
Submitted:
10 May 2023
Posted:
11 May 2023
You are already at the latest version
Abstract
Potato tubers are susceptible to wounding during post-harvest processes, leading to quality decline, perishability and large economic losses. In this study, the potato cultivar, ‘Longshu No.7’, was foliar-sprayed with 3% chitosan (w/v) three times during the pre-harvest period after flowering to evaluate the effect of foliar spraying with chitosan on suberization processing at wounds of harvested potato tubers. Our results demonstrate that foliar-sprayed with chitosan significantly reduced wound-induced fresh weight loss and dry rot disease index by 37.34% and 41.60% on the 28 day after wounding, respectively. Foliar-sprayed with chitosan accelerated the deposition of suberin polyphenolics and lignin at the wound sites of potato tubers with the formation of thicker cell layers. This occurred with increased localized activities of key enzymes in the suberin polyphenolics and lignin pathways, including phenylalanine ammonia lyase, 4-coumaryl-coenzyme A ligase, cinnamyl alcohol dehydrogenase and peroxidase (33.90–64.32%), as well as the contents of cinnamic acid, sinapic acid, flavonoids, lignins and total phenolics (19.70–23.46%) at wound sites of potato tubers on the 7 day after wounding. Our results indicated that foliar application of chitosan accelerated wound-induced suberization of potato tubers and could mitigate post-harvest product damages.
Keywords:
chitosan
; foliar application
; lignin
; potato
; suberin polyphenolics
; wound-induced suberization
1. Introduction
Potato (Solanum tuberosum L.) is the world’s fourth largest food crop and an important raw material in food industrial processing, with an annual production reaching 18 million tons in China (potato grain production is converted to 20% of the fresh weight of potatoes as the standard yield; data from 2021 statistics of the National Bureau of China). About 70–80% of the total production of potato tubers is stored after harvest for consumption over the following year [1]. However, the primary periderm of potato tuber can be fragile and susceptible to peeling, scraping and friction injuries during harvesting, post-harvest transportation and storage operations [2]. The epidermal wounds are main channels for pathogen infection and water loss, leading to a decline in product qualities and increased product decay during storage [3]. In China, the incidence of potato tuber rot during storage was reported to be on average between 10–30% and up to 60%, which represents huge economic losses [4]. Injury to the tuber periderm leads to secondary periderm formation as protection against pathogen infestations and water loss [5]. This healing process involves wound-induced suberization at the wound site, which occurs with the localized synthesis and deposition of suberin polyphenolic (SPP) and lignin. However, this process can take 2-3 weeks to complete, which is sufficient time for substantial water loss, opportunistic pathogen entry, disease incidence and product loss during storage [5]. The study of methods to accelerate wound-induced suberization is therefore of economic importance [3,6].
Chitosan, a deacetylation product of chitin (poly-β-(1, 4)-N-acetyl-D-glucosamine) which is widely found in the exoskeletons of crustaceans, insects and the cell walls of fungi. Chitosan functions as elicitors in the plant system for the induction of antimicrobial activities and innate disease resistance [7,8]. Chitosan is non-toxic, edible and biodegradable, and has been commercially registered for production in some counties for use in food preservation [9]. Reports have shown that soaking or coating treatments of plant products by chitosan can effectively control the occurrence of postharvest decay in some fruits and vegetables [10,11,12]. Significantly, some studies have shown that pre-harvest foliar application of chitosan can also effectively enhance plant stress resistance and reduce the occurrence of field diseases in wheat and maize [13,14]. For fruit and vegetable preservation, Tian et al. (2019) found that foliar spraying of navel oranges with chitosan improved their fruit disease resistance and reduced the incidence of post-harvest fruit decay [15]. Li et al. (2021) reported that pre-harvest spraying of chitosan promoted the accumulation of SPP in muskmelon fruits and enhanced the rate of wound healing at the fruit surface [16]. Chitosan treatment also induced lignin production in suspension cells of Pinus elliottii and callus of Oryza sativa [17,18]. However, the effect of foliar application of chitosan on wound-induced suberization of potato tuber after harvest has not been reported. In this study, S. tuberosum L. cv. Longshu No.7 was used to explore the effects of pre-harvest foliar spraying of chitosan on the suberization of wounded tubers after harvest, the underlying biochemical mechanism involved, and to provide a theoretical basis for the use of this treatment to maintain product quality during commercial potato tuber storage.
2. Materials and Methods
2.1. Materials
Chitosan (poly-β-(1,4)-2-amino-D-glucose) with a degree of deacetylation of ≥ 90% was purchased from WN Group of Publishers Ltd. (Mansouriah block1, Paris, France). 1,3,5-Trihydroxybenzene was purchased from Beijing Regen Biotechnology Co. Ltd. Standards for cinnamic acid and sinapic acid were purchased from Yuanye Biotechnology Co. Ltd.
Fusarium sambucinum was isolated from potato tissues exhibiting typical symptoms of dry rot. The identity of the pathogen was verified from its rDNA-ITS sequence and the pathogenicity was verified in potato tubers according to Koch's rules. The pathogen was preserved in Potato Dextrose Agar medium (PDA) at 4 °C and cultured on PDA medium for 7 days at 28 °C before use.
2.2. Experimental design
Virus-free seed potatoes of S. tuberosum L. cv. Longshu No.7, provided by the Potato Institute of the Gansu Academy of Agricultural Sciences (Lanzhou, China), were planted in an open field on April 20, 2019 in Nangou Village, Huichuan Town, Weiyuan County, Dingxi City, Gansu Province (35°06′30ʺ N, 103°58′15 ʺ E, 2260 m above sea level). Potatoes were planted in six rows per plot, with an inter-row spacing of 60 cm and plant spacing of 30 cm, and 210 plants were sown in each plot. Potato plants were sprayed evenly (1 L/30 plants) by hand sprayer with either water (control) or 3% (w/v) chitosan at the flowering stage, the tuber bulking stage and 2 weeks before harvesting. The experiment was conducted in a completely randomized group arrangement with three replications [19]. Tubers were developed to maturity and harvested on October 10, 2019, dried in the sun for 4 h, then packed into standard corrugated cartons. The tubers used for physiological and biochemical analyses were stored refrigerated (5 ± 2 °C, RH 80–90%) until use.
2.3. Methods
2.3.1. Artificial wounding of potato tubers
Three batches of 300 potato tubers from each of the chitosan and control treatments of similar size, free of mechanical damage, pests and diseases were selected, rinsed under running water and soaked in 1% sodium hypochlorite for 10 min. After air drying, the periderm was cut along the equatorial plane with a stainless steel blade to a depth of 0.3–0.5 mm according to Zheng et al. (2020) [6]. The wounded tubers were then stored in perforated black polyethylene preservation bags in the dark at 20 ± 2 °C with a RH of 65–75%.
2.3.2. Determination of dry rot disease index and loss of fresh weight in wounded potato tubers.
The loss of tuber fresh weight and the disease index of dry rot were determined according to Zheng et al. (2020) [6]. Using sterile water containing 0.01% (w/v) Tween 80, spore suspensions (1 × 106 spores/mL) were prepared from 7 day old F. sambucinum solid PDA cultures. On days 7, 14 and 28 after injury, 20 μL of the F. sambucinum spore suspension were evenly applied at the surface of wounded tubers, which were air-dried, placed in perforated black polyethylene preservation bags and stored in a ventilated storage room in the dark (20 ± 2 °C, RH 75–85%) for 7 days before determining the disease incidence [19]. Ninety tubers were used for each treatment and randomly divided into three groups of 30 as three biological replicates. The severity of dry rot was graded based on the percentage tubers displaying visible fungal on the tuber surface[19]. The disease index was then calculated from the following equation with three biological replicates.
To determine the fresh weight loss, ninety artificially wounded tubers were selected from each treatment group and randomly divided into three biological replicates of thirty tubers for storage under the conditions described above. The fresh weight loss was calculated according to the following equation 7, 14 and 28 days after wounding. The experiment was repeated three times.
2.3.3. Microscopic observation of SPP and lignin deposition at the wound site of potato tubers
The vertical wound surface of the tuber was cut into 0.2–0.3 mm slices of ca. 1 cm2 using a sterile scalpel, which were washed three times with distilled water to remove starch particles. Lignin was stained by the phloroglucinol-hydrochloric acid method [6] and observed using a microscope (CX21FS1C Olympus, Japan). The deposition of SPP was monitored from its autofluorescence using a fluorescence microscope (Shimadzu RF-5301 PC, Tokyo, Japan), with the excitation wavelength and the emission wavelength at 340-390 nm and 420 nm, respectively [20]. IS Capture software (Tucsen, China)were used to determined the thickness of the cell layer of lignin and SPP [6].
2.3.4. Sample collection
The suberized tissues sampling were collected from the wounded site after 0, 1, 3, 5 and 7 days of artificial injury according to the method of Ge et al. (2021) with some modification [19]. Briefly, the sample was taken 3 mm around and below the wound site. Samples were snap-frozen in liquid nitrogen and ground into powder with a grinding mill (IKA M20, IKA-Werke GmbH & Co. KG, Germany), then stored at −80 ℃ until further analysis.
2.3.5. Determination of enzyme activities in SPP and lignin anabolism
For the determination of the relative activities of phenylalanine aminolase (PAL), 4-coumaroyl-coenzyme A ligase (4CL), cinnamyl alcohol dehydrogenase (CAD) and peroxidase (POD), 0.5 gram (FW) of sampled tissue was homogenized on ice with 2 mL of extraction reagents and was centrifuged at 4 ℃ and 10,000 g for 10 min, the supernatant used as the enzyme crude extract. The activities of enzyme were analyzed with microplate assay kits as per the manufacturer’s instructions (Comin Bio. Co., Ltd., Suzhou, China). Crude extracts were placed on ice and tested within 20 minutes. 10 μL crude extract and 190 μL reaction reagent were added and mixed as the reaction system for measuring enzyme activity. The extraction and reaction reagents were provided by kits. Colorimetric determination was made by ultraviolet & visible spectrophotometer (Shimadzu UV-2450, Tokyo, Japan). Using BSA as a standard, the protein content of tissue extracts (mg.mL-1) was determined with the Bradford assay [21]. The activity of enzyme was expressed as U activity mg-1 protein. One unit of PAL and POD enzymes was defined as that required to achieve an increase in the absorbance of 290 nm of 0.1.min-1.mL-1. One unit of 4CL was defined as that required for the production of 1 nmoL of 4-coumaryl coenzyme A per minute and one unit of CAD enzyme activity was defined as that required for the generation of 1 nmol of Nicotinamide Adenine Dinucleotide Phosphate per minute. All enzyme activities presented were determined from three biological replicates.
2.3.6. Determination of metabolic contents of Suberization
The content of phenolic acid monomers (cinnamic acid, sinapic acid) was determined according to Gruz et al., (2008) with minor modifications [22]. Briefly, 1 g of the frozen tissue homogenate was extracted by ultrasonication in 3 mL 70% (v/v) methanol for 30 min and centrifuged twice at 8,000 × g for 20 min. The supernatant was concentrated in vacuum concentrator (EYELA UT2000, Tokyo, Japan) and dissolved in 1 mL of a 70:30:1 mixture of methanol, ultrapure water, and glacial acetic acid, respectively and filtered through a 0.22 μm nylon filter membrane (Biosharp, Shushan District, China). Quaternary gradient ultrafast liquid chromatography (ACQUITY Arc, Waters, USA) and Symmetry® C18 column (4.6 mm × 250 mm, 5 μm) were used to analyze the filtrate. The analysis conditions used were as described in Zhu et al. (2022) [20], where cinnamic and sinapic acids detected at 276 nm and 325 nm, respectively, and identified from their retention times relative to those of their pure standards. The content of these phenolic acid monomers was calculated from their standard curves and was expressed as μg.g-1 (FW). The relative contents of lignin, total phenols and flavonoids were measured according to Ge et al. (2021)[19], where the contents of lignin and total phenols were expressed as OD280.g-1 (FW), and the content of flavonoid was expressed as OD325.g-1 (FW)
2.4. Statistical analysis
All data are presented as the average of at least 3 biological replicas ± SD or ± SE. Significance testing used Duncan's multiple difference significance analysis with SPSS 19.0 software (Chicago, IL, USA), and a P < 0.05 as the threshold. All graphs were produced using Origin 2023 (OriginLab, Northampton, USA).
3. Results
3.1. Foliar spraying of chitosan treatment reduces the effects of wounding on tuber fresh weight loss and dry rot development
Fresh weight loss and dry rot disease development are both promoted by tuber wounding. The loss of fresh weight in wounded tubers gradually increased with increasing healing time. However, the fresh weight loss in wounded tubers of the control group was significantly higher (5.49%) than that in the chitosan-sprayed group (3.44%) after 28 days of storage, representing a ca. 37.34% reduction in fresh weight loss after wounding (P < 0.05; Figure 1A). Typical dark brown spots symptomatic of dry rot were observed on the surface of injured tubers inoculated with F. sambucinum. In the control group, the disease index of dry rot increased rapidly from the 7th (ca. 10%) to the 28th day of incubation (ca. 73%). In contrast, the dry rot index in wounded tubers of the chitosan-sprayed group was consistently and significantly lower(P < 0.05) than the control group by 28.80%–41.60% (Figure 1B). These results demonstrate that the foliar spraying of chitosan significantly reduced water loss and the development of dry rot in wounded potato tubers after harvested.
3.2. Effect of foliar spraying of chitosan on lignin and SPP accumulation at the wound site of potato tuber
The SPP and lignin layers are important components of the wound periderm in wounded potato tubers. As shown in Figure 2A, tubers from the foliar spraying of chitosan group showed an increased deposition of lignin at the wound site relative to the control as early as 3 days after injury. After 7 days of injury, the lignin content of the treated group increased by 23.46% (Figure 3C) and the thickness of the cell layer by 26.54% compared to the control (Figure 3B). Similarly, tuber wounding induced the localized deposition of SPP which increased over 7 days (Figure 2B). After 7 days, tubers from the chitosan treatment group showed larger increases in SPP fluorescence intensity and fluorescent cell layer thickness (27.27%; Figure 3A) at the wound site relative to control tubers. These results indicated that the foliar spraying of chitosan significantly accekerated the deposition of SPP and lignin at the wounds of potato tubers.
3.3. Effect of foliar spraying of chitosan on key enzyme activities of SPP and lignin anabolism during wound-induced suberization
The activities of key enzymes in both the SPP and lignin synthesis pathways were determined. Prior to wounding, there were no significant differences (P ≥ 0.05) in the activities of PAL, 4CL or POD between the control and chitosan treatment groups. However, the activity of CAD was 17.61% relatively higher in tubers of the chitosan treatment group at the time of harvest (Figure 4).
Following wounding, the activities of the four key enzymes showed a gradual increase in the control group. However, these enzyme activities in tubers of the chitosan treatment group were all higher than those of the control group (Figure 4). Notable relative differences included a larger increase in PAL activity 0–3 days post-injury (Figure 4A) and two peaks of 4CL activity at 3 and 7 days after wounding (Figure 4B). The activity of CAD at the wound site was 48.42% higher than that of the control after 1 day of wounding and was maintained higher over the following 6 days (Figure 4C). POD activities showed a continual increase after wounding, with the highest activity at 7 days (Figure 4D). Relative to the control group, the activities of PAL, 4CL, CAD and POD of the chitosan treatment group were 35.02%, 55.74%, 64.32% and 33.9% higher, respectively, after 7 days of wounding (Figure 4). These results indicated that the foliar spraying of chitosan significantly enhanced the localized mobilization of PAL, 4CL, CAD and POD activities in response to potato tuber wounding after harvest.
3.4. Effects of the foliar spraying of chitosan on the contents of phenolic acid monomers, flavonoids and total phenols in tubers during wound-induced suberization
SPPs are polymerized from different phenolic acid monomers, of which cinnamic acid and sinapic acid are the main constituents. Total phenols and flavonoids are the sum of phenolic acid in tubers. The data on day 0 showed that the foliar spraying of chitosan had no significant effect on the contents of cinnamic acid, sinapic acid, or total phenols (Figure 5A–C), but resulted in a significantly higher flavonoid content relative to the control (P< 0.05; Figure 5D). Following tuber wounding, the contents of phenolic acids and flavonoids showed an increasing trend (Figure 5), and the contents of sinapic acid and cinnamic acid increased rapidly, reaching a maximum on day 5 and day 7, respectively. In the chitosan treated group, the contents of both phenolic acid monomers were significantly higher relative to the control group (Figure 5A,B). In tubers of the chitosan treatment group, the total phenolic content was significantly higher than that of the control group at 3–7 days after wounding (Figure 5C), whereas the flavonoid content was not only relatively higher directly after harvesting, but was induced to higher levels after wounding with further increases over time (Figure 5D). Compared with the control group, the content of cinnamic acid, sinapic acid, total phenols and flavonoids in the tubers from the chitosan-treated group increased by 20.34%, 20.50%, 22.86% and 19.70%, respectively, on day 7 after wounding (Figure 5). The above results indicated that the foliar spraying of chitosan promoted a higher rate of synthesis of cinnamic acid, sinapic acid, total phenols and flavonoids in potato tubers after harvest during wound-induced suberization.
4. Discussion
Pre-harvest treatments including seed dipping or foliar spraying with selected agents has been shown to enhance stress resistance of fruits and vegetables crops during plant cultivation, reduce damages to product quality from aspects of post-harvest processes and improve post-harvest preservation of freshness of fruits and vegetables, all of which have great potential for application in agricultural production [23,24,25,26,27]. It has been known that potato tubers decay easily in water, so harvested tubers are not suitable for preservative treatment with water solution after harvest, potato tubers are not suitable for post-harvest treatments involving their immersion and there is a lack of fumigation treatments in current potato production practices. Our results showed that foliar spraying of chitosan for three times during potato tuber development were effective in reducing fresh weight loss and dry rot disease development in wounded tubers during the post-harvest period (Figure 1). The reduced fresh weight loss is likely to be a direct result of the accelerated formation of the wound periderm [19]. Potato tubers are susceptible to Fusarium spp. and F. sambucinum is the dominant causal pathogens of dry rot during potato storage [4]. The pre-harvest foliar spraying of chitosan significantly impaired the development of dry rot in injured tubers, which can be related to the accelerated suberization of the wound periderm and its provision of an enhanced protective barrier to pathogen infection. Similar post-harvest protective effects have been reported for potatoes using stroby (kresoxim-methyl) [19].
Wound-induced tissue suberization is a complex biological process involving increases in precursor phenylpropanoid synthesis and SPP and lignin formation in potentially dedicated pathways [5,16,28]. The phenylpropanoid pathway is an important secondary metabolic pathway closely related to plant immune system, large amounts of phenolic acids are biosynthesized in this pathway [29]. PAL is the key rate-limiting enzyme in the phenylpropanoid pathway and catalyzes the deamination of phenylalanine to produce trans-cinnamic acid [30], which undergoes a series of catalytic transformations to generate many phenolic compounds, including p-coumarate, caffeic, ferulic acid, and sinapic acid [28]. The phenolic acid monomers are catalyzed to hydroxylated phenolic acids which are polymerized in the presence of POD to produce SPP [31]. Hydroxylated phenolic acids are also catalyzed by 4CL to acetylate to produce p-coumaroyl-CoA, feruloyl-CoA, sinapic-CoA, etc., which are further catalyzed in the presence of CAD to produce monolignols, including p-coumaryl, coniferyl and sinapyl alcohols, which are subsequently polymerized by POD into lignin [28,30,31]. The increased availability of phenolic substrates has been associated with an accelerated deposition of SPP and lignin during the wound-induced suberization process [4,20]. Phenolic acids provide the precursor substrates for the synthesis of SPP and lignin, which provide important waterproofing properties and a protective barrier against pathogen infection in the periderm of potato tubers [32]. It has been shown that chitosan treatment can enhance fruit resistance by activating the activity of defense enzymes related to the lignin synthesis pathway [15,16,23] and promote lignin synthesis and prolong fruit shelf life in citrus [33,34], pears [35], grapes [36], muskmelons [16], and bamboo shoots [37]. These effects occurred in foliar spraying of chitosan in our study. Our results showed that pre-harvest foliar spraying of chitosan enhanced the wound-induction of PAL, 4CL, CAD and POD activities (Figure 4) and promoted increased contents of two phenolic acid monomers in wounded sites of potato tubers (Figure 5A,B). Key enzymes activity of the phenylpropanoid pathway, SPP and lignin anabolism were activated, which were associated with an accelerated SPP and lignin deposition in the suberizing wound periderm (Figure 2 and Figure 3).
Many phenolic acids and the derivatives of phenylpropanoid pathway contribute to the total phenolic content in plant tissues [30,31].Total phenols and flavonoids synthesized can have antioxidant and antimicrobial activities, which inhibit the pathogen expansion in the host [7,23,28,38]. The accumulation of total phenols and flavonoids, which could contribute to the improved fruits and vegetables storage performance through potential antimicrobial and antioxidant activities. Li et al. (2021) found that a pre-harvest chitosan treatment increased the content of flavonoids during wound healing of postharvest melon fruits and reduced the disease index of fruit [16]. Cui et al. (2020) reported that pre-harvest chitosan spray promoted the postharvest synthesis of phenolic acids in apricot with the enhancement the fruit antioxidant capacity [23]. Potato varieties with high contents of total phenols tend to have a higher disease resistance [6]. Pre-harvest chitosan spray induced the accumulation of total phenols and flavonoids in muskmelon and apricot, together with an enhanced fruit resistance to infection and extended shelf-life [16,23]. Flavonoids can have high antioxidant capacity and antimicrobial activity in plant products, which can effectively improve plant resistance to infection by pathogens [39,40]. In this study, the foliar spraying of chitosan led to an increased flavonoid content in tubers at harvest and a relatively larger increase in flavonoid levels after tuber wounding (Figure 5D), as well as increased the content of total phenols after wounded in potaot tubers (Figure 5 C), which are consistent with an improvement in antioxidant capacity, the inhibition of pathogen infection and the reduction of disease index.
The pre-harvest chitosan treatment presented here involves foliar spraying potato plants with 3% (w/v) chitosan at the flowering period, tuber enlarging period and two weeks before tubers harvest. The efficacy of this treatment in the improvement of post-harvest potato tuber performance was successfully field-tested in 2017 and 2018. A similar method was developed for the pre-harvest spraying of muskmelon with chitosan in the author's team [16]. As for the regulatory mechanism underlying the beneficial effects of the foliar spraying of chitosan on the wound-induced suberization of potato tubers, further research at the gene level of priming is needed.
5. Conclusions
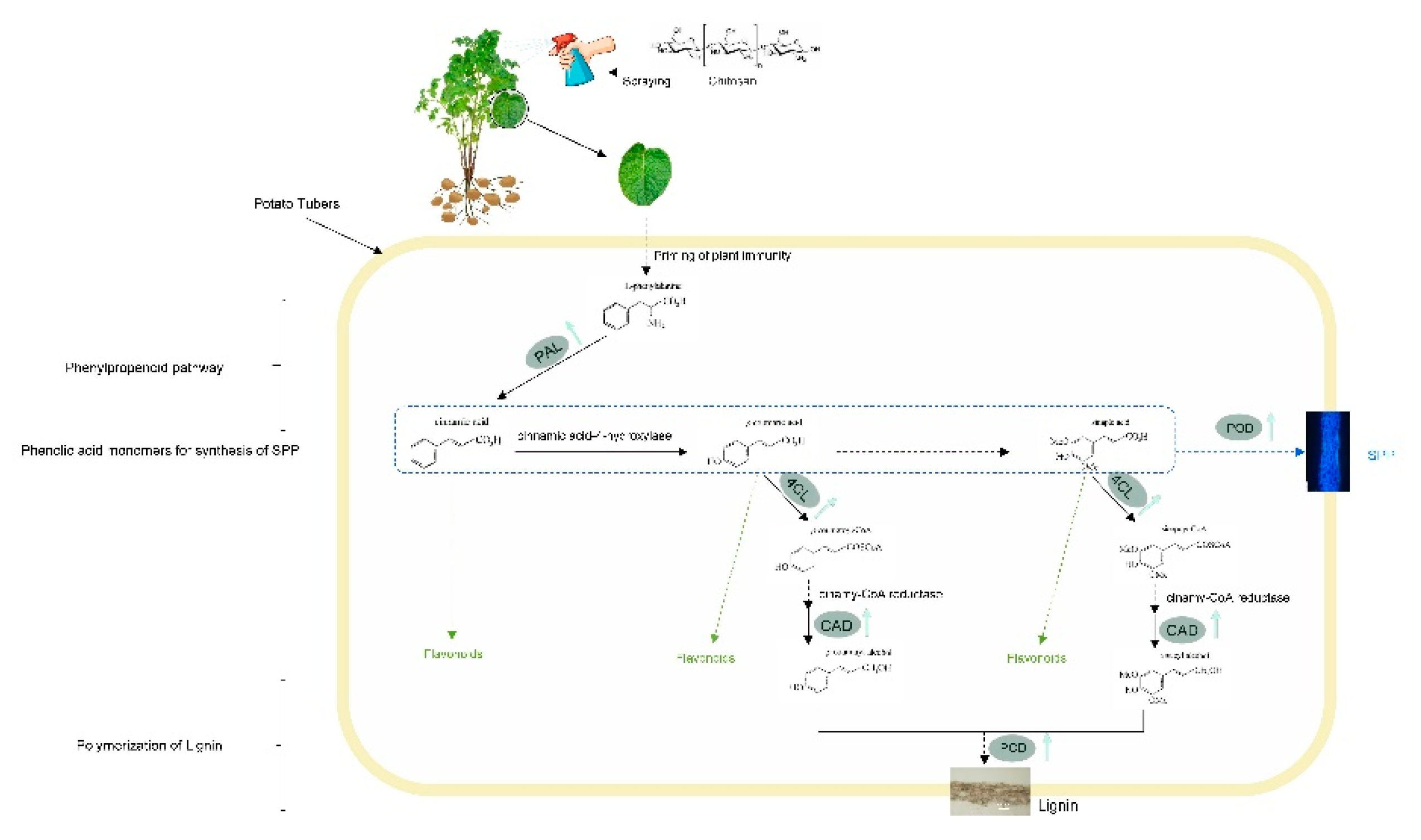
Foliar spraying of chitosan on potato plants enhanced and accelerated wound periderm formation in post-harvest potato tubers, with a reduction in wound-facilitated water loss and dry rot disease development. This occurred with increased wound-induced activities of the phenylpropanoid pathway during potato tuber harvest and the post-harvest wound healing period. The enhanced activities of PAL, 4CL, CAD and POD were associated with an increased content of phenolic compounds at the wound site and an accelerated deposition of SPP and lignin domains of the periderm during wound healing, indicating a causal relationship between the chitosan syrays, the enhanced response of the phenylpropanoid pathway to wounding and accelerated wound periderm formation. The possible mode of foliar spraying of chitosan in accelerating wound periderm formation of potato tubers is illustrated in Figure 6. Our study provides an alternative and eco-friendly treatment for the preservation of commercial potato tuber qualities after harvest.
Author Contributions
Conceptualization, Y.Z. (Yan Zhu) and Y.Z. (Yuanshou Zhao); methodology, W.Z., F.Z. and J.Y.; software, W.S. and W.G.; validation, X.G. and R.C.; formal analysis, F.C. and Y.Z. (Yuanshou Zhao); investigation, X.G. and W.G.; resources, F.C.; data curation, W.S. and J.Y.; writing—original draft preparation, Y.Z. (Yan Zhu) and W.Z.; writing—review and editing, Y.Z. (Yan Zhu) and W.S.; visualization, R.C. and F.Z.; project administration, Y.Z. (Yan Zhu); funding acquisition, Y.Z. (Yan Zhu) and F.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Young Doctor Fund Project of Gansu Provincial Education Department (2022QB-077), the National Natural Science Foundation of China (31760473), and the Excellent Postgraduate Innovation Star Project of Gansu Province (2023CXZX-687).
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
This work was supported by the Young Doctor Fund Project of Gansu Provincial Education Department (2022QB-077), the National Natural Science Foundation of China (31760473), and the Excellent Postgraduate Innovation Star Project of Gansu Province (2023CXZX-687). We thank International Science Editing for editing this manuscript.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Sun, J.; Wang, X.; Huang, Z.; Sun, H.; Cheng, Q.; Zhu, M. Classification and integration of storage and transportation engineering technologies in potato producing areas of China. J. Agric. Sci. Technol. 2017, 18, 710–718. [Google Scholar]
- Campilho, A.; Nieminen, K.; Ragni, L. The development of the periderm: the final frontier between a plant and its environment. Curr. Opin. Plant Biol. 2020, 53, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Bhardwaj, V.; Kaur, K.; Kukreja, S.; Goutam, U. Potato Periderm is the First Layer of Defence against Biotic and Abiotic Stresses: a Review. Potato Res. 2020, 64, 131–146. [Google Scholar] [CrossRef]
- Du, M.; Ren, X.; Sun, Q.; Wang, Y.; Zhang, R. Characterization of Fusarium spp. Causing Potato Dry Rot in China and Susceptibility Evaluation of Chinese Potato Germplasm to the Pathogen. Potato Res. 2012, 55, 175–184. [Google Scholar] [CrossRef]
- Lulai, E.C.; Neubauer, J.D. Wound-induced suberization genes are differentially expressed, spatially and temporally, during closing layer and wound periderm formation. Postharvest Biol. Technol. 2014, 90, 24–33. [Google Scholar] [CrossRef]
- Zheng, X.; Jiang, H.; Bi, Y.; Wang, B.; Wang, T.; Li, Y.; Gong, D.; Wei, Y.; Li, Z.; Prusky, D. Comparison of wound healing abilities of four major cultivars of potato tubers in China. Postharvest Biol. Technol. 2020, 164. [Google Scholar] [CrossRef]
- Al-Mokadem, A.Z.; Alnaggar, A.E.-A.M.; Mancy, A.G.; Sofy, A.R.; Sofy, M.R.; Mohamed, A.K.S.H.; Abou Ghazala, M.M.A.; El-Zabalawy, K.M.; Salem, N.F.G.; Elnosary, M.E.; Agha, M.S. Foliar application of chitosan and phosphorus alleviate the potato virus Y-induced resistance by modulation of the reactive oxygen species, antioxidant defense system activity and gene expression in potato. Agronomy. 2022, 12, e3064. [Google Scholar] [CrossRef]
- Duan, C.; Meng, X.; Meng, J.; Khan, I.H.; Dai, L.; Khan, A.; An, X.; Zhang, J.; Huq, T.; Ni, Y. Chitosan as A Preservative for Fruits and Vegetables: A Review on Chemistry and Antimicrobial Properties. J. Bioresour. Bioprod. 2019, 4, 11–21. [Google Scholar] [CrossRef]
- Miranda-Castro, S.P. Application of Chitosan in Fresh and Minimally Processed Fruits and Vegetables. Academic Press. 2016, 67–113. [Google Scholar]
- Romanazzi, G.; Sanzani, S.M.; Bi, Y.; Tian, S.; Martínez, P.G.; Alkan, N. Induced resistance to control postharvest decay of fruit and vegetables. Postharvest Biol. Technol. 2016, 122, 82–94. [Google Scholar] [CrossRef]
- Niu, X.; Zhu, L.; Xi, L.; Guo, L.; Wang, H. An antimicrobial agent prepared by N-succinyl chitosan immobilized lysozyme and its application in strawberry preservation. Food Control. 2019, 108, 106829. [Google Scholar] [CrossRef]
- Romanazzi, G.; Moumni, M. Chitosan and other edible coatings to extend shelf life, manage postharvest decay, and reduce loss and waste of fresh fruits and vegetables. Curr. Opin. Biotechnol. 2022, 78, 102834. [Google Scholar] [CrossRef] [PubMed]
- Deshaies, M.; Lamari, N.; Ng, C.K.Y.; Ward, P.; Doohan, F.M. The impact of chitosan on the early metabolomic response of wheat to infection by Fusarium graminearum. BMC Plant Biol. 2022, 22, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Hassan, E.O.; Shoala, T.; Attia, A.M.F.; Badr, O.A.M.; Mahmoud, S.Y.M.; Farrag, E.S.H.; EL-Fiki, I.A.I. Chitosan and nano-chitosan for management of Harpophora maydis: Approaches for investigating antifungal activity, pathogenicity, maize-resistant lines, and molecular diagnosis of plant infection. J. Fungi. 2022, 8, 509. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.G.; Ru, P.; Yao, S.; Deng, L.; Zeng, K. Pichia membranaefaciens combined with chitosan treatment induces resistance to anthracnose in Citrus fruit. Food Sci. 2019, 40, 228–237. [Google Scholar]
- Li, Z.; Xue, S.; Xu, X.; Wang, B.; Zheng, X.; Li, B.; Xie, P.; Bi, Y.; Prusky, D. Preharvest multiple sprays with chitosan accelerate the deposition of suberin poly phenolic at wound sites of harvested muskmelons. Postharvest Biol. Technol. 2021, 179. [Google Scholar] [CrossRef]
- Lesney, M.S. Growth responses and lignin production in cell suspensions of Pinuse elliottii 'elicited' by chitin, chitosan or mycelium of Cronartium quercum f. sp. fusiforme. Plant Cell Tissue Org. 1989, 19, 23–31. [Google Scholar] [CrossRef]
- Notsu, S.; Saito, N.; Kosaki, H.; Inui, H.; Hirano, S. Stimulation of Phenylalanine Ammonia-lyase Activity and Lignification in Rice Callus Treated with Chitin, Chitosan, and Their Derivatives. Biosci. Biotechnol. Biochem. 1994, 58, 552–553. [Google Scholar] [CrossRef]
- Ge, X.; Zhu, Y.; Li, Z.; Bi, Y.; Yang, J.; Zhang, J.; Prusky, D. Preharvest multiple fungicide stroby sprays promote wound healing of harvested potato tubers by activating phenylpropanoid metabolism. Postharvest Biol. Technol. 2021, 171, 111328. [Google Scholar] [CrossRef]
- Zhu, Y.; Zong, Y.; Liang, W.; Kong, R.; Gong, D.; Han, Y.; Li, Y.; Bi, Y.; Prusky, D. Sorbitol immersion accelerates the deposition of suberin polyphenolic and lignin at wounds of potato tubers by activating phenylpropanoid metabolism. Sci. Hortic. 2022, 297, 110971. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Gruz, J.; Novák, O.; Strnad, M. Rapid analysis of phenolic acids in beverages by UPLC–MS/MS. Food Chem. 2008, 111, 789–794. [Google Scholar] [CrossRef]
- Cui, K.; Shu, C.; Zhao, H.; Fan, X.; Cao, J.; Jiang, W. Preharvest chitosan oligochitosan and salicylic acid treatments enhance phenol metabolism and maintain the postharvest quality of apricots (Prunus armeniaca L.). Sci. Hortic. 2020, 267, 109334. [Google Scholar] [CrossRef]
- Benkeblia, N.; Tennant, D.P.F.; Jawandha, S.K.; Gill, P.S. Preharvest and harvest factors influencing the postharvest quality of tropical and subtropical fruits. Postharvest Biology and Technology of Tropical and Subtropical Fruits; Yahia, E.M., Ed.; Woodhead Publishing Limited: Sawston, Cambridge CB22 3HJ, UK, 2011; 142e, pp. 112-141. [Google Scholar]
- Wójcik, P. Effects of preharvest sprays of iodine, selenium and calcium on apple biofortification and their quality and storability. PLOS ONE 2023, 18, e0282873. [Google Scholar] [CrossRef]
- Zheng, J.; Chen, H.; Wang, T.; Mustafa, G.; Liu, L.; Wang, Q.; Shao, Z. Quality Improvement of Tomato Fruits by Preharvest Application of Chitosan Oligosaccharide. Horticulturae 2023, 9, 300. [Google Scholar] [CrossRef]
- Fu, E.; Zhang, Y.; Li, H.; Wang, X.; Zhang, H.; Xiao, W.; Chen, X.; Li, L. Chitosan Reduces Damages of Strawberry Seedlings under High-Temperature and High-Light Stress. Agronomy 2023, 13, e517. [Google Scholar] [CrossRef]
- Lulai, E.C.; Campbell, L.G.; Fugate, K.K.; McCue, K.F. Biological differences that distinguish the 2 major stages of wound healing in potato tubers. Plant Signal. Behav. 2016, 11, e1256531. [Google Scholar] [CrossRef]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant. 2010, 3, 2–20. [Google Scholar] [CrossRef]
- Bernards, M.A.; Razem, F.A. The poly(phenolic) domain of potato suberin: a non-lignin cell wall bio-polymer. Phytochemistry 2000, 57, 1115–1122. [Google Scholar] [CrossRef]
- Han, Y.; Yang, R.; Wang, Q.; Wang, B.; Prusky, D. Sodium silicate promotes wound healing by inducing the deposition of suberin polyphenolic and lignin in potato tubers. Front. Plant Sci. 2022, 13, 942022. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Deng, L.; Zhou, Y.; Ming, J.; Yao, S.; Zeng, K. Wound healing in citrus fruit is promoted by chitosan and Pichia membranaefaciens as a resistance mechanism against Colletotrichum gloeosporioides. Postharvest Biol. Technol. 2018, 145, 134–143. [Google Scholar] [CrossRef]
- Gao, Y.; Kan, C.; Wan, C.; Chen, C.; Chen, M.; Chen, J. Quality and biochemical changes of navel orange fruits during storage as affected by cinnamaldehyde -chitosan coating. Sci. Hortic. 2018, 239, 80–86. [Google Scholar] [CrossRef]
- Kou, X.-H.; Guo, W.-L.; Guo, R.-Z.; Li, X.-Y.; Xue, Z.-H. Effects of Chitosan, Calcium Chloride, and Pullulan Coating Treatments on Antioxidant Activity in Pear cv. “Huang guan” During Storage. Food Bioprocess Technol. 2013, 7, 671–681. [Google Scholar] [CrossRef]
- Hu, B.; Lin, L.; Fang, Y.; Zhou, M.; Zhou, X. Application of Chitosan-Lignosulfonate Composite Coating Film in Grape Preservation and Study on the Difference in Metabolites in Fruit Wine. Coatings 2022, 12, 494. [Google Scholar] [CrossRef]
- Yang, H.; Zheng, J.; Huang, C.; Zhao, X.; Chen, H.; Sun, Z. Effects of Combined Aqueous Chlorine Dioxide and Chitosan Coatings on Microbial Growth and Quality Maintenance of Fresh-Cut Bamboo Shoots (Phyllostachys praecox f. prevernalis.) During Storage. Food Bioprocess Technol. 2015, 8, 1011–1019. [Google Scholar] [CrossRef]
- Hahlbrock, K.; Grisebach, H. Enzymic Controls in the Biosynthesis of Lignin and Flavonoids. Annu. Rev. Plant Physiol. 1979, 30, 105–130. [Google Scholar] [CrossRef]
- Cascaes, M.M.; Guilhon, G.M.S.P.; Zoghbi, M.d.G.; Andrade, E.H.A.; Santos, L.S.; da Silva, J.K.R.; Uetanabaro, A.P.T.; Araújo, I.S. Flavonoids, antioxidant potential and antimicrobial activity of Myrcia rufipila mcvaugh leaves (myrtaceae). Nat. Prod. Res. 2021, 35, 1717–1721. [Google Scholar] [CrossRef]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef]
Figure 1.
Effects of the foliar spraying of chitosan on (A) the fresh weight loss and (B) the development of dry rot disease in wounded potato tubers.
Figure 1.
Effects of the foliar spraying of chitosan on (A) the fresh weight loss and (B) the development of dry rot disease in wounded potato tubers.
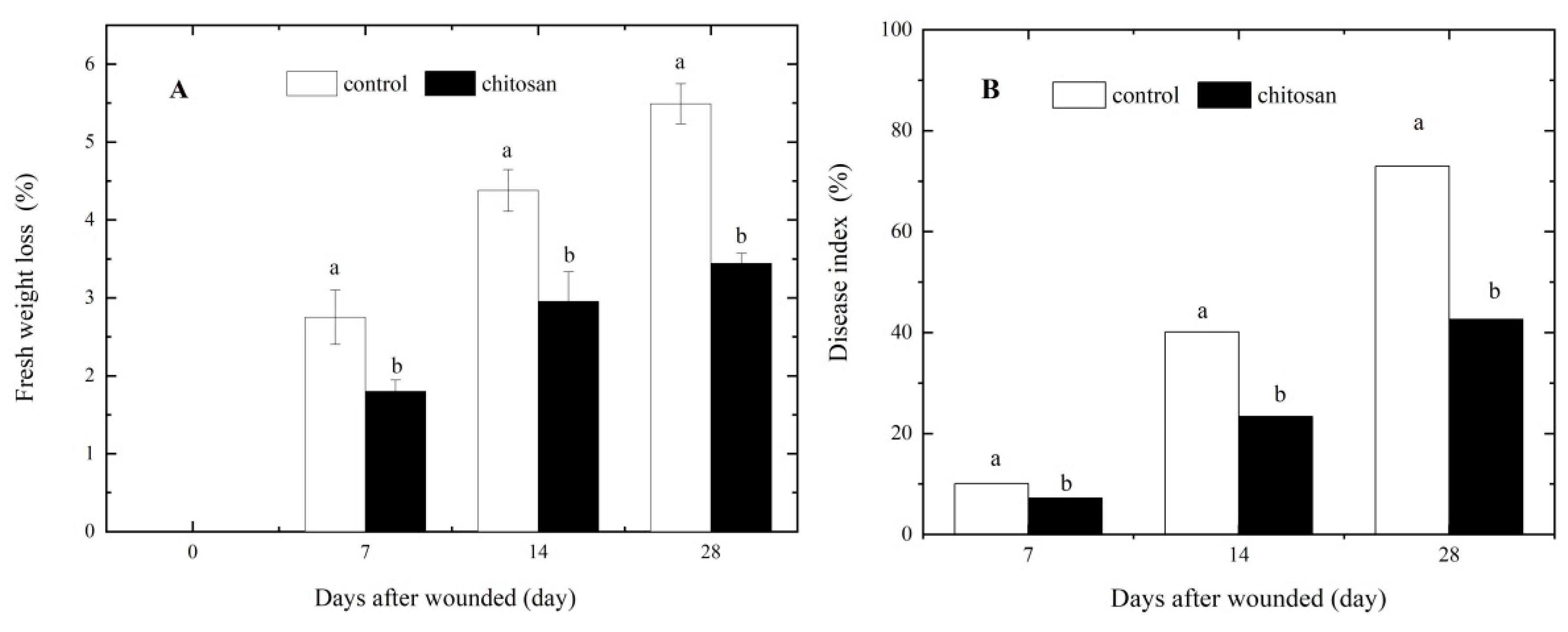
Figure 2.
Effect of foliar spraying of chitosan on the localized accumulation of (A) lignin and (B) SPP in wounded potato tubers. Black arrows indicate deposition of lignin at the wound sites and white arrows indicate deposition of SPP at the wound sites, respectively.
Figure 2.
Effect of foliar spraying of chitosan on the localized accumulation of (A) lignin and (B) SPP in wounded potato tubers. Black arrows indicate deposition of lignin at the wound sites and white arrows indicate deposition of SPP at the wound sites, respectively.
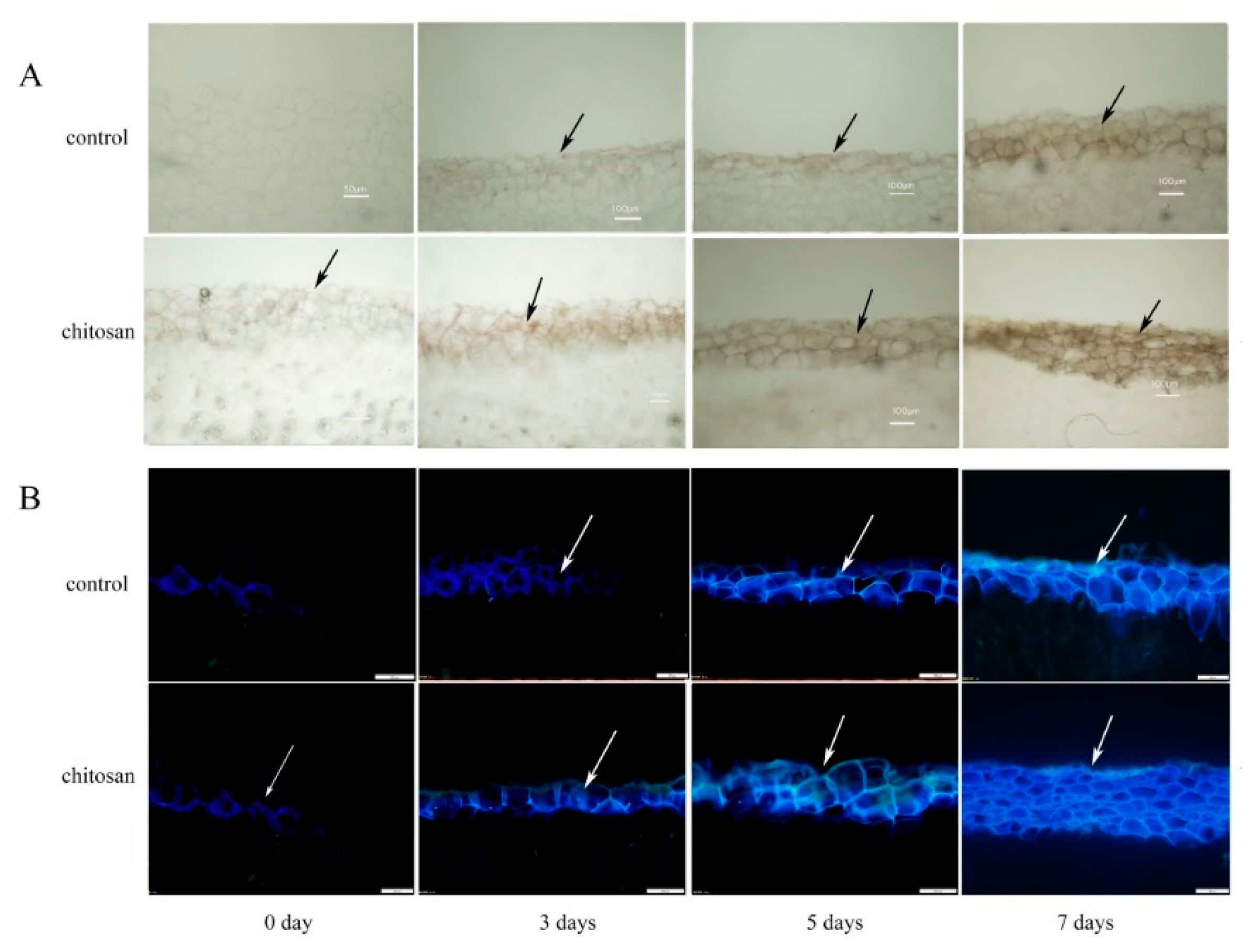
Figure 3.
Effect of foliar spraying of chitosan on (A) the thickness of the SPP cell layer, (B) thickness of the lignin cell layer at the wound site, and (C) the content of lignin. Lowercase letters indicate a significant difference between the different treatment groups (P < 0.05).
Figure 3.
Effect of foliar spraying of chitosan on (A) the thickness of the SPP cell layer, (B) thickness of the lignin cell layer at the wound site, and (C) the content of lignin. Lowercase letters indicate a significant difference between the different treatment groups (P < 0.05).
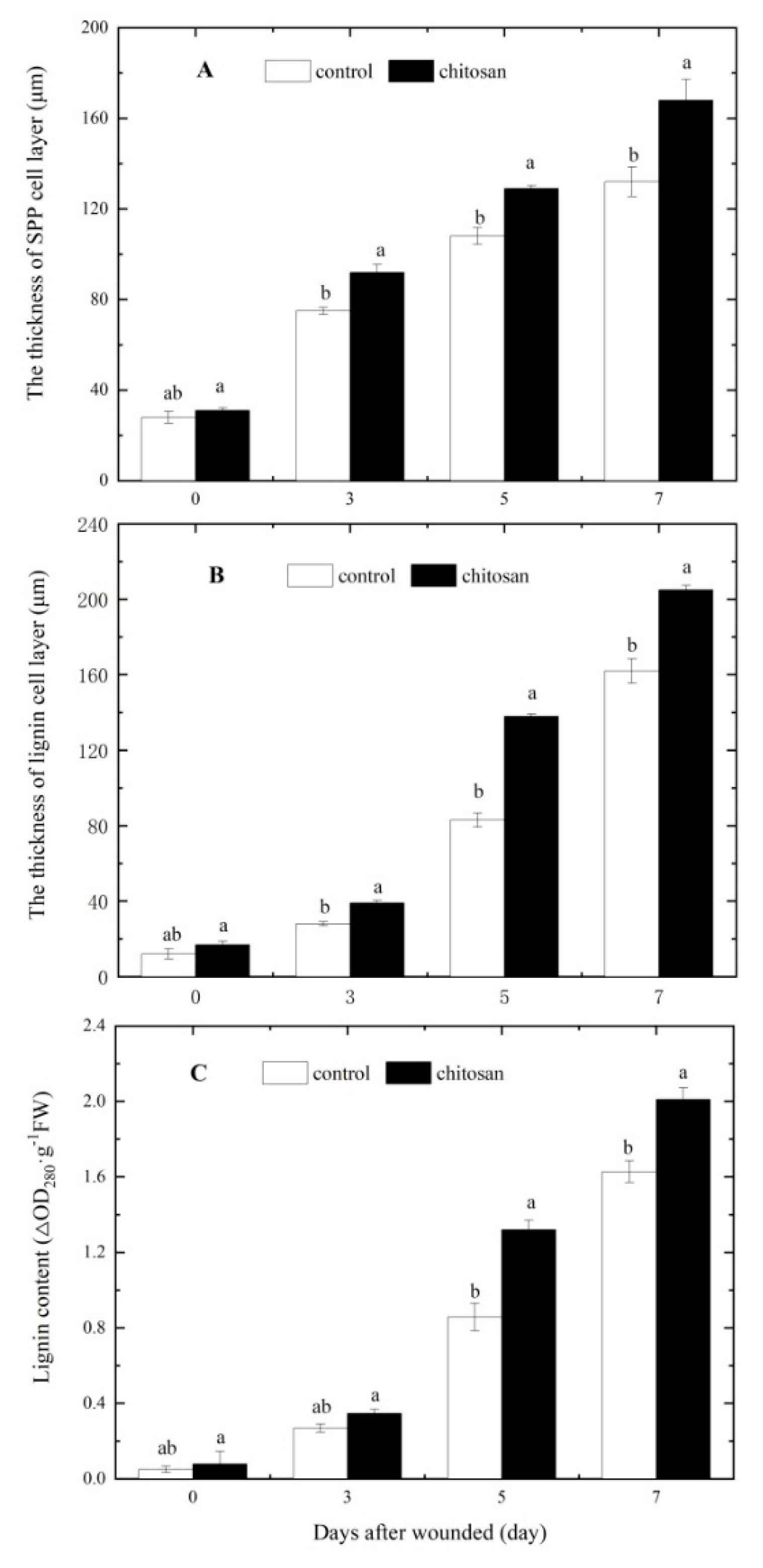
Figure 4.
Effects of the foliar spraying of chitosan on enzyme activities related to SPP and lignin synthesis at the wound site of potato tubers. The activities of (A) PAL, (B) 4CL, (C) CAD and (D) POD. Capital letters indicate significant difference between the control and chitosan treatment (P < 0.05). Lowercase letters indicate significant difference between time points in the same treatment group (P < 0.05).
Figure 4.
Effects of the foliar spraying of chitosan on enzyme activities related to SPP and lignin synthesis at the wound site of potato tubers. The activities of (A) PAL, (B) 4CL, (C) CAD and (D) POD. Capital letters indicate significant difference between the control and chitosan treatment (P < 0.05). Lowercase letters indicate significant difference between time points in the same treatment group (P < 0.05).
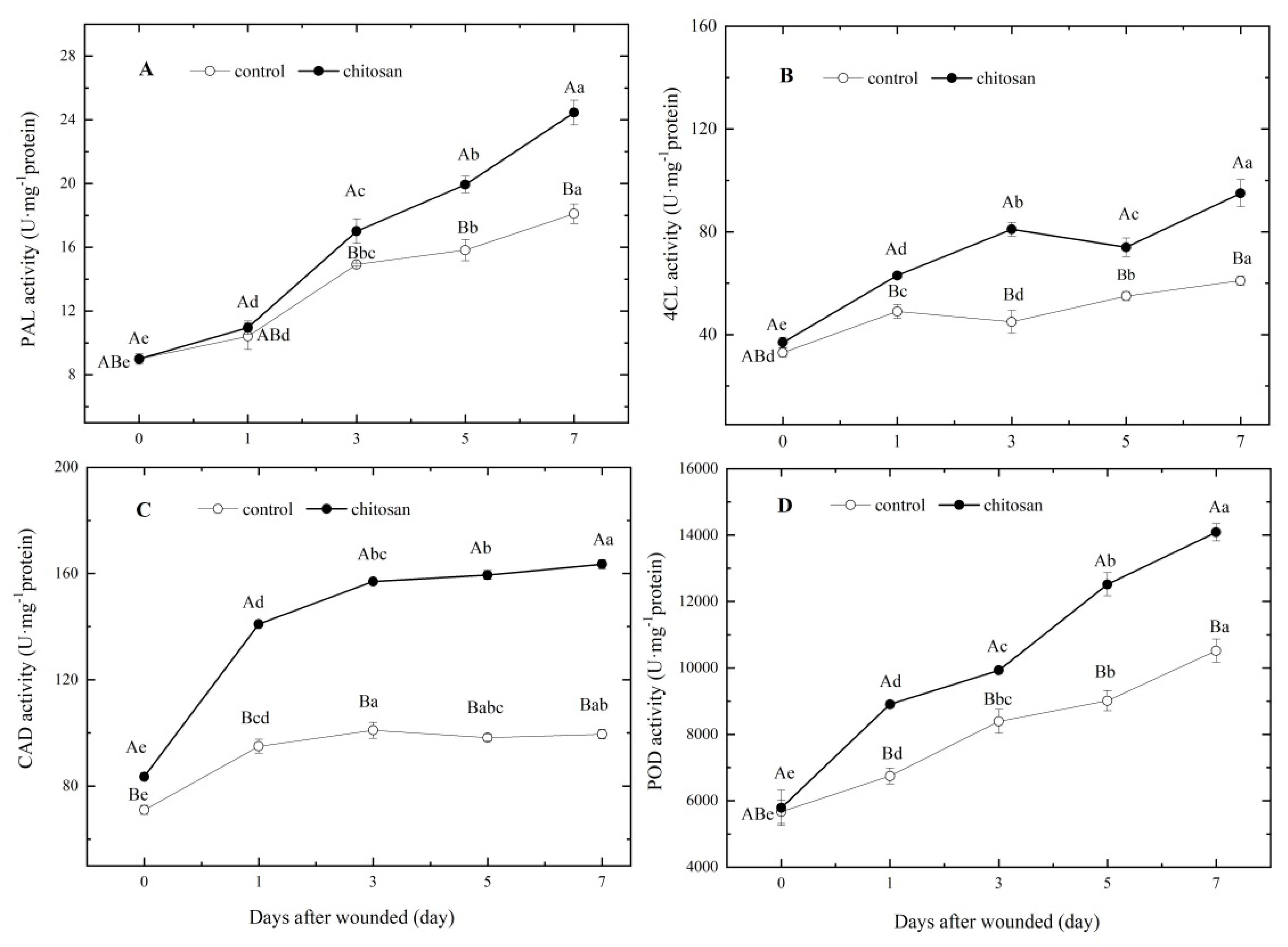
Figure 5.
Effect of foliar spraying of chitosan on the contents of (A) cinnamic acid, (B) sinapic acid, (C) total phenolics and (D) flavonoids at the wound site of potato tubers. Capitals letters indicate significant (P < 0.05) differences between chitosan and control treatments. The lowercase letters indicate significant (P < 0.05) differences between the time points of a treatment group.
Figure 5.
Effect of foliar spraying of chitosan on the contents of (A) cinnamic acid, (B) sinapic acid, (C) total phenolics and (D) flavonoids at the wound site of potato tubers. Capitals letters indicate significant (P < 0.05) differences between chitosan and control treatments. The lowercase letters indicate significant (P < 0.05) differences between the time points of a treatment group.
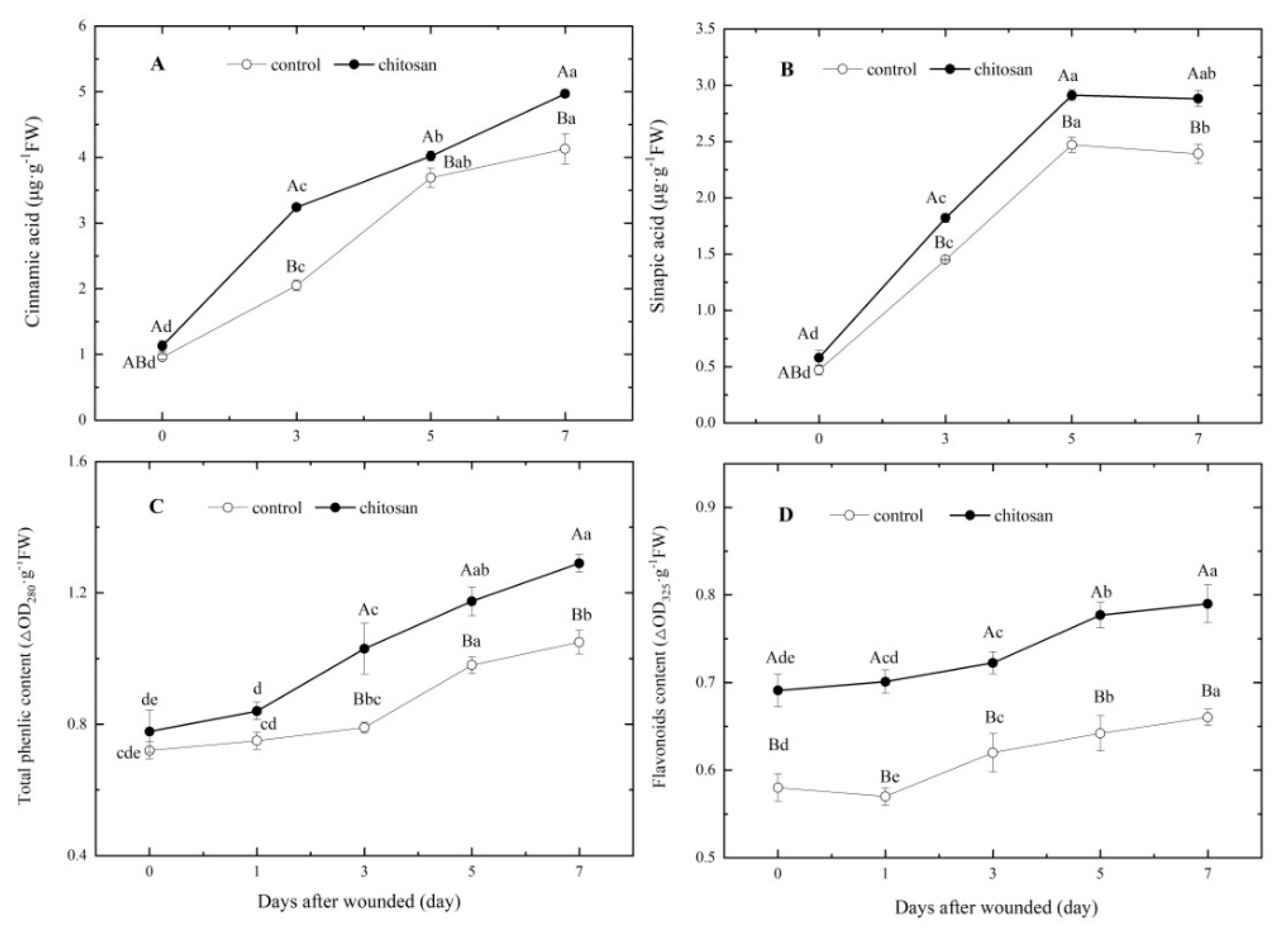
Figure 6.
Foliar spraying of chitosan accelerated the deposition of suberin polyphenolic and lignin at wounds of potato tubers by eliciting phenylpropanoid metabolism, polymerization of SPP and lignin. Reactions denoted by solid lines are known, whereas those denoted by dashed lines are hypothetical metabolic steps catalyzed by multiple enzymes. Black arrows represent the flow of material that syntheizes lignin, blue dashed line in the box and blue arrow represent the flow of material that syntheizes SPP, and green arrows represent the synthesis of flavonoids in the simulation of a potato tubers, respectively. PAL, phenylalanine ammonialyase; 4CL, 4-coumaric acid coenzyme A ligase; CAD, cinnamoyl alcohol dehydrogenase; POD, peroxidase; SPP, Suberin polyphenolic.
Figure 6.
Foliar spraying of chitosan accelerated the deposition of suberin polyphenolic and lignin at wounds of potato tubers by eliciting phenylpropanoid metabolism, polymerization of SPP and lignin. Reactions denoted by solid lines are known, whereas those denoted by dashed lines are hypothetical metabolic steps catalyzed by multiple enzymes. Black arrows represent the flow of material that syntheizes lignin, blue dashed line in the box and blue arrow represent the flow of material that syntheizes SPP, and green arrows represent the synthesis of flavonoids in the simulation of a potato tubers, respectively. PAL, phenylalanine ammonialyase; 4CL, 4-coumaric acid coenzyme A ligase; CAD, cinnamoyl alcohol dehydrogenase; POD, peroxidase; SPP, Suberin polyphenolic.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



