
Submitted:
09 February 2023
Posted:
15 February 2023
You are already at the latest version
Abstract
In this review, the effect of colostrum on immune system development in buffalo calves was examined. The neonatal period is critical for buffalo calves and is characterized by a high mortality rate (more than 40%); the intake of colostrum improves the immune system of calves, increasing their chances of survival. Mainly in intensive farms, the availability of good quality colostrum is necessary, thus, good quality colostrum is often stored to provide newborn calves which cannot be fed by their mothers. The buffalo breeding is constantly expanding in Italy, mainly thanks to the Mozzarella cheese production that represents an excellence of the “Made in Italy” and is exported worldwide. High calf mortality rates directly affects the profitability of the business. Compared with other species, researches on buffalo colostrum are scarce; nevertheless it is fundamental to improve colostrum characteristics and management to guarantee buffalo calves health and reduce their mortality rate.
Keywords:
colostrum
; buffalo
; colostrum management
; colostrum quality
1. Introduction
Buffalo newborns are more susceptible to infections than those of other animal species, thus, colostrum intake plays a decisive role [1]. Compared to cow [2], sheep [3] and goat [4,5], research on buffalo colostrum is limited, mainly regarding its quality. High calf mortality rates directly affect the profitability of farms, whether the herd is destined for meat or milk production. Proper feeding of colostrum to the newborn calf is crucial; several studies have shown that incorrect feeding and incorrect colostrum management do not allow the calf to form an efficient immunity. The National Database of the Zootechnical Registry statistics, from 2017 to 2022, show an average mortality rate 41.11% in the first months of life (0-6 months) of buffaloes [6].
1.1. Buffalo breeding
Buffalo breeding has high economic, cultural and historic importance in the world, mainly in developing or developed countries. In Italy, most of the farms are located in Campania region with 58.4% of herds and 66.6% of animals [7] mainly milked for the production of "Mozzarella di Bufala Campana DOP" [8]. This is an example of how quality and tradition represent identity, but also income and employment. The system along the certified supply chain generates a direct turnover of almost 600 million euros with a remarkable effect on the local economy. “Mozzarella di Bufala Campana DOP” is, after the “Grana Padano”, “Parmigiano Reggiano” and “Gorgonzola”, the fourth Italian DOP in terms of quantity, with an annual production of over 54,000 tons (2021) of which as much as 35% exported (France and Germany alone account for more than half of the quantities exported) [9]. In addition, in recent years the interest in buffalo meat is also increasing in Italy, due to its nutritional characteristics [10,11,12].
1.2. Buffalo physiology
Buffaloes, as all ruminant species, has the syndesmochorial placenta which prevents the passage of antibodies from the mother to the newborn, thus, calves are very vulnerable at birth being agammaglobulinemic or hypogammaglobulinemic with little to no transplacental passage of immunoglobulins (mainly IgG) [13]. In the first days of life, the immune system needs to be guaranteed by the absorption of colostral immunoglobulins, to ensure passive protection against infectious diseases (diarrhoea, respiratory diseases, etc.) and for the future health and growth of the calf [14]. During pregnancy, the passage of nutrients to the fetus occurs by the placentomes, consisting of cotyledons, portions of the placenta, and caruncles, portions of the uterine horns. Placentomes do not allow the passage of antibodies: this is the reason why the colostrum plays a very important role for the newborn [15].
Colostrogenic transformation starts many weeks prior to calving when immunoglobulins are transported from the serum to the mammary gland [16]. Late in the pregnancy, this process continues, but it ends abruptly when the calf is born. Mammary gland secretory epithelial receptors allow the transfer of immunoglobulins from maternal blood to the mammary gland under the influence of lactogenic hormones and regional regulatory factors [17]. Ruminant colostrogenesis begins in the last weeks of pregnancy and ends immediately post-calving [18]. The blood cells and other colostral components migrate to the mammary gland [19,20], where their phenotype and function are altered [21,22]. During colostrogenesis, the transfer of cellular and non-cellular components from the maternal circulation into colostrum occur in the mammary gland. The transfer of immunoglobulins (Ig) from maternal circulation to mammary secretions, begins 3–4 weeks before parturition under endocrine control [23]. It is essential that the newborn calf receives the correct amount of colostrum, a natural source of macro and micro-nutrients [24] and peptides with antimicrobial activity and growth factors [25]. The concentration of immunoglobulins represents the main factor to assess the quality of colostrum. The permeability of the gut decreases over the first 24 h after calving, so, the calf needs to take colostrum within 4-6 hours of age [26]. Squillacioti et al. [27] focused this issue on aquaporins, which are membrane channel protein that regulate water permeability in many tissues [28], presuming that their manifestation in the gut of buffalo calves is affected by a proper ingestion of colostrum after delivery.
2. Colostrum
2.1. Characteristics and composition
To favor healthy growth, newborn calves need nutrients like fat, proteins, vitamins, minerals, lactoferrin, immune cells, cytokines and immunoglobulins (Ig) [29]. The mean chemical composition of buffalo colostrum is reported in Table 1.
Calves assimilate colostrum nutrients during the first hours of life [31]. Changes in colostrum composition depend on many factors, such as the time passed since calving, parity (i.e., the primiparae have different immune contents), age of the animal, dry period feeding and length of the dry period of cows [32]. There are differences in management and nutrient concentrations between small and large farms [33], and there are also differences in the composition of colostrum between geographic zones (high and low altitude) [34]. Colostrum contains 30-200 g/L proteins [35] mainly represented by antibodies [36,37]. Fat is the most variable constituent [38]. El-Fattah et al. [39] showed that the concentration of total protein and whey protein, composed of β-lactoglobulin, α-lactalbumin, proteose peptone, serum albumin, and immunoglobulins in that sequence of abundance [40] did not differ between buffalo and cow colostrum, while fat and total solids were significantly higher in buffalo colostrum. Another factor affecting colostrum composition is the breeding season of buffalo [41], mainly concerning fat content, fatty acids profile and cholesterol [42]. The total protein and Ig content in the first colostrum is higher in buffalo cows than in dairy crossbred cows [43].
The high protein content is mainly due to the large amount of immunoglobulin [44]; through blood analysis, bioactive compounds in colostrum can be directly observable, as is the case for immunoglobulin G1 [45], growth hormone (GH), prolactin (PRL), insulin and glucagon [46]. Colostrum contains insulin-like growth factors (IGF-1 and IGF-2) in high quantities and can improve the development and function of the gastrointestinal tract of newborn calves, and stimulate tissue, body growth and development in newborn calves [47,48].
Furthermore, colostrum also has an important immunoregulating function and modifies the gut microflora [49,50]. Antigenic glycopeptides [51] and macrophage-activating oligosaccharides [52], similar to those found in bovine and human colostrum, were identified in buffalo colostrum [53]. Ashok et al. [54] focused on whey proteins from buffalo colostrum and analyzed the effect of peptides derived from peptic-digestion on DNP-induced oxidative stress on blood components. It may be inferred that the combination of empirical and bioinformatic approaches facilitated the unravelling of various functionalities of early milk peptides that contribute to protect and improve neonatal health during the early part of its growth and development [54].
The minerals in ruminant colostrum (Ca, Mg, P and Na) decreased gradually from the fifth day after delivery [55]. The intensive mineral requirement of the newborn calf may account for the high level of ash content in the colostrum [56]. Colostrum also has a thermoregulation function to the survival of neonatal ruminants [57]. Colostrum energy content may affect the thermoregulation and fatty acid oxidation necessary to sustain gluconeogenesis [58]. Newborn calves, fed on higher volumes of colostrum, exhibited increased thermoregulatory responses, improved growth performance, and immunity [59]. Another function of colostrum is the laxative one, as it allows the calf to start forming its immunity by contributing to the proper expulsion of meconium [60].
2.2. Colostrum quality
Differences among species influence the quality of colostrum which also depends by parity, animal health and farm management. The first 24–48 h after calving are representative of the colostrum quality in terms of IgG concentration [61]. After 24 hours, the gut no longer absorbs immunoglobulins, which remain active in the intestinal lumen acting as a local defense mechanism against agents that may have been ingested orally. To determine the colostrum quality, the parameters of colostrum (i.e., crude protein, total protein, gamma-glutamyltransferase) might be used. Their activities could be used to determine the passive immunity transfer of calves [62]. The enzyme gamma-glutamyltransferase is a cell membrane-bound enzyme located on the outer surface of cell membranes. It transports and uses Glutathione (GSH), a tripeptide with antioxidant action, as a source of amino acids for cell metabolism [63]. Results of Pero et al. [64], showed that glutathione is secreted in buffalo colostrum and that the enzyme GGT uses it as a substrate for its activity.
2.3. Colostrum analysis
The bovine colostrum quality can be estimated by the colostrometer which measures the specific gravity related to the total gamma-globulin concentration and enables an estimation of quantity on the basis of a statistical correlation. In the buffalo, a non-significant correlation from the specific gravity and total gamma-globulin was seen [65]. The reason lies in the different colostrum composition between cow and buffalo colostrum, with higher concentrations of fat and proteins in buffalo colostrum that affect the colostrometer readings. Within all parameters evaluated as possible markers for buffalo colostral quality, the enzyme GGT appeared with the highest correlation observed between its activity and the concentrations of gamma globulins [65].
Younger buffalo cows have lower immunoglobulin content in the colostrum than to older ones [66], the latter being exposed to different environmental conditions over time, which determined a resistance to pathogens. According to Pritchett et al. [67] animals at second lactation show significantly lower IgG concentration than those at the third lactation. The number of bio-active components in colostrum decreases after hours from birth. Colostrum has also been reported to have also anti-inflammatory [68], anticancer [69], antimicrobial [70], and nutraceutical [71] properties.
Compared to bovine colostrum, buffalo colostrum shows higher concentrations of lactose, ash, total solids, fat, vitamin E, phosphorus and IGF-1 and lower of vitamin A, Mg, K, Na, Zn, and lactoferrin [72]. Colostrum contains other bioactive compounds, such lactoferrin (LF), lysozyme, and growth factors [73] which will be discussed later.
Colostrum pasteurization used to prevent pathogens contaminations, could cause undesirable changes in nutritional, functional and physicochemical properties [74,75,76], mainly with temperatures above 60 °C, which leads to the denaturation of bioactive proteins [77,78]. Pasteurization of colostrum is affected either for long (at 63 °C for 30 min) or short period (at 72 °C for 15 s) [79,80,81,82]. To preserve its bioactivity and to improve the quality and shelf life of cow and buffalo colostrum, the combination of light pasteurization (at 57 °C for 30 min) and freeze-drying could be used. Moreover, buffalo colostrum shows higher bioactivity than that of bovine [73].
3. Passive immunity transfer (PIT)
The calf is axenic, it must acquire passive immunity via colostrum [38].
The timing of colostrum intake, the method and volume of colostrum administration, the presence of the mother, and respiratory acidosis of the newly born are linked to its absorption in calves [83]. Colostrum-transmitting antibodies can protect the calf for up to six weeks. During this period, in the environment, the animal contacts infectious agents, which gradually stimulate the development of its immune system [84]. If the calf does not receive immunoglobulins, it may have problems, and this can even cause a high mortality rate of calves on the farm [85,86]. The onset of immunocompetence (active immunity) occurs after three weeks of age, when the passive coverage of the antibodies taken up via colostrum begins to decrease [87].
Feeding colostrum at the right time is crucial for the calf’s health because the highest calf’s ability to absorb immunoglobulins from colostrum decreases as early as six hours after their birth [88]. During the dry period, two months before delivery, tight junctions present at the level of the mammary gland epithelium, are not yet perfectly developed so that the mammary gland epithelium is still loose [89]. This allows macro-molecules (soluble immune factors, anti- bodies, and blood cells) that are present in the mother’s blood to pass between the secreting cells and arrive directly in the alveolar lumen. This also occurs for antibodies during fetal life when the animal is preparing for birth. During that period, a direct passage of antibodies into the alveolar lumen can occur, since the newborn calf, during the first two days of life, has not yet fully developed the intestinal tight junctions, which allow the absorption of macromolecules through the intestinal wall [90,91]. With regard to the immunocompetence of calves, Barmaiya et al. [92] deduced that the amounts of IgM, IgG and IgA in serum samples from buffalo calves are lower than the amounts from cow calves.

Table 2.
Total Ig (globular proteins involved in the immune response) concentration in buffalo colostrum is 54.0 mg/ml in Water Buffalo colostrum [93].
Table 2.
Total Ig (globular proteins involved in the immune response) concentration in buffalo colostrum is 54.0 mg/ml in Water Buffalo colostrum [93].
| Immunoglobulin | % |
|---|---|
| IgG (%) | 86 |
| IgA (%) | 8 |
| IgM (%) | 6 |
Source: Matte et al. 1982 [93].
The absorption capacity of immunoglobulins in the buffalo calf can be determined by quantitatively assessing the concentrations in the blood plasma and its volume. Indeed, the absorption efficiency of immunoglobulins decreases over time [92], in particular, if administered 6 h after birth [94], about 66% are recorded in the plasma, after 12 h about 50% and so on [95]. Dang et al. 2007 [96] observed increased activity in colostrum samples from buffalo collected on day one after calving; that decreased significantly by day seven.
Table 3.
Total Immunoglobulin in colostrum of Murrah buffaloes after calving.
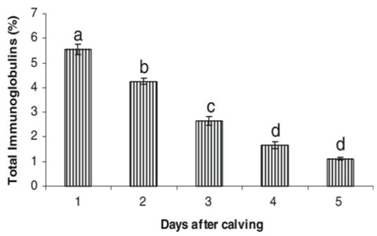 |
Source: Dang et al. 2008 [97].
In particular, IgG is a globular protein and has important effects against enteric and respiratory infectious diseases [98,99]. Interestingly, a recent study showed that buffalo colostrum has useful therapeutic proteins that can also be used in humans [100].
With regard to intestinal absorption of colostrum in calves, Pero et al. reported [101], that CaSR (Ca2+- sensing receptor) and Na+K+ATPase play a critical role identifying the intestinal characteristics that enable the absorption of the colostrum’s nutrients into the body.
Research has shown that colostrum can be improved by supplementing vitamins before birth. The impact of antioxidant vitamins on mineral and Ig secretion in colostrum was studied [102], showing that, thanks to some vitamin supplementation, the buffalo voluntarily takes in more feed before birth, so, there is an increase in weight before birth and, consequently, also the colostrum is of higher quality [103]. Moreover, Infascelli et al. [104] showed that integrating aloe into buffalo feed improves passive immune transfer in newborn buffalo calves, as it increases the immunological properties of the colostrum. Colostrum from mothers fed with aloe during their last months of pregnancy showed an increase in colostrum IgG content. Results indicated that supplementing aloe to the dry ration of mothers can improve colostrum immunological qualities, which in turn improves passive transfer in newborn calves. Furhermore, it has been observed that supplementation of vitamins A and E has a positive influence on the colostrum composition of Murrah buffaloes [105].
4. Evaluation and equipment
In the previous paragraphs, we described how colostrum quality characteristics play an essential role for calves’ health and survival. Therefore, it may be important to carry out an evaluation of colostrum before giving it to a newborn. The measurement of the IgG concentration makes it possible to evaluate the quality of the colostrum and to monitor colostrum feeding practices. The first assessment is visual: the colostrum appears yellow and very thick and creamy. The yellow colour is due to the presence of maternal antibodies [106].
Afterwards, assessment tools need to be used, such as refractometer or colostrometer, that show the content of IgG. The colostrometer measures specific gravity and, using a colour scale calibrated in milligrams per millilitre (mg/mL) of immunoglobulins (Ig), it converts specific gravity to Ig concentration. Generally, colostrum that tests "green" contains > 50 mg/mL of Ig, "yellow" contains 20 to 50 mg/mL, and "red" contains < 20 mg/mL of Ig. [107]. The use of the colostrometer is a commonly practiced procedure, but it suffers from poor accuracy and wide variability with relatively low correlation to the concentration of immunoglobulins. On the contrary, the refractometer is more accurate, and it presents a Brix scale. The refractive index of the serum estimates the protein concentration, which is an indication of the concentration of immunoglobulins [108]. The Brix value of 22% corresponds to 50 mg/mL, and with such value colostrum can be considered of high quality. A lot of studies explored the utility of the Brix refractometer for monitoring colostrum management programs and guaranteeing the passive immunity transfer (PIT) in bovines. The digital Brix refractometer seems to be an affordable tool for use in buffalo farms, allowing producers to use the same device for monitoring both colostrum quality and the success of passive transfer in calf serum. However, further studies evaluating a higher number of animals would be necessary to set specific cut point values [109], because there are studies that were carried out only to evaluate the buffalo’s colostrum quality, the thermal stability and some physico-chemical characteristics [110]. Brix is a good screening parameter for management purposes to see the colostrum quality because there is a correlation between refractive index and IgG concentration.
5. Colostrum management and the impact on calves
5.1. Colostrum Storage
It is common practice to refrigerate high quality colostrum to make it available when necessary. Colostrum should be collected to refill the colostrum bank with new supplies. It is advised to give colostrum right away after it has thawed. After freezing, the content of immunoglobulins doesn’t change, but it needs attention in the phase of defrosting [111]. High temperatures and incorrect agitation can compromise the composition. Colostrum pasteurization is not typically advised to decrease colostrum bacterial numbers as it is linked to immunoglobulin degradation and consistency issues [112]. The way colostrum is administered should be taken into account because it might affect the timing of the first feeding, the amount ingested, and the effectiveness of Ig absorption [113]. Colostrum should be kept chilled to prevent bacterial growth if it is not fed within two hours of collection. So, after evaluation and if it exceeds 3-4 liters, colostrum is stored in freezers, in thermal bags where the buffalo number and the correspondent brix can be identified.
The storage conditions affect the colostrum meaningfully. For example, bacteria and pH are significantly altered and changes are more rapid when colostrum is stored at > 4°C [114]. There are studies which sustain that colostrum can be kept in plastic containers at room temperature, but the shelf life is just three days due to deterioration and fermentation [115]. It has also been studied that the viability of cellular components and immunoglobulins at a temperature of 4°C resists up to a week [116]. Colostrum production rapidly ceases during parturition, and its composition drastically varies with time, making it necessary to collect colostrum from suitable donors as soon as possible [117].
5.2. Benefits of colostrum: infection prevention
Calves are extremely vulnerable and they are exposed to a high risk of infection. The intake of colostrum prevents contamination by viruses, parasites and bacteria. This is due to the presence of certain components, such as lysozyme, that provide the calf with many benefits. Lysozyme is an enzyme with antibacterial action produced by macrophages; its action focuses on the destruction of peptidoglycans that build the cell wall of gram-positive bacteria. The ingestion of colostrum, that contains high amounts of lysozyme, can contribute to the prevention of neonatal infections [118,119]. Enteropathogens are the main culprits causing calf diarrhoea during the animal’s first 30 days [120]. For example, rotaviruses are present in the environment, and they are very contagious. Adult animals are its main source of infection for young calves, especially between 1 and 3 weeks. The virus is fecally transmitted, and calves are often infected by contact with other calves primarily or secondarily through objects, feed, and water. In addition, rotaviruses have a wide host range, infecting many animal and human species [121,122]. Moreover, Toxocara Vitulorum, a parasitosis that mainly affects buffalo calves by localizing in the small intestine causes obstructive phenomena that can result in failure to absorb nutrients from the feed and subsequent decreased growth, diarrhoeic phenomena and, in some cases, death of the calf [123]. Trans-colostral and trans-milk transmission is also recorded for this species, in fact, in buffaloes the larvae become encysted at the muscle level becoming hypobiotic larvae and only shortly before parturition will reactivate by migrating into the mammary gland and colostrum [124]. Furthermore, regarding Escherichia coli, calves are most vulnerable to contract Enterotoxigenic Escherichia coli (ETEC) infection, which causes watery diarrhoea. Following ingestion, ETEC colonizes the gut by adhering to the enterocytes (intestinal epithelial cells) of the intestinal villi and infects the gut epithelium. Due to the low pH, the distal part of the small intestine offers the best conditions for ETEC colonization (less than 6.5), so it infected small intestine and causes secretory diarrhoea in calves [125].
6. Conclusions
Colostrum plays a fundamental role in the survival of buffalo calves. The purpose of this review was to analyze the benefits of passive immunity transfer provided to calves through colostrum administration and the benefits of the latter. Therefore, it appears critical to explore the managerial aspect in order to prevent risks to the newborn. Current studies aim to improve the quality of colostrum, in addition to the methods already used to strengthen the immune system of the calf, such as vaccination of mothers close to delivery, or, for example, the administration of supplement in pregnant buffaloes in order to improve the colostrum quality. It should not be underestimated that several buffalo pathogens are transmissible via colostrum, such as Rotavirus or Toxocara Vitulorum that is also a zoonosis therefore representing a risk for humans. Buffalo farming is constantly growing, in particular in southern Italy, which constitutes a relevant part of the economic landscape for the Mozzarella cheese production. Nevertheless, the studies about this topic are still scarce, and research in the buffalo field needs to be carried out in a more detailed way.
Author Contributions
Conceptualization D.L., E.P. and R.T.; validation F.I., P.L. and N.M.; investigation D.L and E.P.; data curation D.L., S.M. and N.M.; writing—original draft preparation D.L., E.P. and R.T.; writing—review and editing N.M. and P.I.; visualization D.L. and E.P; supervision P.L.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this review are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Singh, A., Ahuja, S. P., Singh, B. Individual variation in the composition of colostrum and absorption of colostral antibodies by the precolostral buffalo calf. J. Dairy Sci. 1993, 76(4), 1148–1156. [CrossRef]
- McGrath, B., Fox, P., McSweeney, P., Kelly, A. Composition and properties of Bovine Colostrum: A Review. J. Dairy Sci. Tech. 2015, 96(2), 133–158. [CrossRef]
- Agenbag, B., Swinbourne, A. M., Petrovski, K., van Wettere, W. H. E. J. Lambs need colostrum: A Review. Livestock Sci. 2021, 251, 104624. [CrossRef]
- Mehra, R., Sangwan, K., Garhwal, R. Composition and Therapeutic Applications of Goat Milk and Colostrum. J. Dairy Sci.Tech. 2021, 10.2, 1-7. [CrossRef]
- Zarrilli, A., Micera, E., Lacarpia, N., Lombardi, P., Pero, M. E., Pelagalli, A., d’Angelo, D., Mattia, M., Avallone, L. Evaluation of goat colostrum quality by determining enzyme activity levels. Livestock Prod. Sci. 2003, 83(2-3), 317–320. [CrossRef]
- Statistics BDN https://www.vetinfo.it/j6_statistiche/#/report-pbi/12.
- Statistics BDN 31/12/22 https://www.vetinfo.it/j6_statistiche/#/report-pbi/1.
- Reg. EC No. 1107 of 12 June 1996 registration of geographical indications and The Commission’s proposal for a Council Directive on the approximation of the laws, regulations and administrative provisions of the Member States relating to the classification of designations of origin within the framework of the procedure laid down in Article 17 of Regulation (EEC) No. Council Regulation. 2081/92 Official Journal No L 148, 21/06/1996 P. 0001-0010.
- Processing Nomisma on MDB Campania 2022 producers survey in collaboration with the Consortium of protection; provisional figure calculated on 80% of the certified volumes.
- Infascelli F., Gigli S., Campanile G. Buffalo meat production: Performance infra vitam and quality of meat. Vet Res Com. 2004, 28, 143–148. [CrossRef]
- Calabrò, S., Cutrignelli, M.I., Gonzalez, O.J., Chiofalo, B., Grossi, M., Tudisco, R., Panetta, C., Infascelli, F., Meat quality of buffalo young bulls fed faba bean as protein source. Meat Sci. 2014, 96(1), 591–596. [CrossRef]
- Cutrignelli, M. I., Calabrò, S., Tudısco, R., Chiofalo, B., Musco, N., Gonzalez, O. J., Grossi, M., Monastra, G., Infascelli, F., Conjugated linoleic acid and fatty acids profile in buffalo meat. Buffalo Bulletin 32(Special Issue 2), 2013, 1270–1273.
- Souza, D. C., Silva, D. G., Rocha, T. G., Monteiro, B. M., Pereira, G. T., Fiori, L. C., Viana, R. B., Fagliari, J. J. Serum biochemical profile of neonatal buffalo calves. Arq. Bras. Med. Vet. e Zootec. 2019, 71(1), 187–196. [CrossRef]
- Wooding, F. B. P., Morgan, G., Adam, C. L. Structure and function in the ruminant synepitheliochorial placenta: Central role of the trophoblast binucleate cell in deer. Microsc. Res. and Tech. 1997, 38(1-2), 88–99. [CrossRef]
- Wooding, F. B. P. The synepitheliochorial placenta of ruminants: Binucleate cell fusions and hormone production. Placenta 1992, 13(2), 101–113. [CrossRef]
- Barrington, G. M., McFadden, T. B., Huyler, M. T., Besser, T. E. Regulation of colostrogenesis in cattle. Livestock Prod. Sci. 2001, 70(1-2), 95–104. [CrossRef]
- Nguyen, D. N., Currie, A. J., Ren, S., Bering, S. B., Sangild, P. T. Heat treatment and irradiation reduce anti-bacterial and immune-modulatory properties of bovine colostrum. J. Funct. Foods 2019, 57, 182–189. [CrossRef]
- Brandon, M. R., Watson, D. L., Lascelles, A. K. The mechanism of transfer of immunoglobulin into mammary secretion of cows. Aust. J. Exp. Biol. Med. Sci. 1971, 49(6), 613–623. [CrossRef]
- Chernishov, V.P., Slukvin II. Mucosal immunity of the mammary gland and immunology of mother/newborn interrelation. Arch. Immunol. Ther. Exp. (Warsz) 1990, 38, 145–164.
- Norcross, N.L. Secretion and composition of colostrum and milk. J. Am. Vet. Med. Assoc. 1982, 181(10), 1057-1060.
- Reber, A. J., Lockwood, A., Hippen, A. R., Hurley, D. J. Colostrum induced phenotypic and trafficking changes in maternal mononuclear cells in a peripheral blood leukocyte model for study of leukocyte transfer to the neonatal calf. Vet. Immunol. Immunopathol. 2006, 109(1-2), 139–150. [CrossRef]
- Parmely, M.J., Reath D.B., Beer A.E., et al. Cellular immune responses of human milk T lymphocytes to certain environ- mental antigens. Transplant. Proc. 1977, 91477–1483.
- Watson, D. L., Lascelles, A. K. Seasonal changes of buffalo colostrum: physicochemical parameters, fatty acids and cholesterol variation. Aust. J. Exp. Biol. Med. Sci. 1971, 49(6), 613–623. [CrossRef]
- Anantakrishnan, C. P., Bhale Rao, V. R., Paul, T. M., Rangaswamy, M. C. The component fatty acids of buffalo colostrum fat. J. Bio. Chem. 1946, 166(1), 31–33. [CrossRef]
- Stelwagen, K., Carpenter, E., Haigh, B., Hodgkinson, A., Wheeler, T. T. Immune components of bovine colostrum and MILK1. J. Anim. Sci. 2009, 87(suppl_13), 3–9. [CrossRef]
- Weaver, D. M., Tyler, J. W., VanMetre, D. C., Hostetler, D. E., Barrington, G. M. Passive transfer of Colostral immunoglobulins in calves. J. Vet. Intern. Med. 2000, 14(6), 569–577. [CrossRef]
- Squillacioti, C., De Luca, A., Pero, M. E., Vassalotti, G., Lombardi, P., Avallone, L., Mirabella, N., Pelagalli, A. Effect of colostrum and milk on small intestine expression of AQP4 and AQP5 in newborn buffalo calves. Res. Vet. Sci. 2015, 103, 149–155. [CrossRef]
- Pelagalli, A., Squillacioti, C., De Luca, A., Pero, M. E., Vassalotti, G., Lombardi, P., Avallone, L., Mirabella, N. Expression and localization of aquaporin 4 and aquaporin 5 along the large intestine of colostrum-suckling buffalo calves. Anatom. Hist. Embr. 2015, 45(6), 418–427. [CrossRef]
- Zarcula S. ,Cernescu H. , Mircu C. ,Tulcan C. ,Morvay A., Baul S. , Popovici D. Influence of Breed, Parity and Food Intake on Chemical Composition of First Colostrum in Cow. Anim. Sci. and Biotech. 2010, 43(1).
- Coroian, A., Erler, S., Matea, C. T., Mireșan, V., Răducu, C., Bele, C., Coroian, C. O. Seasonal changes of buffalo colostrum: Physicochemical parameters, fatty acids and cholesterol variation. Chem. Central J. 2013, 7(1). [CrossRef]
- Roy. J. H. B., 1990. The calf. Butterworths. Wiley & Sons, Incorporated, John.
- Werner, Andrea. Experimentelle Untersuchungen zur Eignung der[gamma]-Glutamyltransferase-Aktivität im Blut von Kälbern zur Überprüfung der Kolostrumversorgung. Diss. Hannover, Tierärztl. Hochsch. 2003.
- Kehoe, S. I., Jayarao, B. M., Heinrichs, A. J. A survey of bovine colostrum composition and colostrum management practices on Pennsylvania Dairy Farms. J. Dairy Sci. 2007, 90(9), 4108–4116. [CrossRef]
- Mehra, R., Kumar, S., Verma, N., Kumar, N., Singh, R., Bhardwaj, A., Nayan, V., Kumar, H. Chemometric approaches to analyze the colostrum physicochemical and immunological (IGG) properties in the recently registered Himachali Pahari Cow Breed in India. LWT 2021, 145, 111256. [CrossRef]
- Korhonen, H., Pihlanto, A. Technological options for the production of health-promoting proteins and peptides derived from milk and colostrum. Curr. Pharm. Des. 2007, 13(8), 829–843. [CrossRef]
- Ashok, N. R., Aparna, H. S. Empirical and bioinformatic characterization of Buffalo (bubalus bubalis) colostrum whey peptides & their angiotensin I-converting enzyme inhibition. Food Chem. 2017, 228, 582–594. [CrossRef]
- Playford, R. J., Weiser, M. J. Bovine colostrum: Its constituents and uses. Nutrients 2021, 13(1), 265. [CrossRef]
- Parrish, D. B., Wise, G. H., Hughes, J. S., Atkeson, F. W. Properties of the colostrum of the dairy cow. V. Yield, specific gravity and concentrations of total solids and its various components of colostrum and early milk. J. Dairy Sci. 1950, 33(6), 457–465. [CrossRef]
- Abd El-Fattah, A. M., Abd Rabo, F. H. R., EL-Dieb, S. M., El-Kashef, H. A. Changes in composition of Colostrum of Egyptian Buffaloes and holstein cows. BMC Vet. Res. 2012, 8(1). [CrossRef]
- Kilara, A., Vaghela, M. N. Whey proteins. Prot. Food Proc. 2004, 72–99.
- Barile, V. L. Improving reproductive efficiency in female buffaloes. Livestock Prod. Sci. 2005, 92(3), 183–194. [CrossRef]
- Coroian, A., Erler, S., Matea, C. T., Mireșan, V., Răducu, C., Bele, C., Coroian, C. O. Seasonal changes of buffalo colostrum: Physicochemical parameters, fatty acids and cholesterol variation. Chem. Cent. J. 2013, 7(1). [CrossRef]
- Singh, A., Ahuja, S. P., Singh, B. Individual variation in the composition of colostrum and absorption of colostral antibodies by the precolostral buffalo calf. J. Dairy Sci. 1993, 76(4), 1148–1156. [CrossRef]
- Wang, J., He, Y., Pang, K., Zeng, Q., Zhang, X., Ren, F., Guo, H. Changes in milk yield and composition of colostrum and regular milk from four buffalo breeds in China during lactation. J. Sci. Food and Agr. 2019, 99(13), 5799–5807. [CrossRef]
- Barrington, G. M., Parish, S. M. Bovine neonatal immunology. Vet. Clin. North Am. Food Anim. Prac. 2001, 17(3), 463–476. [CrossRef]
- Blum, J. W. Nutritional physiology of neonatal calves*. J. Anim. Physiol. Anim. Nutr. (Berl.) 2006, 90(1-2), 1–11. [CrossRef]
- Penchev, G.I. Differences in chemical composition between cow colostrum and milk. Bulg. J. Vet. Med. 2008, No 1, 3−12.
- Blum, J. W., & Hammon, H. Colostrum effects on the gastrointestinal tract, and on nutritional, endocrine and metabolic parameters in neonatal calves. Livestock Prod. Sci. 2000, 66(2), 151–159. [CrossRef]
- Scammell A.W. Production and uses of colostrum. Aust. J. Dairy Technol. 2001, 56, 74.
- Golinelli L.P., Del Aguila E.M., Paschoalin V.M.F., Silva J.T., Conte-Junior C.A. Functional aspect of colostrum and whey proteins in human milk. J. Human Nutr. Food Sci. 2014, 2, 1035.
- Aparna H.S, Veeresh S. Purification of an Antigenic Glycopeptide from buffalo colostrum February. J. Food Sci. Technol. 2001, 38(5).
- Aparna, H. S., & Salimath, P. V. Galactose terminated oligosaccharides activate macrophage respiratory burst. Nutr. Res. 1994, 14(3), 433–444. [CrossRef]
- Chougule, R. A., Aparna, H. S. Characterization of β-lactoglobulin from Buffalo (bubalus bubalis) colostrum and its possible interaction with erythrocyte lipocalin-interacting membrane receptor. J. Biochem. 2011, 150(3), 279–288. [CrossRef]
- Ashok, N. R., Vivek, K. H., Aparna, H. S. Antioxidative role of Buffalo (bubalus bubalis) colostrum whey derived peptides during oxidative damage. Intern. J. Peptide Res. Therap. 2018, 25(4), 1501–1508. [CrossRef]
- Tsioulpas, A., Grandison, A. S., Lewis, M. J. Changes in physical properties of bovine milk from the colostrum period to early lactation. J. Dairy Sci. 2007, 90(11), 5012–5017. [CrossRef]
- Doreau, M. Le lait de jument et sa production: Particularités et facteurs de variation. Le Lait 1994, 74(6), 401–418. [CrossRef]
- Rowan, T. G. Thermoregulation in neonatal ruminants. BSAP Occasional Publication 1992, 15, 13–24. [CrossRef]
- Quigley, J. D., Drewry, J. J. Nutrient and immunity transfer from cow to calf pre- and post-calving. J. Dairy Sci. 1998, 81(10), 2779–2790.
- Silva, F. L., Miqueo, E., Silva, M. D., Torrezan, T. M., Rocha, N. B., Salles, M. S., Bittar, C. M. Thermoregulatory responses and performance of dairy calves fed different amounts of Colostrum. Animals 2021, 11(3), 703. [CrossRef]
- Das, L.K., Behera, S., Colostrum: A wonder nutrition for newborn calves of cattle and buffaloes. Indian Farmer 2015, 2(3), 165-170.
- Gay C.C. The Role of Colostrum in Managing Calf Health. Diary Split Session I 1983.
- Aydogdu, U., Guzelbektes, H. Effect of colostrum composition on passive calf immunity in primiparous and Multiparous Dairy Cows. Veterinární Medicína 2018, 63(No. 1), 1–11. [CrossRef]
- Owen, W.; Griffith, R. J.; Meister, A. Transport of gamma-glutamyl amino acids: role of glutathione and gamma-glutamyl transpeptidase. Proc. Natl. Acad. Sci. U S A. 1979, 76, 6319-6322.
- Pero, M. E., Pelagalli, A., Lombardi, P., Avallone, L. Short communication: Glutathione concentration and gamma-glutamyltransferase activity in Water Buffalo Colostrum. J. Anim. Phys Anim. Nutr. 2010, 94(5), 549–551. [CrossRef]
- Lombardi, P., Avallone, L., Pagnini, U., D’Angelo, D., Bogin, E. Evaluation of buffalo colostrum quality by estimation of enzyme activity levels. J. Food Prot. 2001, 64(8), 1265–1267. [CrossRef]
- Morrill, K. M., Conrad, E., Lago, A., Campbell, J., Quigley, J., Tyler, H. Nationwide evaluation of quality and composition of colostrum on dairy farms in the United States. J. Dairy Sci. 2012, 95(7), 3997–4005. [CrossRef]
- Pritchett, L. C., Gay, C. C., Besser, T. E., Hancock, D. D. Management and production factors influencing immunoglobulin G1 concentration in colostrum from Holstein Cows. J. Dairy Sci. 1991, 74(7), 2336–2341. [CrossRef]
- Støy, A. C., Heegaard, P. M. H., Thymann, T., Bjerre, M., Skovgaard, K., Boye, M., Stoll, B., Schmidt, M., Jensen, B. B., Sangild, P. T. Bovine colostrum improves intestinal function following formula-induced gut inflammation in preterm pigs. Clin. Nutr. 2014, 33(2), 322–329. [CrossRef]
- Agarwal, P., Gupta, R.A review on anticancer property of colostrum. Res. Rev.-J. Med. Heal. Sci 2016, 5, 1-9.
- JSM Biotechnol Bioeng 2016, 3(4), 1063.
- Ahmadi, M. Benefits of Bovine Colostrum in Nutraceutical Products. J. Agroalimentary Process. Technol. 2011, 17, 42–45.
- Abd El-Fattah, A. M., Abd Rabo, F. H. R., EL-Dieb, S. M., El-Kashef, H. A. Changes in composition of Colostrum of Egyptian Buffaloes and holstein cows. BMC Vet. Res. 2012, 8(1), 19. [CrossRef]
- Salar, S., Jafarian, S., Mortazavi, A., Nasiraie, L. R. Effect of hurdle technology of gentle pasteurisation and drying process on bioactive proteins, antioxidant activity and microbial quality of cow and buffalo colostrum. Intern. Dairy J. 2021, 121, 105138. [CrossRef]
- Borad, S. G., Singh, A. K.Colostrum immunoglobulins: Processing, preservation and application aspects. Intern. Dairy J. 2018, 85, 201–210. [CrossRef]
- Liu, Y., Zhang, W., Han, B., Zhang, L., Zhou, P. Changes in bioactive milk serum proteins during milk powder processing. Food Chem. 2020, 314, 126177. [CrossRef]
- Liu, Y., Zhang, W., Zhang, L., Hettinga, K., Zhou, P. Characterizing the changes of bovine milk serum proteins after simulated industrial processing. LWT 2020, 133, 110101. [CrossRef]
- Gelsinger, S. L., Jones, C. M., Heinrichs, A. J. Effect of colostrum heat treatment and bacterial population on immunoglobulin G absorption and health of neonatal calves. J. Dairy Sci. 2015, 98(7), 4640–4645. [CrossRef]
- Xiong, L., Li, C., Boeren, S., Vervoort, J., Hettinga, K. Effect of heat treatment on bacteriostatic activity and protein profile of bovine whey proteins. Food Res. Intern. 2020, 127, 108688. [CrossRef]
- Chatterton, D. E., Aagaard, S., Hesselballe Hansen, T., Nguyen, D. N., De Gobba, C., Lametsch, R., Sangild, P. T. Bioactive proteins in bovine colostrum and effects of heating, drying and irradiation. Food & Function 2020, 11(3), 2309–2327. [CrossRef]
- Donahue, M., Godden, S. M., Bey, R., Wells, S., Oakes, J. M., Sreevatsan, S., Stabel, J., Fetrow, J. Heat treatment of colostrum on commercial dairy farms decreases colostrum microbial counts while maintaining colostrum immunoglobulin G concentrations. J. Dairy Sci. 2012, 95(5), 2697–2702. [CrossRef]
- McMartin, S., Godden, S., Metzger, L., Feirtag, J., Bey, R., Stabel, J., Goyal, S., Fetrow, J., Wells, S., Chester-Jones, H. Heat treatment of bovine colostrum. I: Effects of temperature on viscosity and immunoglobulin G level. J. Dairy Sci. 2006, 89(6), 2110–2118. [CrossRef]
- McGuirk, S. M.,Collins, M. Managing the production, storage, and delivery of Colostrum. Vet. Clin. North Am. Food Anim. Prac. 2004, 20(3), 593–603. [CrossRef]
- Weaver, D. M., Tyler, J. W., VanMetre, D. C., Hostetler, D. E., Barrington, G. M. Passive transfer of Colostral immunoglobulins in calves. J. Vet. Internal Med. 2000, 14(6), 569–577. [CrossRef]
- Radostits, O,M., Blood D.C., Gay C.C. Veterinary medicine: A textbook of the diseases of cattle, sheep, pigs, goats and horses. J. Eq. Vet. Sci. 2000, 20(10), 625.
- McGuire T. C., Pfeiffer N. E., Weikel J. M., Bartsch R. C. Failure of colostral immunoglobulin transfer in calves dying from infectious disease. J. Am. Vet. Med. Assoc. 1976, 169(7), 713-8.
- Chase, C. C. L., Hurley, D. J., Reber, A. J. Neonatal immune development in the calf and its impact on vaccine response. Vet. Clin. North Am. Food Anim. Pract. 2008, 24(1), 87–104. [CrossRef]
- WittumT. E., PerinoL. J. Passive immune status at postpartum hour 24 and long-term health and performance of calves. “Am J Vet Res” Jour. 1995 56(9), 1149-1154. [CrossRef]
- Puppel, K., Gołębiewski, M., Grodkowski, G., Slósarz, J., Kunowska-Slósarz, M., Solarczyk, P., Łukasiewicz, M., Balcerak, M., Przysucha, T. Composition and factors affecting quality of Bovine Colostrum: A Review. Animals 2019, 9(12), 1070. [CrossRef]
- Stelwagen, K., Singh, K. The role of tight junctions in mammary gland function. J. Mammary Gland Biol. Neoplasia 2013, 19(1), 131–138. [CrossRef]
- Stelwagen, K., Carpenter, E., Haigh, B., Hodgkinson, A., Wheeler, T. T. Immune components of bovine colostrum and MILK1. J. Anim. Sci. 2009, 87(suppl_13), 3–9. [CrossRef]
- Tong, F., Wang, T., Gao, N. L., Liu, Z., Cui, K., Duan, Y., Wu, S., Luo, Y., Li, Z., Yang, C., Xu, Y., Lin, B., Yang, L., Pauciullo, A., Shi, D., Hua, G., Chen, W.-H., Liu, Q. The microbiome of the Buffalo Digestive Tract. Nat. Commun., 2022 13(1),823. [CrossRef]
- Barmaiya, S., Aditi Dixit, Mishra, A., Jain, A.K., Gupta, A., Paul, A., Quadri, A.M., Madan, A.M., Sharma I. J. Quantitation of serum immunoglobulins of neonatal buffalo calves and cow calves through elisa and page: status of immune-competence. Buffalo Bulletin 2009 28(2), 85-94.
- Matte, J. J., Girard, C. L., Seoane, J. R., Brisson, G. J. Absorption of colostral immunoglobulin g in the newborn dairy calf. J. Dairy Sci. 1982, 65(9), 1765–1770. [CrossRef]
- Burton, J. L., Kennedy, B. W., Burnside, E. B., Wilkie, B. N.,Burton, J. H. Variation in serum concentrations of Immunoglobulins G, a, and M in Canadian holstein-friesian calves. J. Dairy Sci. 1989, 72(1), 135–149. [CrossRef]
- Besser, T. E., Garmedia, A. E., McGuire, T. C., Gay, C. C. Effect of colostral immunoglobulin G1 and immunoglobulin M concentrations on immunoglobulin absorption in calves. J. Dairy Sci. 1985, 68(8), 2033–2037. [CrossRef]
- Dang, A.K., Kapila, S., Tomar, P., Singh, C. Immunity of the buffalo mammary gland during different physiological stages. Asian Australasian J Anim Sci 2007a, 20, 1174–1181. [CrossRef]
- Dang, A. K., Kapila, S., Purohit, M., Singh, C. Changes in colostrum of murrah buffaloes after calving. Trop. Anim. Health Prod. 2008, 41(7), 1213–1217. [CrossRef]
- Dang, A. K., Kapila, S., Purohit, M., Singh, C. Changes in colostrum of murrah buffaloes after calving. Trop. Anim. Health Prod. 2008, 41(7), 1213–1217. [CrossRef]
- Bulter, J.E. Bovine immunoglobulins: an augmented review. Vet. Immunol. Immunopath. 1983, 4, 43–152. [CrossRef]
- Gopal, P. K., Gill, H. S. Oligosaccharides and glycoconjugates in bovine milk and colostrum. British J. Nutr. 2000, 84(S1), 69–74. [CrossRef]
- Pero M.E., Luca A., Mastellone V., Mirabella N., Lombardi P., Tudisco R., Infascelli F., Avallone L., Romero F. Distribution of Ca2+-sensing receptor (CaSR) and Na-K-ATPase in the gastrointestinal tracts of neonatal calves after colostrum ingestion. Revista Veterinaria. 2010, 21 Issue 1, 620-622.
- Bhanu, L. S., Amano, M., Nishimura, S.-I., Aparna, H. S. Glycome characterization of immunoglobulin G from Buffalo (bubalus bubalis) colostrum. Glycoconj. J. 2015, 32(8), 625–634. [CrossRef]
- Sikka P. Lal D. Khanna S. Tomer A.K.S. Sethi R.K. Studies on colostrum minerals and their transfer to suckling calves in buffaloes- Ind. J. Anim. Sci. 2006, 76 Issue 6, 480 - 483.
- Infascelli, F., Tudisco, R., Mastellone, V., Cutrignelli, M., Lombardi, P., Calabrò, S., Gonzalez, O. J., Pelagalli, A., Grossi, M.,d’Angelo, D., Avallone, L. Diet Aloe Supplementation in Pregnant Buffalo Cows Improves Colostrum Immunoglobulin Content. Revista Veterinaria. 2010, 21 Issue 1, 151-153.
- Mudgal, V., Bharadwaj, A., Verma, A. K. Vitamins supplementation affecting colostrum composition in Murrah Buffaloes. Ind J An Res, 2021 (OF). 55(8),900-904. [CrossRef]
- Ojha, B. K., Dutta, N., Pattanaik, A. K., Singh, S. K., Narang, A. Effect of pre-partum strategic supplementation of concentrates on colostrum quality and performance of buffalo calves. Anim. Nutr. Feed Technol. 2015, 15(1), 41. [CrossRef]
- Qureshi, T. M., Yaseen, M., Nadeem, M., Murtaza, M. A., Munir, M. Physico–chemical composition and antioxidant potential of Buffalo Colostrum, transition milk, and mature milk. J. Food Proc. Pres. 2020, 44(10). [CrossRef]
- Heinrichs, J., Jones, C. Colostrum management tools: hydrometers and refractometers. Penn State Extension. 2011.
- Thornhill, J. B., Krebs, G. L., Petzel, C. E. Evaluation of the brix refractometer as an on-farm tool for the detection of passive transfer of immunity in dairy calves. Aust. Vet. J. 2015, 93(1-2), 26–30. [CrossRef]
- Giammarco, M., Chincarini, M., Fusaro, I., Manetta, A. C., Contri, A., Gloria, A., Lanzoni, L., Mammi, L. M., Ferri, N., Vignola, G. Evaluation of brix refractometry to estimate immunoglobulin G content in buffalo colostrum and neonatal calf serum. Animals 2021, 11(9), 2616. [CrossRef]
- El-Loly, M. M., Mansour, A. I. A. Relationship between the values of density and immunoglobulin concentrations of buffalo’s colostrum and their thermal stability. J. Med. Sci. 2013, 13(8), 723–729. [CrossRef]
- McGuirk, S. M., Collins, M. Managing the production, storage, and delivery of Colostrum. Veterinary Clinics of North America: Food Anim. Prac. 2004, 20(3), 593–603. [CrossRef]
- Godden, S. M., Smith, S., Feirtag, J. M., Green, L. R., Wells, S. J., Fetrow, J. P.Effect of on-farm commercial batch pasteurization of Colostrum on Colostrum and serum immunoglobulin concentrations in dairy calves. J. Dairy Sci. 2003, 86(4), 1503–1512. [CrossRef]
- Godden, S. Colostrum management for dairy calves. Vet. Clin. North Am. Food Anim. Prac. 2008, 24(1), 19–39. [CrossRef]
- Cummins, C., Lorenz, I., Kennedy, E. Short communication: The effect of storage conditions over time on bovine colostral immunoglobulin G concentration, bacteria, and ph. J. Dairy Sci. 2016, 99(6), 4857–4863. [CrossRef]
- Foley, J. A., Otterby, D. E. Availability, storage, treatment, composition, and feeding value of surplus colostrum: A Review. J. Dairy Sci. 1978, 61(8), 1033–1060. [CrossRef]
- Manohar A.A., Williamson M., Koppikar G.V. Effect of storage of colostrum in various containers. Indian Pediatr. 1997, 34, 293–5.
- Piantedosi, D., Servida, F., Cortese, L., Puricelli, M., Benedetti, V., Di Loria, A., Manzillo, V. F., Dall’Ara, P., & Ciaramella, P. Colostrum and serum lysozyme levels in Mediterranean Buffaloes (bubalus bubalis) and in their newborn calves. Vet. Rec. 2010, 166(3), 83–85. [CrossRef]
- Paulik, S., Slanina, L., Polacek, M. Lysozyme in the colostrum and blood of calves and dairy cows. Vet. Med. (Praha) 1985, 30, 21-28.
- Foster, D. M., Smith, G. W. Pathophysiology of diarrhea in calves. Vet. Clin. North Am. Food Anim. Pract. 2009, 25(1), 13–36. [CrossRef]
- Khattar, S., Pandey, R. Cell culture propagation of calf rotavirus and detection of rotavirus specific antibody in colostrum and milk of cows and buffaloes. Rev. Sci. Tech. 1990, 9(4), 1131–1138. [CrossRef]
- Geletu, U. S., Usmael, M. A., & Bari, F. D. Rotavirus in calves and its zoonotic importance. Vet. Med. Intern. 2021, 1–18. [CrossRef]
- Rast, L., Lee, S., Nampanya, S., Toribio, J.-A. L., Khounsy, S., & Windsor, P. A. Prevalence and clinical impact of Toxocara vitulorum in cattle and buffalo calves in northern Lao PDR. Trop. Anim. Health Prod. 2012, 45(2), 539–546. [CrossRef]
- Rajapakse, R. P. V. J., Lloyd, S., Fernando, S. T. Toxocara vitulorum: Maternal transfer of antibodies from Buffalo Cows (bubalis bubalis) to calves and levels of infection with T vitulorum in the calves. Res. Vet. Sci. 1994, 57(1), 81–87. [CrossRef]
- Nagy, B., Fekete, P. Z. (Enterotoxigenic escherichia coli in veterinary medicine. Inter. J. Med. Microb. 2005., 295(6-7), 443–454. [CrossRef]
Table 1.
Composition of Water Buffalo colostrum.
| Nutrient | Mean ± SD |
|---|---|
| Fat (%) | 11.31 ± 0.39 |
| Protein (%) | 8.73 ± 0.15 |
| Lactose (%) | 3.73 ± 0.02 |
| Total Solids (%) | 25.31 ± 0.02 |
| Ash (%) | 0.94 ± 0.02 |
| pH | 6.01 ± 0.01 |
Source: Coroian et al. [30].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



