Submitted:
10 February 2026
Posted:
11 February 2026
You are already at the latest version
Abstract
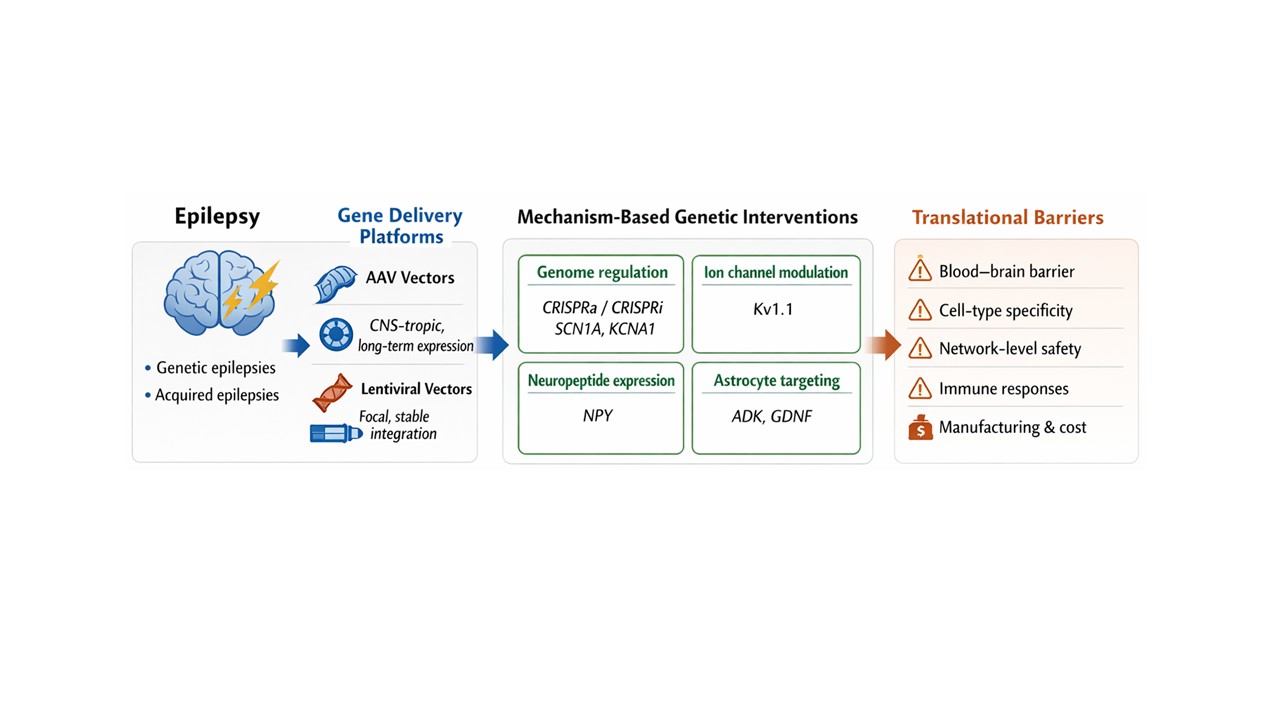
Epilepsy affects more than 50 million individuals worldwide, and approximately one-third of patients remain refractory to existing antiseizure medications. Advances in gene therapy and genome editing have opened new possibilities for disease-modifying interventions that directly target the molecular and circuit-level mechanisms underlying epileptogenesis. Recent progress in central nervous system tropic viral vectors, non-viral delivery systems, and programmable genome-editing technologies has enabled precise manipulation of neuronal and glial function in preclinical epilepsy models. Strategies range from restoration of haploinsufficient genes implicated in monogenic epilepsies, such as SCN1A in Dravet syndrome, to modulation of neuronal excitability through engineered ion channels, neuropeptides, and astrocyte-based approaches. In parallel, CRISPR-derived platforms, including transcriptional activation and repression systems, base editing, and prime editing, offer new avenues for regulating gene expression in post-mitotic neurons without introducing double-strand DNA breaks. Despite these advances, significant translational challenges remain, including efficient and cell-type-specific delivery, long-term safety, and the risk of network-level side effects in the epileptic brain. This review critically examines recent gene therapy and genome-editing approaches for epilepsy, highlights key technological and biological barriers to clinical translation, and discusses emerging strategies that may enable durable and targeted treatments for drug-resistant epilepsies.
Keywords:
epilepsy
; gene therapy
; genome editing
; CRISPR-based technologies
Introduction
Epilepsy is a chronic neurological disorder characterized by recurrent, unprovoked seizures resulting from pathological neuronal hyperexcitability and network synchronization. It affects more than 50 million individuals worldwide and encompasses a broad spectrum of etiologies, including genetic, structural, metabolic, immune, infectious, and idiopathic causes (Stafstrom and Carmant, 2015; Perucca et al., 2020; Thijs et al., 2019). Despite the availability of numerous antiseizure medications, approximately one-third of patients develop drug-resistant epilepsy, a condition associated with increased morbidity, cognitive impairment, psychiatric comorbidities, and elevated mortality risk (Sheng et al., 2018). For these patients, surgical resection or neuromodulation may offer benefit, yet many are not eligible or fail to achieve durable seizure freedom, highlighting a critical unmet clinical need (Simonato et al., 2013; Street et al., 2023).
Gene therapy has emerged as a compelling therapeutic strategy for epilepsy because seizure disorders frequently arise from well-defined molecular and cellular dysfunctions that are amenable to genetic modulation. Advances in viral vector engineering, particularly the development of adeno-associated virus (AAV) platforms, have enabled long-term gene expression in post-mitotic neurons and glial cells with a favorable safety profile (Kay et al., 2001; Naldini, 2015; Hudry and Vandenberghe, 2019). The clinical success of AAV-mediated gene therapies in neurological and neuromuscular disorders, including spinal muscular atrophy and inherited retinal dystrophies, has further accelerated interest in applying these technologies to epilepsy (Mendell et al., 2017; Russell et al., 2018; Deverman et al., 2018).
Both monogenic and acquired epilepsies present rational targets for gene-based interventions. In monogenic epileptic encephalopathies, loss-of-function or gain-of-function variants in ion channel and synaptic genes directly disrupt neuronal excitability and inhibitory control (Perucca et al., 2020; Lopez-Santiago and Isom, 2019). Dravet syndrome, caused primarily by haploinsufficiency of the SCN1A gene encoding the Nav1.1 sodium channel, exemplifies this paradigm and has become a leading model for precision gene therapy approaches in epilepsy (Gataullina and Dulac, 2017; Colasante et al., 2020). In acquired focal epilepsies, such as temporal lobe epilepsy, maladaptive changes in excitatory and inhibitory balance, neurotransmitter signaling, and glial homeostasis provide additional opportunities for targeted genetic modulation (Simonato et al., 2013; Snowball et al., 2019).
Beyond classical gene replacement, genome-editing and gene-regulation technologies have fundamentally expanded the therapeutic landscape for epilepsy. CRISPR-based systems enable programmable manipulation of endogenous genes, allowing either disruption, correction, or transcriptional modulation without the need for full-length transgene delivery (Doudna and Charpentier, 2014; Hsu et al., 2014). Importantly, transcriptional activation and repression platforms based on catalytically inactive Cas9 (CRISPRa/i) permit reversible and tunable regulation of neuronal genes without introducing double-strand DNA breaks, a critical safety consideration in post-mitotic neurons (Swiech et al., 2015; Staahl et al., 2017; Colasante et al., 2020). More recently, base editing and prime editing technologies have enabled precise correction of pathogenic point mutations with reduced genotoxic risk, further expanding the applicability of genome editing to developmental and epileptic encephalopathies (Gaudelli et al., 2017; Anzalone et al., 2019; Chen and Liu, 2023).
Despite these advances, significant translational challenges remain for gene therapy in epilepsy. Efficient and cell-type–specific delivery across the blood–brain barrier, long-term safety and immunogenicity, network-level consequences of sustained gene modulation, and the high cost of vector manufacturing all represent major obstacles to widespread clinical implementation (Nayak and Herzog, 2010; Ylä-Herttuala, 2020; Street et al., 2023). Addressing these challenges will be essential to move epilepsy gene therapies from experimental interventions toward routine clinical practice.
In this review, we critically examine recent advances in gene therapy and genome-editing strategies for epilepsy, with a particular emphasis on delivery platforms, molecular targets, and genome-regulation technologies that have demonstrated efficacy in preclinical models. We discuss how biological constraints unique to the epileptic brain have shaped therapeutic design and highlight emerging approaches that may enable durable, precise, and safe treatments for drug-resistant epilepsies.
2. Vector Platforms for Epilepsy Gene Therapy
Effective gene therapy for epilepsy critically depends on delivery systems capable of achieving precise, durable, and cell-type specific transgene expression within the central nervous system (CNS) (Simonato et al., 2013; Hudry and Vandenberghe, 2019). Unlike peripheral disorders, epilepsy presents unique challenges for gene delivery, including the blood–brain barrier, the predominance of post-mitotic target cells, and the need to modulate defined neuronal and glial populations without disrupting broader network stability (Kantor et al., 2014; Street et al., 2023). Consequently, vector selection is not merely a technical consideration but a central determinant of therapeutic efficacy and safety (Simonato et al., 2013).
2.1. Viral Vectors for CNS Delivery
Among available platforms, adeno-associated viruses (AAVs) have emerged as the most widely used vectors for epilepsy gene therapy due to their favorable safety profile, long-term transgene expression, and ability to transduce non-dividing neurons (Kay et al., 2001; Naldini, 2015; Hudry and Vandenberghe, 2019). AAV-mediated gene delivery has been particularly successful in preclinical epilepsy models, where sustained modulation of neuronal excitability is required to achieve durable seizure suppression (Wykes et al., 2012; Snowball et al., 2019; Colasante et al., 2020). The development of CNS-tropic AAV serotypes and engineered capsids has further expanded their utility, enabling broader brain distribution and improved penetration across the blood-brain barrier (Deverman et al., 2016; Hsu et al., 2020; Mathiesen et al., 2023).
For epilepsy applications, vector tropism is a critical consideration. Many therapeutic strategies aim to selectively target inhibitory interneurons, astrocytes, or discrete excitatory neuronal populations, necessitating the use of cell-type–specific promoters or enhancer elements in combination with appropriate AAV serotypes (Dimidschstein et al., 2016; Kantor et al., 2014). For example, AAV9 and engineered variants have demonstrated robust transduction of neurons throughout the brain following systemic or intracerebroventricular delivery, while interneuron-selective regulatory elements have enabled preferential targeting of GABAergic circuits implicated in seizure generation (Deverman et al., 2016; Dimidschstein et al., 2016). However, widespread CNS transduction also raises concerns regarding off-target effects and excessive inhibition, underscoring the need for precise spatial and cellular control (Simonato et al., 2013; Street et al., 2023).
Cargo capacity remains an important limitation of AAV-based approaches, particularly for epilepsy-associated genes such as SCN1A, which exceed the packaging limits of single AAV vectors (Colasante et al., 2020). This constraint has driven the development of alternative strategies, including transcriptional activation of endogenous genes and dual-vector systems, although the latter suffer from reduced co-transduction efficiency (Colasante et al., 2020; Staahl et al., 2017). These challenges have directly shaped the evolution of genome-regulation approaches in epilepsy, favoring methods that modulate native gene expression rather than relying on full-length gene replacement (Colasante et al., 2020; Street et al., 2023).
CNS-tropic AAV serotypes used in neurological gene therapy are summarized in Table 1.
2.2. Lentiviral Vectors and Stable Gene Integration
Lentiviral vectors offer the advantage of stable genomic integration and sustained transgene expression, making them attractive for chronic neurological disorders requiring long-term modulation (Kay et al., 2001; Naldini, 2015). In epilepsy research, lentiviral vectors have been extensively used in focal delivery paradigms, particularly in rodent models of temporal lobe epilepsy, where localized expression of therapeutic genes such as engineered potassium channels has produced prolonged seizure suppression (Wykes et al., 2012; Snowball et al., 2019). The ability to deliver larger genetic payloads compared with AAV vectors further expands the range of potential targets (Kohn et al., 2020).
Despite these advantages, the integration of lentiviral vectors into the host genome raises safety concerns, including insertional mutagenesis, which may limit their translational applicability for epilepsy (Sadelain, 2004; Naldini, 2015). While the development of self-inactivating lentiviral constructs has substantially improved safety profiles, the irreversible nature of genomic integration remains a consideration, particularly in disorders where fine-tuned regulation of neuronal activity is essential (Li and Samulski, 2020). As a result, lentiviral approaches in epilepsy are currently viewed as powerful experimental tools and potential candidates for highly localized therapies rather than broadly deployable clinical platforms (Simonato et al., 2013).
2.3. Non-Viral Delivery Systems
Non-viral delivery platforms, including lipid nanoparticles and polymer-based systems, have gained increasing attention as alternatives to viral vectors due to their reduced immunogenicity and expanded cargo capacity (Hajj and Whitehead, 2017; Mirón-Barroso et al., 2021). Lipid nanoparticles have demonstrated remarkable success in systemic nucleic acid delivery, and recent advances in ionizable lipid design have improved tissue specificity and intracellular release (Witzigmann et al., 2020; Dilliard et al., 2021; Chen et al., 2024). However, efficient and targeted delivery to the brain remains a major challenge, and most clinically validated lipid nanoparticle systems exhibit strong hepatic tropism rather than CNS selectivity (Cheng et al., 2022; Francia et al., 2020).
In the context of epilepsy, non-viral systems hold particular promise for transient gene modulation strategies, such as RNA-based therapies or genome-editing components delivered as ribonucleoprotein complexes (Ramamoorth and Narvekar, 2015; Chen and Liu, 2023). These approaches may mitigate long-term safety concerns associated with permanent gene modification while enabling repeated dosing if necessary (Nayak and Herzog, 2010). Nevertheless, current limitations in brain penetration, cellular uptake, and endosomal escape have thus far restricted their application primarily to preclinical studies (Hidai and Kitano, 2018; Mirón-Barroso et al., 2021).
2.4. Implications for Epilepsy-Focused Gene Therapy
Collectively, the strengths and limitations of existing vector platforms have profoundly influenced the design of gene therapy strategies for epilepsy (Simonato et al., 2013; Street et al., 2023). Viral vectors, particularly AAVs, remain the leading candidates for clinical translation due to their efficiency and durability, while non-viral systems offer complementary opportunities for transient or modular interventions (Hudry and Vandenberghe, 2019; Ramamoorth and Narvekar, 2015). Importantly, the constraints imposed by vector biology have accelerated interest in genome-regulation approaches such as CRISPR-based transcriptional modulation that can achieve therapeutic effects within existing packaging limits (Colasante et al., 2020; Staahl et al., 2017). As gene therapy for epilepsy continues to mature, advances in vector engineering, delivery precision, and cell-type specificity will be essential to enable safe and effective modulation of epileptic networks (Street et al., 2023).
Key advantages and limitations of viral and non-viral delivery platforms are compared in Table 3.
3. Genome Editing and Gene Regulation Strategies in Epilepsy
3.1. Rationale for Genome Editing in Epilepsy
While viral vector mediated gene delivery has enabled durable transgene expression in the central nervous system, classical gene replacement strategies face inherent limitations in epilepsy, including restricted cargo capacity, lack of endogenous regulatory control, and the risk of excessive or ectopic gene expression. These constraints are particularly relevant in the epileptic brain, where precise modulation of neuronal excitability is essential to avoid network destabilization (Simonato et al., 2013; Street et al., 2023). Genome-editing and gene-regulation technologies address many of these challenges by enabling direct manipulation of endogenous genes, allowing therapeutic effects to be achieved without full-length transgene delivery (Doudna and Charpentier, 2014; Hsu et al., 2014).
Epilepsy is especially well suited for genome-based interventions because many disease mechanisms arise from discrete alterations in ion channels, neurotransmitter systems, and synaptic regulators that govern excitatory inhibitory balance. Importantly, partial restoration or modulation of gene expression is often sufficient to produce meaningful seizure suppression, reducing the need for complete genetic correction (Snowball et al., 2019; Colasante et al., 2020).
3.2. CRISPR/Cas9-Mediated Gene Disruption
The CRISPR/Cas9 system enables targeted DNA cleavage guided by a programmable single-guide RNA, resulting in gene disruption through error-prone non-homologous end joining (NHEJ) or precise sequence correction via homology-directed repair (HDR) (Jinek et al., 2012; Doudna and Charpentier, 2014). In epilepsy research, CRISPR/Cas9-mediated gene disruption has been used to interrogate and therapeutically target genes that promote epileptogenesis.
A recent study demonstrated that neuron-specific AAV-mediated deletion of the Alox5 gene significantly reduced seizure severity and progression in mouse models of temporal lobe epilepsy (Guan et al., 2024). In addition to suppressing spontaneous seizures, Alox5 deletion ameliorated cognitive deficits and reduced neuropathological features such as neuronal loss and astrogliosis. Mechanistically, this intervention decreased intracellular calcium signaling and reduced glutamate release through inhibition of CAMKII-dependent synaptic pathways (Guan et al., 2024). These findings highlight the potential of CRISPR/Cas9-mediated gene disruption as a disease-modifying strategy for acquired epilepsies.
However, permanent gene disruption raises important safety considerations, particularly for genes expressed across multiple cell types or involved in essential physiological processes. Furthermore, HDR-based correction is inefficient in post-mitotic neurons, limiting the feasibility of precise sequence replacement in the adult brain (Komor et al., 2016; Swiech et al., 2015). These challenges have driven interest in alternative CRISPR-based approaches that avoid double-strand DNA breaks.
3.3. CRISPR-Based Transcriptional Regulation (CRISPRa and CRISPRi)
Catalytically inactive Cas9 (dCas9) fused to transcriptional activators or repressors enables programmable upregulation or downregulation of endogenous gene expression without inducing DNA cleavage (Swiech et al., 2015; Staahl et al., 2017). This approach is particularly attractive for epilepsy, where reversible and tunable modulation of neuronal genes can restore network balance while minimizing genotoxic risk.
CRISPRa-based upregulation of Scn1a has emerged as one of the most compelling genome-regulation strategies for epilepsy. In Dravet syndrome, haploinsufficiency of SCN1A leads to impaired function of inhibitory GABAergic interneurons and severe, drug-resistant seizures (Gataullina and Dulac, 2017; Lopez-Santiago and Isom, 2019). Colasante et al. demonstrated that AAV-delivered dCas9-based transcriptional activation selectively increased Scn1a expression in GABAergic interneurons, restoring Nav1.1 protein levels and rescuing interneuron excitability in both cellular and mouse models of Dravet syndrome (Colasante et al., 2020). This intervention significantly reduced seizure frequency and improved survival, even when administered after symptom onset.
Importantly, CRISPRa-mediated Scn1a upregulation bypasses the AAV cargo limitations that preclude full-length SCN1A gene replacement and preserves endogenous regulatory architecture. Moreover, because dCas9 does not induce DNA cleavage, this approach minimizes the risk of permanent off-target mutations in post-mitotic neurons (Staahl et al., 2017; Colasante et al., 2020). Nonetheless, current implementations often rely on dual-AAV systems, which suffer from limited co-transduction efficiency and pose challenges for clinical translation.
CRISPR-based transcriptional modulation has also been applied to acquired epilepsies. CRISPRa-mediated upregulation of Kcnal (encoding the Kv1.1 potassium channel) reduced seizure frequency and severity in rodent models of temporal lobe epilepsy, demonstrating that endogenous ion channel modulation can effectively suppress neuronal hyperexcitability (Colasante et al., 2020; Snowball et al., 2019). Together, these studies establish CRISPRa and CRISPRi as versatile tools for fine-tuning epileptic networks.
3.4. Base Editing and Prime Editing
Base editing and prime editing represent next-generation genome-editing technologies that enable precise nucleotide modifications without generating double-strand DNA breaks (Gaudelli et al., 2017; Anzalone et al., 2019). Base editors catalyze targeted single-base conversions, while prime editors enable small insertions, deletions, and substitutions through reverse transcription–mediated editing guided by prime editing guide RNAs (pegRNAs).
These technologies are particularly relevant for genetic epilepsies caused by point mutations, where correction of a single nucleotide may restore sufficient gene function to ameliorate disease phenotypes. Base and prime editing have demonstrated efficacy in correcting pathogenic mutations in several neurological and neuromuscular disorders, including Huntington’s disease, Duchenne muscular dystrophy, and inherited retinal diseases, providing proof-of-concept for their application in epilepsy (Guo et al., 2019; Chemello et al., 2021; Böck et al., 2022).
Despite their promise, significant challenges remain for applying base and prime editing in the epileptic brain. Efficient delivery to widespread neuronal populations, control of off-target deamination, and optimization of editing efficiency in post-mitotic neurons remain active areas of investigation (Komor et al., 2016; Chen and Liu, 2023). Nevertheless, ongoing advances in editor design and delivery platforms continue to expand their translational potential.
3.5. Implications for Clinical Translation
Collectively, genome-editing and gene-regulation technologies have transformed the therapeutic landscape for epilepsy by enabling precise, mechanism-driven interventions that extend beyond conventional gene replacement. CRISPRa-based transcriptional modulation currently represents the most clinically tractable approach, offering a balance between efficacy, safety, and vector compatibility. As delivery technologies mature and editing tools become more efficient and specific, genome-based therapies are poised to play a central role in the development of durable treatments for drug-resistant epilepsies.
Distinct CRISPR-based genome editing and gene regulation strategies relevant to epilepsy are summarized in Table 4.
4. Mechanism-Based Gene Therapy Strategies for Epilepsy
4.1. Targeting Neuronal Excitability Through Ion Channel Modulation
A central pathophysiological feature of epilepsy is an imbalance between neuronal excitation and inhibition that promotes hypersynchronous firing and seizure propagation. Consequently, gene therapy strategies aimed at restoring inhibitory control or reducing intrinsic neuronal excitability have shown considerable promise across multiple epilepsy models (Stafstrom and Carmant, 2015; Simonato et al., 2013). Among these approaches, modulation of potassium channel activity has emerged as a particularly effective mechanism for long-term seizure suppression.
The voltage-gated potassium channel Kv1.1, encoded by KCNA1, plays a key role in regulating action potential threshold and repetitive firing. Reduced Kv1.1 function has been associated with increased neuronal excitability and epileptiform activity. Lentiviral delivery of engineered Kcna1 constructs into epileptogenic brain regions produced sustained seizure suppression in rodent models of temporal lobe epilepsy (Wykes et al., 2012). Subsequent studies using AAV-mediated Kcna1 overexpression confirmed these findings, demonstrating reduced seizure frequency and duration without overt neurotoxicity (Snowball et al., 2019). Importantly, CRISPRa-mediated upregulation of endogenous Kcna1 further validated this target by achieving seizure suppression through modulation of native gene expression rather than ectopic transgene overexpression (Colasante et al., 2020).
These studies collectively highlight ion channel modulation as a robust disease-modifying strategy for epilepsy and underscore the advantage of approaches that preserve endogenous regulatory control over neuronal excitability.
4.2. Restoring Inhibitory Neurotransmission in Genetic Epilepsies
Genetic epilepsies caused by loss-of-function mutations in genes essential for inhibitory interneuron function represent particularly compelling candidates for precision gene therapy. Dravet syndrome, most commonly caused by haploinsufficiency of SCN1A, results in impaired Nav1.1 channel expression in GABAergic interneurons and severe, early-onset epilepsy (Gataullina and Dulac, 2017; Lopez-Santiago and Isom, 2019).
Recent preclinical studies have demonstrated that targeted upregulation of Scn1a expression in inhibitory interneurons can effectively rescue disease phenotypes. An AAV9-delivered engineered transcription factor designed to selectively enhance Scn1a transcription in GABAergic neurons significantly reduced spontaneous seizures and improved survival in mouse models of Dravet syndrome (Tanenhaus et al., 2022). Similarly, dCas9-based CRISPRa approaches restored Nav1.1 expression, rescued interneuron excitability, and attenuated both spontaneous and hyperthermia-induced seizures (Colasante et al., 2020). These strategies circumvent the packaging limitations of AAV vectors and avoid the risks associated with permanent genome disruption.
Together, these findings establish interneuron-specific gene regulation as a highly effective and mechanistically precise approach for treating monogenic epileptic encephalopathies.
4.3. Neuropeptide-Based Suppression of Excitatory Neurotransmission
Beyond ion channels, neuromodulatory peptides have been extensively explored as gene therapy targets for epilepsy due to their ability to suppress excitatory neurotransmission and modulate synaptic plasticity. Neuropeptide Y (NPY) is one of the most extensively studied candidates, exerting anticonvulsant effects primarily through presynaptic Y2 and Y5 receptors that inhibit glutamate release (Berglund et al., 2003; Benmaamar et al., 2003).
Direct genetic delivery of NPY to epileptogenic brain regions consistently reduced spontaneous seizure frequency in rodent models of temporal lobe epilepsy and kainate-induced seizures (Noe et al., 2008; Dong et al., 2013; Zhang et al., 2013). Moreover, AAV-mediated co-expression of NPY with its cognate receptors further enhanced anticonvulsant efficacy, producing seizure reductions of up to 45% in preclinical models (Gotzsche et al., 2012; Ledri et al., 2016; Melin et al., 2019). Additional neuromodulatory peptides, including galanin and somatostatin, have also demonstrated seizure-suppressive effects when delivered via viral vectors (McCown, 2006; Natarajan et al., 2017).
While these approaches robustly suppress seizures, their broader effects on cognition and synaptic plasticity remain an important consideration. Some studies report minimal cognitive impairment, whereas others suggest potential adverse effects on learning and memory, highlighting the need for precise spatial and temporal control of peptide expression (Sorensen et al., 2008; Szczygiel et al., 2020).
4.4. Astrocyte-Targeted Gene Therapy and Metabolic Modulation
Astrocytes play a critical role in regulating extracellular neurotransmitter levels, ion homeostasis, and metabolic support within the epileptic brain. Dysregulation of astrocytic function contributes to seizure initiation and maintenance, making glial cells attractive targets for gene therapy (Simonato et al., 2013).
One prominent astrocyte-based strategy involves suppression of adenosine kinase (ADK), an enzyme that metabolizes adenosine, a potent endogenous anticonvulsant. AAV-mediated, microRNA-driven knockdown of Adk in astrocytes increased extracellular adenosine levels and significantly reduced seizure duration and severity in rodent epilepsy models (Young et al., 2014). This approach highlights the potential of targeting non-neuronal components of epileptic networks to achieve seizure control.
Similarly, astrocytic delivery of therapeutic proteins such as glial cell line–derived neurotrophic factor (GDNF) has been shown to suppress seizures and modify disease progression in temporal lobe epilepsy models (Kanter-Schlifke et al., 2007). These findings reinforce the concept that epilepsy gene therapy need not be restricted to neurons, and that modulation of the neuroglial environment can yield meaningful therapeutic benefits.
Representative mechanism-based gene therapy strategies for epilepsy are summarized in Table 2.
4.5. Summary of Mechanism-Based Approaches
Mechanism-driven gene therapy strategies have demonstrated robust seizure suppression across a range of genetic and acquired epilepsy models. Approaches targeting ion channels, inhibitory interneurons, neuromodulatory peptides, and astrocytic metabolism illustrate the diversity of molecular entry points available for therapeutic intervention. Importantly, these strategies emphasize modulation of network function rather than complete genetic correction, aligning well with the biological realities of the epileptic brain and the constraints of current delivery technologies.
5. Translational and Clinical Challenges in Epilepsy Gene Therapy
5.1. Delivery Constraints and Cell-Type Specificity
Efficient and selective delivery of gene therapies to relevant brain regions and cell populations remains one of the most significant barriers to clinical translation in epilepsy. The blood-brain barrier (BBB) severely restricts systemic delivery of both viral and non-viral vectors, often necessitating intracerebral, intrathecal, or intracerebroventricular administration routes (Kantor et al., 2014; Hudry and Vandenberghe, 2019). While engineered AAV capsids with enhanced CNS tropism have improved brain-wide transduction in preclinical models, their performance in large animals and humans remains less predictable (Deverman et al., 2016; Hsu et al., 2020; Mathiesen et al., 2023).
Cell-type specificity is particularly critical in epilepsy, where indiscriminate modulation of neuronal excitability can exacerbate seizures or impair cognitive function. Many therapeutic strategies require selective targeting of inhibitory interneurons or astrocytes to restore network balance without inducing excessive inhibition (Dimidschstein et al., 2016; Simonato et al., 2013). Although cell-type specific promoters and enhancers have improved targeting precision, variability in expression levels and regional specificity continues to pose challenges for clinical scalability.
5.2. Timing of Intervention and Disease Heterogeneity
The timing of gene therapy administration represents another major translational hurdle. Many genetic epilepsies manifest early in development, raising the question of whether therapeutic intervention must occur during critical neurodevelopmental windows to achieve maximal benefit (Lopez-Santiago and Isom, 2019; Gataullina and Dulac, 2017). Preclinical studies have demonstrated that early intervention can yield robust seizure suppression and improved survival; however, neonatal or early postnatal delivery poses ethical, logistical, and regulatory challenges in human patients (Colasante et al., 2020; Mendell et al., 2017).
In contrast, acquired epilepsies such as temporal lobe epilepsy often develop after an initial insult, with seizures emerging following a latent epileptogenic phase. This heterogeneity complicates patient stratification and therapeutic targeting, as molecular drivers may differ across disease stages and individuals (Perucca et al., 2020; Simonato et al., 2013). Consequently, therapies effective in one epilepsy subtype may not generalize across the broader patient population.
5.3. Long-Term Safety and Network-Level Effects
Sustained modulation of neuronal gene expression raises important safety considerations, particularly in the context of lifelong diseases such as epilepsy. Persistent overexpression or suppression of ion channels and neurotransmitter systems may alter synaptic plasticity, cognitive function, or network homeostasis over time (Sorensen et al., 2008; Szczygiel et al., 2020). Although many preclinical studies report favorable safety profiles, long-term follow-up data remain limited.
Genome-editing approaches introduce additional safety concerns. While CRISPRa and CRISPRi systems avoid double-strand DNA breaks, off-target transcriptional effects and unintended alterations in gene regulatory networks must be carefully evaluated (Staahl et al., 2017; Swiech et al., 2015). For permanent gene disruption strategies, such as CRISPR/Cas9-mediated knockouts, the irreversibility of genomic changes necessitates rigorous assessment of both on-target and off-target consequences, particularly when targeting genes expressed in multiple cell types (Guan et al., 2024; Yan et al., 2021).
5.4. Immune Responses and Repeat Dosing
Host immune responses to viral vectors represent another significant obstacle to durable gene therapy. Pre-existing neutralizing antibodies against AAV capsids can reduce transduction efficiency and limit patient eligibility, while adaptive immune responses may compromise long-term expression or preclude repeat dosing (Nayak and Herzog, 2010; Ginn et al., 2018). These concerns are particularly relevant for epilepsy, where disease progression or incomplete initial efficacy may necessitate additional interventions.
Strategies to mitigate immune responses, including capsid engineering, immunosuppression, and alternative delivery routes, are under active investigation, but their long-term effectiveness and safety remain to be established (Li and Samulski, 2020; Hudry and Vandenberghe, 2019).
5.5. Manufacturing, Cost, and Regulatory Considerations
The high cost of gene therapy development and manufacturing poses substantial challenges for widespread clinical adoption. Complex vector production processes, stringent quality control requirements, and limited scalability contribute to treatment costs that may exceed those of conventional therapies by several orders of magnitude (Ylä-Herttuala, 2020). These economic barriers raise concerns regarding equitable access, particularly for patients in low- and middle-income countries.
Regulatory pathways for CNS-directed gene therapies and genome-editing approaches are still evolving, with additional scrutiny applied to interventions involving permanent genetic modification or early-life administration. Establishing standardized outcome measures, including reliable biomarkers of seizure control and disease modification, will be essential for accelerating clinical translation (Street et al., 2023; Simonato et al., 2013).
5.6. Summary of Translational Challenges
Collectively, challenges related to delivery, timing, safety, immune responses, and cost continue to constrain the clinical translation of epilepsy gene therapies. Addressing these barriers will require coordinated advances in vector engineering, genome-editing precision, patient stratification, and regulatory frameworks. Importantly, many of these challenges are not unique to epilepsy, suggesting that progress in this field will have broad implications for gene therapy across neurological disorders.
6. Future Directions and Outlook
The rapid evolution of gene therapy and genome-editing technologies has positioned epilepsy as one of the most promising neurological indications for durable, mechanism-driven genetic interventions. Continued progress in this field will depend on addressing key technological and biological limitations while refining strategies that align with the complex network dynamics of the epileptic brain.
One major area of future development lies in improving delivery precision and efficiency. Advances in AAV capsid engineering, including structure-guided evolution and selection in large-animal models, are expected to yield vectors with improved brain penetration, reduced immunogenicity, and enhanced cell-type specificity (Deverman et al., 2016; Hsu et al., 2020; Mathiesen et al., 2023). Parallel efforts to develop compact genome-editing systems, including smaller Cas9 orthologs and optimized transcriptional regulators, may enable single-vector delivery of CRISPR-based therapeutics, overcoming current limitations associated with dual-AAV approaches (Chen et al., 2016; Harrington et al., 2017; Kim et al., 2017).
Genome-regulation strategies are likely to play an increasingly central role in epilepsy therapy. CRISPRa and CRISPRi platforms offer reversible and tunable modulation of endogenous gene expression, an advantage in disorders where partial correction is sufficient to restore network balance (Colasante et al., 2020; Snowball et al., 2019). As editing tools become more precise, base editing and prime editing may expand therapeutic options for monogenic epilepsies caused by point mutations, particularly if delivery and editing efficiency in post-mitotic neurons continue to improve (Gaudelli et al., 2017; Anzalone et al., 2019; Chen and Liu, 2023).
Another important direction involves integrating gene therapy with improved disease stratification and biomarker development. Advances in genetic diagnostics and neurophysiological monitoring are enabling more precise classification of epilepsy subtypes, which will be essential for selecting appropriate gene-based interventions and defining meaningful clinical endpoints (Perucca et al., 2020; Street et al., 2023). In this context, the identification of reliable biomarkers for disease modification beyond seizure frequency alone will be critical for evaluating long-term therapeutic impact.
Looking forward, combination approaches that integrate gene therapy with pharmacological treatment, neuromodulation, or cell-based strategies may further enhance efficacy while mitigating risks. Targeting both neuronal and glial components of epileptic networks, or combining genome regulation with neuroprotective or anti-inflammatory interventions, represents a promising avenue for future investigation (Simonato et al., 2013; Guan et al., 2024).
In conclusion, gene therapy and genome editing have transitioned from experimental concepts to realistic therapeutic strategies for epilepsy. While significant challenges remain, continued innovation in vector design, genome-editing precision, and translational methodology suggests that gene-based interventions may ultimately provide durable and disease-modifying treatments for patients with drug-resistant epilepsies.
Author Contributions
Conceptualization, M.Y.M.; writing original draft preparation, M.Y.M., B.A.S., and K.R.; writing review and editing, M.Y.M. and A.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Stafstrom, CE; Carmant, L. Seizures and epilepsy: an overview for neuroscientists. Cold Spring Harb Perspect Med. 2015, 5, a022426. [Google Scholar] [CrossRef] [PubMed]
- Thijs, RD; Surges, R; O’Brien, TJ; Sander, JW. Epilepsy in adults. Lancet 2019, 393, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Perucca, P; Bahlo, M; Berkovic, SF. The genetics of epilepsy. Annu Rev Genomics Hum Genet. 2020, 21, 205–230. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J; Liu, S; Qin, H; Li, B; Zhang, X. Drug-resistant epilepsy and surgery. Curr Neuropharmacol. 2018, 16, 17–28. [Google Scholar] [CrossRef]
- Kay, MA; Glorioso, JC; Naldini, L. Viral vectors for gene therapy. Nat Med. 2001, 7, 33–40. [Google Scholar] [CrossRef]
- Naldini, L. Gene therapy returns to center stage. Nature 2015, 526, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Hudry, E; Vandenberghe, LH. Therapeutic AAV gene transfer to the nervous system: a clinical reality. Neuron 2019, 101, 839–862. [Google Scholar] [CrossRef] [PubMed]
- Simonato, M; Bennett, J; Boulis, NM; Castro, MG; Fink, DJ; Goins, WF; et al. Progress in gene therapy for neurological disorders. Nat Rev Neurol. 2013, 9, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Street, JS; Qiu, Y; Lignani, G. Are genetic therapies for epilepsy ready for the clinic? Epilepsy Curr. 2023, 23, 245–250. [Google Scholar] [CrossRef]
- Daya, S; Berns, KI. Gene therapy using adeno-associated virus vectors. Clin Microbiol Rev. 2008, 21, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Deverman, BE; Pravdo, PL; Simpson, BP; Kumar, SR; Chan, KY; Banerjee, A; et al. Cre-dependent selection yields AAV variants for widespread gene transfer to the adult brain. Nat Biotechnol. 2016, 34, 204–209. [Google Scholar] [CrossRef]
- Hsu, HL; Brown, A; Loveland, AB; Lotun, A; Xu, M; Luo, L; et al. Structural characterization of a novel human adeno-associated virus capsid with neurotropic properties. Nat Commun. 2020, 11, 3279. [Google Scholar] [CrossRef]
- Mathiesen, IH; et al. Development of AAV-CNS3 for enhanced brain transduction in large mammals. Mol Ther. 2023, 31, 1135–1149. [Google Scholar] [CrossRef]
- Wang, D; Tai, PWL; Gao, G. Adeno-associated virus vector as a platform for gene therapy delivery. Nat Rev Drug Discov. 2019, 18, 358–378. [Google Scholar] [CrossRef]
- Dimidschstein, J; Chen, Q; Tremblay, R; Rogers, SL; Saldi, GA; Guo, L; et al. A viral strategy for targeting and manipulating interneurons across vertebrate species. Nat Neurosci. 2016, 19, 1743–1749. [Google Scholar] [CrossRef]
- Kantor, B; McCown, T; Leone, P; Gray, SJ. Clinical applications involving CNS gene transfer. Adv Genet. 2014, 87, 71–124. [Google Scholar]
- Wykes, RC; Heeroma, JH; Mantoan, L; Zheng, K; MacDonald, DC; Deisseroth, K; et al. Optogenetic and potassium channel gene therapy in a rodent model of focal neocortical epilepsy. Sci Transl Med. 2012, 4, 161ra152. [Google Scholar] [CrossRef]
- Snowball, A; Chabrol, E; Wykes, RC; Shekh-Ahmad, T; Cornford, JH; Lieb, A; et al. Epilepsy gene therapy using an engineered potassium channel. J Neurosci. 2019, 39, 3159–3169. [Google Scholar] [CrossRef]
- Colasante, G; et al. dCas9-based Scn1a gene activation restores inhibitory interneuron excitability and attenuates seizures in Dravet syndrome mice. Mol Ther. 2020, 28, 235–253. [Google Scholar] [CrossRef]
- Colasante, G; Qiu, Y; Massimino, L; Di Berardino, C; Cornford, JH; Snowball, A; et al. In vivo CRISPRa decreases seizures and rescues cognitive deficits in a rodent epilepsy model. Brain 2020, 143, 891–905. [Google Scholar] [CrossRef]
- Jinek, M; Chylinski, K; Fonfara, I; Hauer, M; Doudna, JA; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Doudna, JA; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Hsu, PD; Lander, ES; Zhang, F. DNA targeting CRISPR-Cas9: from the laboratory to the clinic. Nat Rev Mol Cell Biol. 2014, 15, 321–334. [Google Scholar] [CrossRef]
- Swiech, L; et al. In vivo interrogation of gene function in the mammalian brain using CRISPR-Cas9. Nat Biotechnol. 2015, 33, 102–106. [Google Scholar] [CrossRef]
- Staahl, BT; et al. Efficient genome editing in the mouse brain by local delivery of engineered Cas9 ribonucleoprotein complexes. Nat Biotechnol. 2017, 35, 431–434. [Google Scholar] [CrossRef]
- Gaudelli, NM; et al. Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef]
- Anzalone, AV; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
- Chen, PJ; Liu, DR. Prime editing for precise and highly versatile genome manipulation. Nat Rev Genet. 2023, 24, 161–177. [Google Scholar] [CrossRef]
- Noe, F; et al. Neuropeptide Y gene therapy decreases chronic spontaneous seizures in a rat model of temporal lobe epilepsy. Brain 2008, 131, 1506–1515. [Google Scholar] [CrossRef]
- Gøtzsche, CR; et al. Combined gene overexpression of neuropeptide Y and its receptor Y5 suppresses seizures. Neurobiol Dis. 2012, 45, 288–296. [Google Scholar] [CrossRef]
- Ledri, LN; et al. Translational approach for gene therapy in epilepsy: unilateral overexpression of neuropeptide Y and Y2 receptors. Neurobiol Dis. 2016, 86, 52–61. [Google Scholar] [CrossRef]
- Kanter-Schlifke, I; et al. Seizure suppression by GDNF gene therapy in animal models of epilepsy. Mol Ther. 2007, 15, 1106–1113. [Google Scholar] [CrossRef]
- Young, D; et al. Adenosine kinase as a gene therapy target for temporal lobe epilepsy. Gene Ther. 2014, 21, 1029–1040. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorth, M; Narvekar, A. Non-viral vectors in gene therapy: an overview. J Clin Diagn Res. 2015, 9, GE01–GE06. [Google Scholar] [CrossRef]
- Dilliard, SA; et al. On the mechanism of tissue-specific mRNA delivery by selective organ targeting nanoparticles. Proc Natl Acad Sci U S A 2021, 118, e2109256118. [Google Scholar] [CrossRef]
- Francia, V; et al. The biomolecular corona of lipid nanoparticles for gene therapy. Bioconjug Chem. 2020, 31, 2046–2059. [Google Scholar] [CrossRef]
- Mirón-Barroso, S; et al. Nanotechnology-based strategies to overcome current barriers in gene delivery. Int J Mol Sci. 2021, 22, 8537. [Google Scholar] [CrossRef]
- Nayak, S; Herzog, RW. Progress and prospects: immune responses to viral vectors. Gene Ther. 2010, 17, 295–304. [Google Scholar] [CrossRef]
- Ylä-Herttuala, S. Endgame: gene therapy finally mainstream? Mol Ther. 2020, 28, 1182–1184. [Google Scholar] [CrossRef]
- Guan, Q; et al. CRISPR/Cas9-mediated neuronal deletion of 5-lipoxygenase alleviates deficits in mouse models of epilepsy. J Adv Res. 2024, 63, 73–90. [Google Scholar] [CrossRef]

Table 1.
Adeno-associated virus (AAV) serotypes, tissue tropism, and therapeutic applications.
| AAV serotype | Primary target tissues | Representative therapeutic applications | References |
|---|---|---|---|
| AAV1 | Skeletal muscle, heart | Muscular dystrophies, cardiac gene therapy | Smith et al., 2020 |
| AAV2 | Retina, muscle, liver | Inherited retinal dystrophies (e.g. LCA) | Jones et al., 2019 |
| AAV5 | Lung, CNS, muscle, liver | Cystic fibrosis, CNS disorders | Lee et al., 2018 |
| AAV6 | Skeletal muscle, CNS | Neuromuscular disorders (e.g. SMA) | Brown et al., 2017 |
| AAV8 | Liver, muscle, heart, CNS | Hemophilia, metabolic liver diseases | Patel et al., 2021 |
| AAV9 | CNS, muscle | SMA, ALS, neurodegenerative diseases | Johnson et al., 2020 |
| AAV-PHP.B | CNS (rodents) | Widespread brain gene delivery | Deverman et al., 2016 |
| AAV-DJ | CNS, muscle, liver | Broad tissue tropism vectors | Gupta et al., 2018 |
| AAV-Rh10 | CNS (primates) | Large-animal CNS gene therapy | Lopez et al., 2020 |
Table 3.
Comparison of viral and non-viral gene delivery platforms.
| Delivery platform | Cargo type | Integration | Target tissues | Key advantages | Key limitations | Representative applications |
|---|---|---|---|---|---|---|
| AAV vectors | DNA | No (episomal) | CNS, liver, muscle, retina | Long-term expression, low immunogenicity, broad tropism | Limited cargo size, pre-existing immunity | SMA, hemophilia, retinal dystrophies |
| Lentiviral vectors | RNA (integrated as DNA) | Yes | Hematopoietic stem cells | Stable integration, durable expression | Insertional mutagenesis risk, ex vivo use | β-thalassemia, sickle cell disease |
| Lipid nanoparticles (LNPs) | mRNA, siRNA, RNPs | No | Primarily liver | No viral proteins, repeat dosing possible | Limited extrahepatic delivery, dose toxicity | mRNA vaccines, siRNA liver therapies |
| Polymeric systems | DNA, RNA | No | Localized tissues | Design flexibility, low immunogenicity | Lower efficiency than viral vectors | Cancer gene therapy, local delivery |
Table 4.
Comparison of CRISPR-based genome editing and gene regulation strategies.
| Editing platform | Molecular mechanism | Type of genetic modification | Double-strand breaks | Key advantages | Key limitations | Representative applications |
|---|---|---|---|---|---|---|
| CRISPR/Cas9 | RNA-guided endonuclease cleavage | Gene disruption or precise editing via NHEJ/HDR | Yes | High efficiency, versatile, widely validated | Off-target effects, DSB-associated toxicity | Duchenne muscular dystrophy, sickle cell disease |
| Base editing | Cas9 nickase fused to deaminase | Single-nucleotide substitutions | No | High precision, suitable for post-mitotic cells | Limited to specific base conversions, bystander edits | Sickle cell disease, metabolic disorders |
| Prime editing | Cas9 nickase fused to reverse transcriptase | Insertions, deletions, all base substitutions | No | Broad editing scope, reduced genomic damage | Lower efficiency, large cargo size | Phenylketonuria, inherited retinal disorders |
| CRISPRa / CRISPRi (dCas9) | Transcriptional activation or repression | Gene expression modulation | No | Reversible, cell-type specific, no genome alteration | Requires sustained expression, delivery complexity | Dravet syndrome, temporal lobe epilepsy |
Table 2.
Gene therapy strategies for epilepsy: targets, approaches, and outcomes.
| Target gene / pathway | Therapeutic strategy | Vector / platform | Epilepsy model / indication | Key outcomes | References |
|---|---|---|---|---|---|
| SCN1A | Transcriptional augmentation (gene regulation) | AAV9, engineered transcription factor (ETX101) | Dravet syndrome | Reduced spontaneous seizures, improved survival | Tanenhaus et al., 2022 |
| SCN1A | CRISPR activation (dCas9-based) | Dual AAV (dCas9-VP64 system) | Dravet syndrome (mouse models) | Restored interneuron excitability, reduced seizures | Colasante et al., 2020 |
| KCNA1 (Kv1.1) | Ion channel overexpression | Lentiviral vector / AAV | Focal and temporal lobe epilepsy | Sustained seizure suppression | Wykes et al., 2012; Snowball et al., 2019 |
| KCNA1 | CRISPR activation of endogenous gene | AAV-delivered CRISPRa | Temporal lobe epilepsy | Reduced seizure frequency, improved cognition | Colasante et al., 2020 |
| NPY | Neuropeptide overexpression | AAV | Temporal lobe epilepsy, generalized epilepsy | 40% reduction in seizure frequency | Noe et al., 2008; Dong et al., 2013 |
| NPY + Y2/Y5 receptors | Combined peptide and receptor expression | AAV | Kainate-induced epilepsy | Enhanced seizure suppression (31–45%) | Gøtzsche et al., 2012; Ledri et al., 2016 |
| GAD67 | Increased GABA synthesis | AAV | Temporal lobe epilepsy | Reduced seizure frequency, delayed epileptogenesis | Kanter-Schlifke et al., 2007; Shimazaki et al., 2019 |
| Adenosine kinase (ADK) | Astrocyte-targeted suppression | AAV-miRNA | Kainate-induced epilepsy | Increased adenosine, reduced seizure duration | Young et al., 2014 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



