
Submitted:
06 February 2026
Posted:
09 February 2026
You are already at the latest version
Abstract
This study explored the Ultrastructural and oxidative stress-related genes and proteins changes in the laminar tissue of dairy cows with oligofructose (OF)-induced laminitis. Twelve clinically healthy, non-pregnant Chinese Holstein cows were randomly allocated into two groups: OF-overload group (n = 6) and Control group (n = 6). 17g/kg BW of oligofructose (OF) dissolved in 20 mL/kg BW of deionized water was provided to the OF-treated group, while the control group received 20 mL/kg BW of deionized water via a stomach tube. Laminar tissue samples were collected after euthanizing cows at 72 h. We observed that the gene and protein expression of Nrf2, Ho1, and Nqo1 significantly decreased, while keap1 significantly increased in the OF group as compared to the control group. Moreover, the distribution of Keap1 expression significantly increased, while that of the Nrf2 significantly decreased in the OF group relative to the control group. However, in the OF group, the lamina densa appears thick and damaged, with interrelated collagen fibers, and lightly stained; number of hemidesmosomes on the cell membrane decreased; the distance between the basal cells and the epidermal lamellae increased; epidermal basal cells have deformed nuclei with reduced chromatin than in control cows. In conclusion, unbalanced gene and protein status may be the stem cause for the epidermal detachment which confirmed the increased oxidative stress in the OF group.
Keywords:
acute laminitis
; dairy cows
; laminar tissue
; oligofructose
; oxidative stress
; ultrastructure
1. Introduction
Bovine laminitis is a hoof disease that causes lameness [1]. Laminitis was first identified as diffuse aseptic pododermatitis, which is characterized by inflammation of the dermal layers within the hoof [2]. The cause of laminitis is multifactorial with excessive ingestion of fast-digesting carbohydrates being one of the major contributing factors [3]. There have been many economic losses caused by laminitis that may dramatically limit the growth of dairy farming. In the past decade, many studies have focused on the pathophysiology of milk cow laminitis [4,5], metabolomics [6], histology [7], and proteomics [8]. However, due to differences in clinical investigations and a lack of valid laboratory models [9], the root cause and pathophysiology of acute bovine laminitis remain unknown. In clinical practice, bovine laminitis is classified as a secondary cause of several inflammatory illnesses, including ruminal acidosis, mastitis, metritis, and gram-negative pleuropneumonia [10]. By simulating such clinical conditions, investigational models of dairy cow laminitis have been formulated. The OF-induced model is more precise and more frequently applied than the some other models of inducement [11,12], and confirmed that this model has identical clinical presentations and typical histopathological changes to the acute laminitis cases [13,14]. Previously, the use of the OF-induced model in equine laminitis studies was more prevalent than in cow laminitis [15,16]. Thus, this study was based on the successful induction of laminitis in dairy cows via oligofructose, for the first time to investigate the oxidative stress response at the gene and protein level in the laminar tissue of the dairy cow’s claw.
The researchers have discovered the evidence that dairy cow laminitis influences the structural integrity of the lamellae. These tissues are pertinent to offer proper orientation of the third phalanx (P3) and transmit major mechanical forces to the hoof when they attach P3 to the hoof capsule [17]. The lamellae structure is organized into epidermal lamellae and dermal lamellae, linked by contact zone called the basement membrane (BM) [5]. When epidermal and dermal lamellae are disrupted, P3 sinks and turns within the capsule resulting in severe lameness and pain [18]. The extracellular matrix (ECM) is a main component of the BM, which is a boundary between two different layers. The two principal histological alterations that are considered to result in epidermal detachment in bovine laminitis are BM destruction and separation [19,20]. However, the underlying moleculer mechanism remains unclear.
Oxidative damage is initiated by an imbalance in the levels of reactive oxygen species (ROS) and a lack of antioxidant capacity. Increased ROS under oxidative stress causes damage to cellular macromolecules, leading to protein alteration, lipid peroxidation and DNA damage [21,22], resulting in hoof tissue dyskeratosis [23] and programmed chondrocyte death [24]. Current research suggests that oxidative stress is closely related to bovine laminitis [25,26,27]. In the face of deleterious signals, the body had an adjusted oxidant scavenging system. The role of this step has been demonstrated by several recent studies and both Keap1 and Nrf2 are important proteins in cellular defense towards oxidative stress. Second phase enzymatic systems characterized by the presence of Nqo1 and Ho-1 exert antioxidant, antiinflammatory activity in conditions predisposing to oxidative stress [28]. This is mediated by the binding of Nrf2 to an antioxidant response element (ARE) sequence [29]. Downstream genes of Nrf2 such as Ho-1 and Nqo1 also have antioxidant activity [30]. Keap1 is a cytoplasmic chaperone protein and secondary structure Nrf2-regulating domain that is specifically related to the inhibition of Nrf2 by receptors. Nrf2 is negatively regulated by Keap1 [31]. Nrf2 is sequestered in the cytoplasm under basal conditions and attaches to the actin cytoskeleton via Keap1, facilitating its proteasomal degrading. In the presence of oxidative stimulation, Nrf2 and Keap1 dissociate, and then Nrf2 migrates into the nucleus, where Nrf2 triggers the expression of genes encoding important antioxidant molecules and antioxidant enzymes by binding elements of the antioxidant response [29]. Nrf2-mediated regulation of cellular antioxidant and anti-inflammatory mechanisms seems to be significant in protection against oxidative stress [32].
To date, the oxidative stress effectiveness of laminar tissue in lactating cows with OF-induced laminitis have not been reported. Thus, this experiment study was intended to investigate the laminar tissue oxidative stress status with OF-induced bovine laminitis at both gene and protein profiles, with the goal of identifying relevant therapy avenues for this condition in future studies. We suggested that an oral OF challenge induces oxidative stress and ultra-structural alterations in the laminar wall of bovine claw.
2. Materials and Methods
2.1. Experimental Animals
Twelve clinically sound non-pregnant Chinese Holstein cows, which have normal locomotion [33], and have not received a history of claw horn lesions were used in this experiment. Cows body weight ranged from 335-403 kg (379.71±19.87 kg), age ranged from 18-26 months (20.67±3.01 mo) and BCS [34] ranged from 2.7-3.3 (3.00±0.23). All the cows selected in this experiment bought from the Qingxi dairy farm in Xiangfang District Harbin, P.R. China. The study’s cows were placed in the animal shelter with a rubber floor for thirty days prior to the trial. The cows had unlimited access to assorted forages ad libitum and a sufficient supply of clean drinking water. The experimental cows were recorded for their hoof temperature, body temperature, and blood pressure, as well as walk for 5 min, daily to assess their health.
2.2. Experimental Design and Treatment
Twelve dairy cows were randomly categorized into two groups, including: OF-treated group (n = 6) and Control group (n = 6). 17g/kg BW of oligofructose (Bailong Biotech, Inc., Dezhou, Shandong, China) dissolved in 20 mL/kg BW of deionized water was administered to the OF-treated group, and 20 mL/kg BW of deionized water was administered to the Control group at 0 h using a stomach tube (length 2.2 m, diameter 25 mm) in compliance with the protocol reported by [11,12]. Oligofructose (5%) was provided orally once daily for 3 days. Cow were trained to the clinical examination before receiving the advanced oral dose of OF.
Claudication assessments were performed at -72, 0, 6, 12, 18, 24, 36, 48, 60, and 72 h. During this time, cows experienced to walk and trot in a straight pathway by hand before turning in a small circle on the same ground of the Animal Hospital at Northeast Agricultural University in Harbin, China. Five certified veterinarians rated each cow's claudication scores using the protocol described by Sprecher et al. [33]. Once all certified vets scored a score of ≥2, the cow was classified as lame. On each 6 h, all cows were recommended to the clinical evaluation, such as respiratory rate, heart frequency range, the rectal temperature, eating routine, and feces consistency, hoof coronary band, weight shift, diastolic blood pressure, hoof temperature, and rumen movements, hoof discomfort, rumen pH. After 72 h of OF-overloading, each cow was euthanized by overdosing (20 mg/kg) with pentobarbital sodium and phenytoin sodium (Fatal-plus; 20 mg/kg IV) by intravenous injection [13]. In accordance with cow welfare guidelines, supportive therapy with Ringer lactate (15 mL/kg of BW; Heping Animal Medicine Co., Ltd. Harbin, China) at 18 and 24 h and calcium borate (14 mg of Ga/mL; 1.4 mL/kg of BW; Heping Animal Medicine Co., Ltd. Harbin, China) at 18 h was provided post-OF administration.
2.3. Laminar Tissue Sampling
In accordance with the protocol presented by Thoefner et al. [13], the deceased animals' left hind claws were separated in 5-10 min, positioned in an iced pack, and inmmediately brought transported to the laboratory. Laminar tissue was exposed by cutting the hoof wall, and hoof capsule was detached. Subsequently, a laminar tissue was cut into small tissue sections (1-2 mm2). It was immediately frozen in a liquid nitrogen and stored at -80˚C. The whole process was carried out in ice. To avoid tissue exposure, proper gloves and sterile mask were used. Hoof capsule contains the laminar wall tissue which is closely attached to it. The lamella wall typically encompasses the whole of the axial, abaxial, and dorsal side of the claw, extending between 3-7 cm between the coronet and the sole corium. The dissociated lamella wall which is excised 2cm below the coronet encompasses the axial and abaxial elements of the laminar wall tissue. The Ideal laminar wall tissue dimension is of length (6 x 3 cm) and width (3 cm).
2.4. Transmission Electron Microscopic Structure Observation of Laminar Tissue of Dairy Cows
The ultrastructure of hoof tissue in dairy cows with laminitis was observed using a transmission electron microscope. The precise operations steps were as follow: The small pieces of hoof tissue were removed, fixed in 2.5% glutaraldehyde solution, re-fixed with 1% osmium oxide solution at 4˚C for 2 hours, dehydrated with gradient acetone, and then embedded in 812 epoxy resin. At 70˚C, the tissue block was polymerized in resin for 2 days, and the tissue was cut into 65-70 nm thick slices using a UC6 ultra-microtome and stained with uranyl acetate and lead citrate. An H-7650 electron microscope was then used to observe and measure the ultrastructure of hemidesmosomes, organelles, and nuclei of the basal cells of the epidermis of the hoof leaf, as well as the distance from the basal cells to the compact layer of the hoof leaf. Basement membrane hemidesmosomes counting method is performed as follows: at approximately 30,000 magnifications, 100 μm of hoof tissue basement membrane of each cow in the both groups were continuously photographed, the hemidesmosomes on the basement membrane were observed, and the number of hemidesmosomes per micron was calculated. The distance from the basal cells to the dense layer of the hooves was measured as follows: A measurement point every 1 μm was placed on the 100 μm long basement membrane to measure the distance perpendicular to the direction of the basal cell membrane from the cell membrane to the central area of the hoof dense layer.
2.5. RNA Isolation and cDNA Synthesis
The total RNA was taken from laminar tissue of 12 cows via the RNA Miniprep-Kit (Invitrogen, Carlsbad, USA) in compliance with the directions given by the manufacturer. The tissue samples (100 mg) were mixed with 1 mL TRIzol reagent (Invitrogen, Carlsbad, USA). Then it was put into non-DNAse/RNAse centrifuge and total RNA without DNA, protein and isopropanol precipitates was removed using chloroform and washed with 75% ethanol. The quality and quantity of separated total RNA were assessed with an ultra nucleic acid protein testing kit (NO-ONE, Gene Company, USA). The accuracy of each RNA sample was checked by 1% agarose gel electrophoresis (Bio-Rad Laboratories, Hercules, CA, USA). RNA samples were diluted to 1 μg/ μL through optical density measurement. 1ug total RNA was isolated from each sample in line with protocols established by Prime-Script™ RT Kit manufacturer (Takara, Dalian, Liaoning, China). Complementary DNA (cDNA) was obtained using reverse transcription. During Real-time RT-qPCR analysis, the resulting cDNA was diluted (1:3) with DEPC water and kept at-20˚C till further utilization. DEPC water was prepared using dH2O and DEPC after which it was sterilized to eliminate DEPC.
2.6. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
In this experiment, primers were prepared to identify the genes, including Keap1, Nrf2, Ho1, Nqo1, and GAPDH, which were designed through Shanghai Sheng Gong Biotechnology, Co., Ltd. (BBI Life Sciences, Shanghai, China) (Table 1). The efficiency of each of the primer sequences was assessed using the Blast Computer Program of the (NCBI) National Center of Biotechnology Information database (Bethesda, Maryland, USA).
The RT-qPCR experiment was carried out with the Green chimeric florescence detection procedure of SYBR Premix Ex Taq TM ‖ Kit (Takara, Dalian, Liaoning, China) by the Light Cycler 480 RT-qPCR system (Roche, Germany). The PCR reaction mixture (20 μL final volume) contains 2 μL cDNA template, and 18 µL of the major blend to be used in the PCR. Subsequently the PCR master cocktail was composed of 6.4 μL of DEPC water, 10 μL of SYBR green florescent agent, and 1.6 μL of general primer solution (10 μM each of forward and reverse primers). The end primer concentration was 0.4 μM/ μL. The PCR protocols are as follows: pre-denaturation, 1 cycle, 95°C, 1 min; quantitative estimation, 40 cycles of 95°C 5s and 60°C 1 min; melting curve analysis, 95°C 5s, 60°C 1min, 95°C 1 cycle; cooling, 1 cycle at 50°C for 30s. Ct values of the different genes were calculated using Light cycler 480 software 2.0 (Roche, Germany) and Abs Quant/Fit points method. PCR efficiency of the individual genes was calculated using △△Ct technique with the GAPDH being used as the internal reference gene.
2.7. Western Blot
The conserved laminar tissue (100 mg) was added to RIPA lysis buffer (Beyond Biotech, Shanghai, China). The primary constituents of RIPA buffer were deoxycholate (1%), Triton X-100 (1%), and sodium lauryl sulfate (0.1%). Following the administration of 10 μL of phenylmethylsulfonyl fluoride (protease inhibitor, Beyond Biotech, Shanghai, China), the tissue was processed in a grinder (4°C, 4 min), and then spun (12,000rcf/min, 15 min). The concentration of protein was assessed using the BCA technique. The protein volume was determined in the tissue sample using SDS-PAGE where 10 μL of protein sample was added to the individual well. The overall concentration of protein was estimated as 25 μg. The target proteins were transferred to nitrocellulose (NC) filter membranes after isolation on 10% polyacrylamide gel following the semi-dry method (300 mA, 1.5 h). To block the membrane, it was gently agitated in 5% nonfat milk in TBS (Tris-buffer saline) with 0.1% Tween-20 (TBST) for 2 h at room temperature. The blocked membranes were then subjected to primary antibodies comprising Nrf2 (1:750), Keap1 (1:500), Ho1 (1:500), Nqo1 (1:750), and a 1:2000 dilution of β-actin (BIOSS Antibodies Beijing Biosynthesis Biotechnology Co., LTD. P.R. China), then incubated at 4°C for 12 hours and washed three times with TBST (15 min per wash). It was then combined with secondary antibody (HRP-labeled goat anti-rabbit IgG) at ratio 1:10,000 (in 1 X TBST, BBL Life Sciences, Shanghai, China) and kept at room temperature (2 h) followed by shaking (2 h, room temperature). NC membrane was also rinsed three times with TBST (15 min) through rocking. Then, it was put on the plate of a TANON 5200 exposure apparatus, and filter paper was used to remove the liquid. The ECL liquid (Meilun Biotechnology Co., Ltd., Dalian, China) was then dropped to uniformly overlay the protein bands. An image of the western blot was taken with its corresponding exposure time. ImageJ software was applied to identify the gray value of each protein in each gel, and the gray ratio of the target protein band to the internal reference protein band was determined in order to identify the relative expression of the target protein.
2.8. Immunohistochemistry
Immunohistochemistry was utilized to assess the expression of Keap1 and Nrf2 in laminar tissue. The samples were cut into the proper sizes, fixated (4% paraformaldehyde, 24 h), and then sliced and implanted. The tissue sections were paraffinized by overnight incubation at 80 °C, followed by a 10-minute incubation in 3%t H2O2 in the dark to inactivate endogenous peroxidase activity and antigen retrieval was done in a pressure cooker with sodium citrate buffer. The resulting fragments were incubated with bovine serum albumin (20 min, room temperature), primed overnight (4℃) with a primary antibody mixture (1: 200) dilution against Keap1 and Nrf2, Novus Biologicals, USA), and then incubated with streptavidin-conjugated horseradish peroxidase (30 min, room temperature). This was subsequently glued with neutral glue and stained with hematoxylin and DAB and stored in an oven. Lastly, the stained tissue segment in each group was assessed under a microscope and analyzed using Image-Pro Plus 6.0 software (Media Cybernetics, USA).
2.9. Statistical Analysis
The data analysis was carried out with the GraphPad Prism software (Version 8.01, GraphPad software Inc., San Diego, CA, USA). The analysis of the research data was performed with normal distribution. Changes in relative gene expression were calculated by the 2-△△Ct method. The data met the assumption of Gaussian distribution for both groups using the Shapiro-Wilk test in GraphPad prism. For the evaluation of local histology investigation results, an independent student-t test was employed to evaluate the differences between the two groups. While Bonferroni's multiple comparison tests were done across both groups for the clinical data such as lame scor [35]. Differences were considered when P<0.05. All data were showed as mean ± SD.
3. Results
3.1. Clinical Manifestation of Dairy Cows Laminitis
All dairy cows treated with OF had clinical manifestations of unique acute ruminal and systemic acidosis like persistent profuse diarrhea, in-appetence, absent dietary intake, depression, anorexia, swelled of carpal (tarsal) joints, inflamed hoof coronary band, disturbed weight shift, elevation of heart rate, elevated diastolic blood pressure, high hoof temperature, elevated body temperature, slow respiration, apathy of digital (toe) arteries, hoof pain, reduced rumen pH, intermittent fever, and lameness [25,27,35]. There were no signs of a systemic illness in the control cows. To examine claudication, clinical indications of laminitis were firstly assessed at 24 h following OF overload and continued to change until maximum claudication score of 3-5 at 60 h to 72 h [36] were found that confirm the acute laminitis. These symptoms were all similar to those reported by [11,12,13].
3.2. Oxidative Stress-Associated Genes Expression in Laminar Tissue of Laminitis Dairy Cows
The gene expressions of oxidative stress-associated genes were assessed by RTq-PCR. The data indicated that the expression level of Keap-1 increased significantly in the OF group compared to the control group. However, Nrf-2, Ho-1, and Nqo-1 expression in the OF group was considerably lower than in the control group (Figure 1). These results indicate that oral OF treatment induces oxidative stress in the laminar tissue damage of acute laminitis in dairy cows.
3.3. Oxidative Stress-Associated Proteins Expression in Laminar Tissue of Laminitis Dairy Cows
Western blot examination revealed that the expressions of Nrf2, Nqo1, and Ho1 were considerably lower in the OF group's laminar tissue relative to the control group. However, the Keap1 expression increased significantly in the OF group's laminar tissue compared to the control group (Figure 2). Our research showed that oral OF treatment induced oxidative stress in laminar tissue and suggested the involvement of ROS-mediated inflammation as an important pathway leading to laminitis.
The immunohistochemical analysis of Keap-1 protein expression in cow laminar layers revealed that mean Keap-1 staining within the cytoplasm of laminar tissue in the OF-treated group measured approximately 7.36%, while staining measured around 2.41% in control tissue. Statistically, keap-1 protein expression was significantly enhanced in the cytoplasm of laminar tissue in the OF group compared with the control group (P=0.020) as indicated in Figure 3. This result highlight that increased expression of Keap1 protein may be cause of laminar tissue damage of acute laminitis in dairy cows.
The immunohistochemical findings of Nrf-2 protein expression showed that the average Nrf-2 staining in the nucleus of laminar tissue in the OF group was 0.53 %, while it was 2.29 % in the control tissue. Statistically, Nrf-2 protein expression was highly significantly decreased in the nucleus of laminar tissue in the OF group in contrast to the control group (P=0.0032) as indicated in Figure 4. This may be due to severe oxidative stress in laminar tissue. However, the OF group suggested a gradual decline in nuclear Nrf2 material, suggesting a decrease in the activation of this transcriptional factor in laminar tissue further confirming that oxidative stress engaged in the laminar tissue damage of acute laminitis in dairy cows.
3.5. The Ultramicroscopic Structure Characteristics of Laminar Tissue of Laminitis Dairy Cows
In the control group, the epidermal basal cell's apex in the laminar tissue was blunt. The dense layer of the leaf was closely connected, deeply stained, and completely linear, running parallel to the basal cell membrane of the epidermis. Many hemidesmosomes were evenly distributed on the surface and in the basal cytoplasm of the epidermis, with anchoring filaments existing between the lamina densa and hemidesmosomes. The nuclei of the epidermal basal cells are oval in shape, away from the region of the basement membrane, with clear borders and high chromatin (Figure 5).
In the OF-treated cows, the tip of the basal cells of the epidermis in the laminar tissue was sharpened. The lamina densa appears thick and damaged, with interrelated collagen fibers, and lightly stained. The quantity of hemidesmosomes on the epidermis' basal cell membrane declined, and their distribution was uneven. The distance between the epidermal basal cell and the lamina densa has risen dramatically (Table 2). There are a few cytoskeleton anchoring filaments with damaged basal cell organelles. Similarly, in dairy cows with laminitis, the nuclei of the epidermal basal cells are deformed, unusually close to the region of the basement membrane, with ambiguous borders and reduced chromatin. This confirms the electron-microscopic characteristics of the damaged laminar tissue (Figure 5).
4. Discussion
Acute bovine laminitis, which contributes significantly to the gradual degradation of hoof function and structure, is a constant characteristic of oxidative stress. According to published scientific papers, we observed that dairy cow laminitis model was effectively developed through the OF-overload method, and OF group cows indicated clinical features of distinctive acute ruminal and systemic acidosis symptoms, as well as apparent histological indications in the lamellae, for example BM damage and separation, sinking of the epidermal lamellae, along with changes in basal cell shape [25,35]. We obtained another sample of laminar tissue from previous research to examine Ultrastructural changes and oxidative stress-related indicators at the gene and protein levels during OF-induced dairy cows laminitis.
Research suggests that oxidative stress is closely related to bovine laminitis [25,26,27]. To clarify the effect of oxidative stress and its persistent associated inflammation, ROS induces a compensatory response by inducing Nrf2 and subsequent expression of antioxidant and detoxifying enzymes [37,38]. Indeed, some Nrf2-activating phytochemical anti-inflammatory and antioxidant agents can boost cellular resistance to oxidative and electrophilic insults [39]. Oxidative damage and downregulation of the ROS producing enzyme Nqo1 in the laminar tissue were demonstrated by the OF-treated cows studied 72 h after OF overload. There has been comprehensive research into the interaction of Nrf2-Keap1 pathways with multiple refractory diseases, including lung cancer, pulmonary fibrosis, bronchial asthma, and chronic obstructive pulmonary disease with COPD [40], the effects of laminitis on the Nrf2-Keap1 system and its downstream gene products are unidentified and have been studied for the first time in the OF induced bovine laminitis. Despite the intense oxidative stress that Nrf2 activation and upregulation of its downstream gene products were supposed to induce, the OF-treated group showed a gradual decrease in nuclear Nrf2 material, indicating a decrease in the activation of this transcriptional factor in laminar tissue. This was completed by major downregulation of Nrf2 target gene products, containing Ho1 and Nqo1. This phenomenon indicates that the tendency of lame cows to climb compromises the biological response to the prevailing oxidative stress and its detrimental effects on the residual laminar tissue. A sharp increase in the abundance of Keap1 in the laminar tissue of the cows treated with OF compared with the control cows were accompanied by a paradoxical decrease in Nrf2 activation in the face of severe oxidative stress. A cysteine-rich protein Keap1 that functions as a redox indicator, an inhibitor of Nrf2 nuclear translocation and a facilitator of its proteasomal degradation [41]. The apparent absence of activation of Nrf2 can therefore be due, in part, to an increased abundance of Keap1, taking into account the predominant oxidative stress in the laminar tissue indicated in this research. This is consistent with prior research indicating that the severity of oxidative stress and inflammation in Nrf2 Knockout mice and exacerbated tissue injury were amplified by Nrf2 genetic disruption [42,43]. As seen for the first time here, acquired Nrf2 deficiency may also cause the development of oxidative stress and the progression of tissue damage to the laminar tissue in this model.
According to published studies, scientists typically concur that most cases of dairy cow laminitis arise from sub-acute rumen acidosis caused by the consumption of high-energy diets. Abaker et al. [44] reported that high-grain diet-fed SARA-induced cows show enhanced oxidative stress, as well as alterations in oxidative stress indicators in the liver and plasma and hepatic Nrf2 mRNA expression with increasing LPS transplanted into the bloodstream. When the oxidative stresses go beyond, an antioxidative property increasing hepatic Nrf2 mRNA expression [45]. In addition, Nrf2 genes in liver collected from SARA induced cows tended to decrease with the increased blood MDA [44]. Studies reported that increasing oxidative stress (OS) leads to the up- or down-regulation of hepatic genes related to and induced by OS [44,45]. In our work, Nrf2 also significantly decreases at both mRNA and protein levels as blood MDA increases [25], while Keap1 increased dramatically in the OF group versus the control group.
This study demonstrated the ultrastructure characteristics of laminar tissue in laminitis dairy cows, indicating that adhesion between laminar epidermal basal cells (EBCs) to the basement membranes (BMs) is contingent on the stability of many anchoring plaques called hemidesmosomes (HDs) [46]. On an ultrastructural level, HDs are distinguished by electron-dense cytoplasmic plaques that connect epithelial cells to the basement membrane zone (BMZ) while also acting as signal transducers by connecting intermediate filaments on the plasma membrane's cytoplasmic side with anchoring fibrils on its extracellular side [47]. Loss or damage to HDs has been documented in human bullous pemphigoid illness [48] and equine laminitis [49], resulting in improper cell-BM attachment. This segregation is the hallmark of structural alterations in dairy cows with laminitis. As a result, it is suspected that the shift in the number of HDs may entail the detachment of the basal epithelial cells (epidermal hoof) of laminitic dairy cows from the basement membrane and its deep part (the dermis), which finally leads to the development of laminitis.
In the current study, the quantity of HDs in the laminar tissue of OF group decreased, whereas the distance from the basal cell membrane to the central part of the hoof compact layer increased. Similar findings have been described in the horse’s laminitis [50]. Similarly, research discovered that the appearance of diseased horseshoe leaf tissue under the transmission electron microscope was similar to that of the scoring system established by for the evaluation of laminitis tissue damage, that is, for both EBCs and basal BM separation, the number of missing HDs on the basement membrane is positively correlated with the degree of laminitis tissue damage. It was found to be positively linked with line scores, confirming that ultrastructural HDs loss can cause animal laminitis [49].
Scientists currently believe that hoof separation occurs via two distinct mechanisms. In one case, glucose deprivation facilitated HDs to terminate, resulting in the breakdown of the epidermal basal cell cytoskeleton; in the second case, chemical activation of the epidermal basal cell matrix metalloproteinases (MMPs) destroyed anchoring filaments, rendering HDs in the basal cell plasmalemma intact [51,52]. Although the precise mechanisms underlying changes in HDs on the basement membrane and glucose deprivation remain unknown, the existing literature provides some useful inspiration. In the laminitis model induced by OF, for example, the animals will exhibit obvious symptoms before lameness, including marked hyperglycemia and hypercortisolemia 53. Scholars believe that the hooves are naturally distributed with a dense network of blood vessels that provide energy to the hooves. The laminar tissue, which is highly organized and situated at the end of circulation, has been noted to require higher levels of glucose to maintain structural integrity; alternatively, glucose deficiency results in rapid HDs disassembly [51,52]. It has previously been demonstrated that glucose is required for the integrity of laminar tissue. OF dosing producing considerable hyperglycemia, particularly at high doses, which seems not to be correlated with changes in insulin quantity. The hyperglycemia caused by OF overload may represent the unexpected failure of peripheral glucose absorption. Lamellae that are abruptly incapable to transport glucose may experience HDs disintegration, which would bring to laminar dermo-epidermal separation, the lesion that characterizes laminitis [54].
The present study discovered that OF caused shrinkage and loss of HDs in dairy cow laminar tissue, as well as anchoring fiber rupture. Similar ultrastructural damage can occur in laminar tissue when matrix metalloproteinases are activated. Thus, it appears that the psychophysiology of OF-induced laminitis is caused by one or two processes: the constituent MMPs activation and the glucose scarcity in secondary epidermal lamella (SEL) basal cells. Indirect data suggests that the molecular structure of BMs and anchoring fibrils are MMP substrates [55]. MMP levels were elevated in laminar homogenates from laminitic horses [52,56,57,58]. Recently, in our research group, Ding et al. also discovered that MMP-2 and MMP-9 gene expression enhanced in inflamed dairy cow laminar tissue [59]. These data show that the metalloproteinase family may have a role in the separation and destruction of the basement membrane in laminitic dairy cows.
Figure 6.
Schematic diagram illustrating the proposed oxidative stress pathway in laminar tissue during OF-induced acute laminitis in dairy cows. Following OF-overload, rumen fermentation changes, resulting in rumen acidosis and systemic inflammation. Consequently, the pathway begins with the activation of oxidative stress and accumulation of ROS, a key driver of acute laminitis. Elevated ROS levels increase the expression of Keap1, which sequesters and promotes the degradation of the master antioxidant regulator Nrf2. With Nrf2 activity suppressed, the expression of cytoprotective antioxidant genes is diminished, disabling the tissue's intrinsic defense mechanisms. This weakened cellular protection leads to laminar tissue damage, characterized by collagen fiber disruption (including reduced hemidesmosomes), separation of epidermal layers, thickening of the lamina densa, and eventual detachment of basal cells from the basement membrane. This cascade elucidates the molecular link between systemic oxidative insult and the structural failure characteristic of laminitis. Figure was created with BioRender.com.
Figure 6.
Schematic diagram illustrating the proposed oxidative stress pathway in laminar tissue during OF-induced acute laminitis in dairy cows. Following OF-overload, rumen fermentation changes, resulting in rumen acidosis and systemic inflammation. Consequently, the pathway begins with the activation of oxidative stress and accumulation of ROS, a key driver of acute laminitis. Elevated ROS levels increase the expression of Keap1, which sequesters and promotes the degradation of the master antioxidant regulator Nrf2. With Nrf2 activity suppressed, the expression of cytoprotective antioxidant genes is diminished, disabling the tissue's intrinsic defense mechanisms. This weakened cellular protection leads to laminar tissue damage, characterized by collagen fiber disruption (including reduced hemidesmosomes), separation of epidermal layers, thickening of the lamina densa, and eventual detachment of basal cells from the basement membrane. This cascade elucidates the molecular link between systemic oxidative insult and the structural failure characteristic of laminitis. Figure was created with BioRender.com.
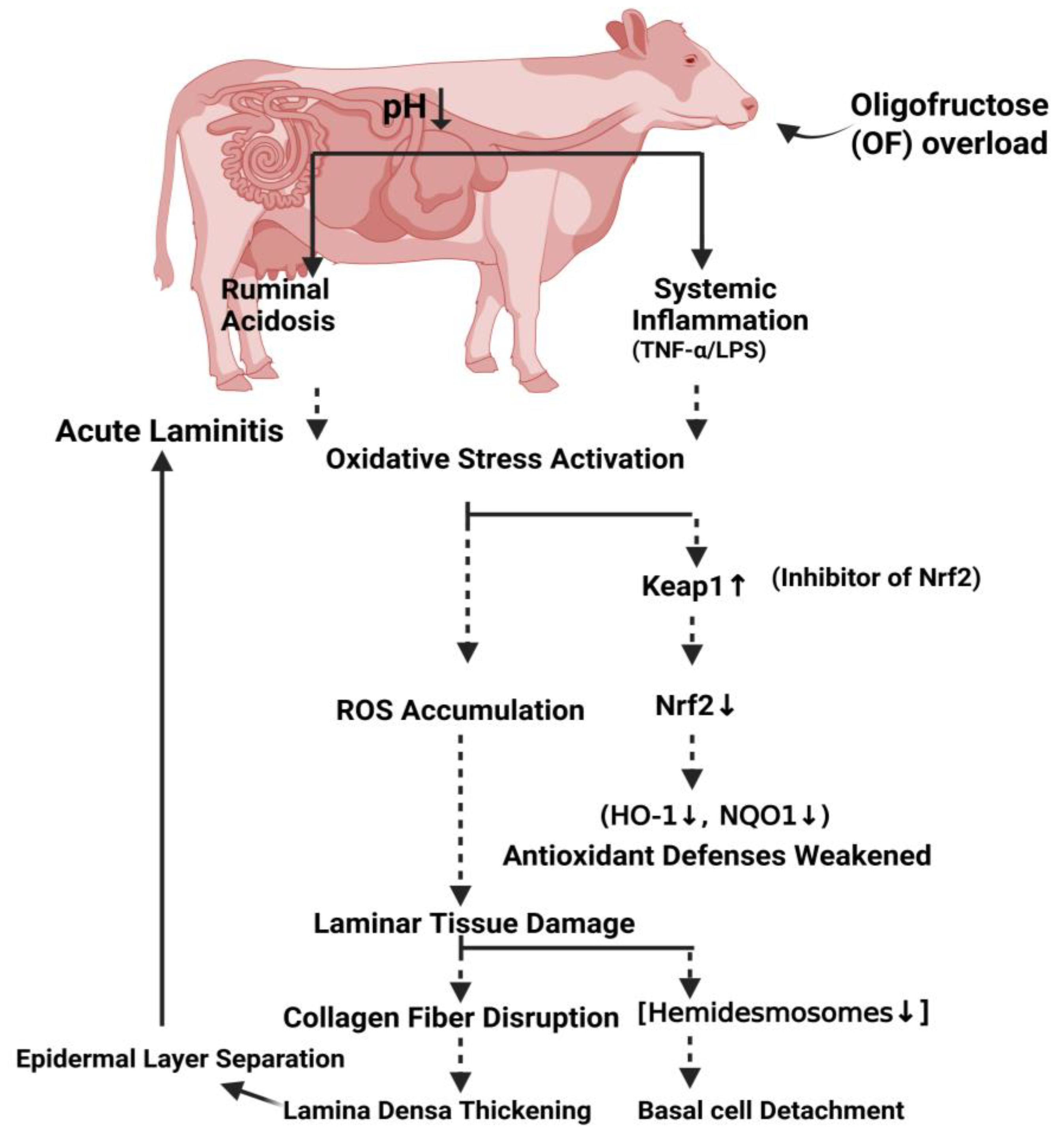
During the standing and walking process, cow hoof tissue is subjected to high loads, causing a significant amount of mechanical stress on the interface between the dermis and epidermis. Histological changes in a laminitis cow, such as the separation of characteristic epidermal basal cells from the basement membrane, combined with the ultrastructural changes, such as the loss of HDs on basement membrane damage, confirmed that HDs play a key role in epidermis-dermis adhesion. Hemidesmosomes and anchoring filament injuries weaken hoof tissue. The mechanical force of load-bearing and mobility acts on the third phalanx, leading it to spin and sink, irritating the sole dermis, and resulting in severe pain and lameness in cows [60]. In summary, dairy cows with OF-induced laminitis had reduced hemidesmosomes numbers, indicating that this was the main factor in the lamina densa's separation from the epidermal basal cell membrane. Additionally, the basal cytoskeleton suspense fibers were highly stained and clumped together, and the cell nucleus had moved to the basement membrane area without a distinct boundary.
While the present study provides novel perspectives into oxidative stress-driven laminar damage in OF-induced dairy cow laminitis, several limitations must be acknowledged. First, in this work, the experimental model utilized OF overload, which causes acute laminitis but doesn't effectively mimic the chronic metabolic stress, such as SARA, that normally occurs in field situations. Second, the present study's sample size (n = 12) was viable to detect significant alterations in the key oxidative stress indicators (Nrf2↓, Keap1↑). However, more cohorts may demonstrate detectable molecular alterations. Third, although the only 72-hour endpoint was validated by maximum laminitis intensity in the preliminary research, the study focused exclusively on the acute phase (72 h post-induction) of oligofructose-induced laminitis, and thus the long-term oxidative and ultrastructural dynamics remain unexplored. Additionally, the study did not assess potential confounding factors such as individual variation in antioxidant baseline status or hoof biomechanical load, which could influence laminitis progression. Future studies incorporating larger cohorts, multiple time points, and integrative omics approaches are warranted to validate and extend these findings. These limitations, however, present important prospects for improving laminitis therapy techniques that address oxidative stress dysregulation.
5. Conclusions
In conclusion, the disrupted Nrf2 activation in the laminar wall can cause a laminar tissue injury. Therefore, our results show that the unbalanced condition of oxidative stress at both gene and protein level could be a root cause of epidermal attachment failure that plays a central role in the pathogenesis of dairy cow laminitis which directly or indirectly causes damage to laminar tissue in dairy cows. In this study, the Ultrastructural damage of the laminar structural cells during the development of laminitis in dairy cows was also proved when the OF led to shrinkage and loss of HDs in dairy cow laminar tissue and rupture of anchoring fibers. Thus, the results obtained in this work could provide theoretical hints and a foundation of knowledge at the base level to enhance prevention and control measures in the context of dairy cows laminitis. However, more extensive gene and protein-based research is needed to fully comprehend the pathophysiology of dairy cows laminitis.
Author’s Contributions: M.A.H., H.B.W. conceived and conducted the experiments, analyzed the data, and wrote the manuscripts. J.F.D., L.T. and Z.X.H. performed some part of experiment. M.A.H. and J.F.D. evaluated the part of data. H.B.W. and Z.J.T. designed the experiments and participate in the conception and revised the manuscript. H.B.W. and Z.J.T. review and editing the manuscript. All authors read and approved the final this manuscript.
Institutional Review Board Statement
This study and all the procedures were approved in accordance with the rules and regulations of Animal Ethical Review Committee (Approval No: SRM-13) at the Department of Veterinary Surgery, College of Veterinary Medicine, Northeast Agricultural University Harbin, P.R. China.
Informed Consent Statement
All animals in this study were purchased from the Qingxi dairy farm in Xiangfang District Harbin, P.R. China. Owner of dairy farm were agreed for animal to participate in research study. He was agreed for animals to collect tissue samples and any further educational purposes. All the procedures were followed by Animal Ethical rules of College of Veterinary Medicine, Northeast Agricultural University Harbin, China.
Data Availability Statement
All data generated or analyzed during the study are included in this published article.
Acknowledgments
This work was supported by the National Key R&D Program of China (Project No.2017YFD0502200) and Heilongjiang Provincial Funding for National Subjects (GX18B023). The funders had no role in the study’s design, data collection and analysis, decision to publish, or preparation of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Tuniyazi, M.; Tang, R.; Hu, X.; Zhang, N. Methylated tirilazad may mitigate oligofructose-induced laminitis in horses. Front. Microbiol. 2024, 15, 1391892. [Google Scholar] [CrossRef]
- Passos, L.T.; Bettencourt, A.F.; Ritt, L.A.; Canozzi, M.E.A.; Fischer, V. Systematic review of the relationship between rumen acidosis and laminitis in cattle. Res. Vet. Sci. 2023, 161, 110–117. [Google Scholar] [CrossRef]
- Serİn, H.; Körez, M.K. Laminitis in cattle: A bibliometric analysis. Res. Pract. Vet. Anim. Sci. 2024, 2024, 104. [Google Scholar] [CrossRef]
- Bojkovski, J.; Nedić, S.; Arsić, S.; Vujanac, I.; Prodanović, R.; Mitrović, A.; Đurić, M.; Bugarski, D.; Panousis, N.K; Kalaitzakis, E.; Ninković, M. Pathogenesis of laminitis in dairy cows. Vet. Žur. Republik. Srpsk. 2023, 23, 307–317. [Google Scholar] [CrossRef]
- Boosman, R.; Nemeth, F.; Gruys, E. Bovine laminitis: clinical aspects, pathology and pathogenesis with reference to acute equine laminitis. Vet. Q. 1991, 13, 163–171. [Google Scholar] [CrossRef]
- Momcilovic, D.; Herbein, J.H.; Whittier, W.D.; Polan, C.E. Metabolic alterations associated with an attempt to induce laminitis in dairy calves. J. Dairy Sci. 2000, 83, 518–525. [Google Scholar] [CrossRef]
- Danscher, A.M.; Toelboell, T.H.; Wattle, O. Biomechanics and histology of bovine claw suspensory tissue in early acute laminitis. J. Dairy Sci. 2010, 93, 53–62. [Google Scholar] [CrossRef]
- Dong, S.W.; Zhang, S.D.; Wang, D.S.; Wang, H.; Shang, X.F.; Yan, P.; Yan, Z.T.; Yang, Z.Q. Comparative proteomics analysis provide novel insight into laminitis in Chinese Holstein cows. BMC Vet. Res. 2015, 11, 161. [Google Scholar] [CrossRef]
- Randall, L.V.; Green, M.J.; Huxley, J.N. Use of statistical modelling to investigate the pathogenesis of claw horn disruption lesions in dairy cattle. Vet. J. 2018, 238, 41–48. [Google Scholar] [CrossRef]
- Greenough, P.R. Bovine laminitis and lameness: a hands on approach. Els. Health Sci. 2007. [Google Scholar]
- Thoefner, M.B.; Pollitt, C.C.; Van-Eps, A.W.; Milinovich, G.J.; Trott, D.J.; Wattle, O.; Andersen, P.H. Acute bovine laminitis: A new induction model using alimentary oligofructose overload. J. Dairy Sci. 2004, 87, 2932–2940. [Google Scholar] [CrossRef] [PubMed]
- Danscher, A.M.; Enemark, J.M.D.; Telezhenko, E.; Capion, N.; Ekstrom, C.T.; Thoefner, M.B. Oligofructose overload induces lameness in cattle. J. Dairy Sci. 2009, 92, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Thoefner, M.B.; Wattle, O.; Pollitt, C.C.; French, K.R.; Nielsen, S.S. Histopathology of oligofructose-induced acute laminitis in heifers. J. Dairy Sci. 2005, 88, 2774–2782. [Google Scholar] [CrossRef] [PubMed]
- Mendes, H.M.; Casagrande, F.P.; Lima, I.R.; Souza, C.H.; Gontijo, L.D.; Alves, G.E.; Vasconcelos, A.C.; Faleiros, R.R. Histopathology of dairy cows' hooves with signs of naturally acquired laminitis. Pesquisa. Vet. Brasil 2013, 33, 613–619. [Google Scholar] [CrossRef]
- Leise, B.S.; Faleiros, R.R.; Watts, M.; Johnson, P.J.; Black, S.J.; Belknap, J.K. Laminar inflammatory gene expression in the carbohydrate overload model of equine laminitis. Equine. Vet. J. 2011, 43, 54–61. [Google Scholar] [CrossRef]
- Dern, K.; Van Eps, A.; Wittum, T.; Watts, M.; Pollitt, C.; Belknap, J. Effect of continuous digital hypothermia on lamellar inflammatory signaling when applied at a clinically-relevant timepoint in the oligofructose laminitis model. J. Vet. Int. Med. 2018, 32, 450–458. [Google Scholar] [CrossRef]
- Raber, M.; Lischer, C.J.; Geyer, H.; Ossent, P. The bovine digital cushion – a descriptive anatomical study. Vet. J. 2004, 167, 258–264. [Google Scholar] [CrossRef]
- Vermunt, J. “Subclinical” laminitis in dairy cattle. N. Z. Vet. J. 1992, 40, 133–138. [Google Scholar] [CrossRef]
- Alvergnas, M.; Strabel, T.; Rzewuska, K.; Sell-Kubiak, E. Claw disorders in dairy cattle: Effects on production, welfare and farm economics with possible prevention methods. Livest. Sci. 2019, 222, 54–64. [Google Scholar] [CrossRef]
- Potterton, S.L.; Bell, N.J.; Whay, H.R.; Berry, E.A.; Atkinson, O.C.; Dean, R.S.; Main, D.C.; Huxley, J.N. A descriptive review of the peer and non-peer reviewed literature on the treatment and prevention of foot lameness in cattle published between 2000 and 2011. Vet. J. 2012, 193, 612–616. [Google Scholar] [CrossRef]
- Caliri, A.W.; Tommasi, S.; Besaratinia, A. Relationships among smoking, oxidative stress, inflammation, macromolecular damage, and cancer. Mutation Res./Rev. Mutation Res. 2021, 787, 108365. [Google Scholar] [CrossRef] [PubMed]
- Kıran, T.R.; Otlu, O.; Karabulut, A.B. Oxidative stress and antioxidants in health and disease. J. Lab. Med. 2023, 47, 1–11. [Google Scholar] [CrossRef]
- Tomlinson, D.J.; Mülling, C.H.; Fakler, T.M. Invited review: formation of keratins in the bovine claw: roles of hormones, minerals, and vitamins in functional claw integrity. J. Dairy Sci. 2004, 87, 797–809. [Google Scholar] [CrossRef] [PubMed]
- Heinecke, L.F.; Grzanna, M.W.; Au, A.Y.; Mochal, C.A.; Rashmir-Raven, A.; Frondoza, C.G. Inhibition of cyclooxygenase-2 expression and prostaglandin E2 production in chondrocytes by avocado soybean unsaponifiables and epigallocatechin gallate. Osteoarthritis Cartilage 2010, 18, 220–227. [Google Scholar] [CrossRef]
- Hayat, M.A.; Ding, J.; Li, Y.U.; Zhang, X.; Zhang, J.I.; Li, S.; Wang, H.B. Determination of the activity of selected antioxidant enzymes during bovine laminitis, induced by oligofructose overload. Med. Weter. 2020, 76, 289–295. [Google Scholar] [CrossRef]
- Zhao, X.J.; Wang, X.Y.; Wang, J.H.; Wang, Z.Y.; Wang, L.; Wang, Z.H. Oxidative stress and imbalance of mineral metabolism contribute to lameness in dairy cows. Bio. Trace Element Res. 2015, 164, 43–49. [Google Scholar] [CrossRef]
- Li, S.; Ding, J.; Jiang, L.; Hayat, M.A.; Song, Q.; Li, Y.; Zhang, X.; Zhang, J. Dynamic ROS production and gene expression of heifers blood neutrophil in a oligofructose overload model. Front. Vet. Sci. 2020, 7, 211. [Google Scholar] [CrossRef]
- Hong, Y.; Boiti, A.; Vallone, D.; Foulkes, N.S. Reactive oxygen species signaling and oxidative stress: transcriptional regulation and evolution. Antioxidants. 2024, 13, 312. [Google Scholar] [CrossRef]
- Liu, S.; Pi, J.; Zhang, Q. Signal amplification in the KEAP1-NRF2-ARE antioxidant response pathway. Redox Bio. 2022, 54, 102389. [Google Scholar] [CrossRef]
- He, F.; Ru, X.; Wen, T. NRF2, a transcription factor for stress response and beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef] [PubMed]
- Annie-Mathew, A.S.; Prem-Santhosh, S.; Jayasuriya, R.; Ganesh, G.; Ramkumar, K.M.; Sarada, D.V.L. The pivotal role of Nrf2 activators in adipocyte biology. Pharmacol. Res. 2021, 173, 105853. [Google Scholar] [CrossRef]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An overview of Nrf2 signaling pathway and its role in inflammation. Molecules. 2020, 25, 5474. [Google Scholar] [CrossRef]
- Sprecher, D.E.A.; Hostetler, D.E.; Kaneene, J.B. A lameness scoring system that uses posture and gait to predict dairy cattle reproductive performance. Theriogenol. 1997, 47, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Edmonson, A.J.; Lean, I.J.; Weaver, L.D.; Farver, T.; Webster, G. A body condition scoring chart for Holstein dairy cows. J. Dairy Sci. 1989, 72, 68–78. [Google Scholar] [CrossRef]
- Ding, J.; Li, S.; Jiang, L.; Li, Y.; Zhang, X.; Song, Q.; Hayat, M.A.; Zhang, J.T.; Wang, H. Laminar inflammation responses in the oligofructose overload induced model of bovine laminitis. Front. Vet. Sci. 2020, 7, 351. [Google Scholar] [CrossRef]
- Hayat, M.A.; Ding, J.; Zhang, X.; Liu, T.; Zhang, J.; Bokhari, S.G.; Akbar, H.; Wang, H. Enhanced Autophagy in Damaged Laminar Tissue of Acute Laminitis Induced by Oligofructose Overloading in Dairy Cows. Animals. 2023, 13, 2478. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Jeon, J.H. Recent advances in understanding Nrf2 agonism and its potential clinical application to metabolic and inflammatory diseases. Int. J. Mol. Sci. 2022, 23, 2846. [Google Scholar] [CrossRef]
- Sharma, V.; Mehdi, M.M. Oxidative stress, inflammation and hormesis: the role of dietary and lifestyle modifications on aging. Neurochem. Int. 2023, 164, 105490. [Google Scholar] [CrossRef]
- Tkaczenko, H.; Kurhaluk, N. Antioxidant-rich functional foods and exercise: unlocking metabolic health through Nrf2 and related pathways. Int. J. Mol. Sci. 2025, 26, 1098. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, J.; Liu, R.; Wang, Q.; Qin, S.; Chen, Y.; Li, W. The role of Keap1-Nrf2 signaling pathway in the treatment of respiratory diseases and the research progress on targeted drugs. Heliyon 2024, 10, 18e37326. [Google Scholar] [CrossRef]
- Crisman, E.; Duarte, P.; Dauden, E.; Cuadrado, A.; Rodríguez-Franco, M.I.; López, M.G.; León, R. KEAP1-NRF2 protein–protein interaction inhibitors: Design, pharmacological properties and therapeutic potential. Med. Res. Rev. 2023, 43, 237–287. [Google Scholar] [CrossRef]
- Yoh, K.; Hirayama, A.; Ishizaki, K.; Yamada, A.; Takeuchi, M.; Yamagishi, S.I.; Morito, N.; Nakano, T.; Ojima, M.; Shimohata, H.; Itoh, K. Hyperglycemia induces oxidative and nitrosative stress and increases renal functional impairment in Nrf2-deficient mice. Genes Cells 2008, 13, 1159–1170. [Google Scholar] [CrossRef]
- Yoh, K.; Itoh, K.; Enomoto, A.; Hirayama, A.; Yamaguchi, N.; Kobayashi, M.; Morito, N.; Koyama, A.; Yamamoto, M.; Takahashi, S. Nrf2-deficient female mice develop lupus-like autoimmune nephritis1. Kidney Int. 2001, 60, 1343–1353. [Google Scholar] [CrossRef]
- Abaker, J.A.; Xu, T.L.; Jin, D.; Chang, G.J.; Zhang, K.; Shen, X.Z. Lipopolysaccharide derived from the digestive tract provokes oxidative stress in the liver of dairy cows fed a high-grain diet. J. Dairy Sci. 2017, 100, 666–678. [Google Scholar] [CrossRef]
- Gessner, D.K.; Schlegel, G.; Keller, J.; Schwarz, F.J.; Ringseis, R.; Eder, K. Expression of target genes of nuclear factor E2-related factor 2 in the liver of dairy cows in the transition period and at different stages of lactation. J. Dairy Sci. 2013, 96, 1038–1043. [Google Scholar] [CrossRef] [PubMed]
- Pollitt, C.C. Lamellar function at the cellular level. Equine Laminitis. 2017, 22–38. [Google Scholar]
- Borradori, L.; Sonnenberg, A. Hemidesmosomes: roles in adhesion, signaling and human diseases. Curr. Opi. Cell Bio 1996, 8, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.S.; Mascaó-Jr, J.M.; Liu, Z.; Espana, A.; Diaz, L.A. The desmosome and hemidesmosome in cutaneous autoimmunity. Clin. Exp. Immunol. 1997, 107. [Google Scholar]
- French, K.R.; Pollitt, C.C. Equine laminitis: loss of hemidesmosomes in hoof secondary epidermal lamellae correlates to dose in an oligofructose induction model: an ultrastructural study. Equine Vet. J. 2004, 36, 230–235. [Google Scholar] [CrossRef]
- Wang, L.; Pawlak, E.A.; Johnson, P.J.; Belknap, J.K.; Eades, S.; Stack, S.; Cousin, H.; Black, S.J. Impact of laminitis on the canonical Wnt signaling pathway in basal epithelial cells of the equine digital laminae. PLoS One 2013, 8, e56025. [Google Scholar] [CrossRef]
- French, K.R.; Pollitt, C.C. Equine laminitis: glucose deprivation and MMP activation induce dermo-epidermal separation in vitro. Equine Vet. J. 2004, 36, 261–266. [Google Scholar] [CrossRef]
- Kyaw-Tanner, M.; Pollitt, C.C. Equine laminitis: increased transcription of matrix metalloproteinase-2 (MMP-2) occurs during the developmental phase. Equine Vet. J. 2004, 36, 221–225. [Google Scholar] [CrossRef]
- Johnson, P.J. Endocrine and metabolic dysregulation in laminitis: Role of corticosteroids. Equine Laminitis. 2017, 141–148. [Google Scholar]
- Pass, M.A.; Pollitt, S.; Pollitt, C.C. Decreased glucose metabolism causes separation of hoof lamellae in vitro: a trigger for laminitis? Equine Vet. J. 1998, 30, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Giannelli, G.; Falk-Marzillier, J.; Schiraldi, O.; Stetler-Stevenson, W.G.; Quaranta, V. Induction of cell migration by matrix metalloprotease-2 cleavage of laminin-5. Sci. 1997, 277, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Pawlak, E.A.; Johnson, P.J.; Belknap, J.K.; Alfandari, D.; Black, S.J. Expression and activity of collagenases in the digital laminae of horses with carbohydrate overload-induced acute laminitis. J. Vet. Int. Med. 2014, 28, 215–222. [Google Scholar] [CrossRef]
- Loftus, J.P.; Johnson, P.J.; Belknap, J.K.; Pettigrew, A.; Black, S.J. Leukocyte-derived and endogenous matrix metalloproteinases in the lamellae of horses with naturally acquired and experimentally induced laminitis. Vet. Immunol. Immunopathol. 2009, 129, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Clutterbuck, A.L.; Harris, P.; Allaway, D.; Mobasheri, A. Matrix metalloproteinases in inflammatory pathologies of the horse. Vet. J. 2010, 183, 27–38. [Google Scholar] [CrossRef]
- Ding, J.; Shi, M.; Wang, L.; Qi, D.; Tao, Z.; Hayat, M.A.; Liu, T.; Zhang, J.T.; Wang, H. Gene expression of metalloproteinases and endogenous inhibitors in the lamellae of dairy heifers with oligofructose-induced laminitis. Front. Vet. Sci. 2020, 7, 597827. [Google Scholar] [CrossRef]
- Laat, M.A.D.; Pollitt, C.C. Ultrastructural examination of basement membrane pathology in horses with insulin-induced laminitis. Domestic Animal Endocrinol. 2019, 69, 30–34. [Google Scholar] [CrossRef]
Figure 1.
RT-qPCR results of mRNA concentration of oxidative stress-related genes including, Keap1, Nrf2, Ho-1, and Nqo1 in laminar tissue of the both groups. “* “show (P<0.05); “**” show (P<0.01).
Figure 1.
RT-qPCR results of mRNA concentration of oxidative stress-related genes including, Keap1, Nrf2, Ho-1, and Nqo1 in laminar tissue of the both groups. “* “show (P<0.05); “**” show (P<0.01).
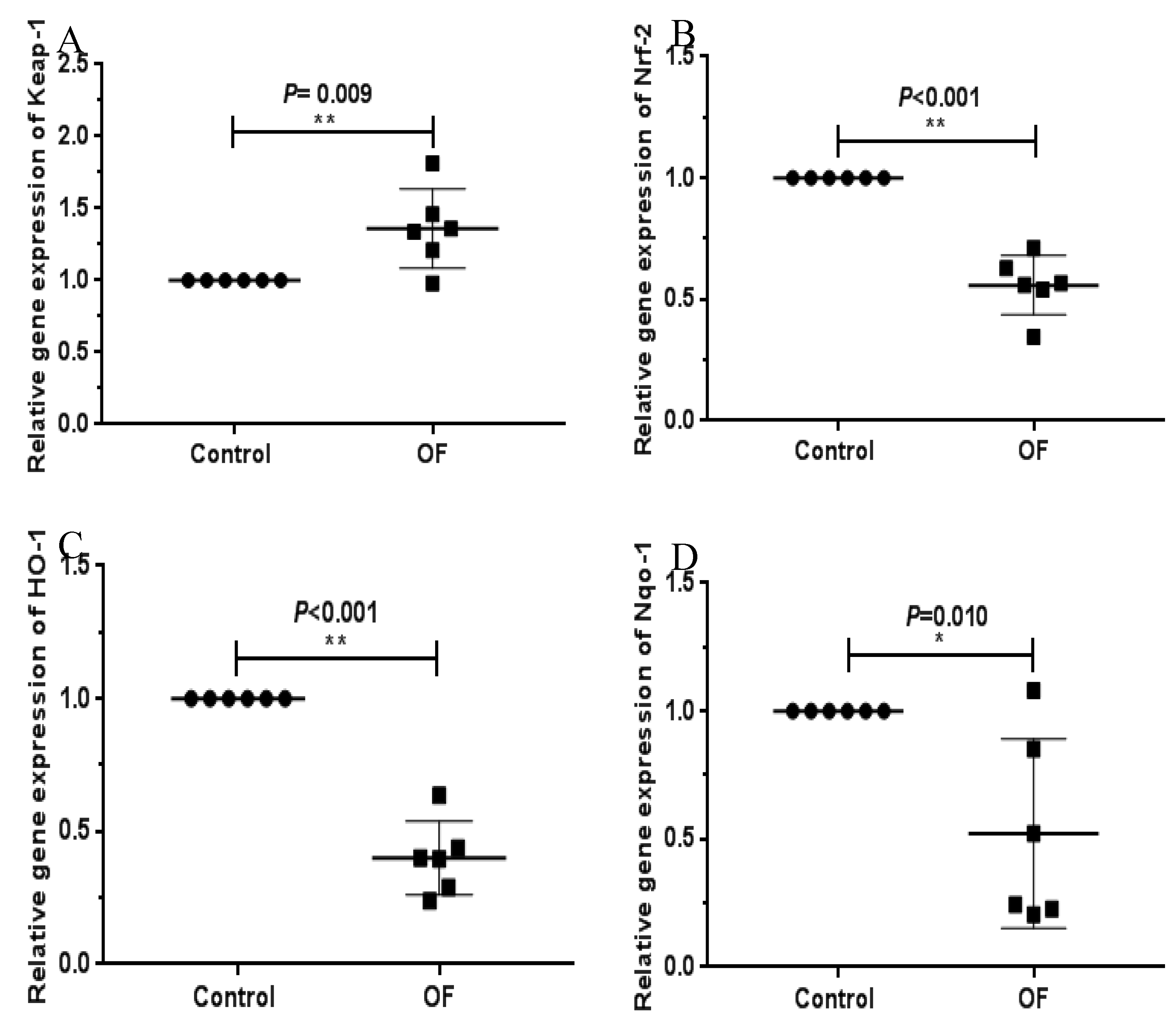
Figure 2.
Western blot results of oxidative stress-related proteins expression including Keap1, Nrf2, Ho1, and Nqo1 in the laminar tissue of the both groups. “* “show (P<0.05); “**” show (P<0.01).3.4. Immuno-expression of Keap1 and Nrf2 Proteins in Laminar Tissue of Laminitis Dairy Cows
Figure 2.
Western blot results of oxidative stress-related proteins expression including Keap1, Nrf2, Ho1, and Nqo1 in the laminar tissue of the both groups. “* “show (P<0.05); “**” show (P<0.01).3.4. Immuno-expression of Keap1 and Nrf2 Proteins in Laminar Tissue of Laminitis Dairy Cows
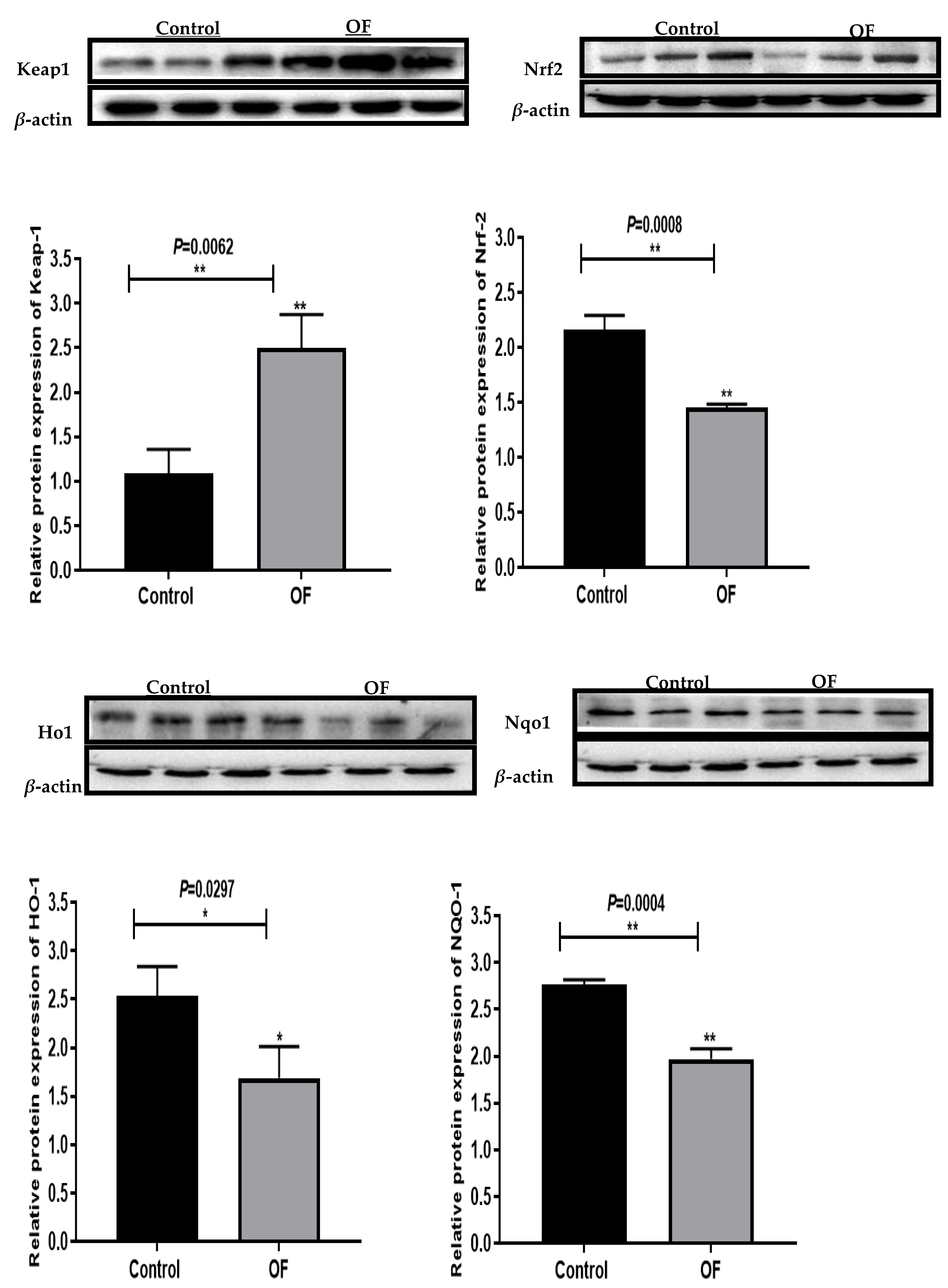
Figure 3.
Immunohistochemical staining of Keap1 in laminar tissues: Scale=200μm, 200X, (A, B, and C) control cows; (D, E, and F) OF-treated cows. EC, epidermal cels of epidermal lamellae; DC, dermal cells of dermal lamellae. “*” show (P<0.05).
Figure 3.
Immunohistochemical staining of Keap1 in laminar tissues: Scale=200μm, 200X, (A, B, and C) control cows; (D, E, and F) OF-treated cows. EC, epidermal cels of epidermal lamellae; DC, dermal cells of dermal lamellae. “*” show (P<0.05).
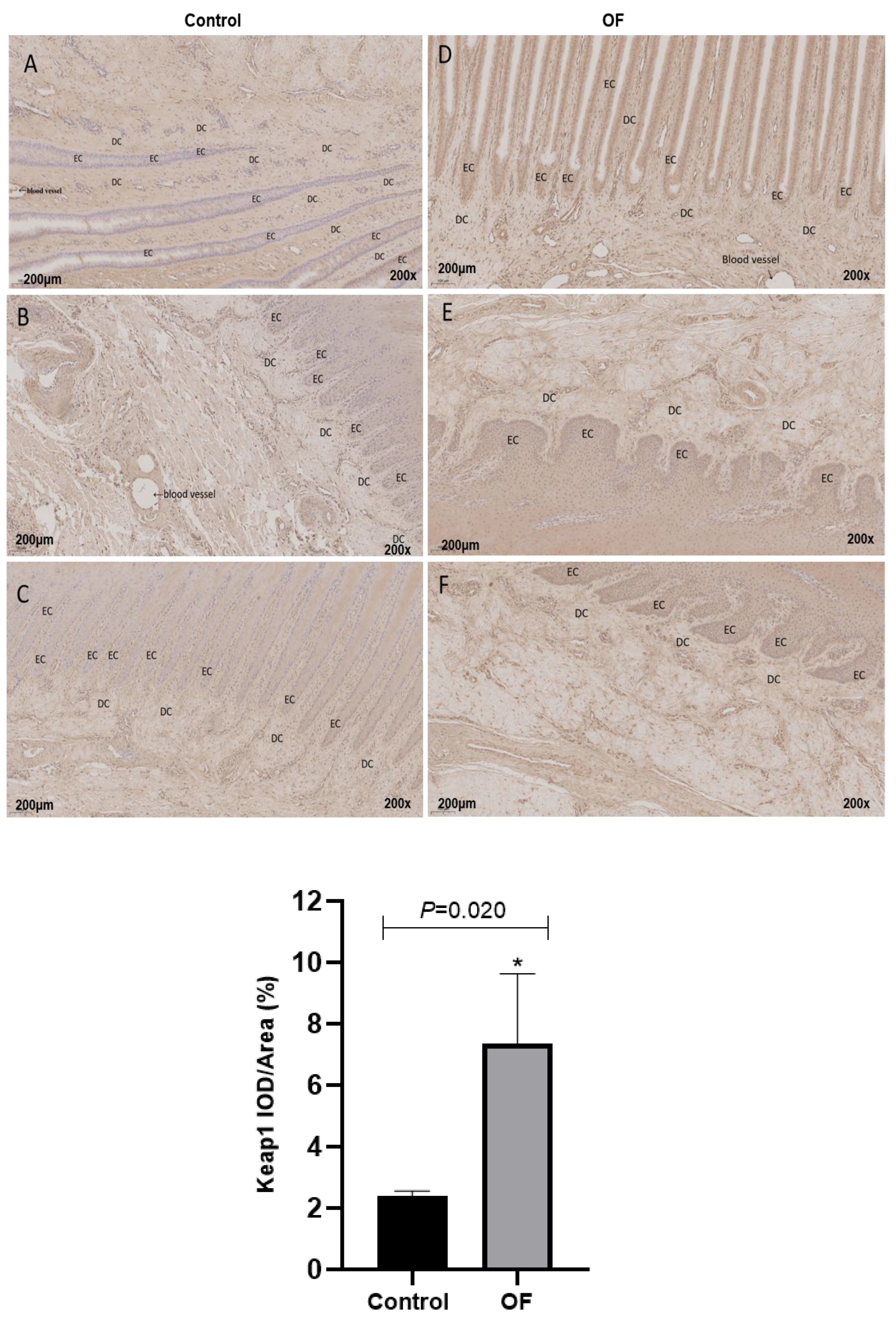
Figure 4.
Immunohistochemical staining of Nrf2 in laminar tissues: Scale=200μm, 200X, (A, B, and C) control cows; (D, E, and F) OF-treated cows. DC, dermal cells of dermal lamellae. “**” show (P<0.01).
Figure 4.
Immunohistochemical staining of Nrf2 in laminar tissues: Scale=200μm, 200X, (A, B, and C) control cows; (D, E, and F) OF-treated cows. DC, dermal cells of dermal lamellae. “**” show (P<0.01).
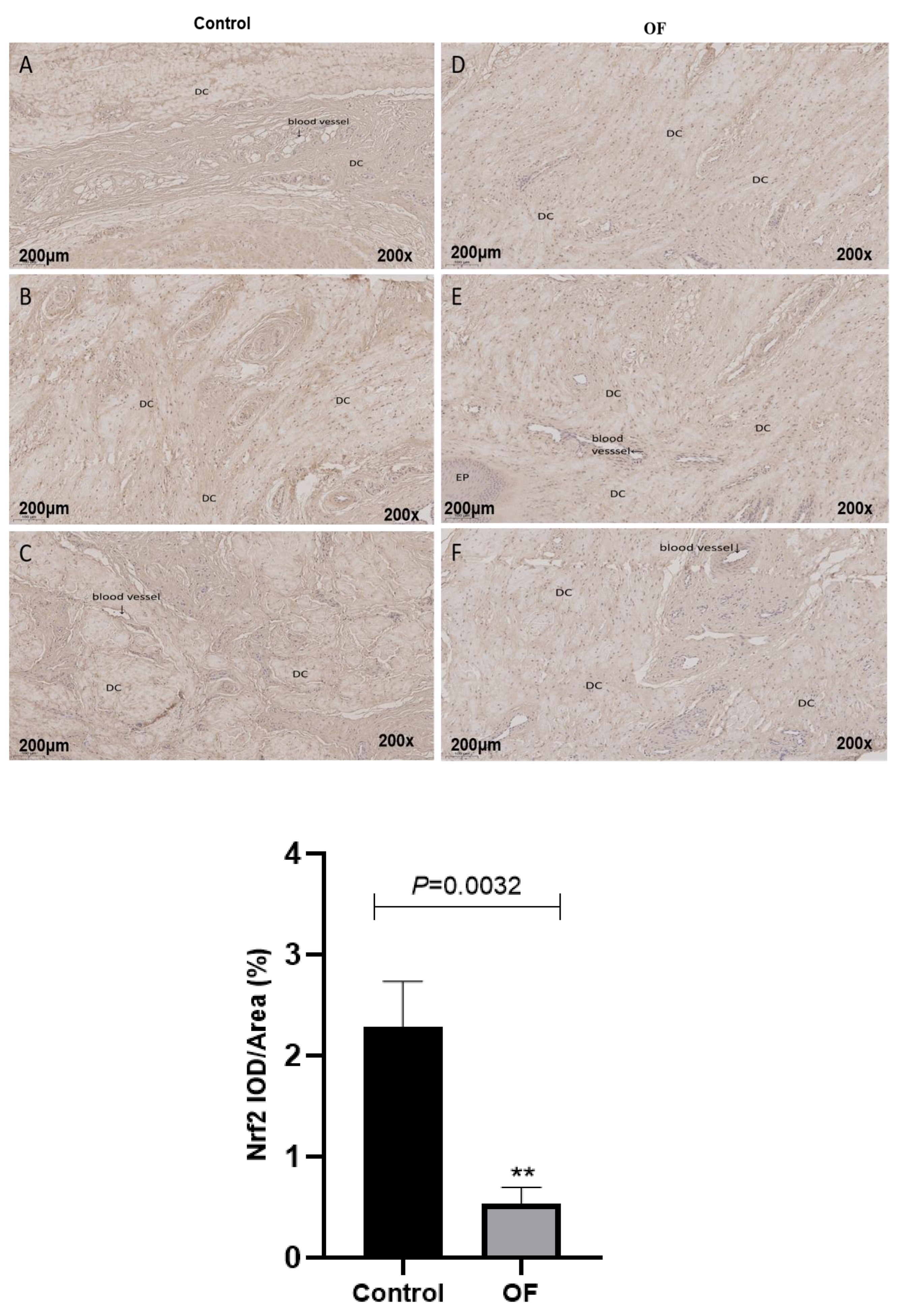
Figure 5.
Ultramicroscopic characteristics of laminar tissue of control and OF-treated cows. A,B: control group, C,D: OF-treated group. EBC: epidermal basal cell; N: nucleus; PD: primary dermal; SD: secondary dermal; Tip: the top of the epidermal basal cell; hemidesmosomes (*); the separation between lamina densa and epidermal basal cell (arrowhead).
Figure 5.
Ultramicroscopic characteristics of laminar tissue of control and OF-treated cows. A,B: control group, C,D: OF-treated group. EBC: epidermal basal cell; N: nucleus; PD: primary dermal; SD: secondary dermal; Tip: the top of the epidermal basal cell; hemidesmosomes (*); the separation between lamina densa and epidermal basal cell (arrowhead).
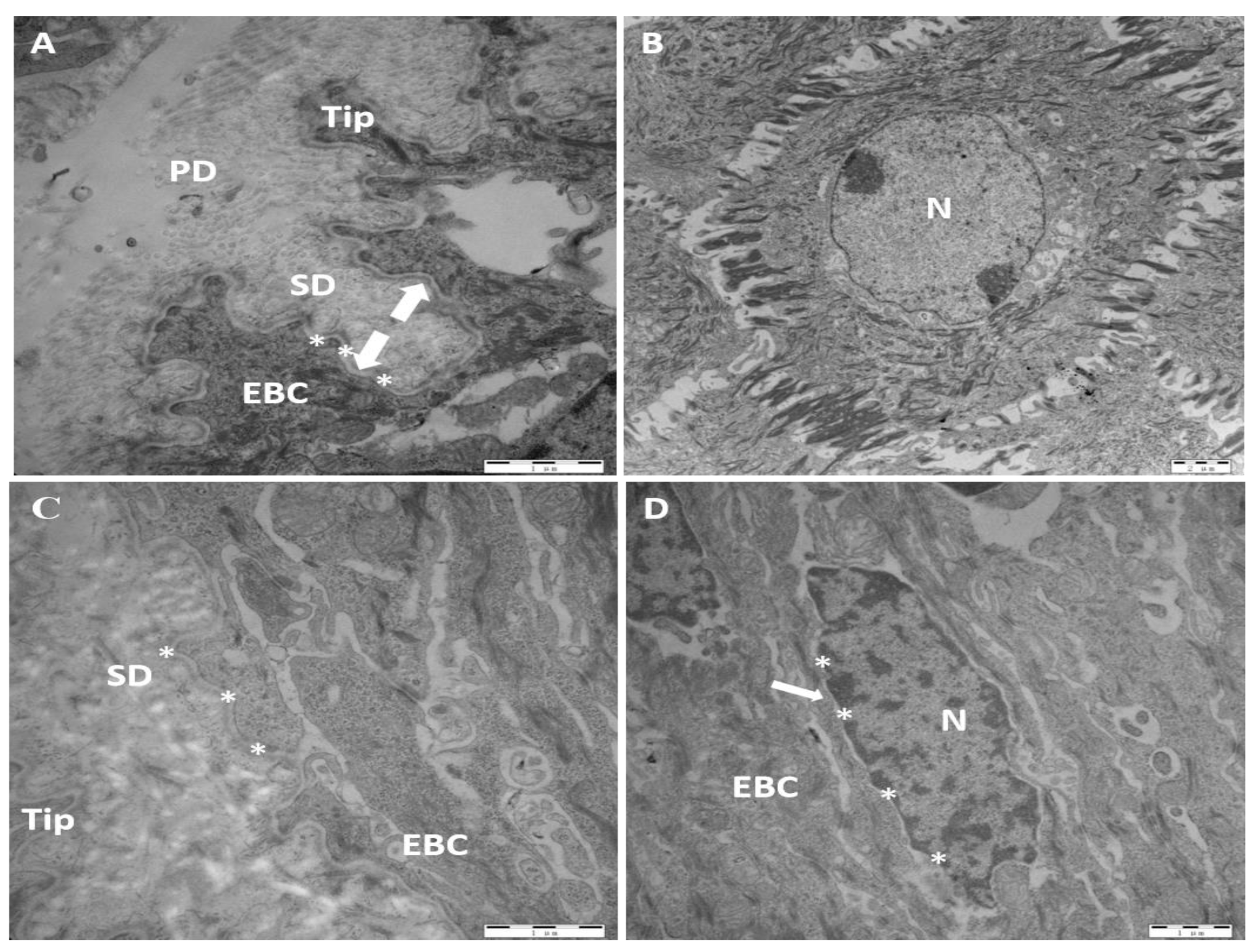

Table 1.
Primer Sequences.
| Genes | RefSeq accession No. | Primer sequences (5`-3`) |
|---|---|---|
| Keap1 | NM_001101142.1 | Forward: GGGCTACGACGGTCACACATTC Reverse: ATTCGGGTCACCTCGCTCCAG |
| Nrf2 | NM_001011678.2 | Forward: ACCACCCTGAAAGCACAACAGC Reverse: GAGTGGTCTGGTGATGCCATGC |
| Nqo1 | NM_001034535.1 | Forward: AGCGGCTCCATGTACTCTCTGC Reverse: TCCTCGGGAGTGTGCCCAATG |
| Ho1 | NM_001014912.1 | Forward: GCAGGCACCAGAGCTTCACAG Reverse: GAGGACCCATCGCAGGAGAGG |
| GAPDH | DQ402990 | Forward: GGGTCATAAGTCCCTCCACGA Reverse: GGTCATAAGTCCCTCCACGA |
Table 2.
Mean number/μm of hemidesmosomes on epidermal basal cells (EBC) basement membrane (BM) and mean distance from the EBC plasmalemma to the center of the lamina densa in laminitic dairy cows (x ± SD).
Table 2.
Mean number/μm of hemidesmosomes on epidermal basal cells (EBC) basement membrane (BM) and mean distance from the EBC plasmalemma to the center of the lamina densa in laminitic dairy cows (x ± SD).
| Group | Number of hemidesmosomes per μm on the basal cell membrane (n=100 μm, counts) | Distance from the basal cell membrane of the epidermis to the center of the laminar dense zone (n=100, μm) |
| Control | 3.831 ± 1.189 | 0.068 ± 0.022 |
| OF-treated | 1.532 ± 1.066** | 0.098 ± 0.029** |
**indicated P<0.01, an extremely significant difference.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



