
Submitted:
01 April 2025
Posted:
02 April 2025
You are already at the latest version
Abstract
Treating endometritis is one of the major challenges faced by veterinarians. The effect of post-insemination intrauterine infusion Wharton’s jelly mesenchymal stromal/stem cell-derived conditioned medium (WJ-MSC-CM) on the uterine response and pregnancy outcomes in problem mares was evaluated in this study. The cycles of 11 problem mares were randomly treated seven/eight hours after insemination with intrauterine infusion of 20 ml of WJ-MSC-CM (TRT) or non-conditioned Ringer’s lactate solution (CTR). Before (PRE) and 12 hours after (POST) treatment, an ultrasonographic examination and a low-volume flush (LVF) for efflux clarity evaluation and bacteriological and cytological analysis were made. Pregnancy diagnosis was performed at 14 days and confirmed at 60 days after ovulation. A decrease (P<0.05) in polymorphonuclear neutrophils infiltration and a trend toward significance in LVF efflux clarity (P<0.1) were observed in TRT cycles. Only a trend toward significance was observed comparing pregnancy frequencies between the two groups (CTR 14% vs. TRT 54%), the pregnancy rate of TRT cycles was comparable to that of normal mares (P>0.05) and higher (P<0.05) than that reported for problem mares in the literature. Post-insemination uterine infusion of WJ-MSC-CM in mares restores pregnancy rates to normal levels, by its anti-inflammatory effect.
Keywords:
equine
; endometritis
; mesenchymal stromal/stem cells
; secretome
; conditioned medium
1. Introduction
Endometritis, an infection and/or inflammation of the endometrium, is the leading cause of subfertility and the third most common disease affecting horses [1,2]. It can have infectious and non-infectious etiologies, which often coexist in clinical practice [3]. Post-breeding inflammation occurs in all mares within 30 minutes after insemination [4,5]. This physiological response facilitates the clearance of seminal plasma, excess sperm, microorganisms, and debris from the uterine lumen in preparation for embryo implantation [6]. However, mares susceptible to endometritis exhibit a delayed onset and a prolonged uterine inflammatory response [7,8,9]. This leads to an excessive accumulation of polymorphonuclear neutrophils (PMNs) and intrauterine fluid persisting for up to 96 h or longer post-breeding, adversely affecting embryonic survival and pregnancy establishment [10,11]. While pregnancy rates range from 40% to 65% in healthy mares, they drop below 22% in cases of severe endometrial inflammation [12,13,14]. Given that the equine embryo migrates from the oviduct to the uterine lumen between 144 and 168 h after ovulation [15], inflammation must resolve within a limited timeframe. Moreover, a compromised innate immune response in affected mares hampers microbial clearance, increasing the risk of persistent infection and chronic inflammation [16]. Current treatment strategies aim to mitigate the uterine inflammation and provide antimicrobial support, often in combination with mechanical clearance methods, such as uterine lavages ecbolic administration [17,18,19]. However, conventional therapies demonstrate inconsistent efficacy, and the rising incidence of antimicrobial resistance has fueled interest in alternative approaches [19,20]. Regenerative therapies, including mesenchymal stromal/stem cells (MSCs) and their derivatives, are emerging as promising candidates due to their immunomodulatory and antimicrobial properties [21].
MSCs are self-renewing cells capable of differentiating into multiple mesodermal and ectodermal tissues. They secrete cytokines, growth factors, and extracellular matrix molecules that regulate hematopoiesis, angiogenesis, and immune and inflammatory responses. They are also involved in regenerating and remodeling injured tissues through their trophic activities, such as inhibiting apoptosis [22,23,24]. MSCs can be isolated from various tissues, including bone, cartilage, tendon, muscle, adipose tissue, and fetal membranes [25]. MSCs derived from birth-associated tissues, such as the placenta and the umbilical cord/Wharton’s jelly (WJ-MSCs), exhibit superior proliferative capacity, particularly under hypoxic conditions [26]. Although evidence remains limited, numerous studies have demonstrated the antimicrobial potential of MSCs both in vitro and in vivo [27,28,29]. This antimicrobial activity is attributed to the secretion of antimicrobial peptides (AMPs), which disrupt the cell membranes of bacteria, as well as fungi, yeasts, and viruses [30,31]. Additionally, MSCs exert indirect antimicrobial effects by modulating immune responses, enhancing phagocytosis, and regulating cytokine secretion [32]. Recently, MSCs-derived products, particularly conditioned medium, have gained interest as a cell-free therapy due to their safety, ease of storage, and transportability [33]. MSC-conditioned medium (MSC-CM) is a biological solution containing the secreted factors produced by MSCs during their growth in culture [34].
MSCs secretome includes microvesicles and exosome, which mediate many of the therapeutic effects attributed to MSCs [35]. Importantly, MSC-CM provides similar anti-inflammatory and antimicrobial benefits while reducing risks such as immune rejection, tumorigenesis, and thrombosis [34,35,36].
This study aimed to evaluate the impact of post-insemination intrauterine infusion of Wharton’s jelly mesenchymal stromal/stem cell-derived conditioned medium (WJ-MSC-CM) on pregnancy outcomes in problem mares, by investigating antimicrobial and anti-/pro-inflammatory effects.
2. Materials and Methods
2.1. Animals
This study involved Standardbred mares from a single farm in the Campania Region (Italy), all reproductively managed by the same practitioners during the 2023 breeding season. Mares were selected based on the following criteria: i) failure to conceive during the previous breeding season or after at least two insemination attempts with proven fertile semen in the same year; ii) designation for artificial insemination with cooled semen from stallions of proven fertility; iii) presence of endometrial inflammation or positive bacteriology during post-insemination evaluation; iv) absence of other reproductive disorders as determined by transrectal palpation, ultrasound examination of the genital tract, and vaginal examination at the beginning of the study. A total of eleven mares, aged between 4 and 20 years (median 5), including 6 primiparous and 5 pluriparous, were selected.
2.2. Study Design
Mares were randomly assigned to either the control (CTR) or treatment (TRT) cycle. If they did not become pregnant, they were subsequently assigned to the alternate cycle. If they became pregnant, they were not treated again. The intrauterine infusions were performed 7-8 h after artificial insemination with cooled semen. Transrectal ultrasound evaluation of the reproductive tract for uterine edema and fluid accumulation and a uterine low-volume flush (LVF) was performed immediately before (PRE) and 12 h after (POST) intrauterine infusion. The LVF was analyzed for bacteriology, cytology, and interleukin-10 (IL-10) concentrations. Pregnancy was diagnosed by ultrasonography 14 days after ovulation and confirmed at 60 days.
2.3. Preparation of the Conditioned Medium
Umbilical cords (UCs) were collected immediately after foaling from mares hospitalized for attended delivery at the Perinatology and Reproduction Unit of the Equine Clinical Service, Department of Veterinary Medical Sciences (DIMEVET, University of Bologna, Italy). Ony UCs from healthy mares with normal pregnancies, eutocic deliveries, and healthy foals were collected, as described by Lanci et al. (2023) [37].
Immediately after spontaneous rupture, the UC portion closest to the foal, characterized by an abundant amount of WJ, was collected. Samples were stored in Dulbecco’s phosphate buffered saline (DPBS) containing 100 IU/ml penicillin and 100 mg/ml streptomycin, at 4 °C for at the latest maximum of 12 h, and transferred to the Animal Reproduction and Biotechnology Laboratory of the Equine Clinical Service (DIMEVET, University of Bologna, Italy). MSCs were isolated from the Wharton’s Jelly and expanded in vitro following the protocols described by Iacono et al. (2012) [38], then frozen as described by Merlo et al. (2016) [39]. Cells were stored for at least 4 months in liquid nitrogen before thawing for conditioned medium preparation. WJ-MSCs were thawed, platelet in T75 flask at a density of 1x106 cells/flask, and cultured in Dulbecco’s Minimum Essential Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) (Gibco, Thermo Fisher Scientific, Italy) until 80-90% confluence. Cells were then washed three times with DPBS and the culture medium was replaced with sterile Ringer’s Lactate solution (for IV administration, Galenica Senese, Italy). After 24 h of culture, the solution was recovered and centrifuged at 4000g for 30 minutes at 25°C, to remove cellular debris. The supernatant was collected as WJ-MSC-CM and frozen at -80°C until use.
2.4. Insemination and Treatment
The estrous stage was monitored by transrectal ultrasound examination twice a week. In the presence of a pre-ovulatory follicle (35x35 mm) and endometrial edema (score 2/5 to 4/5) [28,29], ovulation was induced with an intravenous injection of 2000 IU of human chorionic gonadotropin (Corulon®, Intervet Italia, Italy.). Artificial inseminations with cooled semen from stallions of proven fertility, selected based on owner preference, were performed 24 h after ovulation induction using an artificial insemination catheter. After 7-8 h, mares received an intrauterine infusion of either 20 ml of Ringer’s Lactate (CTR group) or WJ-MSC-CM (TRT group) via a sterile insemination catheter.
Transrectal ultrasound evaluation and LVF were performed immediately before (PRE) and 12 h after (POST) intrauterine infusion. Fluid accumulation > 2 cm and uterine status were recorded. Uterine edema was assessed using a scoring system ranging from 1 to 3, as proposed by Samper (2010) [40].
Before insemination, intrauterine infusion and LVF, the perineal area and vulva were washed with warm water and scrubbed three times with povidone-iodine at 1% (Betadine®, Lombarda, Italy). LVF was performed using a sterile insemination cannula to introduce 120 ml of sterile Ringer’s Lactate into the uterus, which was then recovered in a sterile Falcon tube using 60 ml syringes. Macroscopic evaluation of the efflux clarity was performed immediately, scoring from 0 to 3 as described by LeBlanc (2011) [41], based on the amount of cells, mucus, or debris observed by rotating the tube while holding it up to the light. Samples were then transported to the laboratory at 4°C within 8 h.
2.5. Analysis of LVF
In the laboratory, a portion of the LVF was aseptically extracted for the bacteriological examination, while the remainder was centrifuged. Bacteriological investigation was performed at the Bacteriology Laboratory of the Department of Veterinary Medicine and Animal Production (University of Naples “Federico II”), inoculating each aliquot in Brain Heart Infusion (BHI) broth and incubated aerobically at 37°C for 24 hours [42]. After the overnight incubation, turbid broths were streaked on different agar plates for Gram-positive and Gram-negative bacteria isolation (Columbia CNA agar with 5% sheep blood, Mac Conkey agar, Mannitol salt agar), which were incubated for additional 24 hours at 37°C. The bacterial identification was performed by using matrix-assisted laser desorption/ionization–time of flight mass spectrometry (MALDI-TOF MS) (Bruker Daltonics Inc., Bremen, Germany), following the manufacturer’s guidelines.
After cold centrifugation of the LVF (1000 g for 20 minutes), the supernatant was cryopreserved at -80°C for evaluation within 2 months of the anti-inflammatory cytokine IL-10 concentration using a horse-specific ELISA kit (enzyme-linked immunosorbent assay) (Horse IL-10 ELISA kit, AssayGenie, Ireland). The supernatant was first pre-treated to expose the antigen binding site and the resulting sample, appropriately diluted, was plated in a 96-well plate supplied by the kit and incubated at 37°C. After 90 minutes, the Biotin-labeled antibody was incubated at 37°C for 1 h. Following 3 washes with Wash buffer, the HRP-Streptavidin Conjugate was added to each well and incubated at 37°C for 30 minutes. After 5 washes, TMB (5,5’-Tetramethylbenzidine) was added, and the plate was re-incubated at 37°C for approximately 10 minutes, until a blue colour developed. This colour, upon addition of the Stop solution, turned yellow, and the plate was immediately read with a spectrophotometer (Thermo Fisher Scientific, USA) at 450 nm. IL-10 concentration, expressed in pg/mL, was calculated using a calibration curve.
For cytology, the pellet from the centrifuged LVF was collected with a swab and smeared onto two slides. The slides were stained using Diff-Quick staining (Bio-Optica, Italy) and evaluated with an optical microscope (Nikon Eclipse E600, Italy). For each sample, 10 fields at 1000x magnification (high power fields: HPF) of both slides were assessed. Cells were categorized into endometrial epithelial cells, polymorphonuclear neutrophils (PMNs), and other inflammatory cells (eosinophils, lymphocytes, macrophages). Additionally, for each field, the presence of mucus, debris, and blood contamination (erythrocytes) was assessed, and the ratio of PMNs to total (epithelial) cells (PMN/EP) was calculated. Mares with 0-2 PMNs/HPF were classified as normal (0), with 3 to 5 PMNs were classified as having moderate inflammation (1), while mares with more than 5 PMNs/HPF were classified as having severe inflammation (2) [13,43].
2.6. Statistical Analysis
All results were collected in an Excel® (Microsoft) file and then imported into IBM SPSS version 29 (IBM Corporation, Milan, Italy) for statistical analysis. The data were expressed as mean and standard deviation (SD). Uterine edema, efflux clarity, and inflammation scores, PMN/HPF, PMN/EP and IL-10 concentrations were compared between cycles (CTR vs TRT) before and after (PRE vs POST) intrauterine infusion and within each cycle between PRE and POST using the Wilcoxon test. The frequencies of POST fluid accumulation and pregnancy in CTR and TRT cycles were compared with the Chi-square test or Fisher exact test for a small sample size. The binomial test was used to compare the expected with the observed frequency of positive pregnancies. Expected frequencies were calculated based on the literature [12,13,14], using 65% for normal mares and 22% for problem mares. Significance was set at P <0.05, and tendencies were ascribed for P values between 0.05 and 0.1.
3. Results
A total of 19 cycles were included in the study; in 8 cycles, mares were infused with unconditioned Ringer’s Lactate (CTR), while in 11 cycles, mares were treated with WJ-MSC-CM (TRT).
3.1. Clinical Finding
The uterine edema scores significantly decreased between PRE and POST in both CTR (PRE 3.0 ± 1.2 vs POST 2.0 ± 1.1, P<0.05) and TRT cycles (PRE 3.0 ± 0.8 vs POST 2.1 ± 0.9 P ≤ 0.01). Efflux clarity scores of LVF in CTR cycles were 2.7 ± 0.8 at PRE and 1.8 ± 1.3 at POST, whereas in TRT cycles, they were 2.0 ± 0.8 at PRE and 1.0 ± 1.1 at POST. No significant differences were found between CTR and TRT at either timepoint. However, while no differences were observed between PRE and POST in CTR cycles, TRT cycles showed a tendency toward significance (P=0.06) for improved efflux clarity.
Endometrial inflammation scores evaluated by cytology were PRE 1.0 ± 1.1 vs POST 1.3 ± 0.8 in CTR and PRE 1.5 ± 0.8 vs POST 1 ± 0.9 in TRT, showing no differences between untreated and treated cycles in both PRE and POST time points. However, as shown in Figure 1, the number of PMN/HPF and the PMN/EP ratio significantly decreased (P<0.05) between PRE and POST only in TRT cycles.
Uterine fluid was detected in POST evaluations in 12.5 % (1/8) of CTR cycles and 18% (2/11) of TRT cycles, with no significant differences between groups.
3.2. Bacterial Isolation and Identification
Table 1 presents the bacteriology results of each mare in both control and treated cycles at PRE and POST and the pregnancy outcomes. For all isolated bacteria, MALDI-TOF MS identification gave a log(score) ≥ 2.0, confirming a highly reliable species identification. In control cycles, LVF culture PRE was positive in all cases, and remained positive in POST, with the same isolated bacteria; except in one case where two different bacterial species were isolated between PRE (Deftia tsurunatensis) and POST (Escherichia coli). In particular, Escherichia coli (E. coli) was the most detected bacterium (6/8; 75%), being isolated in both PRE and POST samplings from five out of eight mares (62.5%) and recovered only once alone (ID 8) in POST. Among the five mares, E. coli was isolated as a single bacterial species in two cases (ID 2, 6), whereas in three cases (ID 1, 3, 7), it was identified together with Streptococcus equi subsp. zooepidemicus, Streptococcus equinus, and Enterococcus faecalis, respectively. Among the 11 treated cycles, two were negative in both PRE and POST analysis, seven tested positive for the same bacteria in both PRE and POST analyses, one case showed two different bacterial species isolated in PRE and POST, and two cycles were positive only in POST. Similarly, in treated cycles, E. coli was the most frequently identified bacterium (7/11; 63.6%). In contrast, the other recovered bacterial species exhibited a high degree of variability in isolation. One out 8 cycles with positive bacteriology resulted in a pregnancy, while five out of seven TRT-positive cycles resulted in pregnancy.
3.3. IL-10 Concentrations
Figure 2 shows IL-10 concentrations in the LVFs of control and treated cycles PRE and POST intrauterine infusion of either unconditioned or conditioned Ringer’s Lactate. Comparing PRE and POST, IL-10 concentrations in LVF increased (P < 0.05) following intrauterine infusion in TRT cycles but not in CTR ones. Furthermore, at the POST time point, IL-10 concentrations in LVF were significantly higher (P < 0.05) in the treated cycles compared to the CTR cycles.
3.4. Pregnancy Rates
Pregnancy outcomes for each mare are presented in Table 1. A trend toward significance (P=0.08) was observed when comparing pregnancy frequencies between treated cycles (6/11, 54%) and control cycles (1/8, 14%). When comparing the observed pregnancy frequencies with those expected in normal mares, no differences were found for treated cycles, whereas a significant difference was observed for CTR (P<0.05). Moreover, the pregnancy frequency in CTR did not differ from the expected percentages in problem mares, while the frequency in TRT was significantly higher than that of problem mares (P<0.05).
4. Discussion
This study tested for the first time the therapeutic potential of intrauterine infusion of WJ-MSC-CM in the treatment of endometritis in mares.
MSCs and their derivatives have been recognized for their immunoregulatory properties, making them valuable in treating acute and chronic inflammations [34]. The properties of WJ-MSCs make them an alternative source of stem cells for regenerative medicine. Unlike embryonic stem cells, no ethical concerns are associated with their clinical application. Due to their embryonic nature, WJ-MCs exhibit higher expression of pluripotency markers such as NANOG, Oct 3/4, and SOX2 compared to adult MSCs [44,45]. Furthermore, WJ-MSCs demonstrate a higher proliferation rate, longer longevity, greater differentiation potential, immune privilege, and lower immunogenicity than adult MSCs [46]. It has been observed that MSCs from embryonic tissues, compared to those from bone marrow, enhance Treg cell activity and exhibit greater immunosuppressive effects [47,48]. The therapeutic effects of MSCs primarily occur through paracrine signaling of their secretome [49], which consists of soluble factors (growth factors, cytokines, chemokines, and enzymes) and extracellular vesicles (EVs) such as exosomes and microvesicles (MVs) containing lipids, proteins, RNA and DNA subtypes [50]. Its benefits include minimizing the risk of survival issues or complications caused by improper differentiation of the cells within the host tissue, all while preserving their therapeutic potential [51,52].
MSCs-based therapies have been proposed for treating endometritis due to their ability to modulate the inflammatory response induced by the presence of sperm, debris, and bacteria during mating/insemination. In this study, the pregnancy rate following WJ-MSC-CM (54%) treatment was comparable to that of normal mares as reported in the literature, even if the difference between control and treated cycles only showed a trend toward significance. Indeed, the pregnancy rate in CTR remained at levels consistent with those reported for problem mares [12,13,14]. Positive results on pregnancy seem to be linked to the type of MSCs or derivatives used. Indeed, MSCs and their derivatives from perinatal tissues (embryo, placenta, umbilical cord) seem to have greater immunomodulatory properties than MSCs derived from adult tissues [53,54]. The beneficial effects of MSCs and their derivatives on fertility may be attributed to their role in facilitating paracrine communication between the conceptus and endometrium during early implantation and embryo development [55]. In cases of endometritis, MSCs release growth factors that enhance endometrial cell proliferation and stimulate angiogenesis, ultimately restoring fertility [56,57]. Prior studies have demonstrated that intrauterine administration of extracellular vesicles from amniotic MSCs over multiple cycles before-insemination in mares with chronic endometritis restored maternal-fetal communication and resulted in pregnancy [55]. Similarly, intrauterine therapy with MSCs of embryonic origin or their extracellular vesicles in mares susceptible to PBIE has been associated with higher pregnancy rates [53].
The primary mechanism of action of WJ-MSC-CM observed in this study was an increase in IL-10 concentrations post-treatment, which was not detected in CTR. This is noteworthy because IL-10 is an essential anti-inflammatory cytokine for the immune response, capable of reducing the transcription of pro-inflammatory cytokines by monocytes and macrophages [58]. Our findings align with the literature, as MSCs from bone marrow, adipose tissue, and amniotic origin have been demonstrated to induce IL-10 production both in vivo and in vitro [59,60,61]. MSCs administration has been demonstrated to reduce IL-6 concentration around 6 h post-insemination while increasing the levels of the anti-inflammatory cytokine IL-10 and IL-1RA in the uterus of susceptible mares at both 6 and 24 post-insemination [59]. As in our study, in a study by Lange-Consiglio et al. (2023) [60], the IL-10 concentration in the intrauterine fluid of the control group remained constant after insemination; on the contrary, the insemination with semen supplemented with extracellular vesicles from amniotic MSCs, IL-10 levels significantly increased 6 h post-insemination and remained unchanged up to 24 h. The increase of IL-10 and the decrease of IL-6 following MSC treatments confirm their mechanism of action: they can regulate the immune response via paracrine communication. Despite the different timings, the effect of MSC derivatives consistently appears to increase in IL-10 after insemination. Woodward et al. (2013) [11] demonstrated that 6 hours after insemination, susceptible mares have a reduced expression of the anti-inflammatory cytokines IL-10 and IL-1RA (IL-1RN), which are necessary to resolve inflammation. The ability of intrauterine WJ-MSC-CM infusion post-insemination to increase IL-10 concentrations could help address this deficiency in problem mares.
Nearly all mares in this study exhibited moderate to severe endometrial inflammation on cytology at the time of the intrauterine infusion. The increase in IL-10 in treated cycles was associated with a reduction in PMN infiltration and PMN/EP ratio, along with a trend toward improved LVF efflux clarity. However, there was no significant reduction in inflammation score.
The lack of significant change in some inflammatory parameters may be attributed to the small sample size and the timing of treatment and sample collection. Previous studies have observed reductions in inflammatory markers when MSCs or their derivatives were administrated before insemination and evaluated 24 h post-insemination (i.e. 48 h after treatment). For instance, a decrease in intrauterine fluid accumulation was observed 24 h after insemination in mares treated with adipose-derived MSCs or extracellular vesicles derived from amniotic MSCs [53,61]. Ferris et al. (2014) [59] observed a reduction of PMNs and the cytokine IL-1 in the LVF of mares treated with MSCs 24 h before insemination. Positive outcomes on inflammation and fertility have also been observed following the addition of MSCs or conditioned medium directly into semen [60,61]. MSCs and their derivatives had no negative effects on semen and instead led to a reduction of PMNs in the endometrium, intrauterine fluid accumulation, and pro-inflammatory cytokines 24 h after insemination.
Previous reports have indicated that intrauterine treatment of susceptible mares with MSCs or derivatives before or with insemination plays a positive role in immunomodulation, preventing excessive inflammatory responses and improving fertility [53,59,60,61]. It must be noted that MSCs and derivatives from different tissue types (adipose, bone marrow, placental) have been used in those studies, and a direct comparison of the results is not possible. However, our results represent the first step in supporting the hypothesis that mares can also be treated 6-8 hours after insemination. To likely demonstrate a significant reduction in inflammation score, the LVF for cytology should be conducted at least 48 hours after treatment.
The endometrial bacteria isolated in this study align with those commonly found in equine post-insemination endometritis: streptococci, coliforms, Pseudomonas Aeruginosa, and Staphylococcus aureus [18,62,63,64,65]. Among these, Streptococcus equi subspecies zooepidemicus and E. coli are the primary pathogens associated with endometritis [42,66,67,68,69] and, overall, E. coli was the most frequently identified bacterium in this study, consistently appearing across various samples and treatment conditions. Although bacteria were still isolated after treatment, almost all mares with positive bacteriology that received WJ-MSC-CM became pregnant, suggesting a potential antimicrobial effect.
Several studies have demonstrated the antimicrobial properties of MSC both in vitro and in vivo [27,31,33]. MSCs secrete antimicrobial peptides (AMPs) that disrupt bacterial membranes, inhibit nucleic acid synthesis, or interact with bacterial receptors [28]. AMPs such as cathelicidins, defensins, lipocaines and hepcidins target bacteria, fungi, yeasts, and viruses [29]. MSCs also exhibit indirect antimicrobial mechanisms by coordinating pro- and anti-inflammatory elements of the immune system, increasing phagocytosis, and modulating cytokine secretion [32]. MSCs reduce the migration of pro-inflammatory cells and immunoregulatory factors, and express immunosuppressive molecules such as IL-17, which increases neutrophil phagocytosis, and indoleamine 2,3-dioxygenase (IDO) [29]. The latter works by degrading the essential amino acid tryptophan, necessary for pathogens’ proliferation, defeating bacterial replication and inducing a broad-spectrum antimicrobial activity [70]. Meisel et al. (2011) [31] evaluated the antimicrobial efficacy of MSCs in vitro, finding a broad-spectrum effect. Marx et al. (2020) [27] demonstrated that equine MSCs secretome is effective against bacteria in biofilms, including the antibiotic-resistant methicillin-resistant Staphylococcus aureus, via secretion of active proteases that destabilize biofilms by protein degradation, resulting in increased antibiotic effectiveness. The lack of negativization after WJ-MSC-CM infusion can be explained by the timing, as the WJ-MSC-CM may not have had sufficient time, 12 h, to reduce the bacterial load. In in vitro studies, a reduction in microbial load was observed after 16 and 24 h [27,31,71]. Cortes-Araya et al. (2018) [33] also demonstrated that in vitro, at least 16 h are required for the equine conditioned medium to reduce E. coli growth. The timing used in this study was dictated by field conditions, as it corresponded to the time of ovulation monitoring.
However, in three cases after WJ-MSC-CM intrauterine infusion a positive bacteriology or other bacteria was isolated. It is not possible to determine whether the presence of these bacteria was an expected outcome of the intrauterine infusion, as no change in microbial presence was observed in most cases. This suggests that the detected bacteria may not be directly associated with the infusion procedure.
5. Conclusions
In conclusion, post-insemination uterine infusion of WJ-MSC-CM in mares restores pregnancy rates to normal levels. The primary mechanism of action of WJ-MSC-CM is its anti-inflammatory effect achieved through increased IL-10 concentrations, a cytokine essential for modulating the inflammatory response. Additionally, although no direct antimicrobial effect was demonstrated, the successful pregnancy WJ-MSC-CM-treated cycles suggests a potential antimicrobial role. Further studies are needed to confirm these findings and establish an optimal treatment protocol.
Author Contributions
Conceptualization: C.D.P, B.M., E.I. and M.M.P.; methodology, C.D.P, B.M., E.I. and M.M.P; validation, B.M., and M.M.P.; formal analysis, C.D.P. and B.M.; investigation, F.P.N, C.L., V.L., and N.C.; resources, E.I., L.D.M., and S.D.; data curation, C.D.P. and E.A.; writing—original draft preparation, C.D.P. and E.A.; writing—review and editing, B.M. and M.M.P.; supervision, B.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Animal Welfare Committee of the University of Naples Federico II (OPBA) (protocol PG/2022/0025536 of March 08, 2022) and conducted in accordance with the European Code of Good Veterinary Practice (FVE), the European Directive EU/2010/63, and Italian regulations (D.lgs. 26–March 04, 2014).
Informed Consent Statement
Written informed consent for participation in this study was obtained from the owner before entering the study.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CM | Conditioned medium |
| CTR | Control group |
| DMEM | Dulbecco’s Minimum Essential Medium |
| DPBS | Dulbecco’s phosphate buffered saline |
| E. coli | Escherichia Coli |
| EP | Epithelial cells |
| HPF | High power field |
| IL-10 | Interleukin-10 |
| PMN | Polimorphonuclear neutrophils |
| POST | After treatment |
| PRE | Before treatment |
| TRT | Treated group |
| UC | Umbilical cord |
| WJ-MSC-CM | Wharton’s jelly mesenchymal stromal/stem cell-derived medium |
References
- Troedsson, M.H.T. Uterine Clearance and Resistance to Persistent Endometritis in the Mare. Theriogenology 1999, 52, 461–471. [CrossRef]
- Traub-Dargatz, J.L.; Salman, M.D.; Voss, J.L. Medical Problems of Adult Horses, as Ranked by Equine Practitioners. J Am Vet Med Assoc 1991, 198, 1745–1747. [CrossRef]
- Canisso, I.F.; Segabinazzi, L.G.T.M.; Fedorka, C.E. Persistent Breeding-Induced Endometritis in Mares - a Multifaceted Challenge: From Clinical Aspects to Immunopathogenesis and Pathobiology. Int J Mol Sci 2020, 21. [CrossRef]
- Alghamdi, A.S.; Foster, D.N.; Carlson, C.S.; Troedsson, M.H.T. Nitric Oxide Levels and Nitric Oxide Synthase Expression in Uterine Samples from Mares Susceptible and Resistant to Persistent Breeding-Induced Endometritis. American Journal of Reproductive Immunology 2005, 53, 230–237. [CrossRef]
- Katila, T. Onset and Duration of Uterine Inflammatory Response of Mares after Insemination with Fresh Semen. Biol Reprod 1995, 52, 515–517. [CrossRef]
- Troedsson, M.H.T.; Liu, I.K.M.; Crabo, B.G. Sperm Transport and Survival in the Mare. Theriogenology 1998, 49, 905–915. [CrossRef]
- Troedsson, M.H.T. Therapeutic Considerations for Mating-Induced Endometritis. Pferdeheilkunde Equine Medicine 1997, 13, 516–520. [CrossRef]
- Troedsson, M.H.; Liu, I.K. Uterine Clearance of Non-Antigenic Markers (51Cr) in Response to a Bacterial Challenge in Mares Potentially Susceptible and Resistant to Chronic Uterine Infections. J Reprod Fertil Suppl 1991, 44, 283–288.
- Carnevale, E.M.; Ramirez, R.J.; Squires, E.L.; Alvarenga, M.A.; Vanderwall, D.K.; McCue, P.M. Factors Affecting Pregnancy Rates and Early Embryonic Death after Equine Embryo Transfer. Theriogenology 2000, 54, 965–979. [CrossRef]
- Bucca, S.; Carli, A.; Buckley, T.; Dolci, G.; Fogarty, U. The Use of Dexamethasone Administered to Mares at Breeding Time in the Modulation of Persistent Mating Induced Endometritis. Theriogenology 2008, 70, 1093–1100. [CrossRef]
- Woodward, E.M.; Christoffersen, M.; Campos, J.; Betancourt, A.; Horohov, D.; Scoggin, K.E.; Squires, E.L.; Troedsson, M.H.T. Endometrial Inflammatory Markers of the Early Immune Response in Mares Susceptible or Resistant to Persistent Breeding-Induced Endometritis. Reproduction 2013, 145, 289–296. [CrossRef]
- Rota, A.; Furzi, C.; Panzani, D.; Camillo, F. Studies on Motility and Fertility of Cooled Stallion Spermatozoa. Reproduction in Domestic Animals 2004, 39, 103–109. [CrossRef]
- Riddle, W.T.; LeBlanc, M.M.; Stromberg, A.J. Relationships between Uterine Culture, Cytology and Pregnancy Rates in a Thoroughbred Practice. Theriogenology 2007, 68, 395–402. [CrossRef]
- Kareskoski, M.; Venhoranta, H.; Virtala, A.M.; Katila, T. Analysis of Factors Affecting the Pregnancy Rate of Mares after Inseminations with Cooled Transported Stallion Semen. Theriogenology 2019, 127, 7–14. [CrossRef]
- Freeman, D.A.; Weber, J.A.; Geary, R.T.; Woods, G.L. Time of Embryo Transport through the Mare Oviduct. Theriogenology 1991, 36, 823–830. [CrossRef]
- Christoffersen, M.; Woodward, E.M.; Bojesen, A.M.; Petersen, M.R.; Squires, E.L.; Lehn-Jensen, H.; Troedsson, M.H.T. Effect of Immunomodulatory Therapy on the Endometrial Inflammatory Response to Induced Infectious Endometritis in Susceptible Mares. Theriogenology 2012, 78, 991–1004. [CrossRef]
- Friso, A.M.; Segabinazzi, L.G.T.M.; Cyrino, M.; Correal, S.B.; Freitas-Dell’Aqua, C.P.; Teoro do Carmo, M.; Dell’Aqua, J.A.; Miró, J.; Papa, F.O.; Alvarenga, M.A. Periovulatory Administration of Firocoxib Did Not Alter Ovulation Rates and Mitigated Post-Breeding Inflammatory Response in Mares. Theriogenology 2019, 138, 24–30. [CrossRef]
- Leblanc, M.; Causey, R. Clinical and Subclinical Endometritis in the Mare: Both Threats to Fertility. Reproduction in Domestic Animals 2009, 44, 10–22. [CrossRef]
- Scoggin, C.F. Endometritis: Nontraditional Therapies. Veterinary Clinics of North America - Equine Practice 2016, 32, 499–511. [CrossRef]
- Troedsson, M.H.T.; Nielsen, J.M. Non-Antibiotic Treatment of Equine Endometritis. Pferdeheilkunde 2018, 34, 17–22. [CrossRef]
- Del Prete, C.; Montano, C.; Cocchia, N.; de Chiara, M.; Gasparrini, B.; Pasolini, M.P. Use of Regenerative Medicine in the Treatment of Endometritis in Mares: A Systematic Review and Meta-Analysis. Theriogenology 2024, 227, 9–20. [CrossRef]
- Caplan, A.I. What’s in a Name? Tissue Eng Part A 2010, 16, 2415–2417. [CrossRef]
- Caplan, A.I.; Correa, D. The MSC: An Injury Drugstore. Cell Stem Cell 2011, 9, 11–15. [CrossRef]
- Sorrell, J.M.; Baber, M.A.; Caplan, A.I. Influence of Adult Mesenchymal Stem Cells on In Vitro Vascular Formation. https://home.liebertpub.com/tea 2009, 15, 1751–1761. [CrossRef]
- Mambelli, L.I.; Mattos, R.C.; Winter, G.H.Z.; Madeiro, D.S.; Morais, B.P.; Malschitzky, E.; Miglino, M.A.; Kerkis, A.; Kerkis, I. Changes in Expression Pattern of Selected Endometrial Proteins Following Mesenchymal Stem Cells Infusion in Mares with Endometrosis. PLoS One 2014, 9, e97889. [CrossRef]
- Hass, R.; Kasper, C.; Böhm, S.; Jacobs, R. Different Populations and Sources of Human Mesenchymal Stem Cells (MSC): A Comparison of Adult and Neonatal Tissue-Derived MSC. Cell Communication and Signaling 2011, 9, 1–14. [CrossRef]
- Marx, C.; Gardner, S.; Harman, R.M.; Van de Walle, G.R. The Mesenchymal Stromal Cell Secretome Impairs Methicillin-Resistant Staphylococcus Aureus Biofilms via Cysteine Protease Activity in the Equine Model. Stem Cells Transl Med 2020, 9, 746–757. [CrossRef]
- Silva-Carvalho, A.É.; Cardoso, M.H.; Alencar-Silva, T.; Bogéa, G.M.R.; Carvalho, J.L.; Franco, O.L.; Saldanha-Araujo, F. Dissecting the Relationship between Antimicrobial Peptides and Mesenchymal Stem Cells. Pharmacol Ther 2022, 233, 108021. [CrossRef]
- Hosseiniyan Khatibi, S.M.; Kheyrolahzadeh, K.; Barzegari, A.; Rahbar Saadat, Y.; Zununi Vahed, S. Medicinal Signaling Cells: A Potential Antimicrobial Drug Store. J Cell Physiol 2020, 235, 7731–7746. [CrossRef]
- Cai, Y.; Li, J.; Jia, C.; He, Y.; Deng, C. Therapeutic Applications of Adipose Cell-Free Derivatives: A Review. Stem Cell Res Ther 2020, 11, 1–16. [CrossRef]
- Meisel, R.; Brockers, S.; Heseler, K.; Degistirici; Bülle, H.; Woite, C.; Stuhlsatz, S.; Schwippert, W.; Jäger, M.; Sorg, R.; et al. Human but Not Murine Multipotent Mesenchymal Stromal Cells Exhibit Broad-Spectrum Antimicrobial Effector Function Mediated by Indoleamine 2,3-Dioxygenase. Leukemia 2011 25:4 2011, 25, 648–654. [CrossRef]
- Alcayaga-Miranda, F.; Cuenca, J.; Khoury, M. Antimicrobial Activity of Mesenchymal Stem Cells: Current Status and New Perspectives of Antimicrobial Peptide-Based Therapies. Front Immunol 2017, 8, 248157. [CrossRef]
- Cortés-Araya, Y.; Amilon, K.; Rink, B.E.; Black, G.; Lisowski, Z.; Donadeu, F.X.; Esteves, C.L. Comparison of Antibacterial and Immunological Properties of Mesenchymal Stem/Stromal Cells from Equine Bone Marrow, Endometrium, and Adipose Tissue. Stem Cells Dev 2018, 27, 1518–1525. [CrossRef]
- Pawitan, J.A. Prospect of Stem Cell Conditioned Medium in Regenerative Medicine. Biomed Res Int 2014, 2014, 965849. [CrossRef]
- Kim, H.O.; Choi, S.M.; Kim, H.S. Mesenchymal Stem Cell-Derived Secretome and Microvesicles as a Cell-Free Therapeutics for Neurodegenerative Disorders. Tissue Eng Regen Med 2013, 10, 93–101. [CrossRef]
- Harrell, C.R.; Jankovic, M.G.; Fellabaum, C.; Volarevic, A.; Djonov, V.; Arsenijevic, A.; Volarevic, V. Molecular Mechanisms Responsible for Anti-Inflammatory and Immunosuppressive Effects of Mesenchymal Stem Cell-Derived Factors. Adv Exp Med Biol 2019, 1084, 187–206. [CrossRef]
- Lanci, A.; Merlo, B.; Grandis, A.; Mariella, J.; Castagnetti, C.; Iacono, E. Gross and Histological Examination of Wharton’s Jelly in the Equine Umbilical Cord. Theriogenology 2023, 209, 184–192. [CrossRef]
- Iacono, E.; Brunori, L.; Pirrone, A.; Pagliaro, P.P.; Ricci, F.; Tazzari, P.L.; Merlo, B. Isolation, Characterization and Differentiation of Mesenchymal Stem Cells from Amniotic Fluid, Umbilical Cord Blood and Wharton’s Jelly in the Horse. Reproduction 2012, 143, 455–468. [CrossRef]
- Merlo, B.; Pirondi, S.; Iacono, E.; Rossi, B.; Ricci, F.; Mari, G. Viability, in Vitro Differentiation and Molecular Characterization of Equine Adipose Tissue-Derived Mesenchymal Stem Cells Cryopreserved in Serum and Serum-Free Medium.
- Samper, J.C. A Review of a Practitioner’s Perspective on Endometrial Edema. Pferdeheilkunde 2010, 26, 14–18. [CrossRef]
- Leblanc, M.M.; Act, D. How to Perform and Interpret Findings From a Low-Volume Uterine Flush. 2011.
- Nocera, F.P.; D’eletto, E.; Ambrosio, M.; Fiorito, F.; De Martino, L.; De Martino, L. Occurrence and Antimicrobial Susceptibility Profiles of Streptococcus Equi Subsp. Zooepidemicus Strains Isolated from Mares with Fertility Problems. Antibiotics (Basel) 2021, 11. [CrossRef]
- Cocchia, N.; Paciello, O.; Auletta, L.; Uccello, V.; Silvestro, L.; Mallardo, K.; Paraggio, G.; Pasolini, M.P. Comparison of the Cytobrush, Cottonswab, and Low-Volume Uterine Flush Techniques to Evaluate Endometrial Cytology for Diagnosing Endometritis in Chronically Infertile Mares. Theriogenology 2012, 77, 89–98. [CrossRef]
- Nekanti, U.; Mohanty, L.; Venugopal, P.; Balasubramanian, S.; Totey, S.; Ta, M. Optimization and Scale-up of Wharton’s Jelly-Derived Mesenchymal Stem Cells for Clinical Applications. Stem Cell Res 2010, 5, 244–254. [CrossRef]
- Higuchi, O.; Okabe, M.; Yoshida, T.; Fathy, M.; Saito, S.; Miyawaki, T.; Nikaido, T. Stemness of Human Wharton’s Jelly Mesenchymal Cells Is Maintained by Floating Cultivation. Cell Reprogram 2012, 14, 448–455. [CrossRef]
- Kim, D.W.; Staples, M.; Shinozuka, K.; Pantcheva, P.; Kang, S.D.; Borlongan, C. V. Wharton’s Jelly-Derived Mesenchymal Stem Cells: Phenotypic Characterization and Optimizing Their Therapeutic Potential for Clinical Applications. International Journal of Molecular Sciences 2013, Vol. 14, Pages 11692-11712 2013, 14, 11692–11712. [CrossRef]
- Mareschi, K.; Castiglia, S.; Sanavio, F.; Rustichelli, D.; Muraro, M.; Defedele, D.; Bergallo, M.; Fagioli, F. Immunoregulatory Effects on T Lymphocytes by Human Mesenchymal Stromal Cells Isolated from Bone Marrow, Amniotic Fluid, and Placenta. Exp Hematol 2016, 44, 138-150.e1. [CrossRef]
- Bárcia, R.N.; Santos, J.M.; Filipe, M.; Teixeira, M.; Martins, J.P.; Almeida, J.; Água-Doce, A.; Almeida, S.C.P.; Varela, A.; Pohl, S.; et al. What Makes Umbilical Cord Tissue-Derived Mesenchymal Stromal Cells Superior Immunomodulators When Compared to Bone Marrow Derived Mesenchymal Stromal Cells? Stem Cells Int 2015, 2015, 583984. [CrossRef]
- Fong, C.Y.; Gauthaman, K.; Cheyyatraivendran, S.; Lin, H.D.; Biswas, A.; Bongso, A. Human Umbilical Cord Wharton’s Jelly Stem Cells and Its Conditioned Medium Support Hematopoietic Stem Cell Expansion Ex Vivo. J Cell Biochem 2012, 113, 658–668. [CrossRef]
- Daneshmandi, L.; Shah, S.; Jafari, T.; Bhattacharjee, M.; Momah, D.; Saveh-Shemshaki, N.; Lo, K.W.H.; Laurencin, C.T. Emergence of the Stem Cell Secretome in Regenerative Engineering. Trends Biotechnol 2020, 38, 1373–1384. [CrossRef]
- Chronopoulos, A.; Kalluri, R. Emerging Role of Bacterial Extracellular Vesicles in Cancer. Oncogene 2020 39:46 2020, 39, 6951–6960. [CrossRef]
- Kalluri, R.; LeBleu, V.S. The Biology , Function , and Biomedical Applications of Exosomes. Science 2020, 367. [CrossRef]
- Hollinshead, F.K.; Hanlon, D.W.; Hou, W.; Tasma, Z.; Damani, T.; Bouma, G.J.; Murtazina, D.A.; Chamley, L. Use of Equine Embryo -Derived Mesenchymal Stromal Cells and Their Extracellular Vesicles as a Treatment for Persistent Breeding-Induced Endometritis in Susceptible Mares. J Equine Vet Sci 2024, 139, 105079. [CrossRef]
- Laroye, C.; Boufenzer, A.; Jolly, L.; Cunat, L.; Alauzet, C.; Merlin, J.L.; Yguel, C.; Bensoussan, D.; Reppel, L.; Gibot, S. Bone Marrow vs Wharton’s Jelly Mesenchymal Stem Cells in Experimental Sepsis: A Comparative Study. Stem Cell Res Ther 2019, 10. [CrossRef]
- Lange-Consiglio, A.; Funghi, F.; Cantile, C.; Idda, A.; Cremonesi, F.; Riccaboni, P. Case Report: Use of Amniotic Microvesicles for Regenerative Medicine Treatment of a Mare With Chronic Endometritis. Front Vet Sci 2020, 7, 529611. [CrossRef]
- Zhang, S.; Chuah, S.J.; Lai, R.C.; Hui, J.H.P.; Lim, S.K.; Toh, W.S. MSC Exosomes Mediate Cartilage Repair by Enhancing Proliferation, Attenuating Apoptosis and Modulating Immune Reactivity. Biomaterials 2018, 156, 16–27. [CrossRef]
- Abdelnaby, E.A.; Abdallah, A.N.; Anwar, I.M.; El-Tookhy, O.S.; Shamaa, A.A. The Therapeutic Effect of Stem Cell-Derived Exosomes in the Treatment of Chronic Endometritis as Assessed by Histopathological, Doppler and Hormonal Expression in Arabian Mares. Equine Vet Educ 2024, 36, 347–356. [CrossRef]
- Cyktor, J.C.; Turner, J. Interleukin-10 and Immunity against Prokaryotic and Eukaryotic Intracellular Pathogens. Infect Immun 2011, 79, 2964–2973. [CrossRef]
- Ferris, R.A.; Frisbie, D.D.; McCue, P.M. Use of Mesenchymal Stem Cells or Autologous Conditioned Serum to Modulate the Inflammatory Response to Spermatozoa in Mares. Theriogenology 2014, 82, 36–42. [CrossRef]
- Lange-Consiglio, A.; Gaspari, G.; Funghi, F.; Capra, E.; Cretich, M.; Frigerio, R.; Bosi, G.; Cremonesi, F. Amniotic Mesenchymal-Derived Extracellular Vesicles and Their Role in the Prevention of Persistent Post-Breeding Induced Endometritis. Int J Mol Sci 2023, 24, 5166. [CrossRef]
- Tongu, E.A. de O.; Segabinazzi, L.G.T.M.; Alvarenga, M.L.; Monteiro, A.; Papa, F.O.; Alvarenga, M.A. Allogenic Mesenchymal Stem Cell-Conditioned Medium Does Not Affect Sperm Parameters and Mitigates Early Endometrial Inflammatory Responses in Mares. Theriogenology 2021, 169, 1–8. [CrossRef]
- Canisso, I.F.; Stewart, J.; Coutinho da Silva, M.A. Endometritis: Managing Persistent Post-Breeding Endometritis. Vet Clin North Am Equine Pract 2016, 32, 465–480. [CrossRef]
- Walter, J.; Neuberg, K.P.; Failing, K.; Wehrend, A. Cytological Diagnosis of Endometritis in the Mare: Investigations of Sampling Techniques and Relation to Bacteriological Results. Anim Reprod Sci 2012, 132, 178–186. [CrossRef]
- Beltaire, K.A.; Cheong, S.H.; Coutinho da Silva, M.A. Retrospective Study on Equine Uterine Fungal Isolates and Antifungal Susceptibility Patterns (1999-2011). Equine Vet J Suppl 2012, 44, 84–87. [CrossRef]
- Del Prete, C.; Nocera, F.P.; Piegari, G.; Palumbo, V.; De Martino, L.; Cocchia, N.; Paciello, O.; Montano, C.; Pasolini, M.P. Use of Cytobrush for Bacteriological and Cytological Diagnosis of Endometritis in Mares. Vet World 2024, 17, 398. [CrossRef]
- Albihn, A.; Båverud, V.; Magnusson, U. Uterine Microbiology and Antimicrobial Susceptibility in Isolated Bacteria from Mares with Fertility Problems. Acta Vet Scand 2003, 44, 121–129. [CrossRef]
- Frontoso, R.; De Carlo, E.; Pasolini, M.P.; van der Meulen, K.; Pagnini, U.; Iovane, G.; De Martino, L. Retrospective Study of Bacterial Isolates and Their Antimicrobial Susceptibilities in Equine Uteri during Fertility Problems. Res Vet Sci 2008, 84, 1–6. [CrossRef]
- Nocera, F.P.; Capozzi, L.; Simone, D.; Pizzano, F.; Iovane, V.; Bianco, A.; Parisi, A.; De Martino, L. Multi-Locus Sequence Typing and in Vitro Antimicrobial Resistance of Equine Streptococcus Equi Subspecies Zooepidemicus Strains. Vet Res Commun 2024, 48, 215–224. [CrossRef]
- Nocera, F.P.; Ambrosio, M.; Conte, A.; Di Palma, T.; Castaldo, S.; Pasolini, M.P.; Fiorito, F.; De Martino, L. Importance of Broth-Enrichment Culture in Equine Endometritis Diagnosis. New Microbiol 2021, 44, 19–23.
- Däubener, W.; Schmidt, S.K.; Heseler, K.; Spekker, K.H.; MacKenzie, C.R. Antimicrobial and Immunoregulatory Effector Mechanisms in Human Endothelial Cells Indoleamine 2,3-Dioxygenase versus Inducible Nitric Oxide Synthase. Thromb Haemost 2009, 102, 1110–1116. [CrossRef]
- Marx, C.; Gardner, S.; Harman, R.M.; Wagner, B.; Van de Walle, G.R. Mesenchymal Stromal Cell-Secreted CCL2 Promotes Antibacterial Defense Mechanisms through Increased Antimicrobial Peptide Expression in Keratinocytes. Stem Cells Transl Med 2021, 10, 1666–1679. [CrossRef]
Figure 1.
PMN and PMN/EP in the cytology of low volume flush of cycles before (PRE) and after (POST) intrauterine infusion of either unconditioned Ringer’s Lactate (CTR=8) or Wharton’s jelly mesenchymal stromal/stem cell-derived conditioned Ringer’s Lactate (TRT=11). Asterisks indicate significant differences at P <0.05 (*) between groups within each time point or between time points within each group.
Figure 1.
PMN and PMN/EP in the cytology of low volume flush of cycles before (PRE) and after (POST) intrauterine infusion of either unconditioned Ringer’s Lactate (CTR=8) or Wharton’s jelly mesenchymal stromal/stem cell-derived conditioned Ringer’s Lactate (TRT=11). Asterisks indicate significant differences at P <0.05 (*) between groups within each time point or between time points within each group.
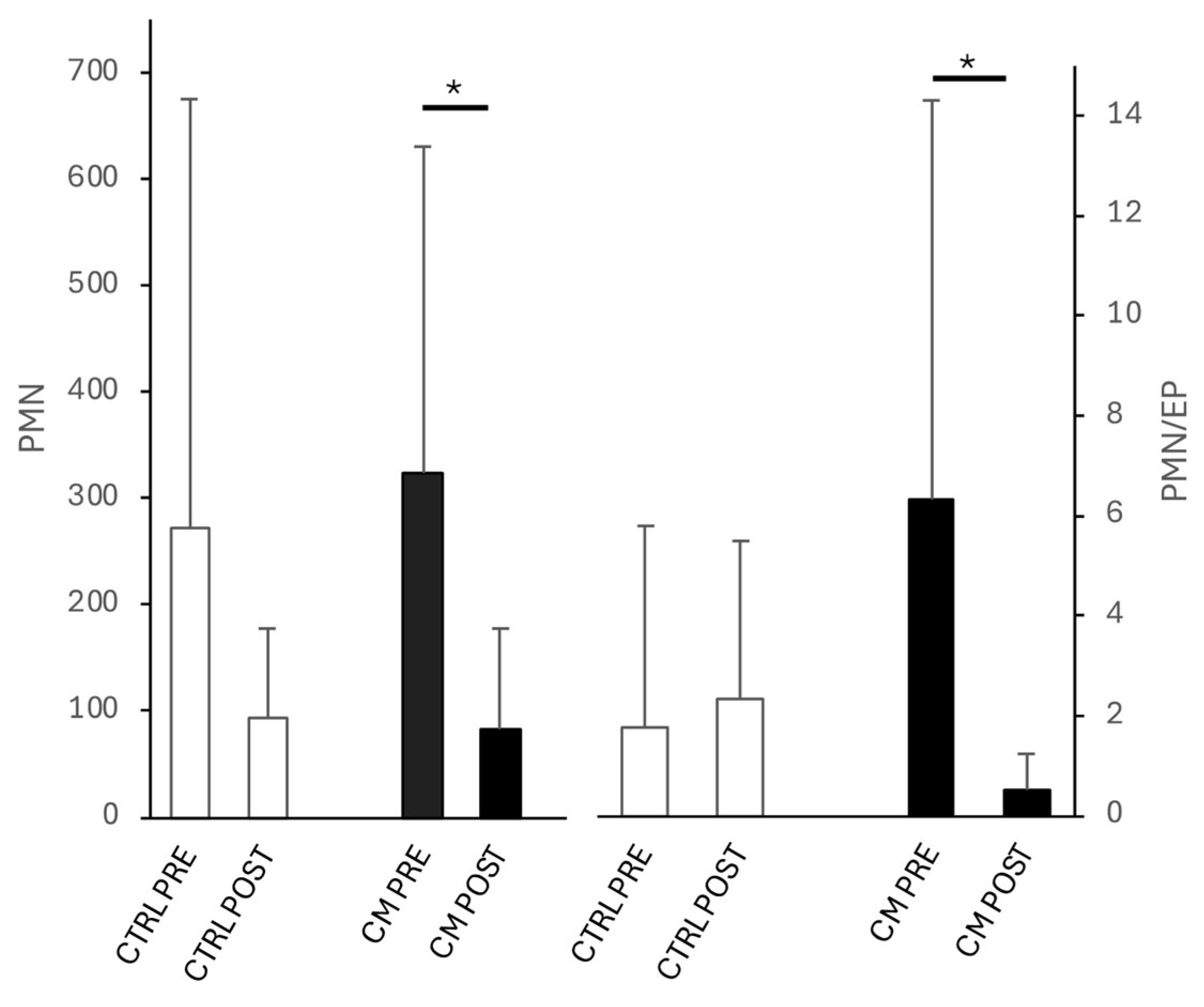
Figure 2.
IL-10 concentrations in low volume flush of cycles before (PRE) and after (POST) intrauterine infusion of either unconditioned Ringer’s Lactate (CTR; n= 7) or Wharton’s jelly mesenchymal stromal/stem cell-derived conditioned Ringer’s Lactate (TRT; n=9). Asterisks indicate significant differences at P≤0.05 (*) between groups within each time point or between time points within each group.
Figure 2.
IL-10 concentrations in low volume flush of cycles before (PRE) and after (POST) intrauterine infusion of either unconditioned Ringer’s Lactate (CTR; n= 7) or Wharton’s jelly mesenchymal stromal/stem cell-derived conditioned Ringer’s Lactate (TRT; n=9). Asterisks indicate significant differences at P≤0.05 (*) between groups within each time point or between time points within each group.
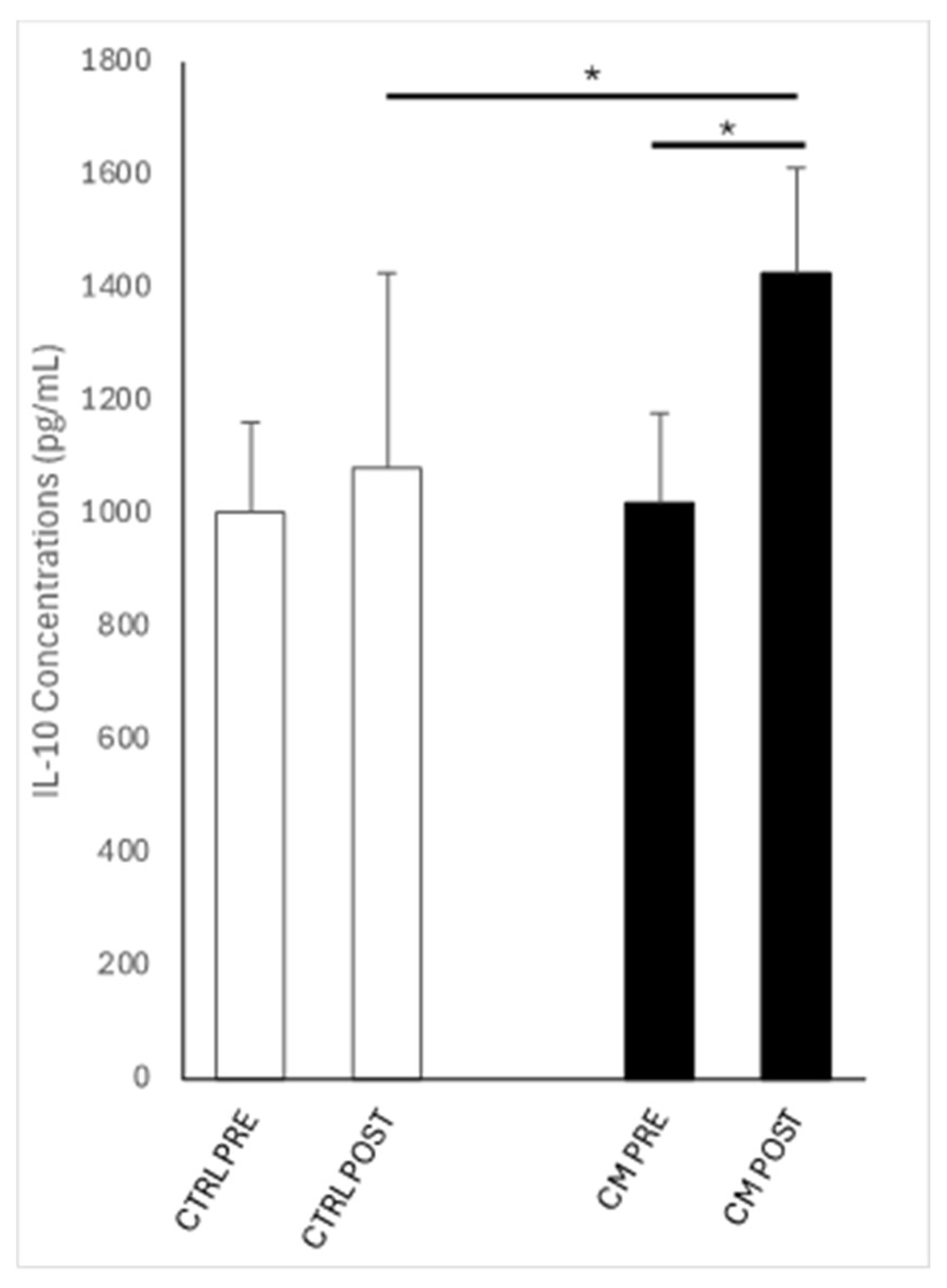

Table 1.
Bacteriology results of low volume flush of each mare in both control (CTR) and Wharton’s jelly mesenchymal stromal/stem cell-derived conditioned Ringer’s Lactate treated cycles (TRT) before (PRE) and after (POST) intrauterine infusion and pregnancy outcomes.
Table 1.
Bacteriology results of low volume flush of each mare in both control (CTR) and Wharton’s jelly mesenchymal stromal/stem cell-derived conditioned Ringer’s Lactate treated cycles (TRT) before (PRE) and after (POST) intrauterine infusion and pregnancy outcomes.
| ID | Cycle | PRE | POST | Pregnancy |
|---|---|---|---|---|
| 1 | CTR |
Streptococcus equi subsp. zooepidemicus; Escherichia coli |
Streptococcus equi subsp. zooepidemicus; Escherichia coli | Y |
| 2 | Escherichia coli | Escherichia coli | N | |
| 3 | Streptococcus equinus; Escherichia coli | Streptococcus equinus; Escherichia coli | N | |
| 4 | Pseudomonas putida | Pseudomonas putida | N | |
| 5 | Staphylococcus aureus | Staphylococcus aureus | N | |
| 6 | Escherichia coli | Escherichia coli | N | |
| 7 | Enterococcus faecalis; Escherichia coli | Enterococcus faecalis; Escherichia coli | N | |
| 8 | Deftia tsurunatensis | Escherichia coli | N | |
| 1 | TRT |
Escherichia coli; Streptococcus equi subsp. zooepidemicus |
Escherichia coli; Streptococcus equi subsp. zooepidemicus |
N |
| 2 | Streptococcus equinus | Streptococcus equinus | Y | |
| 3 | Escherichia coli; Staphylococcus schleiferi | Escherichia coli; Staphylococcus schleiferi | Y | |
| 4 |
Escherichia coli; Enterococcus faecalis; Staphylococcus aureus |
Escherichia coli; Enterococcus faecalis; Staphylococcus aureus |
Y | |
| 5 | Staphylococcus aureus; Streptococcus dysgalactiae; Escherichia coli |
Staphylococcus aureus; Streptococcus dysgalactiae; Escherichia coli |
Y | |
| 6 |
Streptococcus dysgalactiae; Klebsiella pneumoniae |
Streptococcus dysgalactiae | N | |
| 7 | Pseudomonas spp. | Escherichia coli | Y | |
| 8 | - |
Escherichia coli; Streptococcus equi subsp zooepidemicus |
N | |
| 9 | - | Escherichia coli; | N | |
| 10 | - | - | Y | |
| 11 | - | - | N |
CTR: control cycle; TRT: treated cycle; Y: yes; N: no.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



