
Submitted:
28 February 2026
Posted:
04 March 2026
You are already at the latest version
Abstract
Cephalonomia stephanoderis (Hymenoptera: Bethylidae) is a solitary ectoparasitoid of Coffee Berry Borer (CBB) that is established in the field in Puerto Rico but poorly studied. Five coffee farms in Maricao, Lares, Utuado, Adjuntas, and Jayuya were surveyed monthly from February 2022 to January 2023 for evidence of C. stephanoderis. On each collection date, fifty dry and fifty mature ripe coffee berries were collected from the tree and held for emergence of CBB adults and parasitoids. From January to April 2023, we collected 50 dry berries (“raisins”) from the same sites but placed each berry in a separate tube to count the number of C. stephanoderis and the number of unparasitized CBB adults which emerged so that we could estimate percent parasitism. In 2022, results showed that dry berries produced significantly more parasi-toids than mature berries. Infested coffee berries from Adjuntas contained the most C. stephanoderis and coffee berries from Jayuya had the fewest. Temperature negatively affects the parasitoid population, while relative humidity and alti-tude effects were not significant. In 2023, the parasitoid population was highest in February and decreased steadily until the end of sampling in April. The percentage parasitism ranged from 2.79% to 4.97% across collection sites. The potential for augmentation biological control for C. stephanoderis and classical biological control via the introduction of other CBB parasitoids are discussed.
Keywords:
Cephalonomia stephanoderis
; Bethylidae
; coffee berry borer
; parasitoid
; phenology
; Puerto Rico island
1. Introduction
The coffee berry borer (CBB) is the world’s primary pest in the coffee industry, and estimated annual losses are about $500 million [1]. In Puerto Rico, this pest arrived in 2007; since then, decreasing production has been evident [2]. On the island, the control of CBB has been done mainly through the entomopathogenic fungus Bauveria bassiana with both the commercial strain (Mycotrol®) and local strains of the fungus [3,4]. However, other biological controls for CBB have been successfully used in several countries, such as the larval-pupal parasitoids Cephalonomia stephanoderis Betrem and Prorops nasuta Waterston (Hymenoptera: Bethylidae) and the adult parasitoid Phymastichus coffea (Hymenoptera: Eulophidae) [5]. These parasitoids have been mass-produced in laboratories on coffee beans [6,7] or artificial diets [8,9,10,11].
One of the most well studied parasitoids of CBB is Cephalonomia stephanoderis Betrem a solitary ectoparasitoid reported for the first time in the Ivory Coast [12]. The adult parasitoid measures around 2 mm for females but males are somewhat smaller, and both have a shiny black color, transparent wings with reduced venation and hairs on the margins [13], and a rectangular-shaped head [14]. This species is synovigenic, with 48 to 70 days of longevity depending on environmental conditions [8,14,15]. Infante et al., [13] described the life stages and morphology of the wasp, and other others have examined host discrimination [16,17], parasitism behaviors [18], adult performance [19,20], oviposition and fecundity [21,22], response to volatiles [23,24], temperature-dependent development [25], and interactions with other parasitoids [26].
In addition to parasitism of the pupal and larval stages, C. stephanoderis is an efficient predator feeding on these life stages as well. In the laboratory, C. stephanoderis can oviposit from one to three eggs per day, and the same quantity can be killed via host feeding [14,22,27,28]. In the field, Aristizabal et al. [29] found significant predatory activity against CBB which contributed significantly to controlling the pest.
After some observations of C. stephanoderis performance against CBB in laboratory and in field-controlled conditions, several countries have introduced this parasitoid to attempt to control the CBB pest, and massive Classical biological control introductions and mass releases of C. stephanoderis have been conducted worldwide [27,30,31], and several countries, including India, Brazil and Mexico, have reported successful application in the field [32,33,34,35]. In Colombia, augmentation biological control resulted in 49% reduction in the CBB population [29]. A parasitism rate of 35% in the field was observed in India. These results suggest this parasitoid has considerable potential as a natural enemy of CBB [36]. The success or failure of the establishment of natural enemies released in the field depends on many environmental factors (temperature, altitude, humidity, etc.), and these factors can affect the performance and success of parasitoids in controlling the pest [37,38).
Cephalonomia stephanoderis is established in Puerto Rico, having been first recorded parasitizing coffee berry borer in 2009 [39,40]. Subsequent studies in 2013-14 showed field parasitism of 23% [41,42] but nothing has been reported since that time. In this study we surveyed coffee farms across the island to determine the presence or absence of C. stephanoderis and current parasitism rates.
2. Materials and Methods
2.1. Sampling Dry Versus Mature Berries for Parasitoids
To determine C. stephanoderis’s presence in the field, we sampled coffee berries infested by CBB in Puerto Rico from February 2022 to April 2023. We surveyed five coffee farms in the coffee crop area. The coffee farms were located in Maricao (18°10’8”N, 66°54’21”W, 800 masl), Jayuya (18°11’21”N, 66°34’26”, 750 masl), Adjuntas-Experimentatl Station of the University of Puerto Rico (18º10’12” N, 66º47’44”, 620 masl), Utuado (18°17’17”N, 66°46’12”W, 390 masl), and Lares (18°16’18”N, 66°54’18”W, 340 masl) municipalities. The area sampled was around 500 m2 in each location, and by a zigzag transect, we tried not to get more than ten berries from the same tree. The predominant coffee cultivars, Coffea arabica, sampled were “Limaní,” “Fronton,” “Caturra,” “Catuai,” “Borbon,” and “Tipica.” We collected infested “dry” (raisins) and “mature” (ripe) coffee berries. The “dry” berries refer to the berries with dark color without the presence of berry pulp, and “mature” berries refer to berries that are full size and green or red with CBB infested. Both berry maturity stages had CBB in positions C and D (according to Bustillo et al., 1998 [43]) which ensured that CBB adults were well established inside with progeny suitable for parasitism by C. stephanoderis.
We randomly collected 50 CBB-infested coffee berries in a plastic bag monthly, adding a label with information about the places sampled. In the laboratory, berries were placed in a plastic container (11.30cm Ø on the top, 8.5 cm Ø at the bottom, and 8 cm high). A paper towel was placed at the bottom of the container to absorb moisture and avoid excess humidity, and on the top, the container had a lid covered in part of the area with fine mesh to allow ventilation. To avoid fungal development in the mature berries, these were cleaned, and infested berries were first sterilized by dipping in a 2% sodium hypochlorite solution for 10 minutes and then rinsing them with distilled water and dried on paper towels. Then, the containers were placed inside an incubator for development of any parasitoids under controlled conditions (25+2 ºC, 75+5 RH, and photoperiod of 12:12 L:D). Containers with berries were checked for wasps every two days for about two months following field collection. A stereomicroscope (Leica EZ4W) was used to count and identify the parasitoids. Some of the live parasitoids were used to initiate a colony in the laboratory, and the rest were placed in Eppendorf tubes (1 ml) with 70% alcohol for reference. The five coffee farms in the different municipalities were sampled for one year from February 2022 to January2023. Climatic data (temperature and relative humidity) were obtained from a weather station at the Adjuntas Experimental Station belonging to the University of Puerto Rico and reflected seasonal changes in temperature and RH for all farms rather than farm-specific differences.
2.2. Sampling to Estimate Percent Parasitism
From February 2023 to April 2023, we collected CBB-infested berries differently so that we could estimate percent parasitism. We collected only dry berries from coffee plants because these were known to have more parasitoids from 2022 samples. In the same way as before and at the same locations. We collected 50 infested berries monthly from the same five coffee farms, but this time, we placed each berry in a glass tube (150 x 20 mm) with a lid and held them under controlled conditions (25+2 ºC, 75+5 RH, and photoperiod of 12:12 L:D) as described above. Every three days we checked the tubes to observe newly emerged parasitoids. After one month, we dissected the berries to find any remaining parasitoids and counted the numbers of CBB adults. The rationale was that each CBB individual inside the coffee berry either became an adult beetle or was parasitized and yielded a wasp.
Therefore, the equation to calculate parasitism was:
Where PP= parasitism percentage, Np= Number of parasitoids and NCBB= Number of coffee berry borer adults collected from infested berries.
CBB mortality due to predation by C. stephanoderis in the coffee berry was not evaluated and may have added a significant.
2.3. Statistical Analysis
Data on weather conditions was recorded to analyze if there was a relationship between temperature, relative humidity, and altitude and the number of C. stephanoderis collected from infested berries. We analyzed the differences between dry and mature berries with C. stephanoderis emerging through the Wilcoxon non-parametric t-test since the assumptions of normality were not accomplished. Generalized linear models (GLM) were used to observe differences in the number of C. stephanoderis collected by location and in dry and mature berries at each location over time. We use Poisson error distributions with a logarithm link function for parasitoid counts from the collected berries. For parasitism estimates, percentages were used, and all analyses were run using R software version 4.2.2.
3. Results
3.1. Cephalonomia Stephanoderis from Dry Versus Mature Coffee Berries
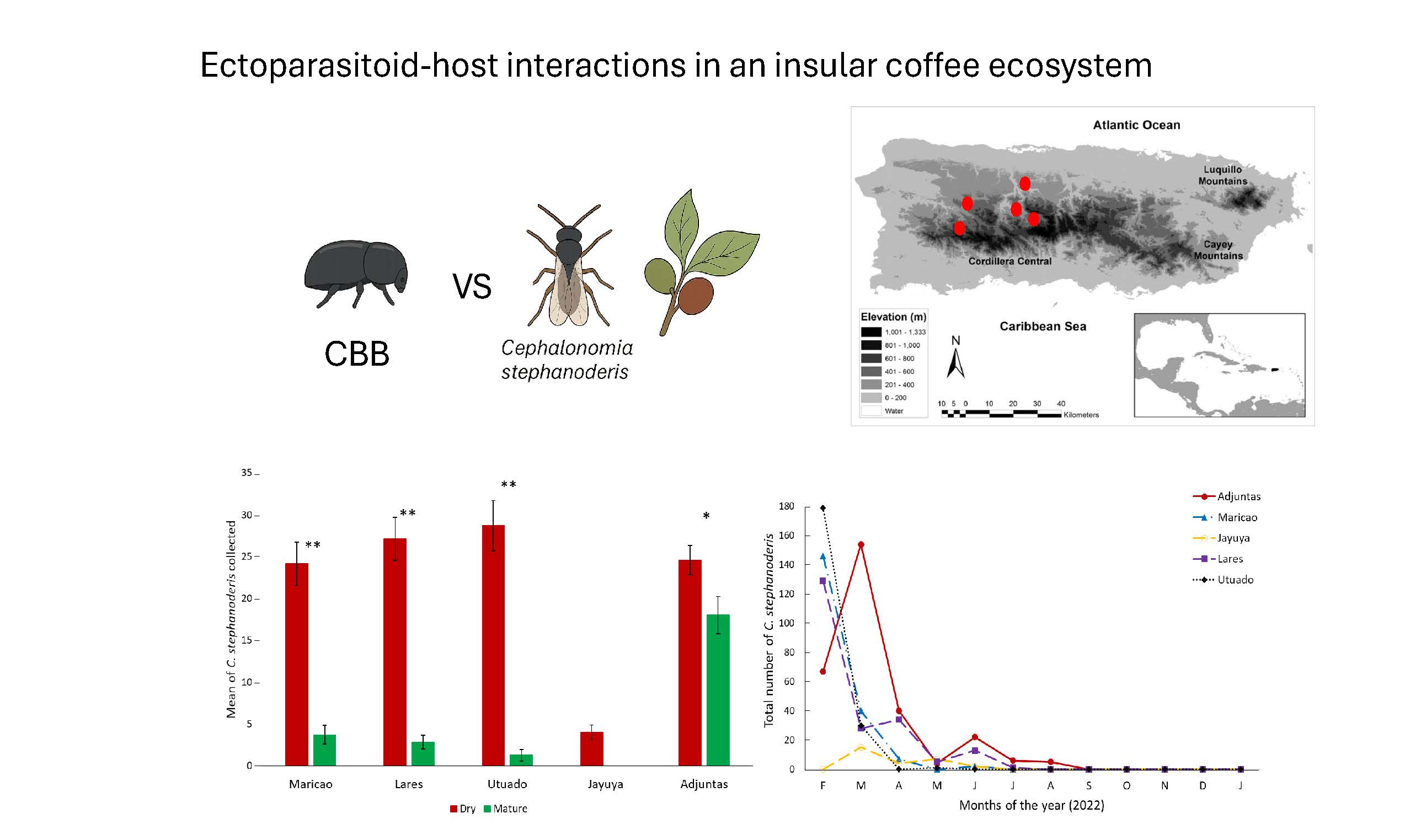
We collected 6,000 CBB-infested berries from the five commercial coffee farms (Figure 1); of this total, 3,000 were from dry berries, while 3,000 were mature (ripe) berries, and all had CBB in positions C and D inside the fruit. Cephalonomia stephanoderis was the only parasitoid found in the sampled coffee berries. In total, 941 adult C. stephanoderis were collected, with 760 (80.7%) of individuals emerging from dry coffee berries, and 181 (19.2%) emerging from mature coffee berries, which was a significant difference (W=832.5, P=0.004). The higher number of parasitoids in dry berries than mature berries is likely due to higher numbers of CBB in dry berries given the longer period of infestation and reproduction.
3.2. Total of Parasitoids Produced per Location
When the numbers of C. stephanoderis adults were analyzed per place (Maricao, Lares, Utuado, Jayuya, and Adjuntas), we found that Adjuntas (42.5 individuals) on average had a more significant number of Cs (z=64.922, P=≥0.0001) and Jayuya (4) was the lowest place having parasitoids (z=-11.983, P=≥0.0001), while Maricao (27.8), (z=-4.644, P=≥0.0001), Lares (30) (z=-3.924, P=≥0.0001) and Utuado (30) (z=-11.925, P=≥0.0001) had a similar number of the parasitoids.
3.2.1. Parasitoids Produced per Location in Dry and Mature Coffee Berries
When we analyzed the means in dry and mature berries containing C. stephanoderis parasitoids in each farm, we observed that the type of grain per location was affecting the results (z=67.509, P<0.0001). In every farm, the number of parasitoids collected was significantly higher in dry coffee berries, and fewer parasitoids were founded in mature coffee berries for Maricao (z=-6.473, P<0.0001), Lares (z=-7.385, P<0.0001), Utuado (z=-7.756, P<0.0001) and Adjuntas (z=-2.922, P<0.007); while for Jayuya (z=-0.001, P>0.05) there were no differences due to no parasitoids being collected from mature berries (Figure 2).
3.3. Parasitoids Abundance over Time at Each Location
Analyzing the total number of wasps found throughout the year per location, we observed that Adjuntas was the place where we could find more individuals in March (154), while Utuado (179), Lares (129), and Maricao (146), February was the highest number of individuals found, and for Jayuya March was the time when more parasitoids were found (15). However, this place had the lowest number of parasitoids (Figure 3). The peak of C. stephanoderis highest number was collected in February and March, depending on the location, and the number decreased until August and disappeared from September until January for all the locations.
3.4. Parasitoids Abundance over Time
Results about the average C. stephanoderis collected during the different months of the year show that February has the highest number of parasitoids collected during the year (z=107.5, P<0.0001), followed by March (z=-9.16, P<0.0001) and April (z=-15.68, P<0.0001). The months with parasitoids collected less than five on average were May (z=-13.96, P<0.0001), June (z=-15.73, P<0.0001), July (z=-11.37, P<0.0001), and August (z=-10.38, P<0.0001) (Figure 4). For the rest of the year, no parasitoids were found.
3.5. Correlation Between Parasitoid Abundance and Environmental Factors
When the correlation between the number of C. stephanoderis individuals against temperature, relative humidity, and altitude was analyzed, only temperature has a negative effect on the presence of the parasitoid, suggesting that when the temperature is increased, the number of parasitoids decreases (Table 1 and Figure 5). The temperature throughout the year varied from 17.2 to 23 °C, but C. stephanoderis were collected between 19.6 and 23 °C on average; the relative humidity ranged from 86.6 to 91.9% on average during the year, and the altitude varied from 350 to 800 masl, and we found individuals in all the locations.
3.6. Percentage of Parasitism by Cephalonomia stephanoderis
For the percentage of parasitism, berries collected from January to April 2023 were used and we found a maximum number of 87 CBB per coffee berry, while the minimum was 1, but on average, it varies from 5.5 to 13.6 individuals per coffee berry through the year. Then, the percentage of parasitism was 0% for January, where no C. stephanoderis individuals collected, while for February, March, and April, parasitism percentages were 3.49%, 2.79%, and 4.97%, respectively (Table 2). For the percentage of February, it is important to mention that only one place (Lares) had parasitism, and no parasitoids were found in the four remaining locations. In March, we could find parasitism in four of the five locations, while in April, only Utuado had no parasitism (Figure 6).
The correlation between the number of CBB collected monthly and the number of C. stephanoderis was not significant (R=0.86, R2 =0.74, P=0.13), meaning that the parameters to express the close relationship between these two species need to be better studied to influence the number of parasitoids.
4. Discussion
Parasitoids produced on dry coffee berries overcome the number of wasps found on mature coffee berries, and it could be related to the number of hosts per coffee berry. Mathieu [44] found that CBB is in a phase of multiplication where all types of coffee berries (green, red, and dry coffee berries) can be found, and the number of CBB is lower, while the number of CBB is increased inside the dry coffee berries. It coincides with Balakrishnan & Sreedharan [45], who found that the activity of the parasitoid C. stephanoderis was low in ripe fruit (red coffee berries) and high in leftover dry berries, confirming our results. A previous study by Gonzalez-Cardona [41] also found that in Puerto Rico, no natural enemies were in green and ripe coffee berries infested by CBB; this result contrasts with our findings in as much as we found few parasitoids. It could be because the green and mature coffee berries infested with CBB are in the reproduction phase but not with the highest number of CBB life stages that allow the highest reproduction of the wasp. Ticheler [12] mentioned that C. stephanoderis can attack CBB in the red berries with about 27% and 50% for the dry coffee berries because the dry coffee berries contain more live stages of CBB available for reproduction. The author suggests that in an ideal coffee crop plantation, C. stephanoderis can decrease the level of the CBB population at the end of the harvest season since raisins contain more parasitoids. The percentage parasitism estimated from our study underestimates the total impact of C. stephanoderis as host predation on larvae and pupa can be significant mortality factor and was not measured.
Despite the coffee crop area in Puerto Rico sharing similar weather conditions among the municipalities sampled, these areas showed differences in the number of wasps found, and some authors explain that as the result of different microclimates that generate different conditions for the development of different communities of plants [46]. Also, the management of pests can affect the results of the data collected. For example, the location of Adjuntas had a higher number of C. stephanoderis, and it might be because it is located in the Experimental Substation of Adjuntas (EEA) area belonging to the University of Puerto Rico, where there previously was an effort to collect dry coffee berries infested with CBB by Dr. Fernando Gallardo (UPRM), and the parasitoid wasps from these collections, C. stephanoderis, were released back in the area of the experimental substation, and this could affect the wasp population abundance living in the area. On average, in Adjuntas, we found 42.5 individuals, and in the same area, a previous study including another location (Maricao) reported 225 individuals of C. stephanoderis for three years, meaning a 0.6% of parasitized coffee berry fruit in average [41], which is lower than our results. It is important to mention that in a fourth of the five areas sampled, only Jayuya was the farm where no direct management practices for CBB control, excluding end-of-season sanitation, were applied, and it was the place where a smaller number of individuals were found. In contrast, the rest of the farms used insecticides and fungi B. bassiana to control CBB and other pests. Landis and Menalled [47] mention that pesticide application is a key factor that limits parasitoid effectiveness. It contrasts with our results, where we found a bigger number of parasitoids in the farms where farmers apply insecticides (Adjuntas, Lares, Maricao, and Utuado). In comparison, we found fewer individuals in places where no insecticides were applied (Jayuya). Maricao, Lares, and Utuado had a similar number of individuals of C. stephanoderis. These farms have management for CBB, and no management to increase the number of natural enemies is used as that used in Adjuntas.
As expected, per each location, the number of individuals of C. stephanoderis found in dry coffee berries surpassed the number of parasitoids in mature coffee berries, confirming some observations done by Aristizabal et al. [31], who mention parasitism is higher in dry coffee fruits. In their study, they found 10.1% on average. This also confirms that this parasitoid is more abundant in dry coffee berries on the coffee plant than in ripe coffee berries since the number of CBB life stages for reproduction is higher [12,46].
According to Hassell [48], parasitism depends mainly on host density, but it is more difficult to evaluate in the field. In this case, we found C. stephanoderis is present inside berries during the period from February to August, and it coincides with Mathieu et al. in New Caledonia [44], who described the period from January to July as the survival phase for CBB, where the quantity of CBB in the field is higher in this period. Also, their findings showed that CBB is found in minor quantities in the period, which they called the multiplication phase of CBB, from October to January. Here in our study, the C. stephanoderis parasitoid was absent, indicating that the species needs the host’s availability to be present. Since the multiplication phase is during this time, the number of CBB stages is probably low. This is probably why the parasitoid is absent from the sampling records. In the case of Puerto Rico, specifically, sanitary harvest is recommended from January to February, after the harvest season when some dry berries remain there [49]. However, our findings show that C. stephanoderis was collected in significant quantities during these months. Then, to maintain and strengthen the population persistence of the parasitoid in the coffee area, maintaining the coffee dry berries on the crop field should be important to increase their population and contribute to the CBB control. This can be ineffective in the control management of CBB.
Our findings show no differences in the number of C. stephanoderis along the altitude, but the altitude of our collection locations was between 340 and 800 masl. Even though Mariño et al. [50], in Puerto Rico, found the highest infestations of CBB at places above 400 masl, we could find parasitism below this altitude. Ijala et al. [51] found high abundance (around 75 individuals on average) at medium altitude (1500-1679 masl), and Maldonado & Benavides [52], in Colombia, collected coffee samples in 17 municipalities that were situated between 1150 and 1840 masl no C. stephanoderis was found. These suggest that different factors can affect the development of the species.
The temperature had a negative effect on the parasitoid C. stephaoderis, with more individuals with temperatures around 20 °C, and similar results were found by Ijala et al. [51], who mentioned that a high abundance of the parasitoids was influenced by shade trees, which influenced the temperature that was from 22 to 25 °C. The authors mention that temperature has a negative effect on the abundance of the parasitoid C. stephanoderis, and this agrees with our results, where temperature above 22 °C in the field showed a smaller number of the parasitoid.
The highest percentage of parasitism found in our study is 4.97%, while Gallardo-Covas in 2019 [42] reported 23% parasitism in Puerto Rico. Part of this result can be attributed to meteorological phenomena such as hurricanes. In September 2022, hurricane Fiona affected the island and probably affected the development of the CBB populations and then the parasitism in the field.
However, our results are in the range of parasitism of C. stephanoderis that several authors reported previously between 0.7% and 24% [30,31,35,53,54]. In the best cases of parasitism for C. stephanoderis, some authors found above 30% [29,36,45]. Moreover, in controlled cage environments, the results improve, showing a reduction of the CBB population by 65% [29]. The parasitoid population in Puerto Rico seems to be well established in natural conditions since 2009 and positively affecting the control of CBB. It contradicts Quintero et al. [35], who said that after some time, the population of parasitoids introduced to new places tends to decrease and, in some cases, to disappear.
The number of CBBs per coffee berry reaches up to 87 individuals, almost double the quantity compared with Mariño et al. [50], since these authors found a maximum number of 47 individuals. It could be because the samples they collected were from ripe berries from August to November, while our data comes from the dry coffee berries in March, where the remaining dry coffee berries are in the field and are supposed to have a higher number of CBB life stages.
As studied by Benavides et al. [55], the parasitism of C. stephanoderis is related to the number of the host. In our result, we found that the correlations between the number of the parasitoid do not increase as the number of the CBB did rise. It contrasts with Lauziere et al. [56], who suggested that the efficacy of the parasitoid depends on the seasonal variation of its host density since, in laboratory conditions, C. stephanoderis delays oogenesis and extends the pre-oviposition time at low host densities.
Cephalonomia stephanoderis populations in the field are modulated by different environmental variables, making it challenging to understand insect performance. However, as much information as we can gather on the behavior and affecting factors, managing the pest with the help of such natural enemies could be improved.
Augmentation of biological control of C. stephanoderis could improve the management and decrease populations of CBB, and a possible introduction of new parasitoids such as Phymasticus coffea could improve the control of the pest.
Author Contributions
Conceptualization, J.C.V.R. and M.C.V.H.; methodology, M.C.V.H. and R.R.B.; formal analysis, M.C.V.H.; resources, J.C.V.R. and P.F.; writing—original draft preparation, M.C.V.H.; writing—review and editing, M.C.V, H., J.C.V.R., P.F.; supervision, J.C.V.R. and P.F.; funding acquisition, J.C.V.R. All authors have read and agreed to the published version of the manuscript.”.
Funding
This work was funded by United States Department of Agriculture Area Wide project (NACA 58-2040-6-007) a cooperative agreement (AP19PPQS&T00C181) with the United States Department of Agriculture, Animal and Plant Health Inspection Service (APHIS) and CEQUIS, University of Puerto Rico, Agricultural Experimental Station. This material was made possible, in part, by a Cooperative Agreement from the United States Department of Agriculture’s Animal and Plant Health Inspection Service (APHIS). It may not necessarily express APHIS’ views.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
To the farmers who allow us to collect infested coffee berries in their crops. We thank Paola Agosto for her help with the laboratory and some field collections, and I thank Roberto Soto and Roger Lawrie for their help with some field collections. Mention of trade names or commercial products in this publication is solely for providing specific information and does not imply recommendation or endorsement by the USDA.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Vega F. E., Franqui R.A., Benavides P. Scientific note: The presence of the coffee berry borer, Hypothenemus hampei, in Puerto Rico: fact or fiction? J. Insect Sci. 2002, 2,1-3. [CrossRef]
- NAPPO- Norteamerican Organization of Plant Protection 2007. Detecciones de la Broca del Café Hypothenemus hampei in Puerto Rico, Estados Unidos. Oficial Pest Report. https://pestalerts.org/es/official-pest-report/detecciones-de-la-broca-del-caf-hypothenemus-hampei-en-puerto-rico-estados. (May 2023).
- Bayman P., Mariño Y.A., Garcia-Rodriguez N.M., Oduardo-Sierra O.F., Rehner S. Local isolates of Bauveria bassiana for control of the coffee berry borer Hypothenemus hampei in Puerto Rico: Virulence, efficacy and persistence. J. Biol.Control 2021,155,1-10. [CrossRef]
- Aristizabal A.L.F., Johnson M., Shriner S., Hollingsworth R., Manoukis N.C., Myers R., Bayman P., Arthus S.P. Integrated Pest Management of Coffee Berry Borer in Hawaii and Puerto Rico: current Status and Prospects. Insects 2017, 8,1-16. [CrossRef]
- Infante F. Pest Management Strategies Against the coffee Berry Borer (Coleoptera: Curculionidae: Scolytinae). J. Agric. Food Chem. 2018, 66, 5275-5280. [CrossRef]
- Orozco-Hoyos J. A guide to the rearing of the parasitoid Phymastichus coffea for control of the coffee berry borer. The Commodities Press, Cali, Colombia, 2002; pp.19.
- Portilla R.M. & Bustillo P.A. Nuevas investigaciones de Hypothenemus hampei y de sus parasitoides Cephalonomia stephanoderis y Prorops nasuta. Rev. Colomb. Entomol. 1995, 21, 25-33.
- Portilla M & Streeett D. Producción masiva automatizada de la Broca del Café Hypothenemus hampei (Ferrari) (Coleoptera: Scolytidae) y de sus parasitoides sobre dietas artificiales. Sist. Agroeco. Mod. Biomatematic 2008,1, 9-24.
- Portilla M. (1999). Mass rearing technique for Cephalonomia stephanoderis (Hymenoptera: Bethylidae) on Hypothenemus hampei (Coleoptera: Scolytidae) developed using Cenibroca artificial diet. Rev. Colomb. Entomol 1999, 25, 57–66. [CrossRef]
- Bustillo A.E., Orozco H.J., Benavides M.P., Portilla R.M. Producción masiva y uso de parasitoides para el control de la broca del café, Hypothenemus hampei, en Colombia. Cenicafé, 1996; 47, 215-230.
- Brun L.O., Gaudichon V., Wigley P.J. An artificial diet for continuous rearing of the coffee berry borer, Hypothenemus hampei (Ferrari) (Coleoptera: Scolytidae). Insect Sci. Appl.1993, 14,585-587. [CrossRef]
- Ticheler J. Etude analytique del’épidémiologie du scolyte des graines de café Stephanoderis hampei Ferr. en Côte d’Ivoire. Mededelingen Landbouwhogeschool Wageningen 1961, 61, 1-49.
- Infante F., Valdez J., Penagos D.I., Barrera J.F. Description of the life stages of Cephalonomia stephanoderis (Hymenoptera: Bethylidae), a parasitoid of Hypothenemus hampei (Coleoptera: Scolytidae). Vedalia 1994, 1, 13-18.
- Abraham Y.J., Moore D., Godwin G. Rearing and aspects of biology of Cephalonomia stephanoderis and Prorops nasuta (Hymenoptera: Bethylidae) parasitoids of the coffee berry borer, Hypothenemus hampei (Coleoptera: Scolytidae). Bull. Entom. Res. 1990, 80, 121-128. [CrossRef]
- Garcia H.M., Duque E.T.C., Rodriguez V.Y., Borrero R.Y., Cabrera R.D., Campo C.Y. Reproducción, ciclo biológico y relación sexual de Cephalonomia stephanoderis Betrem en condiciones de laboratorio en Cuba. Fitosanidad 2008, 12, 227-232.
- Alonso J., Cabrera M., Barrera J.F. The importance of the Parasitoid Egg in Host Discrimination by Cephaolnomia stephanoderis, and Ectoparasitoid of the Coffe Berry Borer Hypothenemus hampei. Biocontrol Sci. Techn. 1998, 8, 153-162.
- Barrera J.F., Gomez J., Alauzet C. Evidence for a marking pheromone in host discrimination by Cephalonomia stephanoderis (Hym.: Bethylidae), Short Note. Entomophaga 1994, 39, 363-366. [CrossRef]
- Lauziere I., Perez-Lachaud G., Brodeur J. Behaviour and Activity Pattern of Cephalonomia stephanoderis (Hymenoptera: Bethylidae) attacking the Coffee Berry Borer, Hypothenemus hampei (Coleoptera: Scolytinae). J. Insect Behav 2000, 13, 1-21. [CrossRef]
- Alonso C.P.L., Rodriguez A.L., Monzon R.C.E., Delgado T.R., Perez R.Y. Evaluation of the parameters of quality and biological effectiveness of Cephalonomia stephanoderis Betrem under Cienfuegos province conditions. Rev. Científ. Agroecosist. 2022, 10, 130-138.
- Gomez J., Barrera J. F., Leido P., Valle J. Influence of age and diet on the performance of Cephalonomia stephanoderis (Hymenoptera, Bethylidae) a parasitoid of the coffee berry borer, Hypothenemus hampei (Coleoptera, Curculionidae). Rev. Brasil. Entom. 2012, 56, 95-100. [CrossRef]
- Lauziere I., Perez-Lachaud G., Brodeur J. Importance of nutrition and host availability on oogenesis and oviposition of Cephalonomia stephanoderis (Hymenoptera: Bethylidae). Bull. Entom. Res. 2001, 91, 185-191. [CrossRef]
- Lauziere I., Perez-Lachaud G., Brodeur J. Effect of female body size and adult feeding on the fecundity and longevity of the parasitoid Cephalonomia stephanoderis Betrem (Hymenoptera: Bethylidae). Ann. Entomol. Soc. Am. 2000, 93, 103-109. [CrossRef]
- Chiu-Alvarado P. & Rojas J.C. Behavioural responses of bethylid parasitoid species of the coffee Berry borer to chemicals cues from host and non-host dust/frass. Biocontrol 2011, 56,45-53. [CrossRef]
- Gomez J., Barrera J.F., Rojas J.C., Macias-Samano J., Liedo J.P., Cruz-Lopez L., Badii M.H. Volatile compounds released by disturbed females of Cephalonomia stephanoderis (Hymenoptera: Bethylidae): a parasitoid of the coffee berry borer Hypothenemus hampei (Coleoptera: Scolytidae). Fla. Entomol. 2005, 88, 180-187. [CrossRef]
- Infante F., Luis J.H., Barrera J.F., Gomez J., Castillo A. Thermal constants for preimaginal development of the parasitoid Cephalonomia stephanoderis Betrem (Hymenoptera: Bethylidae). Can. Entomol 1992, 124, 935-941. [CrossRef]
- Infante F., Mumford J., Baker P., Barrera J., Fowler S. Interspecific competition between Cephalonomia stephanoderis and Prorops nasuta (Hym., Bethylidae), parasitoids of the coffee berry borer, Hypothenemus hampei (Col., Scolytidae). J. Appl. Entomol. 2001,125, 63-70. [CrossRef]
- Damon A. & Valle J. Comparison of two release techniques for the use of Cephalonomia stephanoderis (Hymenoptera: Bethylidae), to control the coffee berry borer Hypothenemus hampei (Coleoptera: Scolytidae) in Soconusco, southeastern Mexico. Biol. Control 2002, 24, 117-127.
- Lauziere I., Brodeur J., Perez-Lachaud G. Host stage selection and suitability in Cephalonomia stephanoderis Betrem (Hymenoptera: Bethylidae), a parasitoid of the coffee Berry borer. Biol. Control 2001, 21, 128-133. [CrossRef]
- Aristizabal A.L.F., Bustillo P.A.E., Baker P.S., Orozco J.H., Chaves B.C. Depredatory effects of the parasitoid Cephalonomia stephanoderis on the immature stages of Hypothenemus hampei under field conditions. Rev. Colomb. Entomol. 1998, 24, 35-41. [CrossRef]
- Vijayalakshmi C.K., Rahiman P.A., Sreedharan K., Selvakumar M. Field evaluation of coffee Berry Borer parasitoid Cephalonomia stephanoderis (Betrem) in the coffee tracts of Wayanad, Kerala. International Scientific Association of Coffee in British Library Conference Proceedings, Bangalore, India, pp. 1282-1283.
- Aristizabal A.L.F., Baker P.S., Orozco J.H., Chaves B.C. Parasitism of Cephalonomia stephanoderis Betrem on populations of Hypothenemus hampei (Ferrari) at infestation low levels in the field. Rev. Colomb. Entomol. 1997, 23, 157-164. [CrossRef]
- Aristizabal A.L.F., Salazar H.M., Mejía C.G., Jimenez M., Bustillo A.E. and Arthurs S.P. Establishment of exotic parasitoids of the coffee berry borer Hypothenemus hampei (Coleoptera: Curculionidae: Scolytinae) in Colombia through farmer participatory research. Int. J. Trop. Insect Sci. 2012, 32, 24-31. [CrossRef]
- Souza M.S., Texeira C.A.D, Azevedo C.O., Costa V.A., Costa J.N.M. Occurrence of Cephalonomia stephanoderis Betrem (Hymenoptera: Bethylidae) in Coffee Plantations of the Brazilian Amazonia, Scientific note. Neotrop. Entomol. 2006, 35, 560-562. [CrossRef]
- Bustillo A.E., Cárdenas R., Posada F.J. Natural Enemies and Competitors of Hypothenemus hampei (Ferrari) (Coleoptera: Scolytidae) in Colombia. Neotrop. Entomol. 2002, 31, 635-639. [CrossRef]
- Quintero H.C., Bustillo P.A.E., Benavides M.P., Chaves C.B. Evidences of the establishment of Cephalonomia stephanoderis and Prorops nasuta (Hymenoptera: Bethylidae) in coffee plantations in the department of Nariño, Colombia. Rev. Colomb. Entomol. 1998, 24, 141-147. [CrossRef]
- Irulandi S., Samuel S.D., Kumar P.K.V., Sreedharan K. Parasitism by the exotic parasitoid, Cephalonomia stephanoderis Betrem (Hymenoptera: Bethylidae) on coffee berry borer, Hypothenemus hampei (F.) on Pulney Hills, Tamil Nadu. Hexapoda, 2009, 16, 32-35.
- Mailafiya D.M., Le Ru B.P., Kairu E.W., Calatayud P.A., Dupas S. Factors affecting stem borer parasitoid species diversity and parasitism in cultivated and natural habitats. Environ. Entomol. 2010, 39, 57-67. [CrossRef]
- Bourchier R.S., & Smith, S.M. Influence of environmental conditions and parasitoid quality on field performance of Trichogramma minutum. Entomol. Exp. Appl. 1996, 80, 461-468. [CrossRef]
- Abreu R.E. & Gallardo C.F. Presencia de Cephalonomia stephanoderis (Hymenoptera: Bethylidae) parasitoide de la broca del café, Hypothenemus hampei (Coleoptera: Curculionidae) en Puerto Rico. Proceedings of the Sociedad Puertoriqueña de Ciencias Agrícolas 2011, Ponce, Puerto Rico, pp. 9.
- Gallardo-Covas F. Biological control of insect pests in Puerto Rico. J. Agric Univ. Puerto Rico 2017, 101, 153-163. [CrossRef]
- González-Cardona O.P. Dinámica Poblacional de la Broca del Café Hypothenemus hampei (Ferrari) y sus Enemigos Naturales en el Cultivo de Café Coffea arabica L. en Puerto Rico. Master thesis, Master degree, Mayaguez, Puerto Rico, 2013.
- Gallardo-Covas F. (2019). Augmentation and conservation of Cephalonomia stephanoderis Betrem for the biological suppression of the coffee berry borer, Hypothenemus hampei Ferrari in Puerto Rico. Memorias Reunión de la Empresa de Café. Puerto Rico, 46p.
- Bustillo A.E., Cárdenas R., Villalba D., Benavides P., Orozco J., Posada F.J. (1998). Manejo Integrado de la Broca del café Hypothenemus hampei (Ferrari) en Colombia. Cenicafe-Boletín informativo sobre la broca del café, 1998, 134p.
- Mathieu F., Brun L.O., Frerot B., Suckling D.M., Frampton C. Progression in field infestation is linked with trapping of coffee berry borer, Hypothenemus hampei (Col., Scolytinae). J. Appl. Entomol. 1999, 123, 535-540. [CrossRef]
- Balakrishnan M.M., & Sreedharan, K. Establishment of Cephalonomia stephanoderis, the introduced parasitoid of coffee berry borer in Coorg District, Karnataka. In Proceedings of the 15th Plantation Crops Symposium Placrosym XV, Mysore, India, 10-13 December, 2002 (pp. 504-508).
- Bernaschini M.L., Rossetti M.R., Valladares G., Salvo A. Microclimatic edge effects in a fragmented forest: disentangling the drivers of ecological processes in plant-leafminer-parasitoid food webs. Ecol. Entomol. 2021, 46, 1047-1058. [CrossRef]
- Landis D. & Menalled F.D. Ecological considerations in the conservation of effective parasitoid communities in agricultural systems. In Conservation biological control, Barbosa P. Ed..Academic Press, London, UK, 1998; pp. 101-121.
- Hassell, M. The spatial and temporal dynamics of host-parasitoid interactions. 1 st ed.; Oxford University Press: New York, USA, 2000; p.198.
- Gallardo-Covas F. & Gonzalez-Cardona O.P., (2015). Manejo Integrado de la Broca del Café en Puerto Rico. Guía Tecnica, Departamento de Agricultura: San Juan, Puerto Rico , USA, 2015; p. 12.
- Mariño Y.A., Vega V.J., Garcia J.M., Verle R. J.C., Garcia N.M., Bayman P. The Coffee Berry Borer (Coleoptera: Curculionidae) in Puerto Rico: Distribution, Infestation, and Population per Fruit. J. Insect Sci. 2017, 17, 1-8. [CrossRef]
- Ijala A.R., Kyamanywa S., Cherukut S., Sebatta C., Karungi J. Parasitism of Hypothenemus hampei (Coleoptera: Scolytidae) in different farming systems and altitudes of Mount Elgon, Uganda. J. Appl. Entomol. 2019, 89,1-10. [CrossRef]
- Maldonado L.C.E. & Benavides M.P. Evaluación del Establecimiento de Cephalonomia stephanoderis y Prorops nasuta, Controladores de Hypothenemus hampei, en Colombia. Cenicafe 2007, 58, 333-339.
- Souza M.S., Almeida S.A., Teixeira C.A.D., Costa J.N.M. Parasitism in the population of the coffee berry borer Hypothenemus hampei (Ferrari) (Coleoptera: Curculionidae) by the parasitoid Cephalonomia stephanoderis Betrem (Hymenoptera: Bethylidae). EntomoBrasilis 2014, 7, 178-182. [CrossRef]
- Salazar H.M. & Baker P.S. Impacto de liberaciones de Cephalonomia stephanoderis sobre poblaciones de Hypothenemus hampei. Cenicafe 2002, 53, 306-316.
- Benavides M.P., Bustillo P.A.E., Montoya E.C. Advances on the use of the parasitoid Cephalonomia stephanoderis for the control of the coffee berry borer Hypothenemus hampei. Rev. Colomb. Entomol. 1994, 20, 247-253. [CrossRef]
- Lauziere I., Perez-Lachaud G., Brodeur J. Influence of host density on the reproductive strategy of Cephalonomia stephanoderis, a parasitoid of the coffee berry borer. Entomol. Exp. Appl. 1999, 92, 21-28. [CrossRef]
Figure 1.
Map of places of Cephalonomia stephanoderis collection in Puerto Rico.

Figure 2.
Mean (±SE) of Cephalonomia stephanoderis collected in dry and mature coffee berries in five different places, asterisk (*) mean significative differences (P<0.05).
Figure 2.
Mean (±SE) of Cephalonomia stephanoderis collected in dry and mature coffee berries in five different places, asterisk (*) mean significative differences (P<0.05).

Figure 3.
Total individuals of Cephalonomia stephanoderis collected through the year from February 2022 to January 2023 per location.
Figure 3.
Total individuals of Cephalonomia stephanoderis collected through the year from February 2022 to January 2023 per location.

Figure 4.
Mean (±SE) of the total of Cephalonomia stephanoderis collected through the year in five locations and temperature (line with dots), relative humidity (line with triangles) per month during the year. Different letters mean significant differences (P<0.0001).
Figure 4.
Mean (±SE) of the total of Cephalonomia stephanoderis collected through the year in five locations and temperature (line with dots), relative humidity (line with triangles) per month during the year. Different letters mean significant differences (P<0.0001).

Figure 5.
Correlation between the total number of Cephalonomia stephanoderis collected from February to August 2022 and temperature (°C).
Figure 5.
Correlation between the total number of Cephalonomia stephanoderis collected from February to August 2022 and temperature (°C).

Figure 6.
Percentage of parasitism of Cephalonomia stephanoderis collected through the year from January to April 2023 per location.
Figure 6.
Percentage of parasitism of Cephalonomia stephanoderis collected through the year from January to April 2023 per location.


Table 1.
Coefficient of correlation and determination to climatic variables compared against the number of Cephalonomia stephanoderis individuals from February to August 2022.
Table 1.
Coefficient of correlation and determination to climatic variables compared against the number of Cephalonomia stephanoderis individuals from February to August 2022.
| Climatic variables | Correlation coefficient (r) | Determination coefficient (r2) | Pvalue1 |
| Temperature | -0.84 | 0.714 | 0.01 |
| Relative Humidity | 0.202 | 0.041 | 0.66 |
| Altitude | 0.07 | 0.005 | 0.73 |
1 P values of <0.05 are significantly different.
Table 2.
Percentage of parasitism per month during the year 2023, the minimum, maximum, and average number of CBB found in each berry collected, and a total of CBB collected.
Table 2.
Percentage of parasitism per month during the year 2023, the minimum, maximum, and average number of CBB found in each berry collected, and a total of CBB collected.
| Month |
% parasitism |
Total number of Cs collected | Average number of Cs collected per site | Total number of CBB | Max. CBB per coffee berry | Average of CBB per coffee berry |
| January | 0 | 0 | 0 | 1393 | 47 | 5.5 |
| February | 3.49 | 92 | 18.4 | 2538 | 65 | 10.5 |
| March | 2.79 | 98 | 19.6 | 3404 | 87 | 13.6 |
| April | 4.97 | 168 | 33.6 | 3207 | 57 | 12.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



