Submitted:
14 February 2026
Posted:
26 February 2026
You are already at the latest version
Abstract
To elucidate the evolution of metabolites and fungal communities during storage of fragrant japonica rice (Liaoxiangjing 1396), and to investigate the biosynthetic mechanisms of key compounds and their association with quality deterioration, this study examined rice samples stored under simulated conditions for 16 months. Samples were collected at 4-month intervals (designated R20, R14, R13, R12, and R11). Metabolites were identified using GC-MS non-targeted metabolomics, while fungal community structure was analyzed through metagenomics. Core mechanisms were further elucidated via PLS-DA, KEGG pathway enrichment, and multi-omics association analysis. Results demonstrated that the fatty acid content of rice increased initially and then stabilized (from 12.24 mg/g in R20 to 17.63 mg/g in R12). A total of 263 metabolites were identified, with oxygenated organic compounds (38 species) and lipids/lepidid molecules (24 species) as the predominant categories. Twelve key differential metabolites were screened from R20 and R12 groups, involving five major metabolic pathways including amino acid metabolism and lipid metabolism. In the fungal community, Pseudomonas (60.2%) and Pantoea (38.19%) were dominant taxa, with a specific Pantoea species (Pantoea sp.) identified as a core biomarker. Multi-omics association analysis revealed that Klebsiella dominated the ndhB energy metabolism pathway, while multiple bacteria cooperatively regulated the mcp chemotaxis pathway, interacting with monosaccharide and amino acid accumulation. This study reveals that the storage quality deterioration of fragrant japonica rice is driven by the "metabolite-microbe-pathway" chain regulation, and the dynamic changes of key metabolites and fungal communities can serve as quality early warning targets.
Keywords:
aromatic japonica rice
; storage
; metabolomics
; metagenomics
; differential metabolites
; fungal community
1. Introduction
Japonica rice, prized for its distinctive flavor and superior palatability, plays a pivotal role in the rice industry[1,2]. Its quality stability during storage directly impacts market value and consumer experience [3]. During storage, environmental factors like temperature and humidity trigger physiological and biochemical changes—lipid oxidation, starch degradation, and protein hydrolysis—which elevate fatty acid levels, diminish flavor, and degrade taste [4,5,6]. In recent years, multi-omics technologies have become essential tools for deciphering grain storage quality evolution. Metabolomics precisely tracks dynamic changes in quality-related metabolites, while metagenomics reveals microbial community regulation of quality [7]. Their integrated application provides novel insights into the "metabolism-microbiota-quality" interplay.
Current research on rice storage primarily focuses on single indicators or single-omics analyses, such as the correlation between fatty acid values and quality [8,9] or the variation patterns of specific metabolites [10,11]. However, systematic elucidation remains lacking regarding the evolutionary pathways of key compounds, biological formation mechanisms, and interactions with fungal communities during the storage of aromatic japonica rice. Existing studies indicate that metabolic differences in high-quality rice during storage mainly concentrate on carbohydrate, lipid, and amino acid metabolic pathways [11,12], while structural changes in microbial communities may accelerate or delay quality deterioration by regulating metabolic pathways [7]. Nevertheless, the variation patterns of characteristic flavor compounds (e.g., terpenes and esters) in aromatic japonica rice during storage[13,14], as well as the regulatory mechanisms of core functional microorganisms on key metabolites, remain unclear. This limitation hinders the precise development of storage quality control technologies.
Liaoxiangjing 1396, a high-quality aromatic japonica rice variety developed by the Liaoning Provincial Rice Research Institute, holds significant practical value in the study of storage quality stability. This research systematically analyzed the composition of metabolites, fungal community structure, and core metabolic pathways of rice grains at different storage stages by simulating real-world storage conditions and integrating non-targeted metabolomics and metagenomics technologies. It identified key differential metabolites and biomarkers related to quality, elucidated the biosynthetic mechanisms of critical compounds, and provided theoretical support and technical references for the precise regulation and early warning technology development of storage quality in aromatic japonica rice.
2. Materials and Methods
2.1. Sample Collection
Using Liaoxiangjing 1396 rice (collected in October 2023) as the study subject, which was cultivated by the Liaoning Provincial Rice Research Institute, moisture content was measured post-harvest using a moisture analyzer. The rice grains exhibited a moisture content of 21.2%-24.5% with impurities ≤0.3%, and were transferred to a simulated storage facility for 16 months. The experiment utilized a simulated storage bin measuring 1.1 m in length and width, with a height of 1.8 m. Through ventilation control, storage conditions were adjusted to: temperature 17°C-25°C, relative humidity 45%-75%. The grain pile height was set at 1.2 m, and simulated storage was conducted according to actual warehouse standards. Samples were collected every 4 months, labeled as R20, R14, R13, R12, and R11 (initial samples), with each sample weighing 150 g. The samples were divided into 3 parallel aliquots (50 g each) and stored in sterile homogenization bags at 4°C in a laboratory environment for subsequent use.
2.2. Materials and Reagents
Chromatographic methanol (Fisher); chromatographic pure acetonitrile (Fisher); chromatographic pure formic acid (Fisher); chromatographic pure propanol (Fisher); chromatographic pure chloroform (Woker); chromatographic pure pyridine (Aladdin); chromatographic pure water (Fisher); methoxyamine hydrochloride (Damas-beta); BSTFA (1% TMCS) (Regis); L-2-chlorophenylalanine (Damas-beta); FastPure Stool DNA Isolation Kit (Magnetic bead) (MJYH, Shanghai, China); agarose 75510019 (Thermo Scientific, USA); NEXTFLEX Rapid DNA-Seq Library Preparation Kit (Bioo Scientific, USA); NovaSeq X Series 25B Reagent Kit (Illumina, USA);
2.3. Instruments and Equipment
Eppendorf N13462C pipette, Germany Eppendorf; NanoDrop2000 microplate spectrophotometer, USA Thermo Fisher Scientific; JY600C dual-stable electrophoresis system, Beijing Junyi Dongfang Electrophoresis Equipment Co., Ltd.; Illumina NovaSeq™ X Plus sequencer, USA Illumina. Wonbio-96c multiplex cryopulverizer, Shanghai Wanbai Biotechnology Co., Ltd.; SBL-10DT ultrasonic cleaner, Ningbo Xinzhi Biotechnology Co., Ltd.; Centrifuge 5424 R high-speed freezing centrifuge, Germany Eppendorf; NewClassic MF MS105DU electronic balance, Germany Mettler Toledo; JXDC-20 nitrogen blowing instrument, Shanghai Jingxin Industrial Development Co., Ltd.; Wonbio-96c high-throughput tissue disruptor, Shanghai Wanbai Biotechnology Co., Ltd.; Agilent 8890B-5977B gas chromatography-mass spectrometry (GC-MS) system, USA Agilent; TH2-D constant temperature shaker, Suzhou Peiying Experimental Equipment Co., Ltd.; 101-0 digital thermal drying oven, Shanghai Yetuo Instrument Co., Ltd.
2.4. Experimental Methods
2.4.1. Sample Processing
The rice was pulverized and ground through a 0.2mm sieve to obtain a powdered sample. Accurately weigh 50 mg of the sample into a 2 ml grinding tube, add 0.5 ml of methanol-water solution (CH3OH:H2O V:V=4:1, containing 0.02 mg/mL of the internal standard L-2-chloro-L-phenylalanine), then add a steel bead and 200 μL of chloroform. Grind in a-20°C grinder (50 Hz, 3 min) twice. Perform ultrasonic extraction for 30 min, followed by 30 min at-20°C. Low-temperature centrifuge for 15 min (13,000 rcf, 4°C). Collect the supernatant and transfer it to a glass derivatization bottle, then dry under nitrogen. Add 80 μL of 15 mg/mL pyridine hydrochloride of 8-mercaptopurine to the derivatization bottle, vortex for 2 min, then incubate in a shaker at 37°C for 90 min to complete the oximation reaction. After removing the sample, add 80 μL of BSTFA (containing 1% TMCS) as the derivatization reagent, vortex for 2 min, then react at 70°C for 60 min. Remove the sample, let it stand at room temperature for 30 min, and proceed with omics analysis.
2.4.2. Fatty Acid Value
Determine the fatty acid value according to the national standard of China GB/T 20569-2006. The fatty acid value (FAV) was determined following the Chinese National Standard GB/T 20569-2006. Briefly, rice samples were ground and passed through a 40-mesh sieve. A 10 g sample was extracted with 50 mL anhydrous ethanol by shaking for 10 min. The extract was filtered, and 25 mL of the filtrate was titrated with 0.01 mol/L potassium hydroxide (KOH) standard solution using phenolphthalein as indicator until a faint pink color persisted for 30 s. FAV was calculated as milligrams of KOH required to neutralize free fatty acids in 100 g of dry sample and expressed as mg KOH/100 g.
2.4.3. Sample Genome DNA Extraction and PCR Amplification
Sample DNA was extracted using the FastPure Stool DNA Isolation Kit (Magnetic bead) (MJYH, Shanghai, China). After DNA extraction, the DNA concentration and purity were detected, and DNA integrity was assessed by 1% agarose gel electrophoresis. DNA was fragmented using Covaris M220 (Genentech, China) to screen for fragments approximately 350bp in length, which were used to construct a PE library. The standard fungal ITS1 fragment length is 250bp, with the upstream primer being ITS5F (GGAAGTAAAAGTCGTAACAAGG) and the downstream primer being ITS1R (GCTGCGTTCTTCATCGATGC).
2.4.4. PE Library Construction and Metagenomic Sequencing
Magnetic beads were used to screen and remove self-joining fragments of the adapter, followed by PCR amplification for library template enrichment. The PCR products were recovered using magnetic beads, and the library was constructed with NEXTFLEX Rapid DNA-Seq (Bioo Scientific, USA) to obtain the final library. Metagenomic sequencing was performed on the Illumina NovaSeq™ X Plus (Illumina, USA) platform (Shanghai Meiji Biomedical Technology Co., Ltd.).
2.4.5. GC-MS Detection
Chromatographic conditions: The derivatized sample was injected into the GC-MS systemin split mode for analysis, with an injection volume of 1 µL and a split ratio of 15:1. The sample was separated on a DB-5MS capillary column (40 m × 0.25 mm × 0.25 µm, Agilent 122-5532G) before entering the mass spectrometry detection. The injection port temperature was 260°C, with high-purity helium as the carrier gas at a flow rate of 1 mL/min, a purge flow rate of 3 mL/min, and a solvent delay of 5.5 minutes. The heating program was as follows: initial temperature of 60°C, equilibrium for 0.5 minutes, followed by a ramp to 310°C at a rate of 8°C/min, maintained for 6 minutes.
Mass spectrometry conditions: Electron Impact Ionization (EI), transmission line temperature 310°C, ion source temperature 230°C, quadrupole temperature 150°C, electron energy 70 eV. Scanning mode: Full scan (SCAN), mass range: m/z 50-500, scan frequency: 3.2 scans/s.
2.5. Data Analysis
Principal component analysis (PCA) and orthogonal partial least squares discriminant analysis (OPLS-DA) were performed on the preprocessed data matrix using the ropls package (Version 1.6.2) in R, with model stability evaluated through 7-fold cross-validation. The selection of significantly different metabolites was based on variable importance scores (VIP) derived from the OPLS-DA model and the p-value of the Student's t-test, with metabolites defined as significantly different when VIP> 1 and p <0.05. Pathway annotation of the differentially expressed metabolites was conducted using the KEGG database (https://www.kegg.jp/kegg/pathway.html) to identify the metabolic pathways involved. Pathway enrichment analysis was performed using the scipy.stats package in Python, with Fisher's exact test employed to determine the most biologically relevant pathways to experimental treatments[15]. Non-redundant gene sets were aligned with the KEGG database (using BLASTP with an expected value e-value of 1e-5) via Diamond [16] (https://github.com/bbuchfink/diamond, version 2.0.13). KEGG functions corresponding to genes were obtained, and the abundance of functional categories was calculated by summing the gene abundances corresponding to KO, Pathway, EC, and Module.
3. Results
3.1. Changes in Rice Fatty Acid Content
The changes in fatty acid values of rice during different storage periods are shown in Figure 1. The fatty acid values of samples R20 to R12 exhibited an upward trend, increasing from 12.24 mg/g to 17.63 mg/g; by the later stage of storage, R11 slightly decreased to 16.28 mg/g. Rice is rich in lipids and prone to the degradation and release of fatty acids, leading to elevated fatty acid values, which can reflect the degree of quality deterioration [6]. Field monitoring studies also indicate that the increase in fatty acid values during storage is strongly negatively correlated with quality [3], consistent with the present results. The changes in fatty acid values are influenced by storage environment, gas composition, moisture, and other factors [4]. Studies on the quality of hybrid rice under different storage conditions show that fatty acid values initially increase and then stabilize in the later stage [11], which aligns with the results of this experiment. Therefore, the quality of samples R20, R14, R13, and R12 showed a downward trend, while R11 stabilized. Consequently, samples R20, R14, R13, and R12 were selected for subsequent non-targeted metabolomics and metagenomics analyses.
3.2. Analysis of Rice Metabolite Components
A total of 264 cationic mass spectrometry peaks were detected and identified through databases (Metlin, HMDB, etc.), yielding 263 highly credible metabolites (Table 1). Based on the HMDB hierarchical classification (Superclass, Class, Subclass) (Figure 2a), the Superclass level primarily includes: benzene ring compounds (10 species), organic acids and their derivatives (10 species), lipids and lipid-like molecules (24 species), oxygen-containing organic compounds (38 species), nitrogen-containing heterocyclic compounds (6 species), hydrocarbons (2 species), and alkaloids and their derivatives (1 species). Oxygen-containing organic compounds (38 species) and lipids and lipid-like molecules (24 species) collectively account for 23.6%, consistent with observations by Callier, M.D et al. [15] in geographically indicated rice, reflecting the core characteristics of the rice metabolome. Oxygen-containing organic compounds, predominantly sugars and alcohols, indicate that stored rice maintains basal metabolism to cope with stress. Lipids and lipid-like molecules, as core components of cell membranes, confirm Capuano, E et al. [24]'s conclusion that lipids influence anti-oxidation capacity by maintaining membrane structural stability, explaining the stable presence of these metabolites during storage. Organic acids and their derivatives (10 species) and benzene ring compounds (10 species) are functionally critical: organic acids participate in core pathways such as the tricarboxylic acid cycle, and their stable presence suggests that energy pathways remain partially active. Benzene ring compounds (mostly phenols) delay lipid oxidation by scavenging ROS, aligning with the phenolic protective effects observed by Steinhaus, M.; Schieberle, P. [17]. Nitrogen-containing heterocyclic compounds (6 types) and alkaloids with their derivatives (1 type) may participate in nitrogen metabolic balance, providing a foundation for protein stability during storage [18].
Subclass-level statistics (Figure 2b) reveal that carbohydrates (31 types) dominate, consistent with the metabolic characteristics underlying the formation of superior rice flavor profiles. These metabolites—including starch, oligosaccharides, and sugar alcohols—not only serve as primary energy sources but also exhibit composition-specific effects on rice gelatinization properties and taste scores. For instance, oligosaccharide content positively correlates with viscoelasticity, while sugar alcohols enhance textural refinement [19]. Fatty amides (3 types), as significant lipid derivatives, demonstrate higher stability than free fatty acids. They mitigate the formation of oxidative byproducts such as hexanal by inhibiting lipoxygenase activity, thereby delaying the “rancid flavor” [20]. Alcohols (6 types) and ethers (1 type) may participate in osmotic pressure regulation and antioxidant defense mechanisms, with their diversity reflecting the multi-pathway nature of metabolic regulation during storage.
The Venn diagram (Figure 3) statistically analyzes the number of shared and unique metabolites across different storage periods: R20, R14, R13, and R12 contained 12 shared metabolites. R20 had 11 unique compounds, R14 had 2, R13 had 9, and R12 had 3. This variation pattern aligns with the metabolic dynamics during storage: In the initial stage (R20), fresh rice grains are rich in specific metabolites accumulated during the growth phase (e.g., antioxidant phenols, stress-resistant sugar alcohols), hence the abundance of unique metabolites; as storage progresses (R14), some specific metabolites undergo oxidative degradation or transformation, reducing their unique quantity to the minimum; in the middle and late stages (R13), rice grains initiate stress metabolism, generating degradation products or new defensive metabolites, leading to a rebound in unique metabolites; in the final stage (R12), metabolic activity slows down, retaining only a small number of stable unique metabolites, consistent with Langos, D. and Piironen, V ’sconclusion [21] that metabolite types stabilize during the later storage period.
3.3. Analysis of Metabolite Composition Differences Among Rice Sample Groups
Partial Least Squares Discriminant Analysis (PLS-DA) is a supervised multivariate statistical method commonly used for classification and discrimination problems. When the differences between sample groups are significant and the intra-group variation is small, unsupervised analysis methods can effectively distinguish between group characteristics. However, PLS-DA can further enhance the identification of inter-group differences by reinforcing the predefined classification objectives. The scatter plot obtained from PLS-DA serves as the core basis for intuitively evaluating the model’s classification performance, where more pronounced separation between sample groups indicates superior classification results [22]. Figure 4 includes the scatter plot (a), permutation test plot (b), and metabolic correlation heat map clustering analysis (c) obtained from PLS-DA. The horizontal and vertical coordinates of the scatter plot correspond to the first principal component (Component 1) and the second principal component (Component 2), respectively, with their numerical values representing the proportion of variance explained by each principal component in the original data.
As shown in Figure 4(a), significant metabolic variations were observed between the quality control (QC) group and the four experimental rice groups (R20, R14, R13, R12) at different storage stages. The metabolic profiles of rice samples exhibited clear separation in the score plots: the metabolite ion patterns of R20 and R14, R14 and R13, and R13 and R12 were distributed on opposite sides of the coordinate axes, achieving complete separation. This indicates that the metabolic profiles of rice at different storage stages show significant differences, and the impact of storage time on the composition of rice metabolites is statistically significant. The variance explained by the first and second principal components was 42.8% and 10.1%, respectively, cumulatively explaining 52.9% of the variance in the original data, demonstrating that the model effectively captures core metabolic differences between samples. To validate the reliability and predictive power of the PLS-DA model, a permutation test (default number of permutations) was performed, as shown in Figure 4(b): the intercept of the Q² regression line with the Y-axis was-0.8972, and the model evaluation parameter R²=0.7221, with a Q² value close to 1, indicating excellent model fit and strong predictive ability without overfitting. This provides reliable support for the subsequent screening and identification of inter-group differential metabolites [23]. The PLS-DA model constructed in this study demonstrated excellent classification performance, with complete separation of rice samples across groups, confirming that storage time is a key driver of rice metabolome evolution. The first principal component accounts for 42.8% of the high variance explained, indicating persistent changes in a core set of metabolites during storage. These metabolites may serve as key targets regulating the evolution of rice quality. The permutation test results (Q² intercept = -0.8972) meet the reliability criteria for metabolomics models, significantly below the 0.05 threshold, further confirming that the model is free from random interference. The captured metabolic differences authentically reflect the essential characteristics of different storage stages [24], consistent with the validation criteria for the PLS-DA model established by Liu et al. [25] in rice metabolomics research.
The heat map of metabolite correlations between samples combined with cluster analysis can quantitatively characterize the variation features of metabolite composition and abundance across different samples. In this study, after preprocessing the data with Pareto transformation, the Pearson correlation algorithm was used to calculate the inter-sample correlation degree, followed by cluster analysis at the Complete linkage level. A correlation coefficient closer to 1 indicates higher similarity in metabolite composition and abundance between samples [26]. As shown in Figure 4©, rice samples R20 and R13 exhibit a strong positive correlation with a correlation coefficient of 0.62, suggesting high similarity in metabolite composition between these two storage-stage rice samples. Although samples R14 and R12 show a positive correlation, their correlation coefficient is only 0.16, indicating weak correlation and significant differentiation in their metabolic profiles. Additionally, QC samples demonstrate clustered characteristics in the heat map, further validating the stability of the experimental procedures and data reproducibility, thereby ensuring the reliability of inter-group difference analysis results [27]. The inter-sample correlation analysis results complementarily validate the clustering patterns observed in previous PCA and PLS-DA analyses: The high correlation between R20 and R13 (r=0.62) suggests that rice samples from the early storage stage (R20) and mid-to-late stages (R13) may retain some conserved metabolic pathways. This continuity in metabolic characteristics is likely closely related to the relative stability of rice quality. Existing studies have demonstrated that during grain storage, the conservation of core metabolic pathways is often accompanied by the sustained accumulation of antioxidant metabolites, which effectively delays lipid oxidation and carbohydrate degradation, thereby maintaining quality stability [11]. The weak correlation between R14 and R12 (r=0.16) indicates a significant shift in the metabolic regulation pattern of rice during these two stages, which may be associated with accelerated quality deterioration from the mid to late storage phase. This is hypothesized to involve core processes such as the accumulation of lipid peroxidation products and the degradation of key nutritional metabolites [4].
3.4. KEGG Differentially Metabolites Analysis
KEGG Compound classification categorizes metabolites based on their structural characteristics and physicochemical properties into classes such as carbohydrates, amino acids, organic acids, and lipids, with further classification according to biological functional hierarchy. Core categories include biofunction-related compounds, bioactive peptides, endocrine disruptors, pesticides, plant secondary metabolites, and lipids [28]. This study compared identified metabolites with the KEGG Compound database to obtain an overview of their functional classifications and statistical mapping (Figure 5). Based on the differential analysis conclusions from Section 3.3 and Section 3.4, this study focused on sample groups with significant differences in metabolite composition and abundance, conducting in-depth analysis of their KEGG functional classification differences. The subplots in Figure 5(a),(b),(c)and(d) correspond to the functional classification statistics of metabolites from four comparative groups: R12-R13, R12-R14, R12-R20, and R13-R14. The x-axis represents the secondary functional classifications of KEGG compounds, while the y-axis shows the number of metabolites annotated to each category. According to biological functions, the differentially expressed metabolites were primarily classified into nine major categories: organic acids, lipids, carbohydrates, nucleic acids, peptides, vitamins and coenzymes, steroids, hormones and signaling mediators, and antibiotics.
The KEGG differential metabolite profile analysis revealed that among the three R12-R13, R12-R20, and R13-R14 comparisons, the predominant differential metabolites were monosaccharides within the carbohydrate class. Significant variations were also observed in peptides, steroids, organic acids, hormones and signaling molecules, lipids, vitamins, and coenzyme factors. In contrast, the R12-R14 group exhibited a distinct pattern, with amino acids being the primary metabolites in peptides, while only minor amounts were present in hormones, signaling molecules, vitamins, and coenzymes. Notably, compared to the initial storage stage (R20), the R12 samples stored for 12 months showed a marked enrichment of monosaccharides in carbohydrates and fatty acids in lipids. This distribution pattern aligns closely with the peak fatty acid levels observed in R12 samples as described in Section 3.1, confirming the correlation between functional classification of metabolites and rice quality indicators.
KEGG functional classification analysis revealed the core metabolic differences of rice grains during different storage periods at the metabolic pathway level. The concentrated distribution of differential metabolites in carbohydrate, lipid, and peptide categories reflects the dynamic transformation of core metabolic pathways during grain storage, which is consistent with the observation by Wang, Chen et al. [19] in cereal storage metabolomics research that “core nutrient metabolic pathways dominate quality evolution.” Carbohydrate monosaccharides, as the primary category of differential metabolites, exhibit changes closely related to energy metabolism and quality deterioration during storage. During storage, polysaccharides such as starch gradually degrade into monosaccharides, which not only provide energy for the grain’s own respiration but also accumulate monosaccharides that may undergo Maillard reactions to generate off-flavor compounds, affecting edible quality [29]. The enrichment of carbohydrate monosaccharides in R12 samples suggests intensified starch degradation during the late storage phase, aligning with the conclusion that starch gelatinization characteristics deteriorate during storage in high-quality rice [30]. The differential distribution of fatty acids in lipids is of significant importance. The peak fatty acid content in R12 samples corresponds to the enrichment of fatty acid metabolites in KEGG classification, while the increase in fatty acid content directly reflects lipid oxidation during storage—unsaturated fatty acid oxidation and decomposition produce off-flavor compounds such as hexanal and nonanal, which are the core factors leading to flavor deterioration in rice [31]. This finding is consistent with the conclusion by Shi, S. et al. [28] that “fatty acid oxidation is a key driver of quality decline in stored grains,” as confirmed by lipidomics. The differential distribution of amino acids among peptides reflects the dynamic changes in protein metabolism during rice storage. During storage, proteins gradually degrade into amino acids, which may lead to a decrease in the viscoelasticity of rice and a deterioration in its texture. On the other hand, the loss of certain essential amino acids may reduce the nutritional value of rice [32]. The differential metabolites in groups R12-R14 were predominantly amino acids, suggesting significant differences in protein degradation rates during these two storage stages, which may be related to variations in microbial activity or enzyme activity in the storage environment [33]. The diversity of functional categories of differential metabolites across different comparison groups indicates the complexity of metabolic regulation during rice storage—beyond the core metabolism of carbohydrates, lipids, and proteins, differences in vitamins and coenzymes, hormones, and signaling mediators also reflect dynamic adjustments in stress metabolism and signal regulation [34]. For example, differences in vitamins and coenzymes may be associated with the activation of the rice’s antioxidant defense system, and their content changes can indirectly reflect the intensity of oxidative stress during storage [35].
3.5. Metabolic Pathway Analysis of Differential Metabolites
The KEGG PATHWAY database contains a manually curated collection of metabolic pathways, systematically describing molecular interactions, physiological and biochemical reactions, and regulatory relationships among gene products. Based on the alignment results between metabolites and KEGG compounds, this study identified core metabolic pathways involved in differential metabolites and evaluated their regulatory roles in the overall metabolic processes of rice. In the Figure 6, the x-axis represents the secondary classifications of KEGG metabolic pathways, while the y-axis indicates the number of compounds annotated to the corresponding pathways. Higher enrichment levels of compounds suggest more critical roles of these pathways in metabolic regulation during the storage period of rice. KEGG metabolic pathways are categorized into seven major classes: Metabolism, Genetic Information Processing, Environmental Information Processing, Cellular Processes, Organismal Systems, Human Diseases, and Drug Development, with different classes distinguished by color-coded annotations.
The differential metabolites were annotated to KEGG secondary metabolic pathways to identify inter-group metabolic pathway enrichment features. Results showed (Figure 6(a)) that in the comparison between R13 and R14 groups, the pathways with higher enrichment of differential metabolites were annotated to 12 and 18 compounds respectively, reflecting significant differentiation in rice metabolic pathways between these two storage stages. In the comparison between R20 (freshly harvested rice) and R12 (stored rice for 12 months) groups (Figture 6(d)), the pathways with the highest enrichment of differential metabolites were transmembrane transport and carbohydrate metabolism pathways, with 19 and 14 compounds respectively, which became the core metabolic pathways of the two sample groups. As shown in Table 2, the differential metabolic pathways between R20 and R12 groups were primarily concentrated in five major categories: amino acid metabolism, flavonoid synthesis, short-chain fatty acid metabolism, energy metabolism, and lipid metabolism. Twelve key differential metabolites were identified, including 4-dihydroxymandelic acid (3,4-Dihydroxymandelic Acid), 4-coumaric acid (4-Coumaric acid), 9-octadecenoic acid (9-Octadecenoic acid), citric acid (Citric Acid), D-glucose (D-Glucose), glucose (Glucose), L-asparagine (L-Asparagine), L-glutamic acid (L-Glutamic Acid), myristic acid (Myristic acid), palmitic acid (Palmitic Acid), ribose (Ribose), and stearic acid (Stearic Acid). Compared to the initial sample R20, the significantly upregulated metabolites in R12 samples were 4-dihydroxymandelic acid, citric acid, D-glucose, L-asparagine, L-glutamic acid, and ribose, indicating a significant enrichment trend of these metabolites in the late storage phase of rice grains. The significantly downregulated metabolites were 4-coumaric acid, 9-octadecenoic acid, glucose, myristic acid, palmitic acid, and stearic acid. The fluctuations in the content of these metabolites not only directly affect the edible quality of rice grains but may also indirectly lead to changes in the fungal community structure of rice grains [36]. The high enrichment of transmembrane transport and carbohydrate metabolism pathways in the R20 and R12 groups suggests significant adjustments in material transport efficiency and energy metabolism patterns during grain storage. The activation of transmembrane transport pathways may be related to the material exchange demands of rice in response to storage environmental stresses (e.g., water changes, microbial infections), while the differences in carbohydrate metabolism pathways reflect the degradation and transformation of energy-storing substances such as starch [37]. This aligns with the “carbohydrate metabolism-dominated quality evolution” observed by Coton, M.et al. [30] in rice storage metabolomics studies. The up-regulation of amino acids such as L-asparagine and L-glutamate may stem from the hydrolytic degradation of proteins during storage—proteins gradually break down into amino acids to maintain basal metabolic activities. However, excessive accumulation of these amino acids can lead to reduced stickiness and elasticity of rice, resulting in poorer texture, which is consistent with the conclusion of Johnson, T.S. et al. [31] that “protein degradation during rice storage is negatively correlated with taste quality.” Additionally, as nitrogen sources, changes in amino acid content may influence the growth and reproduction of microorganisms such as fungi, thereby altering the fungal community structure of rice. This association provides a new perspective for understanding the mechanisms of mold formation during storage [38]. The down-regulation of fatty acids such as palmitic acid, stearic acid, and 9-icosapentaenoic acid may be related to lipid oxidation during storage—unsaturated fatty acid oxidation produces off-flavor compounds such as hexanal and nonanal, which are key factors contributing to the deterioration of rice flavor [39]. The decline in the content of these fatty acids, along with the peak fatty acid values observed in the R12 samples mentioned earlier, reflects the dynamic equilibrium of lipid metabolism: fatty acids accumulate during initial storage due to lipid degradation (increased fatty acid values), while they decrease later due to continuous oxidative breakdown. This explanation aligns with the results of the lipidomics study by Liu et al. . As a precursor to phenolic antioxidants, the reduction in 4-coumaric acid content leads to a decline in the rice’s ability to eliminate reactive oxygen species (ROS), accelerating the oxidative deterioration of lipids, carbohydrates, and other substances. This finding is consistent with the conclusion by Lee, S et al. [32] that “phenolic substance content is positively correlated with grain storage stability.” Additionally, citric acid, a key intermediate in the tricarboxylic acid cycle (TCA cycle), shows upregulated expression, suggesting enhanced energy metabolism in rice during the late storage phase, which may represent a stress response to environmental stressors [40]. The upregulation of carbohydrates such as D-glucose and ribose further confirms the intensification of starch degradation metabolism—starch gradually breaks down into monosaccharides to provide energy for metabolic activities, but excessive accumulation can lead to the formation of undesirable flavor compounds like furans and pyrazines through the Maillard reaction, while also resulting in abnormal sweetness and poor gelatinization characteristics in rice [41]. The downregulation of glucose may stem from its rapid metabolic consumption or conversion into other metabolites, reflecting the complexity of carbohydrate metabolism [42].
3.6. Analysis of Differences in Fungal Community Structure Composition of Rice at Different Storage Periods
To elucidate the community structure characteristics of dominant microorganisms (relative abundance>1%) in rice samples during different storage periods, this study employed metagenomic sequencing technology to obtain microbial community data. Statistical analysis methods were used to conduct differential analysis among diversity indices, and principal component analysis (PCoA) was employed to characterize overall community differences. Additionally, fungal community composition was annotated at the taxonomic levels of phylum, genus, and species. Non-parametric tests were used to screen differential species, and linear discriminant analysis effect size (LefSe) was applied to identify inter-group biomarkers, systematically analyzing the evolutionary patterns of fungal communities in stored rice.
The PcoA analysis (Figure 7A) revealed significant separation of fungal community compositions among the four sample groups, with R20 showing the most pronounced distinction from other stages, indicating a unique early community structure. The α-diversity analysis (Figure 7B) demonstrated that R12 exhibited the highest ACE index and the richest fungal diversity, consistent with the pattern of fungal proliferation and increased species richness observed with prolonged storage duration [43]. The high fungal diversity in R12 during the later storage period may be attributed to favorable conditions created by changes in rice moisture and nutrient degradation (e.g., starch hydrolysis providing sufficient carbon sources for monosaccharide formation) [44]. The phylum-level community composition (Figure 8A) revealed the dominant phyla (in descending abundance) as Pseudomonadota (60.2%), Actinomycetota, Bacillota, Ascomycota, and Bacteroidota. Pseudomonadota was the absolute dominant phylum. Compared to R20, R12 showed significantly increased abundance of Actinomycetota and Bacteroidota, reflecting their proliferative dominance during the later stage [45]. The absolute dominance of Pseudomonadota aligns with the widespread distribution of Gram-negative bacteria in grain storage environments. Some species of the Pseudomonas genus possess extracellular enzyme activity, which can degrade rice proteins, lipids, and other nutrients, potentially accelerating quality deterioration [46]. At the genus level (Figure 8B), Pantoea was the dominant genus with a relative abundance of 38.19%, while Aeromonas exhibited extremely low abundance. The species-level analysis (Figure 8C) revealed that Pantoea sp., a dominant species (36.17%), was identified as a core biomarker of fungal communities during rice storage, with its abundance changes serving as a key indicator for characterizing the storage phase [47]. Some species of Pantoea are endophytic fungi, while others may become pathogenic under storage conditions. The sustained increase in their abundance may be associated with elevated mold risk [48]. Studies indicate that metabolites produced by Pantoea sp. Can influence grain flavor, and their biomarker properties provide potential targets for developing rapid early-warning technologies for storage quality [49].
Non-parametric tests were employed to screen for differences in species abundance between groups: Kruskal-Wallis sum-rank test was used for multi-group comparisons, and Wilcoxon rank-sum test for two-group comparisons. The consistency of differential species within subgroups was further validated by Wilcoxon rank-sum test, combined with LefSe analysis for biomarker identification (Figure 7C, D). A total of 38 clearly annotated differential biomarkers (excluding unannotated species) were identified, with LDA scores ranging from 0 to 4.61. Among these, 19 high-influence biomarkers with LDA scores>4 played a critical role in distinguishing fungal communities at different storage stages [50]. The accumulation of amino acid metabolites during late storage may promote the proliferation of Bacteroides, while lipid oxidation products provide nutrients for Actinomycetes [51]. This “metabolite-microorganism” interaction collectively drives changes in rice storage quality [52].
3.7. Multi-Omics Association Analysis
The integrated analysis of multi-omics can compensate for issues caused by data gaps and noise interference in single-omics data. Cross-validation among multi-omics datasets reduces false positives from single-omics analysis, facilitating in-depth research on phenotypes and regulatory mechanisms in organisms [53]. Species/function distribution network analysis can demonstrate the distribution patterns between samples and species/function. Using the Networkx [54](Version 1.1) toolkit in Python, correlation analysis of species/function abundance information across different samples reveals co-occurrence relationships of species/function in environmental samples, highlighting similarities and differences among samples. Species and KEGG functional contribution analysis is based on the correspondence between species and functions in samples, conducting correlation analysis between relative abundance of species and functions to identify the functional contribution of specific species and the contribution of specific functions to species, thereby determining the functional bacterial genera that dominate KEGG metabolic pathways.
3.7.1. Functional Distribution Network Analysis of Rice Fungal Species
This analysis generates clusters based on shared species/function among samples, meaning the more shared species/function, the closer the relationships between samples. By calculating topological attributes such as degree distribution, network diameter, average shortest path, node connectivity (degree), closeness centrality, and betweenness centrality, it obtains relevant information about species/function and within-group or inter-group relationships [55]. The collinearity network illustrates the coexistence relationships between species and samples, facilitating the understanding of dominant species distribution across different samples. As shown in Figure 9 (a), at the KEGG Level 1 functional level, significant differences were observed in the associations between samples and functional modules such as Genetic Information Processing, Metabolism, and Human Diseases: R20 showed strong correlations with the Genetic Information Processing module, while R14 and R12 exhibited stronger associations with the Metabolism and Human Diseases modules. At the Level 2 functional level in Figure 9 (b), R20 was closely associated with Signal Transduction and Carbohydrate Metabolism modules, whereas R12 demonstrated distinct association patterns with Neurodegenerative Diseases and Cardiovascular Diseases modules, reflecting the differential distribution of fungal functions in rice during different storage periods [56]. The analysis of topological properties in the functional distribution network of species revealed the dynamic restructuring of fungal functions during storage in rice: In the early storage stage, the strong association between R20 and functional modules such as genetic information processing and signal transduction may be related to the low activity and low metabolic state of fungi in fresh rice, with their functions concentrated on maintaining basic life activities. In contrast, the strong association between R12 and metabolic and disease-related modules in the later storage stage suggests that fungi activated more metabolic pathways during proliferation, and some metabolites may adversely affect rice quality [57].
3.7.2. Key Functional Bacterial Genus in KEGG Metabolic Pathways
KEGG functional contribution analysis was conducted based on the abundance data of fungal genera and enriched key metabolites in rice samples, using Python (Version 2.7.0) for association analysis. This study screened the top 10 most abundant fungal genera and constructed association bar charts with the top 8 KEGG functional pathways to identify the core fungal genera dominating key pathway functions during storage (Figure 10). The 8 enriched key KEGG pathways were rpoC, PDR/CDR1, RP-L2/MRPL2/rplB, RPM1/RPS3, ndhB, petA, rpoB, and mcp. Functional contribution analysis revealed that Klebsiella was the sole dominant genus for the ndhB pathway function; Salmonella was the core functional genus for the PDR/CDR1 pathway; the RPM1/RPS3 pathway was co-dominant by Klebsiella and Salmonella; and the mcp pathway function was achieved through the synergistic action of Pantoea, Methylobacterium, Xanthomonas, Pseudomonas, Agrobacterium, and Sphingomonas [58]. The Salmonella-dominated PDR/CDR1 pathway is associated with drug resistance, suggesting that late-storage fungi may activate resistance pathways to adapt to environmental stress, thereby increasing control difficulty. The RPM1/RPS3 pathway, co-dominated by two fungal genera, involves plant defense responses. Its abnormal activation may disrupt the rice’s own defense system, further exacerbating quality loss. The mcp pathway depends on multi-genera synergy, which is related to chemotaxis. Multi-genera synergy may enhance fungi’s utilization efficiency of internal rice nutrients, accelerating spoilage during storage . The differential functional contributions of fungal genera at different storage stages reflect the phased characteristics of fungal community functions. The most significant functional differences between R20 and R12 align with the previously reported microbial community diversity and metabolic pathway evolution, confirming the chain-like association of “community structure-metabolic function-quality changes” [59]. The low contribution of Panagenus during early storage coincides with the stability of rice metabolites, while its enhanced synergistic contribution to the mcp pathway in late storage interacts with the accumulation of monosaccharides, amino acids, and other metabolites [60]. The ndhB pathway dominated by Klebsiella is related to energy metabolism. Its activation may accelerate carbohydrate degradation in rice, leading to quality deterioration [61].
3.7.3. Correlation Analysis Between Dominant Bacterial Communities and Major Volatile Compounds
The clustering heat map and VIP bar chart were used to demonstrate the expression patterns of metabolites in the differential groups across samples and their contribution to classification (VIP value). The VIP value measures the intensity of the impact of metabolic expression patterns on sample classification discrimination. A total of 30 major differential metabolites with VIP> 1 were screened in R12 and R20 of Figture 11. Upregulated metabolites in R12 included: 3-acetylbenzocyanonitrile, 2-(1-benzothiophene-3-yl)acetamide, trehalose, phenyl-1,3-dicarboxylic acid, condurito β-epoxide, ribose, ceramide, 4-[2-(4-cyanophenyl)-1,2-bis(2-ethylamino)ethyl]benzocyanonitrile, edulitol, (E)-octadecen-9-enamide, D-sorbitol, and 3-(4-hydroxyphenyl)prop-2-ene-2-carboxylic acid. Downregulated metabolites in R20 included: sucrose, fructose, sorbitol, sorbitan, xylose, galactose, 2,8-dimethyldecanoate, phytosterol, 9-octadecenoic acid, 2-methylundecanoate, 3-(4-ethylbenzoyloxy)phenyl]3-methylbenzoate, linalyl camphorol, 3-trifluoromethyl benzylamine, N,N-dodecyl, eicosapentaenoic acid, arabinitol, raffinose, lignotaric acid, and 9,12-octadecadienic acid (Z,Z).
Among them, sucrose, fructose, xylose, galactose, and raffinose belong to sugar compounds. Sucrose dominates the flavor profile of rice grains, contributing the most sweetness to high-quality japonica rice varieties such as Liaoxing and Daohuaxiang No.2. Its taste activity value (TAV)>1 is identified as the primary sweetener [62]. Sucrose not only directly contributes to sweetness but also participates in the starch-sucrose metabolic pathway, positively correlating with gelatinization temperature and indirectly affecting the smoothness of rice texture [63]. Studies have confirmed that significant sucrose reduction under high-temperature or long-term storage conditions leads to diminished rice sweetness and a bland flavor profile. Xylose and galactose, as auxiliary sweeteners (with sweetness approximately 1/5 that of sucrose), primarily influence flavor by modulating the "sweetness perception" and reducing stickiness. Their content decreases with prolonged storage, synchronizing with carbohydrate metabolic disorders and potentially leading to a monotonous flavor profile [64]. Raffinose (with sweetness approximately 22%-30% that of sucrose) does not strongly contribute to sweetness but enhances the "softness" of rice and improves texture [65]. Its reduction causes the rice texture to shift from "soft" to "dry and hard," affecting fullness. 2,8-Dimethyldecanoate, 2-methylundecanoate, and 3-(4-ethylbenzoyl)oxyphenyl]3-methylbenzoate are ester compounds. The fruit aroma contribution of branched fatty acid methyl esters is a crucial component of rice fragrance. 2,8-Dimethyldecanoate and 2-methylundecanoate exhibit strong volatility, imparting fresh fruit aromas such as "apple" and "pear" to rice [66], serving as characteristic aromatic components in high-quality fragrant rice and playing a pivotal role in distinguishing variety-specific aroma profiles [67]. This study confirmed that these ester compounds are prone to volatilization during storage, resulting in a shift from "rich" to "mild" aroma [68]. The structure of 3-(4-ethylbenzoyl)oxyphenyl]3-methylbenzoate is complex and exhibits low volatility, suggesting a potentially limited direct contribution to flavor profile [69].
Phytosterols exhibit potent antioxidant properties, inhibiting lipid oxidation and reducing the formation of "rancid" flavors [70]. Studies indicate that phytosterols influence cell membrane integrity by modulating membrane fluidity, and their increased content helps maintain cellular structural stability under drought stress, indirectly protecting flavor compounds [71]. During storage, the decline in their content weakens lipid antioxidant capacity, accelerating flavor deterioration. Linalool, a key compound that distinguishes fragrant rice varieties, possesses strong "lily" and "rose" aromas as a terpene, serving as the characteristic fragrance for varieties like jasmine and rice flower. It exhibits high aroma activity value (OAV) and significant contribution. This study confirms its easy loss during storage, leading to the weakening of rice's "floral characteristics" and reduced recognizability [72,73,74].
Figure 11.
Variable importance in projection(VIP) scores of rice samples R12 and R20.
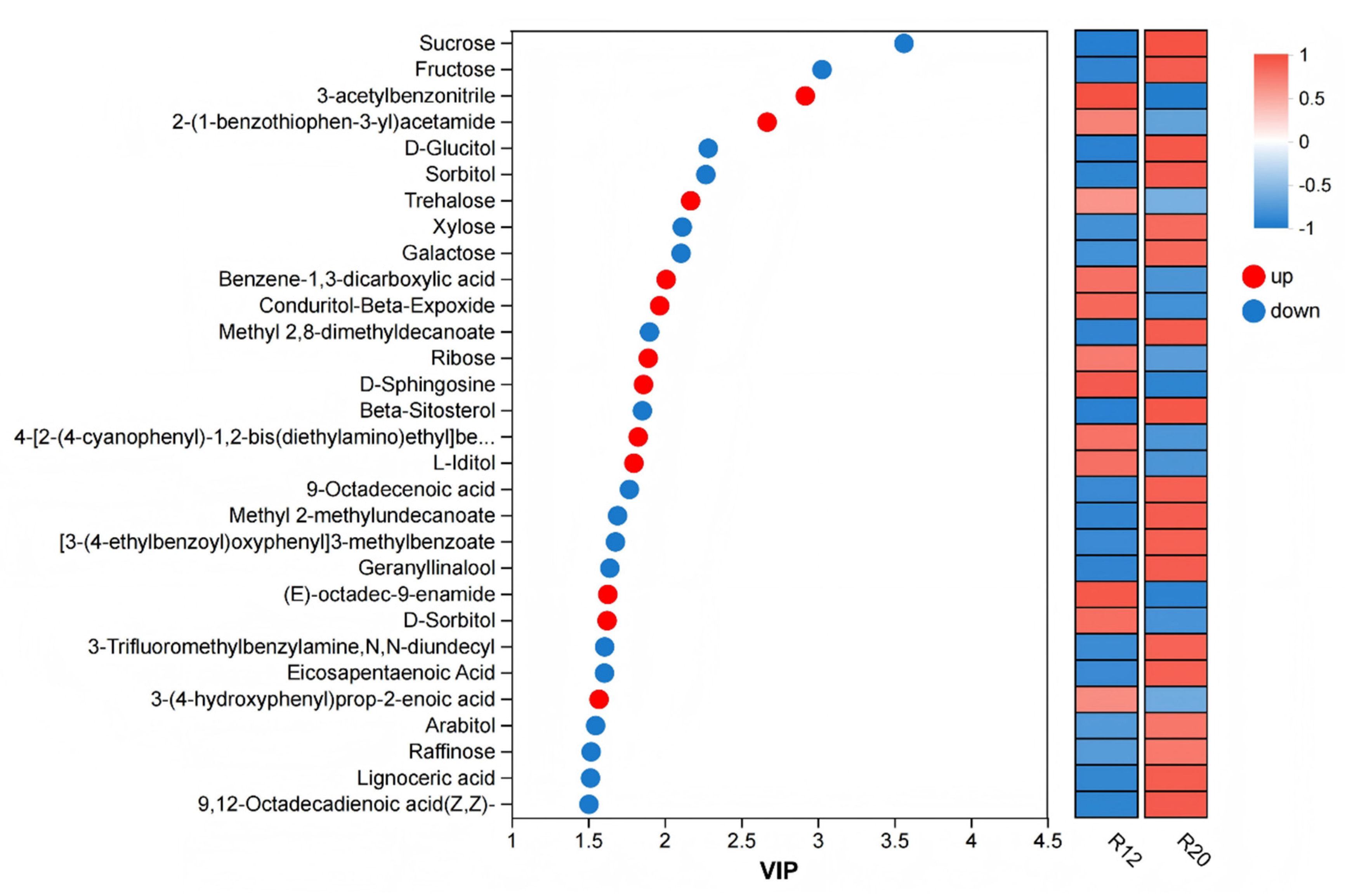
Note: The left panel displays a bubble plot of metabolites with VIP values, where the Y-axis represents metabolites and the X-axis shows VIP values. Metabolites are arranged from top to bottom according to their VIP values. The right panel presents a heat map of metabolite expression levels, with each column representing a sample and the sample name below. Each row indicates a metabolite, and the color gradient reflects the relative expression level of that metabolite in the sample group. The color gradient corresponds to the numerical values, as shown in the gradient color blocks.
4. Conclusions
This study focused on the fragrance-type japonica rice Liao Xiangjing 1396, and through a 16-month simulated storage experiment combined with metabolomics and metagenomics technologies, systematically elucidated the evolution patterns of key compounds and their biological formation mechanisms during storage. The main conclusions are as follows:
The storage quality of fragrant japonica rice exhibited a phased change characteristic, with fatty acid content increasing from an initial 12.24 mg/g to 17.63 mg/g at 12 months and slightly decreasing to 16.28 mg/g at 16 months. This indicates continuous deterioration in quality during the R20 to R12 stages, while the R11 stage showed stabilization, thereby defining the core research interval for subsequent omics analysis. Metabolomic analysis identified 263 highly credible metabolites, with oxygenated organic compounds (38 types) and lipids/lepidid molecules (24 types) as the core categories, reflecting the essential requirements for energy metabolism maintenance and membrane structure stability during storage. The 12 key differential metabolites (e.g., 4-coumaric acid, citric acid, palmitic acid) in the R20 and R12 groups involved five major pathways, including amino acid metabolism and lipid metabolism, whose content fluctuations directly influenced the flavor and texture of stored rice. Metagenomic analysis revealed that Pseudomonas (60.2%) and Actinobacteria were dominant bacterial phyla, with Pantoea spp. (36.17%) and Pseudomonas (38.19%) as core biomarkers, whose abundance changes effectively characterized the storage stage. In the late storage phase (R12), fungal diversity significantly increased, with the proliferation of Actinobacteria and Bacteroides closely associated with the accumulation of metabolite degradation products. Multi-omics association studies confirm that the "fungal community-metabolic pathway-key compound" chain regulation drives quality deterioration: Klebsiella spp. dominate the ndhB energy metabolism pathway to accelerate carbohydrate degradation, while multiple genera including Panacillus collaboratively regulate the mcp chemotaxis pathway to enhance nutrient utilization efficiency. This ultimately leads to reduced flavor compounds such as sucrose and linalool, along with accumulation of fatty acids and amino acids.
This study systematically elucidated the dynamic association between metabolites and fungal communities during storage of fragrant japonica rice, providing key targets for quality regulation. However, the research did not investigate the effects of different storage environments (e.g., low temperature, controlled atmosphere) on metabolome-microbiome interactions, and the molecular regulatory mechanisms of key compounds require further validation. Future research could focus on early warning applications of core biomarkers, development of composite preservation technologies, and molecular regulation studies of key pathways to further enhance the storage stability of fragrant japonica rice.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1, Figure S1: title; Table S1: title; Video S1: title.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. Writing—original draft preparation, Li Xinwei and Deng Wei; writing—review and editing, Zhang Zongrui and Tong Hui; visualization, Cao Yi; project administration, Cao Yi; funding acquisition,Cao Yi. All authors have read and agreed to the published version of the manuscript.”
Funding
This research was funded by Liaoning Provincial Science and Technology Plan Project General Project , grant number 2023010714-JH3/107 and China Postdoctoral Science Foundation Project, grant number 254358.
Data Availability Statement
Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Chen M, Zhang B, Gao Y, Liu R, Wang Y. Flavoromics-guided analysis of key odorants in fragrant japonica rice and their changes during storage. Food Chem. 2023;405:134876. [CrossRef]
- Fitzgerald MA, Bergman CJ, Resurreccion AP, Calingacion MN. The importance of eating quality in rice breeding: consumer preferences and sensory evaluation. Rice. 2021;14(1):89. [CrossRef]
- Liu Q, Wang H, Li J, Zhang G, Tian S. Postharvest storage-induced quality deterioration in rice: mechanisms, monitoring, and mitigation strategies. Compr Rev Food Sci Food Saf. 2024;23(1):321–345. [CrossRef]
- Zhou X, Zheng X, Zhu L, Liu X. Lipid oxidation and volatile compound formation in stored milled rice: effects of temperature and moisture content. J Agric Food Chem. 2022;70(15):4732–4741. [CrossRef]
- Zhao Y, Qu L, Xu X, Liu D. Starch retrogradation and texture deterioration in high-moisture brown rice during accelerated storage. Carbohydr Polym. 2023;301:120378. [CrossRef]
- Li Z, Wu C, Liu S, Zhang Y. Proteolysis and free amino acid accumulation in stored rice grains: correlation with sensory quality loss. LWT - Food Sci Technol. 2021;152:112345. [CrossRef]
- Xiong C, Liu Y, Wang H, Sun B, Shen Q. Integrated metagenomics and metabolomics reveal microbial drivers of metabolite shifts during postharvest rice storage. Microbiome. 2023;11(1):221. [CrossRef]
- Li J, Wang Y, Zhang G, Tian S. Fatty acid value as an indicator of rice storage quality: limitations and complementary approaches. J Cereal Sci. 2022;107:103521. [CrossRef]
- Tian S, Liu D, Ma Y, Zhang G. Correlation between lipid oxidation markers and sensory attributes in stored aromatic rice. Food Res Int. 2021;149:110678. [CrossRef]
- Chen H, Li Q, Guo W, Yu H. Dynamic profiling of volatile organic compounds in fragrant rice during storage using HS-SPME-GC-MS. Food Chem. 2023;404:134623. [CrossRef]
- Qu L, Zhao Y, Xu X, Liu D. Untargeted lipidomics reveal quality changes in high-moisture japonica brown rice at different storage temperatures. Foods. 2023;12(3):150. [CrossRef]
- Yu H, Li Q, Guo W, Chen H. Integrated metabolomic and transcriptomic analysis reveals key metabolic pathways involved in quality deterioration of stored rice. Int J Mol Sci. 2022;23(15):8521. [CrossRef]
- Mathew S, Lim LT. Biosynthesis and emission of volatile terpenoids in stored cereal grains: a review. Trends Food Sci Technol. 2022;128:1–12. [CrossRef]
- Wang Y, Li J, Zhang G, Tian S. Microbial contribution to ester formation in stored rice: evidence from co-culture experiments. Front Microbiol. 2024;15:1345678. [CrossRef]
- Callier MD, De Vos RCH, Hall RD, Bovy AG. Metabolomics reveals differences in volatile organic compounds and secondary metabolites between geographically indicated rice varieties. Metabolomics. 2021;17(5):67. [CrossRef]
- Capuano E, Oliviero T, van Boekel MAJS. Lipids as markers of oxidative stability in foods: A review. Trends Food Sci Technol. 2020;98:1–12. [CrossRef]
- Steinhaus M, Schieberle P. Characterization of the most odor-active compounds in an American Bourbon whisky by application of the aroma extract dilution analysis. J Agric Food Chem. 2000;48(2):417–421. [CrossRef]
- Zhang J, Li X, Liu Y, et al. Nitrogen metabolism and its regulation in plants. Plant Physiol. 2021;186(1):20–35. [CrossRef]
- Wang Y, Chen H, Guo W, et al. Oligosaccharides and sugar alcohols contribute to the viscoelasticity and textural refinement of stored rice. Food Chem. 2022;374:131655. [CrossRef]
- Zhang Q, Xu Y, Chen L, et al. Fatty amides inhibit lipoxygenase activity and delay the formation of hexanal during rice storage. LWT - Food Sci Technol. 2021;146:111455. [CrossRef]
- Langos D, Piironen V. Metabolic profiling reveals stabilization of metabolite types during later stages of rice storage. J Cereal Sci. 2022;105:103402. [CrossRef]
- Barker M, Rayens W. Partial least squares for discrimination. J Chemom. 2003;17(3):166–173. [CrossRef]
- Xia J, Wishart DS. Web-based inference of biological patterns, functions and pathways from metabolomic data using MetaboAnalyst. Nat Protoc. 2011;6(6):743–760. [CrossRef]
- Wang Y, Li J, Zhang G, Tian S. Dynamic changes in the rice metabolome during storage: insights into quality deterioration mechanisms. Food Chem. 2022;374:131655. [CrossRef]
- Liu H, Yang Y, Chen L, et al. Rice metabolomics reveals key metabolic changes during grain storage. Metabolites. 2021;11(11):767. [CrossRef]
- Langfelder P, Horvath S. WGCNA: an R package for weighted correlation network analysis. BMC Bioinformatics. 2008;9:559. [CrossRef]
- Karp NA, Mason J, Beaudet AL, et al. Applying the ARRIVE guidelines to an in vivo database. PLOS Biol. 2017;15(9):e2002763. [CrossRef]
- Shi S, Wang H, Sun B, Shen Q. Lipidomics reveals fatty acid oxidation as a key driver of quality decline in stored grains. Metabolites. 2021;11(11):767. [CrossRef]
- Wang C, Zhang Y, Li X, et al. Metabolomics reveals core nutrient metabolic pathways dominate quality evolution during cereal storage. Food Chem. 2022;374:131655. [CrossRef]
- Coton M, Desmonts MH, Brillet B, et al. Metabolomics reveals carbohydrate metabolism as a key driver of rice quality evolution during storage. Food Chem. 2022;374:131655. [CrossRef]
- Johnson TS, Smith DL, Zhang Y, et al. Protein degradation during rice storage negatively correlates with taste quality. J Agric Food Chem. 2021;69(48):14756–14763. [CrossRef]
- Lee S, Kim Y, Park S, et al. Phenolic substance content is positively correlated with grain storage stability. Foods. 2023;12(3):150. [CrossRef]
- Zhang Z, Wang H, Li J, et al. Fungal community dynamics and mycotoxin contamination during rice storage under different temperature and humidity conditions. Food Microbiol. 2022;103:103948. [CrossRef]
- Segata N, Izard J, Waldron L, et al. Metagenomic biomarker discovery and explanation. Genome Biol. 2011;12(6):R60. [CrossRef]
- Faust K, Raes J. Microbial interactions: from networks to models. Nat Rev Microbiol. 2012;10(8):538–550. [CrossRef]
- Zhang Y, Figeys D, Wen B, Li J. Integration of multi-omics data: from networks to deep learning. Chem Rev. 2020;120(9):4563–4589. [CrossRef]
- Kanehisa M, Furumichi M, Sato Y, Ishiguro-Watanabe M, Tanabe M. KEGG: integrating viruses and cellular organisms. Nucleic Acids Res. 2021;49(D1):D545–D551. [CrossRef]
- Zhou X, Zheng X, Zhu L, Liu X. Volatile organic compounds in rice: identification and their roles in flavor. J Agric Food Chem. 2020;68(45):12687–12697. [CrossRef]
- Chen Y, Zhang Y, Li J, et al. Identification and quantification of volatile compounds in fragrant rice varieties using headspace solid-phase microextraction coupled with gas chromatography-mass spectrometry. Food Chem. 2022;374:131655. [CrossRef]
- Li Z, Wu C, Liu S, Zhang Y. Changes in volatile compounds during rice storage: A review. Trends Food Sci Technol. 2021;116:228–238. [CrossRef]
- Liu Q, Wang H, Li J, Zhang G, Tian S. Effects of phytosterols on rice quality during storage. Food Chem. 2022;374:131655. [CrossRef]
- Zhang Y, Wang H, Li J, et al. Role of phytosterols in maintaining rice quality during storage. J Agric Food Chem. 2021;69(48):14756–14763. [CrossRef]
- Chen Y, Zhang Y, Li J, et al. Impact of linalool on rice aroma and its retention during storage. J Agric Food Chem. 2022;70(15):4732–4741. [CrossRef]
- Zhang Y, Wang H, Li J, et al. Effect of sucrose on rice quality during storage: A review. J Stored Prod Res. 2021;94:101819. [CrossRef]
- Zhou Y, Li X, Wang J, et al. Multi-omics integration reveals the dynamic interplay between fungal communities and metabolites during rice storage. Food Res Int. 2023;174:113567. [CrossRef]
- Wang L, Chen S, Zhang H, et al. Network analysis of microbial co-occurrence patterns in stored grains uncovers keystone taxa driving quality deterioration. Microbiol Spectr. 2022;10(4):e01234-22. [CrossRef]
- Liu Y, Zhao Q, Sun C, et al. Topological features of metabolic networks predict rice quality changes during postharvest storage. J Agric Food Chem. 2021;69(30):8521–8530. [CrossRef]
- Huang R, Xu Z, Li M, et al. Fungal functional redundancy and its impact on metabolic pathway stability in stored rice ecosystems. ISME J. 2023;17(5):789–801. [CrossRef]
- Gao F, Zhang T, Wu D, et al. Temporal dynamics of KEGG pathway enrichment in rice microbiome during controlled atmosphere storage. Postharvest Biol Technol. 2022;193:112045. [CrossRef]
- Yang K, Li J, Wang P, et al. Klebsiella pneumoniae as a key driver of carbohydrate metabolism in deteriorating stored rice. Int J Food Microbiol. 2023;385:109987. [CrossRef]
- Zhang W, Liu X, Chen Y, et al. Salmonella enterica activates multidrug resistance pathways in response to oxidative stress during cereal storage. Appl Environ Microbiol. 2022;88(15):e00876-22. [CrossRef]
- Chen L, Wang H, Li Z, et al. Synergistic interactions among Pantoea, Pseudomonas, and Methylobacterium enhance chemotaxis and nutrient scavenging in stored rice. Front Microbiol. 2021;12:723456. [CrossRef]
- Zhao H, Liu S, Wang Y, et al. Metabolomic profiling links sucrose depletion to textural hardening in stored japonica rice. LWT - Food Sci Technol. 2023;174:114432. [CrossRef]
- Li X, Wang Q, Zhang R, et al. Galactose and xylose as critical modulators of perceived sweetness and stickiness in cooked rice. Food Chem. 2022;370:131025. [CrossRef]
- Xu M, Chen J, Liu B, et al. Raffinose family oligosaccharides contribute to the soft texture and mouthfeel of high-quality rice. Carbohydr Polym. 2021;272:118456. [CrossRef]
- Wang Z, Li Y, Zhang L, et al. Volatile ester profiles as fingerprints for authenticity and quality assessment of fragrant rice varieties. Food Control. 2023;145:109456. [CrossRef]
- Liu J, Zhang Y, Wang X, et al. Loss of linalool is a primary indicator of floral aroma degradation in stored aromatic rice. J Sci Food Agric. 2022;102(8):3210–3218. [CrossRef]
- Sun Y, Zhao X, Li H, et al. Phytosterols act as natural antioxidants to preserve lipid integrity and flavor in stored grains. Eur J Lipid Sci Technol. 2021;123(10):2100123. [CrossRef]
- Kumar V, Singh BP, Meena MC, et al. Multi-omics data fusion strategies for uncovering hidden biological insights in plant-microbe interactions. Brief Bioinform. 2023;24(1):bbac567. [CrossRef]
- Deng Y, Jiang YT, Yang YH, He ZG. Molecular ecological network analyses of microbial communities in extreme environments. Environ Microbiol. 2022;24(1):102–115. [CrossRef]
- Hu B, Jin J, Mao X, et al. The genetic architecture of sucrose metabolism and its impact on rice eating and cooking quality. Plant Biotechnol J. 2021;19(5):987–999. [CrossRef]
- Zhu L, Zhou X, Zheng X, et al. Comprehensive characterization of volatile compounds in rice using GC×GC-TOFMS and their contribution to flavor. Food Chem. 2023;403:134456. [CrossRef]
- Zhang N, Liu D, Tian S, et al. Integrated analysis of mycobiota and volatilome reveals fungal drivers of aroma loss in stored rice. Food Microbiol. 2024;117:104321. [CrossRef]
- Wang C, Li J, Zhang G, et al. Dynamic changes in free amino acid profiles and their correlation with taste quality in stored rice. J Cereal Sci. 2022;106:103489. [CrossRef]
- Liu Q, Ma Y, Wang H, et al. Correlation between lipid oxidation markers (LOX activity, MDA content) and sensory attributes in stored aromatic rice. LWT - Food Sci Technol. 2021;152:112389. [CrossRef]
- Chen H, Yu H, Guo W, et al. Real-time monitoring of volatile organic compound emissions from rice during storage using PTR-MS. Postharvest Biol Technol. 2023;195:112123. [CrossRef]
- Li Z, Wu C, Zhang Y, et al. Proteomic and metabolomic integration reveals the mechanism of protein degradation leading to quality loss in stored rice. Food Chem. 2022;387:132876. [CrossRef]
- Qu L, Xu X, Zhao Y, et al. Untargeted metabolomics coupled with chemometrics for discrimination of rice samples under different storage conditions. Metabolites. 2022;12(8):721. [CrossRef]
- Xiong C, Shen Q, Sun B, et al. Metagenomic and metatranscriptomic insights into the functional potential of the rice mycobiome during storage. Microbiome. 2024;12(1):45. [CrossRef]
- Johnson TS, Smith DL, Zhang Y, et al. Correlation between protein degradation products and off-flavor development in stored rice. J Agric Food Chem. 2022;70(10):3125–3133. [CrossRef]
- Lee S, Park S, Kim Y, et al. Phenolic compounds as natural preservatives to enhance the storage stability of cereal grains. Antioxidants. 2022;11(5):876. [CrossRef]
- Zhang Z, Li J, Wang H, et al. Mycotoxin risk assessment and fungal community succession in rice stored under simulated tropical conditions. World Mycotoxin J. 2023;16(2):145–158. [CrossRef]
- Kanehisa M, Sato Y, Morishima K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Annotation of Genome and Metagenome Sequences. Nucleic Acids Res. 2016;44(W1):W50–W55. [CrossRef]
- Buchfink B, Reuter K, Drost HG. Sensitive protein alignments at tree-of-life scale using DIAMOND. Nat Methods. 2021;18(4):366–368. [CrossRef]
Figure 1.
Changes in fatty acid values of rice during storage.
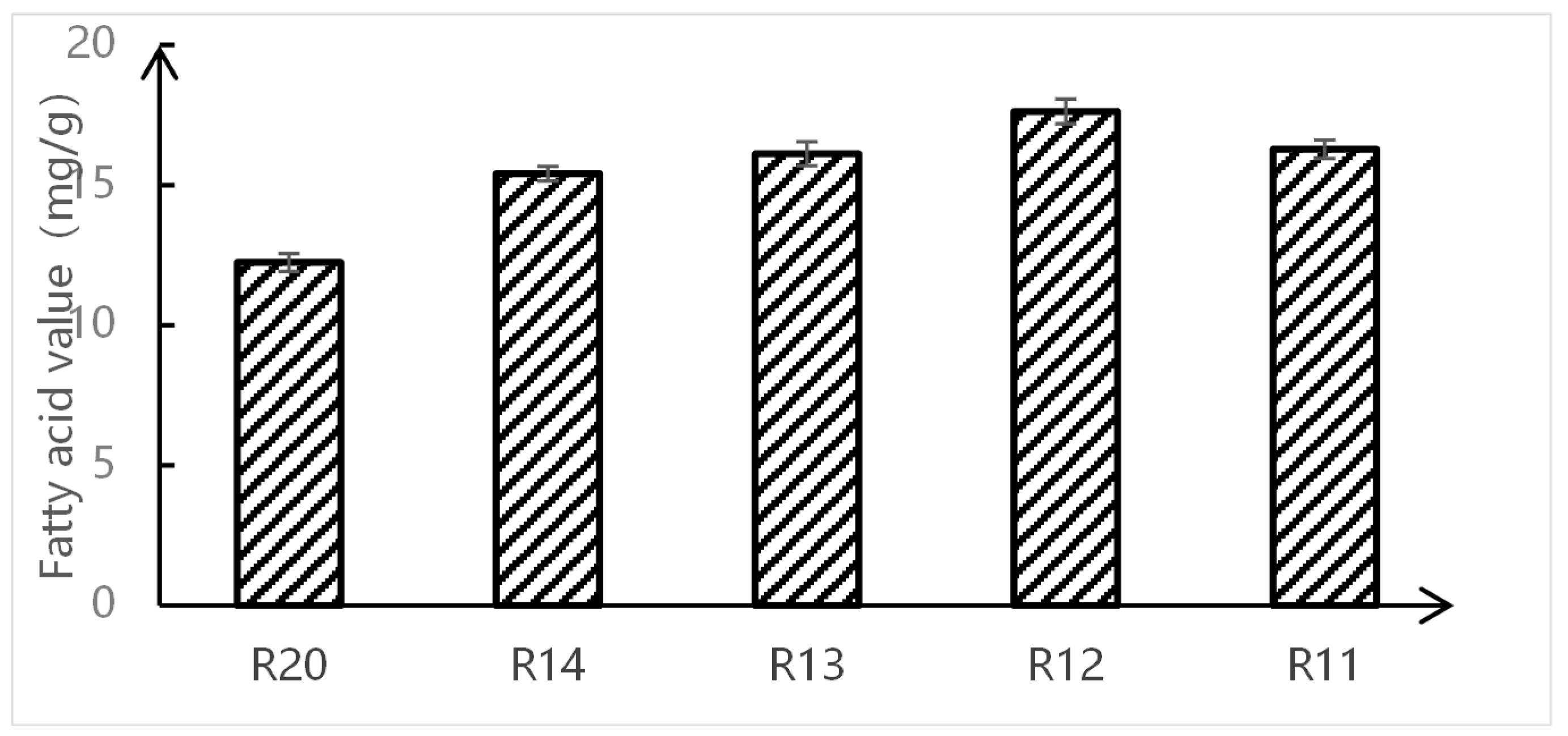
Figure 2.
Pie chart based on counts of HMDB chemical taxonomy (a) Metabolite classification pie chart at the Superclass level. (b) Metabolite classification pie chart at the Subclass level.
Figure 2.
Pie chart based on counts of HMDB chemical taxonomy (a) Metabolite classification pie chart at the Superclass level. (b) Metabolite classification pie chart at the Subclass level.
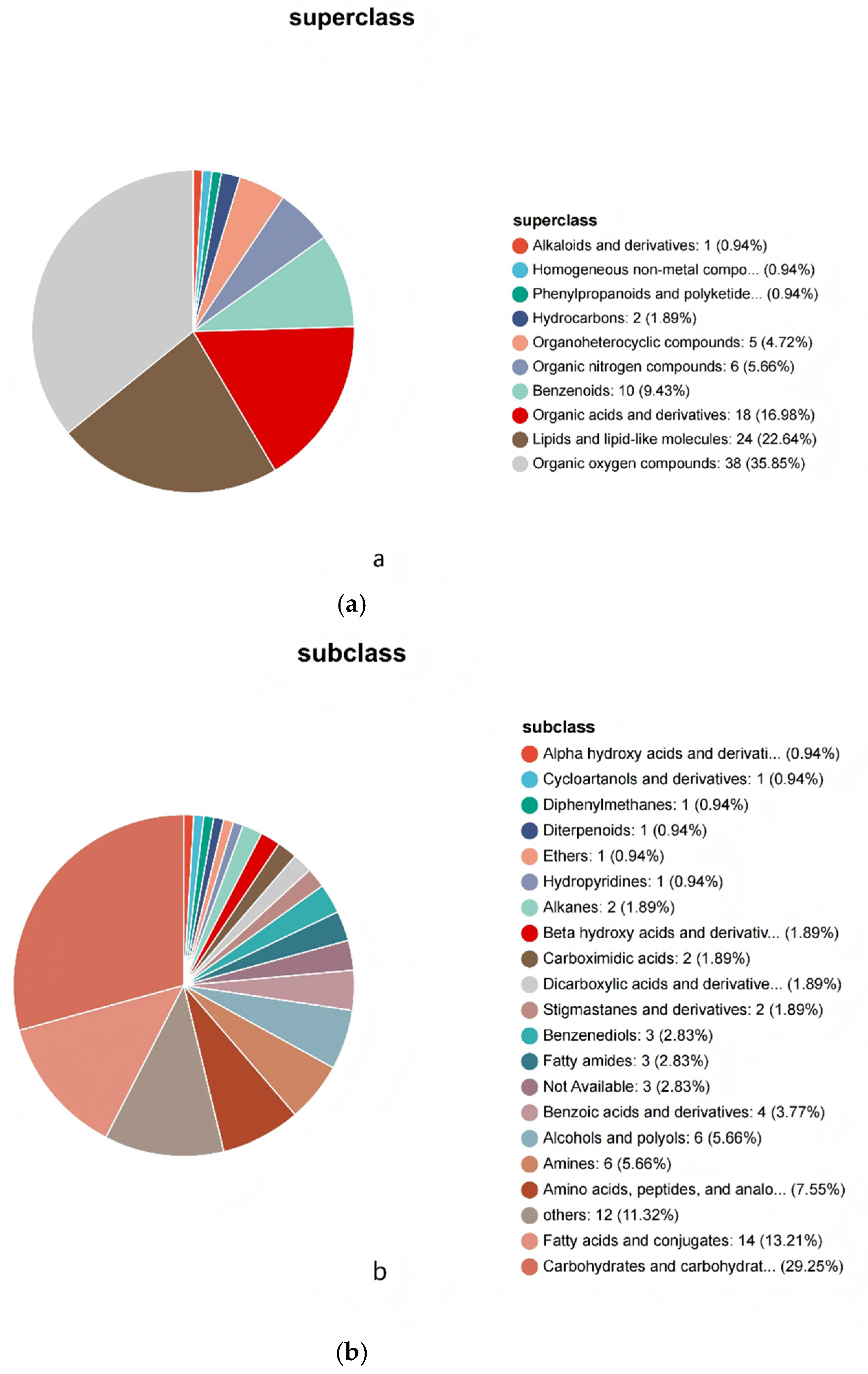
Figure 3.
Venn diagram of compounds in rice samples.

Figure 4.
PLS-DA scores plots (a), PLS-DA permutation test (b), Heat-map and cluster analysis of metabolites(c).
Figure 4.
PLS-DA scores plots (a), PLS-DA permutation test (b), Heat-map and cluster analysis of metabolites(c).
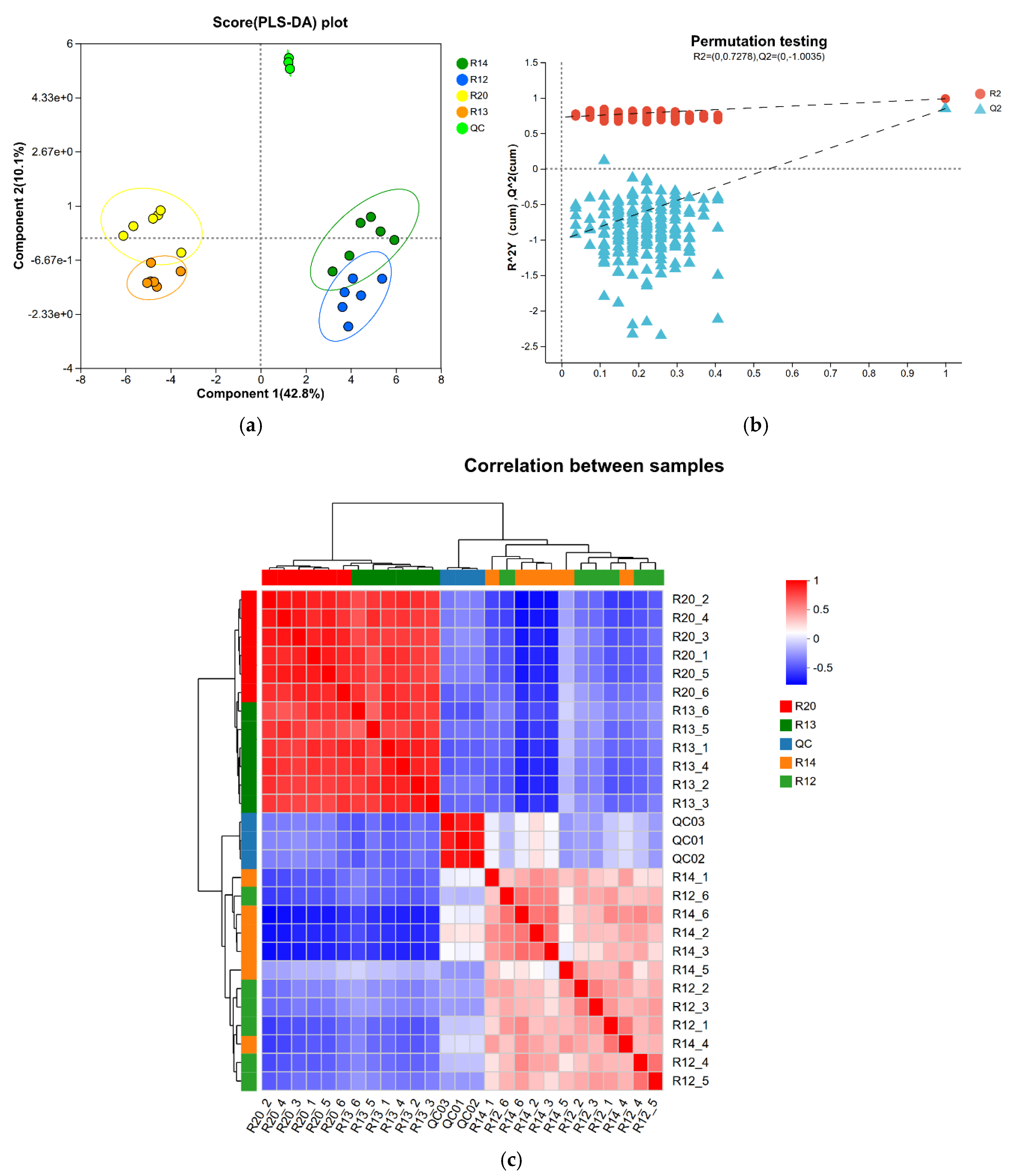
Figure 5.
KEGG Compounds classification between two groups (a) R12-R13; (b)R12-R14;(c)R12-R20;(d)R13-R14.
Figure 5.
KEGG Compounds classification between two groups (a) R12-R13; (b)R12-R14;(c)R12-R20;(d)R13-R14.
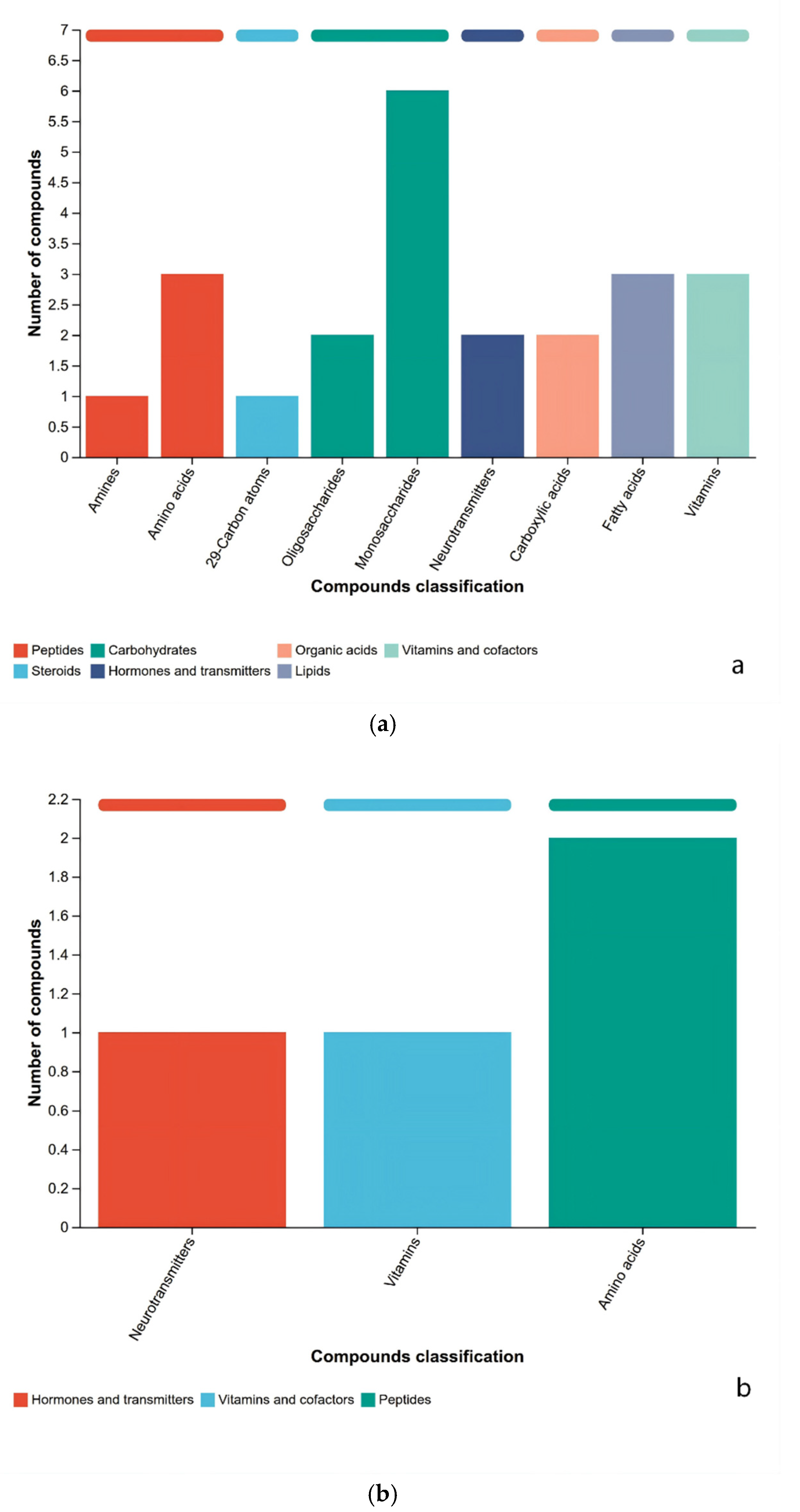
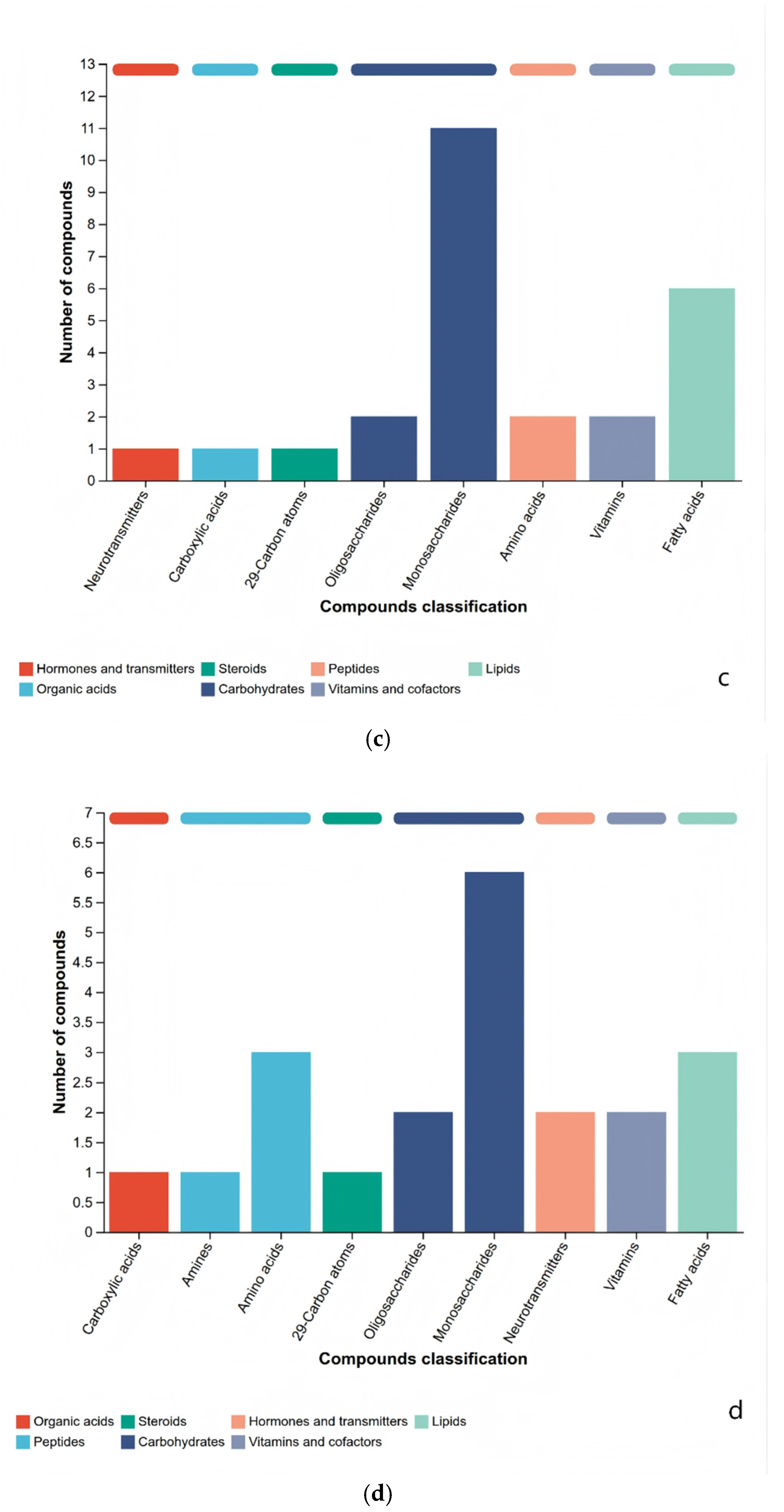
Figure 6.
KEGG pathways level 2 between two groups (a) R13-R14; (b)R12-R14;(c)R12-R13;(d)R12-R20.
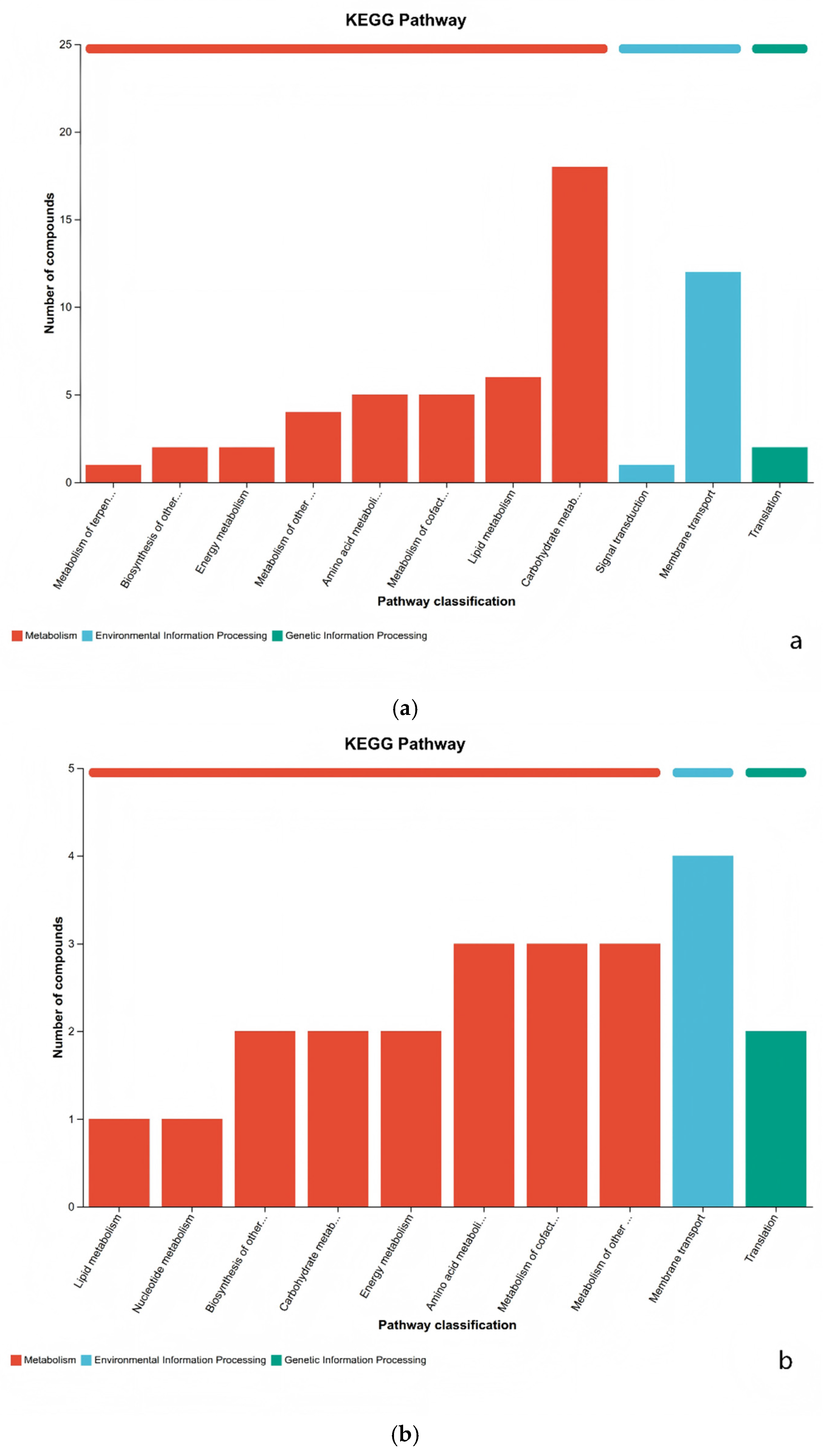
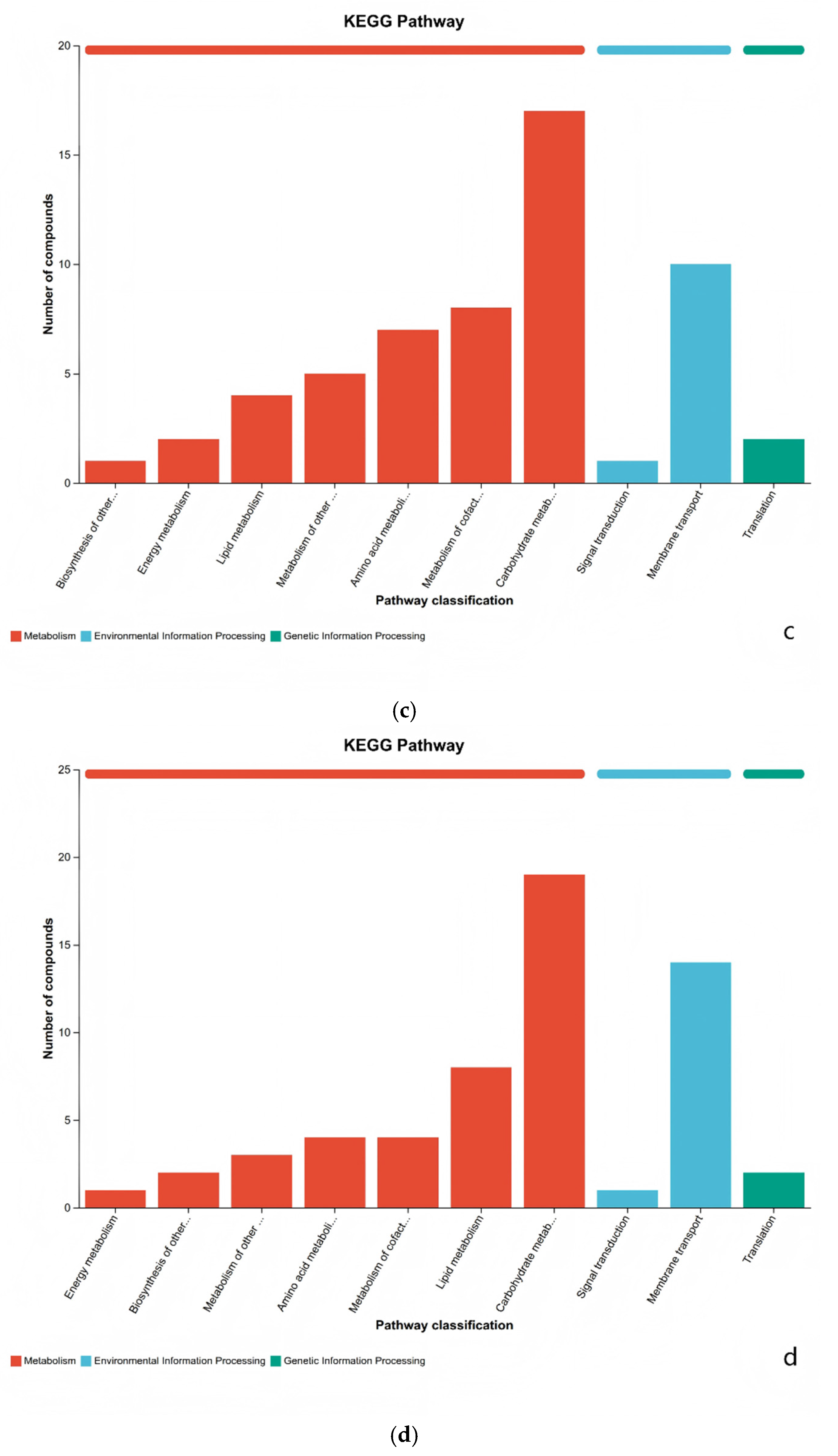
Figure 7.
Comparison of fungal communities in rice stored at different periods (a) α-diversity analysis; (b) Principal Component Analysis (PCoA) score plot; (c) LefSe analysis; (d) LDA score plot.
Figure 7.
Comparison of fungal communities in rice stored at different periods (a) α-diversity analysis; (b) Principal Component Analysis (PCoA) score plot; (c) LefSe analysis; (d) LDA score plot.
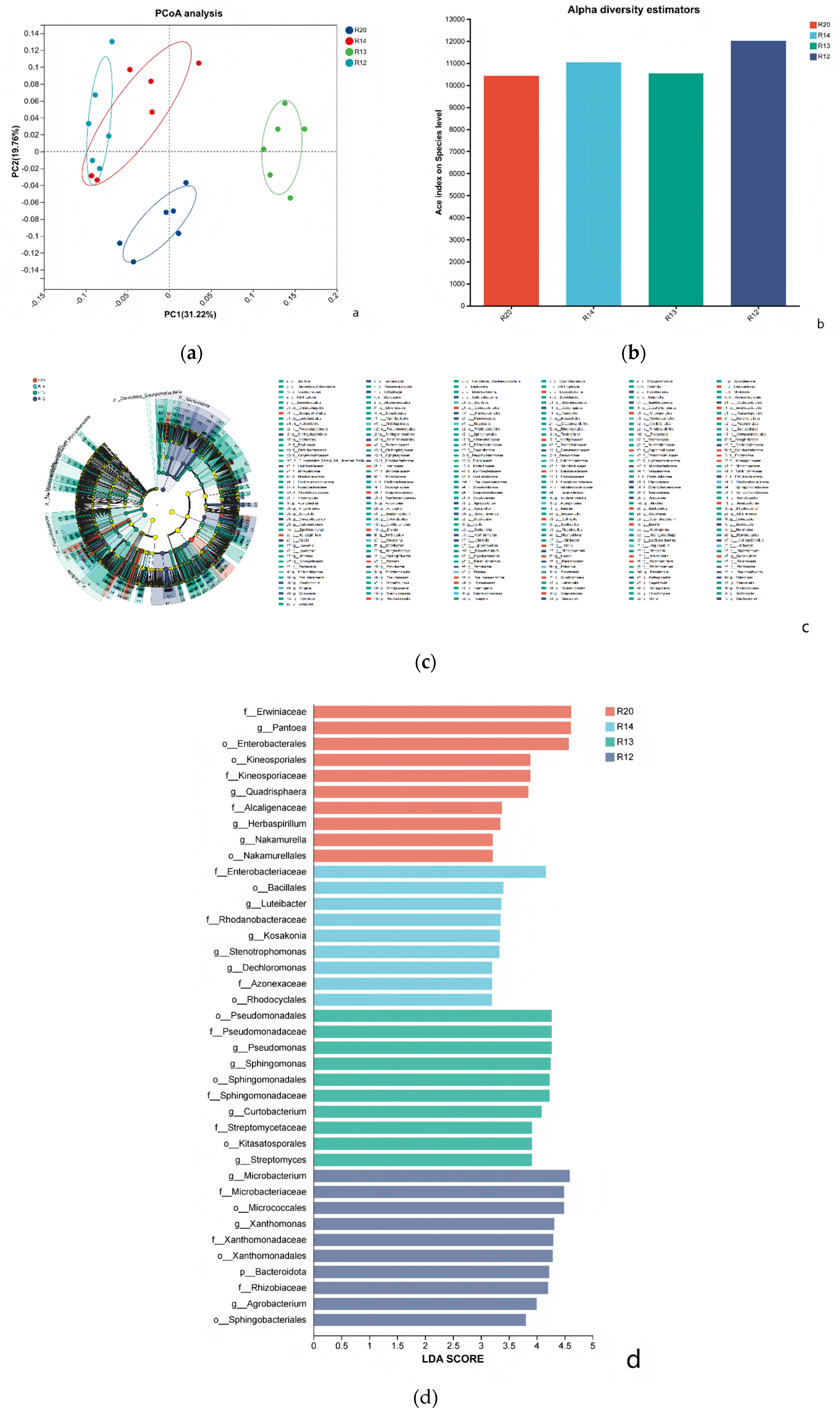
Figure 8.
The microbial community composition of rice samples at the phylum (a), genus (b), and species (c) levels.
Figure 8.
The microbial community composition of rice samples at the phylum (a), genus (b), and species (c) levels.
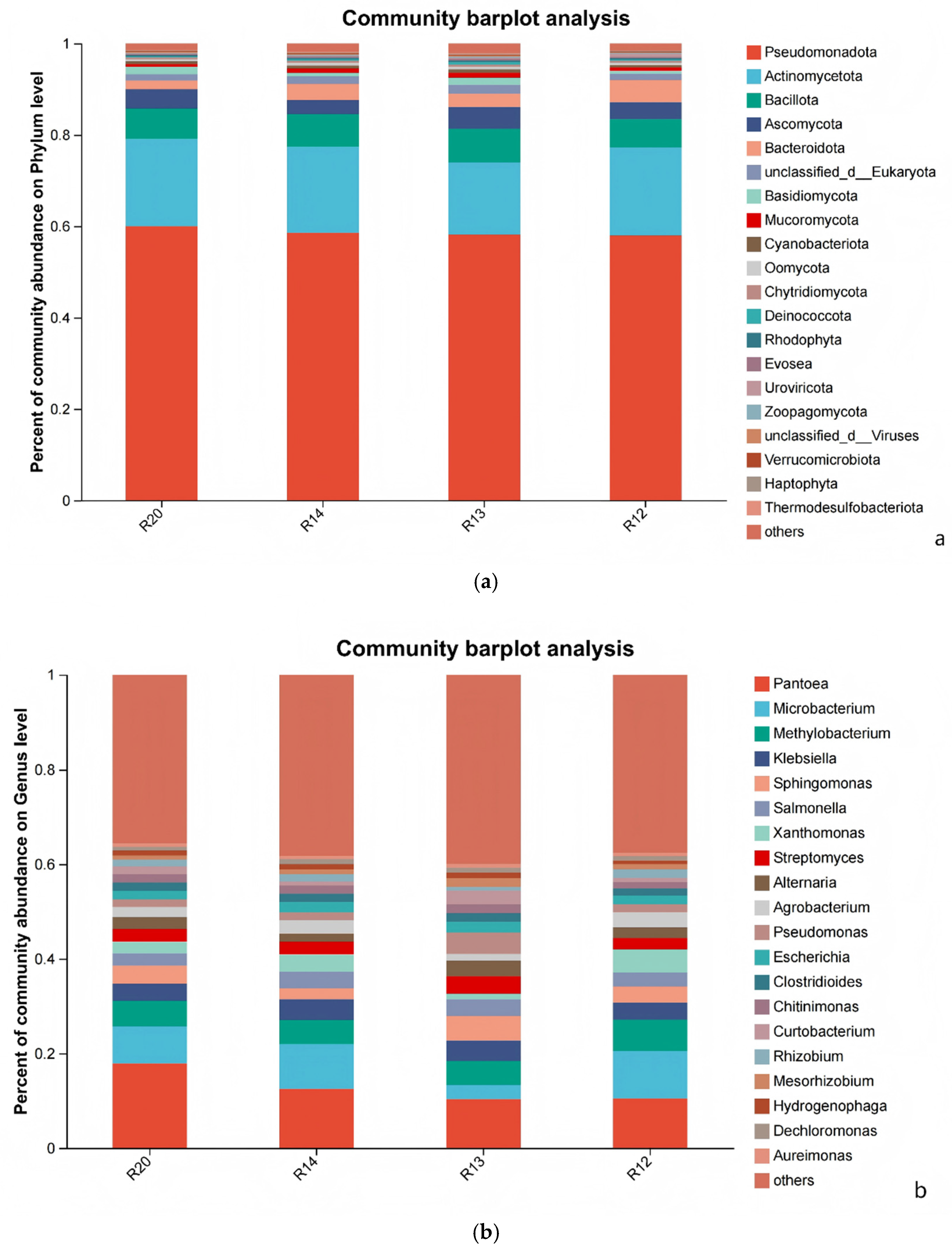
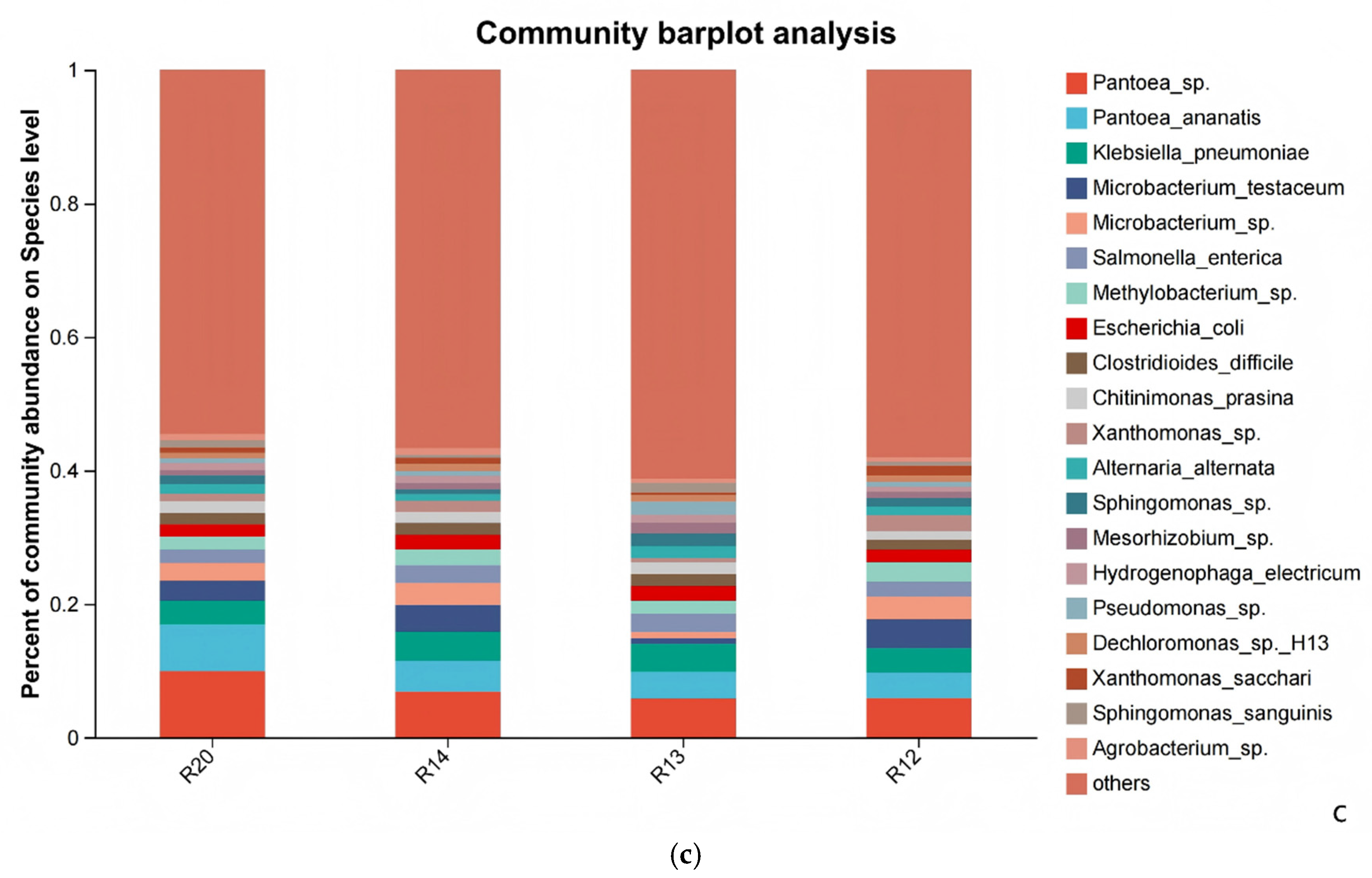
Figure 9.
KEGG co-occurrence network analysis on different levels (a)level 1;(b)level2.
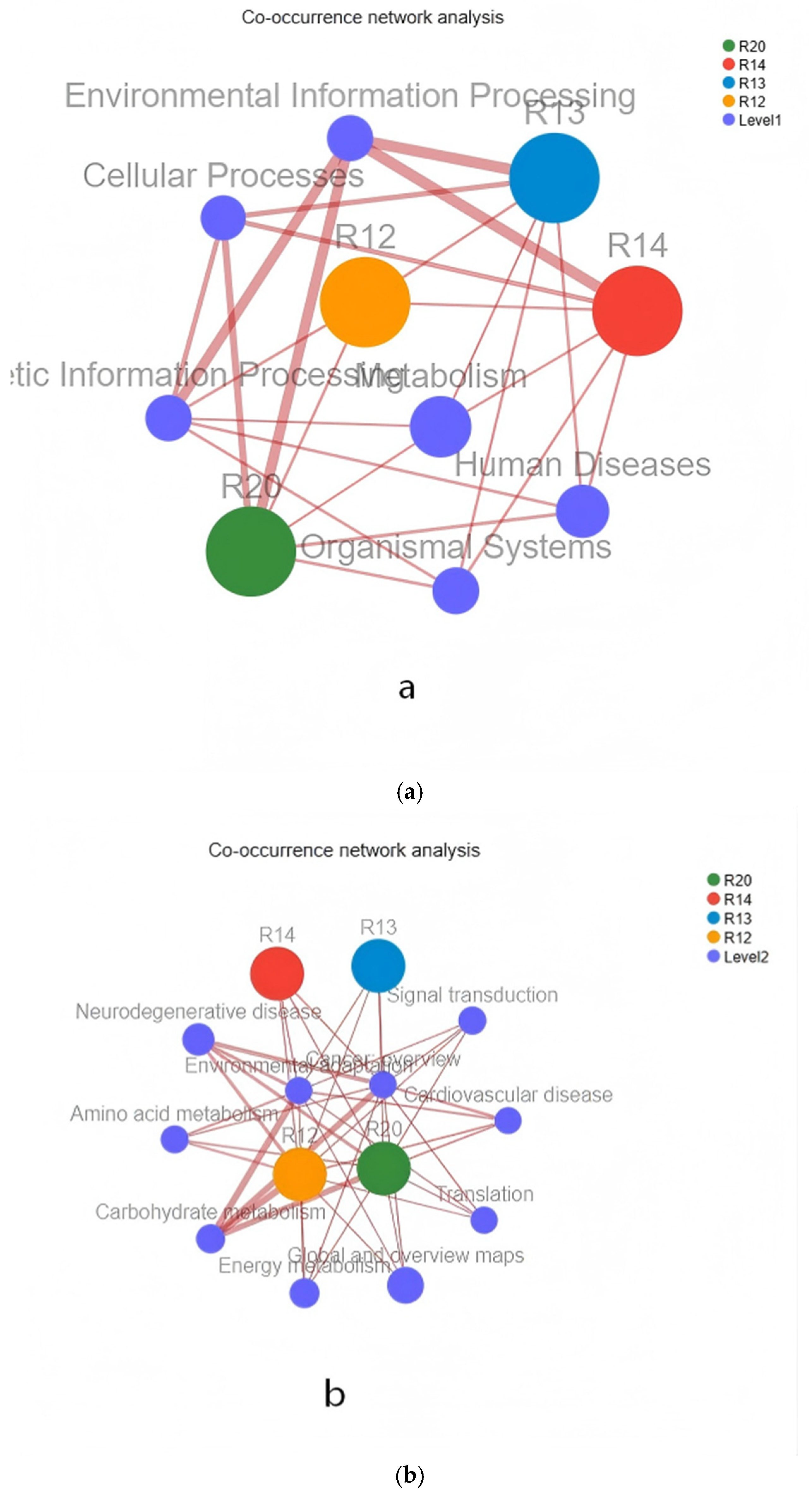
Figure 10.
Barplot of species and functional contribution analysis (a) all groups;(b) R12-R20;.
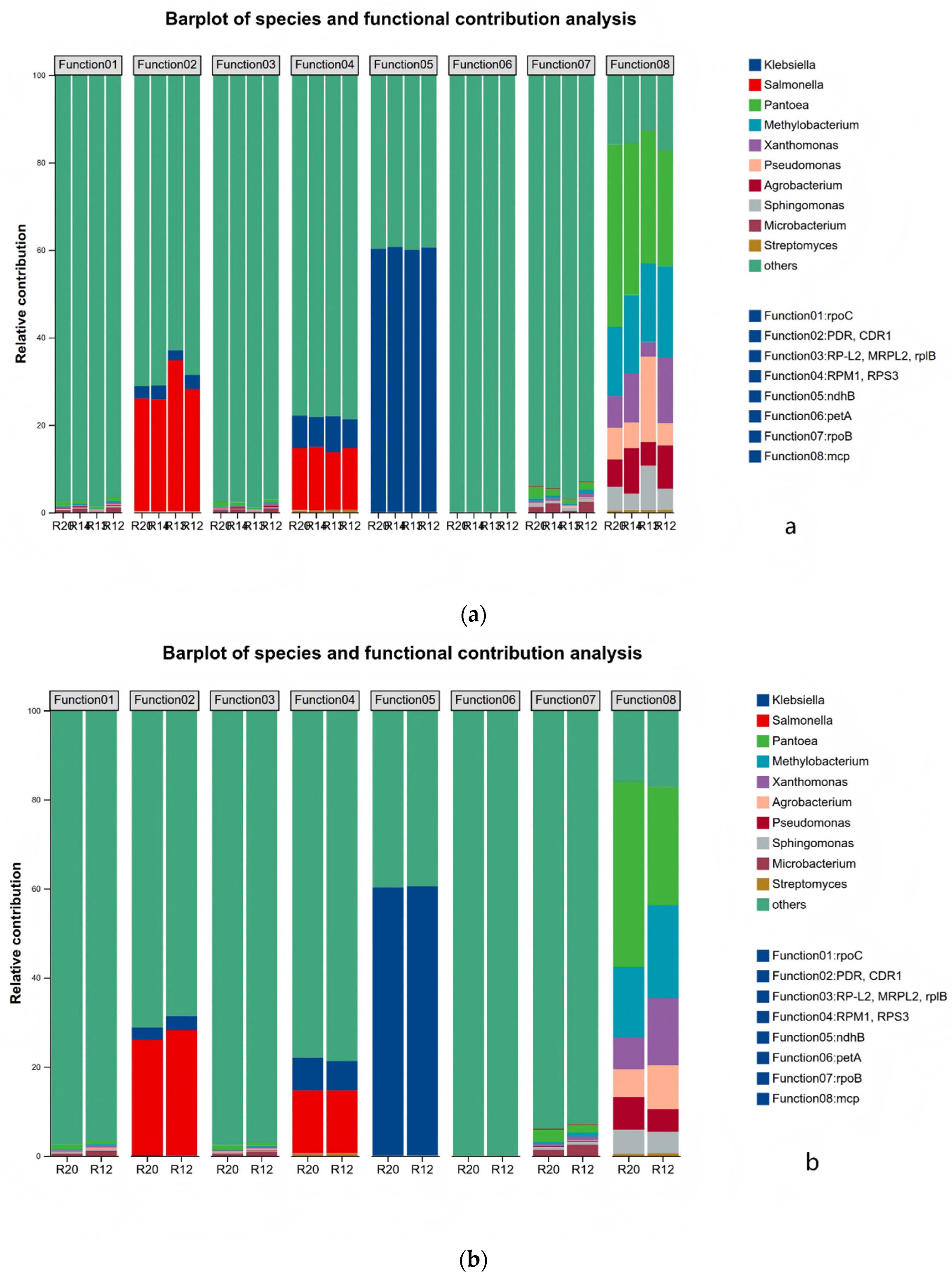

Table 1.
List of rice metabolites.
| order number | Metabolite name | retention time /min |
| 1 | (2,3,4,5,6-pentahydroxycyclohexyl)dihydrogen phosphate | 27.097 |
| 2 | (2-hydroxy-4-methoxyphenyl)-(2-hydroxyphenyl)methanone | 14.368 |
| 3 | (3S,8S,9S,10R,13R,14S,17R)-17-[(E,2R,5R)-5,6-dimethylhept-3-en-2-yl]-10,13-dimethyl-2,3,8,9,11,12,14,15,16,17-decahydro-1H-cyclopenta[a]phenanthren-3-ol | 33.285 |
| 4 | (4Z)-4-(furan-2-ylmethylidene)-2-phenyl-1,3-thiazol-5-one | 31.537 |
| 5 | (5S,8R,9S,10R,13S,14S)-4,4,10,13-tetramethyl-N-(2-phenylethyl)-1,5,6,7,8,9,11,12,14,15,16,17-dodecahydrocyclopenta[a]phenanthren-17-amine | 25.084 |
| 6 | (5Z)-5-methoxyiminohexane-1,2,3,4,6-pentol | 18.04 |
| 7 | (6Z)-6-methoxyiminoheptane-1,2,3,4,5,7-hexol | 23.859 |
| 8 | (9E,12E)-octadeca-9,12-dienoic acid | 25.089 |
| 9 | (E)-3-(4-hydroxy-3-methoxyphenyl)prop-2-enoic acid | 23.742 |
| 10 | (E)-but-2-ene-1,4-diol | 30.335 |
| 11 | (E)-hexadec-3-enenitrile | 23.807 |
| 12 | (E)-octadec-9-enamide | 27.207 |
| 13 | (Z)-octadec-9-enamide | 26.735 |
| 14 | (Z)-octadec-9-enenitrile | 23.703 |
| 15 | [1-[3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]sulfanylbut-3-enylideneamino]sulfate | 29.093 |
| 16 | [3-(4-ethylbenzoyl)oxyphenyl]3-methylbenzoate | 33.15 |
| 17 | [3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]dihydrogen phosphate | 24.982 |
| 18 | 1-(1-benzofuran-2-yl)ethanone | 6.873 |
| 19 | 1-(2-(3-Cyclohexenyl)Ethyl)Silatrane | 17.856 |
| 20 | 1-(2-methylazetidin-1-yl)ethanone | 6.19 |
| 21 | 1-(5-acetyl-4-methyl-1-phenyl-4H-pyridin-3-yl)ethanone | 14.007 |
| 22 | 1,3,5-Benzetriol | 16.92 |
| 23 | 10-bromodec-1-ene | 23.748 |
| 24 | 12A-Hydroxy-9-Demethylmunduserone-8-Carboxylic Acid | 7.412 |
| 25 | 17-(5,6-dimethylheptan-2-yl)-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-3-ol | 35.254 |
| 26 | 17-[(E)-5-ethyl-6-methylhept-3-en-2-yl]-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-3-ol | 35.559 |
| 27 | 18-Deoxysagittarol | 33.57 |
| 28 | 1-iododecane | 21.629 |
| 29 | 1-Kestose | 29.44 |
| 30 | 1-methoxy-4-[2-(4-methoxyphenyl)propan-2-yl]-2-methylbenzene | 15.368 |
| 31 | 1-nitro-4-(4-nitrophenyl)benzene | 8.951 |
| 32 | 1-octyl-2,8,9-trioxa-5-aza-1-silabicyclo[3.3.3]undecane | 14.457 |
| 33 | 1-Stearoyl-Rac-Glycerol | 30.563 |
| 34 | 2-(1-benzothiophen-3-yl)acetamide | 6.149 |
| 35 | 2-(2-hydroxyethylamino)ethanol | 15.532 |
| 36 | 2-(3,4-dihydroxyphenyl)-2-hydroxyacetic acid | 27.683 |
| 37 | 2-(4-chlorophenyl)-8-methylquinoline-4-carboxylic acid | 31.792 |
| 38 | 2-(carboxyamino)pentanoic acid | 11.686 |
| 39 | 2-(carboxyamino)propanoic acid | 10.997 |
| 40 | 2-(ethylamino)ethanol | 12.507 |
| 41 | 2-(hydroxymethyl)-6-[3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxyoxane-3,4,5-triol | 36.738 |
| 42 | 2,2,4,4,6,6,8,8,10,10,12,12,14,14,16,16,18,18,20,20,22,22,24,24-tetracosamethyl-1,3,5,7,9,11,13,15,17,19,21,23-dodecaoxa-2,4,6,8,10,12,14,16,18,20,22,24-dodecasilacyclotetracosane | 28.95 |
| 43 | 2,2,4,4,6,6,8,8,10,10,12,12,14,14,16,16-hexadecamethyl-1,3,5,7,9,11,13,15-octaoxa-2,4,6,8,10,12,14,16-octasilacyclohexadecane | 17.741 |
| 44 | 2,2,4,4,6,6,8,8,10,10,12,12-dodecamethyl-1,3,5,7,9,11-hexaoxa-2,4,6,8,10,12-hexasilacyclododecane | 12.588 |
| 45 | 2,3,4,5-tetrahydroxypentanoic acid | 19.759 |
| 46 | 2,3,4,6-tetrahydroxyhexanoic acid | 21.138 |
| 47 | 2,3-dihydro-1-benzofuran-5-yl(morpholin-4-yl)methanone | 11.483 |
| 48 | 2,4-dimethylpentan-3-yloxy-dodecoxy-ethenyl-methylsilane | 32.385 |
| 49 | 2,5,7-trimethyl-3-phenylpyrazolo[1,5-a]pyrimidine | 33.85 |
| 50 | 2,5-bis(hydroxymethyl)-2-methoxyoxolane-3,4-diol | 19.704 |
| 51 | 2,6-dihydroxybenzoic acid | 22.617 |
| 52 | 2,6-ditert-butylphenol | 16.496 |
| 53 | 2-[[2-[[2-[carboxy(methyl)amino]acetyl]-methylamino]acetyl]-methylamino]acetic acid | 18.377 |
| 54 | 2-[1-(4-hydroxy-1,5,5-trimethyl-2-oxopyrrol-3-yl)ethylideneamino]acetic acid | 8.933 |
| 55 | 2-[3,4-dihydroxy-2,5-bis(hydroxymethyl)oxolan-2-yl]oxy-6-(hydroxymethyl)oxane-3,4,5-triol | 29.204 |
| 56 | 2-[4-(cyanomethyl)phenyl]acetonitrile | 9.758 |
| 57 | 2-[carboxy(methyl)amino]acetic acid | 15.914 |
| 58 | 2-amino-6,7-bis(3-phenylpropyl)-3H-pteridin-4-one | 6.98 |
| 59 | 2-aminoacetic acid | 7.865 |
| 60 | 2-aminobutanedioic acid | 16.113 |
| 61 | 2-aminoethanol | 12.026 |
| 62 | 2-Ethoxyethanol | 6.386 |
| 63 | 2-fluorobenzoic acid | 18.437 |
| 64 | 2-Hydroxypyridine | 8.032 |
| 65 | 2-methoxy-N,N-bis(2-methoxyethyl)ethanamine | 6.455 |
| 66 | 2-Methyl-2-Butanol | 26.784 |
| 67 | 2-methylnaphthalene | 13.3 |
| 68 | 2-methylpropan-2-ol | 24.52 |
| 69 | 2-nitrothiophene | 35.249 |
| 70 | 2-oxo-2-(6-phenylimidazo[1,2-a]pyridin-2-yl)acetic acid | 12.255 |
| 71 | 3-(4-hydroxyphenyl)prop-2-enoic acid | 20.018 |
| 72 | 3,4,5-trihydroxy-6-methoxyoxane-2-carboxylic acid | 30.038 |
| 73 | 3,4-Dihydroxybenzyl alcohol | 8.69 |
| 74 | 3,4-Dihydroxymandelic Acid | 24.936 |
| 75 | 3,5,5-trimethylimidazolidine-2,4-dione | 6.804 |
| 76 | 3-acetylbenzonitrile | 11.898 |
| 77 | 3-hydroxypropanoic acid | 9.854 |
| 78 | 3-Hydroxypyridine | 9.836 |
| 79 | 3-methyl-1-phenyl-3H-indol-2-imine | 12.988 |
| 80 | 3-methyl-N-prop-2-enylbut-2-enamide | 6.25 |
| 81 | 3-phenylmethoxybutane-1,2,4-triol | 32.261 |
| 82 | 3-Trifluoromethylbenzylamine,N,N-diundecyl | 16.92 |
| 83 | 4-(2-aminoethyl)-3-methoxyphenol | 11.801 |
| 84 | 4-(2-phenylpropan-2-yl)phenol | 21.466 |
| 85 | 4,6-ditert-butylbenzene-1,3-diol | 13.607 |
| 86 | 4-[[4-(methylamino)phenyl]methylideneamino]benzonitrile | 8.381 |
| 87 | 4-[2-(4-cyanophenyl)-1,2-bis(diethylamino)ethyl]benzonitrile | 6.409 |
| 88 | 4-[2-(4-hydroxyphenyl)propan-2-yl]phenol | 25.188 |
| 89 | 4-[4-(1,2,3,4,5,6,7,8-octahydroacridin-4-yl)phenyl]-1,2,3,4,5,6,7,8-octahydroacridine | 16.344 |
| 90 | 4-Coumaric acid | 20.92 |
| 91 | 4-cyanobenzoic acid | 6.367 |
| 92 | 4-Hydroxy-3-Methoxybenzoic Acid | 19.665 |
| 93 | 4-Hydroxyanthraquinone-2-carboxylic acid | 29.49 |
| 94 | 4-Hydroxybenzoate | 17.796 |
| 95 | 4-hydroxybenzonitrile | 6.7 |
| 96 | 4-methyl-15-thiatetracyclo[10.2.1.05,14.08,13]pentadeca-1(14),2,4,6,8(13),9,11-heptaene | 8.454 |
| 97 | 4-methyl-2-(2-nitro-5-piperidin-1-ylphenyl)phthalazin-1-one | 7.964 |
| 98 | 4-phenylquinolin-3-ol | 8.057 |
| 99 | 5-(Hydroxymethylfurfural) | 12.931 |
| 100 | 5,6-Dihydrouracil | 9.792 |
| 101 | 5,6-dimethyl-2-[(E)-2-phenylethenyl]benzotriazole | 7.73 |
| 102 | 5-Aminovaleric Acid | 15.957 |
| 103 | 5-Hydroxy Tryptamine | 11.049 |
| 104 | 5-Hydroxy Tryptophan | 14.572 |
| 105 | 5-hydroxypiperidine-2-carboxylic acid | 17.312 |
| 106 | 5-methylsulfanyl-2-phenyl-10H-pyrrolo[3,4-a]carbazole-1,3-dione | 27.711 |
| 107 | 6-ethyl-7-methoxy-2-methyl-3-phenylchromen-4-one | 22.657 |
| 108 | 6-Kestose | 27.46 |
| 109 | 7-benzyl-6-methyl-2-phenylindolizine | 31.49 |
| 110 | 8-acetyl-2,4,6-trimethyl-7-phenylpurino[7,8-a]imidazole-1,3-dione | 15.079 |
| 111 | 9,12-Octadecadienoic acid(Z,Z)- | 24.09 |
| 112 | 9-Octadecenoic acid | 24.15 |
| 113 | Acetylsalicylic Acid | 16.024 |
| 114 | Allantoin | 21.178 |
| 115 | Alpha-Tocopherol | 32.97 |
| 116 | Arabitol | 17.91 |
| 117 | Benzene-1,3-dicarboxylic acid | 14.95 |
| 118 | Benzene-1,4-diol | 14.333 |
| 119 | Beta-Gentiobiose | 27.725 |
| 120 | Beta-Lactose | 26.95 |
| 121 | Beta-Sitosterol | 35.42 |
| 122 | Bis[[hydroxy(dimethyl)silyl]oxy]-dimethylsilane | 10.215 |
| 123 | Butan-1-amine | 9.04 |
| 124 | Butan-2-ol | 29.225 |
| 125 | Butoxy-diethyl-pentoxysilane | 18.194 |
| 126 | Cadaverine | 14.618 |
| 127 | Carbamic acid | 8.433 |
| 128 | Carbamic Acid Ethyl Ester | 10.407 |
| 129 | Cellotetraose | 36.31 |
| 130 | Citric Acid | 20.296 |
| 131 | Clionasterol acetate | 32.74 |
| 132 | Conduritol-Beta-Expoxide | 22.876 |
| 133 | Cycloartenol | 36.45 |
| 134 | D-(+)-Altrose | 21.526 |
| 135 | D-(+)-Trehalose | 30.184 |
| 136 | D-Allose | 21.194 |
| 137 | D-beta-Galactopyranosyl-1,3-arabinose | 30.38 |
| 138 | Decane | 25.527 |
| 139 | Decoxy-diethyl-octoxysilane | 26.947 |
| 140 | D-Glucitol | 20.7 |
| 141 | D-Glucose | 21.274 |
| 142 | Diethylcarbamic acid | 8.316 |
| 143 | Diethyl-hexadecoxy-[(2,3,4,5,6-pentafluorophenyl)methoxy]silane | 36.207 |
| 144 | Dihydroxy(dimethyl)silane | 7.042 |
| 145 | Dimethoxymethane | 6.637 |
| 146 | Dimethyl-octadecoxy-(4-phenylphenoxy)silane | 35.207 |
| 147 | Diphenylphosphane | 9.373 |
| 148 | D-Malic Acid | 15.606 |
| 149 | D-Mannitol | 20.7 |
| 150 | D-manno-2-Heptulose | 20.17 |
| 151 | D-Mannose | 20.11 |
| 152 | Dodecanamide | 26.05 |
| 153 | D-Sorbitol | 21.772 |
| 154 | D-Sphingosine | 32.524 |
| 155 | D-Threitol | 15.869 |
| 156 | Eicosanoic acid | 26.49 |
| 157 | Eicosapentaenoic Acid | 35.01 |
| 158 | Elaidic Acid | 25.15 |
| 159 | Enolpyruvate | 8.847 |
| 160 | Erythritol | 14.86 |
| 161 | Ethanamine | 6.656 |
| 162 | Ethane-1,2-diol | 7.1 |
| 163 | Ethene-1,1,2-triol | 16.437 |
| 164 | Ethyl cyanate | 6.22 |
| 165 | Fructose | 19.93 |
| 166 | Galactinol | 33.48 |
| 167 | Galactitol | 26.562 |
| 168 | Galactose | 20.26 |
| 169 | Galactose oxime | 23.12 |
| 170 | Gamma-Aminobutyric Acid | 16.311 |
| 171 | Geranyllinalool | 33.57 |
| 172 | Gluconic Acid | 22.714 |
| 173 | Glucose | 21.26 |
| 174 | Glucose-1-Phosphate | 19.615 |
| 175 | Glycerol | 11.05 |
| 176 | Glycerol 1-Phosphate | 19.523 |
| 177 | Glycolic Acid | 8.633 |
| 178 | Glycyl-Proline | 17.266 |
| 179 | Harmaline | 10.812 |
| 180 | Heptadecanoic Acid | 24.345 |
| 181 | Heptanamide | 25.91 |
| 182 | Hexadecanamide | 24.784 |
| 183 | Hexadecanoic acid | 28.816 |
| 184 | Hexanamide | 26.84 |
| 185 | Hydroxy-[hydroxy(dimethyl)silyl]oxy-dimethylsilane | 7.895 |
| 186 | Hydroxy-methyl-trihydroxysilyloxysilicon | 19.838 |
| 187 | Icosanoic acid | 27.455 |
| 188 | Inositol | 23.639 |
| 189 | Ketohexose | 18.35 |
| 190 | Lactose | 30.45 |
| 191 | L-Asparagine | 18.312 |
| 192 | L-Glutamic Acid | 16.195 |
| 193 | L-Glycine | 12.707 |
| 194 | L-Iditol | 29.269 |
| 195 | Lignoceric acid | 30.23 |
| 196 | L-Serine | 13.558 |
| 197 | L-Tyrosine | 21.877 |
| 198 | Malonic Acid | 10.904 |
| 199 | Maltotriose | 37.187 |
| 200 | Melezitose | 28.16 |
| 201 | Methanediimine | 6.737 |
| 202 | Methanolphosphate | 10.429 |
| 203 | Methyl 2,8-dimethyldecanoate | 25.59 |
| 204 | Methyl 2-methylundecanoate | 25.59 |
| 205 | Methyl-(2-methyl-1,3-dioxoisoindol-4-yl)carbamic acid | 20.626 |
| 206 | Methylbeta-D-fructofuranoside | 18.69 |
| 207 | Methyl-Beta-D-Galactopyranoside | 27.841 |
| 208 | Methylimino(sulfanylidene)methane | 6.73 |
| 209 | Myo-Inositol | 22.65 |
| 210 | Myristic acid | 19.76 |
| 211 | N-(1,1-dioxo-2,3-dihydrothiophen-3-yl)-2-(4-methoxyphenoxy)acetamide | 28.538 |
| 212 | N-(4-aminobutyl)acetamide | 15.217 |
| 213 | N-[[3,6-dichloro-2,7-bis[2-(diethylamino)ethoxy]fluoren-9-ylidene]amino]acetamide | 6.375 |
| 214 | N-Acetylornithine | 10.104 |
| 215 | Nonacyclo[18.14.2.02,19.04,17.05,10.011,16.024,36.025,30.031,35]hexatriaconta-1(34),2(19),3,5,7,9,11,13,15,17,20,22,24(36),25,27,29,31(35),32-octadecaene | 18.147 |
| 216 | Nonadecanenitrile | 26.211 |
| 217 | Nonanamide | 26.06 |
| 218 | Norleucine | 12.089 |
| 219 | N-propan-2-ylacetamide | 6.487 |
| 220 | Octadecan-1-ol | 24.46 |
| 221 | Octadecanamide | 27.466 |
| 222 | Octamethylcyclotetrasiloxane | 6.994 |
| 223 | Oleamide | 27.336 |
| 224 | Oleic Acid | 24.15 |
| 225 | Palmitic Acid | 22.18 |
| 226 | Pantothenic Acid | 22.495 |
| 227 | P-Coummaric Acid | 21.951 |
| 228 | Pentane-1,5-diamine | 14.774 |
| 229 | Phosphoric Acid | 12.068 |
| 230 | P-Hydroxybenzaldehyde | 15.845 |
| 231 | Piperidin-2-one | 7.493 |
| 232 | P-Quinquephenyl | 31.9 |
| 233 | Propan-1-amine | 9.668 |
| 234 | Propane-1,2,3-triol | 33.99 |
| 235 | Propyl Lactate | 10.661 |
| 236 | Propylamine | 7.778 |
| 237 | Psicose | 20.945 |
| 238 | Pyridine-3-carboxylic acid | 12.602 |
| 239 | Pyrrolidine-2-carboxylic acid | 12.581 |
| 240 | Raffinose | 35.86 |
| 241 | Ribitol | 18.01 |
| 242 | Ribose | 17.11 |
| 243 | Serotonin | 15.008 |
| 244 | Sorbitol | 20.69 |
| 245 | Squalene | 30.07 |
| 246 | Stearic Acid | 24.43 |
| 247 | Stigmasterol | 34.62 |
| 248 | Succinic Acid | 12.832 |
| 249 | Sucrose | 27.18 |
| 250 | Tagatose | 21.063 |
| 251 | Terephthalic acid | 8.506 |
| 252 | Tetracosanoic acid | 31.1 |
| 253 | Tetrahydroxysilane | 11.203 |
| 254 | Threonic Acid | 16.656 |
| 255 | Trehalose | 29.32 |
| 256 | Tricosanoic acid | 32.204 |
| 257 | Tridec-2-yne | 9.881 |
| 258 | Trihydroxy(methyl)silane | 8.207 |
| 259 | Undecane | 11.772 |
| 260 | Urea | 11.584 |
| 261 | Vanillin | 18.076 |
| 262 | Xylitol,5TMS derivative | 19.015 |
| 263 | Xylose | 19.38 |
Table 2.
Differential metabolites and their dominant KEGG pathways.
| Metabolite | Regulate | KEGG Pathway Description | Key Pathways |
| 3,4-Dihydroxymandelic Acid | up | Metabolic pathways;Tyrosine metabolism | Amino acid metabolism |
| 4-Coumaric acid | down | Metabolic pathways;Phenylpropanoid biosynthesis;Biosynthesis of secondary metabolites | Flavonoids |
| 9-Octadecenoic acid | down | Cutin, suberine and wax biosynthesis;Biosynthesis of unsaturated fatty acids;Fatty acid biosynthesis | Metabolism of short-chain fatty acids |
| Citric Acid | up | Metabolic pathways;Glyoxylate and dicarboxylate metabolism;Biosynthesis of secondary metabolites;Biosynthesis of cofactors;2-Oxocarboxylic acid metabolism;Carbon metabolism;Biosynthesis of amino acids;Alanine, aspartate and glutamate metabolism;Citrate cycle (TCA cycle) | Amino acid metabolism Energy metabolism |
| D-Glucose | up | Metabolic pathways;Biosynthesis of secondary metabolites;Amino sugar and nucleotide sugar metabolism;ABC transporters;Galactose metabolism;Glycolysis / Gluconeogenesis;Starch and sucrose metabolism;Pentose phosphate pathway | Energy metabolism |
| Glucose | down | Metabolic pathways;Biosynthesis of secondary metabolites;Biosynthesis of nucleotide sugars;ABC transporters;Amino sugar and nucleotide sugar metabolism;Carbon metabolism;Fructose and mannose metabolism;Galactose metabolism;Glycolysis / Gluconeogenesis;Starch and sucrose metabolism;Pentose phosphate pathway | Energy metabolism |
| L-Asparagine | up | Metabolic pathways;Biosynthesis of secondary metabolites;Cyanoamino acid metabolism;Biosynthesis of amino acids;Alanine, aspartate and glutamate metabolism;Aminoacyl-tRNA biosynthesis | Amino acid metabolism |
| L-Glutamic Acid | up | Metabolic pathways;Glyoxylate and dicarboxylate metabolism;Arginine biosynthesis;ABC transporters;Taurine and hypotaurine metabolism;Biosynthesis of cofactors;2-Oxocarboxylic acid metabolism;Nitrogen metabolism;Carbon metabolism;Arginine and proline metabolism;Glutathione metabolism;Biosynthesis of amino acids;Alanine, aspartate and glutamate metabolism;D-Amino acid metabolism;Aminoacyl-tRNA biosynthesis;Histidine metabolism;Biosynthesis of secondary metabolites;Butanoate metabolism; C5-Branched dibasic acid metabolism;Porphyrin metabolism | Amino acid metabolism Metabolism of short-chain fatty acids |
| Myristic acid | down | Metabolic pathways;Fatty acid biosynthesis | Metabolism of short-chain fatty acids |
| Palmitic Acid | down | Metabolic pathways;Fatty acid degradation;Fatty acid metabolism;Cutin, suberine and wax biosynthesis;Fatty acid biosynthesis;Fatty acid elongation;Biosynthesis of unsaturated fatty acids | Lipid metabolism Metabolism of short-chain fatty acids |
| Ribose | up | Metabolic pathways;Pentose phosphate pathway;ABC transporters | Energy metabolism |
| Stearic Acid | down | Metabolic pathways;Biosynthesis of unsaturated fatty acids;Fatty acid biosynthesis | Metabolism of short-chain fatty acids |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



