Submitted:
10 December 2024
Posted:
10 December 2024
You are already at the latest version
Abstract
Watermelon is an important fruit widely cultivated in tropical and temperate regions across the world. Particularly in China, both the production and consumption of watermelon lead the world. Watermelon is popular all over the world because it is rich in a variety of nutrients. Despite its widespread appeal, watermelon cultivation is frequently challenged by biological and abiotic stresses, which can significantly impair both yield and fruit quality. Grafting is a technique known to enhance the yield and quality of watermelons, yet the identification of suitable rootstocks and a comprehensive understanding of the cytological, physiological, and biochemical aspects of grafting remain elusive. In this study, we had identified the ‘Heiniu’ pumpkin variety as the optimal rootstock for watermelon grafting. Furthermore, we had determined that the most appropriate stages for scion and rootstock grafting are the one-true-leaf stage and the two-true-leaf stage, respectively. During the grafting process, the necrotic layer of the graft junction gradually dissipated, facilitating a tight connection between the scion and rootstock. Concurrently, there was a notable fluctuation in the levels of key plant hormones and protease activity throughout the healing process of the graft union, suggesting that these factors were pivotal to the success of watermelon grafting. Additionally, our findings indicated that grafting can significantly increase the sugar and titratable acid content within watermelons. This research contributes to a deeper comprehension of the histological and physiological mechanisms underlying watermelon grafting. It offers both theoretical insights and practical guidance essential for the advancement of watermelon grafting techniques.
Keywords:
watermelon
; grafting
; cytological
; physiological
; hormones
; protease
1. Intruduction
Watermelon (Citrullus lanatus Thunb. Matsum and Nakai), the third most consumed fruit worldwide, is extensively cultivated in both tropical and temperate regions. In 2021, the global annual production of watermelons reached an impressive 101.63 million tons. China, Turkey, India, Brazil, and Algeria lead the world in watermelon production. Notably, China's annual watermelon production soared to 60.247 million tons in 2021, accounting for a staggering 59.3% of the global output (FAOSTAT 2021). Watermelon is rich in an array of natural substances beneficial to human health, including various natural sugars, organic acids, ascorbic acid (vitamin C), and lycopene. These components contribute to the growing popularity of watermelon in China, where its health benefits are increasingly recognized.
Watermelon cultivation is a battle against a multitude of biotic stresses, including insect pests like spider mites and diseases such as gummy stem blight [1,2,3]. In addition to these biological threats, watermelons also face a barrage of abiotic stresses throughout their life cycle, such as salinity [4,5], cold [6,7], drought [8,9], and flooding [10,11], all of which can severely compromise yield. Developing stress-tolerant varieties is a promising approach to mitigate these issues; however, the limited availability of germplasm resources and the lengthy process of breeding hinder the progress of creating varieties resistant to both abiotic and biotic stresses.
Grafting technology is a proven asexual reproduction method in plants, which involves joining the scions of superior varieties onto robust rootstocks to enhance resistance and quality. This technique is particularly beneficial for watermelon cultivation, as it can significantly increase yield and stress tolerance. Watermelons grafted onto certain rootstocks have been shown to gain resistance against fusarium wilt from bottle gourd (Lagenaria siceraria) and interspecific hybrid squash (Cucurbita moschata × C. maxima) [12,13]. Additionally, resistance to powdery mildew [14,15], melon necrotic spot virus [16], and crown rot [17], can be imparted through grafting onto wild watermelon (Citrullus mucosospermus, C. lanatus var. citroides), bottle gourd rootstocks (Lagenaria siceraria), and interspecific hybrid rootstocks (Cucurbita maxima × Cucurbita moschata). Grafting not only improves the watermelon's disease resistance but also enhances its tolerance to environmental stresses such as cold [18,19], drought [20,21], salinity [22,23], and heavy metals [24,25]. Beyond stress tolerance, grafting also improves watermelon's photosynthetic efficiency and overall biomass [26].
A variety of cucurbitaceae crops are widely employed as rootstocks in watermelon grafting, including bottle gourd (Lagenaria siceraria), pumpkin (Cucurbita pepo L.), Luffa (Luffa aegyptiaca), and wax gourd (Benincasa hispida). Additionally, wild watermelon species such as Citrullus amarus and Citrullus colocynthis are also used as rootstocks [27]. The compatibility between these rootstocks and watermelons is different, leading to diverse survival rates among grafted seedlings. The compatibility process in grafted plants is intricate, involving a series of anatomical, physiological, and biochemical steps, as outlined by Melnyk [28]. This process can be broken down into four clear stages: The first stage is the attachment of the cut tissues between the scion and rootstock; the second, following attachment, cell differentiation and division initiate at the graft junction; the third, auxin and cytokinin responses are activated, which are crucial for promoting vascular reconnection; ultimately, a stable graft union is established by integrating the rootstock and scion.
In the grafting process, a series of hormones and proteases orchestrate the success of the union. Abscisic acid (ABA) plays a dual role, directly engaging in the wound response and indirectly influencing vascular formation [29,30]. Auxin perception stands as a pivotal element for a successful graft, guiding the early stages of tissue integration [31]. Cytokinins (CKs) contribute to the reconnection of vascular tissues, ensuring the flow of nutrients and signals post-graft [32]. Ethylene signaling is crucial for the reorganization of wounded tissues, facilitating the healing process [33]. Gibberellins (GAs) promote cell expansion in the wounded areas, aiding in the physical union of the scion and rootstock [33]. Beyond hormonal regulation, a suite of proteases is essential for the physical restructuring of cells during grafting. These enzymes, primarily involved in cell wall adhesion and reconstruction, include expansin, β-1,3-glucanase, β-1,4-glucanase, and xyloglucan hydrolase [34]. They are crucial for the breakdown and reformation of cell walls, allowing for the successful integration of the graft.
Grafting significantly influences the morphological characteristics and yield of watermelon. The use of Cucurbita pepo L. and interspecific rootstocks can markedly increase yields [35], as well as alter fruit size, rind thickness, and pulp firmness [36]. Furthermore, grafting affects the quality traits of watermelon. For instance, the total sugar content decreases when Lagenaria siceraria and Cucurbita maxima × C. moschata are used as rootstocks [37], while the acid concentration increases with grafting on Citrullus lanatus var. Citroides [38]. Beyond taste, the functional compounds in watermelon, such as β-carotene, citrulline, carotenoids, and ascorbic acid, are significantly enhanced when grafted onto Cucurbita maxima × C. moschata hybrid rootstocks compared to non-grafted watermelons [39]. These improvements can lead to enhanced nutritional value and health benefits for consumers.
Here, we investigated the compatibility between various rootstocks and watermelon by comparing survival rate and seedling index of grafting seedling, and utilized paraffin section to meticulously observe the anatomical structure of graft junction during grafting process. Further, we quantified the content of hormones and proteases activity within the grafted seedlings, measured the total sugar and titratable acid content in the grafted watermelons. Our findings not only reveal the underlying anatomical structures and physiological mechanisms of watermelon grafting but also offer practical insights for optimizing grafting practices.
2. Materials and Methods
2.1. Plant Materials
Our experiment was conducted at the Tropical Crops Resources Institute, Chinese Academy of Tropical Agricultural Sciences, located at 19°28'N, 109°29'E, and 105 meters above sea level. Watermelon ‘Youdu’ (Citrullus lanatus, Fuyou Seedling Co., Ltd., Haikou City, China) was selected as the scion, the interspecific pumpkin ‘Zaoshenxizhen’ and ‘Jinzuan’ (Cucurbita moschata, Hengrui Seed Co., Ltd., Qingzhou City, China), ‘Jinzhen-8’ (Citrullus lanatus, Kerun Agricultural Technology Co., Ltd., Tianjin City, China), ‘Jingxinzhen-2’, ‘Jingxinzhen-3’, ‘Jingxinzhen-9’ and ‘Heiniu’ (Citrullus lanatus, Jinyan Yinong Seed Sci-Tech Co., Ltd., Beijing City, China), and wild watermelon ‘Yongzhen’ (Citrullus lanatus, Jinyan Yinong Seed Sci-Tech Co., Ltd., Beijing City, China) were used as the rootstocks. The seeds for both scion and rootstock were sown into 50-cell plug trays containing commercial “Beilei” matrix (Zhenjiang Beilei Matrix Organic Fertilizer Co., Ltd., Zhenjiang City, China). The ‘amplexiform grafting’ method was employed for all watermelon seedlings, with self-graft or non-grafted watermelon seedlings serving as a control in our experiments. After grafting, the seedlings were transferred to a greenhouse with a temperature of 28 °C during the day and 18 °C at night, and a relative humidity (RH) maintained between 60% and 80%. After a two-week acclimation period, these plants were ready for subsequent experimental procedures.
2.2. Measurements of Plant Growth
We measured the stem thickness using a vernier caliper and the height with a ruler. These measurements were taken for 15 plants per group, and the process was repeated three times. Following the measurements, the grafted seedlings were subjected to a drying process to determine their dry weights (DW). Initially, they were placed in an air oven at 105 °C for 10 minutes, followed by a longer drying period at 70 °C for 72 hours. The survival rates and seedling index of the grafted seedlings were evaluated 10 days post-graft [40]. Survival rate (%) = (survival number / total number of grafted plants) × 100%.
Seedling index = (scion stem thickness / scion height) × plant dry weights (g).
2.3. Microscopic Observation of the Anatomical Structure
The samples were collected at 6, 8, and 10 days post-grafting(dpg) to observe of anatomy structure. At each time point, six grafted seedlings were selected. These samples were fixed with FAA fixative solution and processed into paraffin sections. The sections were carefully cut longitudinally along the vertical direction of the grafting surface, with each section measuring a precise thickness of 12 μm. The sections were subsequently stained with Safranin O-Fast Green, then photographed under Zeiss Axio Imager. A2 fluorescence microscope.
2.4. Quantification of ABA, IAA, ZR, GA3 and ETH
The hormones from the grafted samples were extracted according to the method described previously with a slight modification [41,42]. Briefly, 0.5g grafted samples was homogenized with 5ml 2-propanol/H2O/concentrated HCl (2:1:0.002, vol/vol/vol). After shaking at 4 ℃ for 30min, 5 ml of dichloromethane were added to the extracts, which were then transferred to a refrigerated microcentrifuge and centrifuged at 13,000g for 5 minutes at 4 °C. Following centrifugation, the lower phase was concentrated by using nitrogen flow. The dried samples were dissolved in 0.1 ml ethanol for hormone detection.. The hormones, including ABA, IAA, ZR, GA3 and ETH in the samples were quantified using an ELISA kit in accordance with the manufacturer’s instructions (Shanghai Fantai Biotechnology Co., Ltd., Shanghai city, China).
2.5. Assay of Enzyme Activity
Enzyme extraction from the grafting joints was performed with a slight modification of a previously described method [43]. The samples were ground with pestle in extraction buffer [50 mM Hepes-KOH (pH 7.5), 1 mM EDTA, 5 mM dithioerythritol (DTE), 10% (v/v) glycerol, 0.5 mM phenylmethylsulfonyl fluoride (PMSF), 2 mM benzamidine, 2 mM e-aminocaproic acid]. The homogenized samples were centrifuged at 4 ℃ for 15 min at 13,000 g, then the supernatants were collected for the detection of enzymatic activity. Protein concentration was measured accord to the method of Bradford and Williams[44]. The enzyme activity in the grafted samples and control were assayed by the ELISA kit in accordance with the manufacturer’s instructions (Shanghai Fantai Biotechnology Co., Ltd., Shanghai city, China).
2.6. Sugar Extract and Determination
The sugar of sample was extracted following a modified version of a previously described method [45]. Briefly, 2g samples were ground and homogenized with pestle in 20 mL of 80% ethanol. The homogenates were centrifuged at 5,000 × g for 15 min at 4 ℃, and the 20 μL supernatants were injected into an HPLC system(Agilent 1290 Infinity Series high-pressure liquid chromatography system, USA). The sugar, including fructose, glucose, sucrose, maltose contents were calculated by using standard curves. The total sugar contents was obtained by summing the concentrations of these individual sugars.
2.7. Organic acid Extract and Determination
0.5 g watermelon fruit were ground with pestle in 2.5 mL of ultrapure water. The homogenates were centrifuged at 5,000 × g for 15 min at 4 ℃, and the supernatants were injected into the HPLC system(Agilent 1290 Infinity Series high-pressure liquid chromatography system, USA) with a diode array detector (DAD). The concentrations of organic acids, such as oxalic acid, tartaric acid, malic acid, maleic acid, citric acid, succinic acid, were calculated using standard curves. Total organic acid concentrations were calculated by summing individual concentrations of the various acids detected.
2.8. Statistical Analysis
All experiments were randomly designed and repeated three times, each treatment contained at least 10 plants. Statistical data were analyzed based on one way analysis of variance (ANOVA) and Student’s t-test, Significant differences were indicated by p < 0.05, p < 0.01 and p < 0.001.
3. Results
3.1. Survival Rate and Seedling Index of Watermelon Grafted onto Various Rootstocks
The survival Rates of watermelon grafting onto various pumpkins and wild watermelon ranged from 58.56% to 90.145% (Figure 1C). Among them, ‘Heiniu’ exhibited the highest survival rate, while ‘Yongzhen’ showed the lowest, there was a significant difference between the seedlings grafted onto ‘Yongzhen’ rootstocks and self-rooted grafted seedlings. Additionally, we calculated the seedling index for different rootstocks and self-rooted grafted seedlings, which varied from 0.0494 to 0.135 (Figure 1D), The ‘Jin xin zhen-3’ rootstock had the highest seedling index, followed by ‘Heiniu’ with an index of 0.11, while ‘Yongzhen’ had the lowest. These results showed that the choice of rootstock significantly influences both the survival rate and seedling indexes (Figure 1C and D). By considering both survival rate and seedling index, we selected pumpkin variety ‘Heiniu’ as the optimal rootstock for subsequent research.
3.2. Optimal Seedling Ages for Watermelon Grafting onto ‘Heiniu’ Rootstock
The ‘Heiniu’ was used as rootstock, the survival rates of different seedling ages ‘Youdu’ scions grafting onto ‘Heiniu’ rootstocks of various seedling ages were 86.9%-97.69% (Table1, Figure 2C). The highest survival rate was observed for the 'Youdu' scion at the first true leaf stage grafted onto ‘Heiniu’ rootstock at the second true leaf stage (T5), while the lowest was for the ‘Youdu’ scion at the second true leaf stage grafted onto ‘Heiniu’ rootstock also at the second true leaf stage (T6). Compared to the control (CK), there were extremely significant statistical differences for both T5 and T6. Meanwhile, we recorded the seedling index for the various grafted seedlings, which ranged from 0.061 to 0.088 (Figure 2D). the highest seedling index was the ‘Youdu’ scion at the first true leaf grafting onto ‘Heiniu’ rootstock at the second true leaf (T5), and the lowest was the ‘Youdu’ scion at the first true leaf grafting onto ‘Heiniu’ rootstock at the cotyledon flattening. Based on the the analysis of both survival rates and the seedling indices, we chose the first true leaf and the second true leaf as the optimum seedling ages of scions and rootstocks, respectively.

Table 1.
The grafting combinations of scions and rootstocks in different seedling ages.
| Treatment. | Scion Seedling Age (You Du) | Rootstock seedling age (Hei Niu) | Grafting combination |
| T1 | Cotyledon flattening (Six Days) | Cotyledon flattening (Five Days) | Cotyledon flattening / Cotyledon flattening |
| T2 | The first true leaf (Eight days) | Cotyledon flattening (Five Days) | The first true leaf / Cotyledon flattening |
| T3 | Two true leaf (Ten days) | Cotyledon flattening (Five Days) | Two true leaf / Cotyledon flattening |
| T4 | Cotyledon flattening (Six Days) | Two true leaf (Nine Days) | Cotyledon flattening / Two true leaf |
| T5 | The first true leaf (Eight days) | Two true leaf (Nine Days) | The first true leaf / Two true leaf |
| T6 | Two true leaf (Ten days) | Two true leaf (Nine Days) | Two true leaf / Two true leaf |
| CK | Cotyledon flattening (You Du) | Two true leaf (You Du) | Cotyledon flattening / Two true leaf |
3.3. The Anatomical Structures of Watermelon Graft Union
Extensively previous research has utilized microscopy to examine the anatomical structures of grafted plants [34,46]. Thus, to observe the structural changes at graft junctions union, we generated paraffin sections and employed microscopy to scrutinize the anatomical changes at the graft junctions across various stages. In the control, as the graft junctions developed, parenchyma cells within injured tissues, positioned at different sites along both sides of the necrotic layers, initiated a differentiation process. The mechanically damaged cells were progressively resorbed, leading to the gradual dissolution of the necrotic layers and the concurrent formation of substantial callus tissue. By 10 days post-grafting, the necrotic layers were nearly completely resolved, resulting in a tight apposition of cells from both the rootstock and scion (Fig 3, A-C). Agreement with the controls, the Yd/Hn grafts retained necrotic layers at 6 and 8 days post-grafting. Subsequently, these layers were eliminated, and an intimate contact between the scion and rootstock cells was established (Figure 3, D-F). These results might implicate that by 10 days after grafting, a vascular bridge between the rootstock and scion had been established, and the wound at the graft junction had predominantly healed and consolidated.
3.4. Measurement of Hormone Contents for Watermelon Graft Union During the Grafting Process
Plant hormones play an pivotal role for the grafting process [29,31]. To elucidate the hormonal dynamics during the watermelon grafting process, we quantified hormone levels at the graft junction post-grafting. As shown in Figure 4A, the ABA content in watermelon seedlings initially surged and then declined post-grafting, whereas in grafted seedlings, it exhibited an initial decrease followed by an increase, with a significant even extremely significant difference observed between the two groups (**p < 0.01 and ***p < 0.001).
The IAA content exhibited a decline from 3119.82 μg/g to 1914.96 μg/g (Figure 4B), exhibited a decline from 3119.82 μg/g to 1914.96 μg/g post-grafting. In contrast, grafted seedlings showed an increase from 1813.32 μg/g to 3003.94 μg/g, followed by a decrease to 1604.84 μg/g, with significant difference even extremely significant difference noted between watermelon seedlings and grafted seedlings.
ZR levels showed extremely significant difference between watermelon seedlings and grafted seedlings at 6 and 10 days post-grafting (Figure 4C). In watermelon seedlings, ZR content decreased to 38.84 ng/g from 6 to 8 days and then increased to 75.87 ng/g, while in grafted seedlings, it decreased to 43.96 ng/g and then increased to 60.75 ng/g. Notably, no significant difference was observed between watermelon seedlings and grafted seedlings at 8 days post-grafting.
The GA3 content significantly decreased in both watermelon seedlings and grafted seedlings from 6 to 8 days post-grafting (Figure 4D), then elevated at 10 days. The highest GA3 content were 0.405 ng/g in watermelon seedlings and 0.423 ng/g in grafted seedlings at 6 and 10 days, respectively, while the lowest were 0.24 ng/g in watermelon seedlings and 0.215 ng/g in grafted seedlings at 8 days, respectively, which displayed extremely significant difference between watermelon seedlings and grafted seedlings at 6 and 10 days post-grafting.
ETH levels were significantly higher in grafted seedlings compared to watermelon seedlings at 6 and 8 days post-grafting. ETH content decreased from 11.49 pg/g to 8.63 pg/g in grafted seedlings from 6 to 10 days, while it increased from 9.09 pg/g to 9.87 pg/g in watermelon seedlings. These findings suggested that hormones, particularly ABA, IAA, ZR, GA3, and ETH, play a crucial role in the grafting process of watermelon, with distinct mechanisms of action observed between watermelon seedlings and grafted seedlings.
3.5. Measurement of Enzyme Activities of Watermelon Graft Union During the Grafting Process
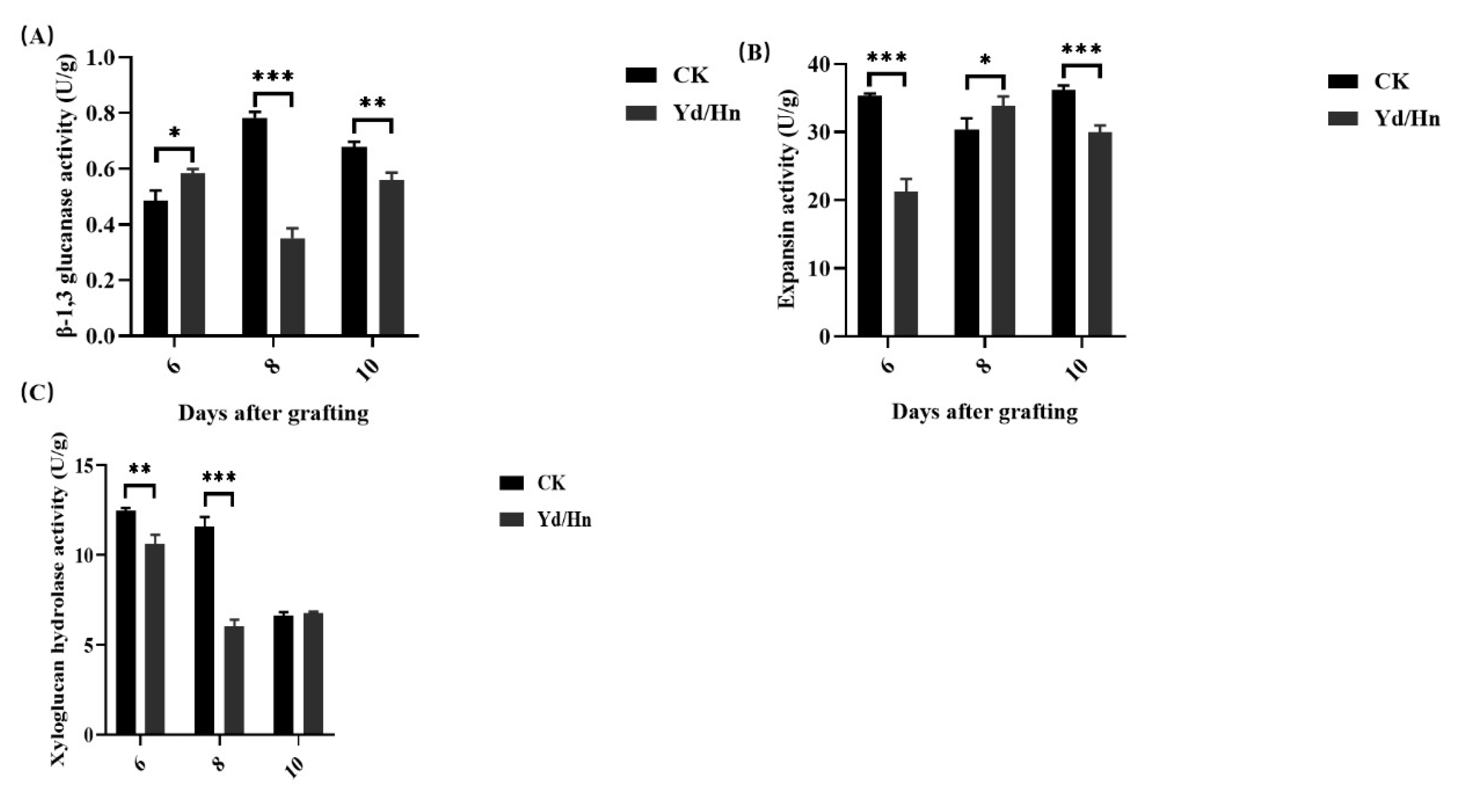
A close relation between glucanase, expansin and grafting has been verified in model plants such as Arabidopsis and rice [34,47]. Nevertheless, the role of these enzymes, including xyloglucan hydrolase (XTH), in the grafting process of watermelon remains elusive. To investigate their potential roles, we quantified the activities of these enzymes at various time points following watermelon grafting. The β-1,3 glucanase activity in watermelon seedlings and grafted seedlings exhibited statistically difference, significant difference and extremely significant difference at 6, 8 and 10 days post-grafting, respectively (Figure 5A). In watermelon seedlings, β-1,3 glucanase activity increased from 0.487 U/g at 6 days to 0.783 U/g at 8 days, then decreased to 0.677 U/g at 10 days. In contrast, in grafted seedlings, the activity started at 0.583 U/g, decreased to 0.35 U/g, and subsequently increased to 0.56 U/g.
Expansin activity in watermelon seedlings decreased from 35.333 U/g to 30.427 U/g and then rose to 30.197 U/g from 6 to 10 days post-grafting (Figure 5B). Compared to the watermelon seedlings, Grafted seedlings exhibited an increase in expansin activity from 21.3 U/g to 33.887 U/g from 6 to 8 days, followed by a decrease to 30.057 U/g at 10 days post-grafting. Statistically difference, significant difference and extremely significant difference were observed between watermelon seedlings and grafted seedlings at 6, 8, and 10 days, respectively.
Watermelon seedlings had significantly higher xyloglucan hydrolase (XTH) activity compared to grafted seedlings (Figure 5C). At 6 days, XTH activities were 12.487 U/g in watermelon seedlings and 10.647 U/g in grafted seedlings. At 8 days, XTH activities were 11.6 U/g and 6.03 U/g, respectively. Notably, at 10 days, there was no significant difference in XTH activities between watermelon seedlings (6.623 U/g) and grafted seedlings (6.75 U/g). These results indicated that the variation trends of these three enzymes levels were inversely related between watermelon seedlings and grafted seedlings during the grafting process, implying grafting may alter these protease synthesis.
3.6. Total Soluble Sugar and Titratable Acid Content in the Grafting Junction Among Watermelon Seedlings and Grafted Seedlings
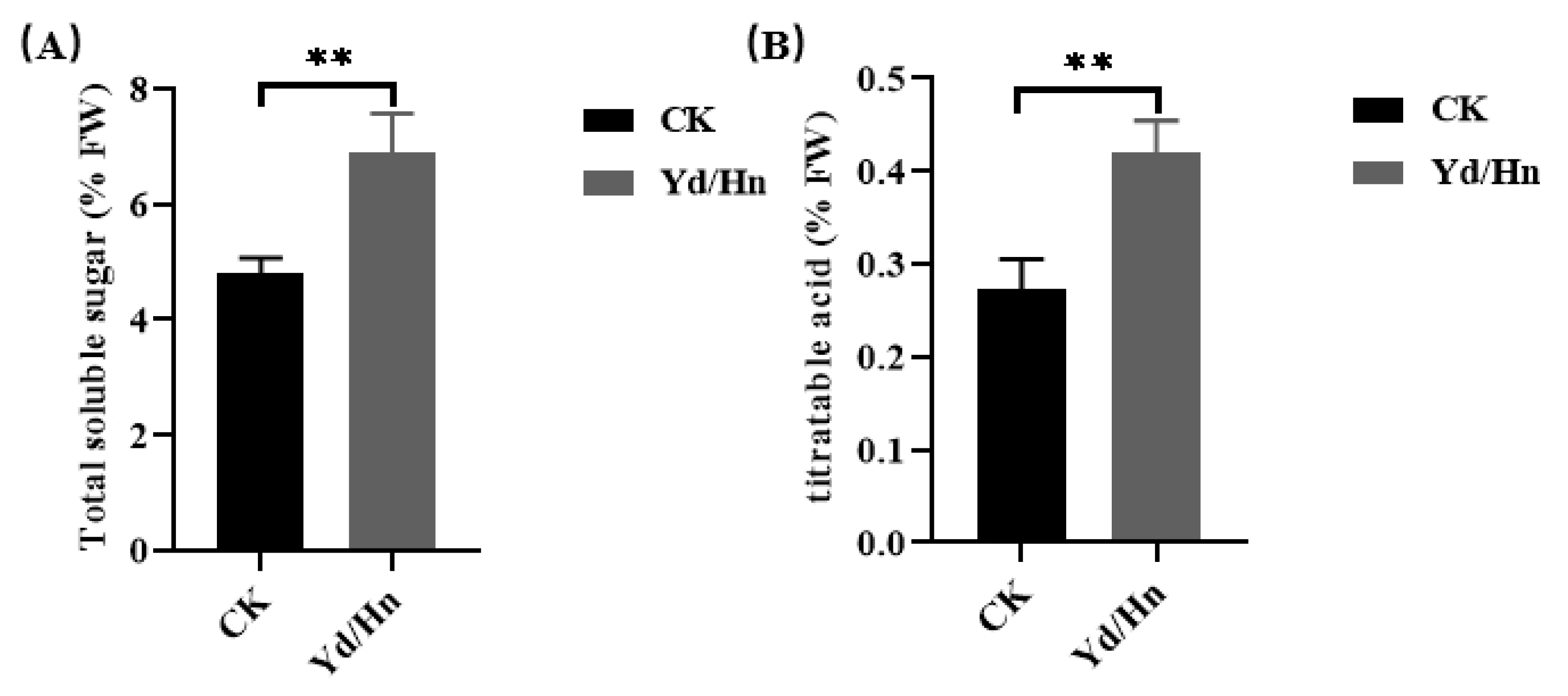
Sugar and acid content are crucial quality characteristics of watermelon, and different rootstock varieties exert varying influences on these components following grafting [39,48]. In our study, we observed a higher total soluble sugar content in grafted seedlings than in watermelon seedlings, the total soluble sugar content were 4.813% and 6.903% in watermelon seedlings and grafted seedlings, respectively. Consistent with the trend observed for total soluble sugar, grafted seedlings exhibited a greater titratable acid content than non-grafted seedlings, with levels at 0.27% and 0.42%, respectively. Consequently, the use of ‘Heiniu’ as a rootstock can enhance the total soluble sugar and titratable acid content in watermelons through grafting, thereby potentially improving their overall quality.
Figure 6.
Total soluble sugar and titratable acid content of watermelon seedlings and grafted seedlings. (A) The total soluble sugar content, (B) the titratable acid content. *, ** and *** indicate significant differences at p < 0.05, 0.01 and 0.001 levels, respectively.
Figure 6.
Total soluble sugar and titratable acid content of watermelon seedlings and grafted seedlings. (A) The total soluble sugar content, (B) the titratable acid content. *, ** and *** indicate significant differences at p < 0.05, 0.01 and 0.001 levels, respectively.
4. Discussion
Grafting is a horticultural technique that has been proven to significantly enhance the yield, quality, and resistance to biotic and abiotic stress of many crops, so it is widely employed in the cultivation of a variety of crops [49,50]. In particular, grafting watermelon onto various pumpkin or wild watermelon rootstocks has been widely utilized in the major watermelon-producing regions around the world. Wild watermelon [citrullus colocynthis (L.) Schrad, ‘Esfahan’] and a commercial interspecific cucurbita hybrid [Cucurbita maxima Duch. × Cucurbita moschata Duch., ‘Shintoza’] serve as rootrocks to inherently boost the resistance in a given setting of drought stress, although this comes at the cost of reduced growth and biomass compared to ungrafted plants [51]. Grafting watermelon cultivar ‘Zaojia 8424’ onto bottle gourd ‘Jingxinzhen 1’ [Lagenaria siceraria] and pumpkin ‘Qingyanzhen 1’ [Cucurbita maxima × C. moschata] remarkablely elevate the fruit flesh, rind firmness, and rind thickness when compared to the non-grafted plants [52]. However, the compatibility between different rootstocks and scions remains unclear. In order to investigate the compatibility, we conducted a survey to assess the survival rates and seedling indices of various rootstocks. Among them, the highest survival rate was the ‘Heiniu’, and the highest seedling index was the ‘Jing Xinzhen-3’. By taking into account both survival rates and seedling indices, we selected ‘Heiniu’ as the optimal rootstock.
The seedling age is an vital factor influencing grafting success, as the compatibility between scion and rootstock varies with their ages, ultimately impacting the survival rate of grafted seedlings. Greenwood et al. demonstrated that the rootstock age significantly impacts vegetative growth, foliar morphology, and reproductive development in grafted red spruce (Picea rubens Sarg.) [53]. Nguyen et al. also revealed that rootstock age has a notable effect on the percentage of graft success and the length of scion sprout[54]. In agreement with the previous studies, our results uncovered that the survival rates and seedling indices arrive at the maximum value when the scion and rootstock age are eight and nine days, respectively. These results suggest that both scion and rootstock age play pivotal roles in the success of grafting. Understanding these age-related factors can help optimize grafting techniques and improve overall grafting outcomes in horticultural practices.
During grafting process, the formation of a necrotic layer at the graft union is a key developmental event, and it is a dynamically changing structure [55]. In the initial stages of grafting, the necrotic layer serves as a barrier, isolating the graft union from the external environment. This isolation is beneficial for wound healing and provides a defense against pathogen invasion [56]. In addition, the differentiation of callus tissue is a crucial step in the development of graft union. This tissue not only connects the scion and rootstock but also absorbs and metabolizes the substances from the necrotic layer, this process allows the cells of the rootstock and scion to re-establish new material connections. At the same time, the callus tissue also provides the potentiality to differentiate into vascular bundles which are essential for connecting the scion and rootstock [57]. In line with these study, our results showed that the necrotic layer recedes as the graft union matures, and necrotic layer essentially disappeared at ten days after grafting, enabling the cells of the rootstock and scion to come into close contact with each other. Therefore, we can consider ten days as a time marker for successful grafting, signifying the point at which the graft union is well-established and the scion and rootstock have fused successfully.
Phytohormones involve every aspect of plant development and resistance to biotic and abiotic stresses. The success of grafting hinges on the regeneration of vascular tissue and the adhesion between the rootstock and scion. Phytohormones are pivotal at various stages of the plant grafting process. Exogenous phytohormones, especially auxin, stimulate differentiation of phloem and xylem at different concentrations [57,58]. The application of 6-benzylaminopurine (6-PA) to apple rootstocks has been shown to restore the growth of secondary shoots, whereas the use of GA4+7 has been found to reduce the percentage of both primary and secondary shoots [59]. Both cytokinin (Kinetin) and indole-3-butryic acid (IBA), when dripped onto the cut surfaces of both rootstock and scion, have been observed to accelerate the rapid proliferation of callus tissue [60]. By monitoring the fluctuations of various hormones during the grafting process, including ABA, IAA, ZR, GA3, and ETH, we discovered that hormone levels vary at different post-grafting intervals. This indicates that hormones are likely to play a significant role in the grafting process of watermelons. Moreover, the contrasting trends in hormone changes between grafted seedlings and the watermelon seedling imply that grafting can influence hormone synthesis.
Grafting in plants is intricately linked to a myriad of physiological and biochemical processes, including injury responses, callus formation, and hormone signal transduction [61]. A multitude of proteins are involved in these processes, encompassing plant tissue adhesion, amino acid metabolism, stress response, DNA damage repair, and plant-microbe interactions [62,63]. Enzymes such as β-1,4-glucanase, β-1,3-glucanase, xyloglucan hydrolase, and expansin, which act as proteases for cell wall modification, are crucial for successful grafting in the majority of crops. The healing process in the TRV-CmGH9B3 melon, which is a crucial gene of β-1,4-glucanase, is slower compared to the wild type [64]. Transcriptome analysis has revealed that the expression of four genes associated with cell wall modification is upregulated following the grafting of tobacco onto Arabidopsis thaliana [34]. In agreement with these reports, our data shows that the activity levels of β-1,3-glucanase, XTH and expansin alter at different days after grafting, with significant differences in enzyme activity observed between grafted seedlings and watermelon seedlings. Based on these observations, we conclude that these three proteases play a critical role in the graft healing process of watermelon.
Watermelon quality encompasses a wide range of characteristics, including physical properties, soluble solid content, titratable acid content, and flavor. Grafting is a horticultural technique that not only enhances resistance to biotic and abiotic stresses but also has the potential to improve the quality of watermelons. In addition to a variety of dissolved proteins, minerals, pigments, and vitamins, sugar and acid are the main components of total soluble solids (TSSs). Most of studies find that the TSSs can be significantly affected by grafting, the TSSs increase when watermelon cvs. Crimson Sweet and Sugar Baby are grafted onto C. maxima × C. moschata hybrid rootstocks [65,66]. However, a few studies manifest that grafting have a negative impact on TSSs, there is a decrease observed post-grafting compared to ungrafted plants [67,68]. In line with these reports, our results revealed that the sugar and titratable acid content of grafted watermelon fruits are significantly higher than those of non-grafted watermelons, indicating that utilizing the pumpkin 'Hei Niu' as rootstock can effectively enhance the quality of watermelon.
5. Conclutions
In our study, we conducted a comprehensive comparison of the survival rate and seedling index for pumpkin and wild watermelon rootstocks following grafting. Through this analysis, we identified ‘Heiniu’ pumpkin as the superior rootstock. Additionally, we determined that the optimal seedling ages for the scion and rootstock are at the stage of the first true leaf and the second true leaf, respectively. Using paraffin sections and microscopic observation, we observed that the necrotic layer between the rootstock and scion had disappeared after 10 days post-grafting, with a significant amount of callus tissue formation. This finding indicates that by this time point, the graft union is well-established, which is crucial for the success of the graft. Throughout the grafting process, we monitored the levels of various hormones, including ABA, IAA, ZR, GA3 and ETH. We noted variations in hormone content. Consistent with hormones content, cell wall modification enzymes, such as β-1,3-glucanase, xyloglucan hydrolase, and expansin, were fluctuant at different days post-grafting. These hormones and proteinase are essential for the successful grafting of watermelon. Furthermore, we found that the quality of watermelon, particularly the sugar and titratable acid content, was significantly improved by using the pumpkin variety ‘Heiniu’ as a rootstock.
Author Contributions
H.H. and G.D. conducted the watermelon grafting experiment, calculated the survival rate and seedling index, determined hormone levels, and drafted the manuscript. X.W. and R.W. were responsible for measuring enzyme activity and sugar content. Y.C., Y.Z. and X.D. conceived the study, participated in its design, and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the Major science and technology projects of Hainan Province (no. ZDKJ2021005), Industrial Technology System of Winter Vegetables of Hainan Province (no. HNARS-05-G07).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schmidt-Jeffris, R. A., Coffey, J. L., Miller, G., & Farfan, M. A. (2021). Residual Activity of Acaricides for Controlling Spider Mites in Watermelon and Their Impacts on Resident Predatory Mites. Journal of Economic Entomology, 114(2), 818-827.
- Gimode, W., Bao, K., Fei, Z., & McGregor, C. (2021). QTL associated with gummy stem blight resistance in watermelon. Theoretical and Applied Genetics, 134(2), 573-584.
- Gusmini, G., Rivera-Burgos, L. A., & Wehner, T. C. (2017). Inheritance of resistance to gummy stem blight in watermelon. HortScience, 52(11), 1477-1482.
- Bantis, F., & Koukounaras, A. (2023). Ascophyllum nodosum and Silicon-Based Biostimulants Differentially Affect the Physiology and Growth of Watermelon Transplants under Abiotic Stress Factors: The Case of Salinity. Plants, 12(3), 433.
- Ye, L., Zhao, X., Bao, E. C., Cao, K., & Zou, Z. R. (2019). Effects of Arbuscular Mycorrhizal Fungi on Watermelon Growth, Elemental Uptake, Antioxidant, and Photosystem II Activities and Stress-Response Gene Expressions Under Salinity-Alkalinity Stresses. Frontiers in Plant Science, 10, 863.
- Nabwire, S., Wakholi, C., Faqeerzada, M. A., Arief, M. A. A., Kim, M. S., Baek, I., & Cho, B. K. (2022). Estimation of Cold Stress, Plant Age, and Number of Leaves in Watermelon Plants Using Image Analysis. Frontiers in Plant Science, 13, 847225.
- Ding, C. Q., Chen, C. T., Su, N., Lyu, W. H., Yang, J. H., Hu, Z. Y., & Zhang, M. F. (2021). Identification and characterization of a natural SNP variant in ALTERNATIVE OXIDASE gene associated with cold stress tolerance in watermelon. Plant Science, 304, 110735.
- Hamurcu, M., Khan, M. K., Pandey, A., Ozdemir, C., Avsaroglu, Z. Z., Elbasan, F., Omay, A. H., & Gezgin, S. (2020). Nitric oxide regulates watermelon (Citrullus lanatus) responses to drought stress. 3 Biotech, 10(11), 494.
- Li, H., Mo, Y. L., Cui, Q., Yang, X. Z., Guo, Y. L., Wei, C. H., Yang, J. Q., Zhang, Y., Ma, J. Q., & Zhang, X. (2019). Transcriptomic and physiological analyses reveal drought adaptation strategies in drought-tolerant and -susceptible watermelon genotypes. Plant Science, 278, 32-43.
- He, N., Umer, M. J., Yuan, P. L., Wang, W. W., Zhu, H. J., Zhao, S. J., Lu, X. Q., Xing, Y., Gong, C. S., Liu, W. G., & Sun, X. W. (2022). Expression dynamics of metabolites in diploid and triploid watermelon in response to flooding. PeerJ, 13814.
- He, N., Umer, M. J., Yuan, P., Wang, W. W., Zhu, H. J., Lu, X. Q., Xing, Y., Gong, C. S., Batool, R., Sun, X. W., & Liu, W. G. (2023). Physiological, biochemical, and metabolic changes in diploid and triploid watermelon leaves during flooding. Frontiers in Plant Science, 14, 1108795.
- Yetisir, H., & Sari, N. (2003). Effect of different rootstocks on plant growth, yield, and quality of watermelon. Australian Journal of Experimental Agriculture, 43, 1269-1274.
- King, S. R., Davis, A. R., Liu, W. G., & Levi, A. (2008). Grafting for disease resistance. HortScience, 43, 1673-1676.
- Kousik, C. S., Mandal, M., & Hassell, R. (2018). Powdery mildew resistant rootstocks that impart tolerance to grafted susceptible watermelon scion seedlings. Plant Disease, 102(7), 1290-1298.
- Mahmud, I., Kousik, C., Hassell, R., Chowdhury, K., & Boroujerdi, A. F. (2015). NMR Spectroscopy Identifies Metabolites Translocated from Powdery Mildew Resistant Rootstocks to Susceptible Watermelon Scions. Journal of Agricultural and Food Chemistry, 63(36), 8083-8091.
- Huitrón-Ramírez, M. V., Huitrón-Ramírez, M. G., & Camacho-Ferre, F. (2009). Influence of grafted watermelon plant density on yield and quality in soil infested with Melon Necrotic Spot Virus. HortScience, 44(7), 1838-1841.
- Kousik, C. S., Donahoo, R. S., & Hassell, R. (2012). Resistance in watermelon rootstocks to crown rot caused by Phytophthora capsici. Crop Protection, 39, 18-25.
- Li, H., Guo, Y. L., Lan, Z. X., Xu, K., Chang, J. J., Ahammed, G. J., Ma, J. X., Wei, C. H., & Zhang, X. (2021). Methyl jasmonate mediates melatonin-induced cold tolerance of grafted watermelon plants. Horticulture Research, 8, 57.
- Guo, Y. L., Yan, J. Y., Su, Z. Z., Chang, J. J., Yang, J. Q., Wei, C. H., Zhang, Y., Ma, J. X., Zhang, X., & Li, H. (2021). Abscisic Acid Mediates Grafting-Induced Cold Tolerance of Watermelon via Interaction With Melatonin and Methyl Jasmonate. Frontiers in Plant Science, 12, 785317.
- Shin, Y. K., Bhandari, S. R., & Lee, J. G. (2021). Monitoring of salinity, temperature, and drought stress in grafted watermelon seedlings using chlorophyll fluorescence. Frontiers in Plant Science, 12, 786309.
- Bikdeloo, M., Colla, G., Rouphael, Y., Hassandokht, M. R., Soltani, F., Soltani, F., Salehi, R., Kumar, P., & Cardarelli, M. (2021). Morphological and Physio-Biochemical Responses of Watermelon Grafted onto Rootstocks of Wild Watermelon [Citrullus colocynthis (L.) Schrad] and Commercial Interspecific Cucurbita Hybrid to Drought Stress. Horticulturae, 7, 359.
- Yang, Y. J., Yu, L., Wang, L. P., & Guo, S. R. (2015). Bottle gourd rootstock-grafting promotes photosynthesis by regulating the stomata and non-stomata performances in leaves of watermelon seedlings under NaCl stress. Journal of Plant Physiology, 186-187, 50-58.
- Yang, Y. J., Lu, X. M., Yan, B., Li, B., Sun, J., Guo, S. R., & Tezuka, T. (2013). Bottle gourd rootstock-grafting affects nitrogen metabolism in NaCl-stressed watermelon leaves and enhances short-term salt tolerance. Journal of Plant Physiology, 170, 653-661.
- Bidabadi, S. S., Abolghasemi, R., & Zheng, S. J. (2018). Grafting of watermelon (Citrullus lanatus cv. Mahbubi) onto different squash rootstocks as a means to minimize cadmium toxicity. International Journal of Phytoremediation, 20, 730-738.
- Nawaz, M. A., Chen, C., Shireen, F., Zheng, Z. H., Jiao, Y. Y., Sohail, H., Sohail, M., Imtiaz, M., Ali, M. A., Huang, Y., & Bie, Z. L. (2018). Improving vanadium stress tolerance of watermelon by grafting onto bottle gourd and pumpkin rootstock. Plant Growth Regulation, 85, 41-56.
- Yang, Y. J., Wang, L. P., Tian, J., Li, J., Sun, J., He, L. Z., Guo, S. R., & Tezuka, T. (2012). Proteomic study participating the enhancement of growth and salt tolerance of bottle gourd rootstock-grafted watermelon seedlings. Plant Physiology and Biochemistry, 58, 54-65.
- Fallik, E., Alkalai-Tuvia, S., Chalupowicz, D., Popovsky-Sarid, S., & Zaaroor-Presman, M. (2019). Relationships between rootstock-scion combinations and growing regions on watermelon fruit quality. Agronomy, 9, 536.
- Melnyk, C. W. (2017). Plant grafting: Insights into tissue regeneration. Regeneration, 4, 3-14.
- Peña-Cortés, H., Fisahn, J., & Willmitzer, L. (1995). Signals involved in wound-induced proteinase inhibitor II gene expression in tomato and potato plants. Proceedings of the National Academy of Sciences, 92, 4106-4113.
- Oh, S., Park, S., & Han, K.-H. (2003). Transcriptional regulation of secondary growth in Arabidopsis thaliana. Journal of Experimental Botany, 54, 2709-2722.
- Melnyk, C. W., Schuster, C., Leyser, O., & Meyerowitz, E. M. (2015). A developmental framework for graft formation and vascular reconnection in Arabidopsis thaliana. Current Biology, 25(11), 1306-1318.
- Parkinson, M., & Yeoman, M. M. (1982). Graft formation in cultured, explanted internodes. New Phytologist, 91(3), 711-719.
- Matsuoka, K., Sugawara, E., Aoki, R., Takuma, K., Terao-Morita, M., Satoh, S., & Asahina, M. (2016). Differential cellular control by cotyledon-derived phytohormones involved in graft reunion of Arabidopsis hypocotyls. Plant and Cell Physiology, 57(10), 2620-2631.
- Notaguchi, M., Kurotani, K. I., Sato, Y., Tabata, R., Kawakatsu, Y., Okayasu, K., Sawai, Y., Okada, R., Asahina, M., Ichihashi, Y., Shirasu, K., Suzuki, T., Niwa, M., & Higashiyama, T. (2020). Cell-cell adhesion in plant grafting is facilitated by β-1,4-glucanases. Science, 369(6501), 698-702.
- Soteriou, G. A., Kyriacou, M. C., Siomos, A. S., & Gerasopoulos, D. (2014). Evolution of watermelon fruit physicochemical and phytochemical composition during ripening as affected by grafting. Food Chemistry, 165, 282-289.
- Huitrón, M. V., Díaz, M., Diánez, F., & Camacho, F. (2007). The effect of various rootstocks on triploid watermelon yield and quality. Journal of Food, Agriculture & Environment, 5(2), 344-348.
- Fredes, A., Roselló, S., Beltrán, J., Cebolla-Cornejo, J., Pérez-de-Castro, A., Gisbert, C., & Picó, M. B. (2017). Fruit quality assessment of watermelons grafted onto citron melon rootstock. Journal of the Science of Food and Agriculture, 97(9), 1646-1655.
- Çandır, E., Yetişir, H., Karaca, F., & Üstün, D. (2013). Phytochemical characteristics of grafted watermelon on different bottle gourds (Lagenaria siceraria) collected from the Mediterranean region of Turkey. Turkish Journal of Agriculture and Forestry, 37(3), 443-456.
- Fallik, E., & Ziv, C. (2020). How rootstock/scion combinations affect watermelon fruit quality after harvest? Journal of the Science of Food and Agriculture, 100(7), 3275-3282.
- Liu, C. J., Lin, W. G., Feng, C. R., Wu, X. S., Fu, X. H., Xiong, M., Bie, Z. L., & Huang, Y. (2021). A new grafting method for watermelon to inhibit rootstock regrowth and enhance scion growth. Agriculture, 11(5), 812.
- Yang, J., Zhang, J., Wang, Z., Zhu, Q., & Wang, W. (2001). Hormonal changes in the grains of rice subjected to water stress during grain filling. Plant Physiology, 127(1), 315-323.
- Pan, X. Q., Welti, R., & Wang, X. M. (2010). Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography-mass spectrometry. Nature Protocols, 5(6), 986-992.
- Ritte, G., Steup, M., Kossmann, J., & Lloyd, J. R. (2003). Determination of the starch-phosphorylating enzyme activity in plant extracts. Planta, 216(3), 798-801.
- Bradford, M. M., & Williams, W. L. (1976). New, rapid, sensitive method for protein determination. Federation Proceedings, 35(3), 274.
- Lee, E. J., Yoo, K. S., Jifon, J., & Patil, B. S. (2009). Characterization of short-day onion cultivars of three pungency levels with flavor precursors, free amino acid, sulfur, and sugar contents. Journal of Food Science, 74(4), C475-C480.
- Devi, P., DeVetter, L., Kraft, M., Shrestha, S., & Miles, C. (2022). Micrographic View of Graft Union Formation Between Watermelon Scion and Squash Rootstock. Frontiers in Plant Science, 13, 878289.
- Reeves, G., Tripathi, A., Singh, P., Jones, M. R. W., Nanda, A. K., Musseau, C., Craze, M., Bowden, S., Walker, J. F., Bentley, A. R., Melnyk, C. W., & Hibberd, J. M. (2022). Monocotyledonous plants graft at the embryonic root-shoot interface. Nature, 602, 280-286.
- López-Galarza, S., Batista, A. S., Pérez, D. M., Miquel, A., Baixauli, C., Pascual, B., Maroto, J. V., & Guardiola, J. L. (2004). Effects of grafting and cytokinin-induced fruit setting on colour and sugar-content traits in glasshouse-grown triploid watermelon. Journal of Horticultural Science & Biotechnology, 79(6), 971-976.
- Khah, M. K. (2011). Effect of grafting on growth, performance, and yield of aubergine (Solanum melongena L.) in greenhouse and open-field. International Journal of Plant Production, 5(3), 359-366.
- Rouphael, Y., Cardarelli, M., & Colla, G. (2008). Yield, mineral composition, water relations, and water use efficiency of grafted miniwatermelon plants under deficit irrigation. HortScience, 43(4), 730-736.
- Bikdeloo, M., Colla, G., Rouphael, Y., Hassandokht, M. R., Soltani, F., Salehi, R., Kumar, P., & Cardarelli, M. (2021). Morphological and Physio-Biochemical Responses of Watermelon Grafted onto Rootstocks of Wild Watermelon [Citrullus colocynthis (L.) Schrad] and Commercial Interspecific Cucurbita Hybrid to Drought Stress. Horticulturae, 7(3), 359.
- Huang, Y., Zhao, L. Q., Kong, Q. S., Cheng, F., Niu, M. L., Xie, J. J., Muhammad, A. N., & Bie, Z. L. (2016). Comprehensive Mineral Nutrition Analysis of Watermelon Grafted onto Two Different Rootstocks. Horticultural Plant Journal, 2(1), 105-113.
- Greenwood, M. S., Day, M. E., & Schatz, J. (2010). Separating the effects of tree size and meristem maturation on shoot development of grafted scions of red spruce (Picea rubens Sarg.). Tree Physiology, 30(4), 459-468.
- Nguyen, V. H., & Yen, C. R. (2018). Rootstock age and grafting season affect graft success and plant growth of papaya (Carica papaya L.) in greenhouse. Chilean Journal of Agricultural Research, 78(1), 59-67.
- Yeoman, M. M., Kilpatrick, D. C., Miedzybrodzka, M. B., & Gould, A. R. (1978). Cellular interaction during graft formation in plants, a recognition phenomenon? Symposium of the Society for Experimental Biology, 32, 139-160.
- Ermel, F. F., Poëssel, J. L., Faurobert, M., & Catesson, A. M. (1997). Early scion/stock junction in compatible and incompatible pear/pear and pear/quince grafts: A histo-cytological study. Annals of Botany, 79(6), 505-515.
- Yang, S. J. (1987). Observation on histological and cellular events of inter-specific grafting (Impatiens walleriana/Impatiens olivieri). Acta Agriculturae Universitatis Pekinensis, 13(2), 359-366.
- Aloni, R. (1987). Differentiation of vascular tissues. Annual Review of Plant Physiology, 38, 179-204.
- Aloni, R. (2001). Foliar and Axial Aspects of Vascular Differentiation: Hypotheses and Evidence. Journal of Plant Growth Regulation, 20(1), 22-34.
- Hooijdonk, V., Woolley, D. J., Warrington, I. J., & Tustin, D. S. (2010). Initial alteration of scion architecture by dwarfing apple rootstocks may involve shoot-root-shoot signalling by auxin, gibberellin, and cytokinin. Journal of Horticultural Science & Biotechnology, 85(1), 59-65.
- Köse, C., & Güleryüz, M. (2006). Effects of auxins and cytokinins on graft union of grapevine (Vitis vinifera L.) New Zealand. Journal of Crop and Horticultural Science, 34(2), 145-150.
- Cookson, S. J., Clemente Moreno, M. J., Hevin, C., Nyamba Mendome, L. Z., Delrot, S., Trossat-Magnin, C., & Ollat, N. (2013). Graft union formation in grapevine induces transcriptional changes related to cell wall modification, wounding, hormone signalling, and secondary metabolism. Journal of Experimental Botany, 64(7), 2997-3008.
- Yang, Y. J., Wang, L. P., Tian, J., Li, J., Sun, J., He, L. Z., Guo, S. R., & Tezuka, T. (2012). Proteomic study participating in the enhancement of growth and salt tolerance of bottle gourd rootstock-grafted watermelon seedlings. Plant Physiology and Biochemistry, 58, 54-65.
- Song, Y., Ling, N., Ma, J. H., Wang, J. C., Zhu, C., Raza, W., Shen, Y. F., Huang, Q. W., & Shen, Q. R. (2016). Grafting resulted in a distinct proteomic profile of watermelon root exudates relative to the un-grafted watermelon and the rootstock plant. Journal of Plant Growth Regulation, 35(4), 778-791.
- Zhu, Y. L., Hu, S. W., Min, J. H., Zhao, Y. T., Yu, H. Q., Irfan, M., & Xu, C.
- (2024). Transcriptomic analysis provides insight into the function of CmGH9B3, a key gene of β-1,4-glucanase, during the graft union healing of oriental melon scion grafted onto squash rootstock. Biotechnology Journal, 19(4), e2400006.
- Petropoulos, S. A., Khah, E. M., & Passam, H. C. (2012). Evaluation of rootstocks for watermelon grafting with reference to plant development, yield, and fruit quality. International Journal of Plant Production, 6(3), 481-492.
- Turhan, A., Ozman, N., Kuscu, H., Serbeci, M. S., & Seniz, V. (2012). Influence of rootstocks on yield and fruit characteristics and quality of watermelon. Horticultural Environment and Biotechnology, 53(3), 336-341.
- Davis, A. R., & Perkins-Veazie, P. (2005). Rootstock effects on plant vigor and watermelon fruit quality. Cucurbit Genetics Cooperative Report, 28-29, 39-42.
Figure 1.
Survival rates and seedling index of watermelon grafted onto different rootstocks (n=100). (A) The grafted watermelon seedlings by using the ‘amplexiform grafting’ method, (B) watermelon growth pictures at two weeks after grafting, (C) wurvival rates and (D) seedling index of grafted seedlings. *, ** and *** indicate significant differences at p< 0.05, 0.01 and 0.001 levels, respectively.
Figure 1.
Survival rates and seedling index of watermelon grafted onto different rootstocks (n=100). (A) The grafted watermelon seedlings by using the ‘amplexiform grafting’ method, (B) watermelon growth pictures at two weeks after grafting, (C) wurvival rates and (D) seedling index of grafted seedlings. *, ** and *** indicate significant differences at p< 0.05, 0.01 and 0.001 levels, respectively.
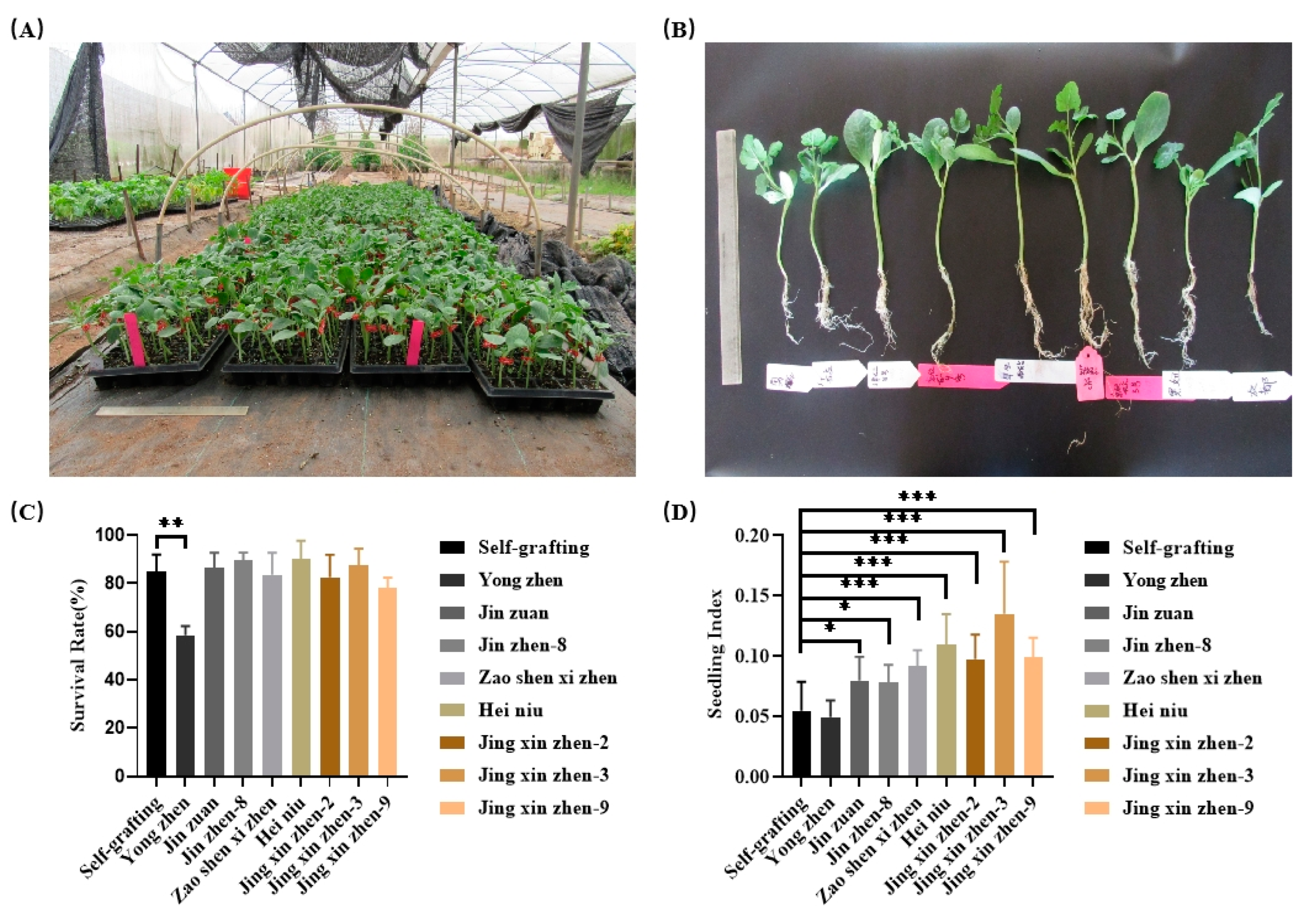
Figure 2.
Survival rates and seedling index of different seedling age scions grafted onto various seedling age rootstocks (n=100). (A) The optimal rootstocks and scions used for grafting, (B) grafted watermelon seedling, (C) survival Rates and (D) seedling index. *, ** and *** indicate significant differences at p< 0.05, 0.01 and 0.001 levels, respectively.
Figure 2.
Survival rates and seedling index of different seedling age scions grafted onto various seedling age rootstocks (n=100). (A) The optimal rootstocks and scions used for grafting, (B) grafted watermelon seedling, (C) survival Rates and (D) seedling index. *, ** and *** indicate significant differences at p< 0.05, 0.01 and 0.001 levels, respectively.
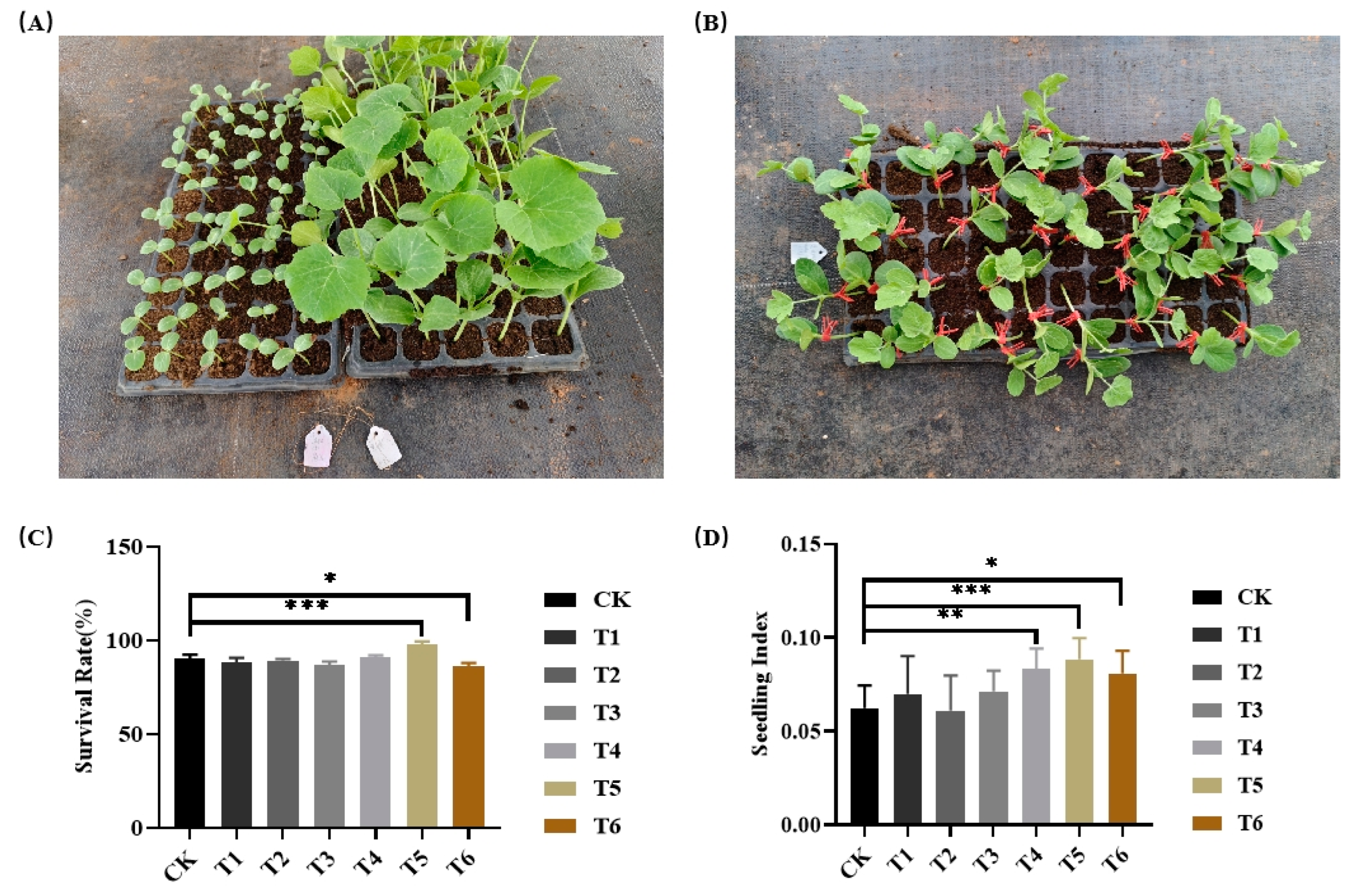
Figure 3.
The concrescence characteristics of watermelon graft junction on the different days after grafting(× 400). (A) Watermelon graft junction on the 6 days, (B) watermelon graft junction on the 8 days, (C) watermelon graft junction on the 10 days in the control after grafting, (D) watermelon graft junction on the 6 days, (E) watermelon graft junction on the 8 days, (F) watermelon graft junction on the 10 days in the Yd/Hn after grafting. H: ‘Heiniu’ rootstock, Y: ‘Youdu’ scion, NL: Necrotic layer, CT: Callus tissue, Bars=500 μm.
Figure 3.
The concrescence characteristics of watermelon graft junction on the different days after grafting(× 400). (A) Watermelon graft junction on the 6 days, (B) watermelon graft junction on the 8 days, (C) watermelon graft junction on the 10 days in the control after grafting, (D) watermelon graft junction on the 6 days, (E) watermelon graft junction on the 8 days, (F) watermelon graft junction on the 10 days in the Yd/Hn after grafting. H: ‘Heiniu’ rootstock, Y: ‘Youdu’ scion, NL: Necrotic layer, CT: Callus tissue, Bars=500 μm.
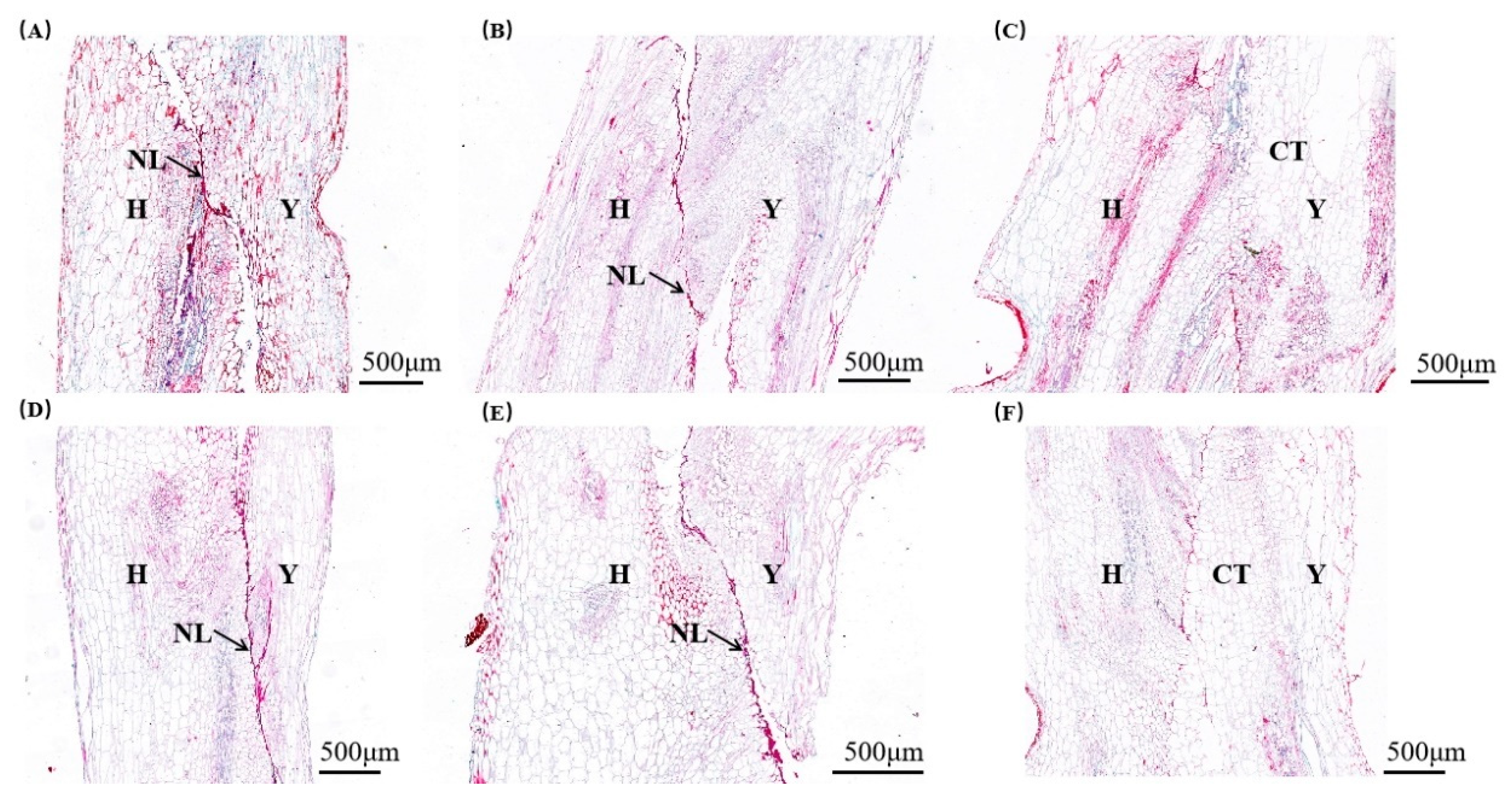
Figure 4.
The hormone contents in watermelon and Hn/Yd seedling at 6, 8,10 DAG. (A) The ABA content, (B) the IAA content, (C) the ZR content, (D) the GA3 content and (D) the ETH content. *, ** and *** indicate significant differences at p < 0.05, 0.01 and 0.001 levels, respectively.
Figure 4.
The hormone contents in watermelon and Hn/Yd seedling at 6, 8,10 DAG. (A) The ABA content, (B) the IAA content, (C) the ZR content, (D) the GA3 content and (D) the ETH content. *, ** and *** indicate significant differences at p < 0.05, 0.01 and 0.001 levels, respectively.
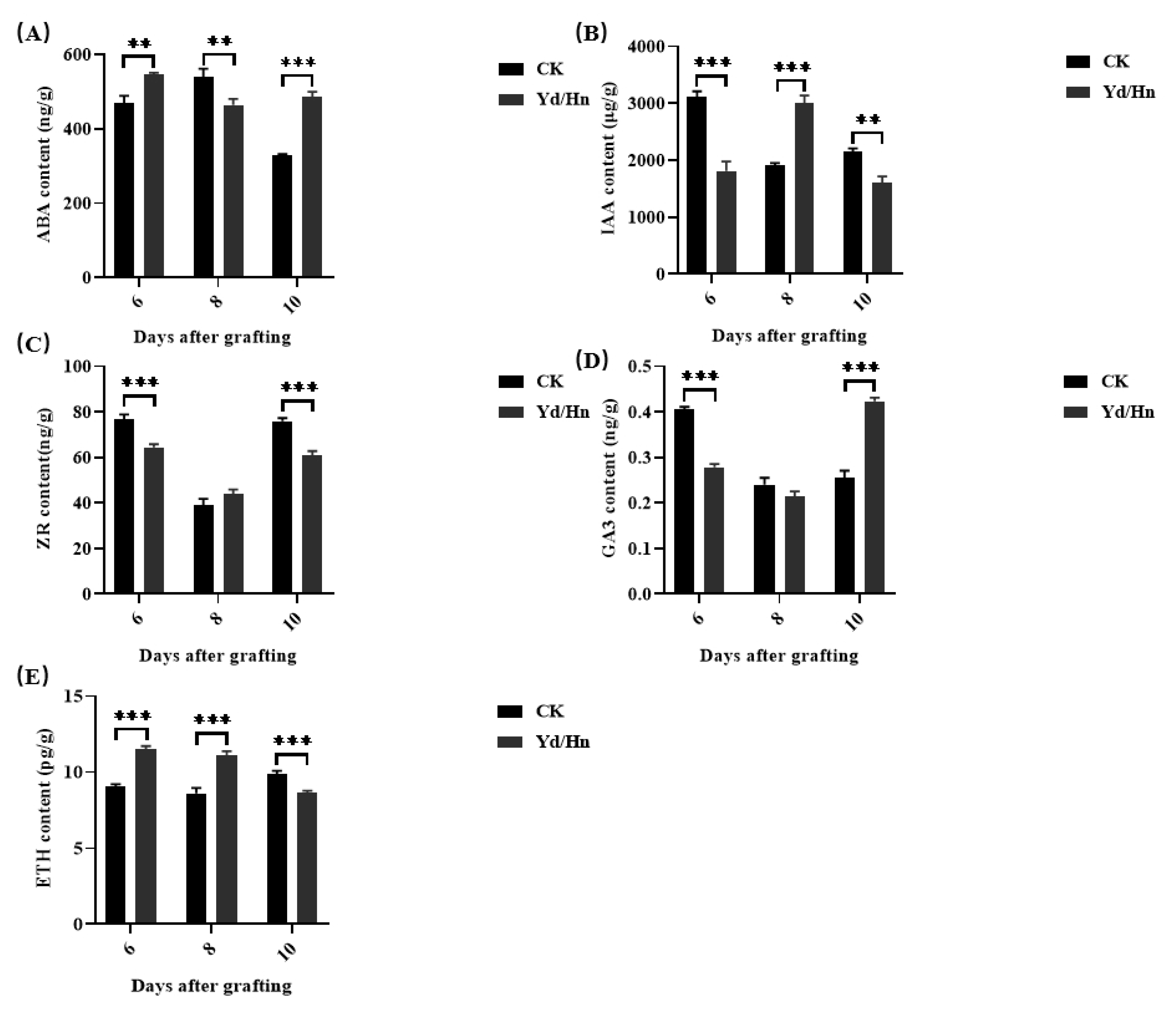
Figure 5.
The protease levels in watermelon seedlingYd/Hn and at 6, 8,10 DAG. (A) The β-1,3 glucanase level, (B) the expansin level and (C) the XTH level. *, ** and *** indicate significant differences at p < 0.05, 0.01 and 0.001 levels, respectively.
Figure 5.
The protease levels in watermelon seedlingYd/Hn and at 6, 8,10 DAG. (A) The β-1,3 glucanase level, (B) the expansin level and (C) the XTH level. *, ** and *** indicate significant differences at p < 0.05, 0.01 and 0.001 levels, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



