
Submitted:
04 February 2026
Posted:
05 February 2026
You are already at the latest version
Abstract
Background: A significant strategy for sustainable management, by decreasing the use of chemical fertilizers, can consist of technology based on suitable levels of plant growth promoting bacteria (PGPB) seed biopriming. The main goal of this work was to demonstrate the positive effect of PGPB as seed biopriming factor on the growth of durum wheat plantlets under stress. Methods: plantlets physiological and biochemical pathways modifications were assessed in controlled growth chamber under salt stress. Results: Seeds bioprimed with bacterial strains, in particular, Bacillus pumilus (MA9), Virgibacillus halodenitrificans (MA14), Bacillus subtilis (MA17), and Bacillus pumilus (MA19), demonstrated the best development of durum wheat seedlings under salt stress. The total length and total dry weight were altogether higher in bioprimed than in unbioprimed plants. The PGPB-bioprimed plants were very sound and hydrated, and the unbioprimed plant leaves were parched within 120 mM NaCl. In contrast to explants without PGPB, all the biochemical variables, for example, auxin, proline, soluble sugars, amino acids, and protein, were essentially improved in bioprimed explants. Biopriming with MA17 fundamentally diminished the OR rate, particularly without salt stress. In any case, this impact of the hindrance of "ORs" was more pronounced under pressure. The explant antioxidant prevention agent pathways (CAT, APX) and the chlorophyll content were improved after seed biopriming with PGPB when explants with unbioprimed seeds had low cell reinforcement pathways under the two treatments, particularly under pressure. The bioprimed plants additionally had higher K+/Na+ proportions and expanded intracellular K+ ion activity. Conclusion: Hence, it can be anticipated that seed biopriming with PGPB is a novel technology that can improve healthy plant development under saline conditions. This study shows that PGPB assume a significant effect in initiating resistance under stressful conditions in plants and can be utilized to develop new bioinoculants to reduce the use of chemical fertilizers.
Keywords:
seed biopriming
; PGPB
; MDA
; proline
; CAT
; APX
; salt stress
; durum wheat
1. Introduction
Plants are exposed to various environmental stresses during growth and development under natural and agricultural conditions. How adverse environments affect plant growth is not only a fundamental scientific question but is also of vital importance for agriculture and food security. Several strategies to alleviate stress-induced adverse effects on plant growth have been described in many comprehensive reviews [1,2,3]. For example, high levels of ethylene that were produced under stressful environments can be reduced by the application of various ethylene inhibitors, such as amino ethoxy vinyl glycine (AVG), cobalt ions (Co2+), and silver ions (Ag+), leading to the enhancement of plant growth by alleviating the adverse effects of high levels of ethylene [4]. However, these agrochemicals are expensive and toxic to humans and soil health [5]. The intensive application of chemical fertilizers to overcome nutrient deficiency and achieve maximum agricultural productivity has led to unanticipated environmental impacts [6]. Currently, there are serious problems related to the management of the water and nutrient supply, so farmers use fertilizers in high quantities to obtain the maximum profit, although these high inputs of fertilizers lead to reductions of soil and groundwater quality [7]. Furthermore, despite increasing crop yields, the use of such chemicals results in a diminished net cash return for farmers. There are also environmental concerns about the persistence of these chemicals in the soil environment [8,9]. The other negative impacts of stresses, such as specific ion toxicity caused by salinity or root desiccation under drought stress, may not be overcome by using these chemicals. Although plants employ some specific mechanisms to combat these stresses, beneficial microbial populations in the rhizosphere also play a significant role in reducing the intensity of stress [10]. Various scientific research works have focused on developing technological approaches for modifying saline soil by rehabilitation measures or adopting biotic approaches using salt-tolerant plant crops, which is not an easy and cost-effective approach for sustainable agriculture [11].In this case, it is urgent to develop appropriate techniques for improved cereal production capacity in inefficiently cultivated or uncultivated areas, such as seed biopriming with diazotrophic PGPB [12]. Promising measures for improving plant health in salinated soils are the use of microbial vectors, which can ameliorate salt stress, promote plant growth [13], and control diseases [12,14,15]. The utilization of plant-associated bacteria that interact with plants by mitigating stress opens a new advanced technology for combating salinity. Many studies have demonstrated that beneficial microbes can enhance plant resistance to adverse environmental stresses, e.g., drought, salinity, nutrient deficiency, and heavy metal contamination. Such inoculants contribute to the development of sustainable agriculture under stressed conditions [16,17,18]. Inoculation of seeds of various crop plants (such as tomato, pepper, canola, bean, and lettuce) with PGPB, can ameliorate the plant tolerance to high levels of salt and leads to enhancement of plant growth by increasing root and shoot growth, dry weight, fruit and seed yields [13,19].
The general impacts of salinity stress on plant growth include hormonal and nutritional imbalance, physiological disorders such as epinasty, abscission, and senescence, and susceptibility to diseases [20,21,22]. Moreover, under salinity stress, ion toxicity occurs, particularly due to excessive amounts of Na+ and Cl−, which cause harmful effects on plant growth and development [23,24]. Certain abnormalities may occur in plants under stress and their intensity increases under such conditions. Plants adopt specific strategies to overcome the negative impact of stress. In most stressful environments, reactive oxygen species (ROS), such as superoxide, hydrogen peroxide, and hydroxyl radicals, which are detrimental to normal plant growth and development, are produced [25,26]. The presence of ROS can cause cellular damage through the oxidation of lipids and proteins, chlorophyll bleaching, and damage to nucleic acids, ultimately leading to cell death [27,28]. Plants develop self-defense mechanisms by producing antioxidant enzymes, such as superoxide dismutase, ascorbate peroxidase, glutathione reductase, and catalase [29,30]. In a saline environment, in addition to nutritional and hormonal imbalances, plant water uptake decreases due to changes in soil water potential [31]. Under such conditions, the accumulation of compatible solutes, namely, proline, glycine betaine, trehalose, polyols, and many others, takes place in the plant body[32,33]. This phenomenon plays an important role in protecting the plant from stress-induced deleterious effects by osmotic adjustment, limiting water loss, and diluting the concentration of toxic ions [34]. The accumulation of compatible solutes enables plants to maintain their osmotic potential for enhanced uptake of water.
In the present study, four Bacillus strains were isolated from the roots of the Halophyte Salicornia brachiata growing in coastal saline soil in Tunisia (34°46’16’’ N10°48’24’’ E). The isolates were checked for improvement of salt stress tolerance of durum wheat under natural and saline conditions. After one month of hydroponic culture, the plants were also tested for physiological and biochemical aspects such as auxin content and lipid peroxidation.
2. Results
2.1. Biochemical Characterization of PGPB Strains
All PGPB strains used for seed biopriming of durum wheat were identified based on16S RNA gene [12] and the Tree of the distance matrix and Distances were calculated using Pairwise Sequence Alignment using Blast and trees were constructed using the Neighbor Joining method (Figure 1). The screening results for PGP traits are duplicated in Tables S1 and S2. The growth capacity under different salt concentrations was studied after 48 h of incubation at 30 °C (Figure S1). All selected strains were able to grow under 125 mM NaCl, beyond which Bacillus pumilus strain MA9 and Virgibacillus halodenitrificans strain MA14 underwent a significant decrease, while the two strains Bacillus subtilis strain MA17 and Bacillus pumilus strain MA19 persisted with significant growth. IAA production (Figure S2), phosphate solubilization (Figure S3), siderophore production and antifungal activity were observed in all isolates of PGPB. All isolates were negative for HCN (Table S1) but positive for ammonia production, EPS secretion, and ACC deaminase production (Table S1). They were tested for the quantitative estimation of IAA in the presence and absence of tryptophan. The results showed that with addition of tryptophan, the production of IAA was higher than with the no addition of tryptophan for all strains in LB medium (Figure S2), but these four isolates were tryptophan dependent in minimal medium (data no shown). With the addition of tryptophan, the production of IAA increased, which, along with phosphate solubilization, was highest in Virgibacillus halodenitrificans strain MA14 and Bacillus pumilus strain MA9, whereas Bacillus subtilis strain MA17 and Bacillus pumilus strain MA19 had equal proportions of phosphate and IAA production (Figure S2; S3).

Table 1.
Seed biopriming of durum wheat plantlets in terms of the percentage increase in biomass by selected PGPB on growth parameters (total length and total dry weight) under nonsaline and saline conditions (in vivo). Two Control SN = untreated plantlets (0 mM); SN+ NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14= Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus pumilus MA19.
Table 1.
Seed biopriming of durum wheat plantlets in terms of the percentage increase in biomass by selected PGPB on growth parameters (total length and total dry weight) under nonsaline and saline conditions (in vivo). Two Control SN = untreated plantlets (0 mM); SN+ NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14= Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus pumilus MA19.
| Treatment | Total length (cm) | % Increase | Total dry mass (mg/plant) | % Increase |
|---|---|---|---|---|
| SN | 12,3333de | - | 53bc | - |
| BiopMA9 | 22b | 78 | 55,3333b | 42 |
| BiopMA14 | 13d | 05 | 46d | 18 |
| BiopMA17 | 24,3333a | 97 | 45,6667d | 17 |
| BiopMA19 | 24,3333a | 97 | 57b | 46 |
| SN+ salt | 7,6667 g | - | 24,3333f | - |
| BiopMA9 | 10f | 30.5 | 65a | 167 |
| BiopMA14 | 10,3333ef | 35 | 36e | 48 |
| BiopMA17 | 19c | 148 | 51,3333c | 111 |
| BiopMA19 | 14,3333d | 87 | 62a | 154.8 |
All the tested PGPB were found to produce phytase, β-glucanase, protease, and amylase activities (Figure 2). The antifungal activity of all strains was checked against Fusarium solani, F. oxysporum, F. graminarum and Rhizoctonia solani using PDA medium (Table S1). The antifungal activities of the tested strains varied according to PGPB and phytopathogenic fungal strains. In fact, Bacillus subtilis strain MA17 is the most effective against all fungal strains and it is the only efficient strain against Fusarium oxysporum. Moreover, no antifungal activity was noted with MA14. However, with Bacillus pumilus strain MA9 and Bacillus pumilus strain MA19, the antifungal activity was highly effective against Rhizoctonia solani but less effective against Fusarium solani (Figure 3).
2.2. Biofilm and Exopolysaccharides Production in Vitro
Biofilm production in vitro was studied using two control strains: E. coli DH5α (negative control) and Staphylococcus aureus sp. (positive control). After 24 h of incubation, the biofilm production capacity, compared with the two positive and negative controls, was more pronounced with Bacillus subtilis strain MA17 and Bacillus pumilus strain MA19 than with the other strains (Figure 4). However, the biofilm production capacities of Bacillus pumilus MA9 and Virgibacillus halodenitrificans MA14, were significantly higher compared to the negative control but lower than the biofilm production capacity of the positive control.
Exopolysaccharides production of selected PGPB strains (Figure 4) revealed that only the two strains, Bacillus pumilus strain MA9 and Bacillus subtilis strain MA17, produce EPS.
2.3. Effect of PGPB on the Morphology and Physiological Parameters of PGPB
The salt sensitivity assay showed that durum wheat var. “aouija” seedling growth was affected at 40 mM and further severely retarded at 125 mM NaCl. Therefore, a 125 mM concentration was selected for all experiments. The present study revealed that durum wheat var. “aouija” could tolerate treated plantlets in hydroponic culture more efficiently when bioprimed with halotolerant PGPB compared with the controls. All PGPB showed better plant growth in hydroponic culture without NaCl. When NaCl was added, the plant length, shoot length, and root length were severely affected in the case of unbioprimed plants, whereas growth was less reduced in bioprimed plants. In the presence of Virgibacillus halodenitrificans strain MA14, Bacillus subtilis strain MA17, and Bacillus pumilus strain MA19, growth was significantly better than that of other strains (Table 1, Figure S3 (A)). Although plants bioprimed with Bacillus pumilus strain MA9 did not show significantly greater growth, the latter was enhanced in comparison to control plants (Table 1). Plant biomass increased at 0 mM in the presence of all the PGPB. However, at 125 mM, plants bioprimed with Virgibacillus halodenitrificans strain MA14, Bacillus subtilis strain MA17, and Bacillus pumilus strain MA19 showed a significantly higher biomass than MA9 (Table 1, Figure S3 (B)).
Water leakage or variation in water retention reflected electrolyte leakage. Under stress and in the absence of any bacterial treatment, plants suffered from turgor and a reduction in ionic potential. Nevertheless, bacterial seed biopriming significantly improved water retention and thus reduced the leakage of electrolytes. The effect of strain on the reduction in intensity of the culture environment was more appreciable for treated plantlets than untreated plantlets, of which Bacillus subtilis strain MA17 presented the most dramatic percentage increase.
2.4. Effect of PGPB Biopriming on the Biochemical Parameters of Plants
Analysis of lipid peroxidation revealed that the level of malondialdehyde (MDA) in untreated bioprimed plantlets with the different strains was decreased considerably compared with untreated plantlets (Figure 5). Results showed also that Bacillus subtilis MA17 strain had the best ability to decrease the content of MDA compared with the cultures bioprimed with other strains. In the case of biopriming alone, the Bacillus subtilis MA17 strain decreased the MDA level by 76.12% compared with the control. Under stressful conditions, biopriming also had a significant effect on lowering the MDA rate in treated plantlets.
Analysis of the leaf proline content in untreated plantlets showed that in the presence of PGPB, its intracellular synthesis was significantly higher than the leaf proline content in the control. Results showed that the leaf proline content was strengthened by Bacillus pumilus MA9 and Bacillus subtilis MA17 strains especially (Figure 6). Thus, biopriming makes it possible to increase proline synthesis both without and with saline stress. Furthermore, its production was considerably improved in salt-treated plantlets compared with untreated plantlets. This effect was most noticeable in Bacillus pumilus strain MA9, in which the leaf content of proline was quadrupled (from 12.93 μg/g FW to 53.6 μg/g FW).
The production rate of amino acid (AA) was more remarkable with MA14 strain, in which the content of AA was tripled, compared to the untreated plantlets (Figure 7A)).
The protein content was significantly reduced in treated plantlets in the absence of any bacterial treatment, whereas in untreated culture, protein expression was normal. Protein expression was more pronounced in treated plantlets than in untreated plantlets, especially after biopriming with Bacillus pumilus MA9, Virgibacillus halodenitrificans MA14, and Bacillus subtilis MA17.
The analysis of total sugars in plant tissue showed that untreated bioprimed plantlet with potent strains had a significant effect on the total sugar content (Figure 8). However, this effect fluctuated according to the strain. Indeed, in the case of unbioprimed seed, the Bacillus pumilus MA19 strain provided the best result compared with the untreated plantlets. The strain with the best effect was, in this case, Bacillus subtilis strain MA17, which showed double the total sugar content. Under stressful conditions, the level of total sugars in bioprimed plantlets was considerably increased compared with the treated plantlets.
The auxin assay results using fresh material showed that biopriming had no significant effect compared with the untreated plantlets, except for the Bacillus pumilus MA9 strain, which showed an improved rate of AIA in plants (Figure 9). Similarly, under stress, only the effect of Bacillus pumilus MA9 strain was more significant than that of Virgibacillus halodenitrificans MA14 strain. However, seed biopriming had no marked effect on IAA cellular secretion with Bacillus subtilis MA17 and Bacillus pumilus MA19 strains compared with the treated plantlets.
The salt-treated plantlets (125 mM) strongly inhibited the formation of chlorophyll pigments. In untreated plantlets, these pigments were produced by the normal metabolism of the plant. Biopriming with PGPB improved the chlorophyll pigment content in untreated as well as under-treated plantlets. Among the other strains, Bacillus subtilis strain MA17 still had the most productive effect on pigmentation under all conditions (Figure 10).
In the present study, we observed a lower Na+ content and a higher K+ content in the treated and bioprimed plantlets. Therefore, higher K+/Na+ ratios in plants were observed due to restricted Na+ uptake and enhanced K+ uptake. Higher Na+ and lower K+ contents were observed in unbioprimed plants at 125 mM, whereas plants bioprimed with Bacillus subtilis MA17 and Bacillus pumilus MA19 strains showed the maximum reduction of Na+ concentrations. The K+ concentration and K+/Na+ ratio was higher at both 0 and 125 mM in the presence of all PGPB (Figure 11A,B). Under salt stress, the K+/Na+ ratio of the bioprimed plants with PGPB was almost double that of the unbioprimed plants.
2.5. Expression of the Antioxidant Enzymatic System
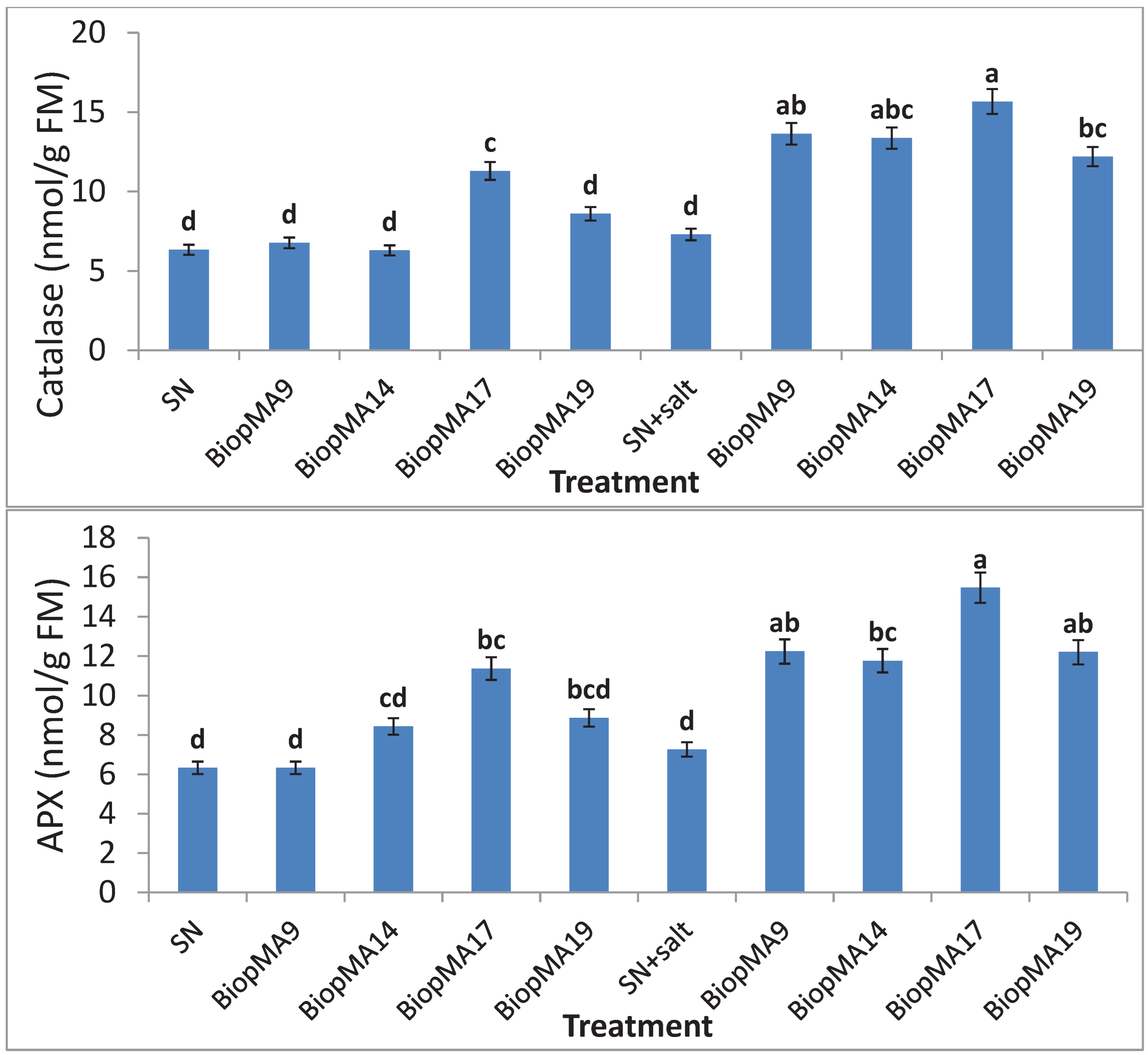
The plants that were not subjected to salinity-treated plantlets showed similar levels of catalase (Figure 12 A) and ascorbate peroxidase activity (Figure 12 B), irrespective of the microbial treatment. Higher catalase and ascorbate peroxidase activities were recorded in the seed-bioprimed plants in comparison to the treated and untreated plantlets. Biopriming with these selected PGPB significantly improved catalase activity in treated and untreated plantlets, and this significant effect was more pronounced in Bacillus subtilis MA17 strain. Similarly, this significant effect of biopriming was observed for ascorbate peroxidase activity. Increased enzyme activities were observed in the bioprimed plants when compared to the control; however, no significant difference was found between the bacterial treatments, with the exception of Bacillus subtilis MA17 strain.
3. Discussion
In this work, the positive effect of PGPB on the physiological and biochemical modifications of wheat plantlets under salt stress was studied. Many reports have underlined the role of Bacillus spp., Virgibacillus spp. and Exigubacterium spp. in promoting plant growth and ameliorating abiotic stress [35]. However, to authors’ knowledge, this is the first study to evaluate the impact of seed biopriming with four Bacillus sp. strains on the physiological and biochemical pathways of Tunisian durum wheat var. “aouija” under stressful culture conditions in a controlled chamber. All the PGPB tested in this study were found to possess ACC deaminase, β-glucanase, and protease activity and produced copious amounts of exopolysaccharides, siderophores, and antifungal compounds, suggesting their possible role in reducing salt-induced damage in plants. Plant responses to salinity stress caused by NaCl have been extensively studied [36]. Under salt stress, plants maintain ion and osmotic homeostasis via ionic signaling perception and adjustment and rapid osmotic stress. Successful colonization establishment is the initial step for plant-bacteria interactions.
According to the present study, the selected strains had a significant power to form a bacterial biofilm in vitro after 24 h of growth (Figure 4), which indicated that these strains formed an adequate microenvironment for their growth and their strong colonization of the plant rhizosphere. Biofilm formation is assumed to be one such strategy for the bacterium to ascertain stable establishment on inert and living surfaces. However, biofilms play potential roles in enabling persistent microorganism establishment [37]. The selected strains used for seed biopriming exhibited many biological activities characterizing all PGPB (Table S1 and Figure 2; 3). Plant growth-promoting bacteria are known to mitigate abiotic stress through various mechanisms, such as increased proline levels [10,38], mitigation of ethylene stress through ACC deaminase production [39,40,41], and exopolysaccharide-mediated reduction of Na+ uptake [37]. In addition, biofilm production after 48 h of incubation was the same in the four strains compared with the positive control. However, compared with the negative control, the ability to adhere to the surface remained stable for the four strains. For this reason, the Bacillus subtilis MA17 and Bacillus pumilus MA19 strains reached their biofilm production threshold faster than the other two strains.
In our study, Salt-treated plantlets reduced the osmotic potential of the growth medium and affected water availability, causing physiological drought in plants. PGPB helps the plant by removing physiological drought under salt stress and increasing the water content in the cell [41]. The higher hydration in bioprimed plants (Figure S3) leads to better water-use efficiency and thus improves photosynthesis and increases biomass [42]. Electrolyte leakage is used to measure the membrane permeability of the cell. Salt causes damage to the plant cell and hence increases its permeability. As a result, the electrolytes present in the membrane leak and accumulate in the surrounding tissues [43]. In the present study, PGPB-bioprimed plants with salt-treated plantlets showed significantly low electrolyte leakage from plant tissue, suggesting that PGPB protected the integrity of the plant cell membrane from the detrimental effect of salt (Figure S3). One reason for this finding may be a change in the saturation pattern of membrane phospholipids upon PGPB biopriming, which in turn causes a reduction in membrane potential [38]. This research work revealed a PGPB-mediated reduction in MDA content in bioprimed plants compared with unbioprimed plants. Thus, along with reduced electrolyte leakage, PGPB also prevented oxidative damage to plants caused by salt stress.
In the control plants, the percentage increase in water content was significantly lower, whereas bioprimed plants showed a higher percentage of water content (Figure S4 A). Analysis of the variances in water retention and electrolyte leakage capacity (Figure S4, A and B) showed that these parameters were proportionately related. Indeed, in the absence of any bacterial treatment, the ability of the plants to overcome the turgor phenomenon decreased significantly with treated plantlets versus untreated plantlets. Thus, in salt-treated plantlets, seed biopriming with these bacteria significantly improved the potential of plants to retain water and further reduced the leakage of any essential nutrients, such as amino acids, sugars, and electrolytes. Malondialdehyde (MDA) results from the decomposition of polyunsaturated fatty acids of cellular membranes and shows a greater accumulation in salt-treated plantlets [44]. Therefore, MDA is a reflection of the extent of the induced treated plantlets as well as the peroxidative damage caused by reactive oxygen species [45]. Plant cell membranes are generally considered primary sites of salt injury. Membrane destabilization is frequently attributed to lipid peroxidation, which can be initiated by activated oxygen species, such as O2-, OH-, or 1O2, or by the action of lipooxygenase [45]. In this study, the decrease in MDA content (Figure 5) in response to salinity in PGPB-treated plants is in accordance with earlier studies correlating the management of lipid peroxidation to improved plantlet tolerance mechanisms [46]. Similar results have been reported for salt tolerance in various plants induced by PGPB [47,48].
Under salinity, high levels of ethylene are produced [16,49] and inhibit root growth, hence affecting some plant metabolic pathways [50]. Auxin and mainly AIA are two growth phytohormones keys. Indeed, auxin stimulates root proliferation, which increases nutrient uptake, thus promoting plant growth [51]. The effectiveness of auxin depends on its concentration. For example, at low concentrations, it improves plant growth [52], while at a high level, it inhibits root growth [53]. In this study, plant AIA secretion depended on strain seed biopriming (Figure 6), in which Bacillus pumilus strain MA9 was the only strain that promoted auxin secretion in plants with freely treated plantlets. However, all the strains subjected to seed biopriming favored production, except Bacillus pumilus strain MA9, which is the most relevant.
Conversely, nonenzymatic antioxidants, such as cysteine, nonprotein thiol, proline, carotenoids, and ascorbic acid, may play a role in inducing resistance to salinity by protecting acid-labile macromolecules against attack by free radicals that are formed during various metabolic reactions leading to oxidative stress [54]. Notably, the enhanced proline content was associated with the plants’ inherent salt tolerance mechanisms (Figure 6). Yoshiba, Kiyosue [55] and Mohamed and Gomaa [56] reported that PGPB inoculation improved the proline content, which might be due to the upregulation of the proline biosynthesis pathway to maintain high proline levels. This phenomenon helps to maintain the cell water status, assisting plants in coping with salinity stress. Many species of gram-positive bacteria have also been shown to increase the proline pool size and accumulate total free amino acids under osmotic stress [47]. Proline and total soluble sugars are very important biochemical indicators of salinity tolerance in plants [57,58]. When plants face salt stress, proline accumulates in the cytosol and plays an important role in cytoplasmic osmotic adjustment [59,60]. Proline also helps the plant cell by stabilizing subcellular structures such as membranes and proteins, scavenging free radicals, and buffering the cellular redox potential under salt stress [61]. Proline, an amino acid, acts as a scavenger for ROS, and its accumulation at a well-defined concentration is a sign of stress [62].
Different amino acids and nitrogen-containing compounds accumulate in higher plants exposed to salt treatment [63,64,65]. However, strain MA14, which showed the most potent effect under untreated plantlets, revealed a significant lowering of the total AA content under salt stress. Under stressful conditions, the AA content was also improved in the presence of PGPB compared with the control without biopriming. Moreover, under stress, the plants expressed many more proteins than the untreated plantlets (Figure 5 B). The biological pathway of amino acid secretion was closely related to the total protein content in bioprimed plants with all the PGPB assessed (Figure 7 A and B). In these two biological pathways, seed biopriming with Virgibacillus halodenitrificans MA14 strain is more remarkable and pronounced under stress. This phenomenon can be explained by the marked transformation of the total AA into protein and can also be justified by the high proteolytic activity in this strain, which makes available the amino acids involved in protein chains and therefore their assimilation by the plant and their transformation into proteins. The total free amino acids are reported to be high in the foliar parts of different salt-tolerant plants [66,67]. Few of these proteins may have a direct correlation with salinity tolerance. Similarly, many proteins that are directly or indirectly related to plant growth promotion have been found to be expressed in rice plants by the interaction of Pseudomonas fluorescens [68].
Salinity causes an imbalance in the ratio of ion homeostasis in the plant system. Plants try to maintain a low salt composition in the cytosol by extrusion through the plasma membrane using the SOS pathway or by scavenging in the vacuole through NHX1 antiprotons. Salinity impedes the ratio of Ca2+ and K+ in the cell. However, an increase in the K+ concentration can alleviate the deleterious effect of salinity on growth and yield [69]. Similarly, several PGPB have been reported to reduce salt toxicity in various other plants by lowering the Na+ concentration and increasing the K+ concentration [70,71,72,73,74]. It is important to note that the effect of biopriming is more pronounced under salt stress. Hence, a clear difference was observed for the untreated plantlets of each strain compared with those exposed to stressful conditions (Figure 11 (A-B)). For example, the two strains Bacillus subtilis strain MA17 and Bacillus pumilus strain MA19 were twice as effective in treated than in untreated plantlets.
The large battery of ROS-scavenging enzymes included in the ‘‘ROS gene network’’ included catalases, superoxide dismutase (SOD), and ascorbate peroxidases. Increased enzyme activities were observed in the bioprimed plants when compared with the control; however, no significant difference was found between the bacterial treatments, with the exception of MA17 (Figure 12 A and B). The higher levels of catalase and ascorbate peroxidase enzymes in the bacteria-bioprimed plants could be associated with the enhanced tolerance toward salinity stress [75,76,77,78].
These hydroponic results indicated that seed biopriming with these PGPB strains improved the tolerance induction, directly or indirectly, of physiological and biochemical pathways and therefore the tolerance of durum wheat "aouija" grown under salt stress conditions.
4. Materials and Methods
4.1. Microorganisms
The present study explores four salt-tolerant Bacillus spp. strains, namely, Bacillus pumilus strain MA9, Virgibacillus halodenitrificans strain MA14, Bacillus subtilis strain MA17, and Bacillus pumilus strain MA19, which were previously isolated from plants roots growing in coastal saline soil (Salicornia brachiata) in Tunisia, and screened for salt tolerance (ranged from 0 to 7 g/l) [12]. .
The 16S rRNA gene sequences of these strains have been submitted to GenBank under accession numbers MT672745, MT672746, MT672747, and MT672748 for Bacillus pumilus strain MA9, Virgibacillus halodenitrificans strain MA14, Bacillus subtilis strain MA17, and Bacillus pumilus strain MA19, respectively [12].
The cultures were maintained on trypticase soy agar under refrigerated conditions. The isolated strains exhibited PGP traits in vitro, such as IAA, phosphate solubilization, siderophore, HCN, antifungal, lytic enzymes, and ammonia production.
4.2. Biofilm and Exopolysaccharides Production
Strains from fresh agar plates were inoculated in 5 mL of tryptic soy broth and incubated for 24 h at 30 °C. Biofilm formation was conducted in 96-well flat-bottom plates (with lids) [79]. E. coli DH5α was used as a negative control, and Staphylococcus aureus M2 was used as a positive control. To visualize biofilm formation, the remaining bacteria attached to the bottom of wells were stained for 15 min at room temperature with 150 mL of 0.2% crystal violet solution prepared in 20% ethanol (v/v) [80]. Subsequently, crystal violet was eliminated, and the excess stain was rinsed three times with sterile water. The microplates were then inverted to remove any dye that was not staining the attached bacteria. Finally, 200 mL of glacial acetic acid was added to each well. The plates were incubated for 1 h at room temperature to promote biofilm bursting and solubilization of crystal violet that had already penetrated the cells. All tests were performed in triplicate. The OD of each well was measured at 570 nm using a microplate’s reader.
The qualitative determination of exopolysaccharide production was performed according to [81]. Each strain was inoculated onto 5 mm diameter discs arranged in modified Burk medium: 2% yeast extract, 1.5% K2HPO4, 0.02% MgSO4, 0.0015% MnSO4, 0.0015% FeS04, 0.003% CaCl2, 1.0% NaCl by addition of 10% sucrose, pH 7.5. Production was characterized by the size of the halo produced and its viscous appearance. The production of EPS was confirmed by mixing a portion of the mucoid substance in 2 mL of absolute ethanol, where the formation of a precipitate indicated the presence of EPS [81].
4.3. Seed Biopriming with PGPB and Salt-Stress Treatment
Durum wheat seeds were collected from three agricultural areas in Tunisia. Seeds were surface sterilized with 0.1% HgCl2 and subsequently rinsed four times with sterilized double-distilled water. PGPB used for seed biopriming was grown in DF salt media at 30 °C at 180 rpm for 24 h. A total of 500 µl of primary culture was bioprimed in 50 ml of LB medium and grown at 30 °C at 180 rpm to an OD 600 of 0.6. Cells were collected at 5,000 rpm for 10 min and resuspended in MS medium to 108 CFU/ml-1. Seeds of uniform size were placed on sterilized cotton in an autoclaved vial after biopriming by diazotrophic PGPB selected for this study for germination. Seven-day-old germinated seedlings were placed on a perforated Eppendorf floated in a glass pot containing 50 ml MS major and minor components in four sets (0 mM NaCl), each with 10 plants. Durum wheat plants were bioprimed with 5 ml of the PGPB culture in two sets. After 4 days of acclimatization with PGPB biopriming, 125 mM NaCl was added to two sets; the other was not exposed to salt. Both bioprimed and control treatments were replicated three times. The plants were grown in a culture room at 25 ± 2 °C under a 16-h/8-h light/dark cycle (350 mmol m-2 s-1 light intensity). The medium in each pot was changed after 7 days. Two pots without PGPB inoculums served as unbioprimed controls for salt-treated and untreated plants.
4.4. Plant Analysis
After a 21-day treatment, the growth parameters of the plants were observed. The fresh weight (FW) and total length of each plant were measured. Each plant was divided into roots and shoots, excluding the endosperm region, and the FW, length, and dry weight (DW) were recorded after drying in an oven at 70 °C for 72 h. The percentage increase in biomass was calculated using the following formula: % increase in biomass= (plant FW - seedling weight)/plant FW * 100
4.4.1. Electrolyte Leakage and Percentage Change in Water Content
Electrolyte leakage was measured according to [82]. One young leaf from the first primary branch toward the distal end of the same size was collected from three plants for each treatment and washed thoroughly with deionized water to remove surface-adhered electrolytes.
The samples were placed in closed vials containing 10 ml of deionized water and incubated at 25 °C on a rotary shaker for 24 h, and the electrical conductivity of the solution (Lt) was determined using Seven Easy conductivity (Metler Toledo AG 8603, Switzerland). Samples were then autoclaved at 120 °C for 20 min, and the final electrical conductivity (L0) was obtained after cooling at 25 °C. The electrolyte leakage was defined as follows:
Electrolyte leakage (%) = (Lt/L0) * 100
The percentage change in water content was calculated as follow [83]:
[(FW – DW)/FW] * 100. Each experiment was replicated three times, and the mean values are provided in Figure 4.
4.4.2. Lipid Peroxidation
Lipid peroxidation was estimated by determining the malondialdehyde (MDA) concentration produced by the thiobarbituric acid (TBA) reaction according to [84]. Leaf material (0.5 g) was homogenized in 15 ml of 80% alcohol. In one set, 1 ml of extract was mixed with 1 ml of 0.5% (w/v) TBA in 20% (w/v) TCA. In another set, TBA was excluded. The mixture was incubated at 90 °C for 30 min and then cooled at room temperature. Samples were centrifuged at 4,000 rpm for 3 min, and the absorbance of the supernatant was read at 400, 532, and 600 nm. The concentration of MDA was calculated with the following formula:
A = [(Abs532+TBA - Abs600+TBA) - (Abs532-TBA - Abs600-TBA)]
B= [(Abs440+TBA _ Abs600-TBA) 0.0571]
MDA equivalents nmol g-1 FW= [(A _ B)] 15.700.106
4.4.3. Auxin Analysis
Auxin analysis was carried out according to [85] using the alcoholic extract of the shoot and root tissue. One milliliter of the alcoholic extract was mixed with 2 ml of Salkowski reagent and incubated at room temperature in the dark for 20 min followed by spectrophotometric analysis at 535 nm.
4.4.4. Biochemical Analysis
The free proline content in the leaves was determined using ninhydrin [86,87] with minor modifications. Plant tissue (100 mg) was homogenized in 1.2 ml of 3% aqueous sulfosalicylic acid and centrifuged at 13,000 rpm for 10 min. After centrifugation, 500 µl of supernatant was increased up to 1 ml with distilled water and reacted with 1 ml of 2% ninhydrin. The mixture was incubated at 90 °C for 1 h. The samples were cooled in an ice bath, and 2 ml of toluene was added and vortexed for 2 min. The upper phase was aliquoted to read the absorbance at 520 nm in a UV-1600 PC spectrophotometer. The proline content was calculated by comparing the absorbance with a standard curve drawn from known concentrations of L-proline (Sigma Aldrich, USA) and expressed as µg/mg FW.
The total amino acid content was determined as described by [58,88] with minor modifications. One milliliter of the plant extract was treated with 1 ml of 0.2 M citrate buffer (pH 5), 1 ml of 80% ethanol, and 1 ml of ninhydrin (1%), followed by incubation at 95 °C for 15 min. The samples were cooled, and the absorbance was read at 570 nm [89] using a Jenway 7305 spectrophotometer.
The total protein content was determined using 0.5 g homogenized plant tissue in 2 ml of extraction buffer [50 mM Tris–HCl (pH 8.3), 1 mM EDTA, 3 mM DTT, 0.08% ascorbic acid, 1 mM PMSF]. The protein was quantified using the Bradford method [90]. The total soluble sugars were analyzed by treating 0.1 ml of the alcoholic extract with 3 ml of freshly prepared anthrone reagent [150 mg anthrone in 100 ml of 72% (v/v) H2SO4] and placed in a boiling water bath for 10 min according to [88,91]. After cooling, the absorbance at 620 nm was determined using a UV–1600 PC spectrophotometer. The calibration curve was generated using glucose (Sigma Aldrich, USA) in the range from 20–400 µg/ml.
The chlorophyll content was determined according to the method of [92] modified by [93]. This method consists of cutting 0.5 g of fresh material from each sample into small segments, homogenizing them in 5 ml of 80% acetone, and leaving them in the freezer overnight. After filtration, the optical density was measured at 663 and 645 nm. The concentrations of chlorophyll a and b were deduced according to the following formula:
Chla (mg / l) = 12.7 OD663 - 2.69 OD645;
Chlb (mg / l) = 22.9 OD645 - 4.68 OD660; Chl total = Chla + Chlb
Where Chla is the concentration of chlorophyll a, Chlb is the concentration of chlorophyll b, and Chl total is the total chlorophyll concentration.
To analyze the ion content, 0.2 g of plant tissue was digested with 4 ml of perchloric acid and nitric acid solution (3:1). The solution was dried on a hot plate and further increased to 25 ml with deionized water and filtered through a 0.2-µm filter. The ion content was measured using an inductively coupled plasma optical emission spectrometer [94].
4.4.5. Studies on Antioxidative Enzymes of Plants
Fresh shoot and root samples (0.5 g) were homogenized in 5 ml of buffer (0.05 M Tris HCl buffer, pH = 7.5, 3 mM MgCl2, 1 mM EDTA) using a mortar and pestle, followed by centrifugation. The enzyme extracts thus obtained were used to determine APX and CAT activities. For determination of APX, 2 mM ascorbate was added to the extraction buffer. For measurement of APX, 0.1 ml of the enzyme extracts was added to the reaction mixture containing 50 mM phosphate buffer, 0.2 ml H2O2 1% (w/v), 0.5 mM ascorbate, and 0.1 mM EDTA. In addition, a decrease in the absorbance of ascorbate (extinction coefficient 2.8 mM-1 cm-1) was observed at 240 nm [95]. The activity of CAT was measured by adding 50 µl of enzyme extract to the reaction mixture containing phosphate buffer and H2O2 and following the decrease in absorbance of H2O2 (extinction coefficient 0.036 mM-1 cm-1) within 1 min at 240 nm [96].
4.5. Statistical Analysis
Each experiment was repeated three times, and the data for ten plants were recorded. The data were analyzed for significance using analysis of variance followed by Duncan’s least significant difference test (p = 0.05) using IBM SPSS Statistics software v. 21.0 (SPSS Inc. Chicago, IL, USA). The critical difference (C.D.) values were calculated at the p = 0.05 level to determine significant differences between the means of different bacterial treatments under both control (0 mM) and 125 mM NaCl stress conditions. Significantly different mean values are indicated by different letters.
5. Conclusion
Seed biopriming with the Bacillus subtilis strain MA17 could protect wheat from salt stress more than the other tested strains. The alleviation of salt stress in wheat could be considered the integration of multiple physiological processes, including the improvement of plant mineral nutrition, increased plant antioxidative capacity and regulation of plant growth by plant hormones (IAAs), siderophores and lytic enzymes or activation of ion transporters leading to an increased ratio of K+/ Na+ in wheat plants. These isolates might be useful in formulating new inoculants with combinations of different mechanisms of action, leading to a more efficient application for salt-treated plantlet adaptation and thereby improving cropping systems. Therefore, it is plausible that PGPB are an effective approach for improving the growth of salt-sensitive plants, a strategy that could be applied for sustainable agriculture.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
The study was conducted with the support of government funding from the Tunisian Ministry of Higher Education and Scientific Research contract program LMBEE_CBS/code: LR15CBS06_2022-2024.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article and in the supplementary materials.
Competing Interests
The authors declare that they have no competing interests.
Authors' Contributions
Experimental design and writing were performed by AHB. AHB, SZ, FE, JM and Man BA conducted the scientific and analytical results. AHB carried out the statistical analysis. Mam BA and FN completed the editing and correction of the manuscript. All authors approved the final version.
Acknowledgments
The authors are very grateful to the two teams from the LBMIE and LMB at the Biotechnology Center of Sfax, Tunisia, for their collaboration. We also dully acknowledge the Pr. Feto Aliye Naser University of Tennessee Health Science Center and Pr. Saîbi Walid Plant Biotechnology Amelioration Lab. Center of Biotechnology of Sfax, for their valuable comments, corrections and proofreading of the manuscript.:
References
- Kaushal, M. and S.P. Wani, Rhizobacterial-plant interactions: strategies ensuring plant growth promotion under drought and salinity stress. Agriculture, Ecosystems & Environment, 2016. 231: p. 68-78. [CrossRef]
- Etesami, H. and B.R. Glick, Halotolerant plant growth–promoting bacteria: Prospects for alleviating salinity stress in plants. Environmental and Experimental Botany, 2020: p. 104124. [CrossRef]
- Gupta, A., et al., Mechanistic insights of plant growth promoting bacteria mediated drought and salt stress tolerance in plants for sustainable agriculture. International Journal of Molecular Sciences, 2022. 23(7): p. 3741.
- Gamalero, E. and B.R. Glick, Bacterial modulation of plant ethylene levels. Plant Physiology, 2015. 169(1): p. 13-22.
- Bernardes, M.F.F., et al., Impact of pesticides on environmental and human health. Toxicology studies-cells, drugs and environment, 2015: p. 195-233.
- Adesemoye, A.O. and J.W. Kloepper, Plant–microbes interactions in enhanced fertilizer-use efficiency. Applied microbiology and biotechnology, 2009. 85(1): p. 1-12.
- Zhu, Z., et al., Microsatellite markers linked to 2 powdery mildew resistance genes introgressed from Triticum carthlicum accession PS5 into common wheat. Genome, 2005. 48(4): p. 585-590. [CrossRef]
- Ahmadi, N., H. Mibus, and M. Serek, Characterization of ethylene-induced organ abscission in F1 breeding lines of miniature roses (Rosa hybrida L.). Postharvest Biology and Technology, 2009. 52(3): p. 260-266.
- Edwards, C., Environmental pollution by pesticides. Vol. 3. 2013: Springer Science & Business Media.
- Etesami, H. and D.K. Maheshwari, Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicology and environmental safety, 2018. 156: p. 225-246. [CrossRef]
- Majeed, A. and S. Siyyar, Salinity Stress Management in Field Crops: An Overview of the Agronomic Approaches, in Plant Ecophysiology and Adaptation under Climate Change: Mechanisms and Perspectives II. 2020, Springer. p. 1-16.
- Hadj Brahim, A., et al., Biopriming of Durum Wheat Seeds with Endophytic Diazotrophic Bacteria Enhances Tolerance to Fusarium Head Blight and Salinity. Microorganisms, 2022. 10(5): p. 970. [CrossRef]
- Lugtenberg, B.J., et al., Plant growth promotion by microbes. Molecular microbial ecology of the rhizosphere, 2013. 2: p. 561-573.
- Lugtenberg, B., et al., Microbial control of plant diseases. Molecular microbial ecology of the rhizosphere, Wiley-Blackwell Publishing, Oxford, 2013: p. 67-91.
- Pliego, C., F. Kamilova, and B. Lugtenberg, Plant growth-promoting bacteria: fundamentals and exploitation, in Bacteria in agrobiology: Crop ecosystems. 2011, Springer. p. 295-343.
- Glick, B.R., et al., Promotion of plant growth by ACC deaminase-producing soil bacteria, in New perspectives and approaches in plant growth-promoting Rhizobacteria research. 2007, Springer. p. 329-339.
- Dodd, I.C. and F. Pérez-Alfocea, Microbial amelioration of crop salinity stress. Journal of Experimental Botany, 2012. 63(9): p. 3415-3428. [CrossRef]
- Berg, G., et al., Biocontrol and osmoprotection for plants under salinated conditions. Molecular microbial ecology of the rhizosphere, 2013. 1: p. 561-573.
- Li, G., et al., Maize endophytic plant growth-promoting bacteria Peribacillus simplex can alleviate plant saline and alkaline stress. International Journal of Molecular Sciences, 2024. 25(20): p. 10870.
- Vimal, S.R., et al., PGPR: An Effective Bio-Agent in Stress Agricultural Management. 2016.
- Dar, Z.M., et al., Review on plant growth promoting rhizobacteria and its effect on plant growth. Journal of Pharmacognosy and Phytochemistry, 2018. 7(3): p. 2802-2804.
- Altuntaş, Ö. and İ.K. Kutsal, Use of Some Bacteria and Mycorrhizae as Biofertilizers in Vegetable Growing and Beneficial Effects in Salinity and Drought Stress Conditions. Physical Methods for Stimulation of Plant and Mushroom Development, 2018: p. 65.
- WANG, L., et al., Effects of NaCl on plant growth, root ultrastructure, water content, and ion accumulation in a halophytic seashore beach plum (Prunus maritima). Pak. J. Bot, 2018. 50(3): p. 863-869.
- Hniličková, H., et al., Effect of salt stress on growth, electrolyte leakage, Na+ and K+ content in selected plant species. Plant, Soil and Environment, 2019. 65(2): p. 90-96.
- Hajiboland, R. and A. Joudmand, The K/Na replacement and function of antioxidant defence system in sugar beet (Beta vulgaris L.) cultivars. Acta Agriculturae Scandinavica Section B–Soil and Plant Science, 2009. 59(3): p. 246-259. [CrossRef]
- Kärkönen, A. and K. Kuchitsu, Reactive oxygen species in cell wall metabolism and development in plants. Phytochemistry, 2015. 112: p. 22-32.
- Tsukagoshi, H., Control of root growth and development by reactive oxygen species. Current Opinion in Plant Biology, 2016. 29: p. 57-63.
- Das, K. and A. Roychoudhury, Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Frontiers in environmental science, 2014. 2: p. 53.
- Latef, A.A.H.A. and H. Chaoxing, Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Scientia Horticulturae, 2011. 127(3): p. 228-233.
- Abd_Allah, E.F., et al., Endophytic bacterium Bacillus subtilis (BERA 71) improves salt tolerance in chickpea plants by regulating the plant defense mechanisms. Journal of Plant Interactions, 2018. 13(1): p. 37-44.
- Acosta-Motos, J.R., et al., Changes in growth, physiological parameters and the hormonal status of Myrtus communis L. plants irrigated with water with different chemical compositions. Journal of plant physiology, 2016. 191: p. 12-21.
- Rajasheker, G., et al., Role and regulation of osmolytes and ABA interaction in salt and drought stress tolerance, in Plant Signaling Molecules. 2019, Elsevier. p. 417-436.
- Wijayasinghe, Y.S., A. Tyagi, and N.K. Poddar, Regulation of cell volume by Osmolytes, in Cellular Osmolytes. 2017, Springer. p. 195-228.
- Ashraf, M., M. Shahbaz, and Q. Ali, Drought-induced modulation in growth and mineral nutrients in canola (Brassica napus L.). Pak J Bot, 2013. 45(1): p. 93-98.
- Allioui, N., et al., Two Novel Bacillus Strains (subtilis and simplex Species) with Promising Potential for the Biocontrol of Zymoseptoria tritici, the Causal Agent of Septoria Tritici Blotch of Wheat. BioMed Research International, 2021. 2021. [CrossRef]
- Zhou, Y., et al., The Effect of Salt-Tolerant Antagonistic Bacteria CZ-6 on the Rhizosphere Microbial Community of Winter Jujube (Ziziphus jujuba Mill.“Dongzao”) in Saline-Alkali Land. BioMed research international, 2021. 2021.
- Sun, Y., et al., The Inhibitory Effects of Ficin on Streptococcus mutans Biofilm Formation. BioMed Research International, 2021. 2021.
- Dimkpa, C., T. Weinand, and F. Asch, Plant–rhizobacteria interactions alleviate abiotic stress conditions. Plant, Cell & Environment, 2009. 32(12): p. 1682-1694. [CrossRef]
- Barnawal, D., et al., 1-Aminocyclopropane-1-carboxylic acid (ACC) deaminase-containing rhizobacteria protect Ocimum sanctum plants during waterlogging stress via reduced ethylene generation. Plant Physiology and Biochemistry, 2012. 58: p. 227-235.
- Singh, S.R., et al., Plant growth-promoting bacteria: an emerging tool for sustainable crop production under salt stress, in Bioremediation of Salt Affected Soils: An Indian Perspective. 2017, Springer. p. 101-131.
- Li, X., et al., A novel PGPR strain Kocuria rhizophila Y1 enhances salt stress tolerance in maize by regulating phytohormone levels, nutrient acquisition, redox potential, ion homeostasis, photosynthetic capacity and stress-responsive genes expression. Environmental and Experimental Botany, 2020: p. 104023.
- Ali, H., et al., Seed priming improves irrigation water use efficiency, yield, and yield components of late-sown wheat under limited water conditions. Turkish Journal of Agriculture and forestry, 2013. 37(5): p. 534-544. [CrossRef]
- Wu, S.S., Enhanced phytoremediation of salt-impacted soils using plant growth-promoting rhizobacteria (PGPR). 2009, University of Waterloo.
- Naidoo, C., Oxidative status and stress associated with cryopreservation of germplasm of recalcitrant-seeded species. 2012.
- Karuppanapandian, T., et al., Reactive oxygen species in plants: their generation, signal transduction, and scavenging mechanisms. Australian Journal of Crop Science, 2011. 5(6): p. 709.
- Khan, M. and S. Panda, Alterations in root lipid peroxidation and antioxidative responses in two rice cultivars under NaCl-salinity stress. Acta Physiologiae Plantarum, 2008. 30(1): p. 81.
- Li, H. and X. Jiang, Inoculation with plant growth-promoting bacteria (PGPB) improves salt tolerance of maize seedling. Russian Journal of Plant Physiology, 2017. 64(2): p. 235-241.
- Khan, M.A., et al., Halotolerant rhizobacterial strains mitigate the adverse effects of NaCl stress in soybean seedlings. BioMed research international, 2019. 2019. [CrossRef]
- Tao, J.-J., et al., The role of ethylene in plants under salinity stress. Frontiers in Plant Science, 2015. 6: p. 1059.
- Shrivastava, P. and R. Kumar, Soil salinity: a serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi journal of biological sciences, 2015. 22(2): p. 123-131.
- Ji, S.H., M.A. Gururani, and S.-C. Chun, Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiological research, 2014. 169(1): p. 83-98.
- Patten, C.L. and B.R. Glick, Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol., 2002. 68(8): p. 3795-3801.
- Xie, H., J. Pasternak, and B.R. Glick, Isolation and characterization of mutants of the plant growth-promoting rhizobacterium Pseudomonas putida GR12-2 that overproduce indoleacetic acid. Current Microbiology, 1996. 32(2): p. 67-71. [CrossRef]
- Choudhary, D.K. and A. Varma, Microbial-mediated induced systemic resistance in plants. 2016: Springer.
- Yoshiba, Y., et al., Regulation of levels of proline as an osmolyte in plants under water stress. Plant and Cell Physiology, 1997. 38(10): p. 1095-1102.
- Mohamed, H. and E. Gomaa, Effect of plant growth promoting Bacillus subtilis and Pseudomonas fluorescens on growth and pigment composition of radish plants (Raphanus sativus) under NaCl stress. Photosynthetica, 2012. 50(2): p. 263-272. [CrossRef]
- Ashraf, M., et al., Inoculating wheat seedlings with exopolysaccharide-producing bacteria restricts sodium uptake and stimulates plant growth under salt stress. Biology and Fertility of soils, 2004. 40(3): p. 157-162.
- Sandhya, V., et al., Effect of plant growth promoting Pseudomonas spp. on compatible solutes, antioxidant status and plant growth of maize under drought stress. Plant Growth Regulation, 2010. 62(1): p. 21-30. [CrossRef]
- Leigh, R.A., N. Ahmad, and R.G.W. Jones, Assessment of glycinebetaine and proline compartmentation by analysis of isolated beet vacuoles. Planta, 1981. 153(1): p. 34-41.
- Upadhyay, S.K., et al., Impact of PGPR inoculation on growth and antioxidant status of wheat under saline conditions. Plant Biology, 2012. 14(4): p. 605-611.
- Ashraf, M. and M. Foolad, Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environmental and experimental botany, 2007. 59(2): p. 206-216.
- Mohammadi, M., et al., Study the effect of 24-epibrassinolide application on the Cu/Zn-SOD expression and tolerance to drought stress in common bean. Physiology and Molecular Biology of Plants, 2020: p. 1-16. [CrossRef]
- Yuan, L., et al., Effects of 24-epibrassinolide on nitrogen metabolism in cucumber seedlings under Ca (NO3) 2 stress. Plant Physiology and Biochemistry, 2012. 61: p. 29-35.
- Mansour, M.M.F., Nitrogen Containing Compounds and Adaptation of Plants to Salinity Stress. Biologia Plantarum, 2000. 43(4): p. 491-500.
- AM, B.N., Oxidative stress markers and antioxidant potential of wheat treated with phytohormones under salinity stress. Journal of Stress Physiology & Biochemistry, 2011. 7(4).
- Nasir, F.A., et al., Free amino acids content in some halophytes under salinity stress in arid environment, Jordan. CLEAN–Soil, Air, Water, 2010. 38(7): p. 592-600.
- Parida, A.K., et al., Physiological, anatomical and metabolic implications of salt tolerance in the halophyte Salvadora persica under hydroponic culture condition. Frontiers in Plant Science, 2016. 7: p. 351.
- Kandasamy, S., et al., Understanding the molecular basis of plant growth promotional effect of Pseudomonas fluorescens on rice through protein profiling. Proteome Science, 2009. 7(1): p. 47.
- Yaghubi, K., et al., Potassium silicate alleviates deleterious effects of salinity on two strawberry cultivars grown under soilless pot culture. Scientia Horticulturae, 2016. 213: p. 87-95.
- Bano, A. and M. Fatima, Salt tolerance in Zea mays (L). following inoculation with Rhizobium and Pseudomonas. Biology and Fertility of Soils, 2009. 45(4): p. 405-413.
- Nadeem, S.M., et al., Variation in growth and ion uptake of maize due to inoculation with plant growth promoting rhizobacteria under salt stress. Soil Environ, 2006. 25(2): p. 78-84.
- Hamdia, M.A.E.-S., M. Shaddad, and M.M. Doaa, Mechanisms of salt tolerance and interactive effects of Azospirillum brasilense inoculation on maize cultivars grown under salt stress conditions. Plant Growth Regulation, 2004. 44(2): p. 165-174.
- Karlidag, H., et al., Plant growth-promoting rhizobacteria mitigate deleterious effects of salt stress on strawberry plants (Fragaria× ananassa). Hortscience, 2013. 48(5): p. 563-567.
- Babu, M., D. Singh, and K. Gothandam, The effect of salinity on growth, hormones and mineral elements in leaf and fruit of tomato cultivar PKM1. J Anim Plant Sci, 2012. 22(1): p. 159-164.
- Sofo, A., et al., Ascorbate peroxidase and catalase activities and their genetic regulation in plants subjected to drought and salinity stresses. International Journal of Molecular Sciences, 2015. 16(6): p. 13561-13578.
- Bianco, C. and R. Defez, Medicago truncatula improves salt tolerance when nodulated by an indole-3-acetic acid-overproducing Sinorhizobium meliloti strain. Journal of experimental botany, 2009. 60(11): p. 3097-3107. [CrossRef]
- Kohler, J., F. Caravaca, and A. Roldán, An AM fungus and a PGPR intensify the adverse effects of salinity on the stability of rhizosphere soil aggregates of Lactuca sativa. Soil Biology and Biochemistry, 2010. 42(3): p. 429-434. [CrossRef]
- El-Esawi, M.A., et al., Azospirillum lipoferum FK1 confers improved salt tolerance in chickpea (Cicer arietinum L.) by modulating osmolytes, antioxidant machinery and stress-related genes expression. Environmental and Experimental Botany, 2019. 159: p. 55-65.
- Zhang, N., et al., Effects of different plant root exudates and their organic acid components on chemotaxis, biofilm formation and colonization by beneficial rhizosphere-associated bacterial strains. Plant and soil, 2014. 374(1-2): p. 689-700.
- Meliani, A. and A. Bensoltane, Biofilm-mediated heavy metals bioremediation in PGPR Pseudomonas. J Bioremediat Biodegrad, 2016. 7(370): p. 2.
- Paulo, E.M., et al., An alternative method for screening lactic acid bacteria for the production of exopolysaccharides with rapid confirmation. Food Science and Technology, 2012. 32(4): p. 710-714.
- LUTTS, S., J.M. KINET, and J. BOUHARMONT, NaCl-induced Senescence in Leaves of Rice ( Oryza sativa L.) Cultivars Differing in Salinity Resistance. Annals of Botany, 1996. 78(3): p. 389-398.
- Zhang, H.-X. and E. Blumwald, Transgenic salt-tolerant tomato plants accumulate salt in foliage but not in fruit. Nature biotechnology, 2001. 19(8): p. 765.
- Hodges, D.M., et al., Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta, 1999. 207(4): p. 604-611. [CrossRef]
- Andreae, W. and M. Van Ysselstein, Studies on 3-indoleacetic acid metabolism. VI. 3-Indoleacetic acid uptake and metabolism by pea roots and epicotyls. Plant physiology, 1960. 35(2): p. 225.
- Bates, L.S., R.P. Waldren, and I. Teare, Rapid determination of free proline for water-stress studies. Plant and soil, 1973. 39(1): p. 205-207.
- Shi, H.-T., et al., Increasing nitric oxide content in Arabidopsis thaliana by expressing rat neuronal nitric oxide synthase resulted in enhanced stress tolerance. Plant and Cell Physiology, 2012. 53(2): p. 344-357. [CrossRef]
- Shi, H., et al., Analysis of natural variation in bermudagrass (Cynodon dactylon) reveals physiological responses underlying drought tolerance. PLoS One, 2012. 7(12): p. e53422.
- Cheng, Z., E. Park, and B.R. Glick, 1-Aminocyclopropane-1-carboxylate deaminase from Pseudomonas putida UW4 facilitates the growth of canola in the presence of salt. Canadian Journal of Microbiology, 2007. 53(7): p. 912-918. [CrossRef]
- Bradford, M.M., A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical biochemistry, 1976. 72(1-2): p. 248-254.
- Irigoyen, J., D. Einerich, and M. Sánchez-Díaz, Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd) plants. Physiologia plantarum, 1992. 84(1): p. 55-60.
- Arnon, D.I., Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant physiology, 1949. 24(1): p. 1.
- Ferus, P. and M. Arkosiova, Variability of chlorophyll content under fluctuating environment. Acta fytotechnica et zootechnica, 2001. 4(2): p. 123-125.
- Chamkouri, N., M. Torabpour, and F. Ghafarizadeh, Microwave assisted digestion coupled with inductively coupled plasma optical emission spectrometry for determining element concentrations in halophytes. Journal of Basic Research in Medical Sciences, 2015. 2(1): p. 49-56.
- Aebi, H., [13] Catalase in vitro, in Methods in enzymology. 1984, Elsevier. p. 121-126.
- Nakano, Y. and K. Asada, Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant and cell physiology, 1981. 22(5): p. 867-880. [CrossRef]
Figure 1.
Distance matrix tree based on 16S rDNA gene sequence alignment of selected PGPB bacteria: Bacillus pumilus strain MA9; Virgibacillus halodenitrificans strain MA14; Bacillus subtilus strain MA17 and Bacillus pumilus strain MA19 and other closely related bacteria.
Figure 1.
Distance matrix tree based on 16S rDNA gene sequence alignment of selected PGPB bacteria: Bacillus pumilus strain MA9; Virgibacillus halodenitrificans strain MA14; Bacillus subtilus strain MA17 and Bacillus pumilus strain MA19 and other closely related bacteria.
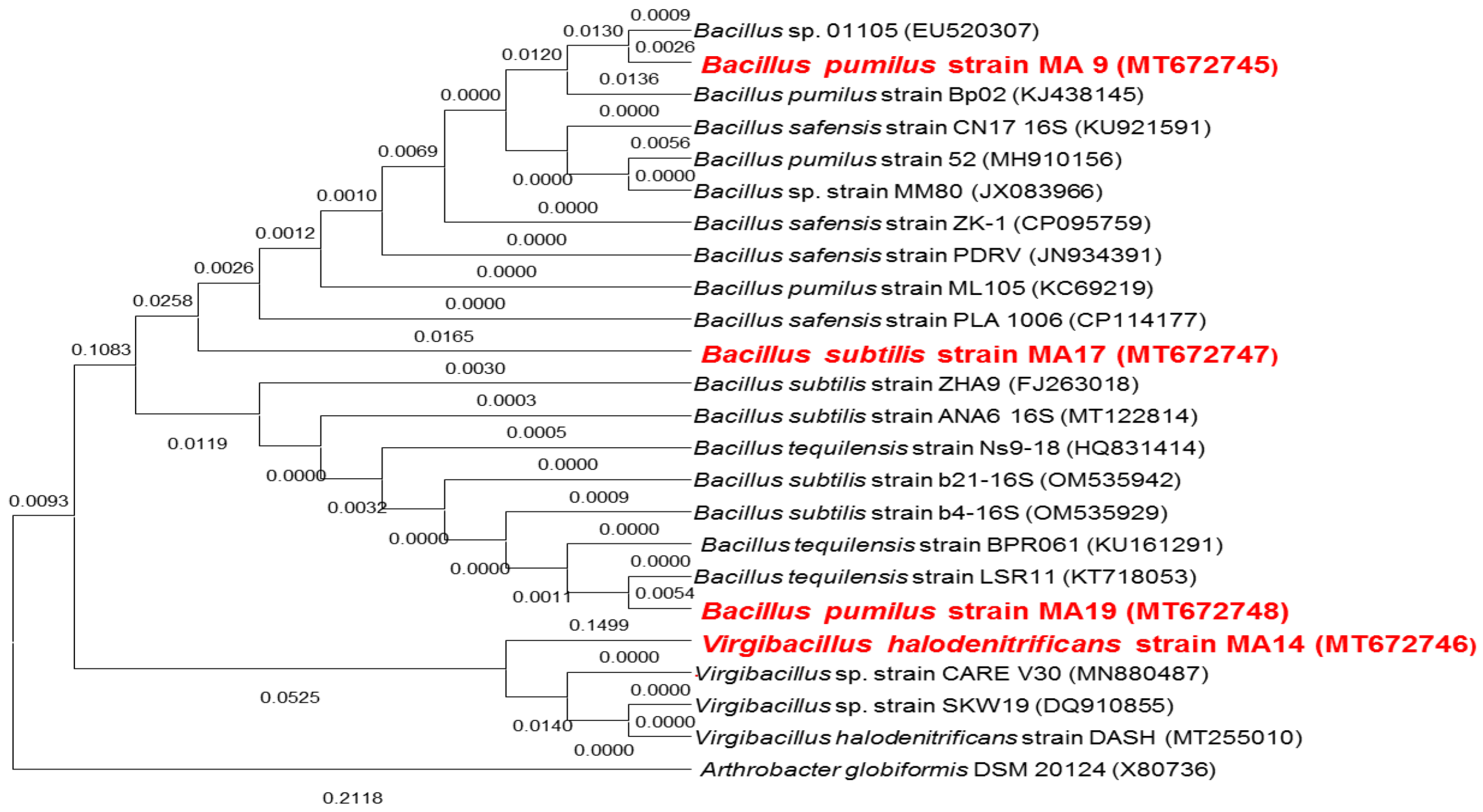
Figure 2.
Production of β-glucanase (A), proteases (B), phytases (C) and amylases (D) in vitro by bacterial culture of: 1: Bacillus pumilus strain MA9, 2: Virgibacillus halodenitrificans strain MA14, 3: Bacillus subtilis strain MA17 and 4: Bacillus pumilus strain MA19 after 7 days at 28 ± 2 °C. The formation of a halo around the culture shows a positive activity.
Figure 2.
Production of β-glucanase (A), proteases (B), phytases (C) and amylases (D) in vitro by bacterial culture of: 1: Bacillus pumilus strain MA9, 2: Virgibacillus halodenitrificans strain MA14, 3: Bacillus subtilis strain MA17 and 4: Bacillus pumilus strain MA19 after 7 days at 28 ± 2 °C. The formation of a halo around the culture shows a positive activity.
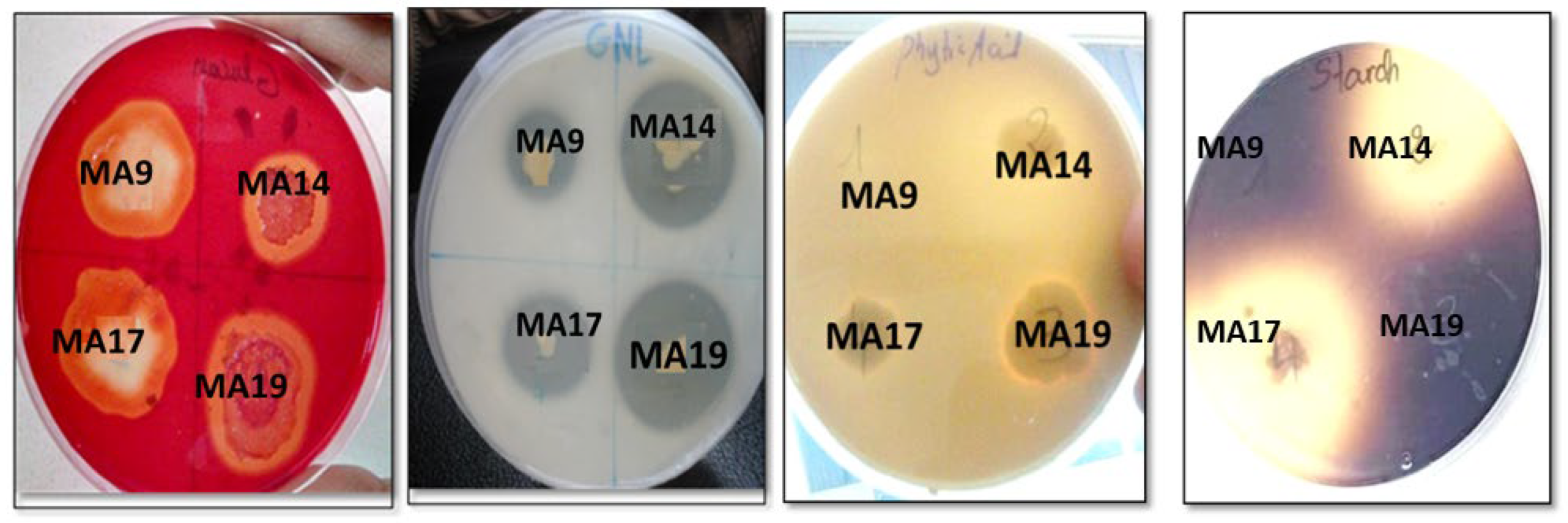
Figure 3.
Biocontrol of selected strains 1: Bacillus pumilus strain MA9; 2: Virgibacillus halodenitrificans strain MA14; 3: Bacillus subtilis strain MA17; 4: Bacillus pumilus strain MA19; against (a) Fusarium oxysporum (b) Fusarium graminearum (c) Fusarium solani, and (d) Rhizoctonia solani.
Figure 3.
Biocontrol of selected strains 1: Bacillus pumilus strain MA9; 2: Virgibacillus halodenitrificans strain MA14; 3: Bacillus subtilis strain MA17; 4: Bacillus pumilus strain MA19; against (a) Fusarium oxysporum (b) Fusarium graminearum (c) Fusarium solani, and (d) Rhizoctonia solani.
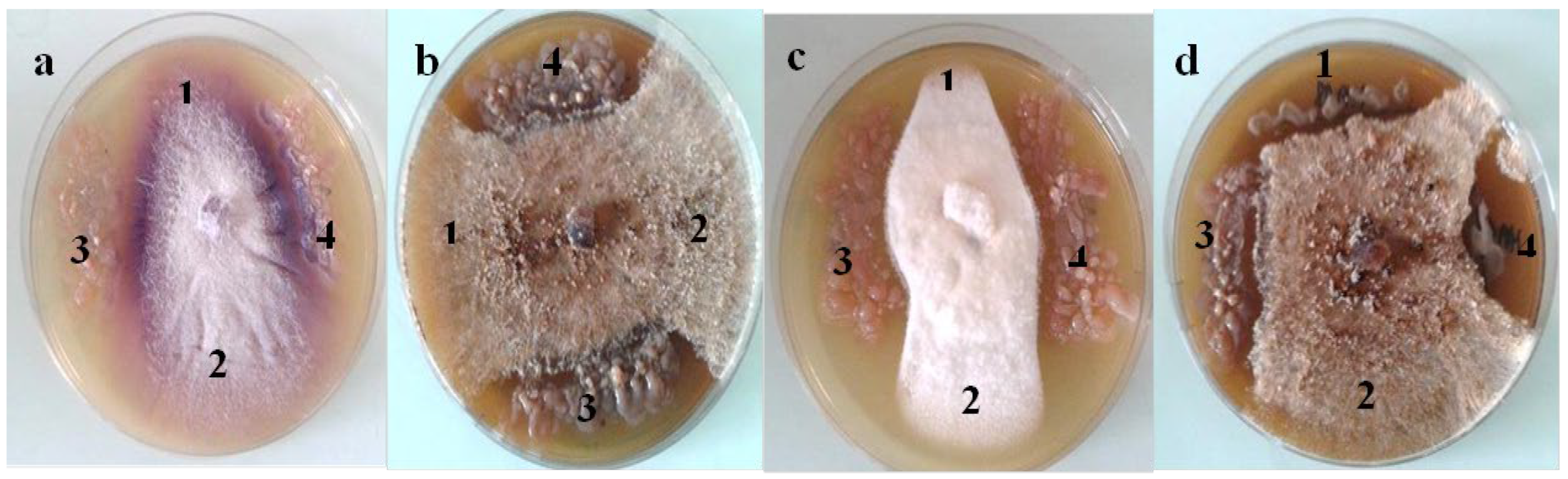
Figure 4.
Biofilm formation and exopolysaccharides production of selected PGPB strains: (MA9) Bacillus pumilus strain MA9; (MA14) Virgibacillus halodenitrificans strain MA14; (MA17) Bacillus subtilus strain MA17 and (MA19) Bacillus pumilus strain MA19.
Figure 4.
Biofilm formation and exopolysaccharides production of selected PGPB strains: (MA9) Bacillus pumilus strain MA9; (MA14) Virgibacillus halodenitrificans strain MA14; (MA17) Bacillus subtilus strain MA17 and (MA19) Bacillus pumilus strain MA19.
Figure 5.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of malonaldehyde MDA content (µmol/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
Figure 5.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of malonaldehyde MDA content (µmol/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
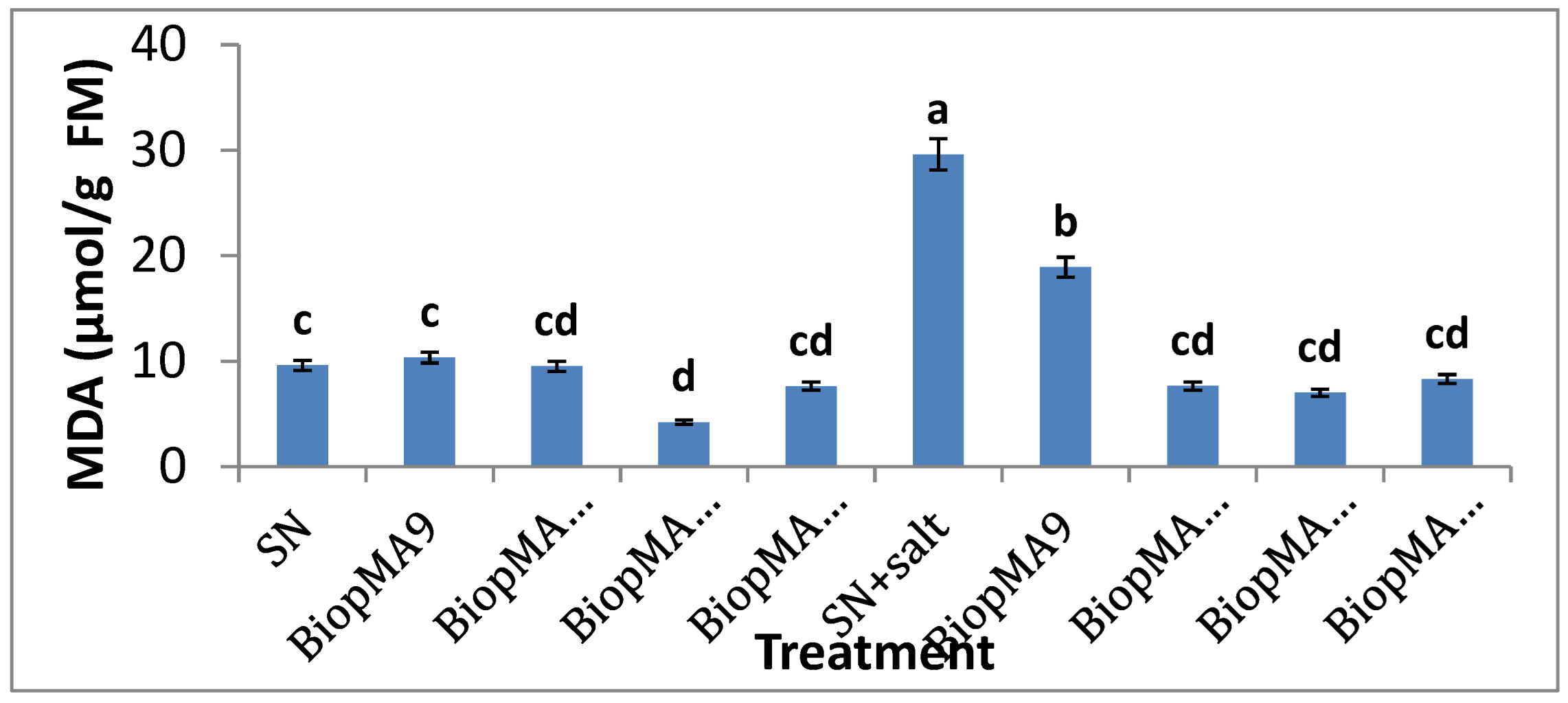
Figure 6.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of proline content (µg/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
Figure 6.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of proline content (µg/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
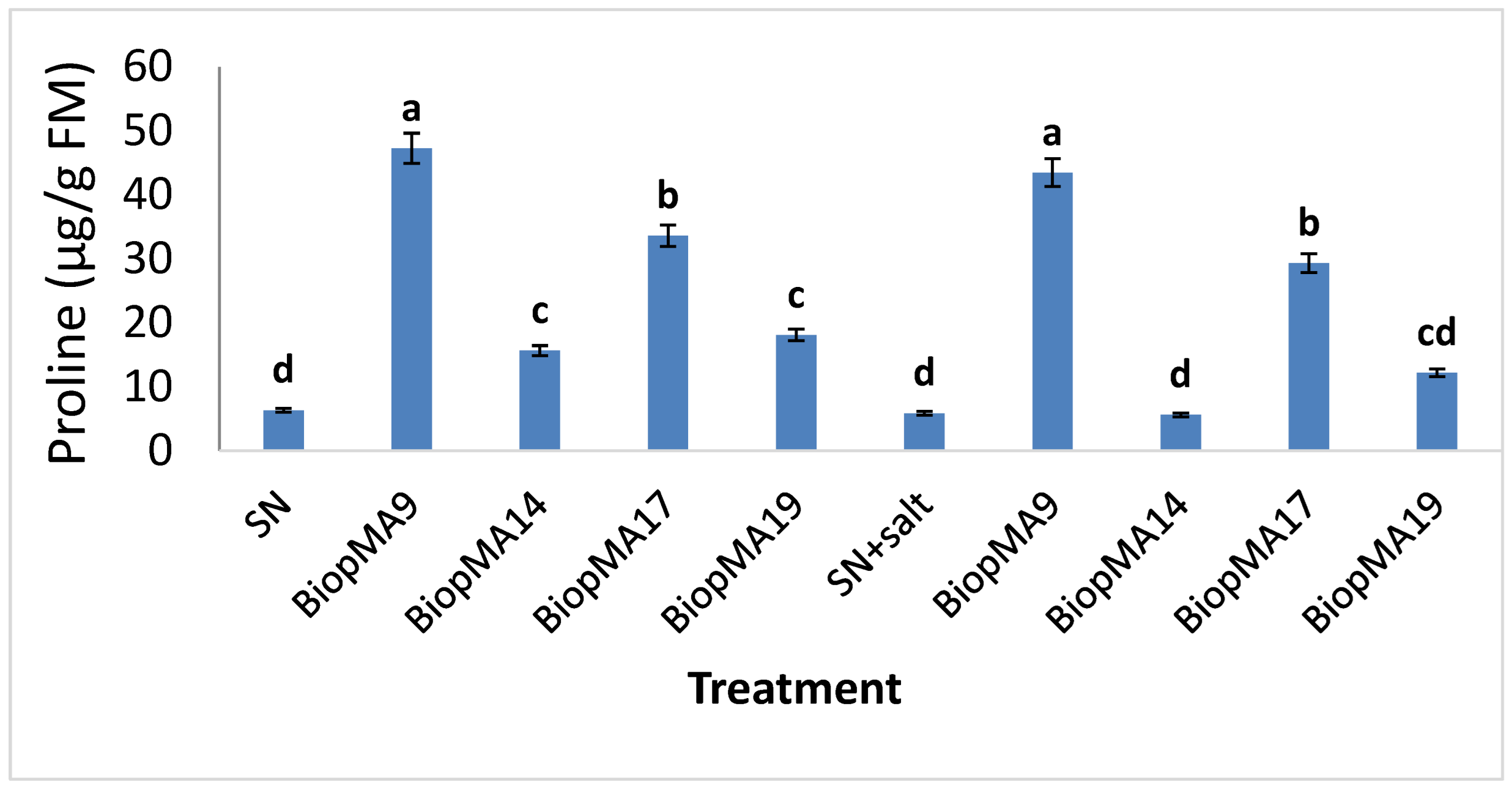
Figure 7.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of amino-acid (µM) (A) and protein content (B) (mg/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
Figure 7.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of amino-acid (µM) (A) and protein content (B) (mg/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
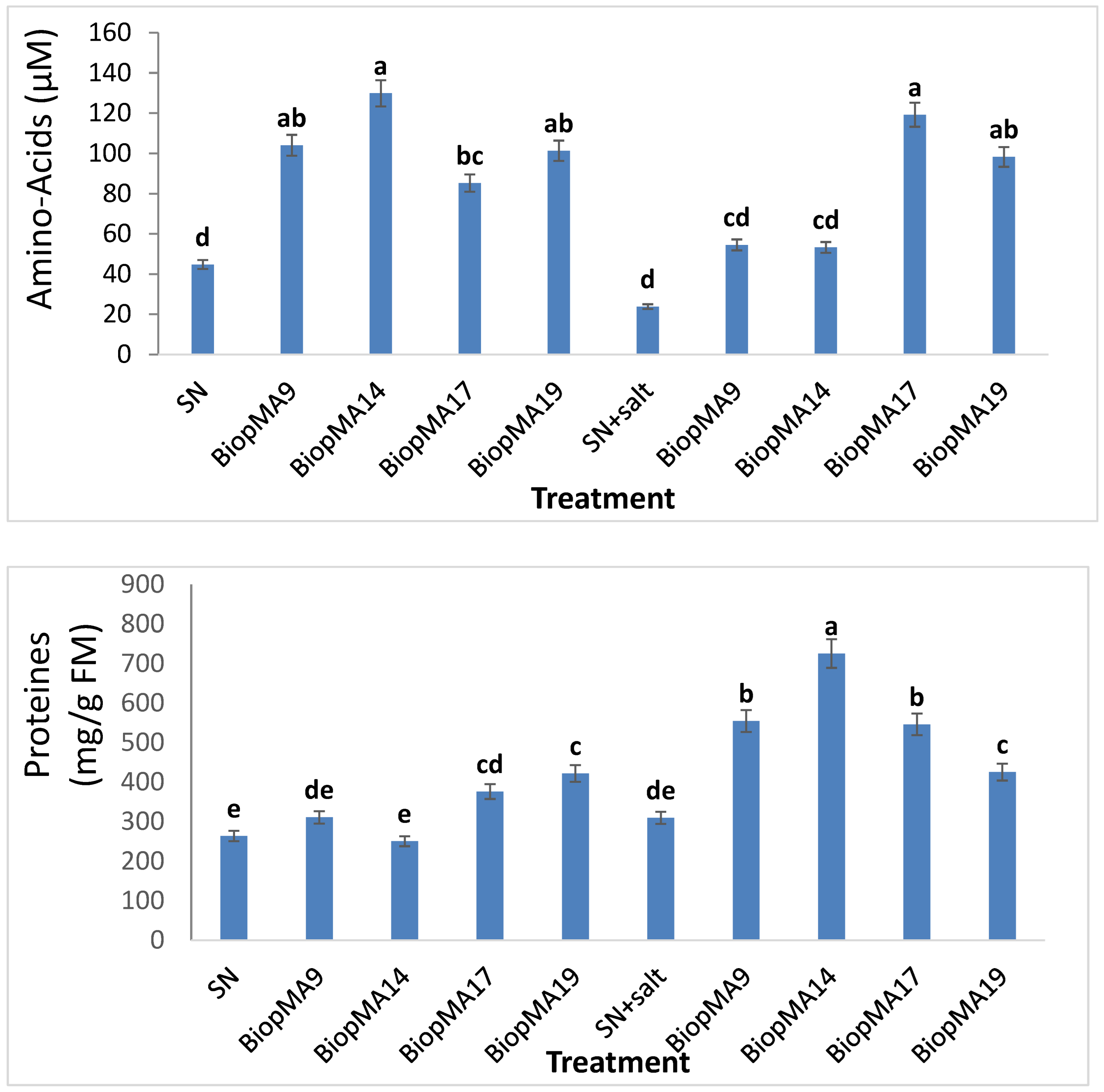
Figure 8.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of total sugar content (mg/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
Figure 8.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of total sugar content (mg/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
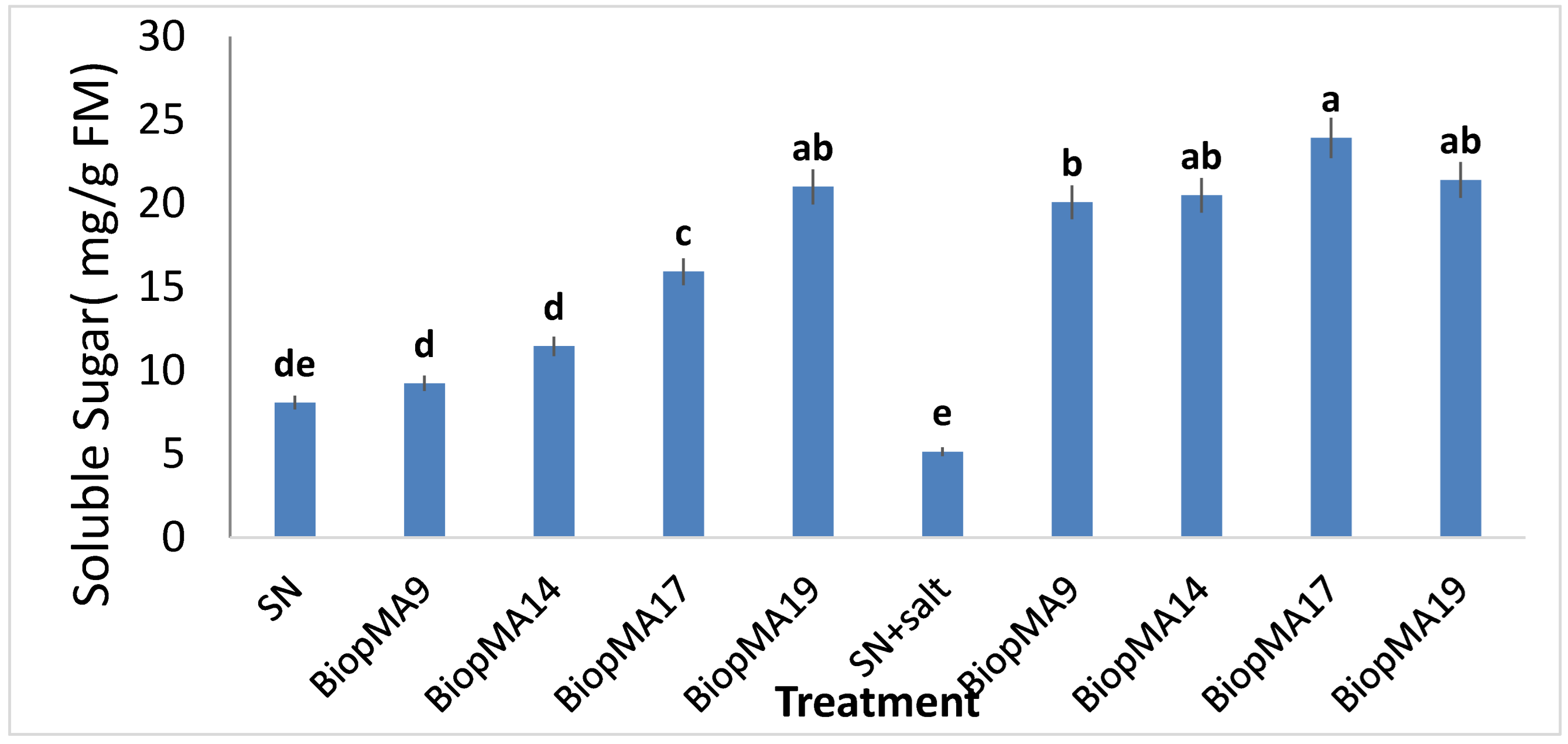
Figure 9.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of auxin AIA content (µmol/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
Figure 9.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of auxin AIA content (µmol/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
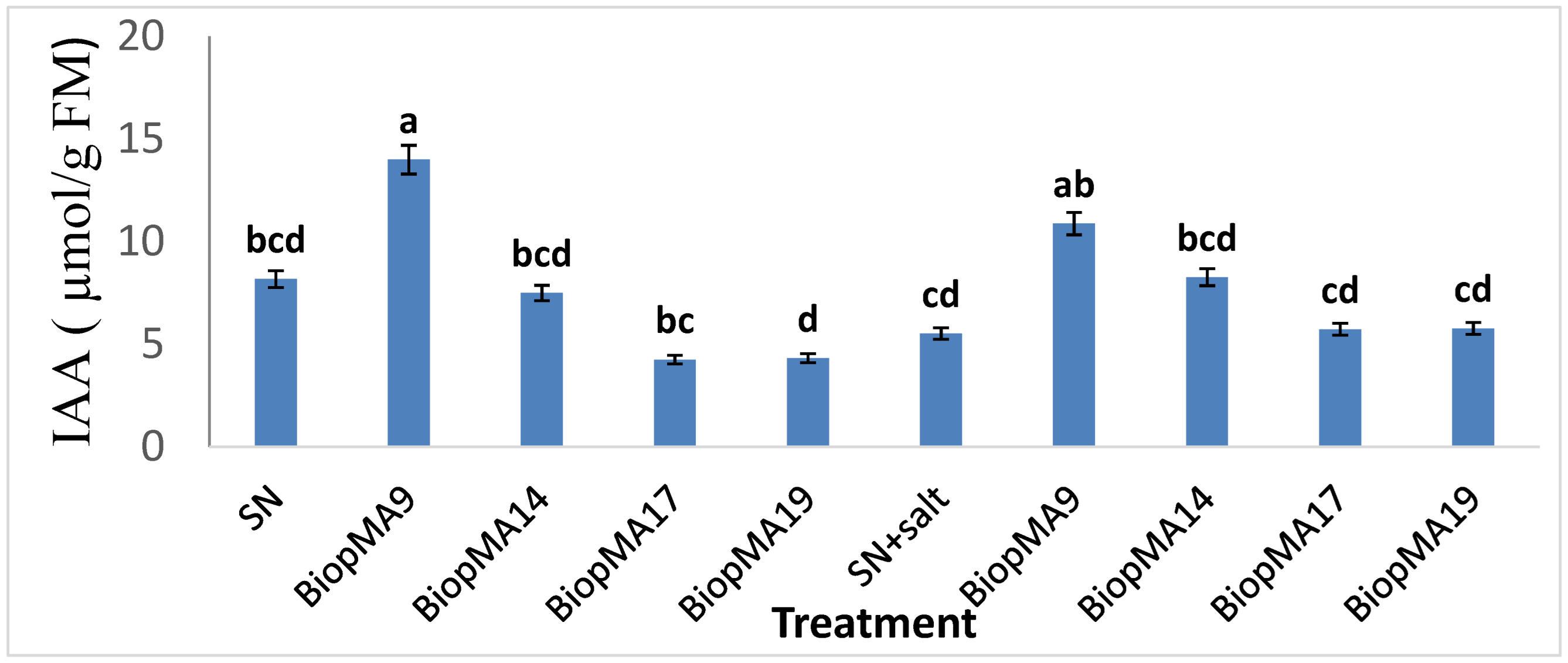
Figure 10.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of the chlorophyll pigment content (nmol/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
Figure 10.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of the chlorophyll pigment content (nmol/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
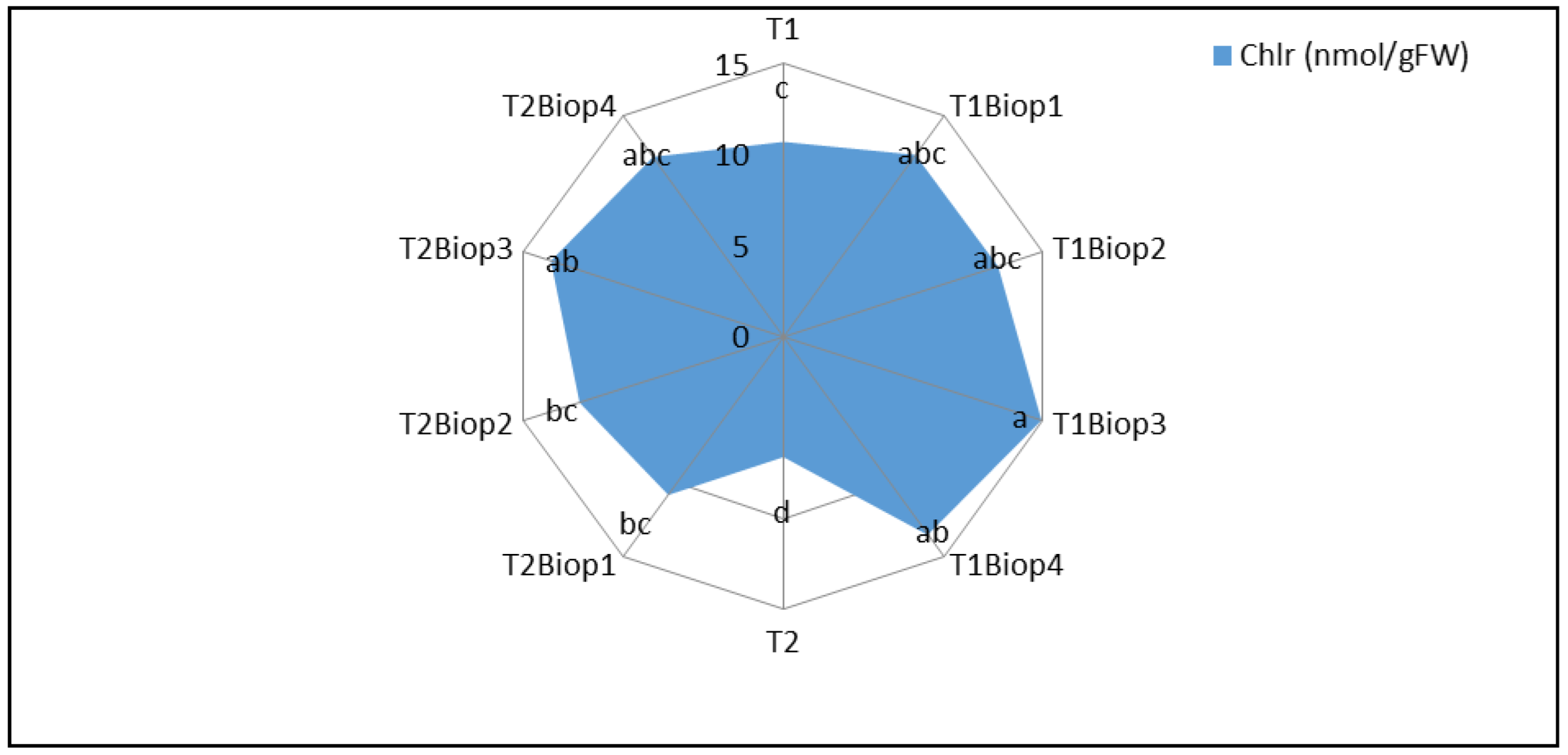
Figure 11.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of the ionic flux of K+ (A) and Na+ (B) (µg/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
Figure 11.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of the ionic flux of K+ (A) and Na+ (B) (µg/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
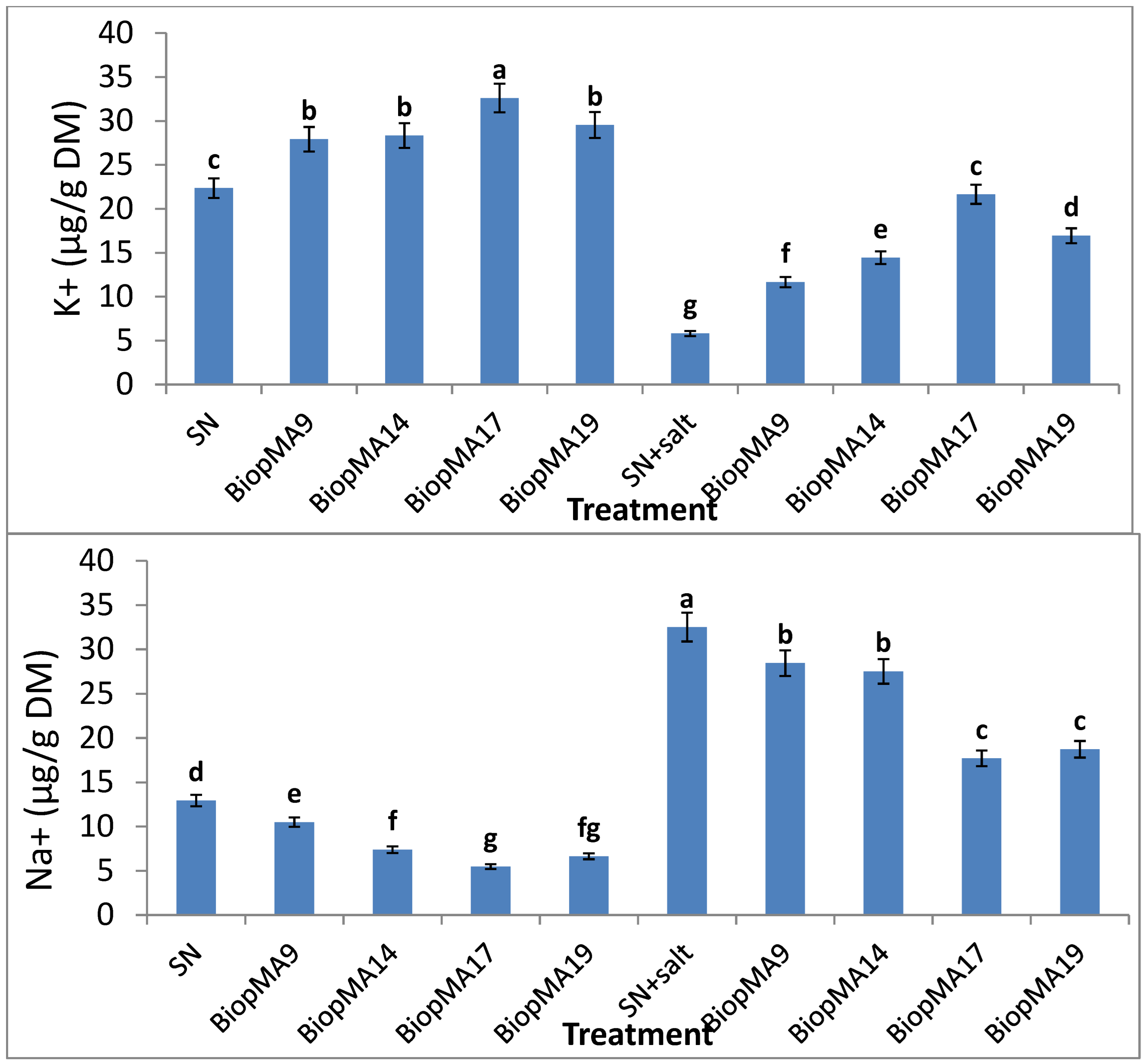
Figure 12.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of the content of the antioxidant enzymes catalase CAT (A) and ascorbate peroxidase APX (B) (nmol/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
Figure 12.
Seed biopriming with selected PGPB of durum wheat plantlets in terms of the content of the antioxidant enzymes catalase CAT (A) and ascorbate peroxidase APX (B) (nmol/g FW) under nonsaline and saline conditions (in planta). Two Control: SN = untreated plantlets (0 mM); SN+NaCl = treated plantlets (125 mM); Biop = biopriming seed; MA9 = Bacillus pumilus MA9; MA14 = Virgibacillus halodenitrificans MA14; MA17 = Bacillus subtilis MA17: MA19 = Bacillus safensis MA19.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



