
Submitted:
01 March 2024
Posted:
04 March 2024
You are already at the latest version
Abstract
Polyphenols and fermentable fibers have shown favorable effects on gut microbiota composition and metabolic function. However, few studies have investigated whether combining multiple fermentable fibers or polyphenols may have additive beneficial effects on gut microbial states. Here, an in vitro fer-mentation model, seeded with human stool combined from 30 healthy volunteers, was supplemented with blends of polyphenols (PP), dietary fibers (FB) or their combination (PPFB) to determine influence on gut bacteria growth dynamics and select metabolite changes. PP and FB blends independently lead to significant increases in the absolute abundance of select beneficial taxa, namely Ruminococcus bromii, Bifidobacterium spp., Lactobacillus spp. and Dorea spp. Total short chain fatty acid concentrations, relative to non-supplemented control (F), increased significantly with PPFB and FB supplementation, but not PP. Indole and ammonia concentrations decreased with FB and PPFB supplementation, but not PP alone while increased antioxidant capacity was only evident with both PP and PPFB. These findings demonstrated that, while the independent blends displayed selective positive impacts on gut states, combination of both blends provided an additive effect. The work outlines the potential of mixed substrate blends to elicit a broader positive influence on gut microbial composition and function to build resiliency toward dysbiosis.
Keywords:
prebiotics
; gut microbiome
; carbohydrate
; supplementation
; bacteria
; extracts
; metabolites
1. Introduction
Gut microbiome community composition and function is influenced by dietary intake, with diet-microbiome interactions linked to both beneficial and detrimental impacts on health outcomes [1]. As such, there is substantial interest in developing dietary strategies that can improve health by favorably modulating the gut microbiota and by promoting the growth of beneficial taxa and production of bacterial-derived metabolites [2]. Of particular interest, are dietary fibers and polyphenols, which have gained traction as nutritional supplements that positively impact gut microbial community dynamics [3,4,5,6]. Large molecular weight polyphenols and fermentable fibers are poorly absorbed by the upper digestive tract, reaching the large intestine with minimal degradation by digestive enzymes [7,8].
The majority of previous studies have focused on individual polyphenol classes (e.g. cranberry proanthocyanidins) and fiber constituents (e.g. resistant starch), which has provided significant data on the effects of individual compounds on gut microbial composition, production of beneficial metabolites, and the reduction of proinflammatory metabolites [9,10]. Studies exploring “whole foods” like fruits, vegetables, and whole grains to understand how natural combinations of polyphenols and fibers positively impact gut microbiota and host gut health have also emerged, without distinction to the specific fiber and/or polyphenol sources [11]. Focused studies understanding how specific blends of polyphenol and fibers, from varying dietary sources, impacts bacteria diversity and metabolite production are limited and strategic rationale for combination of polyphenols and fibers requires additional research to understand the potential for combinatorial effects on gut health. This is of relevance in situations where whole food consumption is not always practical such as a military operational setting.
In the current study, we have explored, through in vitro fermentation, the impact of polyphenol (PP) and fiber (FB) blends on gut microbial dynamics, pro/anti-inflammatory metabolite production, and antioxidant capacity. The PP and FB blends, respectively, were chosen to incorporate compounds with a diversity of structures that, individually, have been shown to beneficially impact the gut microbiota composition and function. The goal was to elicit broader favorable effects on gut microbiota community composition and metabolic activity than any individual fiber or polyphenol source without introducing antagonistic effects. The PP blend contained blueberry, cranberry, green tea, and cocoa powders/extracts providing a diverse range of polyphenol structures which have been shown to positively impact gut states, in particular antioxidant capacity [12,13,14,15]. Constituents of the FB blend were chosen to incorporate a diversity of fiber structures that beneficially impact the gut microbiota composition and function, particularly stimulation of short chain fatty acid (SCFA) and beneficial bacterial taxa [3,16,17]. The FB blend contained high amylose maize starch and two prebiotic substrates, galacto-oligosaccharides (GOS) and oligofructose-enriched inulin, which provided varied polysaccharide types and lengths. An in vitro model, capable of simulating the physiological conditions of the ascending domain of the large intestine [18] was employed to provide broad experimental capacity. This allowed us to study community dynamics and rates of metabolite production as a function of dietary supplementation in a systematic manner to understand blend-specific effects on gut microbial dynamics [19]. The hypothesis was that the combination of the polyphenol and fiber blends would promote synergistic outcomes on the growth of beneficial taxa, production of beneficial metabolites, and reduction of pro-inflammatory compounds and bacterial taxa associated with dysbiosis.
2. Materials and Methods
2.1. Study Population and Sample Collection
Fecal samples were sourced from a previous study, collected from 30 healthy adults (habitual diet control group)18-62 years and without obesity (BMI ≤ 30 kg.m-2) participating in a randomized controlled trial investigating the effects of consuming a diet of military food rations on the gut microbiota (clinicaltrials.gov NCT02423551) [18,20,21,22]. Participants had not used oral antibiotics or had a colonoscopy within three months of study participation, did not have a history of gastrointestinal disease or regular use of medications impacting gastrointestinal function, and were not following a restrictive diet or attempting to lose or gain weight. Additionally, participants were instructed to discontinue use of any probiotic, prebiotic, or other dietary supplements at least two weeks prior to beginning study participation. The study was reviewed and approved by the US Army Research Institute of Environmental Medicine Institutional Review Board, and all participants provided written informed consent prior to participation. Fecal sample collection was adapted from [23] and protocol is described in detail in Pantoja-Feliciano et al. [24].
2.2. Blend Formulations
For supplementation, polyphenols were extracted from four agricultural products; cocoa seed extract (CocoActiv®; 45.1% total phenolics dry wt. Cocoa Extract (CE), Naturex, Avignon, France), wild blueberry powder (4.0% total phenolics dry wt. GAE, Naturex, Avignon, France), cranberry extract (Cystricran®; 57.2% total phenolics dry wt. Gallic Acid Equivalents (GAE), Naturex, Avignon, France), and green tea leaf extract (100% total phenolics dry wt. GAE, Naturex, Avignon, France) were selected. Three fermentable fiber sources were also selected: Orafti® Synergy1 (93.2% oligofructose-enriched inulin by dry wt., Beneo GmbH, Mannheim, Germany), Bimuno-galactooligosaccharides® (85% GOS (w/w), Clasado Biosciences, Reading, UK), Hi-Maize® 260 (59% resistant starch w/w, Ingredion, Inc., Bridgewater, New Jersey, USA). Select polyphenols and fiber sources were combined in ratios comparable to that used in human study referenced above, either in alcohol-solubilized form (polyphenols) or fermentation medium (fibers), to provide a dose of 2g polyphenol/day and 30g fiber/day, respectively (Table S1). Doses were chosen to align with a human study assessing the effects of fermentable fiber and polyphenol supplementation on intestinal barrier function during environmental stress (clinicaltrials.gov identifier: NCT04111263). The supplemented blends were given the designations (PP) for the polyphenol blend and (FB) for the fiber blend. The combination of the two blends was given the designation (PPFB).
2.3. Study Design and Supplementation Parameters
An overview of the fermentation study design can be found in (Figure 1). Parallel batch fermentations (n = 3) were designed to include vessels for blend-specific supplementation, non-supplemented (fecal only, F), and fecal-deficient (medium only, NF) using an HEL BioXplorer 100 (HEL Group, Borehamwood, UK) 8-vessel parallel bioreactor simulating the ascending colon (pH 5.5). The fermentation medium, Colonic Complex Medium (CCM), was prepared based on Macfarlane et al. [25] with slight modifications: addition of resazurin (1 ug/L). The medium was added to the fermentation vessels (125mL/vessel) with pH, temperature, and oxidation-reduction potential (Applikon Biotechnologies, Foster City, CA, USA) probes attached. Vessels were autoclaved for 35 minutes at 120 psig and attached to the bioreactor for overnight equilibration under continuous oxygen-free nitrogen (20 psig, 5 mL/minute), 370C, and constant agitation (450 rpm). After equilibrium, pH values were corrected for calibration drift and pH control initiated (pH 5.5 +/- 0.1) with bioreactor-controlled adjustments using 1N NaOH and 0.2N HCl. Prior to supplementation, CCM volume equivalent to total supplementation volume was removed from each vessel. PP solubilized in methanol was added to the appropriate vessels (5 mL/vessel) using a syringe equipped with an 18-gauge needle and equilibrated overnight. After overnight stabilization, fiber, suspended in CCM, was added to appropriate vessels (9 mL/vessel) and vessels were allowed to stabilize for 30 minutes. Equal volumes of methanol and CCM were added to non-supplemented vessels corresponding to PP and FB respectively.
Fecal samples from 30 participants were thawed and pooled in serum bottles at 20% (w/v) inside an anaerobic chamber (Coy Labs, Grass Lake, MI, USA) to generate fecal inoculum prior to inoculation. The pooled fecal samples simulate a universal gut microbiome and allows for increased diversity in the microbial population by maximizing species lower in abundance from more subjects [26]. After vessel supplementation with PP, FB, or PPFB, vessels were inoculated with pooled stool through headplate septum using syringes equipped with 18-gauge needles. Fecal-deficient control vessels were inoculated with sterile 0.1M phosphate buffer pH 7.2 with 15% (w/v) glycerol. Vessels were sampled at 0, 5, 10 and 24 hours, representing community lag (0hr), logarithmic (5hr, 10hr) and stationary (24hr) growth phases and sample aliquots were stored at -800C for further analysis.
2.4. Targeted Keystone Bacterial Taxa Analysis
DNA extractions were performed using the QIAMP Power Fecal Pro DNA Extraction Kit, (QIAGEN Inc., Germantown, MD, USA). Quantification of DNA (ng/uL) was performed using a Nanodrop One™ instrument (Thermo Fisher Scientific Inc., Waltham, MA, USA). To determine absolute abundance, standard curves were generated using DNA extracted from pure cultures of eight organisms purchased from ATCC (American Type Culture Collection, Manassas, VA, USA): Bifidobacterium animalis subsp. lactis 700451, Lactobacillus reuteri 23272, Eubacterium rectale 33656, Ruminococcus bromii 27255, Akkermansia muciniphilia BAA-835, Blautia hansenii 27752, Dorea spp. BAA-2280, and Faecalibacterium prausnitzii 27768. The specific taxa were selected based on previous studies [18,24]. Briefly, organism specific quantitative polymerase chain reaction (qPCR) primers were selected for the eight organisms included [18,27] (Table S2). Serial dilutions (10-fold) were prepared using DNase and RNase free water and 2X Forget-Me-Not qPCR Master Mix (Biotium, Hayward, CA, USA) was added in the reactions. The iCycler iQ Optical module™ (Bio-Rad Laboratories, Hercules, CA, USA) software was used to quantitate each qPCR reaction. For all reactions, target qPCR efficiency was between 80% to 100% [28]. Genome size for each microorganism was used to calculate copy number for each organism (http://cels.uri.edu/gsc/cndna.html) [29]. Raw results from qPCR were log-transformed to calculate copy number/mL.
2.5. Metabolite Analysis
Fermentate concentrations of acetic, butyric, propionic, valeric, isovaleric and isobutyric acid were analyzed using the approach described by [30,31,32] with slight modifications. Briefly, aliquots were thawed, homogenized, and acidified using 50% sulfuric acid. SCFA were then extracted using diethyl ether by removing the organic layer after centrifugation. Ethyl butyric acid was added as an internal standard before storing at -80⁰C until analysis. SCFA were quantified using an Agilent 7890A GC system with Flame Ionization Detection (Agilent J&W DB-FFAP column dimensions: 60 m x 250 μm, x 0.25 μm, Agilent Technologies, Santa Clara, CA, USA). Calibration standards were included for each fatty acid and used for peak identification and quantification.
2.6. Influence of Supplementation on Indole
Indole concentrations were measured in fermentates in triplicate using the Indole Assay Kit, MAK326, (Sigma-Aldrich Co., St. Louis, MO, USA). Color intensities were quantified at 565 nm, room temperature using Biotek Powerwave HT (Agilent Technologies, Santa Clara, CA, USA). Indole concentrations were calculated from the slope of a standard curve quantified by subtracting the plate blank from the standard values and graphing against indole concentrations in slope-intercept form. The fecal-deficient sample readings were subtracted to determine net concentrations of indole in corresponding fecal vessels.
2.7. Influence of supplementation on Ammonia
Ammonia concentrations were measured in fermentates using the non-enzymatic, Ammonia Assay kit, ab102509 (Abcam Inc., Waltham, MA, USA). Prior to analysis, all samples were filtered using 10 kD spin columns, ab93349, (Abcam Inc. Waltham, MA, USA) to remove proteins and lower background levels of ammonia. Dilutions were required to bring sample concentrations within the range of the standard curve (0 mM – 10 mM). Plates were incubated for 30 minutes at 370C with samples measured in triplicate. Color intensities were quantified at 670 nm using Biotek Powerwave HT (Agilent Technologies, Santa Clara, CA, USA). Interference from reagents present in the ammonium chloride standard was compensated by subtracting standard-deficient wells from all readings. This was important as background readings can be significant in fermentates. Unknown sample values were calculated from the ammonium chloride standard curve slope. The fecal-deficient sample readings were subtracted to determine net concentrations of ammonia in corresponding fecal vessels.
2.8. Influence of Supplementation on Antioxidant Capacity
Antioxidant capacity was measured in fermentates using the Ferric Reducing Antioxidant Power (FRAP) assay kit, MAK369, Sigma-Aldrich Co. (St. Louis, MO, USA). Prior to running the assay, an initial extraction step is required, using an acid-methanol solution (prepared as 70:29.5:0.5 mixture of methanol:ultrapure water:1 M HCl). The acidic conditions allow for the dissociation of Fe3+ from the protein complex. Plates were incubated at 370C for 60 minutes with samples measured in triplicate. Color intensities were quantified at 594 nm using Biotek Powerwave HT (Agilent Technologies, Santa Clara, CA, USA). Antioxidant capacity was calculated using a standard curve, after subtracting the negative control, and plotting them using the slope-intercept form. The raw values for the unknown samples were used to calculate unknown antioxidant capacity from the standard curve.
2.9. Statistical Analysis
All statistical analyses were performed with Rstudio (RStudio 2023.3.0.0; Posit Software PBC, Boston, MA, USA). Changes in metabolite data were analyzed using 2-way repeated measures ANOVA with treatment groups (F, FB, PP, PPFB), timepoints (0, 5, 10 and 24 hours) and their interaction included as within-subjects fixed factors. In cases where the interaction was statistically significant, between-group comparisons were conducted by 1-way repeated measures ANOVA with Bonferroni corrections. The assumption of normality was verified using the Shapiro-Wilk test. Mauchly’s sphericity test was used to verify equal variances. For targeted bacterial taxa analysis and SCFA production, 1-way ANOVA with Tukey-Kramer HSD was used for pairwise comparisons. Statistical significance was defined as p ≤ 0.05. All graphs were generated using JMP® 15 (SAS Institute Inc., Cary, NC, USA).
3. Results
3.1. Effects of Polyphenol and Fiber Supplementation on Bacterial Abundance
Microbial compositional changes of 8 targeted beneficial taxa after polyphenol and fiber supplementation were determined quantitatively using qPCR. Across all the selected taxa, supplementation led to differential changes in abundance, as shown in Figure 2. Supplementation with PP and FB blends led to collective differences in Bifidobacterium spp. abundance relative to F (time-by-treatment interaction, P = 0.047; Figure 2A). Additionally, time-dependent analysis between treatment groups illustrated elevated abundances of Bifidobacterium spp. in PP, FB and PPFB at 5hr and 10hr (p<0.05) compared to F and, for FB and PPFB specifically, at 10hr and 24hr (p<0.05), but a change in abundance was not statistically significant with PP at 24h (Table S3). Significant differences were not observed between supplemented vessels. Lactobacillus spp. in supplemented vessels also showed notable increases in abundance relative to F (time-by-treatment interaction, P = 0.021; Figure 2B) and, more specifically, higher abundances in relation to PP, FB and PPFB at 10hr and 24hr (p<0.02; Table S3) with significant differences observed between PP and PPFB at 24hr (p=0.046). Additionally, there was a significant difference between PPFB and F at 5hr (p=0.032).
Dorea spp. abundance changes were driven by PP and PPFB supplementation (time-by-treatment interaction, P = 0.049; Figure 2C), resulting in sustained abundances in PP and PPFB at 10hr and 24hr (p<0.05) compared to both FB and F. FB supplementation showed similar growth trends to F with decreases in abundance after 5hr. Additionally, differences between PPFB and F were seen at 5hr (p = 0.01) (Table S3). Ruminococcus bromii, showed increased abundance after FB and PPFB supplementation (time-by-treatment interaction, P = 0.002) (Figure 2D). This led to higher abundances of R. bromii in FB and PPFB at 10hr (p<0.01) and 24hr (p<0.05) compared to F. PP supplementation showed similar growth trends to F, and differences were seen between PP and FB at 10hr (p0.017), and between PP and PPFB at 10hr (p=0.015) (Table S3).
For the other four taxa, no significant interaction between time and treatment were observed (Figure S1). However, although a two-way interaction was not evident, interestingly, Faecalibacterium prausnitzii upon PP treatment trended differentially to the other blends at 24hr while the other three organisms converged (Figure S1).
3.2. Effects of Polyphenol and Fiber Supplementation on Metabolite Concentrations
3.2.1. Changes in Total SCFA Concentrations
Mean concentrations of total SCFAs (acetic, propionic, and butyric acid) increased from 0hr to 24hr differently within each treatment group (Figure 3, Table S4). There was a significant time by treatment interaction (P<0.001). Pairwise comparisons between treatment within each time point showed significant increases in FB and PPFB compared to F at 10hr and 24hr (p<0.05, Table S4). However, significant statistical differences were not evident between PP and F (p>0.05, Table S4) across all fermentation time points. Significant differences were not seen between PP and FB compared to PPFB (p>0.05, Table S4) at any residence times. The ratio of acetic acid, propionic acid, and butyric acid was similar across all blends; however, butyric acid was slightly elevated in FB relative to PP and PPFB at 24hr (Table S4). At 10hr, butyric acid proportion in PP were 2-fold and 3-fold higher than that seen in PPFB and FB respectively. Although total SCFA demonstrated statistical significance in select instances, mean concentrations at individual time points for SCFA (acetic, propionic, and butyric acid) were not significantly different across all blends as a function of time (P≥0.05, Table S5).
3.2.2. Changes in Indole Concentrations
Indole analysis was performed to measure protein metabolite production. Mean concentration of indole (n = 3) decreased within F from 0hr to 10hr and then increased at 24hr (Table S6). Indole concentrations were high in both FB and PP compared to F and PPFB at 0hr (Figure 4). Supplementation with each of the three blends lead to notable decreases in indole concentrations at 5, 10 and 24hr (Table S6). There was a significant time by treatment interaction (P<0.001, Figure 4) in indole concentrations. Pairwise comparisons between treatment within each time point showed a significant statistical difference in PP compared to F (p=0.044) at 0hr. At 24hr, there were significant statistical differences between FB (p=0.010), PP (p=0.008), and PPFB (p=0.003) respectively compared to F (Figure 4). Additionally, significant differences between PP and PPFB were seen at 24hr (p= 0.002, Figure 4). Pairwise comparisons across time, within treatment, show significant decreases from 0hr to 24hr in FB (p<0.05) and PPFB (p<0.001, Table S7). The linear slopes depicting rates in indole concentrations over time (Figure 4, inset) shows mean concentrations of indole (uM) decreasing across the 24 hours in the three supplemented blends with similarities between FB and PPFB concentrations, while increasing in non-supplemented samples.
3.2.3. Changes in Ammonia Concentrations
Ammonia concentrations increased within F at 0hr, 5hr and 24hr, with negligible decrease at 10hr (Figure 5, Table S6). Supplementation with PP, FB, and PPFB significantly decreased ammonia concentrations after 24hr of fermentation. There was a significant time by treatment interaction (P<0.001) in ammonia concentrations. Significant differences were observed between FB and F (p=0.020) at 24hr. PP supplementation also led to a significant decrease in ammonia production at 24hr compared to the F (p=0.032). At 24hr, ammonia concentrations were significantly lower in PPFB compared to F (p= 0.016) but without any statistical significance when comparing to FB and PP. Pairwise comparisons across time, within treatment, show that significant differences in ammonia concentrations were not observed from 0hr to 24hr (Table S7). Linear slopes (Figure 5, inset) show positive rates of increase in ammonia production over 24hrs for F and PP; however, supplementation with FB led to a slower rate of ammonia production while PPFB displayed a more rapid decreasing rate in ammonia production.
3.2.4. Changes in Antioxidant Capacity
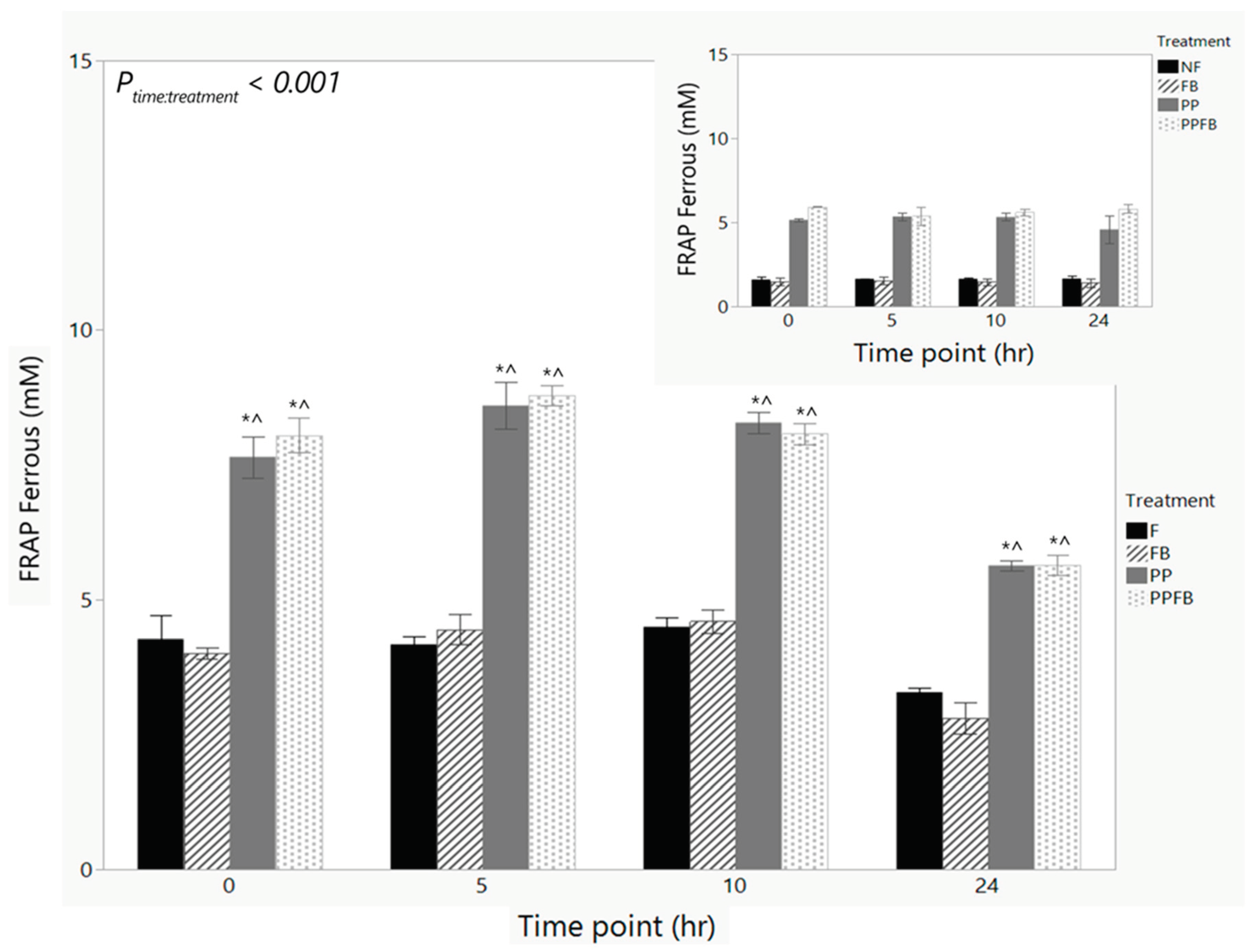
Antioxidant capacity remained constant within F and FB from 0hr to 10hr (Figure 6, Table S6). In PP and PPFB, antioxidant capacity peaked at 5hr and decreased at 10 and 24hr. There was a significant time by treatment interaction (P<0.001) in antioxidant capacity. Pairwise comparisons across treatment show that there were significant differences in PP (p<0.05) and PPFB (p<0.05) respectively, compared to FB and F at each time point from 0hr to 24hr (Figure 6). There were no significant differences between FB and F within each timepoint. There were also no significant differences between PPFB and PP within each time point. The presence of FB in PPFB did not impact antioxidant capacity. Pairwise comparisons across time show that there were significant differences in PP (p=0.012) and PPFB (p=0.043) respectively, between 10hr and 24hr (Table S7). In F, there was a significant decrease in antioxidant capacity at 24hr compared to 0hr (p=0.042). FB did not contribute to significant changes in antioxidant production across time. No significant differences in antioxidant levels were observed in the fecal-deficient, NF samples (Figure 6, inset) across the 24hr fermentation indicating antioxidant capacity changes observed with supplementation is due to the PP and PPFB blends with fecal microbiota present.
4. Discussion
The main aim of this study was to understand the independent and combined effects of supplementing an in vitro model of the large intestine with polyphenol and fiber blends on the growth of beneficial taxa, change in SCFA production, reduction in potentially harmful metabolites, and increase in antioxidant capacity.
Growth dynamics of select gut taxa were driven by supplementation with both FB and PP, impacting eight selected taxa differentially. The selected taxa included some keystone and other lower abundant taxa with well characterized metabolic niches and have been shown to assist in metabolizing complex nutrients from the host diet into metabolites for microbial cross-feeding and host utilization [33]. This included RS degraders Ruminococcus bromii and Blautia coccoides-Eubacterium group; saccharolytic Dorea spp., Lactobacillus spp. and Bifidobacterium spp.; mucin-degrader Akkermansia muciniphila; and butyric acid producers Eubacterium rectale and Faecalibacterium prausnitzii. The taxa changes were seen during the log phases of the fermentation. This was in line with similar in vitro studies comparing the prebiotic effects of individual fibers to a fiber blend [34] with similar changes to select gut bacteria taxa, production of metabolites like SCFAs, and total gas production.
In addition, similar changes were observed when studies supplemented with individual polyphenol sources from our PP blend. Fogliano et al supplemented an in vitro three-stage culture system with cocoa polyphenols leading to the increased abundance of Bifidobacterium spp. and Lactobacillus spp. [35]. Both cranberry and blueberry were included in our polyphenol blend due to their high antioxidant content and diverse polyphenol profiles. Similarly, Ntemiri et al supplemented an in vitro model with polyphenol-rich fractions purified from whole blueberry, leading to significant increases in Bifidobacterium spp. and Feacalibacterium prausnitzii [36]. Solch-Ottaiano et al. showed cranberry polyphenols supplemented in a cross-over study including healthy adults led to increased abundance of Faecalibacterium prausnitzii in subjects [37]. Zhang et al. showed that supplementing fecal fermentations with catechins found in green tea significantly increased the abundance of Bifidobacterium spp., Lactobacillus, and Enterococcus, and increased the production of SCFAs in vitro [38]. These results are consistent with the results seen herein for bacterial taxa changes as a function of PP blend supplementation.
Some studies evaluated polyphenols as part of a whole food, which included fibers naturally present in the food [39]. Our study evaluated the polyphenols and fiber blends both separately and in combination and revealed that the PPFB blend provided an additive positive impact on select taxa when mixing the two blends together in vitro relative to the blends independently. It was important to not immediately assume that the combination of PP and FB would lead to additive effects because there was also a possibility that there could be negative effects associated with combining the two blends due to changes in the competition for resources or potential antimicrobial effects.
Production of beneficial metabolites like short-chain fatty acids (SCFA) are driven by the fermentation of dietary fiber and proteins by gut bacteria. As the primary metabolite of gut bacterial fermentation, SCFAs play a key role in host homeostasis and regulating bacterial community dynamics [40]. In our work, the supplemented blends lead to more significant changes in total SCFA production rather than changes to individual SCFA production. Similar in vitro studies have observed that total concentrations of SCFAs consistently increased when vessels were supplemented with high concentrations of dietary fiber, driven by fiber source and dosage [41]. Of particular interest is butyric acid, which is produced through saccharolytic fermentation by multiple gut commensals including Ruminococcus bromii. Changes in abundance of these taxa have been shown to increase with fiber supplementation accompanied by a related increase in butyric production. FB supplementation was the key driver of increases in total SCFA production. In our study, the increased production of SCFAs in FB supplementation altered butyric concentrations. This is likely linked to the increased abundance of butyric producers such as Rumminococcus upon FB supplementation. It is also well cited in literature that the expected SCFA molar ratio of acetic acid, propionic acid, and butyric acid is 60:20:20 [42]. These ratios were observed in all treatment groups across the 24hr fermentation. Previous in vitro studies showed similar results from fiber supplementation. Wang et al. supplemented in vitro batch cultures with a fructan prebiotic leading to an increase in SCFAs produced [43].
Studies have also shown that polyphenol supplementation benefits the growth of SCFA-producing bacteria in vivo [44] and in vitro [45]. Here, supplementation with PP led to SCFA concentrations approaching significance at the 10hr and 24hr time points relative to F. The lack of significant changes in SCFA production over time warrants further investigation to determine if combining the polyphenols had any antagonistic effects on SCFA production. Changes in SCFA production from PPFB were very similar to those seen in FB with elevated total SCFA and comparable ratios indicating the primary metabolic impact on the community is centered on the FB components. Havlik et al. saw similar effects of mixing fibers with polyphenols in vitro, affecting the production of phenolic metabolites and SCFAs [46].
Like saccharolytic fermentation, protein fermentation by gut bacteria produces SCFAs; however, these metabolites are accompanied by branch-chained fatty acids (BCFA) and pro-inflammatory compounds like amines, ammonia, hydrogen sulfide, indole-compounds, and phenols [47]. More specifically, metabolism of the amino acid tryptophan by gut bacteria generates indole derivatives which can be beneficial (Indole 3-Propionic Acid) or detrimental (Indoxyl Sulfate) to host health [48]. Supplementation with prebiotics have been shown to decrease the production of pro-inflammatory metabolites from different diets in vitro [43]. The increase in saccharolytic activity by gut bacteria counteracts some of the detrimental effects of a proteolytic environment found in various disease states [49]. Both the results for ammonia and indole herein suggest that FB contributed to the decrease in proinflammatory metabolite production. With the additional dietary fiber in FB, a decrease in proinflammatory metabolites was observed most likely due to the lack of amino acids metabolized by proteolytic bacteria [50]. Dietary fiber has been shown to decrease concentrations of ammonia [51] and have an indirect effect on the proliferation of indole-producing bacteria [52].
PP supplementation led to significant decreases in ammonia and indole concentrations compared to F, but less compared to FB supplementation. This could be a result of the short residence time within our batch culture. A fed-batch or continuous culture system may augment the effect, which has been shown in vivo. Goto et al demonstrated that a 6-week supplementation with tea catechins in elderly patients led to significant decreases in fecal ammonia concentrations and other deleterious metabolites [53]. PPFB decreased concentrations of both indole and ammonia similarly to FB in both cases, suggesting that PPFB response is driven by the inclusion of the fiber blend with potential synergistic benefits to decreasing indole concentrations when including the PP blend.
Polyphenols are known for their antioxidant activity while fiber is generally not directly associated with antioxidant production [54]. Antioxidant capacity has a strong link to positive health states by decreasing the presence of reactive oxygen species [55,56]. In this study, FB supplementation did not directly impact the antioxidant capacity. This is supported by other studies regarding an indirect influence as the presence of dietary fiber promotes the growth of beneficial bacteria in the GI tract and the structure of the dietary fibers also have associations to the bioavailability of antioxidant compounds that reach the lower gut [57]. The high antioxidant capacity in both PP and PPFB across all four time points was driven by higher bioavailability to the antioxidant characteristics of PP [58,59]. The decrease in antioxidant capacity at 24hr in PP samples was likely due to further hydrolysis of PP metabolites. Antioxidant capacity was elevated in the PP and PPFB supplemented media and fecal-deficient samples at 0hr likely due to the PP solubilization generating reactive species during the dissolution process. The retention of antioxidant capacity during active growth upon PP blends supplementation is a key finding that shows the ability of PPFB to induce positive gut microbiota states not feasible with FB supplementation independently.
The collective impact of PP and FB, respectively, on select taxa, changes in total amount of metabolic byproducts, decreased production of pro-inflammatory compounds and influence on antioxidant capacity all suggests that PPFB outcomes are not driven by PP or FB but both blends concomitantly. Although a synergistic effect was not evident, the additive outcomes of supplementing with both PP and FB suggests further studies should be considered for microbiome dietary interventions to build toward a healthy, resilient gut microbiome.
5. Conclusions
Blends of select polyphenols and fiber substrates were supplemented into in vitro fecal fermentations to determine the impact of the blends independently and collectively. Supplementation with FB led to increases in Bifidobacterium spp., Lactobacillus spp., and Ruminococcus bromii and, in addition, increases in SCFA production and decreases in indole and ammonia concentrations but did not impact antioxidant production. Supplementation with PP led to increases in Bifidobacterium spp., Lactobacillus spp. and Dorea spp. accompanied by increases in antioxidant concentrations and decreases in indole and ammonia. In general, significant synergistic or antagonistic effects from combining PP and FB were not evident; however, the contributions of both blends provided a beneficial additive effect that suggests that PPFB creates positive effects to gut microbiome states related to inclusion of both PP and FB. The work represents a new supplementation approach of employing tailored blends, rather than individual constituents, for microbiome modulation toward healthy gut states.
6. Study Limitations
The biggest limitation is the gap between in vitro and in vivo relevance. This study doesn’t include host functionality, commonly simulated using intestinal cell culture models. Incorporation of mucin in the growth medium partially simulates the host but it does not imitate the effects of an in vivo mucosal environment on the bacterial community. Passive absorption of metabolites by the human colon is not simulated in our model, and the food supply remains continuous unlike in vivo. Lastly, there is high variability when comparing gut microbe consortiums between individual subjects that adds another layer of complexity. Here, the individual differences were minimized by pooling feces from 30 subjects for a more diverse microbial population within the fermentation inoculum. Although ideal for in vitro studies, the results may not apply across all individuals.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Compositional analysis of four other selected bacteria; Table S1: Constituents in polyphenol and fiber blends and solubilization form; Table S2: Bacterial taxa primer sequences for qPCR; Table S3: Polyphenol and fiber blend supplementation kinetic influence on the abundance of select bacterial taxa - pairwise comparison by treatment across time; Table S4: Differences in total short-chain fatty acid (SCFA) concentrations due to polyphenol and fiber blend supplementation - pairwise comparison by treatment across time; Table S5: Mean short-chain fatty acid (SCFA) concentrations due to polyphenol and fiber blend supplementation; Table S6: Mean concentrations of select microbial metabolites due to polyphenol and fiber blend supplementation; Table S7: Select metabolite pairwise comparison across time by treatment due to polyphenol and fiber blend supplementation.
Author Contributions
Conceptualization, D.A., J.W.S., K.R. and J.P.K.; Methodology, J.W.S., K.R. and J.P.K; Formal analysis, J.A.W. and L.D.; Investigation, L.D., J.A.W., K.K. and J.W.S.; Data curation, J.A.W. and I.P., K.R., and J.P.K; Writing—original draft preparation, J.A.W., I.P., J.W.S.; Writing—review and editing, L.D., K.R., J.P.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the US Defense Health Program; Joint Program Committee 5.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data presented in this study is available on request from the corresponding author.
Acknowledgments
Authors acknowledge Ms. Ava Sargent for her assistance with understanding precedence in literature, Dr. Naomi Balaban for her assistance with molecular measurements, Dr. Syed Hussain for assistance with protocol development, and Ms. Holly McClung and Mr. Nicholes Armstrong for technical assistance during the parent study.
Conflicts of Interest
Authors do not have any conflict of interest for declaration.
References
- Nova, E.; Gomez-Martinez, S.; Gonzalez-Soltero, R. The Influence of Dietary Factors on the Gut Microbiota. Microorganisms 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liang, Q.; Balakrishnan, B.; Belobrajdic, D.P.; Feng, Q.J.; Zhang, W. Role of Dietary Nutrients in the Modulation of Gut Microbiota: A Narrative Review. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: metabolic and health benefits. Br J Nutr 2010, 104 Suppl 2, S1–63. [Google Scholar] [CrossRef]
- Wang, X.; Qi, Y.; Zheng, H. Dietary Polyphenol, Gut Microbiota, and Health Benefits. Antioxidants (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Cheatham, C.L.; Nieman, D.C.; Neilson, A.P.; Lila, M.A. Enhancing the Cognitive Effects of Flavonoids With Physical Activity: Is There a Case for the Gut Microbiome? Front Neurosci-Switz 2022, 16. [Google Scholar] [CrossRef] [PubMed]
- Calatayud, M.; Van den Abbeele, P.; Ghyselinck, J.; Marzorati, M.; Rohs, E.; Birkett, A. Comparative Effect of 22 Dietary Sources of Fiber on Gut Microbiota of Healthy Humans in vitro. Front Nutr 2021, 8, 700571. [Google Scholar] [CrossRef] [PubMed]
- Zaman, S.A.; Sarbini, S.R. The potential of resistant starch as a prebiotic. Crit Rev Biotechnol 2016, 36, 578–584. [Google Scholar] [CrossRef]
- Gowd, V.; Karim, N.; Shishir, M.R.I.; Xie, L.H.; Chen, W. Dietary polyphenols to combat the metabolic diseases via altering gut microbiota. Trends Food Sci Tech 2019, 93, 81–93. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Macfarlane, S. Bacteria, Colonic Fermentation, and Gastrointestinal Health. J Aoac Int 2012, 95, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Duenas, M.; Munoz-Gonzalez, I.; Cueva, C.; Jimenez-Giron, A.; Sanchez-Patan, F.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolome, B. A survey of modulation of gut microbiota by dietary polyphenols. Biomed Res Int 2015, 2015, 850902. [Google Scholar] [CrossRef]
- Tuohy, K.M.; Conterno, L.; Gasperotti, M.; Viola, R. Up-regulating the Human Intestinal Microbiome Using Whole Plant Foods, Polyphenols, and/or Fiber. J Agr Food Chem 2012, 60, 8776–8782. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.Y.; Wang, Y.H.; Lin, Y.; Lang, Y.X.; Li, E.H.; Zhang, X.Y.; Zhang, Q.; Feng, Y.; Meng, X.J.; Li, B. Blueberry polyphenols extract as a potential prebiotic with anti-obesity effects on C57BL/6 J mice by modulating the gut microbiota. J Nutr Biochem 2019, 64, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Bresciani, L.; Di Pede, G.; Favari, C.; Calani, L.; Francinelli, V.; Riva, A.; Petrangolini, G.; Allegrini, P.; Mena, P.; Del Rio, D. In vitro (poly)phenol catabolism of unformulated- and phytosome-formulated cranberry ( Vaccinium macrocarpon ) extracts. Food Res Int 2021, 141. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.Y.; Yang, K.D.; Zhu, J.J. Monitoring the Diversity and Metabolic Shift of Gut Microbes during Green Tea Feeding in an In Vitro Human Colonic Model. Molecules 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Sorrenti, V.; Ali, S.; Mancin, L.; Davinelli, S.; Paoli, A.; Scapagnini, G. Cocoa Polyphenols and Gut Microbiota Interplay: Bioavailability, Prebiotic Effect, and Impact on Human Health. Nutrients 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nature Reviews Gastroenterology & Hepatology 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Birt, D.F.; Boylston, T.; Hendrich, S.; Jane, J.L.; Hollis, J.; Li, L.; McClelland, J.; Moore, S.; Phillips, G.J.; Rowling, M.; et al. Resistant starch: promise for improving human health. Adv Nutr 2013, 4, 587–601. [Google Scholar] [CrossRef]
- Pantoja-Feliciano, I.G.; Soares, J.W.; Doherty, L.A.; Karl, J.P.; McClung, H.L.; Armstrong, N.J.; Branck, T.A.; Arcidiacono, S. Acute stressor alters inter-species microbial competition for resistant starch-supplemented medium. Gut Microbes 2019, 10, 439–446. [Google Scholar] [CrossRef]
- Verbeke, K.; Boobis, A.R.; Chiodini, A.; Edwards, C.; Franck, A.; Kleerebezem, M.; Nauta, A.; Raes, J.; Tuohy, K.; Van Tol, R. Microbial metabolites as potential markers for health benefits of prebiotics. Ann Nutr Metab 2015, 67, 271–272. [Google Scholar]
- Karl, J.P.; Hatch, A.M.; Arcidiacono, S.M.; Pearce, S.C.; Pantoja-Feliciano, I.G.; Doherty, L.A.; Soares, J.W. Effects of Psychological, Environmental and Physical Stressors on the Gut Microbiota. Front Microbiol 2018, 9, 2013. [Google Scholar] [CrossRef]
- McClung, H.L.; Armstrong, N.J.; Hennigar, S.R.; Staab, J.S.; Montain, S.J.; Karl, J.P. Randomized Trial Comparing Consumption of Military Rations to Usual Intake for 21 Consecutive Days: Nutrient Adequacy and Indicators of Health Status. J Acad Nutr Diet 2020, 120, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Karl, J.P.; Armstrong, N.J.; Player, R.A.; Rood, J.C.; Soares, J.W.; McClung, H.L. The Fecal Metabolome Links Diet Composition, Foacidic positive ion conditions, chromatographicallyod Processing, and the Gut Microbiota to Gastrointestinal Health in a Randomized Trial of Adults Consuming a Processed Diet. J Nutr 2022, 152, 2343–2357. [Google Scholar] [CrossRef] [PubMed]
- McInnes, P. Manual of Procedures for Human Microbiome Project Core Microbiome Sampling-Protocol A. 2010, 12.0.
- Pantoja-Feliciano, I.G.; Karl, J.P.; Perisin, M.; Doherty, L.A.; McClung, H.L.; Armstrong, N.J.; Renberg, R.; Racicot, K.; Branck, T.; Arcidiacono, S.; et al. In vitro gut microbiome response to carbohydrate supplementation is acutely affected by a sudden change in diet. Bmc Microbiol 2023, 23. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Macfarlane, S.; Gibson, G.R. Validation of a Three-Stage Compound Continuous Culture System for Investigating the Effect of Retention Time on the Ecology and Metabolism of Bacteria in the Human Colon. Microb Ecol 1998, 35, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, M.; Ramiro-Garcia, J.; Koenen, M.E.; Venema, K. To pool or not to pool? Impact of the use of individual and pooled fecal samples for in vitro fermentation studies. J Microbiol Methods 2014, 107, 1–7. [Google Scholar] [CrossRef]
- Reichardt, N.; Vollmer, M.; Holtrop, G.; Farquharson, F.M.; Wefers, D.; Bunzel, M.; Duncan, S.H.; Drew, J.E.; Williams, L.M.; Milligan, G.; et al. Specific substrate-driven changes in human faecal microbiota composition contrast with functional redundancy in short-chain fatty acid production. Isme J 2018, 12, 610–622. [Google Scholar] [CrossRef]
- Thermo Fisher Scientific. Available online: https://www.thermofisher.com/us/en/home/brands/thermo-scientific/molecular-biology/molecular-biology-learning-center/molecular-biology-resource-library/thermo-scientific-web-tools/qpcr-efficiency-calculator.html (accessed on 15 June, 2022).
- Integrated DNA Technologies. Available online: https://www.idtdna.com/pages/education/decoded/article/calculations-converting-from-nanograms-to-copy-number (accessed on 15, June 2022).
- Patrone, V.; Vajana, E.; Minuti, A.; Callegari, M.L.; Federico, A.; Loguercio, C.; Dallio, M.; Tolone, S.; Docimo, L.; Morelli, L. Postoperative Changes in Fecal Bacterial Communities and Fermentation Products in Obese Patients Undergoing Bilio-Intestinal Bypass. Front Microbiol 2016, 7, 200. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, Y.; Zhang, M.; Pang, X.; Xu, J.; Kang, C.; Li, M.; Zhang, C.; Zhang, Z.; Zhang, Y.; et al. Structural changes of gut microbiota during berberine-mediated prevention of obesity and insulin resistance in high-fat diet-fed rats. Plos One 2012, 7, e42529. [Google Scholar] [CrossRef]
- Blackmore, T.M.; Dugdale, A.; Argo, C.M.; Curtis, G.; Pinloche, E.; Harris, P.A.; Worgan, H.J.; Girdwood, S.E.; Dougal, K.; Newbold, C.J.; et al. Strong stability and host specific bacterial community in faeces of ponies. Plos One 2013, 8, e75079. [Google Scholar] [CrossRef]
- Karl, J.P.; Hatch, A.M.; Arcidiacono, S.M.; Pearce, S.C.; Pantoja-Feliciano, I.G.; Doherty, L.A.; Soares, J.W. Effects of Psychological, Environmental and Physical Stressors on the Gut Microbiota. Frontiers in Microbiology 2018, 9. [Google Scholar] [CrossRef]
- Carlson, J.L.; Erickson, J.M.; Hess, J.M.; Gould, T.J.; Slavin, J.L. Prebiotic Dietary Fiber and Gut Health: Comparing the in Vitro Fermentations of Beta-Glucan, Inulin and Xylooligosaccharide. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Fogliano, V.; Corollaro, M.L.; Vitaglione, P.; Napolitano, A.; Ferracane, R.; Travaglia, F.; Arlorio, M.; Costabile, A.; Klinder, A.; Gibson, G. In vitro bioaccessibility and gut biotransformation of polyphenols present in the water-insoluble cocoa fraction. Mol Nutr Food Res 2011, 55, S44–S55. [Google Scholar] [CrossRef] [PubMed]
- Ntemiri, A.; Ghosh, T.S.; Gheller, M.E.; Tran, T.T.T.; Blum, J.E.; Pellanda, P.; Vlckova, K.; Neto, M.C.; Howell, A.; Thalacker-Mercer, A.; et al. Whole Blueberry and Isolated Polyphenol-Rich Fractions Modulate Specific Gut Microbes in an In Vitro Colon Model and in a Pilot Study in Human Consumers. Nutrients 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Solch-Ottaiano, R.J.; Judkins, T.C.; Matott, S.H.; McDermott, C.E.; Nieves Jr, C.; Wang, Y.; Colee, J.; Tagliamonte, M.S.; Dissanayake, U.; Mai, V.L.; et al. High polyphenolic cranberry beverage alters specific fecal microbiota but not gut permeability following aspirin challenge in healthy obese adults: A randomized, double-blind, crossover trial. J Funct Foods 2022, 99. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, X.L.; Sun, Y.K.; Hu, B.; Sun, Y.; Jabbar, S.; Zeng, X.X. Fermentation in vitro of EGCG, GCG and EGCG3 '' Me isolated from Oolong tea by human intestinal microbiota. Food Res Int 2013, 54, 1589–1595. [Google Scholar] [CrossRef]
- Lakshmanan, A.P.; Mingione, A.; Pivari, F.; Dogliotti, E.; Brasacchio, C.; Murugesan, S.; Cusi, D.; Lazzaroni, M.; Soldati, L.; Terranegra, A. Modulation of gut microbiota: The effects of a fruits and vegetables supplement. Front Nutr 2022, 9, 930883. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.G.; Sencio, V.; Trottein, F. Short-Chain Fatty Acids as a Potential Treatment for Infections: a Closer Look at the Lungs. Infect Immun 2021, 89. [Google Scholar] [CrossRef]
- Fehlbaum, S.; Prudence, K.; Kieboom, J.; Heerikhuisen, M.; van den Broek, T.; Schuren, F.H.J.; Steinert, R.E.; Raederstorff, D. In Vitro Fermentation of Selected Prebiotics and Their Effects on the Composition and Activity of the Adult Gut Microbiota. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat Rev Endocrinol 2015, 11, 577–591. [Google Scholar] [CrossRef]
- Wang, X.D.; Gibson, G.R.; Costabile, A.; Sailer, M.; Theis, S.; Rastall, R.A. Prebiotic Supplementation of In Vitro Fecal Fermentations Inhibits Proteolysis by Gut Bacteria, and Host Diet Shapes Gut Bacterial Metabolism and Response to Intervention. Appl Environ Microb 2019, 85. [Google Scholar] [CrossRef]
- Wu, Z.; Huang, S.; Li, T.; Li, N.; Han, D.; Zhang, B.; Xu, Z.Z.; Zhang, S.; Pang, J.; Wang, S.; et al. Gut microbiota from green tea polyphenol-dosed mice improves intestinal epithelial homeostasis and ameliorates experimental colitis. Microbiome 2021, 9, 184. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Grootaert, C.; Pitart, J.; Vidovic, N.K.; Kamiloglu, S.; Possemiers, S.; Glibetic, M.; Smagghe, G.; Raes, K.; Van de Wiele, T.; et al. Aronia (Aronia melanocarpa) Polyphenols Modulate the Microbial Community in a Simulator of the Human Intestinal Microbial Ecosystem (SHIME) and Decrease Secretion of Proinflammatory Markers in a Caco-2/endothelial Cell Coculture Model. Mol Nutr Food Res 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Havlik, J.; Marinello, V.; Gardyne, A.; Hou, M.; Mullen, W.; Morrison, D.J.; Preston, T.; Combet, E.; Edwards, C.A. Dietary Fibres Differentially Impact on the Production of Phenolic Acids from Rutin in an In Vitro Fermentation Model of the Human Gut Microbiota. Nutrients 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Diether, N.E.; Willing, B.P. Microbial Fermentation of Dietary Protein: An Important Factor in Diet-Microbe-Host Interaction. Microorganisms 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, J. Indole as an intercellular signal in microbial communities. Fems Microbiol Rev 2010, 34, 426–444. [Google Scholar] [CrossRef] [PubMed]
- Makki, K.; Deehan, E.C.; Walter, J.; Backhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Gibson, G.R.; Beatty, E.; Cummings, J.H. Estimation of Short-Chain Fatty-Acid Production from Protein by Human Intestinal Bacteria Based on Branched-Chain Fatty-Acid Measurements. Fems Microbiol Ecol 1992, 101, 81–88. [Google Scholar] [CrossRef]
- Smith, E.A.; Macfarlane, G.T. Enumeration of amino acid fermenting bacteria in the human large intestine: effects of pH and starch on peptide metabolism and dissimilation of amino acids. Fems Microbiol Ecol 1998, 25, 355–368. [Google Scholar] [CrossRef]
- Tennoune, N.; Andriamihaja, M.; Blachier, F. Production of Indole and Indole-Related Compounds by the Intestinal Microbiota and Consequences for the Host: The Good, the Bad, and the Ugly. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Goto, K.; Kanaya, S.; Ishigami, T.; Hara, Y. Effects of tea polyphenols on fecal conditions, part 2. The effects of tea catechins on fecal conditions of elderly residents in a long-term care facility. J Nutr Sci Vitaminol 1999, 45, 135–141. [Google Scholar] [CrossRef]
- Mezes, M.; Erdelyi, M. [Antioxidant effect of the fibre content of foods]. Orv Hetil 2018, 159, 709–712. [Google Scholar] [CrossRef]
- Rajoka, M.S.R.; Thirumdas, R.; Mehwish, H.M.; Umair, M.; Khurshid, M.; Hayat, H.F.; Phimolsiripol, Y.; Pallares, N.; Marti-Quijal, F.J.; Barba, F.J. Role of Food Antioxidants in Modulating Gut Microbial Communities: Novel Understandings in Intestinal Oxidative Stress Damage and Their Impact on Host Health. Antioxidants-Basel 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Perez-Burillo, S.; Rufian-Henares, J.A.; Pastoriza, S. Towards an improved global antioxidant response method (GAR plus ): Physiological-resembling in vitro digestion-fermentation method. Food Chemistry 2018, 239, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Palafox-Carlos, H.; Ayala-Zavala, J.F.; Gonzalez-Aguilar, G.A. The role of dietary fiber in the bioaccessibility and bioavailability of fruit and vegetable antioxidants. J Food Sci 2011, 76, R6–R15. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid Med Cell Longev 2009, 2, 270–278. [Google Scholar] [CrossRef]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24. [Google Scholar] [CrossRef]
Figure 1.
Schematic describing an overview of the experimental workflow. CCM = Complex Colonic Medium.
Figure 1.
Schematic describing an overview of the experimental workflow. CCM = Complex Colonic Medium.
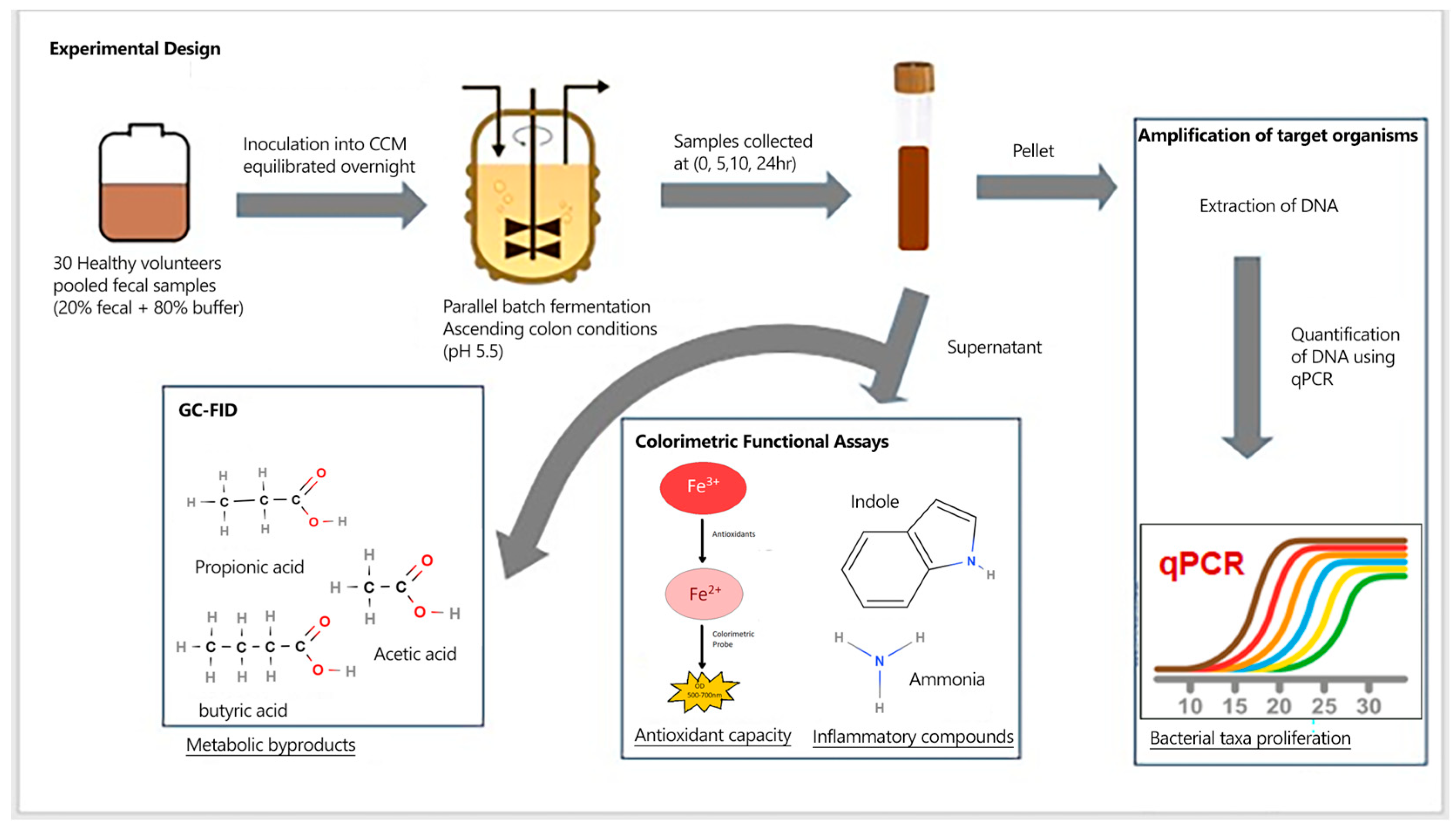
Figure 2.
Compositional analysis of selected bacterial taxa and abundance changes over time due to supplementation. Bifidobacterium spp. (A), Lactobacillus spp. (B), Dorea spp. (C), Ruminococcus bromii (D). As fermentation residence time increases, differential changes occur within the community. F = non-supplemented; PP= polyphenol blend; FB = fiber blend, PPFB – PP and FB blend. Data are Mean ± SEM (n = 3).
Figure 2.
Compositional analysis of selected bacterial taxa and abundance changes over time due to supplementation. Bifidobacterium spp. (A), Lactobacillus spp. (B), Dorea spp. (C), Ruminococcus bromii (D). As fermentation residence time increases, differential changes occur within the community. F = non-supplemented; PP= polyphenol blend; FB = fiber blend, PPFB – PP and FB blend. Data are Mean ± SEM (n = 3).
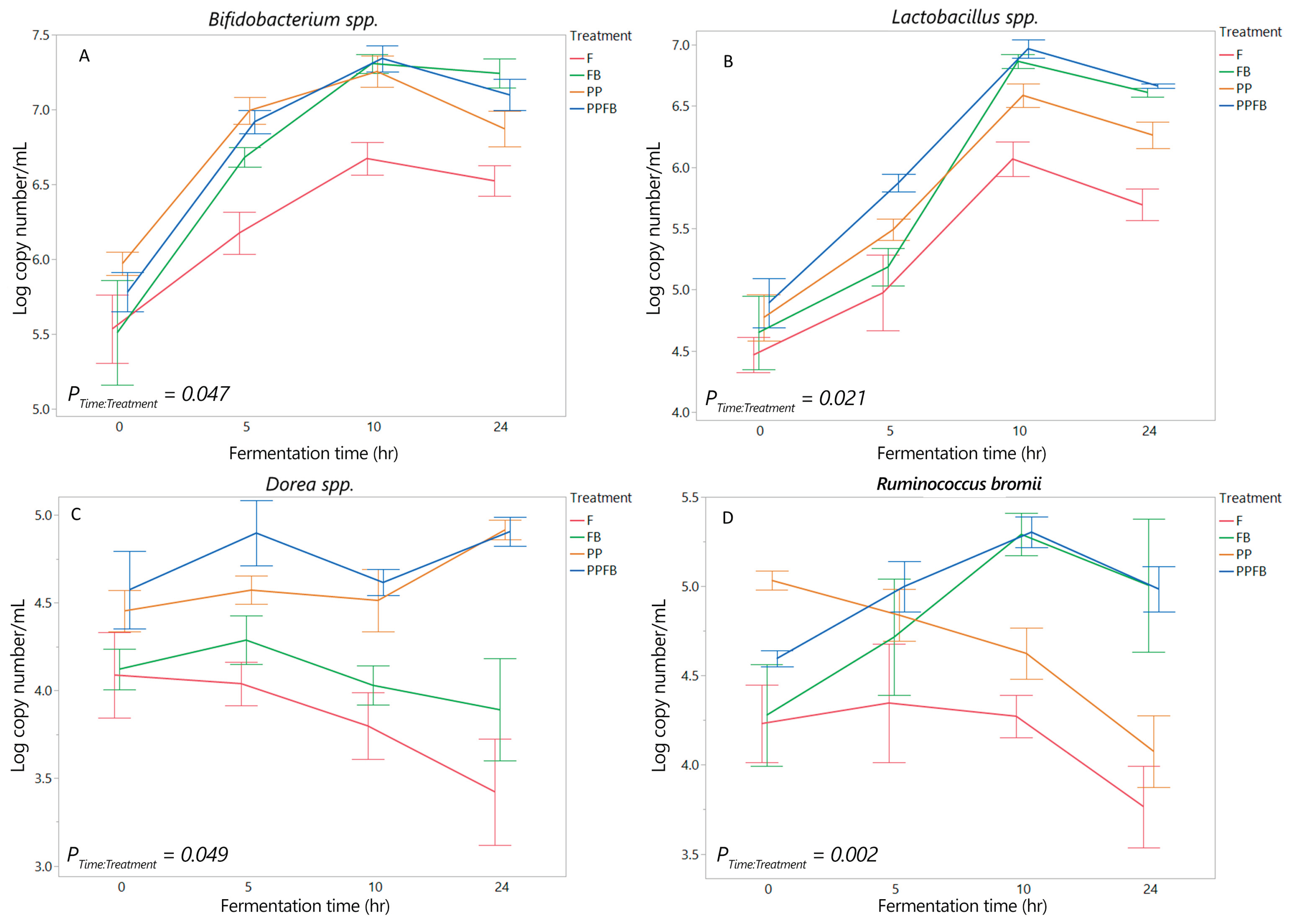
Figure 3.
Mean SCFA concentrations over fermentation residence time during supplementation (n = 3). As fermentation increases, FB blend shows a marked increase in total SCFA, similar to PPFB blend. *Significant differences of each treatment relative to F at the same time point (p<0.05). F = non-supplemented; PP= polyphenol blend; FB = fiber blend, PPFB = PP and FB blend.
Figure 3.
Mean SCFA concentrations over fermentation residence time during supplementation (n = 3). As fermentation increases, FB blend shows a marked increase in total SCFA, similar to PPFB blend. *Significant differences of each treatment relative to F at the same time point (p<0.05). F = non-supplemented; PP= polyphenol blend; FB = fiber blend, PPFB = PP and FB blend.
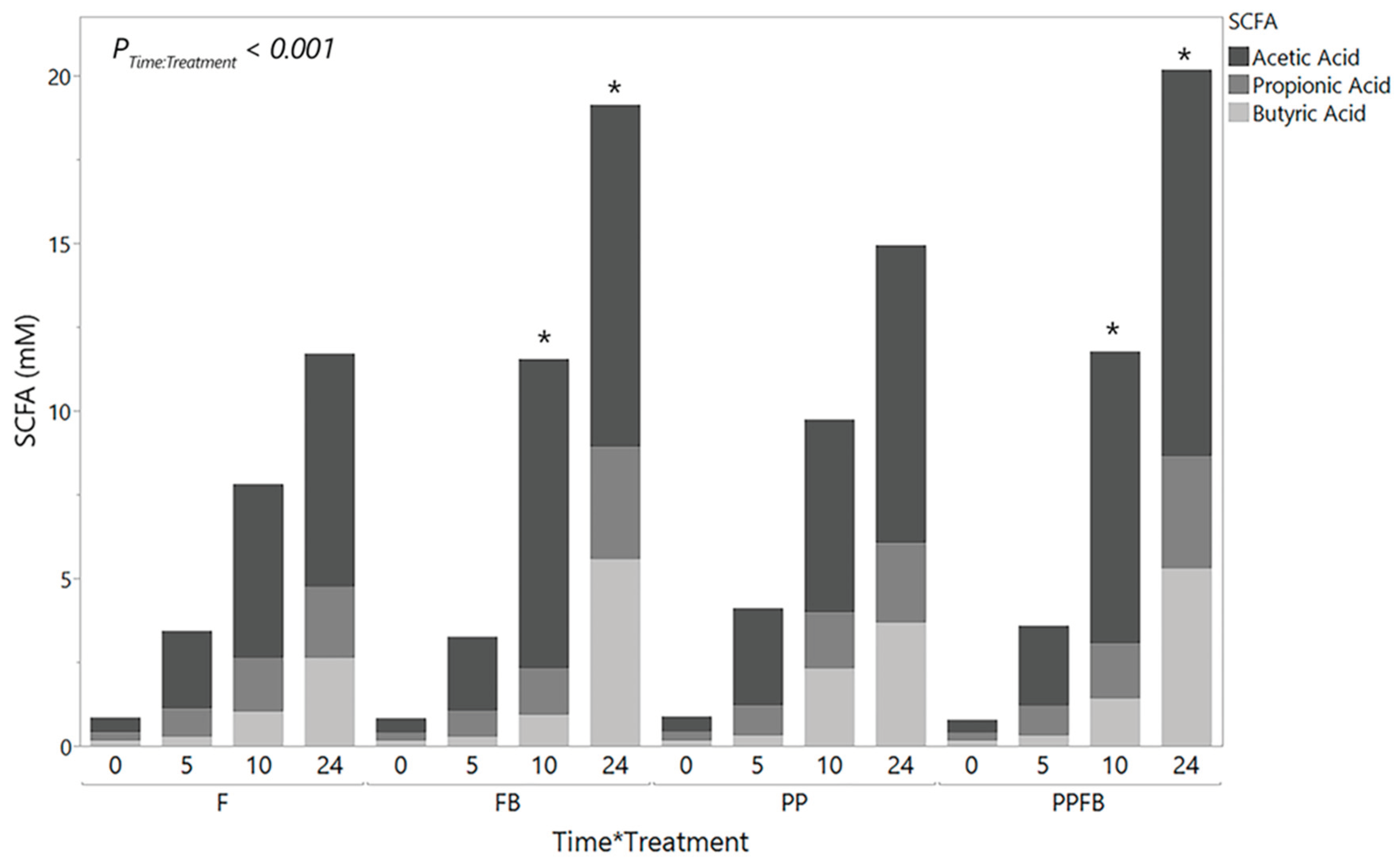
Figure 4.
Net changes in indole concentration over time during supplementation including the corresponding linear slope analysis (inset). As fermentation residence time increases, all three blends show a marked decrease in indole, which are statistically different than F at 24hr. Linear slopes show differences in indole production rate for the FB and PPFB blends relative to F. *Significant differences across treatment compared to F at the same time point (p<0.05). ^Significant differences compared to PP at the same time point (p<0.05). Data are mean ± SEM (n = 3). F = non-supplemented; PP = polyphenol blend; FB = fiber blend, PPFB = PP and FB blend.
Figure 4.
Net changes in indole concentration over time during supplementation including the corresponding linear slope analysis (inset). As fermentation residence time increases, all three blends show a marked decrease in indole, which are statistically different than F at 24hr. Linear slopes show differences in indole production rate for the FB and PPFB blends relative to F. *Significant differences across treatment compared to F at the same time point (p<0.05). ^Significant differences compared to PP at the same time point (p<0.05). Data are mean ± SEM (n = 3). F = non-supplemented; PP = polyphenol blend; FB = fiber blend, PPFB = PP and FB blend.
Figure 5.
Net changes in ammonia concentration over time during supplementation including the corresponding linear slope analysis (inset). As fermentation residence time increases, FB and PPFB blends show a marked decrease in pro-inflammatory marker ammonia that is statistically different to F. Linear slopes show a dramatic rate reduction of ammonia production for PPFB relative to F. *Significant differences across treatment compared to F at same time point (p<0.05). Data are mean ± SEM (n = 3). F = non-supplemented; PP = polyphenol blend; FB = fiber blend, PPFB = PP and FB blend.
Figure 5.
Net changes in ammonia concentration over time during supplementation including the corresponding linear slope analysis (inset). As fermentation residence time increases, FB and PPFB blends show a marked decrease in pro-inflammatory marker ammonia that is statistically different to F. Linear slopes show a dramatic rate reduction of ammonia production for PPFB relative to F. *Significant differences across treatment compared to F at same time point (p<0.05). Data are mean ± SEM (n = 3). F = non-supplemented; PP = polyphenol blend; FB = fiber blend, PPFB = PP and FB blend.
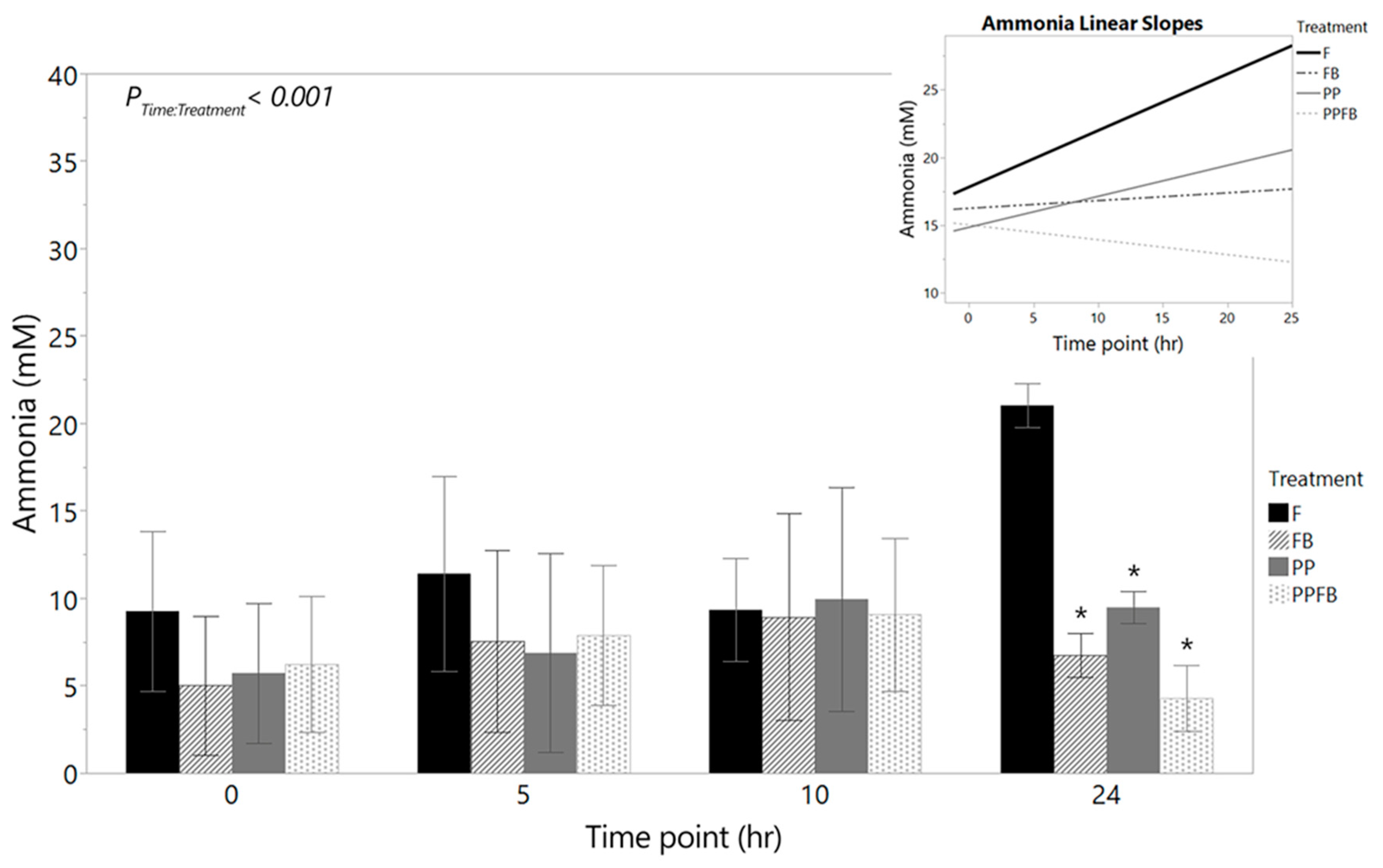
Figure 6.
Change in antioxidant capacity over time during supplementation with corresponding capacity in NF samples (inset). As fermentation residence time increases, concentrations of antioxidant marker remain high in PP and PPFB with a marked decrease at 24hr with statistically significant differences to F. FB shows no significant differences to F. NF samples show no changes in antioxidant capacity across the fermentation. *Significant differences across treatment compared to F at the same time point (p<0.05). ^Significant difference compared to FB at same time point (p<0.05) Data are mean ± SEM (n = 3). F = non-supplemented; PP= polyphenol blend; FB = fiber blend, PPFB = PP and FB blend; NF = fecal-deficient.
Figure 6.
Change in antioxidant capacity over time during supplementation with corresponding capacity in NF samples (inset). As fermentation residence time increases, concentrations of antioxidant marker remain high in PP and PPFB with a marked decrease at 24hr with statistically significant differences to F. FB shows no significant differences to F. NF samples show no changes in antioxidant capacity across the fermentation. *Significant differences across treatment compared to F at the same time point (p<0.05). ^Significant difference compared to FB at same time point (p<0.05) Data are mean ± SEM (n = 3). F = non-supplemented; PP= polyphenol blend; FB = fiber blend, PPFB = PP and FB blend; NF = fecal-deficient.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. Approved for public release. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



