
Submitted:
18 April 2023
Posted:
19 April 2023
You are already at the latest version
Abstract
The corpus luteum is a temporary reproductive endocrine structure established from a ruptured follicle wall after ovulation. In the ovarian cycle of ruminant species, the corpus luteum organ undergoes a repeated pattern of specific cellular proliferation, differentiation, and transformation. The corpus luteum encounters several physiological events, including growth, function, and regression throughout its life span. Also, the corpus luteum comprises granulosa and theca cells and endothelial cells, such as luteal steroidogenic cells and luteal endothelial cells. These cells play an important role in the physiology of the corpus luteum and the maintenance and degeneration of the corpus luteum. Furthermore, the role of reproductive hormones in the ovaries is important. Representative hormones include estrogen, progesterone, prostaglandin F2α, and oxytocin. Understanding their functions is important in studying the physiological phenomena and various mechanisms of the corpus luteum in the ovary. Therefore, the following review will discuss the role of reproductive hormones and luteal cell types in the microenvironment of the corpus luteum in the bovine ovary.
Keywords:
reproductive hormone
; luteal cells
; ovary
; corpus luteum
; cow
1. Introduction
The female reproductive organ, like the ovary, is a dynamic tissue that expresses rapid growth and regression periodically [1,2]. In an ovarian cycle, the repeated series of cellular proliferation, differentiation, and transformation transpire during the follicular development and the formation and regression of the corpus luteum [3]. The corpus luteum, a temporary endocrine gland structure, is the principal function of producing the progesterone hormone required for establishing and maintaining a pregnancy [4,5]. For the life span of the corpus luteum, this transient reproductive gland encounters many physiological events, including growth, function, and regression [3,6]. Interestingly, the establishment of the corpus luteum is maintained by reproductive hormones, which estrogen, progesterone, prostaglandin F2α, and oxytocin regulate regression of the corpus luteum. It is well established that in the female reproductive system, angiogenesis and apoptosis are required for the luteinization and luteolysis of the corpus luteum, respectively [7,8].
The Corpus luteum composes of luteal steroidogenic cells and luteal endothelial cells, and other cell types. Luteal steroidogenic cells are derived from follicular granulosa and theca cells, and luteal endothelial cells originated from neovascularization during the corpus luteum formation in humans and cows [9]. Both luteal cell types have different cell functions and morphological characteristics, and responses to reproductive hormones during the formation and development of the ovarian corpus luteum. Thus, luteal steroidogenic cells and luteal endothelial cells play a role in the formation and regression of the corpus luteum. Taken together, this review aims to propose the functions of the reproductive hormones and luteal cells in the microenvironment of corpus luteum formation and regression.
2. The functions of reproductive hormone in the ovarian corpus luteum
Cattles are polyestrous animals exhibiting estrous behavior that lasts for 18-24 days of every estrous cycle, which consists of a follicular phase (4-6 days) and a luteal phase (14-18 days) [10]. The estrous cycle is coordinated under the actions of the hypothalamic-pituitary-gonadal axis and its regulation of the female reproductive organ, including the uterus and ovary [11]. The mammalian ovary is an extremely dynamic organ composed of a highly organized composite of germ cells (oocyte or eggs) and somatic cells (granulosa cells, thecal cells, and stromal cells) [12,13]. The ovary plays an important role in the orchestration of the estrous cycle by producing several hormones, such as E2, P4, and inhibin [10].
The endocrine system of hormonal regulation in the estrous cycle performs through the mechanism of a positive and negative feedback system, Figure 1. [14]. During the follicular phase, the pituitary hormones, including FSH and LH, whose secretion is provoked by hypothalamus-derived GnRH, provide the major mechanism that regulates follicular selection and dominance via feedback loops with the hypothalamus-pituitary ovarian axis [13]. The beginning of follicle development during the estrous cycle is characterized by the emergence of a follicle cohort consisting of approximately 5-20 primary follicles. It has corresponded to a transient increase in FSH concentrations [15]. FSH is the primary hormone that contributes to the process of follicular growth resulting in the secretion of E2 and inhibin originated from a large dominant follicle chosen from a pool of developing follicles that will be destined to ovulate [16].
The growth of the follicles during the ovarian folliculogenesis process leads to the surrounding layers of it, which are granulosa cells starting the production of estrogen hormone [17]. Granulosa cells are the primary source of estrogen hormone production and also one of the targets of estrogen hormone since they express the estrogen receptors (ERα and ERβ) [13,18]. The production of estrogen hormones within the granulosa cells involves several steps through the secretion of androgens by the thecal cells. They are utilized by the granulosa cells and converted by p450aromates to various estrogen molecules with the main one being estrogen hormones [19]. Estrogen and inhibin produced by the granulosa cells act on the hypothalamic-pituitary system and decrease FSH secretion to the basal level known as a negative ovarian feedback mechanism to restrain the further growth of subordinate follicles [13].
The subordinate follicles are not developing further and undergo a degeneration process known as follicular atresia through the cell apoptosis mechanism [13,20]. On the other hand, the dominant follicle continues growing to become a mature follicle in the face of a low concentration of FSH and concomitantly becomes more responsive to LH [10]. The LH receptors begin to appear on the granulosa cell of the growing dominant follicle. The transition from FSH independency to LH independency occurs for further development of the dominant follicle, resulting in the ovulation event induced by an LH surge [13]. Before ovulation, in which the dominant follicle transforms into a Graafian follicle, the LH surge occurs due to prolonged exposure of estrogen hormone to the anterior pituitary gland in a positive feedback mechanism that stimulates the massive release of LH into the bloodstream [17]. To the releasing oocyte from the follicle, many proteolytic enzymes and prostaglandin are activated, leading to the digestion of the follicle wall collagen resulted in ovarian follicle ruptured of ovulation events [21].
Immediately afterward, the ovulation takes place, and the luteal phase begins to proceed with the corpus luteum formation [10]. During this initial stage of corpus luteum, known as corpus hemorrhagic, the remaining granulosa cells, which are not released along with the oocyte during the ovulation process, enlarge and acquire lutein (carotenoids) that is yellow in color [17]. The formation of the corpus luteum following the preovulatory LH surge involves a sequence of morphologic and biochemical alteration in cells of theca interna and granulosa cells of the preovulatory follicle in which these changes are termed luteinization [20]. Luteinization is a process of cell transformation from the follicular cells into the luteal cells that is characterized by increased steroid hormone production and a transition from generating estrogen to progesterone hormones [3]. Progesterone concentrations start to increase due to the corpus luteum formation in which the granulosa cells and theca cells of the ovulated dominant follicle luteinize [10].
The major function of the corpus luteum is to produce progesterone hormone, which is required for the establishment and maintenance of pregnancy [5]. The progesterone concentration remains elevated after the corpus luteum formation, which acts on the hypothalamic-pituitary system through a negative feedback mechanism that only allows the secretion of greater amplitude. Still, the lesser frequency of LH resulted in an inadequate amount of LH for ovulation of the dominant follicle that simultaneously develops by the release of FSH from the anterior pituitary [10]. The progesterone hormone is required for the endometrial remodeling process, which changes the endometrial lining from a proliferative stage into a secretory phase to prepare the embryo implantation [17]. However, in the absence of fertilization and embryo implantation, the corpus luteum will undergo a self-destruction mechanism as a consequence of the inactivation of precise interaction between pituitary and embryonic gonadotropins, nor intra-luteal autocrine and paracrine signals that regulate the endocrine function of luteal cells [3].
In ruminants and other domestic animals, the luteal regression is provoked by the episodic release of PGF2α from the uterus by reaching the corpus luteum organ through a counter-current system between the vena uterine and arteria ovaries [3]. PGF2α lead to corpus luteum regression by disrupting LH action on the corpus luteum and increasing the secretion of oxytocin by the ovary. The episodic secretion of PGF2α from the uterus is propagated in response to the binding of oxytocin to newly developed receptors in the uterus [10]. The estrogen hormones produced by the ovary contribute to increasing the modulation and development of the number of oxytocin receptors in the uterus. The oxytocin secreted by the corpus luteum promotes the luteolysis process by regulating the double-positive feedback mechanism [22]. Oxytocin plays an important role in amplifying and modulating the course of induced luteolysis as a regulator of the pulsatile amplitude of PGF2α secretion [23]. The secretion of oxytocin in the bovine corpus luteum was discovered to be parallel with the pulsatile release of uterine PGF2α which occurs as a result of oxytocin activating the phosphatidylinositol hydrolysis pathway [24]. The PGF2α was found to have an essential role in inducing the release of luteal oxytocin in the cow corpus luteum [22]. The release of oxytocin from the ovarian corpus luteum that acts on the uterus perpetuates a pulsatile release of PGF2α, allowing luteolysis to occur [25]. In addition, the oxytocin secreted by the posterior pituitary stimulates the release of PGF from the uterine endometrium [26]. During luteolysis, the corpus luteum organ undergoes function and structural regression characterized by the decreasing progesterone production that the luteal cells die through the programmed cell death mechanism [26]. This reduces the circulating progesterone concentration, increasing the estrogen hormone concentration, and the GnRH produced by the hypothalamus stimulates the new estrous cycle [10]. Eventually, the ovary, a cytogenic and hormone-secreting organ, undergoes a repeated pattern of the estrous cycle that involves different functional stages, including the maturation of oocytes, the ovulation event, the corpus luteum formation, and regression [27]. In summary, the key hormones that take an essential part in the regulation of the estrous cycle include progesterone (P4), estrogen (E2), gonadotropin-releasing hormone (GnRH), follicle-stimulating hormone (FSH), luteinizing hormone (LH), inhibins, prostaglandin F2α (PGF2α), and oxytocin [10,17]. Further descriptions of those essential hormones in ovarian regulation are elucidated below.
2.1. Estrogen
Estrogen (E2) is a steroid hormone, the primary female sex hormone that plays a vital role in the estrous cycle [28]. Estrogens are produced fundamentally by the developing follicles in the ovaries and the corpus luteum [29]. The secretion of estrogen from the developing follicle induces the luteinizing hormone (LH) surge that results in the event of ovulation [30]. During the luteal phase, the secretion of estrogen produced by the corpus luteum, concomitantly along with the production of progesterone, plays an essential role in preparing the endometrium for the possibility of an implanted embryo [31]. In addition, the estrogens throughout the parturition process also substantially contribute to increasing oxytocin receptors and promoting the synthesis of prostaglandins [32]. The synthesis of estrogens begins in theca interna cells in the ovary through the synthesis of androstenedione from cholesterol [33]. The androstenedione secreted by the theca interna cells then traverses through the basal membrane into the intracellular cell of the surrounding granulosa cells, where it is converted to estrone or estradiol, either immediately or through testosterone, under the action of aromatase [29]. As a lipophilic molecule, the estrogens loosely cross the cell membrane and interact with a cytoplasmic target cell receptor. The estrogen then binds with the intracellular receptor to form the steroid receptor complex that penetrates the nucleus and modulates the gene transcription [28]. The genomic pathway of estrogen action is initiated by the binding of chemicals or ligands with the nuclear ERs, ERα, and ERβ. The ligand-bound ERs complexes act as transcription factors to up-regulate or down-regulate the transcription of estrogen-responsive target genes [34].
2.2. Progesterone
Progesterone (P4) is a steroid hormone involved in various physiological functions, including the female reproductive cycle, pregnancy, and embryogenesis [35]. Furthermore, progesterone is essential in regulating female fertility, with robust actions throughout the female reproductive axis [36]. Progesterone is a chemical name of 4-pregnene-3,20-dione, a c21 steroid hormone, with a double bond in the A ring and a ketone on carbon 3 [37]. The synthesis and release of progesterone hormones in the theca and luteal cells of the ovary are modulated by LH. There are various genes involved in progesterone synthesis, including steroidogenic acute regulatory protein (STAR), cholesterol side-chain cleavage enzyme (CYP11A1) and its partner's ferredoxin (FDX1) and ferredoxin reductase (FDXR), and 3β-hydroxysteroid dehydrogenase/delta5 delta4- isomerase (HSD3B1 or HSD3B2) [38]. Progesterone is synthesized from pregnenolone, a derivative of cholesterol molecules, under the action of cholesterol side-chain cleavage enzyme and 3-beta-hydroxysteroid dehydrogenase [35]. Progesterone mainly acts through the intracellular progesterone receptor [39]. Once progesterone is bound to the receptor, the progesterone receptor complex translocates to the nucleus. It attaches directly to particular nucleotide sequences of the chromosomal DNA, altering the transcriptional activity of specific genes [38]. When the P4 is absent, the progesterone receptor is bound by chaperone protein (hsp90, hsp70, and hsp59) and is located in either the cytoplasm or the nucleus [37]. In addition, the action of progesterone through the non-genomic mechanism is mediated by the cell membrane receptor called PGRMC1 [40]. The expression of the progesterone cell membrane receptor appears to be regulated under the control of the gonadal steroid hormone [40,41].
2.3. Prostaglandin F2α
Prostaglandin F2α (PGF2α) is an essential regulator of corpus luteum function, uterine contractility, ovulation, and embryo attachment [42]. The PGF2α hormone is a primary hormone that is responsible for inducing corpus luteum regression [43]. The uterine endometrium of the female reproductive organ is the most important source of PGF2α production [44]. During the spontaneous luteolysis of the estrous cycle, the administration of PGF2α causes a dramatic decrease in progesterone secretion and structural degeneration of the corpus luteum [43]. The uterine endometrial PGF2α is secreted into the uterine vein and moved by counter-current transfer to the ovarian artery, the ipsilateral ovary, and the systemic circulation [45]. The plasma profile of PGF2α usually describes the concentration of PGF2α metabolite (PGFM) [46]. The mechanism of PGF2α in inducing the luteal regression is related to the acquisition of luteolytic capacity in the corpus luteum that involves the PGF2α blocking cholesterol transport pathways (StAR, LDL receptor) through PGF2α regulation of LH receptors and DAX1 [47]. The action of PGF2α is regulated by a systemic and a local mechanism to induce corpus luteum regression [44]. The role of PGF2α is primarily mediated by a plasma membrane receptor termed FP receptor, which these prostaglandin receptors are a member of the seven-transmembrane domain receptor [45]. The specific high affinity of PGF2α receptors is available in the corpus luteum at all stages of the estrous cycle, and they are majorly located on large luteal [47]. In the corpus luteum organ, the PGF2α binds to the G-coupled-protein FP receptors leading to an elevation of free intracellular calcium and activation of protein kinase C (PKC) [45]. The activation of FP receptors by the PGF2α in responsive corpus luteum initiates a cascade of intracellular events resulting in luteolysis [44].
2.4. Oxytocin
Oxytocin is a peptide hormone composed of nine amino acids (nano peptide) with a disulfide bridge between cysteine residues 1 and 6 [48]. Oxytocin, an α-Hypophamine, is a neurohypophysial hormone secreted from magnocellular neurosecretory cells of the hypothalamus supraoptic and paraventricular nuclei that are stored in the posterior pituitary until its release into the bloodstream [49]. Those neural secretions of oxytocin from neurosecretory nerve endings are regulated by the electrical activity of the cells in the hypothalamus [50]. However, oxytocin is also identified to be produced by non-neural sources using radioimmunoassay and high-performance liquid chromatography methods [48]. The oxytocin mRNA transcripts and oxytocin receptors have been found to local synthesis within the reproductive tract [51]. The female reproductive tracts that secrete oxytocin hormone include the oviduct, uterus, and corpus luteum organ [48]. Oxytocin also plays an important role in the stimulatory action on the synthesis of PGF2α through the indirect effect of oxytocin [52]. The local secretion of oxytocin outside the neural tissue production termed non-neural oxytocin indicates a paracrine role near the sites of secretion [51]. The oxytocin receptors have been detected and localized within the tissues of the female reproductive tract, suggesting an essential role of the oxytocin receptors as a critical paracrine regulator [49]. The oxytocin receptors are a seven transmembrane-domain polypeptide belonging to the Rhodopsin Type Class-I G-protein coupled receptor family [48]. The oxytocin receptors are coupled with the Gq subtype of G-proteins that bind to GTP and stimulate the activity of the phospholipase-C enzyme that triggers the release of intracellular Ca++, which in turn triggers a variety of cellular events [51].
3. Luteal cell types of the corpus luteum
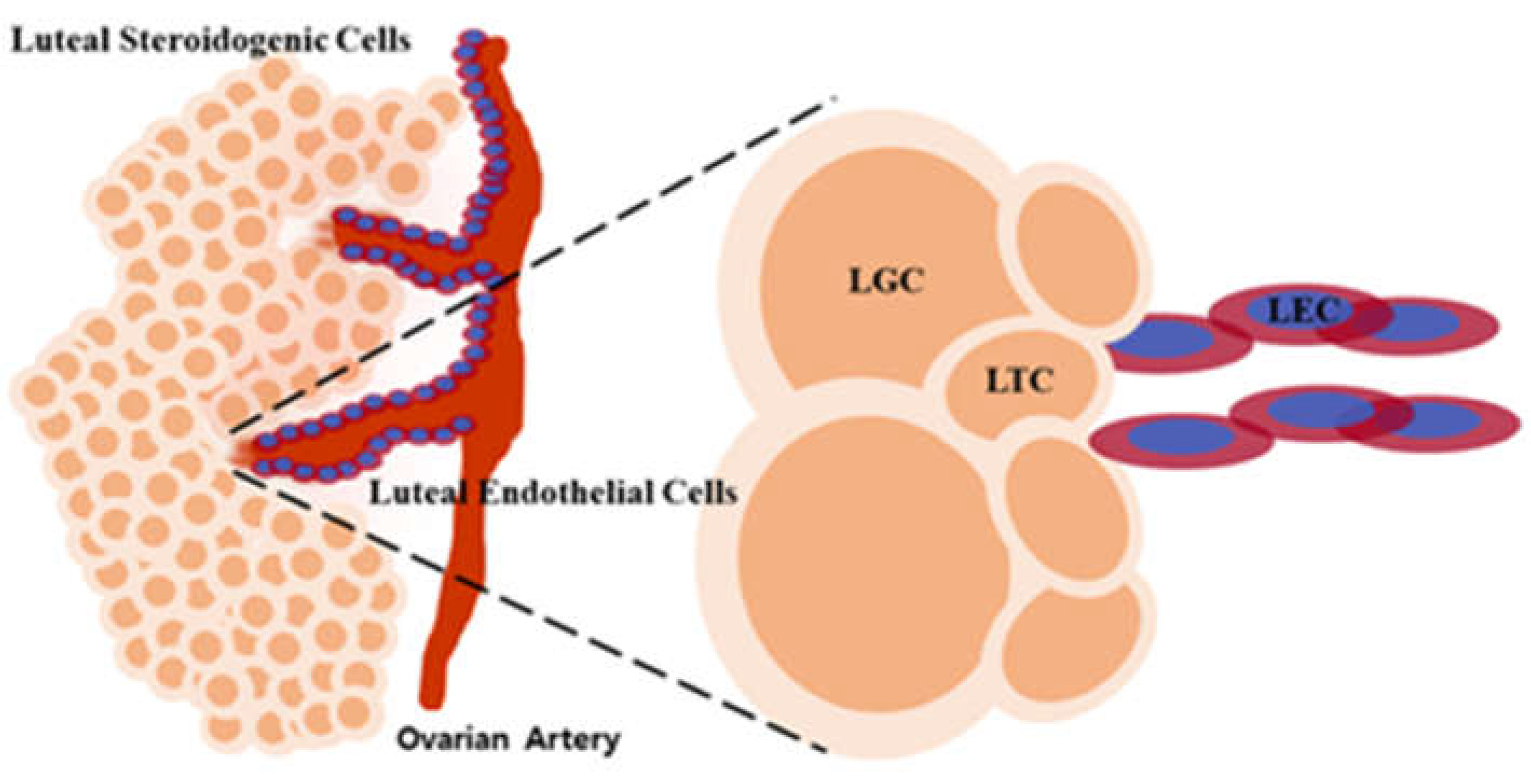
The ovarian corpus luteum also well-known as the yellow body, is a temporary reproductive endocrine gland [53]. The corpus luteum organ has a primary function in synthesizing the progesterone hormone required for the initiation and maintenance of pregnancy [3]. The corpus luteum holds three major stages, including luteinization, maintenance, and degeneration, throughout its lifespan [54]. This transient endocrine gland grows, develops, and reaches structural and functional sophistication by the middle of the luteal phase. It will undergo regression in a non-fertile cycle circumstance [55]. In a normal stage of the luteal phase, the corpus luteum growth and function are significantly related to the luteotropic hormones that promote the increase of structural enlargement in size and support the rise of functional ability in secreting the progesterone hormone [4]. As Figure 2, the corpus luteum is a heterogeneous tissue comprising large luteal cells, small luteal cells, endothelial cells, pericytes, fibroblasts, smooth muscle cells, and other cell types [56]. Those heterogeneous cell populations of corpus luteum can be categorized into two main representatives of cell types: non-steroidogenic cells, such as microvascular cells, connective tissue cells, and immune cells, and steroidogenic cells, such as large luteal cells and small luteal cells [57].
3.1. Luteal steroidogenic cell
There are two primary luteal steroidogenic cells within the corpus luteum organ. Those steroidogenic cells can be differentiated based on the follicular origin source and morphological and biochemical characteristics [4]. Steroidogenic cells are referred to the capacity of cells to deal with steroidogenesis, which is the process of specialized cells in specific tissues synthesis steroid hormones, an essential class of terpene base or small lipid molecules [26]. Those steroid hormones have various functions however are synthesized by biosynthetic pathways that are identical in the initial stages [58]. Generally, the biosynthesis of steroid hormones is controlled mainly by the pituitary tropic hormones, such as luteinizing hormone and follicle-stimulating hormone [5]. In all steroidogenic cells, regardless of the hormones synthesized, the initial step in steroidogenesis is the conversion of cholesterol to the first steroid, which is pregnenolone [59]. This conversion occurs through the actions of the cytochrome P450 side-chain cleavage enzyme, which exists in the inner mitochondrial membrane in all steroidogenic cells [60]. Then, the pregnenolone proceeds in the mitochondrial organelle are converted to progesterone in the microsomal compartment of the steroidogenic cells [61]. The luteal steroidogenic cells within the ovarian corpus luteum synthesize and secrete the progesterone hormone via the conversion of pregnenolone by the 3β-hydroxysteroid dehydrogenase enzyme [5]. Progesterone secreted by the corpus luteum is a critical regulator of normal cyclical reproductive function in the female mammalian species [31]. The luteal steroidogenic cells of the ovarian corpus luteum organ responsible for progesterone secretion are the large and small luteal cells [62]. In addition, the term of large luteal cells and small luteal cells are also referred to as luteal granulosa cells (LGCs) and luteal theca cells (LTCs), respectively [4].
3.2. Luteal granulosa cell
The origin of large luteal cell formation is presumably derived from the follicular granulosa cells [62]. The morphological characteristic of luteal granulosa cells under the microscope level represents that LGCs are polyhedral in shape and usually contain lipid droplets [63]. The size of LGCs identified in the bovine ranges around 20 - 50μm. Furthermore, the feature of LGCs can be confirmed biochemically through the expression level of biomolecules, including the oxytocin mRNA and LH receptor mRNA. The LGCs tend to exhibit a characteristic of a high expression level of oxytocin mRNA [9]. The LGCs mainly secrete the progesterone hormone in large amounts. The LGCs have a characteristic of high basal steroid hormone production but are less or not responsive to the administration of LH [64]. Furthermore, the LGCs undergo hypertrophy, which increases cellular size due to the enlargement of its cell components during corpus luteum formation [65].
3.3. Luteal theca cell
The emergence of small luteal cells appears to be originated mainly from the follicular theca cells [4]. The bovine luteal theca cells were recognized by the size range of roughly about 15 - 19μm [63]. Under the microscope magnification, the luteal theca cells display smaller morphological characteristics and are stained darker than LGCs. The LTCs produce the progesterone hormone, which performs a high response to the administration of LH with a pronounced increase in progesterone synthesis [64]. The biochemical characteristic of LTCs in expressing LH receptor mRNA shows that LTCs tend to exhibit a high expression level of LH receptor mRNA [9]. The progesterone hormone secreted by the LTCs has mainly involved the essential role of LH that directly stimulates the LTCs through activating the protein kinase A second messenger pathway [4]. LTCs play a vital role during corpus luteum formation, in which the proliferation level of LTCs increases throughout the early and developing luteal state, significantly contributing to corpus luteum growth [9]. In addition, the main factor that promotes a robust proliferation process in LTCs during the corpus luteal establishment is the cell proliferation progressive genes expressed more strongly in the LTCs than in LGCs [9].
3.4. Luteal endothelial cell
The luteal endothelial cells own the largest number of corpus luteum cellular composition, accounting for up to 50% of the total luteal tissue [66]. The luteal endothelial cells are categorized into non-steroidogenic cells that cannot produce steroid hormones [65]. The luteal endothelial cells are derived from the migrated endothelial cell originating in the parent vessel walls throughout the corpus luteum formation [66]. The endothelial cells are involved in the development of new capillaries required for supporting the corpus luteum function through the migration from the pre-existing blood vessel [26]. In the corpus luteum establishment, luteal endothelial cells undergo cellular proliferation under the influence of angiogenic factors and hypoxic conditions [67]. The luteal endothelial cells play an essential role as microvascular cells that participate to set up the well-developed capillary bed for establishing an appropriate microenvironment for differentiation and maturation of fully functional corpus luteum [65].
4. Conclusion
The corpus luteum organ is a temporary reproductive gland that undergoes a repeated cycle of formation, function, and regression throughout its lifespan. The major regulator of the ovarian corpus luteum cycle is the endocrine system through the hormonal action of the hypothalamic-pituitary-gonadal axis. On the other hand, the microenvironment within the corpus luteum is considered to play an important role in the regulation of corpus luteum establishment and degeneration throughout its ovarian cycle. The interaction of the cellular milieu around the luteal cells, including the neighboring blood vessel, immune cells, signaling molecule, and extracellular matrix, are essential in many luteal physiological events, including cellular proliferation, differentiation, and transformation for the corpus luteum formation and regression.
Author Contributions
D.H.B and S.L. conceived, structured, and wrote the manuscript. The authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Research Foundation of Korea (NRF 2018R1D1A3B07048167), Republic of Korea.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no competing interests.
References
- Billhaq, D.H. Microenvironment system during the stage of formation and regression of the ovarian corpus luteum in cows. M.S., Kangwon National University, Republic of Korea, February 22, 2020.
- Reynolds, L.P.; Killilea, S.D.; Redmer, D.A. Angiogenesis in the female reproductive system. Faseb J. 1992, 6, 886–892. [Google Scholar] [CrossRef] [PubMed]
- Schams, D.; Berisha, B. Regulation of corpus luteum function in cattle–an overview. Reprod. Domest. Aanim. 2004, 39, 241–251. [Google Scholar] [CrossRef]
- Niswender, G.D.; Juengel, J.L.; Silva, P.J.; Rollyson, M.K.; McIntush, E.W. Mechanisms controlling the function and life span of the corpus luteum. Physiol. Rev. 2000, 80, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Tomac, J.; Cekinović, Đ.; Arapović, J. Biology of the corpus luteum. Period. Biol. 2011, 113, 43–49. [Google Scholar]
- Devoto, L.; Fuentes, A.; Kohen, P.; Céspedes, P.; Palomino, A.; Pommer, R.; Strauss III, J.F. The human corpus luteum: life cycle and function in natural cycles. Fertil. Steril. 2009, 92, 1067–1079. [Google Scholar] [CrossRef]
- Russell, D.L.; Robker, R.L. Molecular mechanisms of ovulation: co-ordination through the cumulus complex. Hum. Reprod. Update 2007, 13, 289–312. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, H.K. Loss luteal sensitivity to luteinizing hormone underlies luteolysis in cattle: A hypothesis. Reprod.. Biol. 2021, 21, 100570. [Google Scholar] [CrossRef]
- Yoshioka, S.; Abe, H.; Sakumoto, R.; Okuda, K. Proliferation of luteal steroidogenic cells in cattle. PLoS One 2013, 8, e84186. [Google Scholar] [CrossRef]
- Forde, N.; Beltman, M.E.; Lonergan, P.; Diskin, M.; Roche, J.F.; Crowe, M.A. Oestrous cycles in Bos taurus cattle. Anim. Reprod. Sci. 2011, 124, 163–169. [Google Scholar] [CrossRef]
- Carletti, M.Z.; Christenson, L.K. MicroRNA in the ovary and female reproductive tract. J. Anim. Sci. 2009, 87, E29–E38. [Google Scholar] [CrossRef]
- Richards, J.S.; Pangas, S.A. The ovary: basic biology and clinical implications. J. Clin. Invest. 2010, 120, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, F.; Inoue, N.; Manabe, N.; Ohkura, S. Follicular growth and atresia in mammalian ovaries: regulation by survival and death of granulosa cells. J. Reprod. Develop. 2012, 58, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Roche, J.F. Control and regulation of folliculogenesis--a symposium in perspective. Rev. Reprod. 1996, 1, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Ginther, O.J.; Bergfelt, D.R.; Beg, M.A.; Kot, K. Role of low circulating FSH concentrations in controlling the interval to emergence of the subsequent follicular wave in cattle. Reproduction 2002, 124, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.G.; Robinson, R.S.; Mann, G.E.; Webb, R. Endocrine and paracrine control of follicular development and ovulation rate in farm species. Anim. Rreprod. Sci. 2004, 82, 461–477. [Google Scholar] [CrossRef]
- Beshay, V.E.; Carr, B.R. Clinical Reproductive Medicine and Surgery; Springer International Publishing: Cham, USA, 2017; pp. 1–17. [Google Scholar]
- Berisha, B.; Pfaffl, M.W.; Schams, D. Expression of estrogen and progesterone receptors in the bovine ovary during estrous cycle and pregnancy. Endocrine 2002, 17, 207–214. [Google Scholar] [CrossRef]
- Robinson, R.S.; Mann, G.E.; Reynolds, T.S.; Lamming, G.E.; Wathes, D.C. Expression of oxytocin, oestrogen and progesterone receptors in uterine biopsy samples throughout the oestrous cycle and early pregnancy in cows. Reproduction 2001, 122, 965–979. [Google Scholar] [CrossRef]
- Berisha, B.; Schams, D. Ovarian function in ruminants. Domest. Anim. Endocrinol. 2005, 29, 305–317. [Google Scholar] [CrossRef]
- Espey, L.L. Current status of the hypothesis that mammalian ovulation is comparable to an inflammatory reaction. Biol. Reprod. 1994, 50, 233–238. [Google Scholar] [CrossRef]
- Stormshak, F. Biochemical and endocrine aspects of oxytocin production by the mammalian corpus luteum. Reprod. Biol. Endocrinol. 2003, 1, 92. [Google Scholar] [CrossRef]
- Kotwica, J.; Skarzynski, D.; Miszkiel, G.; Melin, P.; Okuda, K. Oxytocin modulates the pulsatile secretion of prostaglandin F2αin initiated luteolysis in cattle. Res. Vet. Sci. 1999, 66, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Jamshidi, A.A.; Girard, D.; Beaudry, F.; Goff, A.K. Progesterone metabolism in bovine endometrial cells and the effect of metabolites on the responsiveness of the cells to OT-stimulation of PGF2α. Steroids 2007, 72, 843–850. [Google Scholar] [CrossRef]
- Ohtani, M.; Kobayashi, S.I.; Miyamoto, A.; Hayashi, K.; Fukui, Y. Real-Time Relationships between Intraluteal and Plasma Concentrations of Endothelin, Oxytocin, and Progesterone during Prostaglandin F2α-Induced Luteolysis in the Cow. Biol. Reprod. 1998, 58, 103–108. [Google Scholar] [CrossRef]
- Stocco, C.; Telleria, C.; Gibori, G. The molecular control of corpus luteum formation, function, and regression. Endocr. Rev. 2007, 28, 117–149. [Google Scholar] [CrossRef] [PubMed]
- Plendl, J. Angiogenesis and vascular regression in the ovary. Anat. Histol. Embryol. 2000, 29, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Kiyama, R.; Wada-Kiyama, Y. Estrogenic endocrine disruptors: Molecular mechanisms of action. Environ. Int. 2015, 83, 11–40. [Google Scholar] [CrossRef] [PubMed]
- Young, J.M.; McNeilly, A.S. Theca: the forgotten cell of the ovarian follicle. Reproduction 2010, 140, 489–504. [Google Scholar] [CrossRef]
- Rimon-Dahari, N.; Yerushalmi-Heinemann, L.; Alyagor, L.; Dekel, N. Molecular mechanisms of cell differentiation in gonad development; Springer International Publishing: Cham, USA, 2016; pp. 167–190. [Google Scholar]
- Stouffer, R.L.; Bishop, C.V.; Bogan, R.L.; Xu, F.; Hennebold, J.D. Endocrine and local control of the primate corpus luteum. Reprod. Biol. 2013, 13, 259–271. [Google Scholar] [CrossRef]
- Makieva, S.; Saunders, P.T.; Norman, J.E. Androgens in pregnancy: roles in parturition. Hum. Reprod. Update 2014, 20, 542–559. [Google Scholar] [CrossRef]
- Patel, S.S.; Beshay, V.E.; Escobar, J.C.; Carr, B.R. 17α-Hydroxylase (CYP17) expression and subsequent androstenedione production in the human ovary. Reprod. Sci. 2010, 17, 978–986. [Google Scholar] [CrossRef]
- Moggs, J.G.; Orphanides, G. Estrogen receptors: orchestrators of pleiotropic cellular responses. EMBO Rep. 2001, 2, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Taraborrelli, S. Physiology, production and action of progesterone. Acta Obstet. Gynecol. Scand. 2015, 94, 8–16. [Google Scholar] [CrossRef]
- Kubota, K.; Cui, W.; Dhakal, P.; Wolfe, M.W.; Rumi, M.K.; Vivian, J.L.; Soares, M.J. Rethinking progesterone regulation of female reproductive cyclicity. Proc. Natl. Acad. Sci. U.S.A. 2016, 113, 4212–4217. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Miyagawa, S.; Iguchi, T. Handbook of Hormones, 1st ed.; Academic Press: Cambridge, USA, 2015; p. 507. [Google Scholar]
- Davis, J.S.; LaVoie, H.A. Molecular Regulation of Progesterone; Academic Press: Cambridge, USA, 2019; pp. 237–253. [Google Scholar]
- Li, X.; O'Malley, B.W. Unfolding the action of progesterone receptors. J. Biol. Chem. 2003, 278, 39261–39264. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.J. Progesterone receptor membrane component 1 and its role in ovarian follicle growth. Front. Neurosci. 2013, 7, 99. [Google Scholar] [CrossRef] [PubMed]
- Koensgen, D.; Mustea, A.; Klaman, I.; Sun, P.; Zafrakas, M.; Lichtenegger, W.; Sehouli, J. Expression analysis and RNA localization of PAI-RBP1 (SERBP1) in epithelial ovarian cancer: association with tumor progression. Gynecol. Oncol. 2007, 107, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Diaz, F.J.; Anderson, L.E.; Wu, Y.L.; Rabot, A.; Tsai, S.J.; Wiltbank, M.C. Regulation of progesterone and prostaglandin F2α production in the CL. Mol. Cell. Endocrinol. 2002, 191, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Ginther, O.J.; Fuenzalida, M.J.; Shrestha, H.K.; Beg, M.A. The transition between pre-luteolysis and luteolysis in cattle. Theriogenology 2011, 75, 164–171. [Google Scholar] [CrossRef]
- Waclawik, A.; Jabbour, H.N.; Blitek, A.; Ziecik, A.J. Estradiol-17β, prostaglandin E2 (PGE2), and the PGE2 receptor are involved in PGE2 positive feedback loop in the porcine endometrium. Endocrinology 2009, 150, 3823–3832. [Google Scholar] [CrossRef]
- De Rensis, F.; Saleri, R.; Tummaruk, P.; Techakumphu, M.; Kirkwood, R.N. Prostaglandin F2α and control of reproduction in female swine: a review. Theriogenology 2012, 77, 1–11. [Google Scholar] [CrossRef]
- Ginther, O.J.; Shrestha, H.K.; Fuenzalida, M.J.; Imam, S.; Beg, M.A. Stimulation of pulses of 13, 14-dihydro-15-keto-PGF2α (PGFM) with estradiol-17β and changes in circulating progesterone concentrations within a PGFM pulse in heifers. Theriogenology 2010, 74, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Diaz, F.J.; Wiltbank, M.C. Acquisition of luteolytic capacity involves differential regulation by prostaglandin F2α of genes involved in progesterone biosynthesis in the porcine corpus luteum. Domestic animal endocrinology 2005, 28, 172–189. [Google Scholar] [CrossRef] [PubMed]
- Assad, N.I.; Pandey, A.K.; Sharma, L.M. Oxytocin, functions, uses and abuses: a brief review. Theriogenology 2016, 6, 1. [Google Scholar] [CrossRef]
- Froemke, R.C.; Carcea, I. Principles of Gender-Specific Medicine, 3rd ed.; Academic Press: Cambridge, USA, 2017; pp. 161–182. [Google Scholar]
- Salmina, A.B.; Lopatina, O.; Ekimova, M.V.; Mikhutkina, S.V. , Higashida, H. CD38/Cyclic ADP-ribose system: a new player for oxytocin secretion and regulation of social behavior. J. Neuroendocrinol. 2010, 22, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Arrowsmith, S.; Wray, S. Oxytocin: its mechanism of action and receptor signalling in the myometrium. J. Neuroendocrinol. 2014, 26, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Smallman, M.S. Possible Autocrine/Paracrine Action of Progesterone in the Ovine Corpus Luteum. M.S., Oregon State University, USA, June 26, 2018.
- Woad, K.J.; Robinson, R.S. Luteal angiogenesis and its control. Theriogenology 2016, 86, 221–228. [Google Scholar] [CrossRef]
- Jewgenow, K.; Amelkina, O.; Painer, J.; Göritz, F.; Dehnhard, M. Life cycle of feline corpora lutea: histological and intraluteal hormone analysis. Reprod. Domest. Anim. 2012, 47, 25–29. [Google Scholar] [CrossRef]
- Yanhong, Y.; Yan, C.; Sijiu, Y. Histological characteristics of corpus luteum in yak during early pregnancy. In Proceedings of the 2004 International Congress on Yak, Chengdu, China, 19 September 2004. [Google Scholar]
- O'Shea, J.D.; Rodgers, R.J.; D'occhio, M.J. Cellular composition of the cyclic corpus luteum of the cow. Reproduction 1989, 85, 483–487. [Google Scholar] [CrossRef]
- Spanel-Borowski, K. Five different phenotypes of endothelial cell cultures from the bovine corpus luteum: present outcome and role of potential dendritic cells in luteolysis. Mol. Cell. Endocrinol. 2011, 338, 38–45. [Google Scholar] [CrossRef]
- Mishra, S.R.; Palai, T.K. Steroidogenesis in luteal cell: A critical pathway for progesterone production. J. Invest. Biochem. 2015, 4, 170–172. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, Z.; Shen, W.J.; Azhar, S. Cellular cholesterol delivery, intracellular processing and utilization for biosynthesis of steroid hormones. Nutr. Metab. 2010, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- LaVoie, H.A. The Life Cycle of the Corpus Luteum; Springer International Publishing: Cham, USA, 2017; pp. 37–55. [Google Scholar]
- Papadopoulos, V.; Miller, W.L. Role of mitochondria in steroidogenesis. Best Pract. Res. Clin. Endoc. Metab. 2012, 26, 771–790. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.D. Models of luteinization. Biol. Reprod. 2000, 63, 2–11. [Google Scholar] [CrossRef]
- Lei, Z.M.; Chegini, N.; Rao, C.V. Quantitative cell composition of human and bovine corpora lutea from various reproductive states. Biol. Reprod. 1991, 44, 1148–1156. [Google Scholar] [CrossRef]
- Christenson, L.K.; Devoto, L. Cholesterol transport and steroidogenesis by the corpus luteum. Reprod. Biol. Endocrinol. 2003, 1, 1–9. [Google Scholar] [CrossRef]
- Shrestha, K.; Rodler, D.; Sinowatz, F.; Meidan, R. The Ovary, 3rd ed.Academic Press: Rehovot, Israel, 2019; pp. 255–267. [Google Scholar]
- Shirasuna, K.; Akabane, Y.; Beindorff, N.; Nagai, K.; Sasaki, M.; Shimizu, T.; Miyamoto, A. Expression of prostaglandin F2α (PGF2α) receptor and its isoforms in the bovine corpus luteum during the estrous cycle and PGF2α-induced luteolysis. Domest. Anim. Endocrinol. 2012, 43, 227–238. [Google Scholar] [CrossRef]
- Okuda, K.; Nishimura, R. The Life Cycle of the Corpus Luteum; Springer International Publishing: Cham, USA, 2017; pp. 23–36. [Google Scholar]
Figure 1.
Scheme of hormonal feedback mechanism on the ovarian cycle. The reproductive hormones perform a positive and negative feedback mechanism through the endocrine system that regulates the ovarian cycle. During the follicular phase, estrogen controls the secretion of pituitary hormones, including FSH and LH through negative and positive feedback mechanisms. After the ovulation event, progesterone becomes a major hormone that acts on the hypothalamic-pituitary system through a negative feedback mechanism during the luteal phase. However, If the pregnancy does not occur, the PGF2α hormones produced by the uterine will induce the corpus luteum luteolysis, and the oxytocin produced by the corpus luteum will promote the secretion of PGF2α through the double-positive feedback mechanism that allows the new ovarian cycle begins.
Figure 1.
Scheme of hormonal feedback mechanism on the ovarian cycle. The reproductive hormones perform a positive and negative feedback mechanism through the endocrine system that regulates the ovarian cycle. During the follicular phase, estrogen controls the secretion of pituitary hormones, including FSH and LH through negative and positive feedback mechanisms. After the ovulation event, progesterone becomes a major hormone that acts on the hypothalamic-pituitary system through a negative feedback mechanism during the luteal phase. However, If the pregnancy does not occur, the PGF2α hormones produced by the uterine will induce the corpus luteum luteolysis, and the oxytocin produced by the corpus luteum will promote the secretion of PGF2α through the double-positive feedback mechanism that allows the new ovarian cycle begins.
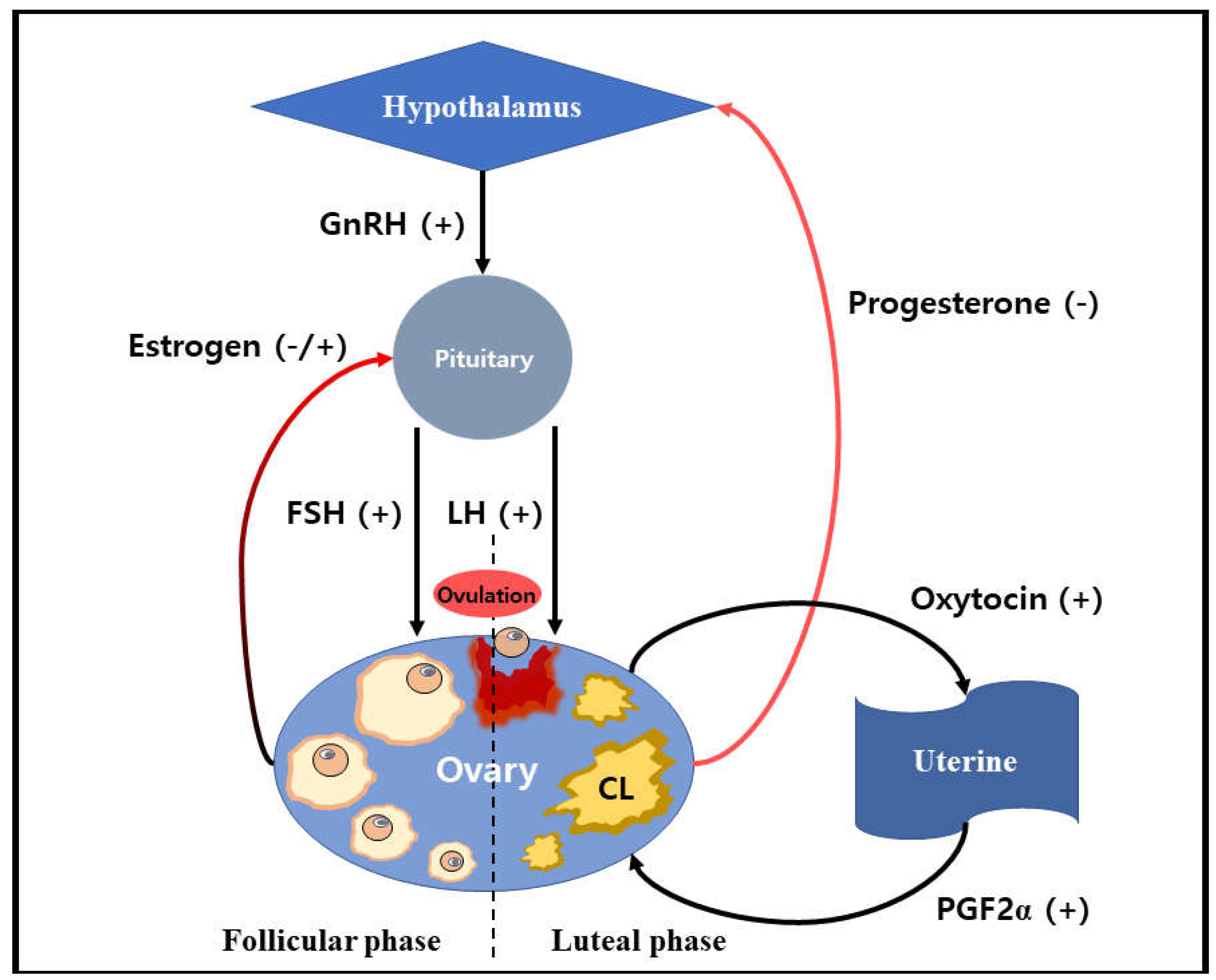
Figure 2.
Diagram of luteal cell types in ovarian corpus luteum of bovine. The Corpus luteum has composed of luteal steroidogenic cells and luteal endothelial cells. LGC, luteal granulosa cell; LTC, luteal theca cell.
Figure 2.
Diagram of luteal cell types in ovarian corpus luteum of bovine. The Corpus luteum has composed of luteal steroidogenic cells and luteal endothelial cells. LGC, luteal granulosa cell; LTC, luteal theca cell.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



