicutes abundance reduced. An increase in phylum Actinobacteriota and Proteobacteria abundance in CON and RB4 pigs was also observed.
From the top genera (Table S1), Megasphaera, Streptococcus, Anaerovibrio, Rumminococcaceae unclassified, Erysipelotrichaceae unclassified, Bacillus unclassified, Terrisporobacter and Clostridiaceae unclassified abundance were significantly influenced by the diets (P < 0.05) and the gut locations (P < 0.02) examined. The mean relative abundance of eleven genera (e.g., Megasphaera, Selenomonadaceae unclassified, Phascolarctobacterium, Firmicutes unclassified, Bacteria unclassified, Erysipelotrichaceae unclassified, Negativibacillus, Anaerovibrio), functionally recognized as lactate utilizing bacteria (LUB) were higher (P < 0.05) in the caecum, but comparable in the jejunum and ileum. Likewise, lower (P < 0.05) Streptococcus, Lactococcus, Lactobacillales unclassified, Streptococcaceae unclassified, and Bacilli unclassified abundance (mostly lactic acid producing bacteria –LAB) was observed in the caecum compared to the jejunum and ileum.
3.2. Differential abundant genera modulated by the diets
Differential abundance at the genus level and distribution per diet was computed by Welch’s t-test and FDR-corrected in STAMP. The result indicated an increased (P < 0.05) Megasphaera, Selenomonadaceae_unclassified and Veillonellaceae_unclassified abundance in pigs fed CON diet, Erysipelotrichaceae_unclassified, Clostridiaceae_unclassified and Rumminococcaceae_unclassified in ZNO pigs, while Bacilli_unclassified and Anaerovibrio increased in RB2 and RB4 pigs respectively. Further analyses of each gut location with inter-diet comparisons as shown in Figure 4, presented decrease in Veillonellaceae_unclassified and Selenomonadaceae_unclassified abundance in the jejunum of RB2 and RB4 pigs, compared to CON and ZNO pigs, whereas in the ileum, only RB2 pigs showed an increased Terrisporobacter abundance relative to CON pigs. The caecum was enriched with nine genera (e.g., Romboutsia, Clostridiaceae_unclassified, Terrisporobacter, Candidatus_Soleaferrea, Muribaculaceae_ge and Clostridium_sensu_stricto_1) in ZNO pigs compared to CON but diminished in genus Selenomonadaceae_unclassified. Pigs fed red beetroot diets (RB2, RB4) had increased caecal Selenomonadaceae_unclassified and/or Anaerovibrio abundance relative to ZNO pigs.
3.3. Metabolite profile and association with gut microbial composition
Short chain fatty acid (SCFA) profile followed the expected pattern of increased levels in the lower gut (caecum and colon), including the fecal samples. Nonetheless, the experimental diets influenced SCFA levels observed in these locations (Table 3). Concomitant with the species richness in the gut locations, SCFA levels increased significantly in the jejunum of RB2 pigs, reduced in the ileum of ZNO pigs, but was comparable in the caecum across the diet groups. Overall, total SCFA levels reduced significantly (P = 0.01) in RB4 and ZNO pigs, as was for most SCFA (e.g., acetate, propionate, and butyrate).
Similarly, the trend of high jejunal bile acid concentration (approx. 1 to 3-fold) and levels observed in other locations examined is not biologically relevant (Table 4). Pigs fed RB2 diet had higher (P < 0.05) TCA, GCDCA, CA, GLTCA, CDCA and DCA compared to other diets, but equivalent total and unconjugated bile acids (CA, DCA, CDCA and LCA) levels with CON pigs. Conversely, bile acid concentration reduced in ZNO (TCDCA, TDCA, GCDCA, GDCA and CDCA) and RB4 (TCA, GLTCA, DCA, CA) pigs relative to CON, which cumulatively ensued lesser total and unconjugated bile acids levels.
Although, total SCFA in CON and RB2 pigs was higher, caecal SCFA levels were comparable across the diet, acetate and propionate levels correlate significantly to bacteria (e.g., Firmicutes_unclassified, Mitsuokella, Megasphaera, Streptococcus, Streptococcceae_unclassified, Anaerovibrio, Lactobacillus and Selenomonadaceae unclassified) abundance in the caecum for CON and RB pigs, but closely associated with the jejunum and ileum (e.g., Phascolarctobacterium, Bacteria_unclassified) genera abundance in ZNO and RB4 pigs. Across the gut locations, Faecalibacterium, Blautia, Clostridia_unclassified, Clostridiaceae_unclassified, Dialister, Olsenella, Selenomonadaceae_unclassified, Veillonellaceae_unclassified, and Firmicutes _unclassified are examples of genera significantly associated with butyrate in ZNO and RB4 pigs (Figure 5a,b), most of which were associated with ileal butyrate in RB2 pigs although not significant (Figure S1b).
Associations between the gut genera abundance and bile acid levels are presented in Figure S2. Focusing on the unconjugated (CA, CDCA, LCA) and conjugated (GCA, TCA, GDCA, GCDCA, TCDCA, TDCA, GLTCA, THCA, GHDCA) bile acids, in the jejunum, CA, CDCA and total bile acid levels were strongly associated with most genera in RB2, unlike ZNO and RB4 pigs (Figure 6). Conjugated bile acids were significantly associated with the ilea genera (e.g., Bacteria_unclassified, Selenomonadaceae_unclassified and Veillonellaceae_unclassified) abundance in the CON and RB pigs, however most bacteria (e.g., Lactobacillus, Lactococcus, Streptococcaceae_unclassified, Firmicutes_unclassified, Terrisporobacter, RF39_ge) were associated with, unconjugated, conjugated, and total bile acids in the ileum of ZNO pigs. A significant correlation was observed in the caecum between the bacteria genera (Dialister, Streptococcus, Lactobacillaes_unclassified, Streptococcaceae_unclassified, Lactococcus, and Selenomonadaceae_unclassified) and unconjugated bile acids (CA, CDCA) in CON, RB2 and RB4 pigs but not in ZNO pigs.
3.4. Predicted gut microbiota functions
Compared to ZNO pigs, pathways enabling bacteria response and adaptation to environmental changes (e.g., biofilm formation, flagella assembly and two-component system) were upregulated in the caecum of pigs on CON diet. Also, pigs fed RB4 diet had pathways influencing lipid metabolism (inositol phosphate, glycerol-phospholipid, fatty acid degradation, chloroalkane and chloroalkene degradation) enhanced. Aside from these, there were no variations between the diets in the functional pathway predictions from the jejunal and ilea microbiota.
4. Discussion
Current reports on the health potential of red beetroot have necessitated evaluations of its probable effect on the gut microbiota and as an alternative to zinc oxide in weaned pig diet. Red beetroot, a rich source of nutrient, fibre, and bioactive compounds is well recognized for its anti-inflammatory, antioxidant, antimicrobial properties, and prebiotic effect in the gut. Given these benefits, adding red beetroot to weaned pig diet could promote beneficial microbiota modulation of the gut, thus prevent gut dysbiosis and diarrhea post weaning.
Diet, remains an uncontestable factor shaping and modulating the gut microbiota, towards the achievement of gut health and overall wellbeing [60]. Capper, et al. [61] in a controlled clinical trial with healthy humans consuming whole cooked red beetroot, showed gut microbiota modulation with reduced Bacteroidetes, increased alpha diversity and short chain fatty acid (SCFA) levels combined with a normal fecal score. The weaning phase in pig production is of focus here due to the attending economic impact, more so the recent ban on in-feed ZnO which has further exacerbated the health implication on weaned pigs.
Studies on pig gut microbiome have long established that early modulation of the gut microbiota of young pigs is vital to the prevention of post weaning diarrhoea, maturation of the immune system and improvement of growth performance [2,62,63]. Adaptation of the weaned pig gut to a new diet and achievement of a relatively stable gut microbiota 7 to 10 days post weaning is essential for early attainment of a rich-diverse gut microbial composition and gut health [64].
In this present study, 14-day supplementation of weaned pig diet with 2% red beetroot (RB2) influenced the alpha diversity, increasing the species richness of the jejunum compared to other diets. Although, a comparable number of species was observed in the caecum of all the pigs. Reduced bacteria species in the jejunum [65,66] or ilea digesta [67] of weaned pigs fed diet containing ZnO have been linked to the antimicrobial and growth-promoting ability of in-feed ZnO. According to Bonetti, Tugnoli, Piva and Grilli [25], in-feed ZnO reduces gut bacteria activity, making available more energy for growth and metabolism.
The caecum had a rich and more diverse bacteria as supposed, whereas the jejunal and ileal microbiota were closely related. This certifies the existence of more unique taxa in the caecum compared to the jejunum and the ileum [68]. Although, RB2 diet compares with ZNO, the latter clearly modulated a diverse caecal microbiota with more distinct bacteria than CON and RB4. This was possibly driven by decreased caecal Firmicutes abundance, causing increased relative mean abundance of other phyla (e.g., Bacteroidota). To the best of our knowledge, this is the first report on supplementation of red beetroot in weaned pig diet, however observations from ZNO diet resonate with previous findings on pharmacological dose of ZnO in weaned pig diet [66], while RB2 diet improved the species richness of the gut.
Meanwhile, increased RB levels did not translate to a diverse caecal microbiota despite increased fibre levels, depicting the gut microbiota acted differently toward the fibre. Besides, the functions of dietary fibre in the gut are largely determined by its source and physicochemical characteristics (e.g., solubility, viscosity, fermentability), which subsequently impacts the gut microbial composition and metabolite output [69]. In this study, pigs fed RB4 had a significant increase in Proteobacteria, reduced SCFA and secondary bile acids. Bacteria in this phylum tend to increase during weaning stress and in pigs on diet rich in protein, fat, and fibre, consequently depleting beneficial bacteria like; Lactobacillus, Lactococcus and Bifidobacterium [70].
Consistently, Firmicutes is the most dominant phylum, accounting for < 95% of all phyla observed in the gut [71,72,73]. However, reduced caecal Firmicutes abundance observed in this current study negate reports of higher populations in the pig caecum [68,74]. Similarly, Actinobacteriota was the second predominant phylum compared to Bacteroidota, reported in most studies. While these phyla are important commensal of the gut, differences in management, experimental diets, sampling age and location used in these studies may explain the observed disparity [4,75]. However, such gut microbiota alterations have recently been attributed to lactate accumulation in the gut.
Wang, et al. [76] confirmed gut microbiota variation from lactate accumulation, where phylum Bacteroidetes and Firmicutes were replaced by Actinobacteria and Proteobacteria, with concomitant reduction in butyrate and propionate production. Proteobacteria (e.g., Campylobacter and Salmonella species) utilize lactate under microaerophilic conditions, to produce carbon dioxide and water [77] and have predominantly been linked to gut perturbations mostly associated with diarrhea.
The small intestine is usually dominated by lactic acid bacteria -LAB (e.g., Lactobacilli, Lactococcus, Streptococcus, Bifidobacterium etc.) and is responsible for lactate production through various biochemical pathways [78]. Lactate prevents the growth of pathogenic organisms by lowering the gut pH, but increased levels can be harmful, causing alterations in gut microbiota, toxicity, and pathogenic colonization of the gut. To corroborate this claim, high ilea lactate levels (mM) were observed in CON (96.76) and RB4 (76.85) pigs, but levels in ZNO (44.72) and RB2 pigs (48.83) were similar (unpublished).
It is noteworthy that the gut microbiota employs lactate utilizing bacteria - LUB (from the phylum Firmicutes) with remarkable SCFA producing ability to avert the detrimental effect of lactate accumulation, thereby stabilizing the gut microbiota [79,80]. Hence, a balance between the LAB and LUB (functional groups) in terms of production and utilization of lactate has been found necessary for gut health [77]. Prominent LUB (e.g., Megasphaera, Phascolarctobacterium, Negativibacillus and Veillonellaceae) and LAB (e.g., Streptococcus, Lactococcus, Lactobacillales unclassified and Streptococcaceae unclassified) were identified in this study. Importantly, a higher caecal relative LUB abundance but reduced LAB was observed, while the abundance of both bacteria groups was comparable in the small intestine. These genera were associated with SCFA (acetate, propionate, and butyrate) levels across the gut locations (jejunum, ileum, and caecum) as shown by the correlation matrices.
Though the small intestine is not the major site for microbiota fermentation and SCFA production, significant SCFA levels, and correlations with the jejunal and ilea microbiota were observed in RB diets. The nutritional functions of the jejunum with capacity for energy metabolism and fibre fermentation have been confirmed in many studies, while the gut microbiota metabolite impact on the jejunal immune system, barrier function and cell proliferation [81,82]. In addition, the host immune system is regulated by continuous interaction between the gut microbiota and dietary metabolites hence, reduced gut microbiota association with butyrate levels observed in this study may partly be due to host immune responses as well as lactate accumulation on the gut [83]. Moreover, a decline in bacteria sensitivity to metabolite production may have doused a strong correlation between the gut microbiota and butyrate levels in RB2 pigs, unlike in ZNO pigs. Overall, inter-individual variability in response to diet as well as variations in gut microbial composition and function cannot be ruled out.
Bile acids have also been linked to host physiology and immunity via gut microbial metabolism. Diet influences the gut microbiota composition, and bile acid levels through bile salt hydrolyzing bacteria (BSHB) species (e.g., Clostridium spp., Lactobacillus, Bifidobacterium, and Enterococcus) that possess inducible genes responsible for the conversion of primary bile acids to secondary bile acids [84,85]. However, interactions between the bile acid and the gut microbiota can be severely impaired in the event of gut dysbiosis at weaning. Song, et al. [86] observed dietary supplementation with CDCA, a natural primary bile acid in animal bile, improved growth performance and reduced diarrheal incidence in weaned pigs.
Generally, RB2 diet increased individual primary and secondary (CA, CDCA and DCA) unconjugated bile acid levels compared to CON. DCA (deoxycholic acid) was mainly observed in the colon and feces, hence not correlated with the gut microbiota abundance. Another study by Tian, et al. [87] confirmed a higher and potent antibacterial activity in unconjugated bile acids compared to their other counterpart, and the sensitivity of bile acids to Gram-positive bacteria than Gram-negative was also demonstrated. The jejunal bile acid profile was strongly associated with the jejunal microbiota of RB2 pigs unlike ZNO and RB4 pigs, but same as observed in the ileum for ZNO pigs. Most bacteria in this region (small intestine) are usually resistant to bile acids, offering protection against pathogenic invasion [88]. Reduced bile acid levels in the gut have been implicated in bacterial overgrowth and inflammation [11]. Conversely, across the diet groups in the caecum, very few genera (e.g., Streptococcus, Lactobacillales unclassified, Lactococcus, Selenomonadaceae_unclassified, and Erysipelotrichaceae_ unclassified) were (associated) involved in bacterial metabolism of the unconjugated bile acids (CA and CDCA). The reasons for reduced bile acid levels with increased red beetroot is not clear. Usually, a high fat diet increases bile acid discharge, increasing circulating bile acid levels. Alteration of secondary bile acids by dietary fibre and an increased Proteobacteria abundance with RB4 are some possible causes of this trend.
Exploring differentially abundant genera in the pigs, lactate utilizers (e.g., Veillonellaceae_ unclassified and Selenomonadaceae_unclassified) increased in the jejunum of CON pigs relative to those fed RB diets, which signifies an increased abundance of lactic acid producing bacteria LAB, and a potential for lactate accumulation in CON pigs. Terrisporobacter, an anaerobic Gram-positive bacterium in the family Peptostreptococcaceae, was differentially increased in the ileum of RB2 pigs compared to CON and associated with butyrate, GDCA and GLTCA. Other compositional differences observed in the cecum include increased gut fermenters (e.g., Romboutsia, Muribauculaceae_ge, Terrisporobacter, Clostridiaceae_unclassified) and decreased Selenomonadaceae_unclassified in ZNO pigs compared to the control. Except for an increased Selenomonadaceae_unclassified, RB2 was not different from ZNO, while RB4 had increased Anaerovibrio inclusive.
Generally, Clostridiaceae_unclassified, Rumminococcaceae_unclassified and Erysipelotrichaceae_unclassified were significantly higher in pigs fed ZNO diet, which coincides with results from [20] in pigs fed 2425 mg/kg dietary zinc. The presence of these strict anaerobes demonstrates a rapid transition of the pig gut microbiota from a (milk-based diet) suckling microbiota to a post weaning (solid-based diet containing complex compounds) microbiota. The preceding genera are linked to bile acid and SCFA production. However, increased Erysipelotrichaceae abundance has been implicated in dysbiosis-related disorders of the gut [89], and in mice post treated with broad spectrum antibiotics (e.g., gentamicin) [90].
Similarly, many studies have confirmed associations between bacteria belonging to this genus and host lipidemic profiles [91,92,93] and cholesterol metabolism [94,95]. This characteristic may be connected to the high systemic and hepatic lipid peroxidation observed in the plasma and liver tissue of pigs in this group (unpublished data), coupled with an increased tendencies for hepatic toxicity and oxidative stress in ZnO fed pigs. This additionally coincides with downregulation of pathways facilitating lipid metabolism for pigs fed ZNO diet compared to RB4.
Moreover, dietary supplementation with quercetin was reported to inhibit Erysipelotrichaceae [96]. Quercetin is a flavonoid (polyphenol) found in fruits and vegetables recognized for its health benefit and potential therapeutic effects. Polyphenol and bioactive pigments (betalains) in red beetroot may be responsible for a decreased abundance of Erysipelotrichaceae_unclassified in RB pigs.
Interestingly, red beetroot has been described effective in preventing lipid peroxidation in membrane decreasing oxidative damage [97], this aligns with observations of upregulated lipid metabolism pathways (i.e., inositol and glycerol-phospholipid metabolism) in the cecum of pigs in this group relative to ZNO pigs. An increased Anaerovibrio abundance; a strictly lipolytic bacteria known for hydrolysis of triglycerides to fatty acids, in pigs fed red beetroot supplemented diet further confirm these inferences. Overall, the predicted functional profile from the microbiota of each gut location (jejunum and ileum) did not differ from each diet, aside what has been earlier described, pathways in response to bacteria adaptation to environmental changes was enhanced by the caecal microbiota of CON pigs.
This study provides baseline outcomes of red beetroot supplementation of weaned pig diet; however, some limitations were identified. The absence of colon and fecal microbiota information hindered correlations with metabolite output. Also, whole red beetroot was used in this study, which represents a cost-effective way of providing red beetroot bioactive compounds (betalains, nitrate and polyphenols) to the pigs while also preserving its fibre content. Hence, the effect observed can only be inferred but not specific to any of these components. Future studies aimed at utilizing these components individually will improve outcomes and aid understanding of their roles in gut microbiota modulation and pig health.
5. Conclusions
Diet remains a viable strategy to modulate the gut microbiota of weaned pig, and red beetroot supplementation provides an avenue to explore its bioactives for pig gut health. From this study, weaned pig diet supplemented with red beetroot at 2% increased the species richness of the gut microbiota. However, inclinations of lactate accumulation were observed with increased RB to 4% (RB4), which was characterized by potential decline in butyrate and propionate and an increased Proteobacteria abundance. The jejunum and ileum microbial compositions were similar across the diet groups, but the cecum was diverse with ZNO diet, relative to RB2, while CON and RB4 were comparable. RB2 diet also increased the gut microbiota metabolites (SCFA and unconjugated bile acids) production in the jejunum and ileum, depicting fore gut fibre fermentation, but butyrate levels were not significantly associated with the gut microbiota as observed in ZNO and RB4. The functional pathway predictions from caecal microbiota were closely associated with the distinct bacteria present in the caecum of the pigs across the diets. Put together, red beetroot has the potential to modulate the gut microbiota of weaned pigs with increased species richness, enhanced lipid metabolism and metabolite production. Future work focused on purified red beetroot components and dosage in weaned pigs are warranted.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Comparison of mean relative abundance of top bacteria genera between diet groups and gut location. Figure S1: Spearman correlation matrices between top abundant bacteria genera and short chain fatty acids in the (a) jejunum (b) ileum. Figure S2: Spearman correlation matrices between top abundant bacteria genera and bile acid levels in the (a) ileum (b) caecum.
Author Contributions
Conceptualization, OOA, HM, CB and ARM.; validation, ARM, CB and KMcD.; formal analysis, OOA, KMcD and CB.; investigation, OOA.; resources, CB and ARM.; data curation OOA, KMcD and CB.; writing—original draft preparation, OOA.; writing—review and editing, OOA, CB, ARM, KMcD and HGH.; visualization, OOA and KMcD.; supervision, CB, ARM and KMcD.; project administration, OOA, HM, CB and ARM.; funding acquisition, OOA, CB and ARM. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the N8AgriFood consortium and a PhD scholarship (OA) from the Commonwealth Scholarship Commission.
Institutional Review Board Statement
The animal study was approved by the Animal Welfare and Ethical Review Committee (AWERC) of the University of Leeds, UK (Approval number: N0. 070510HM).”
Data Availability Statement
The 16S rRNA sequences (fastq files) generated from the study have been deposited with the NCBI Sequence Read Archive (with BioProject ID: PRJNA798387). All data analyzed have been described in the manuscript and other statistical outputs are available in the supplementary files.
Acknowledgments
The authors appreciate the contributions of the technical staff at the National Pig Centre, University of Leeds and members of the animal bioscience and food bioactive research group. We would also like to thank Amy Taylor and Steve Laird for their contributions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, H.B.; Isaacson, R.E. The pig gut microbial diversity: Understanding the pig gut microbial ecology through the next generation high throughput sequencing. Veter- Microbiol. 2015, 177, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Guevarra, R.B.; Lee, J.H.; Lee, S.H.; Seok, M.-J.; Kim, D.W.; Na Kang, B.; Johnson, T.J.; Isaacson, R.E.; Kim, H.B. Piglet gut microbial shifts early in life: causes and effects. J. Anim. Sci. Biotechnol. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.; Ma, S.; Zhu, Z.; Su, Y.; Zoetendal, E.G.; Mackie, R.; Liu, J.; Mu, C.; Huang, R.; Smidt, H.; et al. Age, introduction of solid feed and weaning are more important determinants of gut bacterial succession in piglets than breed and nursing mother as revealed by a reciprocal cross-fostering model. Environ. Microbiol. 2016, 18, 1566–1577. [Google Scholar] [CrossRef] [PubMed]
- Guevarra, R.B.; Hong, S.H.; Cho, J.H.; Kim, B.-R.; Shin, J.; Lee, J.H.; Na Kang, B.; Kim, Y.H.; Wattanaphansak, S.; Isaacson, R.; et al. The dynamics of the piglet gut microbiome during the weaning transition in association with health and nutrition. J. Anim. Sci. Biotechnol. 2018, 9, 54. [Google Scholar] [CrossRef] [PubMed]
- Valeriano, V.; Balolong, M.; Kang, D.-K. Probiotic roles ofLactobacillussp. in swine: insights from gut microbiota. J. Appl. Microbiol. 2017, 122, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yao, W.; Li, J.; Shao, Y.; He, Q.; Xia, J.; Huang, F. Dietary garcinol supplementation improves diarrhea and intestinal barrier function associated with its modulation of gut microbiota in weaned piglets. J. Anim. Sci. Biotechnol. 2020, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Liu, Z.; Wang, C.; Ma, L.; Chen, Y.; Li, T. Effects of Dietary Protein Level on the Microbial Composition and Metabolomic Profile in Postweaning Piglets. Oxidative Med. Cell. Longev. 2022, 2022, 1–13. [Google Scholar] [CrossRef]
- Huting, A.M.; Middelkoop, A.; Guan, X.; Molist, F. Using Nutritional Strategies to Shape the Gastro-Intestinal Tracts of Suckling and Weaned Piglets. Animals 2021, 11, 402. [Google Scholar] [CrossRef]
- Xiong, X.; Tan, B.; Song, M.; Ji, P.; Kim, K.; Yin, Y.; Liu, Y. Nutritional Intervention for the Intestinal Development and Health of Weaned Pigs. Front. Veter- Sci. 2019, 6, 46. [Google Scholar] [CrossRef]
- Vasquez, R.; Oh, J.K.; Song, J.H.; Kang, D.-K. Gut microbiome-produced metabolites in pigs: a review on their biological functions and the influence of probiotics. J. Anim. Sci. Technol. 2022, 64, 671–695. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef]
- Seal, B.S.; Lillehoj, H.S.; Donovan, D.M.; Gay, C.G. Alternatives to antibiotics: a symposium on the challenges and solutions for animal production. Anim. Heal. Res. Rev. 2013, 14, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Slade, R.; Kyriazakis, I.; Carroll, S.; Reynolds, F.; Wellock, I.; Broom, L.; Miller, H. Effect of rearing environment and dietary zinc oxide on the response of group-housed weaned pigs to enterotoxigenic Escherichia coli O149 challenge. Animal 2011, 5, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Becattini, S.; Taur, Y.; Pamer, E.G. Antibiotic-Induced Changes in the Intestinal Microbiota and Disease. Trends Mol. Med. 2016, 22, 458–478. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Xiao, K.; Song, J.; Luan, Z. Effects of zinc oxide supported on zeolite on growth performance, intestinal microflora and permeability, and cytokines expression of weaned pigs. Anim. Feed. Sci. Technol. 2013, 181, 65–71. [Google Scholar] [CrossRef]
- Dong, X.; Xu, Q.; Wang, C.; Zou, X.; Lu, J. Supplemental-coated zinc oxide relieves diarrhoea by decreasing intestinal permeability in weanling pigs. J. Appl. Anim. Res. 2019, 47, 362–368. [Google Scholar] [CrossRef]
- Zhao, Y.; Su, J.-Q.; An, X.-L.; Huang, F.-Y.; Rensing, C.; Brandt, K.K.; Zhu, Y.-G. Feed additives shift gut microbiota and enrich antibiotic resistance in swine gut. Sci. Total. Environ. 2018, 621, 1224–1232. [Google Scholar] [CrossRef]
- Holman, D.B.; Chénier, M.R. Antimicrobial use in swine production and its effect on the swine gut microbiota and antimicrobial resistance. Can. J. Microbiol. 2015, 61, 785–798. [Google Scholar] [CrossRef]
- Pi, Y.; Gao, K.; Peng, Y.; Mu, C.L.; Zhu, W.Y. Antibiotic-induced alterations of the gut microbiota and microbial fermentation in protein parallel the changes in host nitrogen metabolism of growing pigs. Animal 2019, 13, 262–272. [Google Scholar] [CrossRef]
- Starke, I.C.; Pieper, R.; Neumann, K.; Zentek, J.; Vahjen, W. The impact of high dietary zinc oxide on the development of the intestinal microbiota in weaned piglets. FEMS Microbiol. Ecol. 2013, 87, 416–427. [Google Scholar] [CrossRef]
- Pieper, R.; Vahjen, W.; Neumann, K.; Van Kessel, A.G.; Zentek, J. Dose-dependent effects of dietary zinc oxide on bacterial communities and metabolic profiles in the ileum of weaned pigs. J. Anim. Physiol. Anim. Nutr. 2011, 96, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Ciesinski, L.; Guenther, S.; Pieper, R.; Kalisch, M.; Bednorz, C.; Wieler, L.H. High dietary zinc feeding promotes persistence of multi-resistant E. coli in the swine gut. PLOS ONE 2018, 13, e0191660. [Google Scholar] [CrossRef] [PubMed]
- Vahjen, W.; Pieper, R.; Zentek, J. Increased dietary zinc oxide changes the bacterial core and enterobacterial composition in the ileum of piglets1. J. Anim. Sci. 2011, 89, 2430–2439. [Google Scholar] [CrossRef] [PubMed]
- Schokker, D.; Zhang, J.; Vastenhouw, S.A.; Heilig, H.G.H.J.; Smidt, H.; Rebel, J.M.J.; Smits, M.A. Long-Lasting Effects of Early-Life Antibiotic Treatment and Routine Animal Handling on Gut Microbiota Composition and Immune System in Pigs. PLOS ONE 2015, 10, e0116523–e0116523. [Google Scholar] [CrossRef] [PubMed]
- Bonetti, A.; Tugnoli, B.; Piva, A.; Grilli, E. Towards Zero Zinc Oxide: Feeding Strategies to Manage Post-Weaning Diarrhea in Piglets. Animals 2021, 11, 642. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.; Kyvsgaard, N.C.; Battisti, A.; Baptiste, K.E. Environmental and public health related risk of veterinary zinc in pig production - Using Denmark as an example. Environ. Int. 2018, 114, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Yazdankhah, S.; Rudi, K.; Bernhoft, A. Zinc and copper in animal feed – development of resistance and co-resistance to antimicrobial agents in bacteria of animal origin. Microb. Ecol. Health Dis. 2014, 25. [Google Scholar] [CrossRef]
- Bednorz, C.; Oelgeschläger, K.; Kinnemann, B.; Hartmann, S.; Neumann, K.; Pieper, R.; Bethe, A.; Semmler, T.; Tedin, K.; Schierack, P.; et al. The broader context of antibiotic resistance: Zinc feed supplementation of piglets increases the proportion of multi-resistant Escherichia coli in vivo. Int. J. Med Microbiol. 2013, 303, 396–403. [Google Scholar] [CrossRef]
- de Oliveira, S.P.A.; do Nascimento, H.M.A.; Sampaio, K.B.; de Souza, E.L. A review on bioactive compounds of beet (beta vulgaris l. Subsp. Vulgaris) with special emphasis on their beneficial effects on gut microbiota and gastrointestinal health. Critical Rev Food Sci and Nutr 2021, 61, 2022–2033. [Google Scholar]
- Baião, D.D.S.; da Silva, V.; Paschoalin, M. Beetroot, a Remarkable Vegetable: Its Nitrate and Phytochemical Contents Can be Adjusted in Novel Formulations to Benefit Health and Support Cardiovascular Disease Therapies. Antioxidants 2020, 9, 960. [Google Scholar] [CrossRef]
- Ninfali, P.; Angelino, D. Nutritional and functional potential of Beta vulgaris cicla and rubra. Fitoterapia 2013, 89, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Nemzer, B.; Pietrzkowski, Z.; Spórna, A.; Stalica, P.; Thresher, W.; Michałowski, T.; Wybraniec, S. Betalainic and nutritional profiles of pigment-enriched red beet root (Beta vulgaris L.) dried extracts. Food Chem. 2010, 127, 42–53. [Google Scholar] [CrossRef]
- Chhikara, N.; Kushwaha, K.; Sharma, P.; Gat, Y.; Panghal, A. Bioactive compounds of beetroot and utilization in food processing industry: A critical review. Food Chem. 2018, 272, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Gómez, B.; Peláez, C.; Martínez-Cuesta, M.C.; Parajó, J.C.; Alonso, J.L.; Requena, T. Emerging prebiotics obtained from lemon and sugar beet byproducts: Evaluation of their in vitro fermentability by probiotic bacteria. LWT 2019, 109, 17–25. [Google Scholar] [CrossRef]
- Danneskiold-Samsøe, N.B.; Barros, H.D.d.F.Q.; Santos, R.; Bicas, J.L.; Cazarin, C.B.B.; Madsen, L.; Kristiansen, K.; Pastore, G.M.; Brix, S.; Júnior, M.R.M. Interplay between food and gut microbiota in health and disease. Food Res. Int. 2018, 115, 23–31. [Google Scholar] [CrossRef] [PubMed]
- van der Merwe, M. Gut microbiome changes induced by a diet rich in fruits and vegetables. Int J Food Sci Nutr 2021, 72, 665–669. [Google Scholar] [CrossRef]
- Clifford, T.; Howatson, G.; West, D.J.; Stevenson, E.J. The Potential Benefits of Red Beetroot Supplementation in Health and Disease. Nutrients 2015, 7, 2801–2822. [Google Scholar] [CrossRef]
- Singh, B.; Hathan, B.S. Chemical composition, functional properties and processing of beetroot—a review. Int J Sci and Eng Res 2014, 5, 679–684. [Google Scholar]
- Klewicka, E.; Zduńczyk, Z.; Juśkiewicz, J.; Klewicki, R. Effects of lactofermented beetroot juice alone or with n-nitroso-n-methylurea on selected metabolic parameters, composition of the microbiota adhering to the gut epithelium and antioxidant status of rats. Nutrients 2015, 7, 5905–5915. [Google Scholar] [CrossRef]
- Haq, I.U.; Butt, M.S.; Randhawa, M.A.; Shahid, M. Nephroprotective effects of red beetroot-based beverages against gentamicin-induced renal stress. J. Food Biochem. 2019, 43, e12873. [Google Scholar] [CrossRef]
- Mirmiran, P.; Houshialsadat, Z.; Gaeini, Z.; Bahadoran, Z.; Azizi, F. Functional properties of beetroot (Beta vulgaris) in management of cardio-metabolic diseases. Nutr. Metab. 2020, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wootton-Beard, P.C.; Brandt, K.; Fell, D.; Warner, S.; Ryan, L. Effects of a beetroot juice with high neobetanin content on the early-phase insulin response in healthy volunteers. J. Nutr. Sci. 2014, 3, e9. [Google Scholar] [CrossRef] [PubMed]
- da Silva, D.V.T.; Pereira, A.D.; Boaventura, G.T.; Ribeiro, R.S.d.A.; Verícimo, M.A.; de Carvalho-Pinto, C.E.; Baião, D.d.S.; Del Aguila, E.M.; Paschoalin, V.M.F. Short-Term Betanin Intake Reduces Oxidative Stress in Wistar Rats. Nutrients 2019, 11, 1978. [Google Scholar] [CrossRef] [PubMed]
- Pinedo-Gil, J.; Tomás-Vidal, A.; Jover-Cerdá, M.; Tomás-Almenar, C.; Sanz-Calvo, M.Á.; Martín-Diana, A.B. Red beet and betaine as ingredients in diets of rainbow trout (oncorhynchus mykiss): Effects on growth performance, nutrient retention and flesh quality. Arch Anim Nutr 2017, 71, 486–505. [Google Scholar] [CrossRef] [PubMed]
- Pinedo-Gil, J.; Martín-Diana, A.B.; Bertotto, D.; Sanz-Calvo, M. .; Jover-Cerdá, M.; Tomás-Vidal, A. Effects of dietary inclusions of red beet and betaine on the acute stress response and muscle lipid peroxidation in rainbow trout. Fish Physiol. Biochem. 2018, 44, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.; Wu, X.; Chen, D.; Yu, B.; He, J. Adaptation of gut microbiome to different dietary nonstarch polysaccharide fractions in a porcine model. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Howell, K.; Padayachee, A.; Comino, T.; Chhan, R.; Zhang, P.; Ng, K.; Cottrell, J.J.; Dunshea, F.R. Effect of a polyphenol-rich plant matrix on colonic digestion and plasma antioxidant capacity in a porcine model. J. Funct. Foods 2019, 57, 211–221. [Google Scholar] [CrossRef]
- Li, H.; Li, H.; Xie, P.; Li, Z.; Yin, Y.; Blachier, F.; Kong, X. Dietary supplementation with fermented Mao-tai lees beneficially affects gut microbiota structure and function in pigs. AMB Express 2019, 9, 26. [Google Scholar] [CrossRef]
- Sutton, T.; O’neill, H.M.; Bedford, M.; McDermott, K.; Miller, H. Effect of xylanase and xylo-oligosaccharide supplementation on growth performance and faecal bacterial community composition in growing pigs. Anim. Feed. Sci. Technol. 2021, 274, 114822. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for rna-seq data with deseq2. Genome Biol 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Iwai, S.; Weinmaier, T.; Schmidt, B.L.; Albertson, D.G.; Poloso, N.J.; Dabbagh, K.; DeSantis, T.Z. Piphillin: Improved Prediction of Metagenomic Content by Direct Inference from Human Microbiomes. PLOS ONE 2016, 11, e0166104. [Google Scholar] [CrossRef]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.E.; Bedford, M.R.; Miller, H.M. The effects of xylanase on grower pig performance, concentrations of volatile fatty acids and peptide YY in portal and peripheral blood. Animal 2018, 12, 2499–2504. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, W.; Lee, Y.-K.; Xie, J.; Zhang, H. Spatial Heterogeneity and Co-occurrence of Mucosal and Luminal Microbiome across Swine Intestinal Tract. Front. Microbiol. 2018, 9, 48. [Google Scholar] [CrossRef]
- Revelle, W. How to: Use the psych package for factor analysis and data reduction. Evanston, IL: Northwestern University, Department of Psychology 2016.
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Alou, M.T.; Lagier, J.-C.; Raoult, D. Diet influence on the gut microbiota and dysbiosis related to nutritional disorders. Hum. Microbiome J. 2016, 1, 3–11. [Google Scholar] [CrossRef]
- Capper, T.E.; Houghton, D.; Stewart, C.J.; Blain, A.P.; McMahon, N.; Siervo, M.; West, D.J.; Stevenson, E.J. Whole beetroot consumption reduces systolic blood pressure and modulates diversity and composition of the gut microbiota in older participants. NFS J. 2020, 21, 28–37. [Google Scholar] [CrossRef]
- Argüello, H.; Estellé, J.; Leonard, F.C.; Crispie, F.; Cotter, P.D.; O’sullivan, O.; Lynch, H.; Walia, K.; Duffy, G.; Lawlor, P.G.; et al. Influence of the Intestinal Microbiota on Colonization Resistance to Salmonella and the Shedding Pattern of Naturally Exposed Pigs. mSystems 2019, 4, e00021–19. [Google Scholar] [CrossRef] [PubMed]
- De Vries, H.; Smidt, H. Microbiota development in piglets. The suckling weaned piglet 2020, 95. [Google Scholar]
- Chen, L.; Xu, Y.; Chen, X.; Fang, C.; Zhao, L.; Chen, F. The Maturing Development of Gut Microbiota in Commercial Piglets during the Weaning Transition. Front. Microbiol. 2017, 8, 1688. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Chen, Y.; Wang, Z.; Zhou, A.; He, M.; Mao, L.; Zou, H.; Peng, Q.; Xue, B.; Wang, L.; et al. Coated zinc oxide improves intestinal immunity function and regulates microbiota composition in weaned piglets. Br. J. Nutr. 2014, 111, 2123–2134. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Zhu, C.; Chen, S.; Gao, L.; Lv, H.; Feng, R.; Zhu, Q.; Xu, J.; Chen, Z.; Jiang, Z. Dietary High Zinc Oxide Modulates the Microbiome of Ileum and Colon in Weaned Piglets. Front. Microbiol. 2017, 8, 825. [Google Scholar] [CrossRef] [PubMed]
- Namkung, H.; Gong, J.; Yu, H.; de Lange, C.F.M. Effect of pharmacological intakes of zinc and copper on growth performance, circulating cytokines and gut microbiota of newly weaned piglets challenged with coliform lipopolysaccharides. Can. J. Anim. Sci. 2006, 86, 511–522. [Google Scholar] [CrossRef]
- Gao, J.; Yin, J.; Xu, K.; Li, T.; Yin, Y. What Is the Impact of Diet on Nutritional Diarrhea Associated with Gut Microbiota in Weaning Piglets: A System Review. BioMed Res. Int. 2019, 2019, 1–14. [Google Scholar] [CrossRef]
- Awati, A.; Williams, B.A.; Bosch, M.W.; Gerrits, W.J.J.; Verstegen, M.W.A. Effect of inclusion of fermentable carbohydrates in the diet on fermentation end-product profile in feces of weanling piglets1. J. Anim. Sci. 2006, 84, 2133–2140. [Google Scholar] [CrossRef]
- Zijlmans, M.A.; Korpela, K.; Riksen-Walraven, J.M.; de Vos, W.M.; de Weerth, C. Maternal prenatal stress is associated with the infant intestinal microbiota. Psychoneuroendocrinology 2015, 53, 233–245. [Google Scholar] [CrossRef]
- Kim, H.B.; Borewicz, K.; White, B.A.; Singer, R.S.; Sreevatsan, S.; Tu, Z.J.; Isaacson, R.E. Longitudinal investigation of the age-related bacterial diversity in the feces of commercial pigs. Veter- Microbiol. 2011, 153, 124–133. [Google Scholar] [CrossRef]
- Looft, T.; Johnson, T.A.; Allen, H.K.; Bayles, D.O.; Alt, D.P.; Stedtfeld, R.D.; Sul, W.J.; Stedtfeld, T.M.; Chai, B.; Cole, J.R.; et al. In-feed antibiotic effects on the swine intestinal microbiome. Proc. Natl. Acad. Sci. 2012, 109, 1691–1696. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Nie, Y.; Chen, J.; Zhang, Y.; Wang, Z.; Fan, Q.; Yan, X. Gradual Changes of Gut Microbiota in Weaned Miniature Piglets. Front. Microbiol. 2016, 7, 1727. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Xiao, Y.; Chen, J.; Chen, J.; He, X.; Yang, H. Microbial composition in different gut locations of weaning piglets receiving antibiotics. Asian-Australasian J. Anim. Sci. 2016, 30, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R. Feed- and feed additives-related aspects of gut health and development in weanling pigs. J. Anim. Sci. Biotechnol. 2013, 4, 1–1. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.P.; Rubio, L.A.; Duncan, S.H.; Donachie, G.E.; Holtrop, G.; Lo, G.; Farquharson, F.M.; Wagner, J.; Parkhill, J.; Louis, P.; et al. Pivotal Roles for pH, Lactate, and Lactate-Utilizing Bacteria in the Stability of a Human Colonic Microbial Ecosystem. mSystems 2020, 5. [Google Scholar] [CrossRef]
- Louis, P.; Duncan, S.H.; Sheridan, P.O.; Walker, A.W.; Flint, H.J. Microbial lactate utilisation and the stability of the gut microbiome. Gut Microbiome 2022, 3, e3. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014, 8, 1323–1335. [Google Scholar] [CrossRef]
- Brestenský, M.; Nitrayová, S.; Bomba, A.; Patráš, P.; Strojný, L.; Szabadošová, V.; Pramuková, B.; Bertková, I. The content of short chain fatty acids in the jejunal digesta, caecal digesta and faeces of growing pigs. Livest Sci 2017, 205, 106–110. [Google Scholar] [CrossRef]
- Duarte, M.E.; Kim, S.W. Intestinal microbiota and its interaction to intestinal health in nursery pigs. Anim. Nutr. 2022, 8, 169–184. [Google Scholar] [CrossRef]
- Chen, X.; Zheng, R.; Liu, R.; Li, L. Goat milk fermented by lactic acid bacteria modulates small intestinal microbiota and immune responses. J. Funct. Foods 2019, 65, 103744. [Google Scholar] [CrossRef]
- Ivanov, I.I.; Honda, K. Intestinal Commensal Microbes as Immune Modulators. Cell Host Microbe 2012, 12, 496–508. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Xie, G.X.; Jia, W.P. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat Rev Gastro Hepat 2018, 15, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Winston, J.A.; Theriot, C.M. Diversification of host bile acids by members of the gut microbiota. Gut Microbes 2019, 11, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Zhang, F.; Chen, L.; Yang, Q.; Su, H.; Yang, X.; He, H.; Ling, M.; Zheng, J.; Duan, C.; et al. Dietary chenodeoxycholic acid improves growth performance and intestinal health by altering serum metabolic profiles and gut bacteria in weaned piglets. Anim. Nutr. 2021, 7, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Gui, W.; Koo, I.; Smith, P.B.; Allman, E.L.; Nichols, R.G.; Rimal, B.; Cai, J.; Liu, Q.; Patterson, A.D. The microbiome modulating activity of bile acids. Gut Microbes 2020, 11, 979–996. [Google Scholar] [CrossRef]
- Wahlström, A.; Sayin, S.I.; Marschall, H.-U.; Bäckhed, F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab. 2016, 24, 41–50. [Google Scholar] [CrossRef]
- Kaakoush, N.O. Insights into the Role of Erysipelotrichaceae in the Human Host. Front. Cell. Infect. Microbiol. 2015, 5, 84. [Google Scholar] [CrossRef]
- Palm, N.W.; De Zoete, M.R.; Cullen, T.W.; Barry, N.A.; Stefanowski, J.; Hao, L.; Degnan, P.H.; Hu, J.; Peter, I.; Zhang, W.; et al. Immunoglobulin A Coating Identifies Colitogenic Bacteria in Inflammatory Bowel Disease. Cell 2014, 158, 1000–1010. [Google Scholar] [CrossRef]
- Finlayson-Trick, E.C.L.; Getz, L.J.; Slaine, P.D.; Thornbury, M.; Lamoureux, E.; Cook, J.; Langille, M.G.I.; Murray, L.E.; McCormick, C.; Rohde, J.R.; et al. Taxonomic differences of gut microbiomes drive cellulolytic enzymatic potential within hind-gut fermenting mammals. PLOS ONE 2017, 12, e0189404. [Google Scholar] [CrossRef]
- Cuevas-Tena, M.; del Pulgar, E.M.G.; Benítez-Páez, A.; Sanz, Y.; Alegría, A.; Lagarda, M.J. Plant sterols and human gut microbiota relationship: An in vitro colonic fermentation study. J. Funct. Foods 2018, 44, 322–329. [Google Scholar] [CrossRef]
- Pereira, A.C.; Bandeira, V.; Fonseca, C.; Cunha, M.V. Egyptian Mongoose (Herpestes ichneumon) Gut Microbiota: Taxonomical and Functional Differences across Sex and Age Classes. Microorganisms 2020, 8, 392. [Google Scholar] [CrossRef] [PubMed]
- Martínez, I.; Perdicaro, D.J.; Brown, A.W.; Hammons, S.; Carden, T.J.; Carr, T.P.; Eskridge, K.M.; Walter, J. Diet-Induced Alterations of Host Cholesterol Metabolism Are Likely To Affect the Gut Microbiota Composition in Hamsters. Appl. Environ. Microbiol. 2013, 79, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Martínez, I.; Wallace, G.; Zhang, C.; Legge, R.; Benson, A.K.; Carr, T.P.; Moriyama, E.N.; Walter, J. Diet-Induced Metabolic Improvements in a Hamster Model of Hypercholesterolemia Are Strongly Linked to Alterations of the Gut Microbiota. Appl. Environ. Microbiol. 2009, 75, 4175–4184. [Google Scholar] [CrossRef]
- Etxeberria, U.; Arias, N.; Boqué, N.; Macarulla, M.; Portillo, M.; Martínez, J.; Milagro, F. Reshaping faecal gut microbiota composition by the intake of trans-resveratrol and quercetin in high-fat sucrose diet-fed rats. J. Nutr. Biochem. 2015, 26, 651–660. [Google Scholar] [CrossRef]
- Wang, C.-Q.; Yang, G.-Q. Betacyanins from Portulaca oleracea L. ameliorate cognition deficits and attenuate oxidative damage induced by D-galactose in the brains of senescent mice. Phytomedicine 2010, 17, 527–532. [Google Scholar] [CrossRef]
Figure 1.
Alpha diversity indices of pig gut microbiota; (a) Chao1 index (b) Shannon index, showing diet effect on gut species richness and/or diversity. Boxplot represents mean (minimum to maximum) species richness and or evenness from each diet in the gut locations evaluated. Significant difference between the diets linked by a line is indicated by * P < 0.05, ** P < 0.01, *** P< 0.001, ns; - not significant. CON, ZNO, RB2, and RB4 represent the control diet and diet supplemented with zinc oxide, 2% and 4% red beetroot respectively.
Figure 1.
Alpha diversity indices of pig gut microbiota; (a) Chao1 index (b) Shannon index, showing diet effect on gut species richness and/or diversity. Boxplot represents mean (minimum to maximum) species richness and or evenness from each diet in the gut locations evaluated. Significant difference between the diets linked by a line is indicated by * P < 0.05, ** P < 0.01, *** P< 0.001, ns; - not significant. CON, ZNO, RB2, and RB4 represent the control diet and diet supplemented with zinc oxide, 2% and 4% red beetroot respectively.
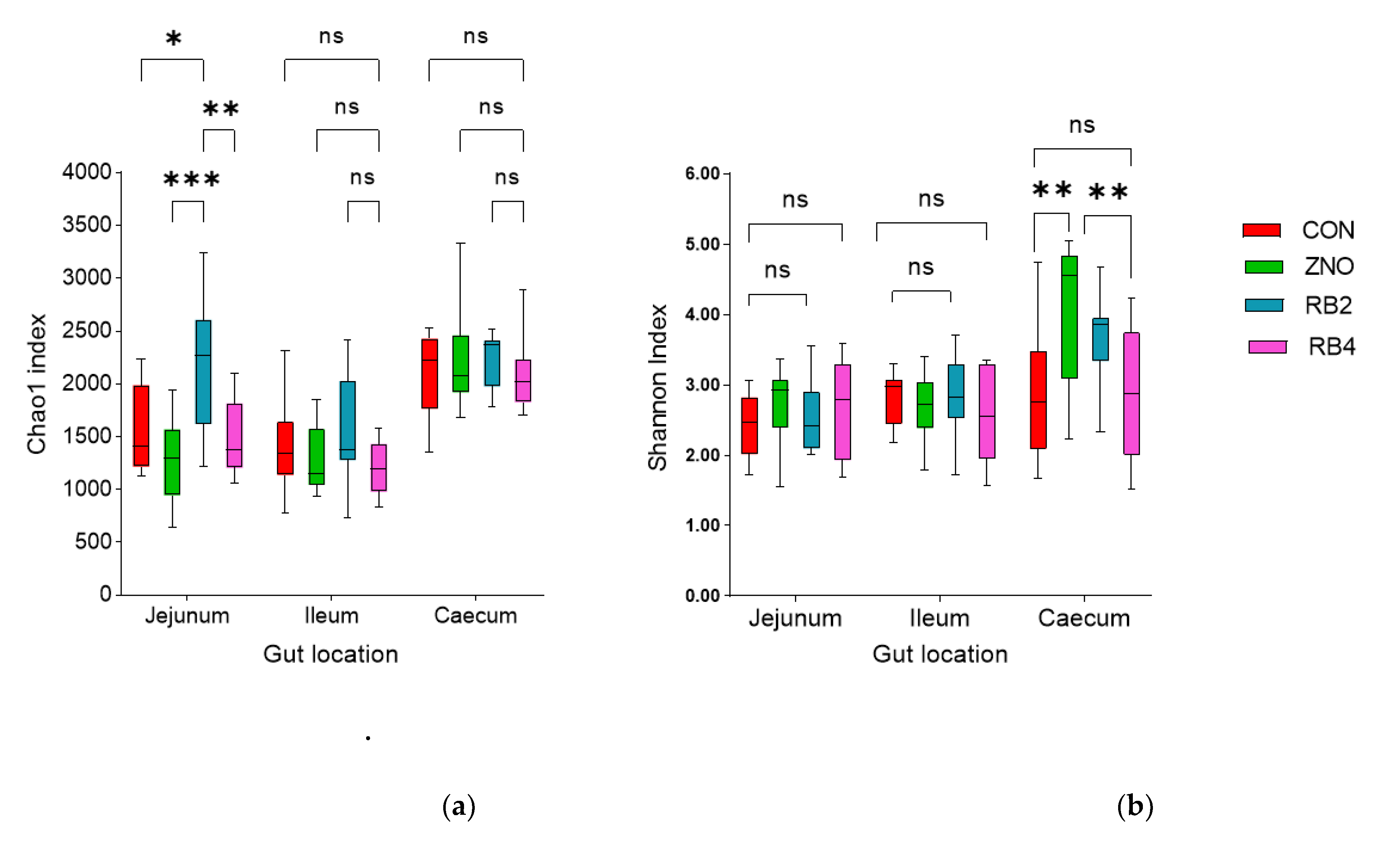
Figure 2.
Non-metric multidimensional scaling (NMDS) plots of Bray Curtis non-phylogenetic distance matrices of gut microbial community of weaned pigs fed different diets (a) distribution of samples by diet and gut location (b) distribution of samples from the caecum. Diets: CON, ZNO, RB2, and RB4 represent; control diet, diet supplemented with zinc oxide, 2% and 4% red beetroot supplemented diets respectively.
Figure 2.
Non-metric multidimensional scaling (NMDS) plots of Bray Curtis non-phylogenetic distance matrices of gut microbial community of weaned pigs fed different diets (a) distribution of samples by diet and gut location (b) distribution of samples from the caecum. Diets: CON, ZNO, RB2, and RB4 represent; control diet, diet supplemented with zinc oxide, 2% and 4% red beetroot supplemented diets respectively.
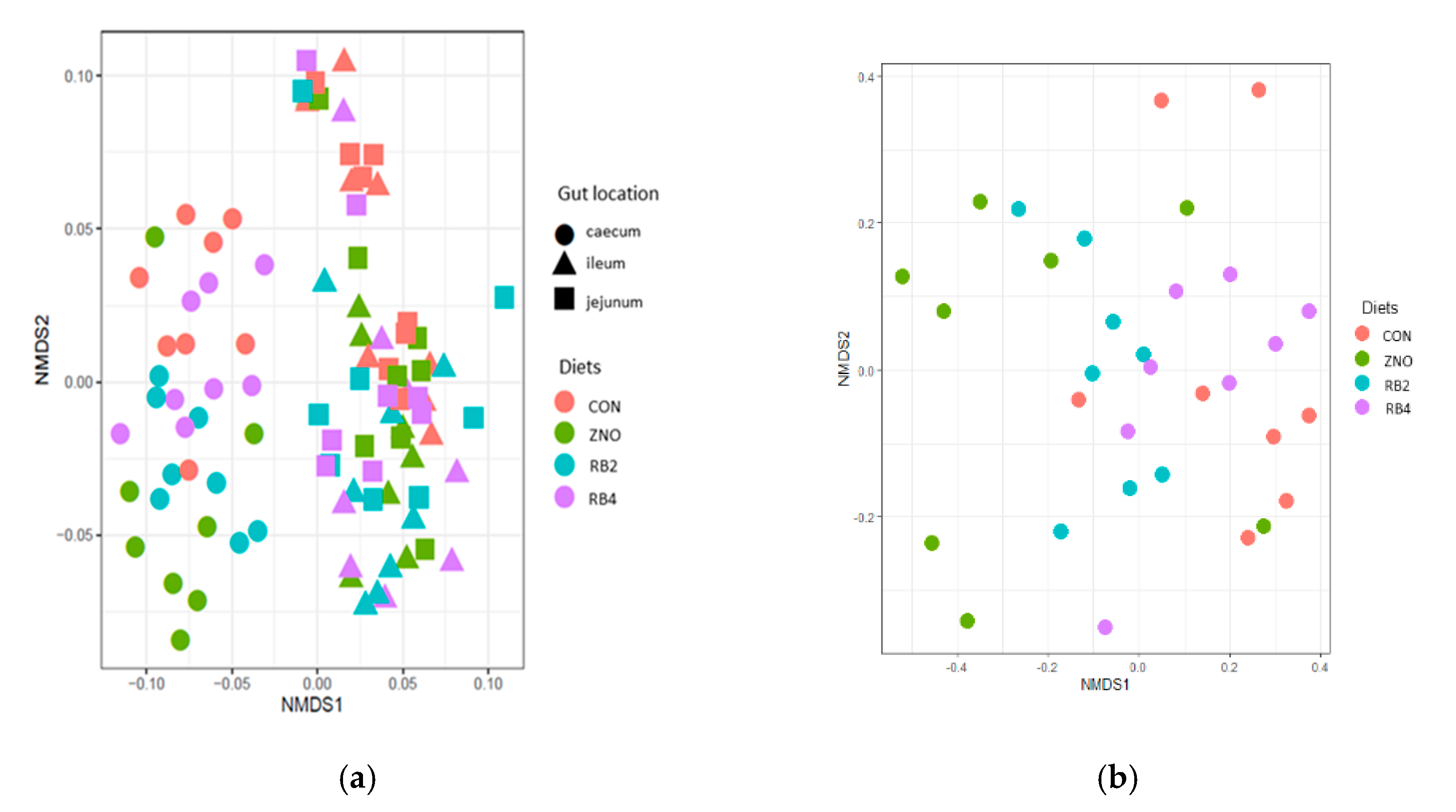
Figure 3.
Mean relative abundance of (a) phyla and (b) genera in pig gut locations with respective diets. CON, ZNO, RB2 and RB4 represent control diet and diets supplemented with zinc oxide, 2% and 4% red beetroot respectively.
Figure 3.
Mean relative abundance of (a) phyla and (b) genera in pig gut locations with respective diets. CON, ZNO, RB2 and RB4 represent control diet and diets supplemented with zinc oxide, 2% and 4% red beetroot respectively.
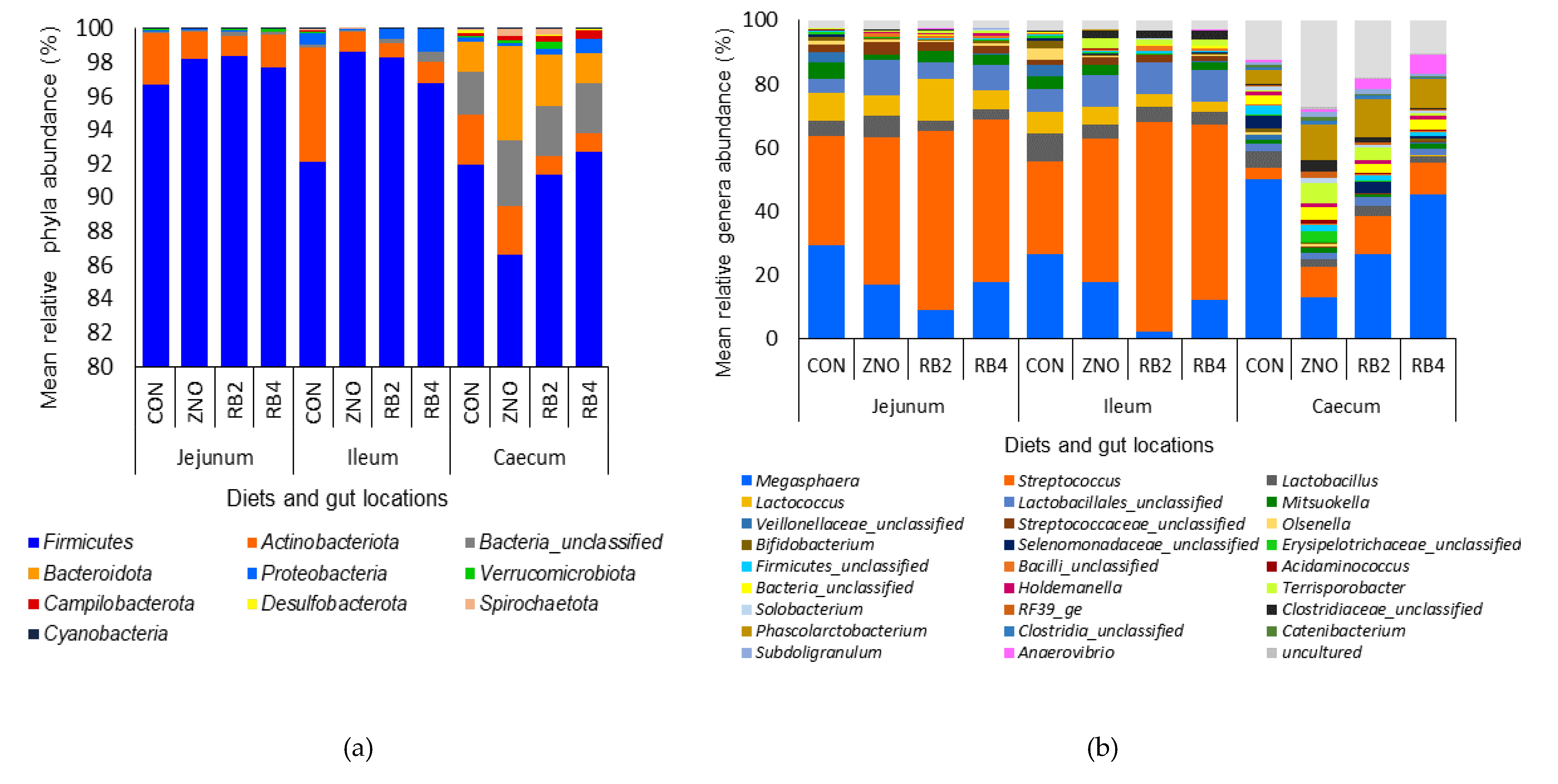
Figure 4.
Differentially abundant genera, comparisons between diets in the gut locations. Only comparisons with significant (P < 0.001) log 2-fold changes are presented. RB diets; red beetroot diets. CON, ZNO, RB2 and RB4 represents; control diet and diets supplemented with zinc oxide, 2% and 4% red beetroot, respectively.
Figure 4.
Differentially abundant genera, comparisons between diets in the gut locations. Only comparisons with significant (P < 0.001) log 2-fold changes are presented. RB diets; red beetroot diets. CON, ZNO, RB2 and RB4 represents; control diet and diets supplemented with zinc oxide, 2% and 4% red beetroot, respectively.
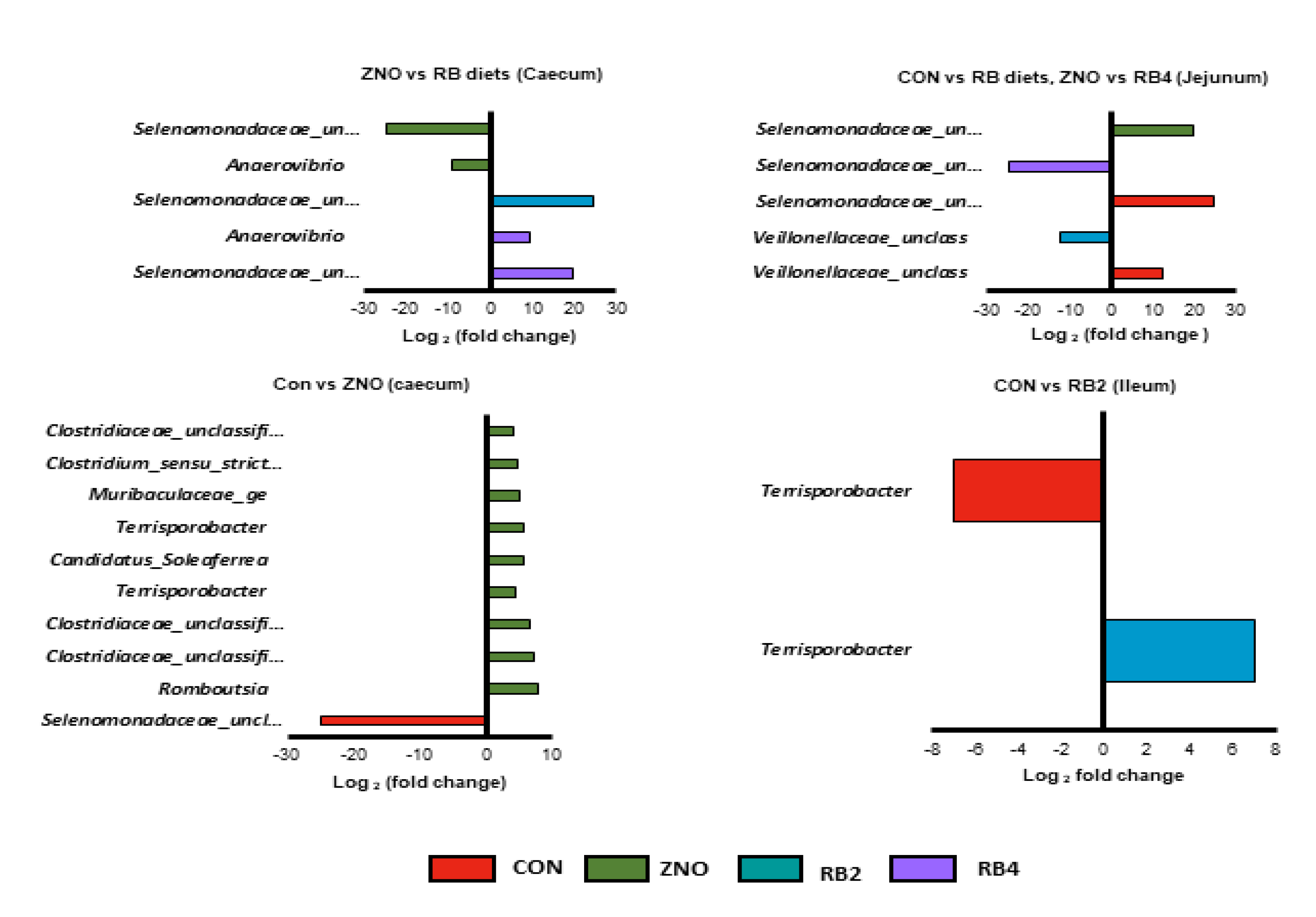
Figure 5.
Spearman correlation analyses between top abundant bacteria genera and (a) caecal short chain fatty acids (b) ileal short chain fatty acids. Omitted fatty acids were not detected in the corresponding location hence not shown, color depth depicts correlation between genera and gut metabolite where red color denotes a positive correlation and blue color a negative correlation. The strength of association between the subjects is indicated by the color intensity, *** P ≤ 0.001, **P ≤ 0.01, *P ≤ 0.05). CON, ZNO, RB2 and RB4 represent control diet and diets supplemented with zinc oxide, 2% and 4% red beetroot respectively.
Figure 5.
Spearman correlation analyses between top abundant bacteria genera and (a) caecal short chain fatty acids (b) ileal short chain fatty acids. Omitted fatty acids were not detected in the corresponding location hence not shown, color depth depicts correlation between genera and gut metabolite where red color denotes a positive correlation and blue color a negative correlation. The strength of association between the subjects is indicated by the color intensity, *** P ≤ 0.001, **P ≤ 0.01, *P ≤ 0.05). CON, ZNO, RB2 and RB4 represent control diet and diets supplemented with zinc oxide, 2% and 4% red beetroot respectively.

Figure 6.
Spearman correlation matrices between jejunal genera abundance and bile acid levels. Omitted bile acid (deoxycholic acid - DCA) was not detected in the jejunum for the pigs hence not shown. Correlation depicted by color depth, where red color denotes a positive and blue color a negative correlation. The strength of association between the subjects is indicated by the color intensity and *** P ≤ 0.001, **P ≤ 0.01, *P ≤ 0.05). CON, ZNO, RB2 and RB4 represent control and diets supplemented with zinc oxide, 2% and 4% red beetroot respectively. Taurohyodeoxycholic acid (THCA), glycohyodeoxycholic acid (GHDCA), taurocholic acid (TCA), glycocholic acid (GCA), taurochenodeoxycholic acid (TCDCA), taurodeoxycholic acid (TDCA), glycochenodeoxycholic acid (GCDCA), glycodeoxycholic acid (GDCA), cholic acid (CA), glycolithocholic acid (GLTHCA), chenodeoxycholic acid (CDCA) and lithocholic acid.
Figure 6.
Spearman correlation matrices between jejunal genera abundance and bile acid levels. Omitted bile acid (deoxycholic acid - DCA) was not detected in the jejunum for the pigs hence not shown. Correlation depicted by color depth, where red color denotes a positive and blue color a negative correlation. The strength of association between the subjects is indicated by the color intensity and *** P ≤ 0.001, **P ≤ 0.01, *P ≤ 0.05). CON, ZNO, RB2 and RB4 represent control and diets supplemented with zinc oxide, 2% and 4% red beetroot respectively. Taurohyodeoxycholic acid (THCA), glycohyodeoxycholic acid (GHDCA), taurocholic acid (TCA), glycocholic acid (GCA), taurochenodeoxycholic acid (TCDCA), taurodeoxycholic acid (TDCA), glycochenodeoxycholic acid (GCDCA), glycodeoxycholic acid (GDCA), cholic acid (CA), glycolithocholic acid (GLTHCA), chenodeoxycholic acid (CDCA) and lithocholic acid.
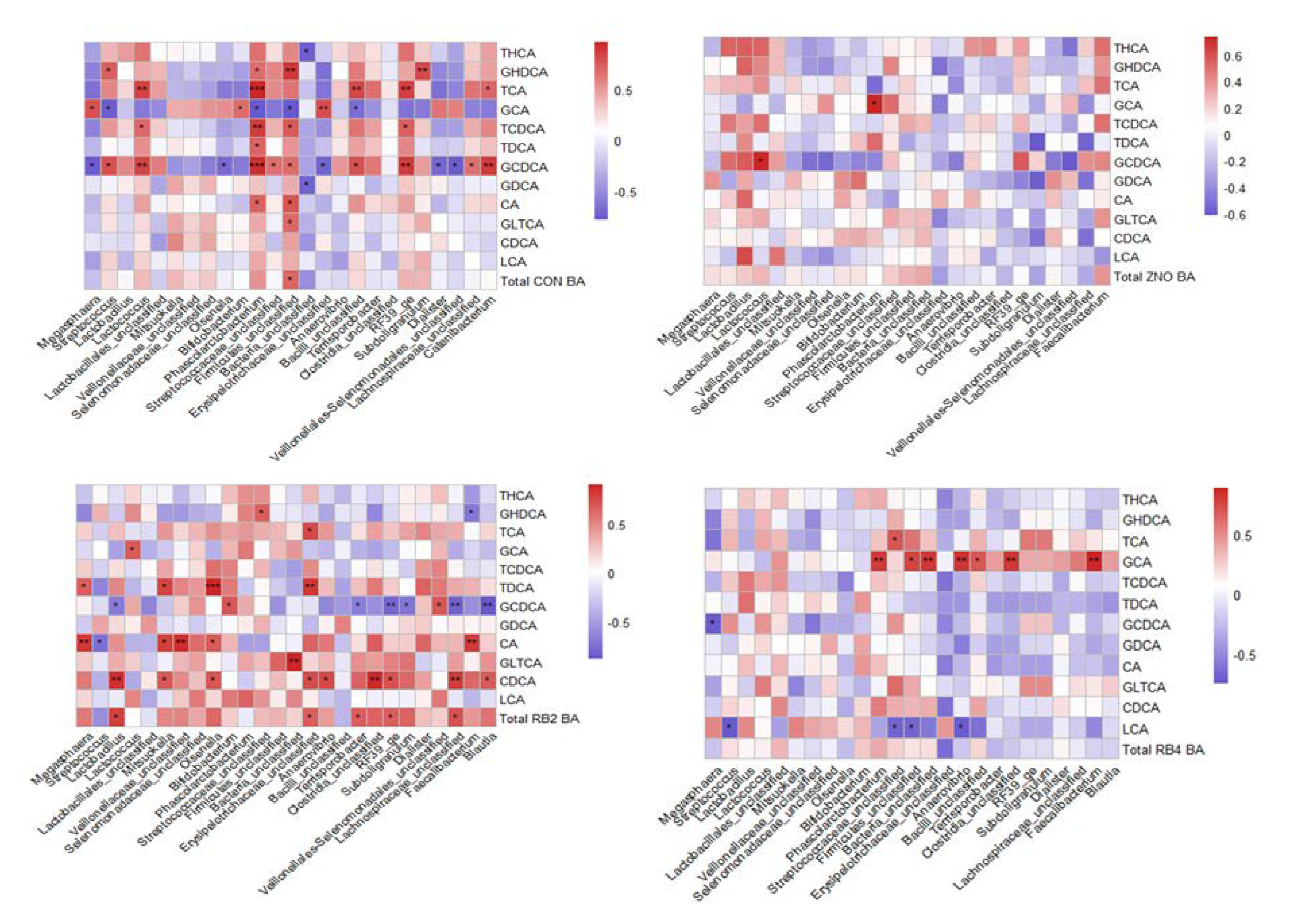

Table 1.
Composition of experimental diets and result of analyzed nutrients.
| Ingredients (%) | Control (CON) | Diet with ZnO (ZNO) | 2% red beetroot supplemented diet (RB2) | 4% red beet root supplemented diet (RB4) |
|---|---|---|---|---|
| Barley | 15.00 | 15.00 | 14.70 | 14.40 |
| Wheat | 28.17 | 28.17 | 27.51 | 26.95 |
| Micronized maize bulk | 2.50 | 2.50 | 2.45 | 2.40 |
| Micronized oats | 5.00 | 5.00 | 4.90 | 4.80 |
| Fishmeal bulk | 6.00 | 6.00 | 5.88 | 5.76 |
| Soya hypro | 18.16 | 18.16 | 17.80 | 17.43 |
| Full fat soybean | 2.50 | 2.50 | 2.45 | 2.40 |
| Pig weaner premix | 0.50 | 0.50 | 0.49 | 0.48 |
| Whey powder bulk | 13.89 | 13.89 | 13.61 | 13.33 |
| Potato protein | 1.60 | 1.60 | 1.57 | 1.54 |
| Sugar/sucrose | 0.63 | 0.63 | 0.61 | 0.60 |
| L-Lysine | 0.28 | 0.28 | 0.28 | 0.27 |
| DL-Methionine | 0.19 | 0.19 | 0.19 | 0.19 |
| L-Threonine | 0.15 | 0.15 | 0.15 | 0.15 |
| L-Tryptophan | 0.02 | 0.02 | 0.02 | 0.02 |
| L-Valine | 0.04 | 0.04 | 0.04 | 0.04 |
| Vitamin E | 0.04 | 0.04 | 0.04 | 0.04 |
| Pan-tek robust | 0.02 | 0.02 | 0.02 | 0.02 |
| Sucram | 0.01 | 0.01 | 0.01 | 0.01 |
| Benzoic acid | 0.50 | 0.50 | 0.49 | 0.48 |
| Pigzin (zinc oxide) | 0.00 | 0.31 | 0.00 | 0.00 |
| Di-calcium phosphate | 1.13 | 1.13 | 1.11 | 1.08 |
| Sodium carbonate | 0.05 | 0.05 | 0.05 | 0.05 |
| Sipernat 50 | 0.31 | 0.00 | 0.30 | 0.30 |
| Red beetroot | 0.00 | 0.00 | 2.00 | 4.00 |
| Soya oil | 3.40 | 3.40 | 3.33 | 3.26 |
| Total (%) | 100 | 100 | 100 | 100 |
| Dry matter (%) | 89.93 | 89.65 | 89.47 | 89.01 |
| Analysed nutrient | ||||
| Ash (%) | 6.80 | 7.50 | 6.70 | 6.60 |
| Ether extract (%) | 6.73 | 6.99 | 6.62 | 5.92 |
| Crude protein (%) | 21.30 | 21.30 | 20.70 | 20.40 |
| Crude fibre (%) | 1.90 | 1.50 | 1.80 | 2.20 |
| Zinc (mg/kg) | 422.00 | 2252.00 | 193.00 | 187.00 |
Table 2.
Comparative analyses of mean relative phyla abundance between diet and gut locations.
|
Phylum |
Diets | Gut locations | dSEM | P value | |||||||
| CON | ZNO | RB2 | RB4 | Jejunum | Ileum | Caecum | fL | eD | gL* D | ||
| Firmicutes | 93.58a | 94.48a | 96.01a | 95.72a | 97.74a | 96.46a | 90.64b | 0.617 | 0.000 | 0.312 | 0.062 |
| Actinobacteriota | 4.25a | 1.88ab | 1.06b | 1.42b | 1.93a | 2.51a | 2.02a | 0.347 | 0.733 | 0.004 | 0.224 |
| Bacteria unclassified | 0.95a | 1.37a | 1.13a | 1.26a | 0.17b | 0.28b | 3.09a | 0.206 | 0.000 | 0.796 | 0.800 |
| Bacteroidota | 0.61a | 1.86a | 1.03a | 0.59a | 0.003b | 0.005b | 3.06a | 0.248 | 0.000 | 0.067 | 0.029 |
| Proteobacteria | 0.31ab | 0.11b | 0.31ab | 0.71a | 0.03b | 0.66a | 0.39a | 0.068 | 0.000 | 0.005 | 0.126 |
| Verrucomicrobiota | 0.09a | 0.06a | 0.18a | 0.08a | 0.11a | 0.04a | 0.16a | 0.026 | 0.187 | 0.389 | 0.132 |
| Campilobacterota | 0.09a | 0.08a | 0.13a | 0.16a | 0.01b | 0.01b | 0.32a | 0.041 | 0.002 | 0.916 | 0.921 |
| Desulfobacterota | 0.07a | 0.01a | 0.03a | 0.04a | 0.004b | 0.001b | 0.11a | 0.010 | 0.000 | 0.230 | 0.213 |
| Spirochaetota | 0.05a | 0.13a | 0.11a | 0.01a | 0.001b | 0.03b | 0.19a | 0.026 | 0.003 | 0.259 | 0.058 |
Data represents mean phyla abundance in each gut location with the different experimental diets, significant difference was indicated by different superscript between diet groups and the gut location. d Standard error of the group mean, e p value for gut location, f p value for diet, g p value for interaction between the gut location and diet. CON, ZNO, RB2 and RB4 represent control diet and diets supplemented with zinc oxide, 2% and 4% red beetroot respectively.
Table 3.
Short chain fatty acid (mM) profile of diet groups and locations evaluated.
| SFCA | Diets | P value | ||||||
| CON | ZNO | RB2 | RB4 | SEM | *L | #D | +L*D | |
| Acetate | 69.14a | 54.76b | 63.81ab | 56.64b | 3.32 | < 0.01 | < 0.05 | > 0.05 |
| Propionate | 23.83a | 17.04d | 20.34b | 19.16c | 1.42 | < 0.05 | < 0.01 | < 0.01 |
| Isobutyrate | 2.59a | 1.60b | 1.79b | 0.86c | 0.36 | < 0.02 | < 0.02 | < 0.02 |
| Butyrate | 9.48a | 6.76b | 8.08ab | 7.09b | 0.61 | < 0.05 | < 0.05 | > 0.05 |
| Isovalerate | 1.11a | 0.71b | 0.67b | 0.44c | 0.14 | < 0.02 | < 0.02 | < 0.05 |
| Valerate | 1.55a | 0.82b | 0.58c | 0.63c | 0.22 | < 0.02 | < 0.02 | < 0.05 |
| Location | ||||||||
| 1Plasma | 1.99 | 1.87 | 2.31 | 1.61 | 0.15 | < 0.01 | > 0.05 | < 0.05 |
| 1Jejunum | 7.40b | 7.88b | 11.28a | 7.08b | 0.97 | < 0.01 | < 0.05 | < 0.05 |
| 1Ileum | 9.21a | 7.91b | 9.82a | 8.45a | 0.42 | < 0.01 | < 0.05 | < 0.05 |
| Caecum | 24.91 | 22.36 | 23.71 | 22.12 | 0.65 | < 0.05 | > 0.05 | < 0.05 |
| Colon | 24.57a | 19.25b | 17.32b | 22.29ab | 1.61 | < 0.05 | < 0.05 | < 0.05 |
| Feces | 39.61a | 22.43c | 30.84b | 23.27c | 4.00 | < 0.05 | < 0.05 | < 0.05 |
| Total SCFA | 107.70a | 81.69b | 95.28a | 84.82b | 5.876 | < 0.05 | < 0.05 | < 0.05 |
Data represent mean SCFA for each diet group and gut location, significant differences were indicated by different superscripts across the rows. CON, ZNO, RB2 and RB4 represent; control diet, diets supplemented with zinc oxide, 2% and 4% red beetroot respectively. 1SCFA levels in these locations were significantly different from levels in the caecum. *p value for effect of location on SCFA levels, #p value for significant effect of diets, +p value interaction between location and diet.
Table 4.
Bile acid profile (nmol/g digesta/feces) from diet groups and locations evaluated.
| Bile acids | Diets | P value | ||||||
| CON | ZNO | RB2 | RB4 | SEM | *L | #D | +L*D | |
| THCA | 40.74a | 27.29b | 16.88c | 26.86b | 4.90 | < 0.01 | < 0.01 | 0.75 |
| GHDCA | 33.78b | 46.08a | 25.76c | 33.45b | 4.20 | < 0.01 | 0.05 | 0.62 |
| TCA | 1.50b | 1.56b | 1.96a | 1.30b | 0.14 | < 0.01 | < 0.01 | < 0.01 |
| GCA | 0.40b | 0.47b | 0.45b | 1.37a | 0.23 | > 0.05 | 0.01 | > 0.05 |
| TCDCA | 21.90a | 8.89c | 8.30c | 15.09b | 3.18 | < 0.01 | < 0.01 | < 0.01 |
| TDCA | 5.11a | 3.42b | 4.07ab | 3.90b | 0.36 | 0.02 | 0.05 | 0.93 |
| GCDCA | 15.42c | 13.88c | 35.91a | 25.40b | 5.10 | < 0.01 | < 0.01 | 0.25 |
| GDCA | 3.57b | 4.91a | 2.95c | 3.26b | 0.43 | < 0.01 | < 0.01 | 0.05 |
| GLTCA | 2.11ab | 1.99b | 3.60a | 1.96b | 0.40 | < 0.05 | < 0.01 | 0.18 |
| CA | 3.80b | 2.59c | 4.23a | 1.63d | 0.59 | < 0.01 | < 0.01 | < 0.01 |
| CDCA | 121.16b | 72.28c | 147.09a | 74.32c | 18.34 | < 0.01 | < 0.01 | < 0.01 |
| DCA | 0.10b | 0.14b | 0.31a | 0.12b | 0.05 | < 0.02 | < 0.02 | 0.20 |
| LCA | 134.25a | 108.90b | 118.20b | 107.66b | 6.13 | < 0.01 | 0.05 | 0.42 |
| Location | ||||||||
| 1Jejunum | 138.20b | 135.03b | 210.83a | 128.68c | 19.32 | < 0.01 | < 0.05 | < 0.05 |
| Ileum | 95.20a | 40.21c | 29.11d | 50.09b | 14.40 | < 0.01 | < 0.01 | < 0.05 |
| Caecum | 23.95a | 17.37b | 15.65b | 12.84c | 2.35 | < 0.01 | < 0.05 | < 0.05 |
| Colon | 59.88a | 28.45c | 46.40b | 21.10d | 8.77 | <0.01 | < 0.05 | < 0.05 |
| Faeces | 66.59c | 71.33b | 67.72c | 83.62a | 3.90 | < 0.01 | < 0.01 | < 0.05 |
| Total unconjugated | 259.30a | 183.91b | 269.83a | 183.74b | 23.41 | < 0.05 | < 0.05 | < 0.05 |
| Total conjugated | 124.52a | 108.48b | 99.88b | 112.60b | 5.12 | < 0.05 | < 0.05 | < 0.05 |
| Total bile acids | 383.82a | 292.39b | 369.71a | 296.33b | 23.98 | < 0.001 | < 0.05 | < 0.01 |
Data represents mean bile acid levels for each diet and location, significant differences indicated by superscripts across the table for the diet groups. CON, ZNO, RB2 and RB4 represent control diet and diets supplemented with zinc oxide, 2% and 4% red beetroot respectively. *p value for effect of location, #p value for significant effect of diets, +p value interaction between location and diet. Taurohyodeoxycholic acid (THCA), glycohyodeoxycholic acid (GHDCA), taurocholic acid (TCA), glycocholic acid (GCA), taurochenodeoxycholic acid (TCDCA), taurodeoxycholic acid (TDCA), glycochenodeoxycholic acid (GCDCA), glycodeoxycholic acid (GDCA), cholic acid (CA), glycolithocholic acid (GLTHCA), chenodeoxycholic acid (CDCA), deoxycholic acid (DCA) and lithocholic acid.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



