
Submitted:
03 February 2026
Posted:
05 February 2026
You are already at the latest version
Abstract
Outbreaks of emerging and re-emerging diseases of both animals and plants are increasing due to climate change, globalization, land-use change, and agricultural intensification. While most pathogen monitoring programs focus on zoonotic threats, wildlife and other organisms in natural habitats can also serve as reservoirs and sentinels for pathogens of agricultural and ecological concern. Plant communities and the pathogens circulating within them are underrepresented in integrated disease monitoring frameworks. This study demonstrates how biodiversity and zoonosis monitoring programs conducted in protected habitats (tallgrass prairies and woodlands) across Illinois, together with insect specimens preserved in biorepositories, can be leveraged to improve knowledge of the identities and ecological associations of a wide range of potential pathogens. We developed an integrative workflow combining taxonomic identification, molecular screening, and epidemiological inference to detect vector-borne plant pathogens from archived insect material. Focusing on Hemiptera (Auchenorrhyncha), we screened specimens for phytoplasmas (Mollicutes), uncultured bacterial plant pathogens transmitted by sap-feeding insects, and characterized host-pathogen associations. At least three distinct phytoplasma strains were detected: ‘Candidatus Phytoplasma asteris’ (16SrI-B), ‘Candidatus Phytoplasma pruni’–related strains (16SrIII), and ‘Candidatus Phytoplasma sacchari’–related strains (16SrXI-H). The latter represents the first documented occurrence of a 16SrXI-H phytoplasma subgroup in Illinois. Overall, we identified five insect specimens harboring phytoplasmas across four preserved sites, all of them were previously unreported insect-phytoplasma associations. These findings demonstrate the value of existing biodiversity infrastructures for proactive surveillance of plant pathogens and extend the One Health paradigm to explicitly include natural ecosystems.
Keywords:
integrated wildlife surveillance
; leafhoppers
; One Health
; plant pathogen
; biodiversity
1. Introduction
One Health (OH) is a simple yet powerful framework for addressing emerging infectious diseases, as it seeks to build consensus around integrated solutions that safeguard health across interconnected biological systems [1]. However, OH has also been criticized as conceptually and operationally incomplete, often remaining anchored to targeted response-based approaches rather than more holistic and proactive system understanding [2]. In 2022, the One Health High-Level Expert Panel proposed a working definition intended to integrate principles from related frameworks such as EcoHealth and Planetary Health. This definition emphasizes the interdependence of human, animal, plant, and environmental health, and calls for coordinated action across sectors, disciplines, and societal levels to promote sustainable development and resilience [3]. Despite its integrative ambition, the operationalization of this definition remains challenging. In particular, its broad wording risks obscuring an epistemic gap between how the biosphere is perceived and managed and what is required to understand its underlying biological functioning. This gap limits critical awareness of systemic drivers of health and disease and fosters the perception of health crises as unpredictable or accidental. Addressing this gap requires critical learning processes, including the re-evaluation of inherited paradigms, context-specific knowledge generation, and the development of operational frameworks that counteract phenomena such as the “epistemology of ignorance” [4]. A further tension facing One Health frameworks is the need to maintain and promote a range of discipline-specific expertise (e.g., taxonomic), while fostering and building capacity for interdisciplinary collaborations in collection and analysis of data. The commonly adopted representation of One Health as a triad linking humans, animals, and the environment further reflects an anthropocentric framing, in which the environment is often reduced to a passive backdrop rather than an active, living system. Building on hybrid perspectives proposed by Furberg [5], we adopt a biosphere-centered view in which humans are embedded within, rather than positioned above, interconnected plant, microbial, animal, and abiotic components (Figure 1). From this perspective, health outcomes emerge from shared ecological space and interdependence, implying that management and surveillance strategies must operate in balance with the functioning of the entire biosphere.
Emerging infectious diseases (EIDs) are increasingly recognized as predictable yet highly complex phenomena, arising from land-use change, agricultural intensification, biodiversity loss, and climate-driven environmental shifts. Rather than being rare or stochastic events, disease emergence is now widely understood as the outcome of altered ecological opportunities that favor pathogen transmission and host switching. From this perspective, the One Health (OH) framework is particularly relevant for complex epidemiological systems such as vector-borne pathogens, whose transmission dynamics are inherently shaped by interactions among hosts, vectors, and the environment. It has been repeatedly documented that many pathogens circulate silently within alternative or secondary hosts, frequently associated with natural or semi-natural ecosystems, where they remain largely undetected. These enzootic or cryptic transmission cycles often persist without notice until pathogens spill over into humans or into species on which humans depend, such as crops or livestock. Within a One Health perspective, effective surveillance must therefore acknowledge the biosphere as an interconnected system and recognize that EIDs more often arise from changes in ecological opportunity rather than true pathogen novelty. Recent studies have highlighted the value of integrated approaches to wildlife disease monitoring. For example, Barroso et al. [6] demonstrated how Integrated Wildlife Monitoring in Europe can identify disease maintenance hotspots and key drivers of interspecific transmission. Similar conceptual and operational needs were emphasized by Cardoso et al. [7] and Machalaba et al. [8], who argued for closer integration between biodiversity science and health surveillance. Despite these advances, biodiversity monitoring and medical surveillance still rarely interact, even though they target the same biological space and often the same organisms.
The disconnection between disease surveillance and biodiversity monitoring is particularly evident for plant diseases. As with many animal (including human) pathogens, plant pathogens are most frequently detected in managed landscapes, such as agricultural fields and urban or peri-urban environments, where their impacts become economically or socially visible. By the time such pathogens are detected, opportunities for outbreak prevention are often limited. In this sense, managed ecosystems function as epidemiological observatories, revealing pathogen dynamics that have long remained undocumented in natural systems. In contrast, natural ecosystems are still poorly characterized from a plant pathogen perspective. Plant pathologists typically sample natural areas opportunistically, if at all, resulting in major knowledge gaps regarding pathogen diversity, host range, and circulation in wild plant and vector communities. This lack of baseline information severely constrains spillover risk assessment and predictive capacity, particularly under ongoing environmental change.
In this context, we aim to address this epistemic gap by proposing a joint surveillance framework that integrates biodiversity monitoring with pathogen detection. By leveraging existing biodiversity inventories, biorepositories, and monitoring programs for zoonotic disease surveillance at the Prairie Research Institute (University of Illinois at Urbana-Champaign), we outline an operational workflow for detecting, characterizing, and contextualizing pathogens circulating in wildlife. Using phytoplasma (non-yet-culturable bacteria within the class Mollicutes) and their vectors as a biological model, we integrate host-pathogen research with natural history collections to evaluate the potential spillover risk and strengthen One Health surveillance in natural and semi-natural ecosystems. Our objective is not to predict emergence events per se, but to improve baseline knowledge of pathogen presence and ecological context as a prerequisite for proactive and preventive One Health implementation.
2. Materials and Methods
2.1. Leveraged Monitoring Programs and Specimen Collection
Independent monitoring programs are conducted annually at the Prairie Research Institute (University of Illinois at Urbana-Champaign) to document both arthropod diversity and pathogen presence in natural and managed ecosystems across Illinois. In this study, we leveraged specimens collected through two distinct, but complementary monitoring efforts led by the Illinois Natural History Survey (INHS): (i) biodiversity surveys conducted by the INHS Leafhopper Lab; and (ii) public health surveillance conducted by the INHS Medical Entomology Lab. The biodiversity surveys focus on sampling auchenorrhynchan insects (Hemiptera) across natural habitats, primarily tallgrass prairies, with main objective of documenting the decline of prairie specialist and endangered species. The samples screened in this study were obtained through of the surveys conducted in 2021 (Figure 2). Specimens are collected using various standard entomological methods, including sweeping and vacuuming of vegetation, night collecting at lights, and in Malaise (flight intercept) traps.
Zoonotic disease surveillance monitoring primarily targets mosquito populations in urban parks, forest preserves, and other natural or semi-natural areas representing a gradient of residential land use and green space. This program aims to investigate vectors of zoonotic pathogens of public health and veterinary concern in Illinois. Mosquitoes and other hematophagous Diptera are collected using multiple trap types baited with dry ice, including CDC light traps, miniature ultraviolet (UV) light traps, as well as BG Sentinel-2 traps supplemented with additional attractant cues (UV-LED light source, BG-Lure® and/or 1-Octen-3-ol). Traps are operated for approximately 24 hours per sampling event, after which specimens were transported on dry ice and stored at −80 °C until processing (see Mackay et al. [9] for full methodological details). In addition to mosquitoes, these traps incidentally collect other insect orders and arthropods. The samples screened in this study were collected from woodland habitats in 2022 and 2024 (Figure 2). Future efforts may expand to screening a larger number of specimens collected during mosquito sampling conducted from 2023 to 2025 at 44 additional natural areas, encompassing a diverse range of habitats across Illinois.
All auchenorrhynchan specimens, including vectors of phytoplasmas (obligate vector-borne plant pathohens), were repurposed for phytoplasma screening and entered a specific workflow (see below).
2.2. Non-Destructive DNA Extraction
Specimens were initially preserved in 96% ethanol and stored in −20°C freezers at INHS facilities. DNA was extracted using a non-destructive lysis protocol allowing molecular screening while retaining voucher specimens for deposit in permanent collections at INHS [10].
Briefly, individual specimens were incubated at 56 °C for two days in a lysis buffer containing Tris-HCl, EDTA, and SDS, supplemented with Proteinase K (20 mg/mL) to facilitate tissue digestion. DNA purification was performed using an iminodiacetic acid–based chelating resin (20% working solution). After removal of the insect body, an equal volume of chelating resin solution was added to the lysate. A final 30-minute incubation at 56 °C completed the extraction. DNA extracts were stored at −20 °C or immediately analyzed for the presence of phytoplasmas.
2.3. Voucher Preparation, Host Association and Metadata Integration
Following DNA extraction, specimens were pinned, labelled, and assigned a unique INHS collection code. Vouchers were subsequently housed in standard entomological drawers within archival-quality collection cabinets at the INHS Insect Collection.
Associated metadata, including collection locality, date, collector, taxonomic identification, preservation status, INHS collection code, and infection status, were recorded in the INHS Insect Collection Database [11]. The digitized host–pathogen associations stored in the collection database constitute a documented, publicly accessible scientific record that can be revised as taxonomy is updated or as additional data become available.
2.4. Real-Time PCR Screening for Phytoplasmas and Conventional PCR Assays
Phytoplasma detection was performed using a universal probe-based quantitative PCR (qPCR) assay targeting the 16S rRNA gene. Reactions consisted of a commercial qPCR master mix containing hot-start Taq DNA polymerase, dNTPs, optimized buffer, and stabilizers. Amplification used a universal primer set (p16S-fw: 5′-CGTACGCAAGTATGAAACTTAAAGGA-3′; p16S-rv: 5′-TCTTCGAATTAAACAACATGATCCA-3′) and a hydrolysis probe labeled with a 5′ FAM reporter and dual 3′ quenchers ZEN and IBFQ (5′-FAM-TGACGGGAC-ZEN-TCCGCACAAGCG-IBFQ-3′). Ultrapure nuclease-free water was used for all reaction mixtures. Each qPCR plate included a phytoplasma-positive control to validate assay performance and monitor amplification efficiency. The reaction mixture without DNA templates were included as negative control.
Amplifications with conventional nested PCR assays were carried out on the 16S–23S ribosomal RNA gene for all qPCR-positive samples, yielding amplicons of about 1240 bp for the subsequent characterization of the phytoplasma strain by sequencing. The universal primer pairs for phytoplasmas P1/P7 [12,13], followed by R16F2n/R2 [14] were used. Tubes with the reaction mixture devoid of DNA templates were included in each experiment as negative control. As a positive control of both qPCR and PCR, we use DNA of the FD (Italian grapevine yellows) phytoplasma, provided by the Viticulture Research Centre (Conegliano, Italy), was used as a positive control in all amplification reactions. The PCR products were analyzed by 1% precast agarose gels with SYBR Safe DNA gel stain electrophoresis.
2.5. Sequencing, Phytoplasma Characterization and Phylogenetic Analysis
Amplicons obtained were purified through the PureLink PCR Purification Kit (Invitrogen, Waltham, MA, USA) and sequenced directly by the commercial sequencing facilities of DNA Service, Roy J. Carver Biotechnology Center, University of Illinois at Urbana-Champaign, US. Bidirectional sequencing of each amplicon was carried out using a primer-walking approach, employing the PCR primers and internal primers (P1/P7 and R16F2n/R2) to obtain complete sequence coverage. Raw sequences were trimmed of the unwanted 5′ and 3′ fragments generally characterized by low sequence quality and assembled using Staden [15]. All sequences obtained in this study were deposited in NCBI under the accession numbers listed in the “Data availability” paragraph and in Table 1.
Potential chimeric sequences were assessed using the DECIPHER package (v3.4) in R [16]. Assembled Sanger consensus sequences were screened using a reference-guided chimera detection approach (FindChimeras) against phytoplasma reference sequences. No chimeric sequences were detected.
Sequences from the same gene were aligned with MEGA7 [17], and the MUSCLE algorithm [18] was used for sequence alignments.
A maximum likelihood phylogenetic analysis was conducted using RAxML (Randomized Axelerated Maximum Likelihood) program [19] under the GTR+Γ+I model of nucleotide substitution, with among-site rate heterogeneity modelled using a gamma distribution and the proportion of invariant sites estimated by maximum likelihood. A maximum-likelihood analysis with thorough bootstrapping was carried out using 1,000 bootstrap replicates, with bootstrap support values mapped onto branch lengths. Reference strains used in the phylogenetic tree are reported in Supplementary Material Table S1.
3. Results
3.1. Phytoplasma Screening and Insect Museum Vouchers
Although the INHS biodiversity and zoonotic disease surveillance programs were designed for distinct ecological and public health objectives, and they are not focused directly on plant pathogen surveillance, the interoperability of curated specimens, associated metadata, and molecular workflows enabled a cross-sector surveillance approach consistent with One Health principles.
All specimens screened in this study (195 individuals) were dry point mounted following DNA extraction and are preserved as vouchers in the INHS Insect Collection. Despite molecular processing, the bodies of all specimens remained largely intact, allowing confident taxonomic assignment at the species level using traditional morphological criteria (Figure 3). Notably, specimens recovered as bycatch from mosquito surveillance traps showed fully intact bodies, enabling both molecular screening and morphological identification. The incidental capture of non-target insects, including Hemiptera, in mosquito surveillance traps has been documented previously and examination of such bycatch can yield valuable information that would otherwise go undetected, including new occurrence and distribution data for non-target taxa such as phytoplasmas [20,21,22,23,24,25].
Among the screened material, 16 specimens tested positive for phytoplasmas by qPCR but only 5 yielded amplifiable DNA for downstream sequencing (Supplementary Material Table S2). These specimens were identified as Macrosteles lepidus (Van Duzee, 1894), Osbornellus clarus Beamer, 1937, Latalus missellus (Ball, 1899), Pendarus punctiscriptus (Van Duzee, 1892), and Stictocephala brevis (Walker, 1851). Representative images of each positive specimen are shown in Figure 3. Voucher specimens are deposited under INHS collection codes 1086365, 1083960, 1083966, 1083969, 1089637 (Table 1).
3.2. BLAST Analysis, Phytoplasma Identification, and Phylogenetic Analysis
Five phytoplasma-positive samples yielded amplicons with sizes ranging from 1242 to 1669 bp (Supplementary Material Table S2). iPhyClassifier and BLAST analysis classified the five phytoplasma strains detected in three distinct ribosomal groups: 16SrI, 16SrIII and 16SrXI. Virtual RFLP analysis of the 16S rRNA gene F2nR2 fragment using iPhyClassifier assigned the phytoplasma detected in O. clarus to 16Sr group I, subgroup B, producing a restriction pattern identical to the reference profile of ‘Candidatus (Ca.) Phytoplasma (P.) asteris’ (GenBank accession AP006628; similarity coefficient = 1.00). Consistent with this classification, BLAST analysis of the 16S rRNA gene sequence revealed highest similarity to members of the ‘Ca. P. asteris’ clade, sharing 99.84% sequence identity to phytoplasma strains previously reported from Vitis vinifera in Pennsylvania, USA (GenBank accessions KX236145 and KX236147), and to a phytoplasma detected in the putative vector Amblysellus necopinus from Mexico (PV564083).
Phylogenetic analysis based on the 16S-23S rRNA gene region placed the phytoplasma sequence detected in O. clarus within the 16SrI clade, clustering with reference strains of ‘Ca. P. asteris’ from the eastern and central United States (Figure 4). The sequence grouped most closely with phytoplasma strains detected in insect hosts from Mexico and United States, A. necopinus and Macrosteles sp. (PV564083 and JF705931). This insect-associated clade was separated from lineages comprising phytoplasmas detected in plant hosts, including Thlaspi arvense, Vitis, potato and coneflower, collected in neighboring states, with strong bootstrap support (99%).
Two phytoplasmas belonging to ‘Candidatus Phytoplasma pruni’-related strains (16SrIII) were independently detected in M. lepidus and S. brevis at two nearby sites in eastern Illinois. Virtual RFLP analysis using iPhyClassifier assigned the phytoplasma detected in M. lepidus to 16SrIII group. The virtual restriction pattern was most similar to the reference profile of subgroup 16SrIII-J (GenBank accession AF147706), with a similarity coefficient of 0.98, indicating that this strain represents a variant of the 16SrIII-J subgroup. Consistent with this classification, BLAST analysis of the 16S rRNA gene sequence revealed highest similarity to phytoplasma strains belonging to the 16SrIII clade, supporting its placement within this group. In particular, this strain shared 98.47% with a strain detected in a wild savanna plant, Aegiphila verticillata, collected in Brazil (KT148597) and assigned to 16SrIII-J subgroup [26], as well as in cultivated Corylus avellana collected in Oregon, USA (AF189288), assigned to 16SrIII-B subgroup [27].
In contrast, virtual RFLP analysis of the phytoplasma detected in S. brevis produced a restriction pattern that was distinct from all previously established 16Sr group III subgroups. The most similar reference pattern corresponded to subgroup 16SrIII-F (GenBank accession AF510724), but with a similarity coefficient of 0.89, below the accepted threshold (≤0.97) for subgroup assignment. This result suggests that the phytoplasma detected in S. brevis may represent a putative new subgroup within the 16SrIII group. BLAST analysis confirmed affiliation with the 16SrIII clade and showed its highest sequence similarity to a strain reported from milkweed collected in New York, USA, assigned to the 16SrIII-F subgroup (HQ589200.1). The strain from M. lepidus and S. brevis showed 98.31% sequence similarity to each other.
Phylogenetic reconstruction indicated that the S. brevis phytoplasma clustered with reference strains assigned to the 16SrIII-F subgroup, whereas the strain detected in M. lepidus formed a sister lineage to this clade (Figure 4).
Two additional phytoplasma strains belonging to the ‘Candidatus Phytoplasma sacchari’-related strains were detected in L. missellus and P. punctiscriptus collected during the same sampling event at a site in western Illinois. BLAST analysis revealed that the phytopalsma sequence detected in L. missellus shared only 95.67% pairwise sequence similarity with a phytopalsma strain previously detected in Psammottetix cephalotes from Germany (HQ589192). Due to insufficient sequence coverage, virtual RFLP analysis could not be reliably performed for the phytoplasma detected in L. missellus, precluding formal subgroup designation for this strain, even though it was assigned to ‘Candidatus Phytoplasma sacchari’-related strain. In contrast, the phytoplasma detected in P. punctiscriptus yielded a longer sequence, allowing robust classification. This strain showed highest sequence similarity (99.78%) to a phytoplasma previously reported from the wild prairie grass Bothriochloa laguroides collected in Texas (OR711913, [28]), the reference strain of subgroup 16SrXI-H. Virtual RFLP analysis using iPhyClassifier produced a restriction pattern identical to the 16SrXI-H reference profile (similarity coefficient = 1.00), confirming assignment of this strain to subgroup 16SrXI-H. Phylogenetic reconstruction further supported this placement, with P. punctiscriptus phytoplasma clustering with reference strains assigned to the 16SrXI-H subgroup, whereas the strain detected in L. missellus diverged from this clade (Figure 4).
3.3. Novel Insect-Phytoplasma Associations in Natural Areas
Macrosteles lepidus is a macropterous Nearctic leafhopper that is widespread and common in eastern United States [29]. According to previous studies conducted in tallgrass prairies in Illinois, this species completes more than two generations per year, is associated with at least two plant families, and is more frequently linked to ecotonal or disturbed habitats or rather than pristine tallgrass prairies. Accordingly, it has been assigned a moderately low coefficient of conservatism [30]. To our knowledge, M. lepidus was not previously reported as a phytoplasma-associated host. Its preferred plant hosts are unknown but likely include sedges or grasses that occur in moist woodlands. In this study, M. lepidus collected in a restored tallgrass prairie mixed with woody areas in Illinois (Vermillion County) is reported for the first time to be infected with a ‘Ca. P. pruni’-related strain. Stictocephala brevis is a macropterous Nearctic membracid that appears to be polyphagous, alternating between woody oviposition hosts and herbaceous foodplants particularly those belonging to Asteraceae and Fabaceae [31]. Similarly, a ‘Ca. P. pruni’-related strain was found infecting S. brevis collected in a similar habitat in another location in the same area. The leafhopper species Osbornellus clarus is reported in Central and North America [32,33], and was found infected with a strain of ‘Ca. P. asteris’. Latalus missellus and Pendarus punctiscriptus are both largely restricted to native grasslands where they feed on perennial grasses although their specific host preferences remain unknown.
4. Discussion
Pathogen vectors have been recognized as key (sentinel) taxa for tracking elusive pathogens in One Health centered surveillance programs [34]. Pathogen elusiveness stems from the complex trophic networks present in most natural ecosystems, as well as diagnostic challenges (including asymptomatic hosts, or low rates of prevalence at which certain pathogens occur), and epistemic gaps. Our findings demonstrate the potential to leverage existing wildlife and biodiversity monitoring infrastructures for proactive surveillance of plant pathogens within natural ecosystems. By linking ecological, taxonomic, and molecular data, our approach extends the One Health paradigm to explicitly include natural ecosystems, advancing early detection and cross-sector surveillance of emerging pathogens at the wildlife-crop interface.
By repurposing specimens and data generated for biodiversity assessment and medical entomology, we expanded plant pathogen screening efforts without additional field sampling, illustrating the value of museum-enabled research infrastructures for integrative pathogen surveillance. Although mosquito surveillance is primarily designed to assess vectors of human health concern, incidental capture of non-target arthropods, including Hemiptera, has been documented previously and shown to provide valuable biodiversity and distributional data when examined systematically [20]. Our findings extend this perspective by demonstrating that such bycatch can also retain sufficient morphological integrity and DNA quality (often preserved by cold-chain handling during arbovirus surveillance) for plant pathogen screening and phylogenetic characterization. This underscores the value of interoperable surveillance infrastructures, where standardized trapping, specimen preservation, and museum curation enable reuse of samples beyond their original purpose. Other examples where such interoperability is relevant include detection of vectors of veterinary concern, such as Ceratopogonid biting midges or Simuliid black flies that are frequently found as by-catch in mosquito collections (Mackay et al., unpublished data). Within a One Health framework, the integration of biodiversity surveys, zoonotic entomology surveillance, and museum collections offers an efficient and scalable approach to detect pathogens across ecological compartments, while maximizing the scientific return of existing monitoring efforts.
Although none of the insect species in which phytoplasma DNA was detected can currently be considered competent vectors, as transmission trials were not conducted, we adopted a strict and standardized workflow to ensure reliable detection of phytoplasmas in insect bodies and to document the presence of phytoplasma strains circulating natural areas in Illinois. While these species may not represent true vectors under natural conditions and may have acquired phytoplasmas incidentally through feeding or probing activities, they can be regarded as biological accumulators that reveal the presence of phytoplasma strains in natural habitats, potentially associated with still unidentified host plants. In phytoplasma research, it is not uncommon for insect species to remain classified as ‘potential’ vectors due to the logistical challenges associated with establishing laboratory colonies and conducting controlled transmission experiments (REF). Indeed, Weintraub et al. [35] reported approximately 200 known and potential phytoplasma vectors, of which transmission competence has been experimentally confirmed for only about half. Furthermore, other authors provided evidence that vector competence in phytoplasmas may be phylogenetically constrained [36], opening avenues for inference based on evolutionary relationships when experimental trials are not feasible. Nevertheless, all insect-based phytoplasma detections reported here should be interpreted with caution and viewed as a foundation for future experimental validation, while they still represent a valuable surveillance tool for uncovering cryptic pathogen circulation and guiding future studies aimed at identifying true vectors and reservoir hosts.
In particular, the close phylogenetic relationship between the phytoplasma detected in Osbornellus clarus and phytoplasmas previously reported from insect hosts associated with ‘Ca. P. asteris’ is consistent with the broad host range and epidemiological versatility that characterize this phytoplasma lineage worldwide [37,38,39]. Osbornellus clarus is considered a polyphagous species, being associated with more than one family of herbaceous host plants, including Asteraceae [40]. This species is not known as an agricultural pest or of vector of plant pathogens. However, other species within the genus Osbornellus are known or suspected phytoplasma vectors. These include the Palearctic Osbornellus horvathi, a competent vector of aster yellow phytoplasma in Sicily (Italy) [41]; the Nearctic Osbornellus borealis, a competent vector of X-disease phytoplasma (16SrIII phytoplasma group) affecting peach in California [42]; Osbornellus dabeki, a potential vector of coconut lethal yellowing phytoplasma (16SrIV phytoplasma group) in Jamaica [43]; and the Nearctic Osbornellus auronitens, a potential vector of an Alder yellows phytoplasma-related strain (16SrV-C phytoplasma) in Switzerland [44]. In our study, the specimen of O. clarus found infected with ‘Ca. P. asteris’ was collected in a relatively closed canopy woody area with moderate-dense understory vegetation in the Sangchris Lake State Park in Central Illinois.
Although some species of Macrosteles are well known as competent vectors of aster yellows phytoplasmas (group 16SrI), other species within this genus, including M. cristata and M. laevis, have been reported as competent vectors of phytoplasmas belonging to subgroup 16SrIII-B (X-disease phytoplasma) in Europe [45]. In the Czech Republic, X-disease phytoplasma has also been detected in the native North American perennial Echinacea purpurea (Asteraceae) cultivated as ornamental plants [46]. In this context, the association between M. lepidus and a ‘Ca. P. pruni’ -related strain (a variant of the 16SrIII-J) detected in a restored tallgrass prairie habitats in Illinois appears biologically plausible. Together, these findings suggest that species of Macrosteles may play a broader role in the ecology and transmission of 16SrIII phytoplasmas across both managed and natural ecosystems. Thus, further investigation into their vector competence and host plant associations may help to better understand the risk factor that this association poses. Stictocephala bisonia was found infected with ‘Ca. P. mali’ in apple crops in Italy [47] and carrots in Serbia [48] and was also reported to have been infected accidentally with stolbur phytoplasma in a vineyard [49] in Italy. Unfortunately, in both cases the phytoplasma load was too low to be able to further characterize the strains. The independent detection of two distinct ‘Ca. P. pruni’ -related strains in M. lepidus and S. brevis at two close sites in eastern Illinois suggests independent epidemiology within the same landscape given that the species belongs to two different families Cicadellidae and Membracidae. Similar sympatric occurrence of divergent ‘Ca. P. pruni’ -related strains has been reported in both plant and insect hosts in western USA and is thought to reflect complex transmission networks involving multiple reservoir plants and vector species [50,51,52]. These findings support the idea and precious evidence for phytoplasma diversity in natural ecosystems being underestimated, particularly when sampling is focused primarily on symptomatic crops rather than insect communities [53].
The detection of ‘Ca. Phytoplasma sacchari’–related strains in L. missellus and P. punctiscriptus from a natural prairie habitat in Illinois is consistent with previous reports of 16SrXI phytoplasmas from non-agricultural systems [28]. Notably, the closest match to the P. punctiscriptus phytoplasma was a strain detected in the wild prairie grass Bothriochloa laguroides in Texas, which was also collected in a natural grassland ecosystem rather than from cultivated crops. For these prairie-associated records, neither the definitive plant reservoirs nor the competent insect vectors have been identified, and their epidemiological cycles remain under investigation. The recurrence of closely related 16SrXI-H phytoplasmas in geographically distant but ecologically similar natural habitats suggests the existence of a cryptic epidemiology involving native plants and insect communities that are largely overlooked by crop-focused surveys.
Although vector competence cannot be inferred from detection alone, the presence of 16SrXI phytoplasmas in leafhopper species collected from natural areas supports the use of insects as sentinels for revealing phytoplasma diversity in unmanaged ecosystems. The divergent and incompletely characterized strain detected in L. missellus further highlights that additional phytoplasma diversity remains to be discovered and will require further investigation through expanded screening and sampling efforts.
Some of the specimens analyzed here were obtained as bycatch from CDC light traps and related suction-based surveillance devices, which are primarily optimized for monitoring host-seeking adult mosquitoes [54] rather than plant-associated Hemiptera. These traps typically use incandescent or long-wave ultraviolet (UVA) light sources, often supplemented with carbon dioxide, which attract a broad range of insects [55], including a wide taxonomic range of non-target species [56,57], but may underrepresent taxa more efficiently sampled through vegetation-based methods, sweep netting, or Malaise traps. In addition, the use of relatively coarse catch container mesh (30-mesh; ~0.6 mm opening) in CDC light traps used during the 2022 and 2024 sampling seasons may have resulted in the loss of some very small-bodied hemipterans, particularly members of the subfamily Typhlocybinae, which are among the smallest phytoplasma-associated leafhoppers. Larger-bodied hemipteran insects that include most confirmed and putative phytoplasma vectors are unlikely to have been affected by this mesh size. In contrast, UV and BG traps deployed in this study employed finer mesh catch containers, improving retention of small-bodied taxa.
Despite these limitations, the approach offers important advantages. Leveraging existing biodiversity and mosquito surveillance infrastructure and systematically screening non-target bycatch enables cost-effective, large-scale detection of phytoplasmas across diverse landscapes without the need for dedicated phytoplasma-focused sampling. This strategy is particularly valuable for detecting pathogen circulation in natural and semi-natural habitats where host plants and vectors remain poorly characterized.
From a One Health perspective, integrating plant pathogen screening into entomological surveillance frameworks traditionally focused on human and animal health or on conservation of biodiversity creates opportunities for early detection, hypothesis generation, and cross-sectoral risk assessment. While targeted vegetation sampling and vector competence studies remain essential for resolving epidemiological cycles, opportunistic analysis of surveillance bycatch provides a powerful complementary tool for understanding pathogen ecology at the interface of plant, insect, and environmental health.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Phytoplasma reference strains used in this paper; Table S2: List of 16 Auchenorrhyncha insects tested positive for phytoplasmas and gel electrophoresis signal after nested PCRs.
Author Contributions
Conceptualization, V.T., A.J.M., C.M.S., C.H.D.; methodology, VT, A.J.M.; formal analysis, V.T.; investigation, V.T., A.J.M.; resources, V.T., A.J.M., C.M.S., C.H.D.; data curation, V.T.; writing—original draft preparation, V.T.; writing—review and editing, A.J.M., C.M.S., C.H.D.; visualization, V.T.; supervision, V.T.; project administration, V.T., C.M.S.; funding acquisition, V.T., C.M.S., C.H.D. All authors have read and agreed to the published version of the manuscript.
Funding
this study was supported by US National Science Foundation Grant DEB-2244871 and the State of Illinois Used Tire Management and Emergency Public Health funds.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequence data generated in this study have been deposited in the National Center for Biotechnology Information (NCBI) database under accession numbers PX789936- PX789940.
Acknowledgments
The first author thanks Matilde Tessitori for useful suggestions in interpreting the identity of a new phytoplasma strain collected in natural areas. The authors thank Aurora Marguccio for conceiving, planning, and creating the graphical abstract; the illustration is an original, hand-drawn artwork. The authors thank Corrado Cara for assistance in sorting Auchenorrhynchan insects from mosquitoes.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- One Health Joint Plan of Action (2022‒2026): Working Together for the Health of Humans, Animals, Plants and the Environment Available online: https://www.who.int/publications/i/item/9789240059139 (accessed on 17 December 2025). [CrossRef]
- Giraudoux, P.; Besombes, C.; Bompangue, D.; Guégan, J.-F.; Mauny, F.; Morand, S. One Health or ‘One Health Washing’? An Alternative to Overcome Now More than Ever. CABI One Health 2022, 2022. [CrossRef]
- Panel (OHHLEP), O.H.H.-L.E.; Adisasmito, W.B.; Almuhairi, S.; Behravesh, C.B.; Bilivogui, P.; Bukachi, S.A.; Casas, N.; Becerra, N.C.; Charron, D.F.; Chaudhary, A.; et al. One Health: A New Definition for a Sustainable and Healthy Future. PLOS Pathogens 2022, 18, e1010537. [CrossRef]
- Stockly, O. The Epistemology of Ignorance. Anthós 2011, 3. [CrossRef]
- Furberg, E. Are We Persons or Animals? Exposing an Anthropocentric Bias and Suggesting a Hybrid View. Ethics, Medicine and Public Health 2017, 3, 279–287. [CrossRef]
- Barroso, P.; Relimpio, D.; Zearra, J.A.; Cerón, J.J.; Palencia, P.; Cardoso, B.; Ferreras, E.; Escobar, M.; Cáceres, G.; López-Olvera, J.R.; et al. Using Integrated Wildlife Monitoring to Prevent Future Pandemics through One Health Approach. One Health 2023, 16, 100479. [CrossRef]
- Cardoso, B.; García-Bocanegra, I.; Acevedo, P.; Cáceres, G.; Alves, P.C.; Gortázar, C. Stepping up from Wildlife Disease Surveillance to Integrated Wildlife Monitoring in Europe. Research in Veterinary Science 2022, 144, 149–156. [CrossRef]
- Machalaba, C.; Uhart, M.; Ryser-Degiorgis, M.-P.; Karesh, W.B. Gaps in Health Security Related to Wildlife and Environment Affecting Pandemic Prevention and Preparedness, 2007–2020. Bull. World Health Organ. 2021, 99, 342-350B. [CrossRef]
- Mackay, A.J.; Cara, C.; Kim, C.-H.; Stone, C.M. Update on the Distribution of Culiseta Melanura in Regions of Illinois with Prior Eastern Equine Encephalitis Virus Activity. Journal of Vector Ecology 2022, 47. [CrossRef]
- Brown, M.E.; Ottati, S.; Trivellone, V. A Non-Destructive, Fast, Inexpensive, Non-Toxic Chelating Resin-Based DNA Extraction Protocol for Insect Voucher Specimens and Associated Microbiomes. Journal of Insect Science 2025, 25, 17. [CrossRef]
- McElrath, T. Illinois Natural History Survey Insect Collection. Illinois Natural History Survey. Occurrence Dataset. Available online: (accessed on 2 January 2026). [CrossRef]
- Deng, S.; Hiruki, C. Amplification of 16S rRNA Genes from Culturable and Nonculturable Mollicutes. Journal of Microbiological Methods 1991, 14, 53–61. [CrossRef]
- Smart, C.D.; Schneider, B.; Blomquist, C.L.; Guerra, L.J.; Harrison, N.A.; Ahrens, U.; Lorenz, K.H.; Seemüller, E.; Kirkpatrick, B.C. Phytoplasma-Specific PCR Primers Based on Sequences of the 16S-23S rRNA Spacer Region. Appl Environ Microbiol 1996, 62, 2988–2993.
- Lee, I.M.; Bertaccini, A.; Vibio, M.; Gundersen, D.E. Detection of Multiple Phytoplasmas in Perennial Fruit Trees with Decline Symptoms in Italy. Phytopathology 1995, 85, 728–735. [CrossRef]
- Bonfield, J.K.; Smith, K. f; Staden, R. A New DNA Sequence Assembly Program. Nucleic Acids Res 1995, 23, 4992–4999.
- Wright, E.S. Fast and Flexible Search for Homologous Biological Sequences with DECIPHER V3. The R Journal 2025, 16, 191–200. [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res 2004, 32, 1792–1797. [CrossRef]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A Graphical Interface and Toolkit for Phylogenetic Analyses Using RAxML. Methods in Ecology and Evolution 2021, 12, 373–377. [CrossRef]
- Hribar, L.J. Expanding Basic Entomological Knowledge by Using Mosquito Surveillance Bycatch. esaa 2020, 113, 439–446. [CrossRef]
- Grootaert, P.; Loziaková Peňazziová, K.; Csank, T.; Pačanská, Z.; Pivka, S.; Oboňa, J. Bycatch with Benefits: CO₂ Traps Reveal New Hybotidae for Slovakia. Biodiversity & Environment 2025, 17, 15–18.
- Mullin, L.J. The Effects of Infusion Water Type and Fermaentation on Mosquito and Non-Target Organism Collected in the CDC’s Autocidal Gravid Ovitrap. J FL Mosq Control Assoc 2021, 67, 23–27. [CrossRef]
- Barringer, L.E.; Bartlett, C.R. Pennsylvania Planthoppers (Hemiptera: Auchenorrhyncha: Fulgoroidea): Relative Abundance and Incidental Catch Using Novel Trapping Methods. Insecta Mundi 2018.
- Urfer, K.; König, N.; Kranz-Baltensperger, Y.; Klopfstein, S.; Niehuis, O.; Von Virag, A.; Sann, M. Reclaiming the Unwanted: Mining Scientific Gold from Multi-Trap Bycatch. AlpEnt 2025, 9, 91–102. [CrossRef]
- Spears, L.R.; Christman, M.E.; Koch, J.B.U.; Looney, C.; Ramirez, R.A. A Review of Bee Captures in Pest Monitoring Traps and Future Directions for Research and Collaboration. Journal of Integrated Pest Management 2021, 12, 49. [CrossRef]
- Munhoz, E.M.; Pereira, T.B.C.; Silva, E.M.S.; Bedendo, I.P. A Subgroup 16SrIII-J Phytoplasma Associated with Aegiphila Verticillata, a Typical Wild Plant of Brazilian Savanna Region, Exhibiting Witches’ Broom Symptoms. Trop. plant pathol. 2016, 41, 52–56. [CrossRef]
- Jomantiene, R.; Postman, J.D.; Montano, H.G.; Maas, J.L.; Davis, R.E.; Johnson, K.B. First Report of Clover Yellow Edge Phytoplasma in Corylus (Hazelnut). Plant Disease 2000, 84, 102–102. [CrossRef]
- Rios, D.; Ueckert, J.; Ong, K.; Barillas, J.R.; Costanzo, S. First Report of a ‘ Candidatus Phytoplasma Sacchari’-Related Strain Associated with Yellowing and Decline of Silver Bluestem in Texas, U.S.A. Plant Disease 2024, 108, 2216. [CrossRef]
- Beirne, B.P. The Nearctic Species Of. The Canadian Entomologist 1952, 84, 208–232. [CrossRef]
- Wallner, A. Evaluating North American Tallgrass Prairie Quality Using the Auchenorrhyncha Quality Index. PhD dissertation, University of Illinois at Urbana-Champaign: Urbana, 2010.
- Kopp, D.D.; Yonke, T.R. The Treehoppers of Missouri: Part 2. Subfamily Smiliinae; Tribes Acutalini, Ceresini, and Polyglyptini (Homoptera: Membracidae). Journal of the Kansas Entomological Society 1973, 233–276.
- Beamer, R.H. A Review of the Genus Osbornellus in the United States and Canada (Homoptera-Cicadellidae). Journal of the Kansas Entomological Society 1937, 10, 89–112.
- DeLong, D.M. Twelve New Species of Osbornellus (Homoptera-Cicadellidae) from Mexico, Guatemala, and Panama. Bulletin of the Brooklyn Entomological Society 1942, 92–101.
- Blondet, L.; Mouanda Sounda, A.; Fritz, M.; Filloux, D.; Yvon, M.; Blanc, S.; Varsani, A.; Niama, R.F.; Leroy, E.M.; Roumagnac, P. Sentinels Tracking Viruses in Various Ecosystems: Towards a One Health Approach. PLoS Pathog 2025, 21, e1013141. [CrossRef]
- Weintraub, P.G.; Trivellone, V.; Krüger, K. The Biology and Ecology of Leafhopper Transmission of Phytoplasmas. In Phytoplasmas: Plant Pathogenic Bacteria - II: Transmission and Management of Phytoplasma - Associated Diseases; Bertaccini, A., Weintraub, P.G., Rao, G.P., Mori, N., Eds.; Springer: Singapore, 2019; pp. 27–51 ISBN 9789811328329.
- Trivellone, V.; Ripamonti, M.; Angelini, E.; Filippin, L.; Rossi, M.; Marzachí, C.; Galetto, L. Evidence Suggesting Interactions between Immunodominant Membrane Protein Imp of Flavescence Dorée Phytoplasma and Protein Extracts from Distantly Related Insect Species. J. Appl. Microbiol. 2019, 127, 1801–1813. [CrossRef]
- Chiykowski, L.N.; Chapman, R.K. Migration of the Six-Spotted Leafhopper in Central North America. Wisconsin Agricultural Experiment Station Research Bulletin 1965, 261, 21–45.
- Clements, J.; Garcia, M.; Bradford, B.; Crubaugh, L.; Piper, S.; Duerr, E.; Zwolinska, A.; Hogenhout, S.; Groves, R.L. Genetic Variation Among Geographically Disparate Isolates of Aster Yellows Phytoplasma in the Contiguous United States. J Econ Entomol 2020. [CrossRef]
- Frost, K.E.; Esker, P.D.; Van Haren, R.; Kotolski, L.; Groves, R.L. Seasonal Patterns of Aster Leafhopper (Hemiptera: Cicadellidae) Abundance and Aster Yellows Phytoplasma Infectivity in Wisconsin Carrot Fields. Environ Entomol 2013, 42, 491–502. [CrossRef]
- Fontes, E.M.G.; Habeck, D.H.; Slansky Jr, F. Phytophagous Insects Associated with Goldenrods (Solidago Spp.) in Gainesville, Florida. Florida Entomologist 1994, 209–221.
- Rizza, S.; Antonella, P.; Vera, D.; Ernesto, R.; Cristina, M.; Tessitori, M. Transmission of ‘Candidatus Phytoplasma Asteris’ (16SrI) by Osbornellus Horvathi (Matsumura 1908) Co-Infected with “Ca. Phytoplasma Phoenicium” (16SrIX). Phytoparasitica 2016, 44, 491–500. [CrossRef]
- Jensen, D.D. Transmission of Peach Yellow Leaf Roll Virus by Fieberiella Florii (Stål) and a New Vector, Osbornellus Borealis DeL. & M.1. J Econ Entomol 1957, 50, 668–672. [CrossRef]
- Ghauri, M.S.K. The Identity of a Suspected Vector of Coconut Lethal Yellowing Disease in Jamaica and Notes on Caribovia Intensa (Walker) (Homoptera: Cicadelloidea). Bulletin of Entomological Research 1980, 70, 411–415. [CrossRef]
- Trivellone, V.; Cao, Y.; Dietrich, C.H. Comparison of Traditional and Next-Generation Approaches for Uncovering Phytoplasma Diversity, with Discovery of New Groups, Subgroups and Potential Vectors. Biology 2022, 11, 977. [CrossRef]
- Brcak, J. Leafhopper and Planthopper Vectors of Plant Disease Agents in Central and Southern Europe. In Leafhopper Vectors and Plant Disease Agents; Academic Press: London, 1979; pp. 97–146.
- Fránová, J.; Špak, J.; Šimková, M. First Report of a 16SrIII-B Subgroup Phytoplasma Associated with Leaf Reddening, Virescence and Phyllody of Purple Coneflower. Eur J Plant Pathol 2013, 136, 7–12. [CrossRef]
- Fischnaller, S.; Parth, M.; Messner, M.; Stocker, R.; Kerschbamer, C.; Janik, K. Surveying Potential Vectors of Apple Proliferation Phytoplasma: Faunistic Analysis and Infection Status of Selected Auchenorrhyncha Species. Insects 2020, 12, 12. [CrossRef]
- Duduk, B.; Peri, P.; Mar, D.; Drobnjakovi, T.; Picciau, L.; Alma, A.; Bertaccini, A. Phytoplasmas in Carrots: Disease and Potential Vectors in Serbia. Bulletin of Insectology 2008, 61, 327–331.
- Quaglino, F.; Sanna, F.; Moussa, A.; Faccincani, M.; Passera, A.; Casati, P.; Bianco, P.A.; Mori, N. Identification and Ecology of Alternative Insect Vectors of ‘Candidatus Phytoplasma Solani’ to Grapevine. Sci Rep 2019, 9, 19522. [CrossRef]
- Molnar, C.; Shires, M.K.; Wright, A.A.; Hoskins, M.C.; Cowell, S.J.; Nikolaeva, E.V.; Knier, R.; Nouri, M.T.; Black, B.; Harper, S.J. Putting ‘X’ into Context: The Diversity of ‘ Candidatus Phytoplasma Pruni’ Strains Associated with the Induction of X-Disease. Plant Disease 2024, 108, 2677–2687. [CrossRef]
- Harper, S.J.; Northfield, T.D.; Nottingham, L.B.; DuPont, S.T.; Thompson, A.A.; Sallato, B.V.; Serban, C.F.; Shires, M.K.; Wright, A.A.; Catron, K.A.; et al. Recovery Plan for X-Disease in Stone Fruit Caused by ‘ Candidatus Phytoplasma Pruni.’ Plant Health Progress 2023, 24, 258–295. [CrossRef]
- Cooper, W.R.; Marshall, A.T.; Lewis, T.M.; Cook, R.; Horton, D.R.; Pitt, W.J.; Corral, C.R.; Jocson, D.M.I.; Gardner, J.D.; Zack, R.S.; et al. Biosystematic Classification of Colladonus Ball Vectors of “Candidatus Phytoplasma Pruni” in the Pacific Northwest (Hemiptera: Cicadellidae: Deltocephalinae). went 2025, 127, 141–161. [CrossRef]
- Trivellone, V.; Wei, W.; Filippin, L.; Dietrich, C.H. Screening Potential Insect Vectors in a Museum Biorepository Reveals Undiscovered Diversity of Plant Pathogens in Natural Areas. Ecol. Evol. 2021, 11, 6493–6503. [CrossRef]
- Duran-Ahumada, S.; Petersen, V.; Futo, M.; Zamy, M.; Pereira, T.; Burini, B.C. The Toolbox for Mosquito Vector Research. Parasites Vectors 2025, 18, 389. [CrossRef]
- Kim, K.; Huang, Q.; Lei, C. Advances in Insect Phototaxis and Application to Pest Management: A Review. Pest Management Science 2019, 75, 3135–3143. [CrossRef]
- Li, C.-X.; Smith, M.L.; Fulcher, A.; Kaufman, P.E.; Zhao, T.-Y.; Xue, R.-D. Field Evaluation of Three New Mosquito Light Traps Against Two Standard Light Traps to Collect Mosquitoes (Diptera: Culicidae) and Non-Target Insects in Northeast Florida. Florida Entomologist 2015, 98, 114–117. [CrossRef]
- Acevedo, N. COMPARISON OF THE CDC LIGHT TRAP AND THE DYNATRAP® DT2000 FOR COLLECTION OF MOSQUITOES IN SEMI-FIELD AND FIELD SETTINGS. J FL Mosq Control Assoc 2021, 67, 69–72. [CrossRef]
Figure 1.
Tethahedrom representation of our proposed vision of One Health to cope with emerging infectious diseases.
Figure 1.
Tethahedrom representation of our proposed vision of One Health to cope with emerging infectious diseases.
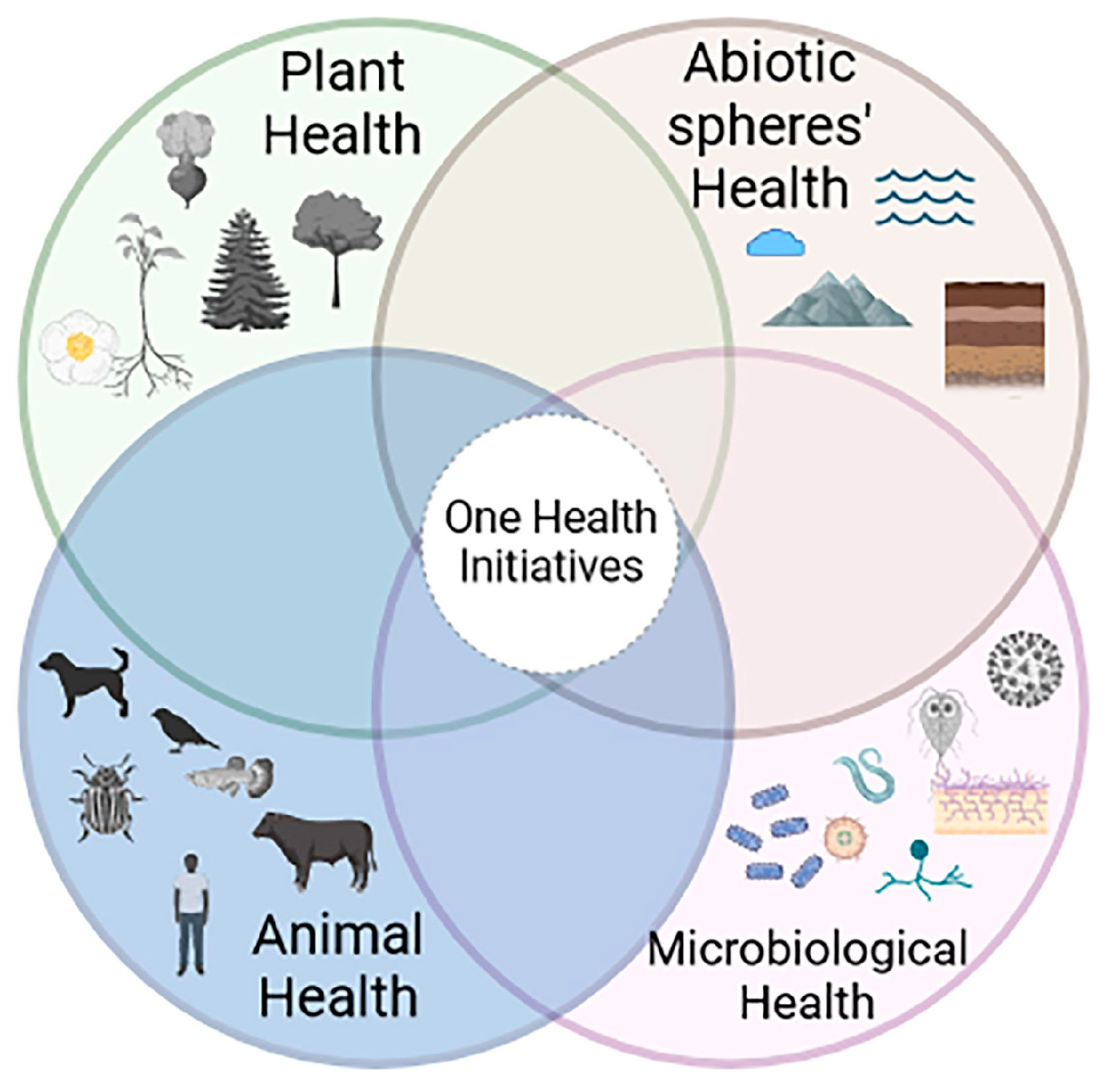
Figure 2.
Distribution of the sites surveyed during the biodiversity surveys in 2021 (green circle) and the public health surveillance surveys in 2022 and 2024 (purple quadrat) in Illinois, USA. The points in red represent the positive specimens, name of the species are plotted. Map created using QGIS 3.10, the inset at the bottom left shows the location of Illinois within the contiguous United States. Major rivers and lakes are shown in light blue.
Figure 2.
Distribution of the sites surveyed during the biodiversity surveys in 2021 (green circle) and the public health surveillance surveys in 2022 and 2024 (purple quadrat) in Illinois, USA. The points in red represent the positive specimens, name of the species are plotted. Map created using QGIS 3.10, the inset at the bottom left shows the location of Illinois within the contiguous United States. Major rivers and lakes are shown in light blue.
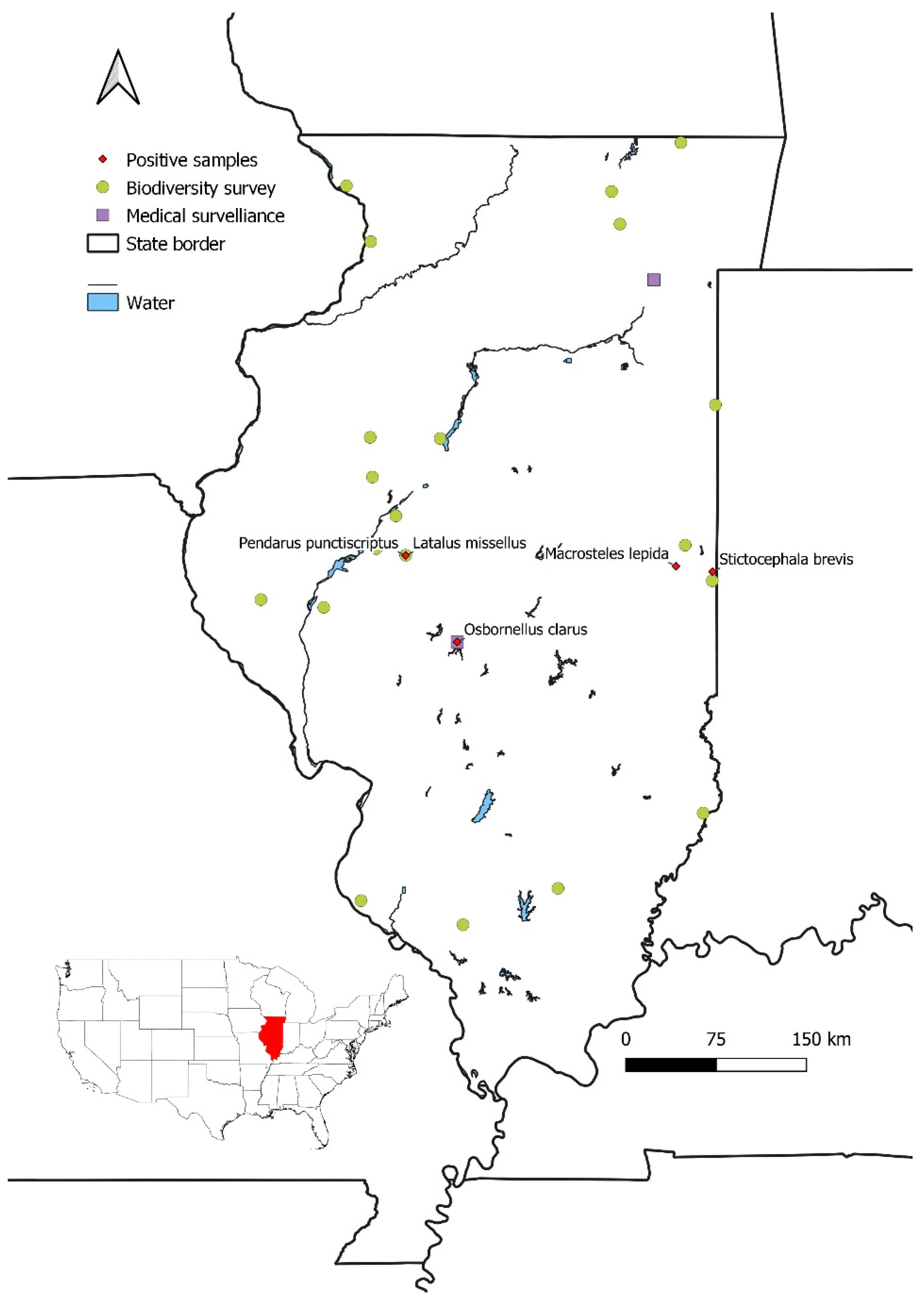
Figure 3.
Dorsal views of the 5 species of leafhoppers that tested positive for phytoplasmas. A, Macrosteles lepidus (Van Duzee) (Illinois Natural History Survey Insect collection code, 1086365); B, Osbornellus clarus Beamer (1083960); C, Latalus missellus (Ball, 1899) (1083966); D, Pendarus punctiscriptus (Van Duzee, 1892) (1083969); E, Stictocephala brevis (Walker, 1851) (1089637). Scale bar 1.0 mm.
Figure 3.
Dorsal views of the 5 species of leafhoppers that tested positive for phytoplasmas. A, Macrosteles lepidus (Van Duzee) (Illinois Natural History Survey Insect collection code, 1086365); B, Osbornellus clarus Beamer (1083960); C, Latalus missellus (Ball, 1899) (1083966); D, Pendarus punctiscriptus (Van Duzee, 1892) (1083969); E, Stictocephala brevis (Walker, 1851) (1089637). Scale bar 1.0 mm.
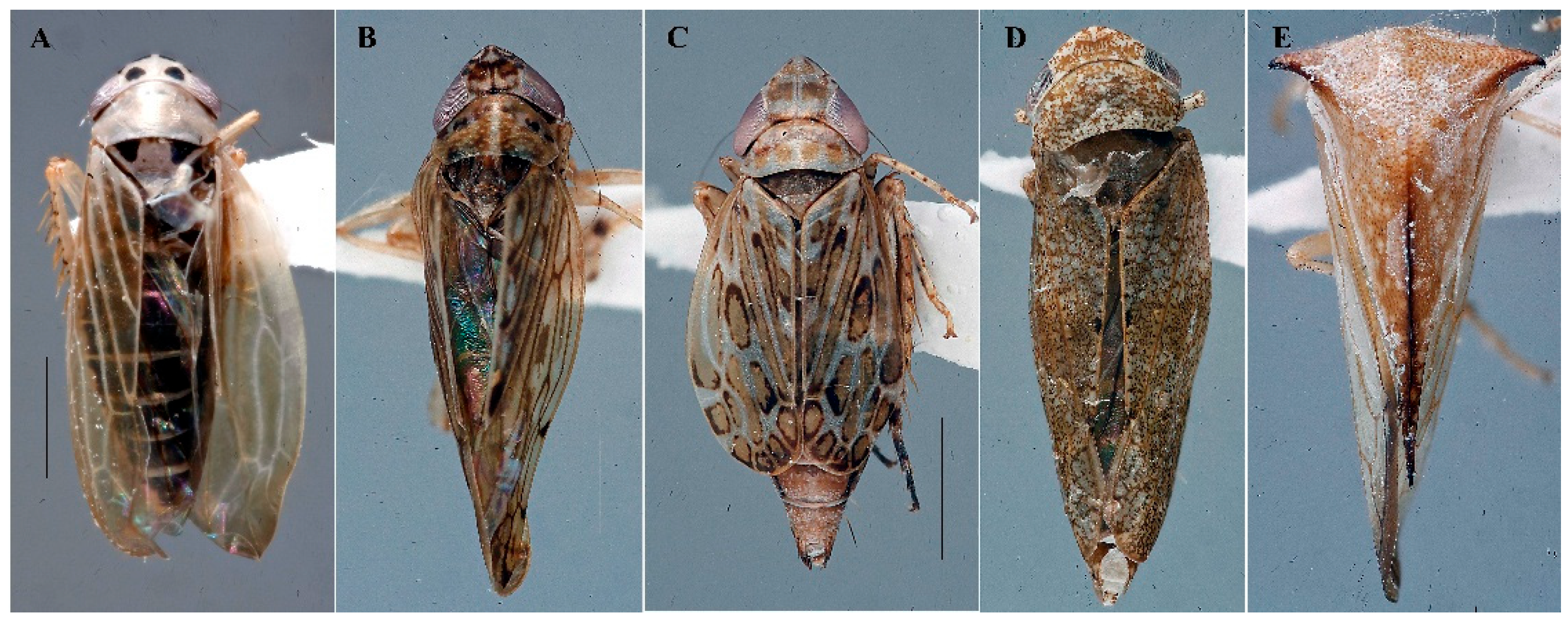
Figure 4.
Phylogenetic tree inferred by Maximum Likelihood (ML) based on 16Sr rRNA gene. Bootstrap values >70% (1,000 replicates) shown. Samples from this study are indicated with an orange square. Acholeplasma palmae was used as outgroup (GenBank accession number NR_029152.1).
Figure 4.
Phylogenetic tree inferred by Maximum Likelihood (ML) based on 16Sr rRNA gene. Bootstrap values >70% (1,000 replicates) shown. Samples from this study are indicated with an orange square. Acholeplasma palmae was used as outgroup (GenBank accession number NR_029152.1).
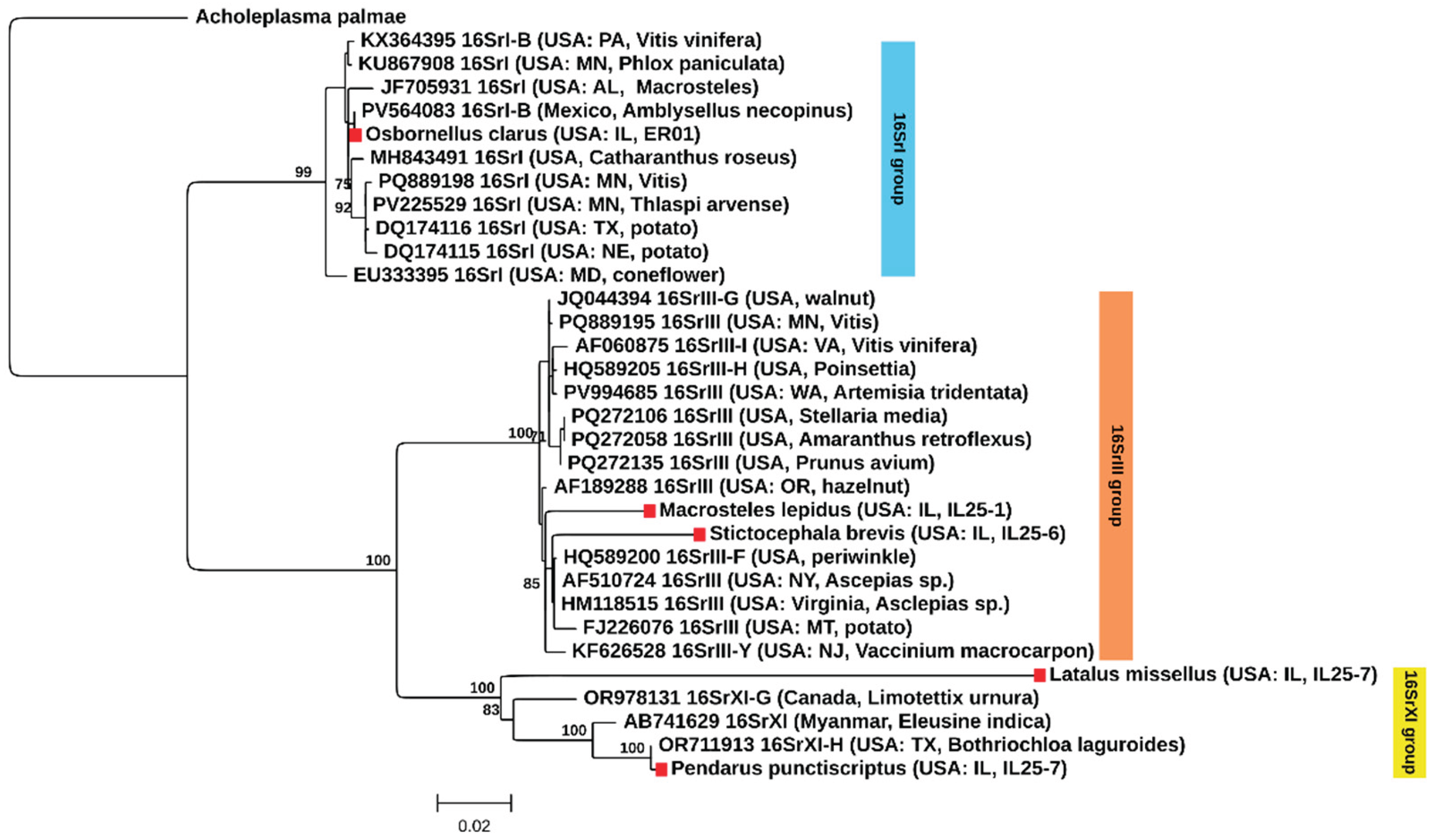

Table 1.
List of samples collected from 2020 to 2025 in XX independent surveys in preserved areas in Illinois.
Table 1.
List of samples collected from 2020 to 2025 in XX independent surveys in preserved areas in Illinois.
| Species | Coordinates | Date | INHS Insect coll. code | Associated Phy. | qPCR Ct value | NCBI accession |
| Macrosteles lepidus1 | 87°50'11"W 40°05'27"N | 06/12/2025 | 1086365 | 16SrIII | 21.97 | PX789936 |
| Osbornellus clarus2 | 89°28'08.9"W 39°39'26.9"N | 09/02/2022 | 1083960 | 16SrI-B | 24.02 | PX789937 |
| Latalus missellus1 | 89°51'12.5"W 40°09'09.6"N |
08/20/2025 | 1083966 | 16SrXI-H | 25.85 | PX789938 |
| Pendarus punctiscriptus1 | 89°51'12.5"W 40°09'09.6"N |
08/20/2025 | 1083969 | 16SrXI-H | 23.21 | PX789939 |
| Stictocephala brevis1 | 87°33'50.9"W 40°03'35.6"N |
07/29/2025 | 1089637 | 16SrIII | 19.08 | PX789940 |
1 INHS Auchenorrhyncha biodiversity survey 2 INHS mosquitoes state monitoring.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



