
Submitted:
02 February 2026
Posted:
03 February 2026
You are already at the latest version
Abstract
Multidrug resistance (MDR) is a major global health challenge, necessitating alternative or adjuvant strategies such as lactoferrin (Lf) and probiotics. Opportunistic bacteria are key drivers of healthcare-associated infections (HAIs) due to their ability to form biofilms and cause persistent infections. Lf, a glycoprotein present in mammalian milk and secretions, exhibits antimicrobial properties as part of innate immunity. Similarly, probiotics—primarily lactic acid bacteria (LAB)—influence pathogen growth and adhesion. This study evaluated the antimicrobial and anti-biofilm effects of purified bovine Lf (bLf) versus commercial Lf, alone and combined with LAB-derived supernatants, against several opportunistic strains. Using qualitative and quantitative methods, we assessed the antimicrobial, anti-adherence, and immunomodulatory activities of bLf (Sigma-Aldrich), commercial Lf (Jarrow Formulas), and LAB supernatants. High-purity bLf demonstrated superior antimicrobial and antibiofilm effects compared to supplement-derived Lf. Furthermore, commercial Lf, combined with LAB metabolites, effectively inhibited biofilm formation, reduced inflammation, and prevented bacterial adhesion to eukaryotic cells. These findings support the potential use of lactoferrin, alone or in synergy with probiotics, as adjuvants or prophylactic agents, particularly for immunocompromised patients, to limit bacterial adherence and prevent chronic infections.
Keywords:
lactoferrin
; antimicrobial activity
; anti-biofilm activity
; probiotics
; opportunistic infections
; immunomodulatory activity
; multi-drug resistance (MDR)
1. Introduction
Antimicrobial Resistance (AMR), including Multi-Drug Resistance (MDR), counts among the top 10 threats to public health and has been described as a “silent pandemic” [1,2,3,4]. MDR has been defined as the decreased sensitivity to ≥ 1 antibiotic agent from ≥ 3 antibiotic classes [5]. More than 10 years ago, The World Health Organization (WHO) warned about entering a period where no antibiotics will longer be effective against multi-resistant microorganisms, known as the “post-antibiotic” era [1,6]. In less than 30 years from now on, it is presumed that more deaths will be due to AMR alone than diabetes and cancer together [1,7]. The administration of broad-spectrum antibiotics, either for empirical or targeted treatment, has led to the dissemination of multiple MDR bacterial strains [3]. The most important pathogens causing MDR infections are: (1) Gram-negative: Acinetobacter baumannii, Pseudomonas aeruginosa, Klebsiella pneumoniae, Escherichia coli; and (2) Gram-positive: Streptococcus pneumoniae, Staphylococcus aureus, Enterococcus faecium, from which the latter two have developed extensive resistance to antibiotics, including methicillin and vancomycin [1,2,4,7,8].
Antibiotic resistance, driven by chromosomal or plasmid-mediated resistance genes, is divided into: (1) natural or constitutive resistance, stemming from the bacterium’s intrinsic properties that contribute to its resistance to the effects of antibiotics; and (2) acquired resistance, caused by multiple mechanisms, such as: decreased membrane permeability and compositional alterations of cytoplasm membrane, resulting in reduced drug quantities entering the cell; activation of efflux pumps; synthesis of enzymes that decompose or block the activity of antibiotics; activation of secondary metabolic pathways; structural modifications of the antibiotic target [2,8].
Though, as a result of the stress caused by adverse conditions including various antimicrobials, bacterial cells have developed a secondary lifestyle of isolated cells, an adaptive state, in large communities known as biofilms [9,10]. The term “biofilm” was firstly defined by Donlan et al. as a sessile, multilayered microbial population consisting of cells permanently attached to one another and to biotic or abiotic substrates, that are incorporated in a self-produced matrix of Extracellular Polymeric Substances (EPS) and exhibit different, specific gene expression profiles and growth rate phenotypes [10,11,12]. Bacterial cells embedded in a biofilm are more resistant than their planktonic counterparts, attributed to “super resistant cells”, tolerant due to their metabolic latency or dormant state that support the survival of biofilms during long-term antibiotic treatment [12,13]. Implementing alternatives or adjuvants to antibiotics, such as probiotics and Lf, stems from the biofilm-included cells possessing a notable, non-inherited resistance or tolerance to antibiotics and immune effectors, leading to chronic and persistent infections [10,12,13,14].
Lactoferrin (Lf) is an iron-chelating 80 kDa glycoprotein purified from bovine milk for the first time 85 years ago [10,15]. It is present in the whey protein fraction of milk, alongside α-lactoalbumin, β-lactoglobulin, lactoperoxidase, serum albumin, and immunoglobulins [16]. Besides mammal milk, Lf has been identified in saliva, semen, tears, bile, intestinal, vaginal and bronchial secretions, where it takes part in the local antipathogenic defense mechanisms [17,18,19]. Bovine Lf (bLf) and human Lf (hLf) are similar in structure, given that the polypeptide chain of bLf is 689 aminoacids and 691 aminoacids long for hLf, allowing for the use of bLf in human therapeutics [20,21]. Past research has underlined that Lf is a multivalent protein, given its antibacterial, antifungal, antiviral, antiparasitic, antioxidant, anti-inflammatory, immune-promoting and antitumoral properties [10,18,22,23,24]. The antimicrobial action of Lf is due to the bacteriostatic and bactericidal mechanisms described against several Gram-positive and Gram-negative pathogens [21,23,25]. The bactericidal action is linked to the N-terminal region of Lf (lactoferricin, Lfcin), as well as the positive charge of the molecule, facilitating the interaction with anionic molecules on bacterial surface, such as lipopolysaccharides (LPS) of Gram-negative bacteria and lipoteichoic acids of Gram-positive bacteria, resulting in membrane permeabilization [21,23,24,25,26]. The bacteriostatic aspect of Lf is arising from its iron-sequestering property, although the microbial metabolism and growth inhibition is transient and can be restored by exogenous iron supplementation [21,25,26] (Figure 1). The interaction between Lf and bacterial membrane porins might be correlated to the antibacterial properties [24]. In the context of the emerging incidence of infections with MDR strains, Lf represents a promising candidate for the management of HAI, enabling its use as an adjuvant to antibiotic therapy [27].
Probiotics, including LAB, are represented by Gram-positive, non-spore-forming, microaerophilic bacteria colonizing the normal gut microbiota that benefit the host [28,29,30]. The immunostimulatory and immunomodulatory effect, anti-infectious activity, mutagenesis-inhibitory action, diminishing blood ammonia and serum cholesterol concentrations, decreasing gut inflammation and stabilization of mucosal barriers can be counted among these benefits [29,31]. The idea about these beneficial microorganisms was underlined by E. Metchnikoff (1907), but the term “probiotic” (“for life”) was coined later in 1953, describing it as bioactive compounds crucial for the proper development of biological systems [29,32]. Probiotics comprise a heterogenous class of bacteria also present in fermented products (cheese, meats, yogurt and other dairy products). The most commonly used probiotic strains belong to the Lactobacillus, Bifidobacterium, Enterococcus, Lactococcus, Pediococcus and Streptococcus genera, as well as yeast strains of Saccharomyces boulardii and, most notably, a strain of Escherichia coli – E. coli Nissle 1917 [30,33,34,35]. Out of these, the most predominant probiotic genera in the human gut and the first to colonize the newborn intestines are Lactobacillus and Bifidobacterium [33,34,36]. The Lactobacillus genus is a member of the Lactobacillaceae family within the Firmicutes phyla and is the most representative bacteria of the LAB group [29,33,34]. Species of Lactobacillus are able to transform hexose carbohydrates into lactic acid, therefore providing unfavorable acidic conditions, preventing the growth and adhesion of pathogenic bacteria [33]. Lactobacilli are considerable antimicrobial agents due to the synthesis and release of benzoic acid and H2O2 which inhibit the colonization of catalase-negative and low pH-sensitive pathogens. Further antibacterial mechanisms of LAB include the antagonism for binding sites and nutrients, interference with the quorum-sensing (QS) system, and secretion of bacteriocins [28,34,37]. These molecules are antibacterial agents with low molecular weight and broad-spectrum activity against multiple enteric microorganisms [34]. The previously described strategies are effective in eradicating infections caused by Clostridium spp. (C. difficile), Salmonella spp., Helicobacter pylori and E. coli [29,31]. Besides, probiotics in general have notable effects on modulating the host’s immune defense by promoting release of anti-inflammatory cytokines and blocking synthesis of pro-inflammatory cytokines [38]. Taking these benefits into consideration, LAB have attracted the interest of researchers for their use as alternatives or adjuvants to antibiotics [34]. In recent times, antibiotic therapy has been administered simultaneously with probiotics to prevent gut dysbiosis caused by antibiotics, even though probiotics alone proved an efficient decolonizing activity against pathogenic bacteria [39].
In recent times, bacterial products as metabolites, non-viable cell fragments and extracellular vesicles (EVs) have been included in the spectrum of probiotics’ action, being classified as postbiotics, which are beneficial to the human organism [40,41,42,43]. Postbiotics have been defined in 2021 by the International Scientific Association of Probiotics and Prebiotics as preparations of “inanimate microorganisms and/or their components that confers a health benefit on the host” [44]. EVs are a group of spheric lipid particles, with diameters within the nanometer scale (20-400 nm) and released by both eukaryotic and prokaryotic cells, including probiotics [41,42,43,45,46]. Initially, it was considered that EVs were exclusively produced by Gram-negative bacteria, given their structural similarity to the outer membrane; though, these structures were observed in probiotic Gram-positive bacteria such as Lb. plantarum, Lb. rhamnosus and Lb. paracasei [40,47,48,49,50]. LAB-derived EVs are transporters of bioactive macromolecules (lipids, nucleic acids, proteins, enzymes, polysaccharides, metabolites), therefore supporting the host’s health by their antibacterial and immunomodulatory activities [40,42,43,47,48,51,52]. In this context, a nanoplatfom formulated with Lb. paracasei EVs was demonstrated to be a novel approach to antimicrobial therapy against Staphylococcus aureus [47]. In other studies, EVs from Lb. plantarum had a notable action against Shewanella putrefaciens, S. aureus and Propionibacterium acnes [40,48]. Lb. rhamnosus, E. coli Nissle 1917 and Lb. plantarum-derived EVs promoted the immunomodulatory release of anti-inflammatory cytokines (IL-1β, IL-6, TNF-α) and inhibition of anti-inflammatory cytokines (IL-10) synthesis in microglial cells and immune effectors such as dendritic cells and macrophages [49,53,54,55]. In addition, EVs from E. coli Nissle 1917 enhanced the bactericidal action of macrophages when infected with E. coli, S. aureus and S. typhimurium [53].
This study aimed to evaluate the inhibitory effect of bLf, in opposition to commercial Lf, upon microbial growth and adhesion of several opportunistic strains. Furthermore, the combinations between commercial Lf and supernatants derived from LAB cultures were tested for the same activities. Finally, the immunomodulatory actions of these combinations against opportunistic infections were measured.
2. Materials and Methods
2.1. Bacterial Strains
The Gram-positive and Gram-negative opportunistic bacterial strains used in the experiments were purchased from American Type Culture Collection (ATCC, Manassas, Virginia, USA) (Staphylococcus aureus ATCC 25923, Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853), as well as isolated from newborn feces (Enterococcus faecium).
Prior to the evaluation, fresh cultures were obtained by transferring and incubating the strains on nutritive agar in Petri dishes at 37oC, for 24h in a LBX Instruments incubator (Labbox, Barcelona, Spain).
2.2. Tested Compounds
The tests were conducted using the following agents and procedures (Table 1):
2.3. Antimicrobial Qualitative Test (Adapted Antibiogram Method)
Prior to the qualitative test, serial binary dilutions were performed to obtain six different concentrations of bLf (1000, 500, 250, 125, 62.5, and 31.25 µg/mL). A standardized inoculum, represented by a 0.5 McFarland bacterial suspension in 0.9% NaCl sterile solution, equivalent to 1.5 x 108 CFU (Colony Forming Units)/mL, was prepared for each strain. The inocula were individually seeded on Petri dishes containing solid medium (nutritive agar), followed by placing 10 µL of each bLf concentration using an Eppendorf pipette (Eppendorf, Hamburg, Germany). After the diffusion of the drops, the Petri dishes were incubated at 37oC, for 18-20h, followed by the measurement of the growth inhibition zone diameters in millimeters (mm) [56]. For each bLf dilution, the experiment was performed in duplicate.
2.4. The Minimal Inhibitory Concentration (MIC) Method
The microdilution method was used to establish the Minimal Inhibitory Concentration (MIC) of the tested compounds. Therefore, the method was carried out using 96-well plates, in sterile nutrient broth liquid media. Serial binary dilutions of each compound were performed in a final volume of 200 µL, producing the following concentrations: bLf (2000, 1000, 500, 250, 125, 62.5, 31.25, 15.63, 7.81 and 3.91 µg/mL); commercial Lf (5000, 2500, 1250, 625, 313.5, 156.25, 78.13, 39.06, 19.53 and 9.77 µg/mL), alone and combined with LAB supernatants (50%, 25%, 12.5%, 6.25%, 3.13%, 1.56%, 0.78%, 0.39%, 0.2%, and 0.1%), and gentamicin (100, 50, 25, 12.5, 6.25, 3.13, 1.56, 0.78, 0.39, and 0.2 µg/mL). Then, a 1/10 dilution of the 0.5 McFarland suspension was executed for each pathogenic strain, resulting in a density of 1.5 x 107 CFU/mL. 10 µL of each diluted bacterial suspensions were then transferred to each well, succeeded by incubating the plates at 37oC, for 24h. Thereafter the MIC values were determined by macroscopical analysis [12,56,57].
2.5. The Minimal Biofilm Eradication Concentration (MBEC) Method
To evaluate the compounds’ anti-biofilm action, the crystal violet microdilution method was used. In short, each compound (bLf, commercial Lf alone and mixed with LAB supernatants, and gentamicin, respectively) was diluted in 96-well plates, in sterile liquid broth, in a final volume of 200 µL. After that, each well was seeded with 10 µL of a suspension with a density of 1.5 x 107 CFU/mL for each strain, followed by incubation of the plates at 37oC, for 24h. Thereafter, the liquid broth was removed and the 96-well plates were washed twice with 150 µL of PBS to eliminate non-adherent bacterial cells. The remaining adherent cells on the plastic substrate were fixed with 150 µL of cold methanol for 5 min. Subsequently to the methanol removal, the bacterial cells were stained with 120 µL of a 1% crystal violet solution for 20 min and the remaining dye was washed off with tap water. The aspect and incorporation of bacterial cells in the biofilms were examined by an inverted optical microscope (200x) (Optika Microscopes, Ponteranica, Italy). The stained biofilms were resuspended in 120 µL of a 33% acetic acid solution and the optical density (OD) of the suspensions was determined spectrophotometrically at a 490 nm using the Synergy HTX ELISA reader associated with the Gen5 2.06 software (BioTek, Winooski, Vermont, USA) [12,56,57].
2.6. Induction of Cytokines Secretion
The colorectal adenocarcinoma HCT-8 cell line (ATCC, Manassas, Virginia, USA) was used to assess the production of cytokines. The cells were maintained in DMEM (Dulbecco’s Modified Eagle Medium):F12 (Thermo Fisher Scientific Inc., Waltham, Massachusetts, USA) supplemented with 10% FBS (Fetal Bovine Serum), in 24-well plates in a volume of 1 mL per well, at 37 °C in a humidified atmosphere with 5% CO2, according to the provider’s instructions [58].
The immunomodulation protocol was adapted by Lepanto et al., 2019 [59], as it follows. Firstly, the HCT-8 cells in every well were pre-treated with a volume of 200 µL commercial Lf and/or LAB supernatants, succeeded by incubating the 24-well plates at 37 °C, 5% CO2, for 2.5 h. Secondly, after the pre-treatment, the cells were stimulated by adding 400 µL of bacterial suspensions of S. aureus ATCC 25923, E. coli ATCC 25922, P. aeruginosa ATCC 27853, and E. faecium at a density of 1.5 x 107 CFU/mL in PBS, then followed by plates’ incubation at 37 °C, 5% CO2, for another 2.5 h. After the immune stimulation, cell supernatants were transferred into Eppendorf 1.5 mL tubes and kept at -20oC [56,59].
2.7. Enzyme-Linked Immunosorbent Assay
The release of anti-inflammatory (IL-10) and pro-inflammatory (IL-6, IL-8, TNF-α) cytokines was quantified using EIAab Science ELISA kits (EIAab Science INC, Wuhan, China), as stated by the manufacturer’s instructions. The assessments were conducted on the supernatants obtained in the previous step, which were pre-treated with commercial Lf and/or LAB supernatants and then stimulated with microbial suspensions (See 2.4.1). The cytokine concentrations were determined by spectrophotometer readings at 450 nm using the Synergy HTX ELISA reader and the Gen5 2.06 software (BioTek, Winooski, Vermont) [56].
2.8. Bacterial Adhesion to Cell Substrate – Adhesion Patterns and Adhesion Index
In order to analyze the adhesion patterns and determine the Adhesion Index (AI%), the HCT-8 cells were maintained in DMEM:F12 supplemented with 10% FBS, in 6-well plates in a volume of 2 mL per well, at 37 °C in a humidified atmosphere with 5% CO2. To every well, 200 µL of commercial Lf and/or LAB supernatants and 400 µL of bacterial suspensions of S. aureus ATCC 25923, E. coli ATCC 25922 and P. aeruginosa ATCC 27853 at a density of 1.5 x 107 CFU/mL in PBS were added. Subsequently, the plates were incubated for 2 h at 37 °C, 5% CO2, then washed twice with PBS, stained for 20 min with a Giemsa solution (Merck, Darmstadt, Germany), then washed with tap water and examined using a Zeiss Primo Star microscope (Zeiss, Oberkochen, Germany). The microscopic examination was used to determine the adhesion patterns and AI%, indicating the percentage of bacteria-adhering HCT-8 cells among total counted HCT-8 cells [56,60,61,62,63].
2.9. Data Analysis
Interleukin concentrations were calculated from the measured absorbance values based on linear regression formulas according to the kit’s manufacturer (EIAab Science INC, Wuhan, China). Detailed investigations were performed by focusing on each tested variant (SNx, SNx + LF or pathogens). Data was pooled for Gram-negative (E. coli ATCC 25922 and P. aeruginosa ATCC 27853) or Gram-positive (S. aureus ATCC 25923 and E. faecalis) pathogens in order to observe the overall effect of a bacterial group.
The absolute interleukin concentrations were used to create bar plots and perform statistical testing. Given that the current research is a preliminary study, statistical significance was not observed for these investigations due to small sample sizes (n = 2 for the experimental variants and n = 1 for the control variants), although trends in the data could be observed.
In order to evaluate the effect of each tested variant, the variant of interest was isolated using calculations such as: Δ(Pathogen control – Cells control) for pathogens effect, Δ(LF – Pathogen control) for LF effect, Δ(SNx – Pathogen control) for the effect of each tested SN, and Δ(SNx + LF – Pathogen control) for the effect of each tested SN+LF. This investigation allowed for a precise evaluation of the variant effect, whilst increasing the sample size from the pooled values corresponding to Gram-negative or Gram-positive bacteria.
Graphical representations and statistical testing were performed in GraphPad Prism version 8.0.2 for Windows, GraphPad Software, Boston, Massachusetts, USA, www.graphpad.com. Error bars for all bar plots were fitted using the mean + SEM (standard error of mean) option. Statistical testing consisted of Kruskal-Wallis ANOVA paired with Dunn’s multiple comparisons post-hoc test. Statistical significance threshold was set at α = 0.05 at 95% confidence.
3. Results
3.1. Assessment of Lactoferrin and/or Lactic Acid Bacteria Supernatants’ Effects Against Bacterial Adhesion and Growth
3.1.1. Effects of Bovine Lactoferrin on Bacterial Growth
In the qualitative method, several bLf (Sigma-Aldrich, USA) concentrations (1000, 500, 250, 125, 62,5 and 31,25 µg/mL) corresponding to the following dilution factors: 1, 1/2, 1/4, 1/8, 1/16, 1/32 were used. The results did not reveal a significant antimicrobial effect of either concentration on the S. aureus ATCC 25923, E. coli ATCC 25922, P. aeruginosa ATCC 27853 and E. faecium strains cultured on solid media. Given that the data obtained in the qualitative testing were moderate, further quantitative testing of the antimicrobial and anti-biofilm effects in liquid media was conducted.
3.1.2. Antimicrobial and Anti-Biofilm Activity of High-Purity Bovine Lactoferrin Compared to Commercial Lactoferrin
The quantitative evaluation of the action upon bacterial growth and adhesion underlined a strain-dependent effect and a more prevalent anti-biofilm activity of both lactoferrin variants.
A significant effect against development of monospecific biofilms was recorded in the treatment of Gram-positive strains (S. aureus ATCC 25923 and E. faecium) with bLf (Sigma-Aldrich, USA), in comparison to growth inhibition. Thus, Minimal Biofilm Eradication Concentration (MBEC) (500 µg/mL for S. aureus ATCC 25923 and 250 µg/mL for E. faecium) values were notably lower in contrast to Minimal Inhibitory Concentration (MIC) values (2000 µg/mL for S. aureus ATCC 25923 and 500 µg/mL for E. faecium). The treatments with commercial Lf (Jarrow Formulas, USA) followed a similar trend for both Gram-positive strains, with MBEC values (1250 µg/mL) beneath MIC (5000 µg/mL). The antimicrobial control (gentamicin, Sigma-Aldrich, USA) had predominantly consistent effects: MBEC = 4 µg/mL and MIC = 4 µg/mL for S. aureus ATCC 25923; MBEC = 12.5 µg/mL and MIC = 25 µg/mL for E. faecium. Our results suggest a higher sensitivity of the Gram-positive strains to bLf (Sigma-Aldrich, USA) since it required lower doses in order to reach MBEC and MIC values (Table 2).
On a microscopic scale, bLf and commercial Lf visibly reduced the adhesion to inert substrates for the S. aureus ATCC 25923 and E. faecium strains. Moreover, bLf induced a similar effect to gentamicin on E. faecium (Figure 2).
In contrast to the Gram-positive strains, one of the Gram-negative pathogens (E. coli ATCC 25922) displayed a higher tolerance to both bLf (Sigma-Aldrich, USA) and commercial Lf (Jarrow Formulas, USA), yielding MIC and MBEC values of 5000 µg/mL and 2000 µg/mL respectively. However, for the other Gram-negative strain (P. aeruginosa ATCC 27853), a similar predominant anti-biofilm effect was noticed when treated with both Lf variants: MIC = 2000 µg/mL and MBEC = 62.5 µg/mL for bLf; MIC = 5000 µg/mL and MBEC = 1250 µg/mL for commercial Lf. Gentamicin (Sigma-Aldrich, USA) provided equal MBEC and MIC values for each strain: 4 µg/mL for E. coli ATCC 25922 and 25 µg/mL for P. aeruginosa ATCC 27853 (Table 3).
Microscopically, the elevated MIC and MBEC for E. coli ATCC 25922 could be connected to its diminished ability to develop biofilms. In the case of P. aeruginosa ATCC 27853, gentamicin and bLf had more visible effects on bacterial adhesion (Figure 3).
Consequently, the results highlighted that the treatments with bLf had a greater capacity to modulate both microbial growth and adhesion to inert substrates than Lf from food supplements. Out of the tested pathogens, the Gram-positive strains were more susceptible to both Lf variants. Regarding the Gram-negative strains, the inhibitory effect was dependent on the tested strain.
3.1.3. Influence of Lactoferrin and/or Lactic Acid Bacteria Supernatants on Biofilm Formation
This experiment was performed to evaluate the antimicrobial and anti-biofilm effects of the association between commercial Lf (Jarrow Formulas, USA) and supernatants containing LAB metabolites against S. aureus ATCC 25923, P. aeruginosa ATCC 27853, E. coli ATCC 25922 and E. faecium. The supernatants from LAB strains were referred to in the experiments as it follows: SN (supernatant) 1 = Lb. rhamnosus MF9; SN2 = Lb. brevis 10; SN3 = Lb. plantarum F.
The variants considered in the testing yielded considerable MBEC values especially on the Gram-positive strains (S. aureus ATCC 25923 and E. faecium) and less significant MIC values for both Gram-negative and Gram-positive bacteria.
Accordingly, for the S. aureus ATCC 25923 strain, SN2 determined a MBEC value equal to the MRS control (50%). When combined with LAB supernatants, a decrease in the MBEC value of commercial Lf (0.08 µg/mL) was perceived in contrast to Lf alone (MBEC = 1250 µg/mL) (Table 4).
When microscopically examined, it has been observed that Lb. brevis 10 supernatant (SN2) had a notable anti-biofilm action against S. aureus ATCC 25923 in opposition to the MRS control. Additionally, the mixtures between commercial Lf (Jarrow Formulas, USA) and SN1, SN2, and SN3 determined lower MBEC values than the MRS + Lf control (Figure 4).
In the case of E. faecium, the MBEC values needed to dissociate the biofilms formed on plastic substrates were lower for SN1 and SN2 (0.2%) compared to the MRS media control (0.39%). In addition, in the presence of LAB supernatants, a lower concentration of commercial Lf (Jarrow Formulas, USA) was required to inhibit biofilm formation (MBEC = 0.31 µg/mL when combined with SN1 and SN2 and MBEC = 0.08 µg/mL when associated with SN3) on E. faecium than Lf alone (MBEC = 1250 µg/mL). The commercial Lf – LAB supernatants association yielded lower MBEC values than the Lf + MRS control (Table 5).
The microscopical analysis of biofilms revealed lower adhesion capacities of E. faecium when treated with the individual LAB supernatants compared to the control. In opposition, commercial Lf (Jarrow Formulas, USA), in the supernatant combination, did not determine considerable MBEC values compared to the control (Figure 5).
3.2. Effects of Lactoferrin and/or Lactic Acid Bacteria Supernatants on Immune Modulation and Bacterial Adhesiveness
3.2.1. Modulation of Cytokine Production
The immunomodulatory effect on HCT-8 cells pre-treated with 25 mg/mL commercial Lf and/or LAB supernatants and subsequently infected with E. coli ATCC 25922, P. aeruginosa ATCC 27853, S. aureus ATCC 25923 and E. faecium was evaluated by quantifying the release of pro- and anti-inflammatory cytokines. The results obtained for every strain were compared to four distinct controls: (1) Lf control, including the bacterial cells treated with commercial Lf and the HCT-8 cells; (2) pathogen control, comprising the bacterial and eukaryotic cells; (3) MRS control, represented by the bacterium, MRS media and HCT-8 cells; (4) cells control, containing the untreated HCT-8 cells.
For IL-6 and TNF-α, no significant data has been recorded regardless of pathogenic strain or treatment. Though, the treatments had varying effects on the release of the pro-inflammatory cytokine IL-8 depending on the compound and pathogen that the eukaryotic cells have been challenged with. Therefore, compared to the untreated control (IL-8 level of 824.81 pg/mL), in the immune challenge with both Gram-negative pathogens (E. coli ATCC 25922 and P. aeruginosa ATCC 27853), the association between Lf and the Lb. rhamnosus MF9 supernatant (SN1) significantly reduced these levels to 421.18 pg/mL and 383 pg/mL respectively, potentially exerting an anti-inflammatory effect. Lf induced a lower pro-inflammatory response mediated by IL-8 (687.09 pg/mL) than P. aeruginosa ATCC 27853 alone (887.55 pg/mL). For E. coli ATCC 25922, Lf induced no significant differences compared to the pathogen alone and untreated control (Figure 6, a). Similar to the Gram-negative pathogens, the Lf and Lb. rhamnosus MF9 supernatant (SN1) combination diminished the IL-8 concentration in the infections with S. aureus ATCC 25923 (535.27 pg/mL) and E. faecium (559.82 pg/mL), as opposed to untreated cells. On top of that, for E. faecium, all LAB supernatants alone and the mixture of Lf and the Lb. brevis 10 supernatant (SN2) had a significant impact on IL-8 levels, generating concentrations between 636.63 – 692.55 pg/mL. Individual Lf administration, in the absence of infection, suppressed inflammation (652.55 pg/mL) compared to untreated cells. For S. aureus ATCC 25923 and E. faecium, Lf determined lower levels of IL-8 (599.82 and 657 pg/mL) compared to the pathogens alone (985.76 and 677.55 pg/mL) and cells control (824.82 pg/mL) (Figure 6, b). No statistically significant differences were observed.
The dataset corresponding to the effect of each tested variant on IL-8 levels following expose to Gram-negative pathogens was correlated with a statistically significant Kruskal-Wallis ANOVA (H = 20.49, p = 0.0001, ndataset = 52). Post-hoc analysis in the form of Dunn’s multiple comparisons test revealed statistically significant differences between the effects of: Gram-negative pathogens and SN1 (mean rank difference = 32.33, p = 0.002, nformer = 6, nlatter = 8), Gram-negative pathogens and SN1 + LF (mean rank difference = 42.58, p = 0.0004, nformer = 6, nlatter = 4), as well as SN3 and SN1 + LF (mean rank difference = 32.25, p = 0.014, nformer = 8, nlatter = 4) (Figure 7).
The dataset corresponding to the effect of each tested variant on IL-8 levels following expose to Gram-positive pathogens was correlated with a statistically significant Kruskal-Wallis ANOVA (H = 26.25, p = 0.0005, ndataset = 52). Post-hoc analysis testing revealed statistically significant differences between the effects of: SN2 and SN1 + LF (mean rank difference = 32.88, p = 0.011, nformer = 8, nlatter = 4), SN3 and SN1+LF (mean rank difference = 32.25, p = 0.014, nformer = 8, nlatter = 4), as well as SN1+LF and SN3+LF (mean rank difference = -37.75, p = 0.012, nformer = 4, nlatter = 4) (Figure 8).
In the assessment of anti-inflammatory cytokine IL-10 secretory pattern, the varied effects of the applied treatments had been observed. When compared to untreated HCT-8 cells (0.98 pg/mL), the Lb. brevis 10 supernatant decreased IL-10 levels (0.78 pg/mL) in cells infected with both Gram-negative strains (E. coli ATCC 25922 and P. aeruginosa ATCC 27853). In the E. coli ATCC 25922 infection on HCT-8, Lb. brevis 10 (SN2) and Lb. plantarum F (SN3) supernatants associated with Lf slightly promoted the secretion of IL-10 (1.12 pg/mL) compared to the control. The supernatant from Lb. rhamnosus MF9 (SN1) significantly uplifted the IL-10 concentration (1.59 pg/mL) in cells infected with P. aeruginosa ATCC 27853, correlating with the possible anti-inflammatory action by suppressing IL-8 release. Lf generated slightly higher levels of IL-10 in E. coli ATCC 25922 (0.84 pg/mL) than those observed in the pathogen control (0.71 pg/mL); for P. aeruginosa ATCC 27853, no significant data has been recorded on the Lf treatment (Figure 8, a). In both Gram-positive bacteria (S. aureus ATCC 25923 and E. faecium), individual and Lf-associated supernatants induced an anti-inflammatory response via release of IL-10. In S. aureus ATCC 25923, compared to untreated cells, all the supernatants, alone and combined with Lf, promoted IL-10 synthesis. Regarding the infection with E. faecium, while SN1 and SN2, both mixed with Lf, determined lower IL-10 expression (0.84 pg/mL) in contrast to control cells, all supernatants, but especially SN2, induced an anti-inflammatory effect (SN1 – 2.27 pg/mL, SN2 – 4.24 pg/mL and SN3 – 1.32 pg/mL). Lf increased the IL-10 concentration in E. faecium and S. aureus ATCC 25923 (1.32 and 0.84 pg/mL) in comparison to the pathogens alone (0.78 and 0.71 pg/mL) (Figure 8, b). No statistically significant differences were observed.
The dataset corresponding to the effect of each tested variant on IL-10 levels following expose to Gram-negative pathogens was correlated with a statistically significant Kruskal-Wallis ANOVA (H = 19.02, p = 0.008, ndataset = 52). Post-hoc analysis testing revealed statistically significant differences between the effects Gram-negative pathogens and SN1 (mean rank difference = -27.92, p = 0.017, nformer = 6, nlatter = 8) (Figure 9).
The dataset corresponding to the effect of each tested variant on IL-10 levels following expose to Gram-positive pathogens was correlated with a statistically significant Kruskal-Wallis ANOVA (H = 20.21, p = 0.005, ndataset = 52). Post-hoc analysis testing revealed statistically significant differences between the effects LF and SN2 (mean rank difference = -24.79, p = 0.016, nformer = 10, nlatter = 8) (Figure 10).
3.2.2. Influence on Adhesion Capacity on Cellular Substrates (Adhesion Patterns and Adhesion Index)
The inhibitory effects of LAB supernatants, alone or associated with commercial Lf, against adhesion on HCT-8 cells were assessed by analyzing the adhesion patterns and Adhesion Indices for all the tested strains (E. coli ATCC 25922, P. aeruginosa ATCC 27853, S. aureus ATCC 25923). For each strain, the results were compared to three distinct controls: (1) adhesion control, comprising the bacterial and eukaryotic cells; (2) MRS control, represented by the bacterium, MRS media and HCT-8 cells; (3) Lf control, including the bacterial cells treated with commercial Lf and the HCT-8 cells.
Out of all the treatments, SN1 (Lb. rhamnosus MF9) and its combination with commercial Lf were effective against every opportunistic bacterial strain considered in this experiment, especially compared to the adhesion and MRS controls. In the case of Gram-negative pathogens (P. aeruginosa ATCC 27853 and E. coli ATCC 25922), commercial Lf was individually effective against adhesion to cellular substrate as well (AI% = 50%). Additionally, for P. aeruginosa ATCC 27853, the mixture between SN3 (Lb. plantarum F) and food supplements-derived Lf had a remarkable anti-adherent effect (AI% = 25%). In contrast, for E. coli ATCC 25922, all the probiotic supernatants, alone and correlated with Lf, were significantly efficient, with the lowest AI% values yielded by SN2 (Lb. brevis 10) and SN3 (equal values of AI% = 15%), along with their association with Lf, both generating an AI% of 10%. Compared to the Gram-negative pathogens, for the Gram-positive strain included in this experiment (S. aureus ATCC 25923), only SN1, alone and mixed with commercial Lf, induced a lower AI% (50%). The obtained data indicates a more prevalent inhibitory effect on bacterial adhesion to cell substrate against the Gram-negative strains (Figure 11).
Three main adhesion patterns (aggregative, diffuse and localized) have been previously described in vitro when infecting eukaryotic cells with bacterial strains [62,63,64]. Adhesion is a key virulence factor and implementing novel agents targeting this mechanism is a valuable option to standard antibiotic therapy as it mitigates the dissemination of MDR mechanisms.
Concerning the adhesion pattern to HCT-8 cells, when untreated and treated with MRS and commercial Lf, P. aeruginosa ATCC 27853 exhibited a tendency to evenly bind to eukaryotic cells, displayed as a diffuse pattern. The treatment with Lb. rhamnosus MF9 supernatant (SN1) potentially induced a stress response, given the transition to a localized pattern. Interestingly, the Lb. brevis 10 supernatant (SN2), alone and mixed with Lf, and the Lb. plantarum F supernatant (SN3) determined a shift to a diffuse-localized pattern, illustrated as the development of fine, but still dispersed microcolonies, which significates that this probiotic strain has an antipathogenic potential, being able to reduce the adherence capacity (Figure 12).
With respect to the adherence capacity of E. coli ATCC 25922 to cellular substrates, compared to the other Gram-negative strain, even though the Adhesion Indices (AI%) were reduced, neither of the considered treatments altered the adhesion patterns, the diffuse pattern being prevalent (Figure 13).
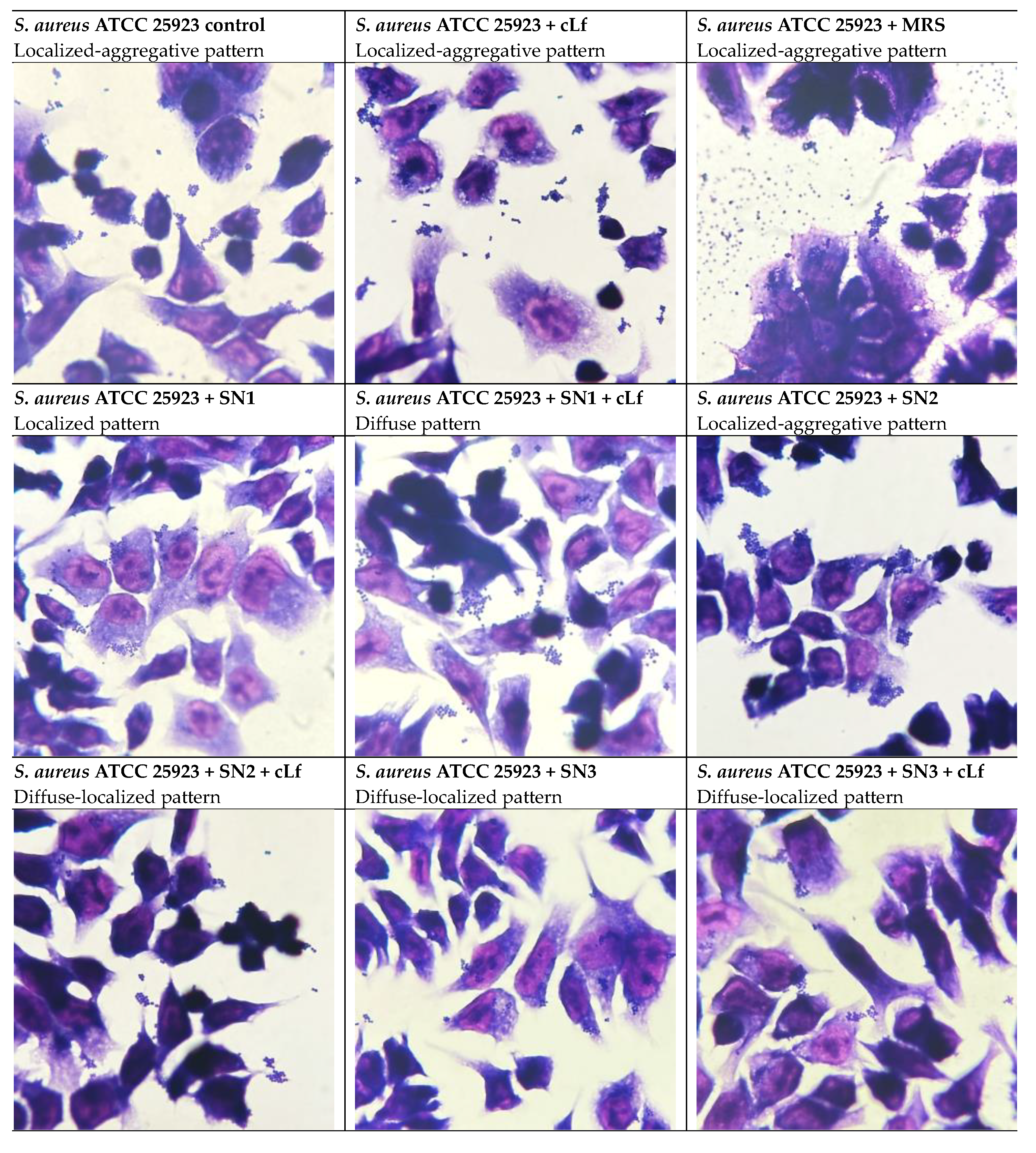
Regarding the ability of S. aureus ATCC 25923 to adhere to HCT-8 cells, the localized-aggregative pattern was identified when untreated and treated with MRS and commercial Lf. A shift to the localized pattern was remarked in the bacterial interaction with the supernatant from Lb. rhamnosus MF9 (SN1), suggesting bacterial clustering in response to the treatment. In association with Lf, SN1 yielded a diffuse pattern, perhaps determining an aggregate breakage. The supernatant obtained from Lb. brevis 10 (SN2) did not alter the colonization pattern, whilst its combination with Lf, as well as individual and Lf-associated Lb. plantarum F supernatant (SN3) produced a diffuse-localized pattern, indicating an anti-virulence/antipathogenic potential of this combination, this effect being underlined by the formation of diffuse cell clusters (Figure 14).
Figure 14.
Diffuse adhesion patterns of E. coli ATCC 25922 to cellular substrates in the presence of all tested combinations, as well in controls (Giemsa staining, 1000x).
Figure 14.
Diffuse adhesion patterns of E. coli ATCC 25922 to cellular substrates in the presence of all tested combinations, as well in controls (Giemsa staining, 1000x).
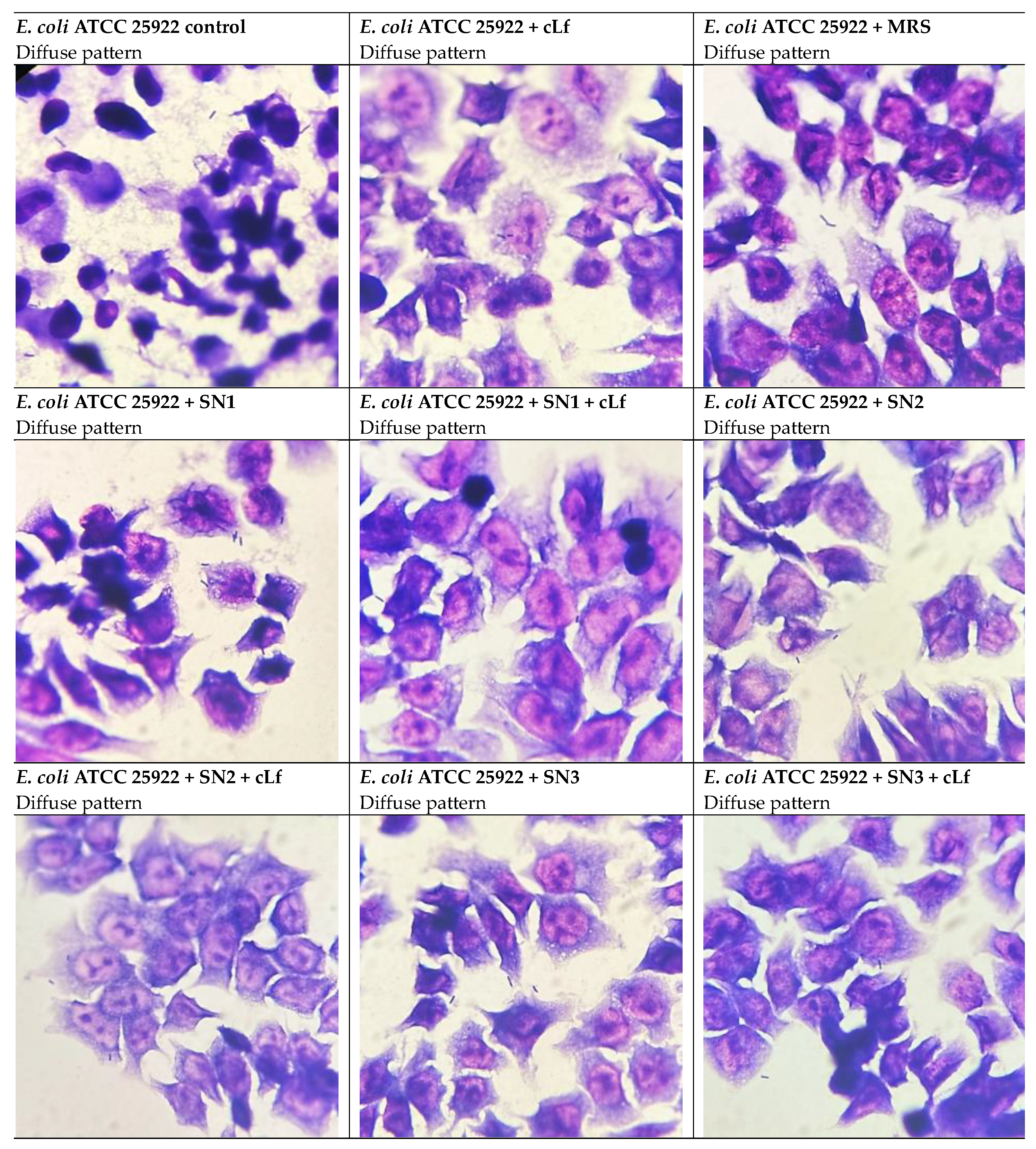
Figure 15.
Adhesion patterns of S. aureus ATCC 25923 to cellular substrates in the presence of tested combinations and in controls (Giemsa staining, 1000x).
Figure 15.
Adhesion patterns of S. aureus ATCC 25923 to cellular substrates in the presence of tested combinations and in controls (Giemsa staining, 1000x).
4. Discussion
The worldwide MDR phenomenon stands from excessive, often times unfounded use of antibiotics in healthcare units and non-clinical settings. Therefore, pathogenic and conditionally pathogenic bacteria adopted several resistance strategies to most antibiotic classes. This enforces the urgency to implement novel therapeutic approaches in order to take action against infections caused by MDR pathogens [8,24,65]. The antimicrobial effect of Lf has been previously characterized, alongside its capacity to block pathogenic bacteria from adhering to different surfaces. For this reason, to reduce persistent infections emerging from biofilm formation, Lf has been studied as an anti-adherent agent [66]. The antibacterial action of Lf has been previously described against several standardized and clinically-isolated bacterial strains, such as S. aureus, E. coli, P. aeruginosa, E. faecium, Vibrio parahaemolyticus, Bacillus subtilis, Porphyromonas gingivalis, Prevotella intermedia, Streptococcus agalactiae, K. pneumoniae and Helicobacter pylori [10,17,25,67,68,69,70,71,72,73,74,75,76,77,78,79]. In addition to Lf, LAB strains could be antibiotic adjuvants arising from their benefits provided for the host, via the production of antimicrobial agents such as lactic acid, H2O2 and bacteriocins [12,80,81]. Probiotic strains represent a promising venue in managing MDR infections stemming from their antimicrobial and anti-biofilm capacities towards Gram-positive and Gram-negative bacteria, by inhibiting adhesion to host cells and QS mechanisms [28,37].
In this context, in our study, we assessed the suppressive effects of high-purity bLf, in opposition to commercial Lf, on bacterial growth and adhesion to both inert and cellular substrates of several representative opportunistic strains. In addition, we tested the aforementioned properties of the combination between commercial Lf and supernatants derived from cultures of Lb. rhamnosus MF9, Lb. plantarum F and Lb. brevis 10. Lastly, we evaluated the potential immunomodulatory effects of these combinations in managing opportunistic infections.
Antimicrobial peptides (AMPs) are part of the innate immunity and are represented by short-chain positively or negatively charged molecules, with proven antibacterial, antifungal, antiviral and antitumoral properties [24,70,82,83]. Lfcin is such an AMP obtained by gastric pepsin digestion from the amino end of bLf, in the form of LfcinB, or hLf, as LfcinH [83]. S. aureus usually resides in the human normal microbiota, but is also an opportunistic pathogen isolated from a range of infections, including chronic skin conditions and subcutaneous, muscular or bone infections. This microorganism is capable of producing exotoxins leading to additional pathogenic processes, such as Toxic Shock Syndrome Toxin-1 (TSST-1), associated with TSS, and exfoliatins, responsible for the scalded skin syndrome [84]. In this sense, the study published by Ulvatne and Vorland aimed to test the inhibitory effects of an AMP resulted from LfcinB, LfcinB(17-31) respectively, against S. aureus ATCC 25923 [82,85]. The data revealed diminished MIC values (30 µg/mL), in support of the inhibitory effect of Lfcins against bacterial growth [82]. Data from another report underline MIC of native-form bLf (partially iron-saturated) values between 125 and 1000 μg/mL against 15 bovine mastitis-causing S. aureus strains, with the most predominant value being 500 μg/mL, roughly similar to our results (2000 μg/mL for bLf) on S. aureus ATCC 25923 [75]. Equivalent to our study design, Jugert et al. tested several commercially available food Lf-containing supplements Lf against 10 S. aureus isolates and obtained a moderate (> 5–8 mm diameter) and strong inhibition (> 8 mm diameter) in more than half of the strains by using the disk diffusion method [86]. In relation to our data on the antibiofilm action of Lf (MBECbLf = 500 µg/mL and MBECcommercial Lf = 1250 µg/mL), is has been reported that 1.0 mg/mL and 10 mg/mL of bLf reduced biomass and viability of MDR S. aureus biofilms after 12 and 24 hours. Thus, by inhibiting further adherence in pre-formed biofilms, bLf has been suggested to be a bacteriostatic rather than bactericidal compound [87]. Lf has also been tested in the form of a hydrolysate and disrupted staphylococcal biofilms on glass surfaces at the concentration of 2.5 mg/mL [88]. This activity of bLf might be a valuable strategy in eradicating biofilms established on medical devices.
Infections produced by E. faecium (i.e., urinary tract infections - UTIs, endocarditis) are a global health concern as a result of extreme usage of antibiotics. MDR enterococci are responsible for a significant proportion of HAI, acquiring tolerance to most antimicrobial drugs [70]. Additionally, virulence factors as capsular polysaccharides and adhesins contribute to antibiotic resistance in E. faecium strains. Resistance to antibiotics and virulence factors can be acquired by horizontal genetic transfer (via mobile genetic elements) between enterococci [89]. Mishra et al. developed narrow-spectrum AMPs exemplified by LfcinB6 5L and 6L derived from the antimicrobial domain of bLf. These peptides provided MIC values ranging between 4–16 μg/mL against several antibiotic-resistant E. faecium clinical isolates and prompted a 90% inhibition of biofilm formation for the E. faecium C68 strain at 32 μg/mL. Moreover, this anti-adherent concentration induced E. faecium membrane depolarization, fluidization and disruption [70]. These studies indicate a higher efficacy of bLf-derived AMPs against Gram-positive bacteria, compared to our data obtained on purified bLf (MIC = 500 µg/mL and MBEC = 250 µg/mL) and commercial Lf (MIC = 5000 µg/mL and MBEC = 1250 µg/mL).
P. aeruginosa is a clinically relevant pathogen due to its QS network, allowing the bacteria to adapt to external stress factors by gene expression regulation [74]. P. aeruginosa is well known for adhering to cell surfaces, which correlates to chronic respiratory tract infections, wound infections, and diminished antibiotic therapy efficacy [69,74]. Our findings coincide with the data obtained by Kamiya et al. The effects of bLf treatment against growth and adhesion to inert substrates of 9 clinically-isolated P. aeruginosa strains were assessed. Similar to our observations, the bacterial adherence was predominantly suppressed, with bLf anti-biofilm concentrations recorded between 0.1-2 mg/mL, compared to the MIC of 16 mg/mL [74]. In contrast, another report focused on the growth inhibitory effects of LfcinB against P. aeruginosa ATCC 27853, generating a MIC of 15.6 μg/mL [90]. The native form of bLf displayed a significant inhibitory influence on P. aeruginosa PTCC 1707 biofilms at 14 mg/mL [91], compared to our observations (MBECbLf = 62.5 µg/mL and MBECcommercial Lf = 1250 µg/mL). Additionally, Ramamourthy et al. tested several bLf-derived peptides against P. aeruginosa PAO1 planktonic cell growth and biofilm formation, highlighting the effectiveness of LfcinB against this pathogen (MIC = 32 µM and MBEC = 128 µM) [69].
E. coli is part of the commensal bacterial communities colonizing the intestinal tract and also responsible for HAI and other opportunistic infections, including wound infections following surgery, UTIs and septicemia. The diseases caused by E. coli stem from different strains, affecting the small [enterotoxigenic E. coli (ETEC), enteropathogenic E. coli (EPEC), enteroaggregative E. coli (EAEC)] and large intestines [enteroinvasive E. coli (EIEC), enterohemorrhagic E. coli (EHEC)]. Due to the increasing resistance in E. coli strains, Lf and its derivatives could be used to alleviate these diarrheagenic conditions [71]. The restraining effect of bLf-resulted AMPs was evaluated for E. coli ATCC 25922 as well; hence, LfcinB induced a MIC value of 30 µg/mL, but also 62.5 µg/mL [82,90]. Moreover, the study conducted by Huertas Méndez et al. aimed to test the antimicrobial action of bLf, LfcinB and several LfcinB-derived short synthetic peptides against E. coli ATCC 11775. The data revealed a greater inhibitory action of Lfcin B compared to bLf, illustrated by MIC values of 100 μg/mL for LfcinB, ranging between 6.2 and 25 μg/mL for LfcinB synthetic peptides and larger than 200 μg/mL in the case of bLf. The elevated inhibitory concentrations of bLf point to a dose-dependent efficacy of the native protein [83]. In addition, the pronounced antimicrobial action of Lf food supplements against 10 E. coli clinical strains was underlined by generating detectable growth inhibition in 90% of the strains (growth inhibition zones higher than 5–8 mm in diameter) [86]. Consistent with our experimental findings (MBECbLf = 2000 µg/mL and MBECcommercial Lf = 2000 µg/mL), when administered at the concentration of 16 mg/mL, bLf determined a modest inhibition towards E. coli ATCC 25922 biofilms [91].
Results of research done by Elass-Rochard at al. highlighted the presence of two binding sites on both hLf and bLf to LPS on the E. coli 055B5 cell wall [92]. Other than LPS, outer membrane porins act as Lf receptors in several Gram-negative bacteria residing in the gut. Several species, such as E. coli, Shigella sp. and Salmonella sp., displayed Lf tolerance due to masking of porin binding sites by the carbohydrate O-antigenic chains of LPS [93]. This binding site shielding could be an explanation for our heterogenous results obtained on Gram-negative strains, the effects of Lf being strain-dependent.
Probiotics, including LAB, are defined as live microorganisms that benefit the host health when administered in proper doses [33,37,80,94]. LAB CFS can be promising adjuvants of antibiotic therapy, proven by the synergistic action of Lb. rhamnosus PTCC 1637 CFS with aminoglycosides as gentamicin and amikacin against P. aeruginosa PTCC 1430 [95]. Complementary to our data, Lb. plantarum and Lb. rhamnosus CFS were shown to prevent the formation of biofilms in several S. aureus strains [96]. A paper published by Sulaiman et al. shed light on the biofilm inhibitory effects of cell-free supernatants from several LAB strains (including Lb. plantarum OBISE A9) on biofilm formation of multiple pathogens: E. coli EAEC DO28J, Pseudomonas aeruginosa EO102, S. aureus OAU AAA 059A. Interestingly, the results point towards a higher susceptibility of E. coli to LAB metabolites, followed by S. aureus and P. aeruginosa. The LAB supernatants had a biofilm inhibitory activity of approximately 24% on S. aureus when used at the concentration of 1% [37]. This is in contrast to our data, since no visible suppressive effect against Gram-negative strains was observed, and only SN2 (Lb. brevis 10) induced a measurable MBEC value (50%) when applied on S. aureus ATCC 25923. In regards to the other Gram-positive strain (E. faecium) included in our study, scientific reports mostly refer to the probiotic or antimicrobial activity of several E. faecium strains of rather than the potentially pathogenic strains [89,97,98]. Future studies could elucidate the inhibitory action of probiotic supernatants on E. faecium growth and biofilm formation.
The association between Lf and LAB has been investigated in prior studies for its antimicrobial activities. In this sense, it has been proven that bLf can act as a prebiotic on several Lactobacillus strains, including Lb. rhamnosus ATCC 53103, in dose-responsive manner, when their growth has been inhibited by prior incubation at a lower temperature [36]. Another in vitro study focused on the antimicrobial synergistic action of supernatants derived from several LAB strains, including Lb. rhamnosus ATCC 53103 and ATCC 7469, combined with apo-bLf (iron-free bLf), on 9 MRSA (methicillin-resistant S. aureus) clinical strains. Out of the probiotic strains, Lb. fermentum ATCC 11739, Bifidobacterium animalis subsp. lactis BCRC 17394 and Bifidobacterium longum subsp. longum ATCC 15707 had a significant impact on bacterial growth, with the first LAB strain generating the most notable synergistical effect when associated with apo-bLf. These results could be attributed to the potentiating action of bLf, facilitating the entrance of LAB metabolites into pathogenic bacteria [23].
To our knowledge, the combination between Lf and LAB CFS has not been tested for its effects on release of cytokines before. Therefore, previous research has only described the immunomodulatory action of Lf and LAB individually. Lf is considered to contribute to the protection of intestinal mucosa against inflammation and infections generated by adhesion of bacterial pathogens [99]. The absence of IL-6 and TNF-α secretion in HCT-8 cells, as observed in our experiments, might be explained by limited innate responsiveness of intestinal epithelial cells. Given that HCT-8 is a transformed epithelial cell line, it might preferentially produce chemokines like IL-8 instead of macrophage-type cytokines like IL-6 and TNF-α [100,101,102]. Our results are in support of the anti-inflammatory effects of Lf, since it suppressed the pro-inflammatory chemokine IL-8 secretion and promoted the anti-inflammatory cytokine IL-10 release in HCT-8 cells challenged with E. coli ATCC 25922, P. aeruginosa ATCC 27853, S. aureus ATCC 25923 and E. faecium. With respect to IL-8, Lf had a greater inhibitory action when administered in parallel with P. aeruginosa ATCC 27853, S. aureus ATCC 25923 and E. faecium. In the case of IL-10, the pre-treatment induced a modest anti-inflammatory response in infections with all pathogens. Our findings are in agreement with the bLf inhibition on IL-8 secretion in THP-1 and Mono Mac 6 cells stimulated with LPS, but do not align with the diminished IL-10 levels in Mono Mac 6 cells observed in the research done by Håversen et al. [103]. In a similar manner, in Caco-2 colon adenocarcinoma cells infected with E. coli HB101, the treatment with native and iron-saturated bLf reduced IL-8 levels [104]. Compared to our data obtained in an immune assay, levels of IL-6, IL-8 and TNF-α in mRNA have been quantified, with bLf inhibiting cytokine mRNA expression in E. coli LF82-infected Caco-2 cells [99]. In agreement with our observations, in another report, IL-6 synthesis was not detectable in Caco-2 cells. Contrary to our findings, the data obtained by Frioni et al. suggest a potential synergism between IFN-γ and bLf in Caco-2 cells infected with the clinical strain E. coli LF82 by highlighting an increase in IL-8 levels in IFN-γ-stimulated cells infected with E. coli. The effects of Lf on inflammation might depend on the pathogen and cell line used [105].
Probiotics, either as whole bacterial cells or culture supernatants, have been studied for their effects on modifying cytokine secretion patterns in eukaryotic cell cultures and in vivo models. It is considered that several Lactobacillus strains might alleviate inflammatory responses in cell cultures infected with pathogenic bacteria by upregulating IL-10 and downregulating TNF-α and IL-8 [106]. Our results demonstrated that CFS derived from Lb. rhamnosus MF9, Lb. brevis 10, Lb. plantarum F cultures, alone and Lf-coupled, could ameliorate inflammation caused by infections with both Gram-positive and Gram-negative strains in HCT-8 cells by suppressing IL-8 levels and promoting IL-10 secretion. Notably, SN1 (Lb. rhamnosus MF9) combined with Lf inhibited IL-8 release induced by both Gram-negative bacteria, as well as S. aureus ATCC 25923. Likewise, the Lb. rhamnosus MF9 CFS promoted the highest IL-10 response in P. aeruginosa ATCC 27853 infected cells, compared to the infection control. In the case of the infection with E. faecium, Lb. brevis 10 most significantly increased IL-10 levels. One report used a TNF-α-based method of inflammation induction in Caco-2 cells co-cultured with Lb. plantarum, highlighting the reduction in IL-8 levels by this LAB strain [107]. In a model of HT-29 colon cancer cells co-cultured with E. coli ATCC 43889 and several lactobacilli strains, the ability of some Lb. plantarum and Lb. rhamnosus strains to inhibit inflammation by lowering E. coli-induced IL-8 release has been observed [106]. When incubated with Lb. rhamnosus KLDS, inhibition of IL-8 production was described in Caco-2 cells challenged with LPS from E. coli 0111:B4 [108]. Pre-treatment with Lb. brevis KU15152 produced an equivalent suppression of IL-8 induced by S. aureus lipoteichoic acid in HT-29 cells [109]. The mRNA levels of pro-inflammatory cytokines have been measured in similar settings; a paper by Huang et al. emphasized the restraining effect of treatment with Lb. rhamnosus ATCC 53103 on P. aeruginosa PAO1-generated IL-8 expression in SW480 colon cancer cells [110]. In another experiment, lack of IL-6 and TNF-α synthesis has been reported in Caco-2 cells. Correlating to our ELISA results, the 4 hours pre-treatment with Lactiplantibacillus plantarum (previously Lb. plantarum) in Salmonella-challenged Caco-2 cells stimulated the mRNA expression of IL-10 compared to the pathogen alone or untreated cells [111]. In support of the in vitro results on pro-inflammatory cytokines inhibition, in ICR mice infected with an ETEC pathotype strain, inoculation of Lb. brevis AY858 decreased IL-6 relative mRNA expression [112]. While previous research on CFS from the strains included in our experiments is scarce, interestingly, in an early study, treatment with cell-free probiotic cultures exhibited pro-inflammatory effects on HeLa cells infected with S. aureus and E. coli strains, by promoting release of IL-6 and IL-8 and blocking synthesis of IL-10 [113]. A supernatant from Lactococcus lactis mitigated IL-8 production in LPS-stimulated HT-29 cells [114]. Zanetta et al. evaluated the beneficial effects of CFS from Lactobacillus johnsonii LJO02 in S. aureus ATCC 43300-infected HaCaT cells and revealed the reduction of IL-6 production in infected and CFS treated cells as opposed to infected cells (control) [115]. Notably, in a recent study, CFS from Lb. rhamnosus displayed inhibitory activities on IL-6, IL-10 and TNF-α release in human P. aeruginosa LPS-stimulated PBMC [116]. When released by activated immune cells, pro-inflammatory cytokines aid in eradication of infections and cancer cells. The immunostimulatory action of several LAB supernatants has been tested in RAW264.7 cells, with results indicating the release of pro-inflammatory molecules (IL-6, TNF-α) [117].
Adherence of pathogenic bacteria to susceptible host cells is the stage that precedes the subsequent steps in the initiation of an infectious process [118]. In our experiments, when combined with LAB CFS, Lf improved the anti-adherence properties in HCT-8 cells infected with Gram-positive and Gram-negative bacteria. With regard to the interaction of opportunistic pathogens with cellular substrates, Berlutti et al. investigated the influence of bLf on adhesion and invasion capacities of P. aeruginosa ATCC 15692, using the A549 lung carcinoma cell line, cultured in monolayers [79]. Interestingly, compared to our results, the adhesiveness (measured as Adhesion Efficiency) of P. aeruginosa was improved after 4 hours of incubation with bLf. Conversely, the incubation of the infected cells with bLf diminished the invasiveness (measured as Invasion Efficiency) of P. aeruginosa [79]. The results of the paper by Kawasaki et al., contrasting with the aforementioned work and consistent with our observations, shed light on the suppressive effect of bLf at 16 mg/mL on the adherence of 8 strains of ETEC and EPEC pathotypes to the JTC-17 cervical cancer cells [77]. The differing results could be correlated to the source of the Lf used, considering that, for the adhesion assay, we used commercial Lf, derived from food supplements which also contain some excipients.
In addition to the anti-adherent properties of Lf, probiotic strains are able to block bacterial adherence and biofilm formation by competing for adhesion sites on eukaryotic cells due to exopolysaccharides and cell wall proteins [118]. The trend among our adherence data is the effect of LAB supernatants on P. aeruginosa ATCC 27853 and E. coli ATCC 25922 rather than S. aureus ATCC 25923. Especially, individual or Lf-combined SN1 (Lb. rhamnosus MF9) was effective in preventing the adherence of all strains. E. coli ATCC 25922 proved to be the most sensitive to all supernatants, most notably to SN2 (Lb. brevis 10) and SN3 (Lb. plantarum F). In resemblance to the immunomodulation assay, these bacteria have been considered both as whole cells and CFS for previous adherence studies. In line with our findings, previous reports have stated the inhibitory action of whole bacterial cells from several Lb. rhamnosus strains (such as GG, GR-1, DSM 20021, DSM7133, ATCC 7469) on adhesiveness of multiple S. aureus, P. aeruginosa and E. coli strains to normal or tumoral eukaryotic cells [38,110,118,119,120,121,122,123,124]. In addition, Lb. plantarum strains (299v, CS24.2, ZLP001, L15, GL17, MKMB01, MKMB02, L33, L125) isolated from different sources, including fermented foods, have been tested for similar activities against E. coli and S. aureus [106,111,124,125,126,127,128]. Khodaii et al. evaluated the effects of bacteria-free supernatants from Lb. plantarum 5C, Lb. brevis 1D and Lb. rhamnosus T against invasiveness of EIEC pathotype in T84 colon cancer cells, since invasion is a significant virulence factor for the evolution of an infection [129]. The Lb. brevis 1D supernatant displayed the most visible anti-invasive effect against E. coli on T84 cells, corresponding to our observations on adhesion inhibition [129]. Furthermore, CFS from Lb. plantarum ATCC 14917 and Lb. rhamnosus ATCC 53103 were demonstrated to prevent adhesion of MSSA (Methicillin-Susceptible S. aureus) to human osteoblasts [96].
Prior studies investigating the influence of Lf and/or LAB CFS on alteration of adhesion patterns are limited. In our experiments, for P. aeruginosa ATCC 27853, compared to the control (diffuse pattern), the diffuse-localized pattern was predominant in most of the tested conditions. Concerning E. coli ATCC 25922, the diffuse adhesion pattern was not modified in either of the experimental conditions. In cells infected with S. aureus ATCC 25923, a similar shift to the diffuse-localized adhesion pattern was noticed. The bacterial capacity to adhere to and enter eukaryotic cells is generally dictated by ≥1 virulence factors, including adhesins [13]. Pili are the most common adhesins of Gram-positive and Gram-negative bacteria, involved in motility, generation of biofilms and conjugation, apart from adherence [130]. The localized adhesion phenotype, characteristic to diarrheagenic E. coli EPEC, is mediated by intimin, an outer membrane adhesin, and is responsible for inducing Attaching and Effacing (A/E) lesions in enterocytes [64,131,132]. Especially in the case of S. aureus ATCC 25923, the transition from a localized-aggregative adhesion to the diffuse-localized pattern might indicate a virulence reduction under treatments with LAB CFS and commercial Lf. Targeting virulence strategies such as bacterial adhesion might be a promising venue in managing MDR infections, since it exerts a low selective pressure for the development of novel MDR mechanisms by withholding the bacterium’s virulence without hindering its viability [130,133].
As a result of their health-promoting benefits, food supplements containing both LAB strains and Lf have been formulated and considered in a series of in vivo studies for modulating the female genital normal microbiota and managing dysbiosis [23,80,81,134,135]. One example is the Respecta®® complex, comprising a combination of two probiotic strains: Lb. acidophilus La-14, Lb. rhamnosus HN001, and bLf RCXTM (50 mg) [80,81,134,135]. A pilot study conducted by De Alberti et al. concluded that the oral administration of the Respecta®® dietary supplement promoted the colonization of vaginal microbiota with the aforementioned strains in the active-treatment group compared to the placebo group [80], leading to the potential application in restoring vaginal microbiota. Another two clinical trials underscored, besides the colonization with both strains, an amelioration of altered female genital microbiota in a cohort of women suffering from bacterial vaginosis which were administered the same Respecta®® complex [134,135].
5. Conclusions
High-purity bovine lactoferrin generated a more notable antimicrobial and anti-biofilm activity as opposed to food supplements-derived lactoferrin, supported by the results obtained in quantitative assessments, indicating that the source of Lf might play an important role in its efficacy. Both lactoferrin variants displayed a more significant modulation of bacterial adhesion to inert substrates, rather than bacterial growth, especially on the tested Gram-positive strains (S. aureus ATCC 25923 and E. faecium). Regarding the Gram-negative pathogens (E. coli ATCC 25922 and P. aeruginosa ATCC 27853), the inhibitory effect of lactoferrin on biofilm generation was dependent on the strain included in the experiment; thus, the treatments were more effective on biofilm eradication in P. aeruginosa ATCC 27853 compared to E. coli ATCC 25922, potentially as a consequence of the structure of Lf receptors on the bacterial membrane. The Gram-positive strains were more susceptible to the combination between commercial lactoferrin and probiotic supernatants. By the addition of Lactic Acid Bacteria metabolites, lactoferrin was more potent against biofilm formation. Cell-free supernatants obtained from Lb. rhamnosus MF9, Lb. brevis 10 and Lb. plantarum F cultures, when administered individually or combined with lactoferrin, modulated the release of pro-inflammatory and anti-inflammatory cytokines, promoting an overall inhibition of inflammation in eukaryotic cells stimulated with bacterial pathogens. Similarly, lactoferrin alone inhibited IL-8 synthesis and stimulated IL-10 release, supporting its quality of an anti-inflammatory agent. The addition of probiotic metabolites to lactoferrin substantially prevented bacterial adhesion to cellular substrates. The adhesiveness of Gram-negative pathogens (P. aeruginosa ATCC 27853 and E. coli ATCC 25922) to eukaryotic cells was more hindered by Lactic Acid Bacteria supernatants in contrast to their Gram-positive counterpart (S. aureus ATCC 25923). The Lb. rhamnosus MF9 supernatant, either alone or coupled with lactoferrin, effectively blocked the adherence of all tested strains. To conclude with, the results of this preliminary study encourage the therapeutic use of lactoferrin and Lactic Acid Bacteria supernatants, individually or combined, as adjuvants or for prophylaxis, especially in immunosuppressed patients, given their abilities to inhibit bacterial growth and adherence, and modulate cytokine release towards eradicating persistent infections.
Author Contributions
Conceptualization, D.A.C., L.M.D. and V.L.; methodology, D.A.C., L.M.D, V.A.B., L.I.M., L.I.V., A.I., and C.B.; validation, L.M.D., Y.K., C.C., C.B., and V.L.; formal analysis, D.A.C. and L.M.D.; investigation, D.A.C., V.A.B., L.M.D, L.I.M., L.I.V., C.C., and A.I.; resources, L.M.D and C.B.; data curation, D.A.C.; writing—original draft preparation, D.A.C., L.M.D, and V.L.; writing—review and editing, L.M.D and V.L.; visualization, D.A.C. and A.I.; supervision, L.M.D., C.B., and V.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| A/E | Attaching and Effacing |
| AI% | Adhesion Index |
| AMPs | Antimicrobial Peptides |
| AMR | Antimicrobial Resistance |
| ATCC | American Type Culture Collection |
| bLf | Bovine lactoferrin |
| CFS | Cell-Free Supernatants |
| CFU | Colony Forming Units |
| DMEM | Dulbecco’s Modified Eagle Medium |
| EPS | Extracellular Polymeric Substances |
| EVs | Extracellular Vesicles |
| FBS | Fetal Bovine Serum |
| HAI | Healthcare-Associated Infections |
| hLf | Human Lactoferrin |
| LAB | Lactic Acid Bacteria |
| Lf | Lactoferrin |
| Lfcin | Lactoferricin |
| LfcinB | Bovine Lactoferricin |
| LfcinH | Human Lactoferricin |
| MBEC | Minimal Biofilm Eradication Concentration |
| MDR | Multi-Drug Resistance |
| MIC | Minimal Inhibitory Concentration |
| MRS | Man-Rogosa-Sharpe |
| MRSA | Methicillin-Resistant Staphylococcus aureus |
| MSSA | Methicillin-Susceptible Staphylococcus aureus |
| PBS | Phosphate Buffered Saline |
| QS | Quorum Sensing |
| SEM | Standard Error of Mean |
| TSST-1 | Toxic Shock Syndrome Toxin-1 |
| UTI | Urinary Tract Infection |
| WHO | World Health Organization |
References
- Tsalidou M, Stergiopoulou T, Bostanitis I, et al. Surveillance of Antimicrobial Resistance and Multidrug Resistance Prevalence of Clinical Isolates in a Regional Hospital in Northern Greece. Antibiotics. 2023;12(11). [CrossRef]
- Saha M, Sarkar A. Review on multiple facets of drug resistance: A rising challenge in the 21st century. J Xenobiot. 2021;11(4):197-214. [CrossRef]
- van Duin D, Paterson DL. Multidrug-Resistant Bacteria in the Community: An Update. Infect Dis Clin North Am. 2020;34(4):709-722. [CrossRef]
- Bharadwaj A, Rastogi A, Pandey S, Gupta S, Sohal JS. Multidrug-Resistant Bacteria: Their Mechanism of Action and Prophylaxis. Biomed Res Int. 2022;2022. [CrossRef]
- Yang AF, Huang V, Samaroo-Campbell J, Augenbraun M. Multi-drug resistant Pseudomonas aeruginosa: a 2019–2020 single center retrospective case control study. Infect Prev Pract. 2023;5(3). [CrossRef]
- Reardon S. WHO warns against “post-antibiotic” era. Nature. Published online April 30, 2014. [CrossRef]
- Mancuso G, Midiri A, Gerace E, Biondo C. Bacterial antibiotic resistance: the most critical pathogens. Pathogens. 2021;10(10). [CrossRef]
- Catalano A, Iacopetta D, Ceramella J, et al. Multidrug Resistance (MDR): A Widespread Phenomenon in Pharmacological Therapies. Molecules. 2022;27(3). [CrossRef]
- Rather MA, Gupta K, Mandal M. Microbial biofilm: formation, architecture, antibiotic resistance, and control strategies. Braz J Microbiol. 2021;52(4):1701-1718. [CrossRef]
- Costea DA, Lazãr V. Lactoferrin-A Multivalent Molecule. Roum Arch Microbiol Immunol. 2024;83(2):108-115. [CrossRef]
- Donlan RM, Costerton JW. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev. 2002;15(2):167-193. [CrossRef]
- Barenji EK, Beglari S, Tahghighi A, Azerang P, Rohani M. Evaluation of Anti-Bacterial and Anti-Biofilm Activity of Native Probiotic Strains of Lactobacillus Extracts. Iran Biomed J. 2024;28(2-3):102-112. [CrossRef]
- Lazar V, Oprea E, Ditu LM. Resistance, Tolerance, Virulence and Bacterial Pathogen Fitness—Current State and Envisioned Solutions for the Near Future. Pathogens. 2023;12(5). [CrossRef]
- Alkhulaifi MM, Alosaimi MM, Khan MS, et al. Assessment of Broad-Spectrum Antimicrobial, Antibiofilm, and Anticancer Potential of Lactoferrin Extracted from Camel Milk. Appl Biochem Biotechnol. 2024;196(3):1464-1480. [CrossRef]
- Sorensen M, Sorensen SPL. The proteins in whey. Comptes rendus. 1939;23(7):55-99. Accessed May 31, 2024. https://www.cabidigitallibrary.org/doi/full/10.5555/19420400573.
- Dyrda-Terniuk T, Pomastowski P. The Multifaceted Roles of Bovine Lactoferrin: Molecular Structure, Isolation Methods, Analytical Characteristics, and Biological Properties. J Agric Food Chem.American Chemical Society. 2023;71(51):20500-20531. [CrossRef]
- Kowalczyk P, Kaczyńska K, Kleczkowska P, Bukowska-Ośko I, Kramkowski K, Sulejczak D. The Lactoferrin Phenomenon—A Miracle Molecule. Molecules. 2022;27(9):2941. [CrossRef]
- Gibbons JA, Kanwar RK, Kanwar JR. Lactoferrin and cancer in different cancer models. Front Biosci. 2011;3:1080.
- Ramírez-Rico G, Drago-Serrano ME, León-Sicairos N, de la Garza M. Lactoferrin: A Nutraceutical with Activity against Colorectal Cancer. Front Pharmacol. 2022;13. [CrossRef]
- Kondapi AK. Targeting cancer with lactoferrin nanoparticles: Recent advances. Nanomedicine.Future Medicine Ltd. 2020;15(21):2071-2083. [CrossRef]
- Gruden Š, Ulrih NP. Diverse mechanisms of antimicrobial activities of lactoferrins, lactoferricins, and other lactoferrin-derived peptides. Int J Mol Sci. 2021;22(20). [CrossRef]
- Ramírez-Sánchez DA, Canizalez-Román A, León-Sicairos N, Pérez Martínez G. The anticancer activity of bovine lactoferrin is reduced by deglycosylation and it follows a different pathway in cervix and colon cancer cells. Food Sci Nutr. 2024;12(5):3516-3528. [CrossRef]
- Chen PW, Jheng TT, Shyu CL, Mao FC. Synergistic antibacterial efficacies of the combination of bovine lactoferrin or its hydrolysate with probiotic secretion in curbing the growth of meticillin-resistant Staphylococcus aureus. J Med Microbiol. 2013;62(PART 12):1845-1851. [CrossRef]
- Brouwer C, Welling MM, Alwasel S, Boekhout T. Potential health benefits of lactoferrin and derived peptides–how to qualify as a medical device? Crit Rev Microbiol. Preprint posted online 2025. [CrossRef]
- Angulo-Zamudio UA, Vidal JE, Nazmi K, et al. Lactoferrin Disaggregates Pneumococcal Biofilms and Inhibits Acquisition of Resistance Through Its DNase Activity. Front Microbiol. 2019;10. [CrossRef]
- Cao X, Ren Y, Lu Q, et al. Lactoferrin: A glycoprotein that plays an active role in human health. Front Nutr. 2023;9. [CrossRef]
- Abd El-Hack ME, Abdelnour SA, Kamal M, et al. Lactoferrin: Antimicrobial impacts, genomic guardian, therapeutic uses and clinical significance for humans and animals. Biomed Pharmacother. 2023;164. [CrossRef]
- Bădăluță VA, Curuțiu C, Dițu LM, Holban AM, Lazăr V. Probiotics in Wound Healing. Int J Mol Sci. 2024;25(11). [CrossRef]
- Sepehr A, Miri ST, Aghamohammad S, et al. Health benefits, antimicrobial activities, and potential applications of probiotics: A review. Medicine (Baltimore). 2024;103(52):e32412. [CrossRef]
- Plaza-Diaz J, Ruiz-Ojeda FJ, Gil-Campos M, Gil A. Mechanisms of Action of Probiotics. In: Adv Nutr. Vol 10. Oxford University Press; 2019:S49-S66. [CrossRef]
- Costa FF, Dias TG, Mendes PM, et al. Antioxidant and Antimicrobial Properties of Probiotics: Insights from In Vitro Assays. Probiotics Antimicrob Proteins. Preprint posted online 2024. [CrossRef]
- Zommiti M, Feuilloley MGJ, Connil N. Update of probiotics in human world: A nonstop source of benefactions till the end of time. Microorganisms. 2020;8(12):1-33. [CrossRef]
- Fijan S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int J Environ Res Public Health. 2014;11(5):4745-4767. [CrossRef]
- Santacroce L, Charitos IA, Bottalico L. A successful history: probiotics and their potential as antimicrobials. Expert Rev Anti Infect Ther. 2019;17(8):635-645. [CrossRef]
- Kruis W, Frič P, Pokrotnieks J, et al. Maintaining remission of ulcerative colitis with the probiotic Escherichia coli Nissle 1917 is as effective as with standard mesalazine. Gut. 2004;53(11):1617-1623. [CrossRef]
- Chen PW, Liu ZS, Kuo TC, Hsieh MC, Li ZW. Prebiotic effects of bovine lactoferrin on specific probiotic bacteria. BioMetals. 2017;30(2):237-248. [CrossRef]
- Sulaiman AA, Adetoye AA, Ayeni FA. Lactic Acid Bacteria and Fermented Maize Supernatant (Omidun) Have Anti-Biofilm Properties Against Staphylococci and Enteroaggregative Escherichia coli Strains. FUDMA J Sci. 2024;7(6):250-260. [CrossRef]
- Palkovicsné Pézsa N, Kovács D, Somogyi F, et al. Effects of Lactobacillus rhamnosus DSM7133 on Intestinal Porcine Epithelial Cells. Animals. 2023;13(19). [CrossRef]
- Rahman MN, Barua N, Tin MCF, Dharmaratne P, Wong SH, Ip M. The use of probiotics and prebiotics in decolonizing pathogenic bacteria from the gut; a systematic review and meta-analysis of clinical outcomes. Gut Microbes. 2024;16(1). [CrossRef]
- Yadav P, Debnath N, Mehta PK, Kumar A, Yadav AK. Assessment of Antimicrobial Potential of Lactiplantibacillus plantarum and Their Derived Extracellular Vesicles. Mol Nutr Food Res. 2025;69(18). [CrossRef]
- Krzyżek P, Marinacci B, Vitale I, Grande R. Extracellular Vesicles of Probiotics: Shedding Light on the Biological Activity and Future Applications. Pharmaceutics. 2023;15(2). [CrossRef]
- Domínguez Rubio AP, D’Antoni CL, Piuri M, Pérez OE. Probiotics, Their Extracellular Vesicles and Infectious Diseases. Front Microbiol. 2022;13. [CrossRef]
- Baek J, Lee S, Lee J, Park J, Choi E, Kang SS. Utilization of Probiotic-Derived Extracellular Vesicles as Postbiotics and Their Role in Mental Health Therapeutics. Food Sci Anim Resour.Korean Society for Food Science of Animal Resources. 2024;44(6):1252-1265. [CrossRef]
- Salminen S, Collado MC, Endo A, et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat Rev Gastroenterol Hepatol.Nature Research. 2021;18(9):649-667. [CrossRef]
- Ciobanasu C. Bacterial Extracellular Vesicles and Antimicrobial Peptides: A Synergistic Approach to Overcome Antimicrobial Resistance. Antibiotics. 2025;14(4). [CrossRef]
- Li M, Mao B, Tang X, et al. Lactic acid bacteria derived extracellular vesicles: emerging bioactive nanoparticles in modulating host health. Gut Microbes. 2024;16(1). [CrossRef]
- Naskar A, Cho H, Kim KS. A Nanocomposite with Extracellular Vesicles from Lactobacillus paracasei as a Bioinspired Nanoantibiotic Targeting Staphylococcus aureus. Pharmaceutics. 2022;14(11). [CrossRef]
- Lee BH, Wu SC, Shen TL, Hsu YY, Chen CH, Hsu WH. The applications of Lactobacillus plantarum-derived extracellular vesicles as a novel natural antibacterial agent for improving quality and safety in tuna fish. Food Chem. 2021;340. [CrossRef]
- Champagne-Jorgensen K, Mian MF, McVey Neufeld KA, Stanisz AM, Bienenstock J. Membrane vesicles of Lacticaseibacillus rhamnosus JB-1 contain immunomodulatory lipoteichoic acid and are endocytosed by intestinal epithelial cells. Sci Rep. 2021;11(1). [CrossRef]
- Chen X, Li Q, Xie J, Nie S. Immunomodulatory Effects of Probiotic-Derived Extracellular Vesicles: Opportunities and Challenges. J Agric Food Chem. 2024;72(35):19259-19273. [CrossRef]
- Ronacher C, Gonzalez CF, Lorca GL. A critical evaluation of methodological and mechanistic insights on probiotic-derived extracellular vesicles. Front Nutr. 2025;12. [CrossRef]
- Liu R. A promising area of research in medicine: recent advances in properties and applications of Lactobacillus-derived exosomes. Front Microbiol. 2024;15. [CrossRef]
- Hu R, Lin H, Li J, et al. Probiotic Escherichia coli Nissle 1917-derived outer membrane vesicles enhance immunomodulation and antimicrobial activity in RAW264.7 macrophages. BMC Microbiol. 2020;20(1). [CrossRef]
- Zhao Q, Lai J, Jiang Y, et al. Lactiplantibacillus plantarum -derived extracellular vesicles alleviate acute lung injury by inhibiting ferroptosis of macrophages. J Nanobiotechnol. 2025;23(1). [CrossRef]
- Yang Y, Li N, Gao Y, et al. The activation impact of lactobacillus-derived extracellular vesicles on lipopolysaccharide-induced microglial cell. BMC Microbiol. 2024;24(1). [CrossRef]
- Badaluta VA, Ionascu A, Ditu LM, et al. Antimicrobial and immunomodulatory activity of natural bioactive compounds – an in vitro investigation with potential applications in managing Hidradenitis Suppurativa. Front Immunol. 2025;16. [CrossRef]
- Van LT, Hagiu I, Popovici A, et al. Antimicrobial Efficiency of Some Essential Oils in Antibiotic-Resistant Pseudomonas aeruginosa Isolates. Plants. 2022;11(15). [CrossRef]
- Neacsu IA, Leau SA, Marin S, et al. Collagen-carboxymethylcellulose biocomposite wound-dressings with antimicrobial activity. Materials. 2021;14(5):1-19. [CrossRef]
- Lepanto MS, Rosa L, Cutone A, et al. Bovine lactoferrin pre-treatment induces intracellular killing of AIEC LF82 and reduces bacteria-induced DNA damage in differentiated human enterocytes. Int J Mol Sci. 2019;20(22). [CrossRef]
- Mihai MM, Bălăceanu-Gurău B, Holban AM, et al. Promising Antimicrobial Activities of Essential Oils and Probiotic Strains on Chronic Wound Bacteria. Biomedicines. 2025;13(4). [CrossRef]
- Ilie CI, Oprea E, Geana EI, et al. Bee Pollen Extracts: Chemical Composition, Antioxidant Properties, and Effect on the Growth of Selected Probiotic and Pathogenic Bacteria. Antioxidants. 2022;11(5). [CrossRef]
- Cravioto A, Tello A, Navarro A, et al. Association of Escherichia coli HEp-2 adherence patterns with type and duration of diarrhoea. Lancet. 1991;337(8736):262-264. [CrossRef]
- Chifiriuc MC, Bleotu C, Pelinescu DR, et al. Patterns of colonization and immune response elicited from interactions between enteropathogenic bacteria, epithelial cells and probiotic fractions. Int J Biotechnol Mol Biol Res. 2010;1(4):47-57. http://www.academicjournals.org/IJBMBR.
- Scaletsky ICA, Pedroso MZ, Oliva CAG, Carvalho RLB, Morais MB, Fagundes-Neto U. A Localized Adherence-Like Pattern as a Second Pattern of Adherence of Classic Enteropathogenic Escherichia coli to HEp-2 Cells That Is Associated with Infantile Diarrhea. Infect Immun. 1999;67(7):3410-3415.
- Hasan TH, Al-Harmoosh RA. Mechanisms of antibiotics resistance in bacteria. Syst Rev Pharm. 2020;11(6):817-823. [CrossRef]
- de Sá Almeida JS, de Oliveira Marre AT, Teixeira FL, et al. Lactoferrin and lactoferricin B reduce adhesion and biofilm formation in the intestinal symbionts Bacteroides fragilis and Bacteroides thetaiotaomicron. Anaerobe. 2020;64. [CrossRef]
- Imoto I, Yasuma T, D’Alessandro-Gabazza CN, et al. Antimicrobial Effects of Lactoferrin against Helicobacter pylori Infection. Pathogens. 2023;12(4). [CrossRef]
- Bolscher JGM, Adão R, Nazmi K, et al. Bactericidal activity of LFchimera is stronger and less sensitive to ionic strength than its constituent lactoferricin and lactoferrampin peptides. Biochimie. 2009;91(1):123-132. [CrossRef]
- Ramamourthy G, Vogel HJ. Antibiofilm activity of lactoferrin-derived synthetic peptides against Pseudomonas aeruginosa PAO1. Biochem Cell Biol. 2021;99(1):138-148. [CrossRef]
- Mishra B, Felix LO, Basu A, et al. Design and Evaluation of Short Bovine Lactoferrin-Derived Antimicrobial Peptides against Multidrug-Resistant Enterococcus faecium. Antibiotics. 2022;11(8). [CrossRef]
- Yen CC, Shen CJ, Hsu WH, et al. Lactoferrin: An iron-binding antimicrobial protein against Escherichia coli infection. BioMetals. 2011;24(4):585-594. [CrossRef]
- Flores-Villaseñor H, Canizalez-Román A, Reyes-Lopez M, et al. Bactericidal effect of bovine lactoferrin, LFcin, LFampin and LFchimera on antibiotic-resistant Staphylococcus aureus and Escherichia coli. BioMetals. 2010;23:569-578. [CrossRef]
- Dashper SG, Pan Y, Veith PD, et al. Lactoferrin inhibits Porphyromonas gingivalis proteinases and has sustained biofilm inhibitory activity. Antimicrob Agents Chemother. 2012;56(3):1548-1556. [CrossRef]
- Kamiya H, Ehara T, Matsumoto T. Inhibitory effects of lactoferrin on biofilm formation in clinical isolates of Pseudomonas aeruginosa. J Infect Chemother. 2012;18(1):47-52. [CrossRef]
- Seraphina K, Furukido R, Isobe N, Nii T, Kurokawa Y, Suzuki N. Effect of lactoferrin on biofilm formation of bovine mastitis-causing Staphylococcus species. J Vet Med Sci. Published online 2025. [CrossRef]
- Wakabayashi H, Yamauchi K, Kobayashi T, Yaeshima T, Iwatsuki K, Yoshie H. Inhibitory effects of lactoferrin on growth and biofilm formation of Porphyromonas gingivalis and Prevotella intermedia. Antimicrob Agents Chemother. 2009;53(8):3308-3316. [CrossRef]
- Kawasaki Y, Tazume S, Shimizu K, et al. Inhibitory effects of bovine lactoferrin on the adherence of enterotoxigenic Escherichia coli to host cells. Biosci Biotechnol Biochem. 2000;64(2):348-354. [CrossRef]
- Lu J, Francis JD, Guevara MA, et al. Antibacterial and Anti-biofilm Activity of the Human Breast Milk Glycoprotein Lactoferrin against Group B Streptococcus. ChemBioChem. 2021;22(12):2124-2133. [CrossRef]
- Berlutti F, Superti F, Nicoletti M, et al. Bovine Lactoferrin Inhibits the Efficiency of Invasion of Respiratory A549 Cells of Different Iron-Regulated Morphological Forms of Pseudomonas Aeruginosa and Burkholderia Cenocepacia. Int J Immunopathol Pharmacol. 2008;21(1):51-59. [CrossRef]
- De Alberti D, Russo R, Terruzzi F, Nobile V, Ouwehand AC. Lactobacilli vaginal colonisation after oral consumption of Respecta®® complex: a randomised controlled pilot study. Arch Gynecol Obstet. 2015;292(4):861-867. [CrossRef]
- Russo R, Superti F, Karadja E, De Seta F. Randomised clinical trial in women with Recurrent Vulvovaginal Candidiasis: Efficacy of probiotics and lactoferrin as maintenance treatment. Mycoses. 2019;62(4):328-335. [CrossRef]
- Ulvatne H, Vorland LH, Uløatne H. Bactericidal Kinetics of 3 Lactoferricins Against Staphylococcus aureus and Escherichia coli. Scand J Infect Dis. 2001;33:507-511.
- Huertas Méndez NDJ, Vargas Casanova Y, Gómez Chimbi AK, et al. Synthetic Peptides Derived from Bovine Lactoferricin Exhibit Antimicrobial Activity against E. coli ATCC 11775, S. maltophilia ATCC 13636 and S. enteritidis ATCC 13076. Molecules. 2017;22(3). [CrossRef]
- Lazăr V, Bădăluță VA, Apetrei RM, Dițu LM. Staphylococcus aureus-A Quiet Human Body Resident and Sometimes a Feared Enemy. J Microbes Health Dis. 2025;1(1):100002.
- Rosa L, Cutone A, Lepanto MS, et al. Physico-chemical properties influence the functions and efficacy of commercial bovine lactoferrins. BioMetals. 2018;31(3):301-312. [CrossRef]
- Jugert CS, Didier A, Plötz M, Jessberger N. Strain-specific Antimicrobial Activity of Lactoferrin-based Food Supplements. J Food Prot. 2023;86(10). [CrossRef]
- Kosznik-Kwaśnicka K, Kaźmierczak N, Piechowicz L. Activity of Phage–Lactoferrin Mixture against Multi Drug Resistant Staphylococcus aureus Biofilms. Antibiotics. 2022;11(9). [CrossRef]
- Quintieri L, Caputo L, Monaci L, Cavalluzzi MM, Denora N. Lactoferrin-derived peptides as a control strategy against skinborne staphylococcal biofilms. Biomedicines. 2020;8(9). [CrossRef]
- Zheng W, Zhang Y, Lu HM, et al. Antimicrobial activity and safety evaluation of Enterococcus faecium KQ 2.6 isolated from peacock feces. BMC Biotechnol. 2015;15(1). [CrossRef]
- Pei J, Xiong L, Wu X, et al. Bovine lactoferricin exerts antibacterial activity against four Gram-negative pathogenic bacteria by transforming its molecular structure. Front Cell Infect Microbiol. 2025;15. [CrossRef]
- Shahidi F, Roshanak S, Javadmanesh A, Tabatabaei Yazdi F, Pirkhezranian Z, Azghandi M. Evaluation of antimicrobial properties of bovine lactoferrin against foodborne pathogenic microorganisms in planktonic and biofilm forms (in vitro). J Verbr Lebensm. 2020;15(3):277-283. [CrossRef]
- Elass-Rochard E, Roseanu A, Legrand D, et al. Lactoferrin-lipopolysaccharide interaction: Involvement of the 28-34 loop region of human lactoferrin in the high-affinity binding to Escherichia coli 055B5 lipopolysaccharide. Biochem J. 1995;312(3):839-845. [CrossRef]
- Naidu AS, Arnold RR. Influence of Lactoferrin on Host-Microbe Interactions. In: Hutchens TW, Lönnerdal B, eds. Lactoferrin: Interactions and Biological Functions. Humana Press; 1997. [CrossRef]
- Mao Y, Wang Y, Luo X, Chen X, Wang G. Impact of cell-free supernatant of lactic acid bacteria on Staphylococcus aureus biofilm and its metabolites. Front Vet Sci. 2023;10. [CrossRef]
- Aminnezhad S, Kermanshahi RK, Ranjbar R. Evaluation of synergistic interactions between cell-free supernatant of Lactobacillus strains and Amikacin and genetamicin against Pseudomonas aeruginosa. Jundishapur J Microbiol. 2015;8(4). [CrossRef]
- Kalaycı Yüksek F, Gümüş D, Gündoğan Gİ, Anğ Küçüker M. Cell-Free Lactobacillus sp Supernatants Modulate Staphylococcus aureus Growth, Adhesion and Invasion to Human Osteoblast (HOB) Cells. Curr Microbiol. 2021;78(1):125-132. [CrossRef]
- Keeratikunakorn K, Kaewchomphunuch T, Kaeoket K, Ngamwongsatit N. Antimicrobial activity of cell free supernatants from probiotics inhibits against pathogenic bacteria isolated from fresh boar semen. Sci Rep. 2023;13(1). [CrossRef]
- Fugaban JII, Bucheli JEV, Holzapfel WH, Todorov SD. Characterization of partially purified bacteriocins produced by enterococcus faecium strains isolated from soybean paste active against listeria spp. and vancomycin--resistant enterococci. Microorganisms. 2021;9(5). [CrossRef]
- Bertuccini L, Costanzo M, Iosi F, et al. Lactoferrin prevents invasion and inflammatory response following E. coli strain LF82 infection in experimental model of Crohn’s disease. Dig Liver Dis. 2014;46(6):496-504. [CrossRef]
- Andrews C, McLean MH, Durum SK. Cytokine tuning of intestinal epithelial function. Front Immunol. 2018;9(JUN). [CrossRef]
- Eckmann L, Jung HC, Schurer-Maly C, Panja A, Morzycka-Wroblewska E, Kagnoff MF. Differential Cytokine Expression by Human Intestinal Epithelial Cell Lines: Regulated Expression of lnterleukin 8. Gastroenterology. 1993;105(6):1689-1697. [CrossRef]
- Schuerer-Maly CC, Eckmann L, Kagnoff MF, Falco MT, Malyt FE. Colonic epithelial cell lines as a source of interleukin-8: stimulation by inflammatory cytokines and bacterial lipopolysaccharide. Immunology. 1994;81:85-91.
- Håversen L, Ohlsson BG, Hahn-Zoric M, Hanson LÅ, Mattsby-Baltzer I. Lactoferrin down-regulates the LPS-induced cytokine production in monocytic cells via NF-κB. Cell Immunol. 2002;220(2):83-95. [CrossRef]
- Berlutti F, Schippa S, Morea C, et al. Lactoferrin downregulates pro-inflammatory cytokines upexpressed in intestinal epithelial cells infected with invasive or noninvasive Escherichia coli strains. In: Biochemistry and Cell Biology. Vol 84. 2006:351-357. [CrossRef]
- Frioni A, Conte MP, Cutone A, et al. Lactoferrin differently modulates the inflammatory response in epithelial models mimicking human inflammatory and infectious diseases. BioMetals. 2014;27(5):843-856. [CrossRef]
- Tuo Y, Song X, Song Y, et al. Screening probiotics from Lactobacillus strains according to their abilities to inhibit pathogen adhesion and induction of pro-inflammatory cytokine IL-8. J Dairy Sci. 2018;101(6):4822-4829. [CrossRef]
- Ko JS, Yang HR, Chang Y, Seo JK, Seo K. Lactobacillus plantarum inhibits epithelial barrier dysfunction and interleukin-8 secretion induced by tumor necrosis factor-a rapid communication. World J Gastroenterol. 2007;13(13):1962-1965.
- Rocha-Ramírez LM, Hernández-Chiñas U, Moreno-Guerrero SS, Ramírez-Pacheco A, Eslava CA. Probiotic properties and immunomodulatory activity of lactobacillus strains isolated from dairy products. Microorganisms. 2021;9(4). [CrossRef]
- Kim WJ, Hyun JH, Lee NK, Paik HD. Protective Effects of a Novel Lactobacillus brevis Strain with Probiotic Characteristics against Staphylococcus aureus Lipoteichoic Acid-Induced Intestinal Inflammatory Response. J Microbiol Biotechnol. 2022;32(2):205-211. [CrossRef]
- Huang FC, Lu YT, Liao YH. Beneficial effect of probiotics on Pseudomonas aeruginosa–infected intestinal epithelial cells through inflammatory IL-8 and antimicrobial peptide human beta-defensin-2 modulation. Innate Immun. 2020;26(7):592-600. [CrossRef]
- Devi MB, Bhattacharya A, Kumar A, et al. Potential probiotic Lactiplantibacillus plantarum strains alleviate TNF-α by regulating ADAM17 protein and ameliorate gut integrity through tight junction protein expression in in vitro model. Cell Commun Signal. 2024;22(1). [CrossRef]
- Han X, Ding S, Ma Y, et al. Lactobacillus plantarum and Lactobacillus brevis Alleviate Intestinal Inflammation and Microbial Disorder Induced by ETEC in a Murine Model. Oxid Med Cell Longev. Published online September 21, 2021:6867962. [CrossRef]
- Iordache F, Iordache C, Chifiriuc MC, et al. Antimicrobial and immunomodulatory activity of some probiotic fractions with potential clinical application. Arch Zootech. 2008;11(3):41-51.
- De Marco S, Sichetti M, Muradyan D, et al. Probiotic cell-free supernatants exhibited anti-inflammatory and antioxidant activity on human gut epithelial cells and macrophages stimulated with LPS. Evid Based Complement Alternat Med. 2018;2018. [CrossRef]
- Zanetta P, Ballacchino C, Squarzanti DF, Amoruso A, Pane M, Azzimonti B. Lactobacillus johnsonii LJO02 (DSM 33828) Cell-Free Supernatant and Vitamin D Improve Wound Healing and Reduce Interleukin-6 Production in Staphylococcus aureus-Infected Human Keratinocytes. Pharmaceutics. 2024;16(1). [CrossRef]
- Bianchi M, Kaya E, Logiudice V, et al. Biotherapeutic potential of different fractions of cell-free supernatants from Lacticaseibacillus rhamnosus against Pseudomonas aeruginosa. Front Cell Infect Microbiol. 2025;15. [CrossRef]
- Lee J, Kim S, Kang CH. Immunostimulatory Activity of Lactic Acid Bacteria Cell-Free Supernatants through the Activation of NF-κB and MAPK Signaling Pathways in RAW 264.7 Cells. Microorganisms. 2022;10(11). [CrossRef]
- Wu Q, Liu MC, Yang J, Wang JF, Zhu YH. Lactobacillus rhamnosus GR-1 ameliorates Escherichia coli-induced inflammation and cell damage via attenuation of ASC-independent NLRP3 inflammasome activation. Appl Environ Microbiol. 2016;82(4):1173-1182. [CrossRef]
- Mohammedsaeed W, McBain AJ, Cruickshank SM, O’Neill CA. Lactobacillus rhamnosus GG inhibits the toxic effects of Staphylococcus aureus on epidermal keratinocytes. Appl Environ Microbiol. 2014;80(18):5773-5781. [CrossRef]
- Spacova I, O’Neill C, Lebeer S. Lacticaseibacillus rhamnosus GG inhibits infection of human keratinocytes by Staphylococcus aureus through mechanisms involving cell surface molecules and pH reduction. Benef Microbes. 2020;11(7):703-715. [CrossRef]
- Zawistowska-Rojek A, Kośmider A, Stępień K, Tyski S. Adhesion and aggregation properties of Lactobacillaceae strains as protection ways against enteropathogenic bacteria. Arch Microbiol. 2022;204(5). [CrossRef]
- Lizardo M, Magalhães RM, Tavaria FK. Probiotic Adhesion to Skin Keratinocytes and Underlying Mechanisms. Biology. 2022;11(9). [CrossRef]
- Batoni G, Kaya E, Catelli E, et al. Lactobacillus Probiotic Strains Differ in Their Ability to Adhere to Human Lung Epithelial Cells and to Prevent Adhesion of Clinical Isolates of Pseudomonas aeruginosa from Cystic Fibrosis Lung. Microorganisms. 2023;11(7). [CrossRef]
- Dhanani AS, Bagchi T. Lactobacillus plantarum CS24.2 prevents Escherichia coli adhesion to HT-29 cells and also down-regulates enteropathogen-induced tumor necrosis factor-α and interleukin-8 expression. Microbiol Immunol. 2013;57(4):309-315. [CrossRef]
- Michail S, Abernathy F. Lactobacillus plantarum Inhibits the Intestinal Epithelial Migration of Neutrophils Induced by Enteropathogenic Escherichia coli. J Pediatr Gastroenterol Nutr. 2003;36(3):385-391. [CrossRef]
- Wang J, Zeng Y, Wang S, et al. Swine-derived probiotic Lactobacillus plantarum inhibits growth and adhesion of enterotoxigenic Escherichia coli and mediates host defense. Front Microbiol. 2018;9(JUN). [CrossRef]
- Alizadeh Behbahani B, Noshad M, Falah F. Inhibition of Escherichia coli adhesion to human intestinal Caco-2 cells by probiotic candidate Lactobacillus plantarum strain L15. Microb Pathog. 2019;136. [CrossRef]
- Kiousi DE, Panopoulou M, Pappa A, Galanis A. Lactobacilli-host interactions inhibit Staphylococcus aureus and Escherichia coli-induced cell death and invasion in a cellular model of infection. Front Microbiol. 2024;15. [CrossRef]
- Khodaii Z, Ghaderian SMH, Mehrabani MN. Probiotic Bacteria and their Supernatants Protect Enterocyte Cell Lines from Enteroinvasive Escherichia coli (EIEC) Invasion. Int J Mol Cell Med. 2017;6(3):183-189. [CrossRef]
- Pecoraro C, Carbone D, Parrino B, Cascioferro S, Diana P. Recent Developments in the Inhibition of Bacterial Adhesion as Promising Anti-Virulence Strategy. Int J Mol Sci. 2023;24(5). [CrossRef]
- Vuopio-Varkila J, Schoolnik GK. Localized Adherence by Enteropathogenic Escherichia coil Is an Inducible Phenotype Associated with the Expression of New Outer Membrane Proteins. J Exp Med. 1991;174:1167-1177.
- Nougayrède JP, Fernandes PJ, Donnenberg MS. Adhesion of enteropathogenic Escherichia coli to host cells. Cell Microbiol. 2003;5(6):359-372. [CrossRef]
- Dehbanipour R, Ghalavand Z. Anti-virulence therapeutic strategies against bacterial infections: recent advances. Germs. 2022;12(2):262-275. [CrossRef]
- Russo R, Karadja E, De Seta F. Evidence-based mixture containing Lactobacillus strains and lactoferrin to prevent recurrent bacterial vaginosis: A double blind, placebo controlled, randomised clinical trial. Benef Microbes. 2019;10(1):19-26. [CrossRef]
- Russo R, Edu A, De Seta F. Study on the effects of an oral lactobacilli and lactoferrin complex in women with intermediate vaginal microbiota. Arch Gynecol Obstet. 2018;298(1):139-145. [CrossRef]
Figure 1.
The antimicrobial effects of Lf (generated with BioRender.com, accessed on October 17 2025).
Figure 1.
The antimicrobial effects of Lf (generated with BioRender.com, accessed on October 17 2025).
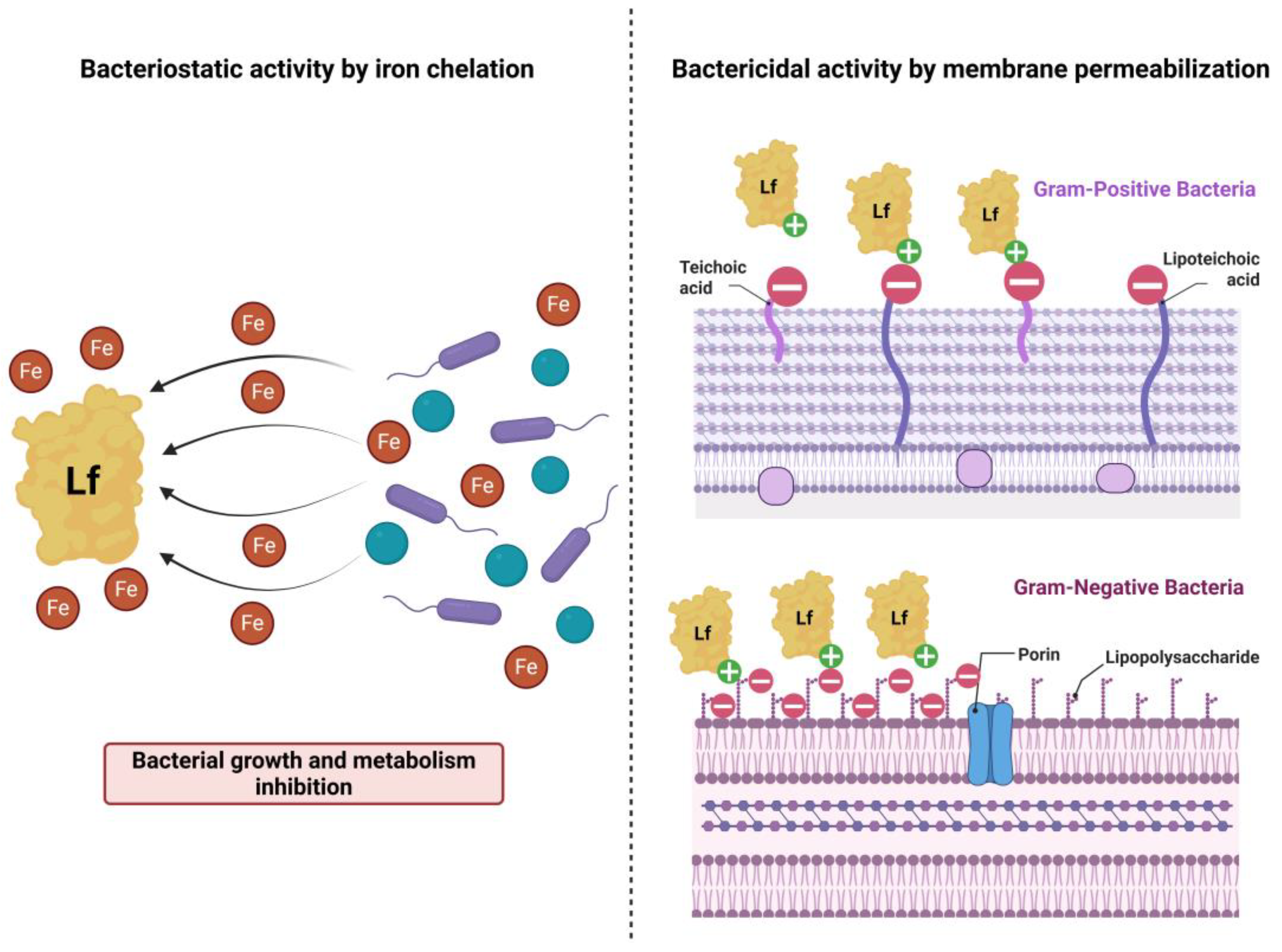
Figure 2.
Microscopic images of the biofilms formed by the Gram-positive strains (S. aureus and E. faecium) treated with bLf, commercial Lf (cLf), gentamicin (Ge) and the growth control (GC) (Crystal violet staining, 200x). The untreated bacteria (GC) generated more compact biofilms, as opposed to the dispersed biofilms resulting from the treatments.
Figure 2.
Microscopic images of the biofilms formed by the Gram-positive strains (S. aureus and E. faecium) treated with bLf, commercial Lf (cLf), gentamicin (Ge) and the growth control (GC) (Crystal violet staining, 200x). The untreated bacteria (GC) generated more compact biofilms, as opposed to the dispersed biofilms resulting from the treatments.
Figure 3.
Microscopic images of the biofilms formed by the Gram-negative strains (E. coli ATCC 25922 and P. aeruginosa ATCC 27853) treated with bLf, commercial Lf (cLf), gentamicin (Ge) and the growth control (GC) (Crystal violet staining, 200x). Regarding P. aeruginosa ATCC 27853, the untreated bacteria (GC) generated more compact biofilms, as opposed to the less dense biofilms resulting from the treatments. The biofilms produced by E. coli ATCC 25922 were generally dispersed in all conditions, with minor differences between treatment settings.
Figure 3.
Microscopic images of the biofilms formed by the Gram-negative strains (E. coli ATCC 25922 and P. aeruginosa ATCC 27853) treated with bLf, commercial Lf (cLf), gentamicin (Ge) and the growth control (GC) (Crystal violet staining, 200x). Regarding P. aeruginosa ATCC 27853, the untreated bacteria (GC) generated more compact biofilms, as opposed to the less dense biofilms resulting from the treatments. The biofilms produced by E. coli ATCC 25922 were generally dispersed in all conditions, with minor differences between treatment settings.
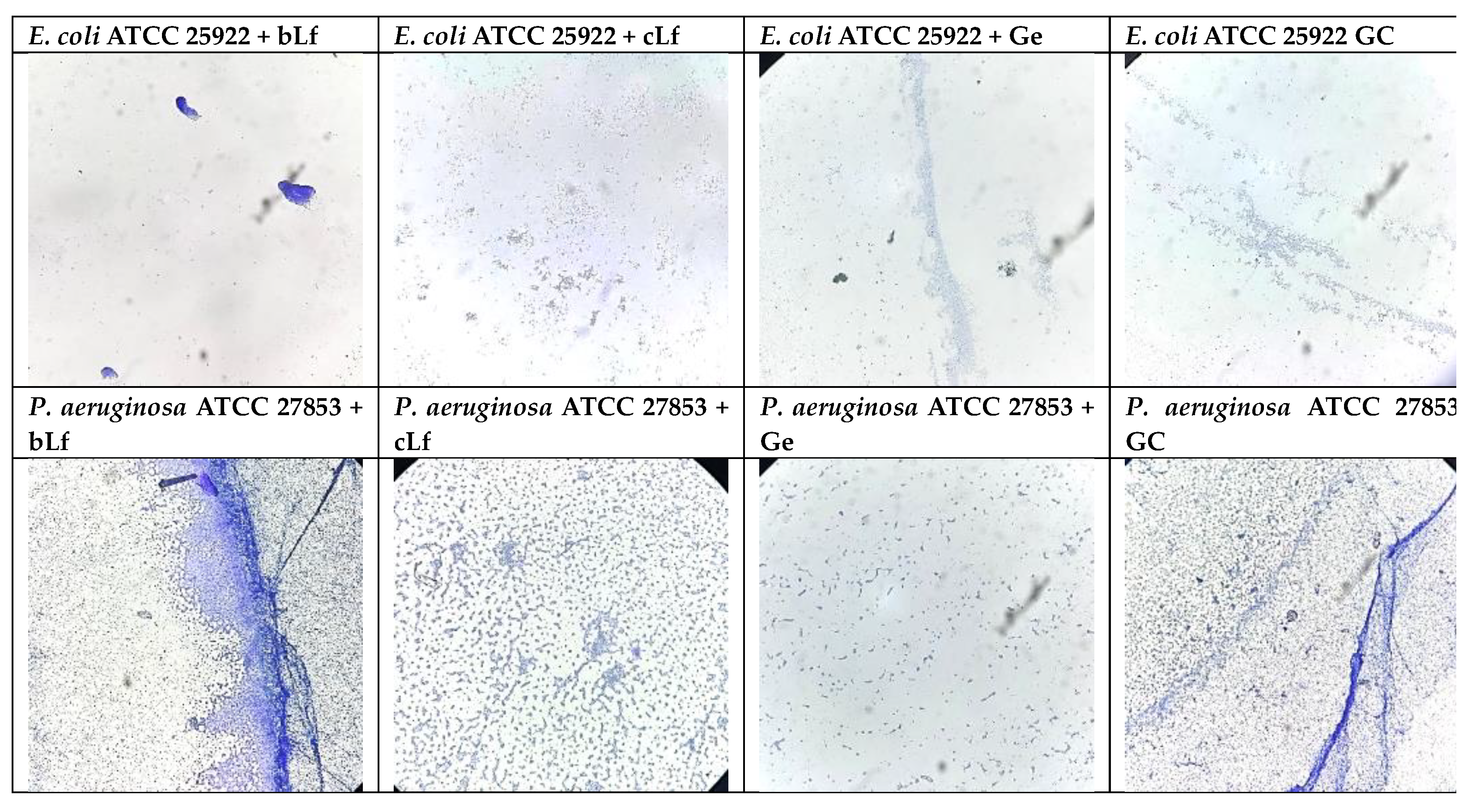
Figure 4.
Microscopic images of the biofilms generated by the S. aureus ATCC 25923 strain treated with LAB supernatants, alone and combined with commercial Lf (Crystal violet staining, 200x). Compared to the dense biofilms observed in the MRS and MRS + cLf wells, the addition of LAB CFS effectively dissociated microbial biofilms.
Figure 4.
Microscopic images of the biofilms generated by the S. aureus ATCC 25923 strain treated with LAB supernatants, alone and combined with commercial Lf (Crystal violet staining, 200x). Compared to the dense biofilms observed in the MRS and MRS + cLf wells, the addition of LAB CFS effectively dissociated microbial biofilms.
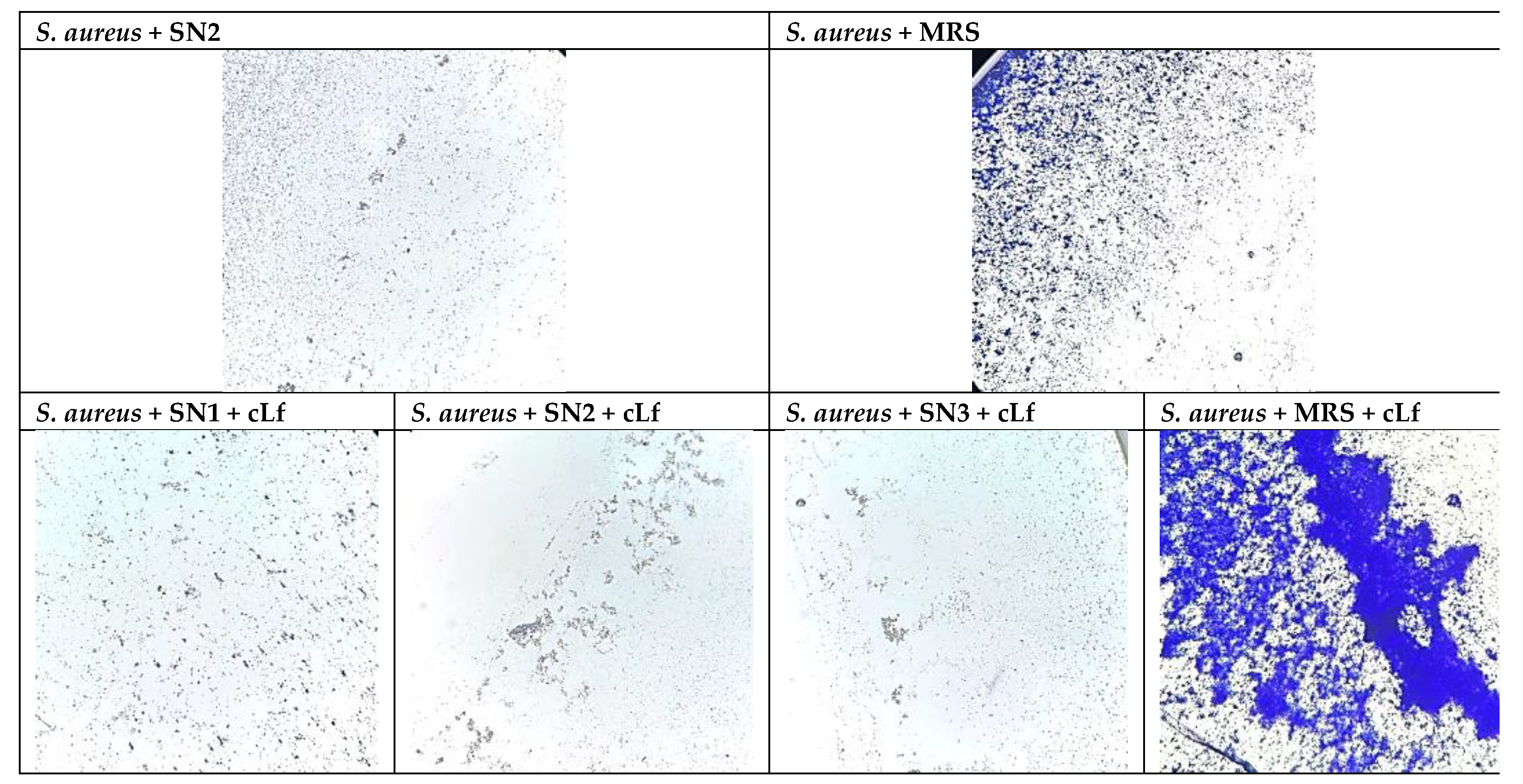
Figure 5.
Microscopic images of the biofilms generated by the E. faecium strain treated with LAB supernatants, alone and combined with commercial Lf (Crystal violet staining, 200x). The biofilms produced by E. faecium were generally dispersed in most of the experimental conditions, with minor differences between each treatment.
Figure 5.
Microscopic images of the biofilms generated by the E. faecium strain treated with LAB supernatants, alone and combined with commercial Lf (Crystal violet staining, 200x). The biofilms produced by E. faecium were generally dispersed in most of the experimental conditions, with minor differences between each treatment.
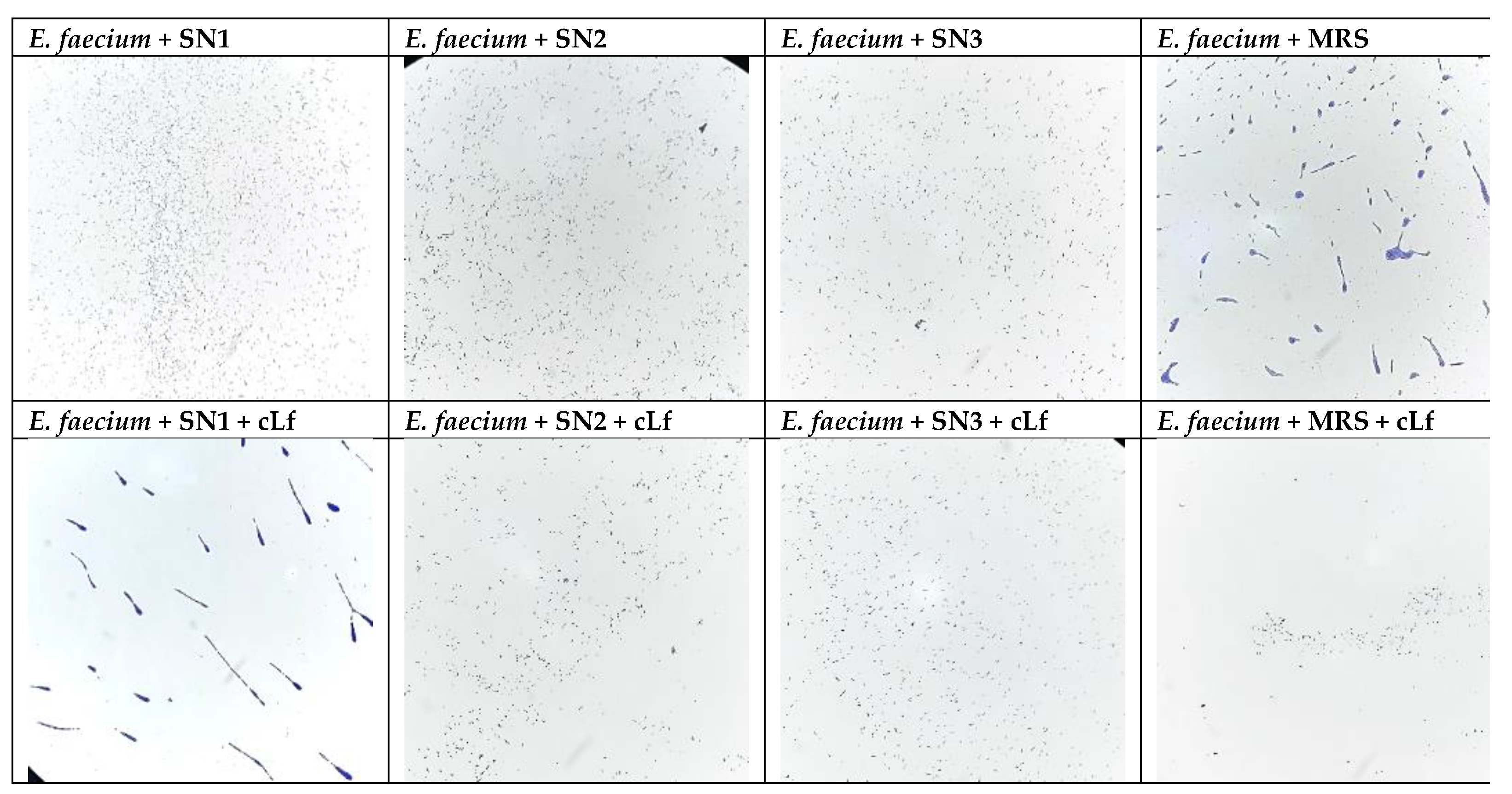
Figure 6.
Graphs illustrating the absolute values of IL-8 levels in HCT-8 cells treated with Lf and/or LAB supernatants and infected with (a) Gram-negative pathogens and (b) Gram-positive pathogens. Statistical significance was not obtained for tested variants pairs due to a low sample sizes (n = 2 for the experimental variants and n = 1 for the control variants).
Figure 6.
Graphs illustrating the absolute values of IL-8 levels in HCT-8 cells treated with Lf and/or LAB supernatants and infected with (a) Gram-negative pathogens and (b) Gram-positive pathogens. Statistical significance was not obtained for tested variants pairs due to a low sample sizes (n = 2 for the experimental variants and n = 1 for the control variants).
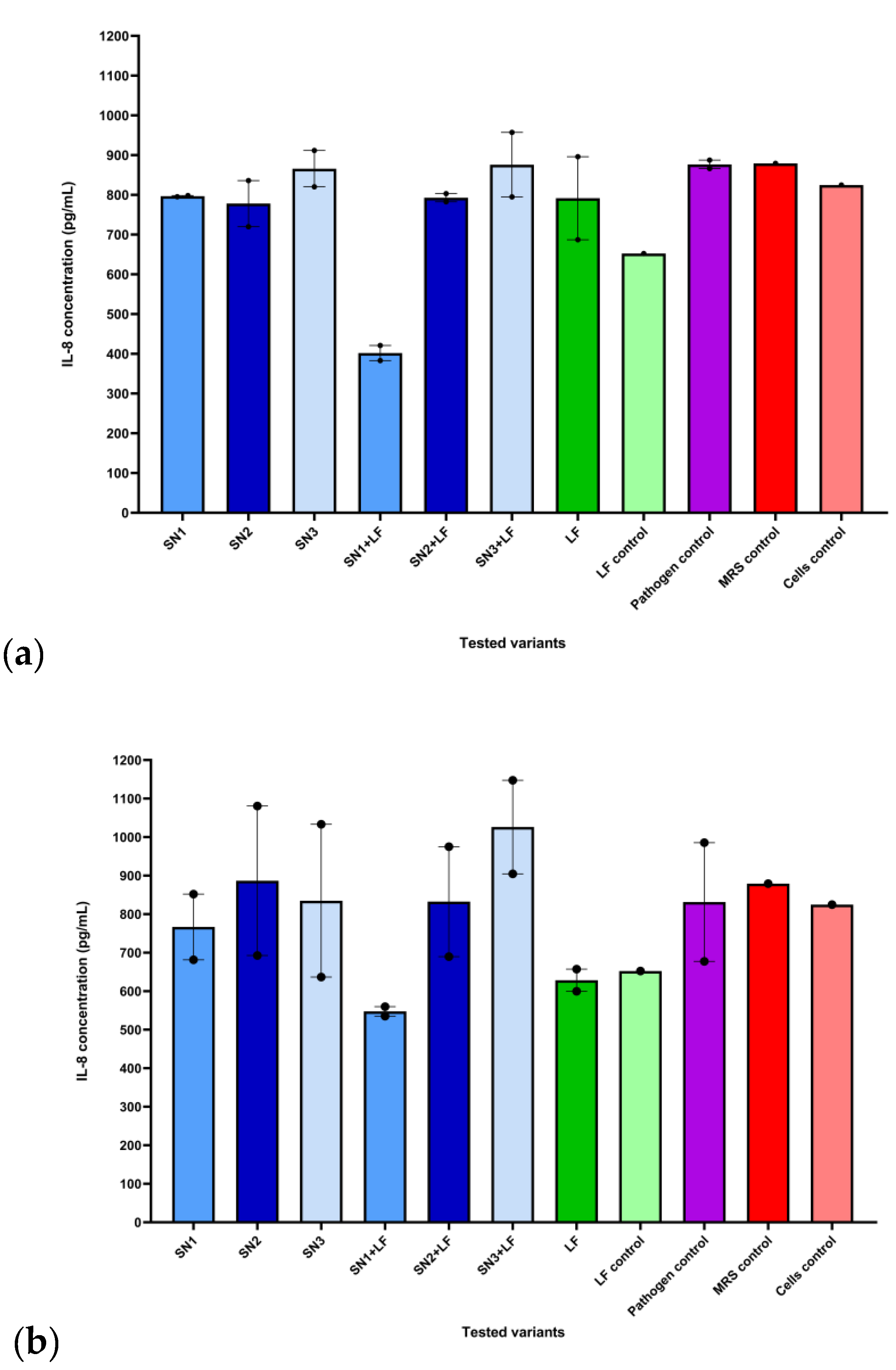
Figure 7.
Graph illustrating the variant effect values of IL-8 levels in HCT-8 cells treated with Lf and/or LAB supernatants and infected with Gram-negative pathogens (* p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001). Sample sizes for the calculated effects of tested variants range between n = 4 (SNx + LF groups) and n = 10 (LF).
Figure 7.
Graph illustrating the variant effect values of IL-8 levels in HCT-8 cells treated with Lf and/or LAB supernatants and infected with Gram-negative pathogens (* p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001). Sample sizes for the calculated effects of tested variants range between n = 4 (SNx + LF groups) and n = 10 (LF).
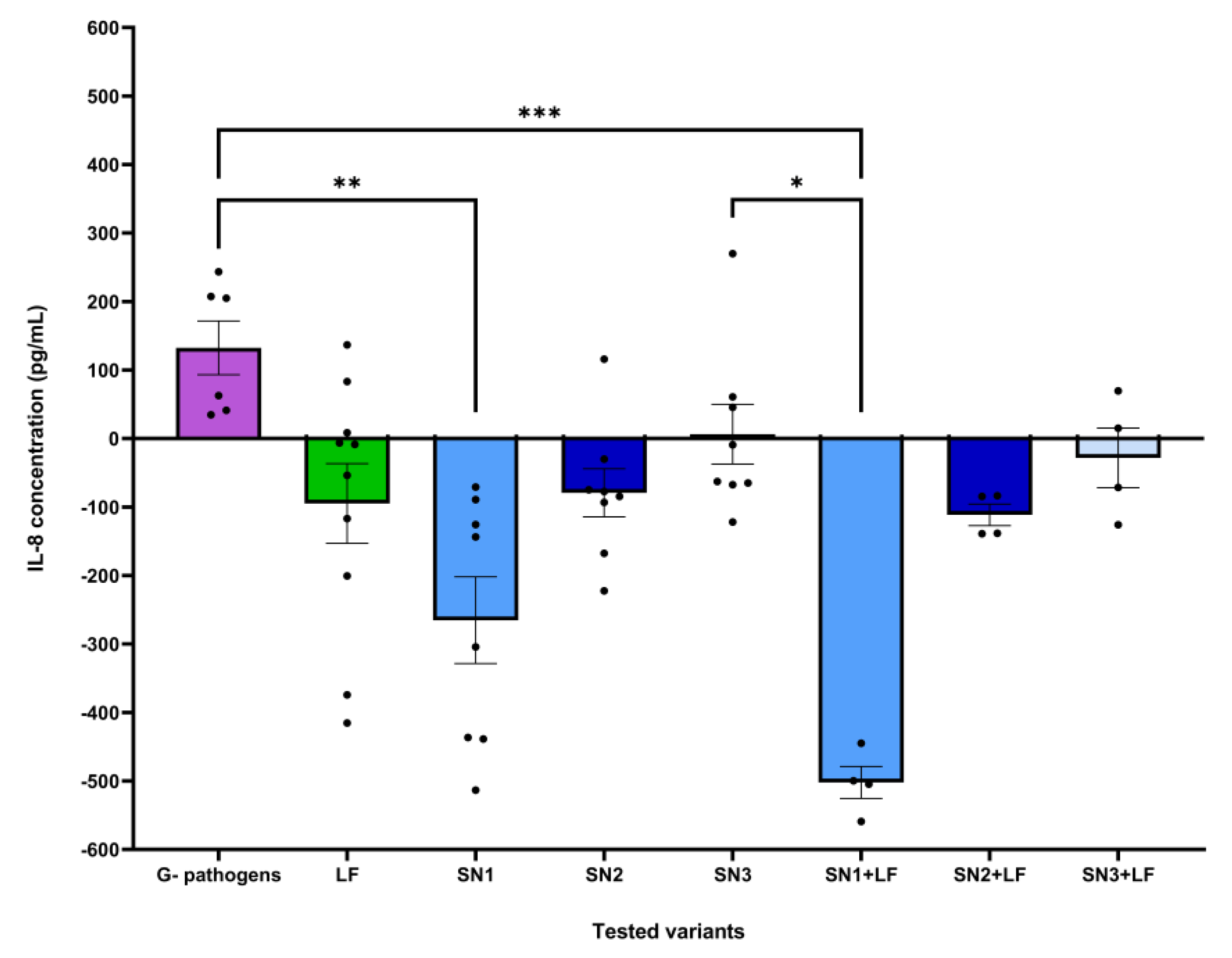
Figure 8.
Graph illustrating the variant effect values of IL-8 levels in HCT-8 cells treated with Lf and/or LAB supernatants and infected with Gram-positive pathogens (* p ≤ 0.05). Sample sizes for the calculated effects of tested variants range between n = 4 (SNx + LF groups) and n = 10 (LF).
Figure 8.
Graph illustrating the variant effect values of IL-8 levels in HCT-8 cells treated with Lf and/or LAB supernatants and infected with Gram-positive pathogens (* p ≤ 0.05). Sample sizes for the calculated effects of tested variants range between n = 4 (SNx + LF groups) and n = 10 (LF).
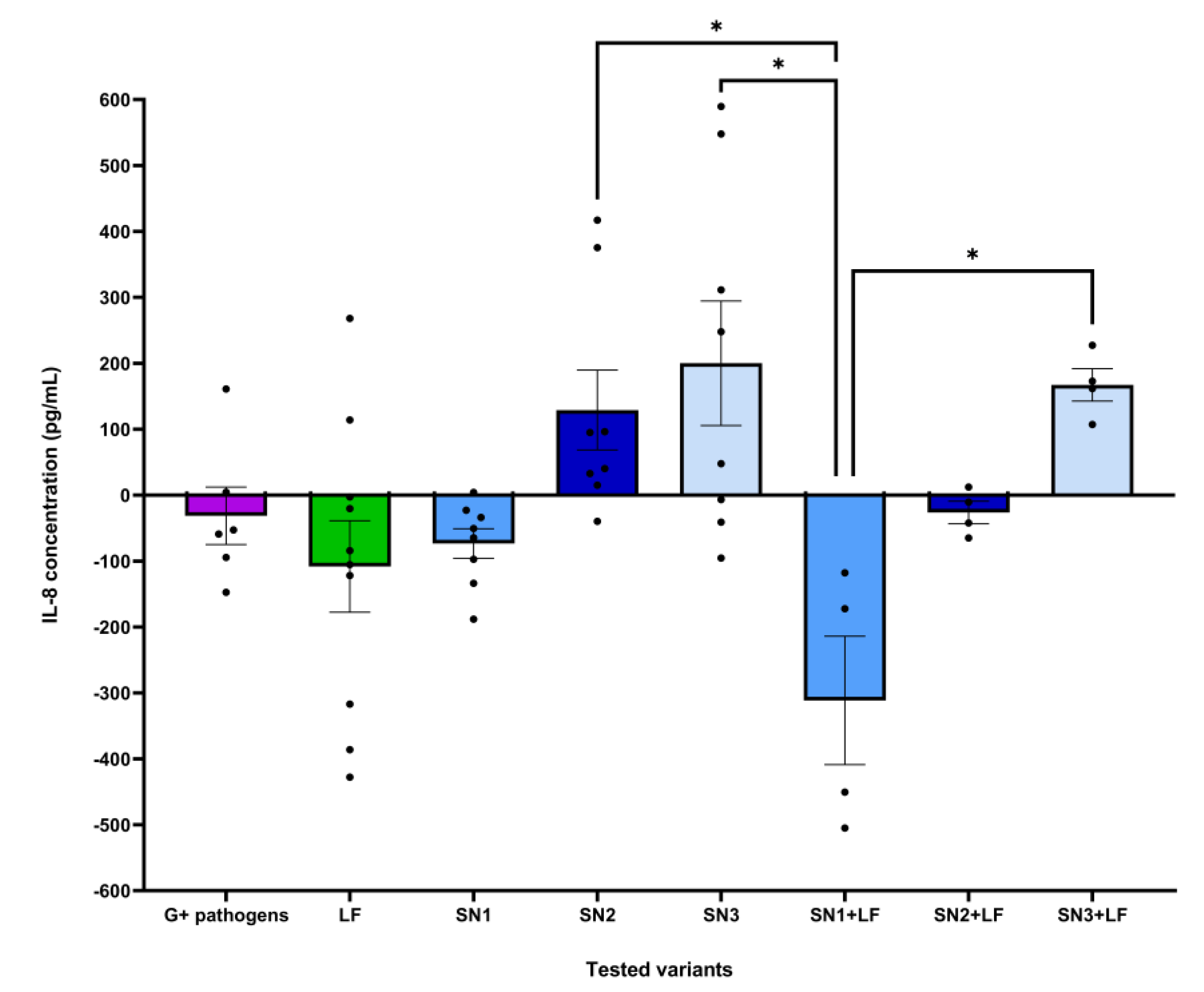
Figure 9.
Graphs illustrating the absolute values of IL-10 levels in HCT-8 cells treated with Lf and/or LAB supernatants and infected with (a) Gram-negative pathogens and (b) Gram-positive pathogens. Statistical significance was not obtained for tested variants pairs due to a low sample sizes (n = 2 for the experimental variants and n = 1 for the control variants).
Figure 9.
Graphs illustrating the absolute values of IL-10 levels in HCT-8 cells treated with Lf and/or LAB supernatants and infected with (a) Gram-negative pathogens and (b) Gram-positive pathogens. Statistical significance was not obtained for tested variants pairs due to a low sample sizes (n = 2 for the experimental variants and n = 1 for the control variants).
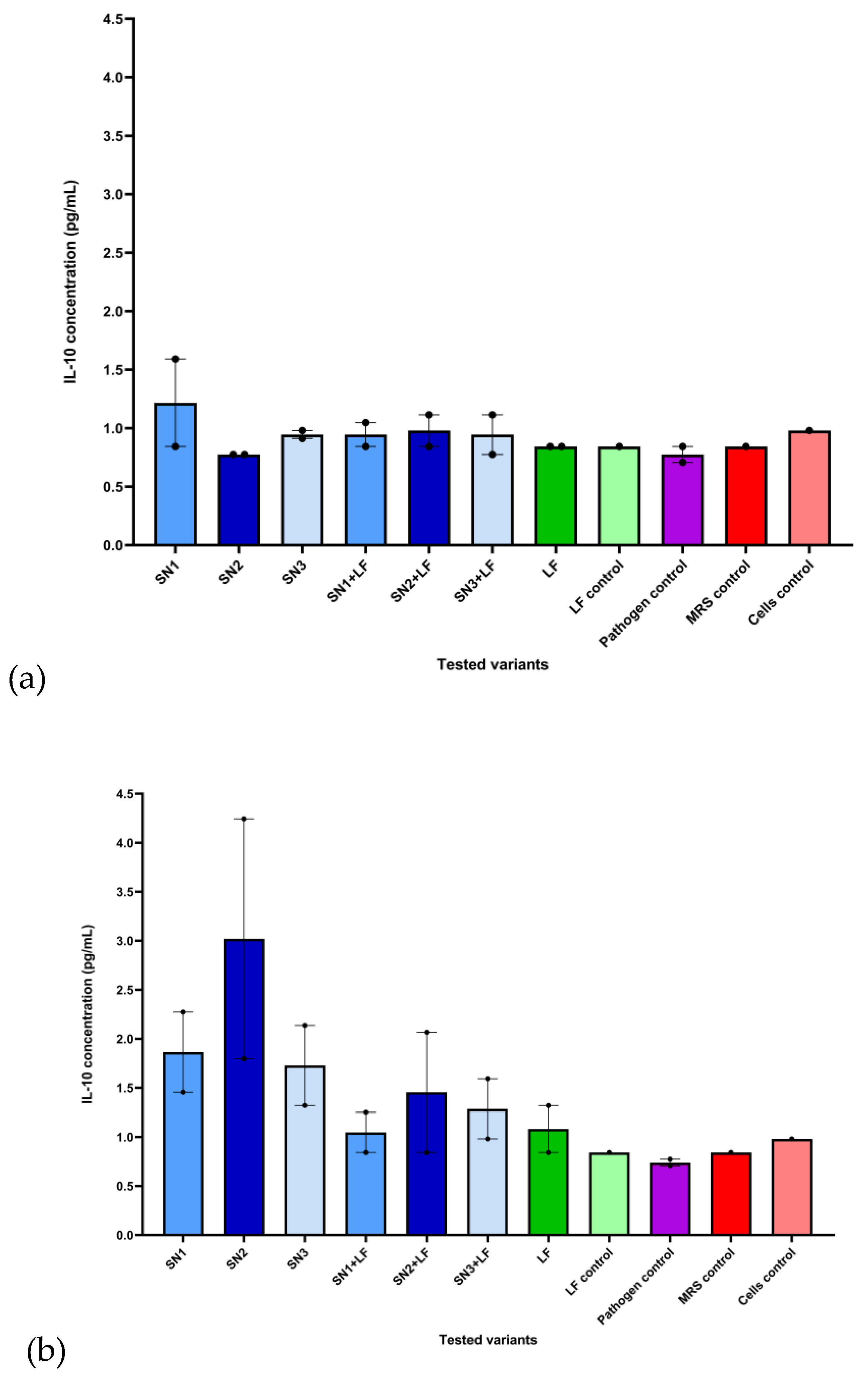
Figure 10.
Graph illustrating the variant effect values of IL-10 levels in HCT-8 cells treated with Lf and/or LAB supernatants and infected with Gram-negative pathogens (* p ≤ 0.05). Sample sizes for the calculated effects of tested variants range between n = 4 (SNx + LF groups) and n = 10 (LF).
Figure 10.
Graph illustrating the variant effect values of IL-10 levels in HCT-8 cells treated with Lf and/or LAB supernatants and infected with Gram-negative pathogens (* p ≤ 0.05). Sample sizes for the calculated effects of tested variants range between n = 4 (SNx + LF groups) and n = 10 (LF).
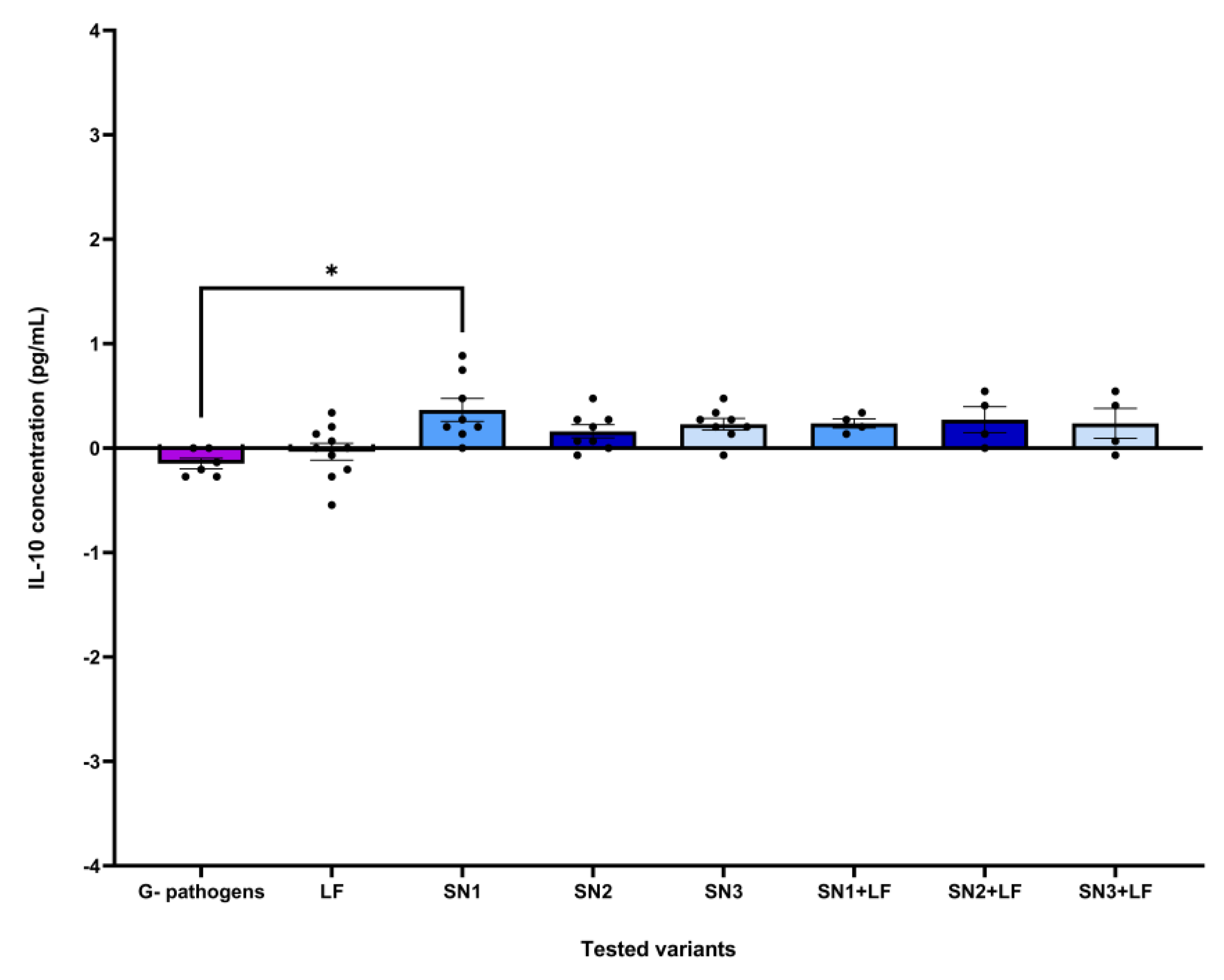
Figure 11.
Graph illustrating the variant effect values of IL-10 levels in HCT-8 cells treated with Lf and/or LAB supernatants and infected with Gram-positive pathogens (* p ≤ 0.05). Sample sizes for the calculated effects of tested variants range between n = 4 (SNx + LF groups) and n = 10 (LF).
Figure 11.
Graph illustrating the variant effect values of IL-10 levels in HCT-8 cells treated with Lf and/or LAB supernatants and infected with Gram-positive pathogens (* p ≤ 0.05). Sample sizes for the calculated effects of tested variants range between n = 4 (SNx + LF groups) and n = 10 (LF).
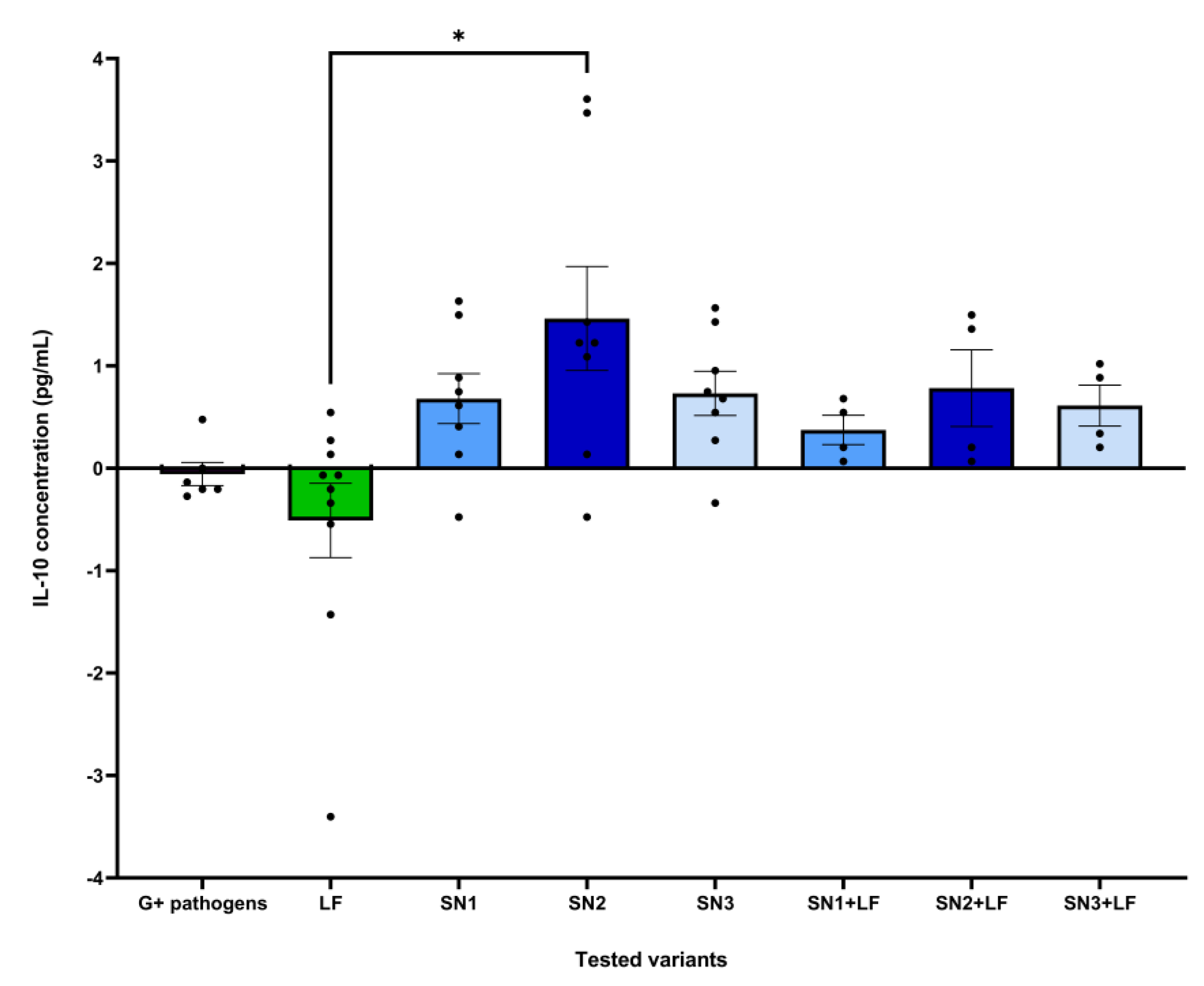
Figure 12.
Graphs illustrating the Adhesion Index (AI%) of (a) P. aeruginosa ATCC 27853, (b) E. coli ATCC 25922 and (c) S. aureus ATCC 25923 to cellular substrates induced by LAB supernatants, alone and associated with commercial Lf (cLf) (* p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001).
Figure 12.
Graphs illustrating the Adhesion Index (AI%) of (a) P. aeruginosa ATCC 27853, (b) E. coli ATCC 25922 and (c) S. aureus ATCC 25923 to cellular substrates induced by LAB supernatants, alone and associated with commercial Lf (cLf) (* p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001).
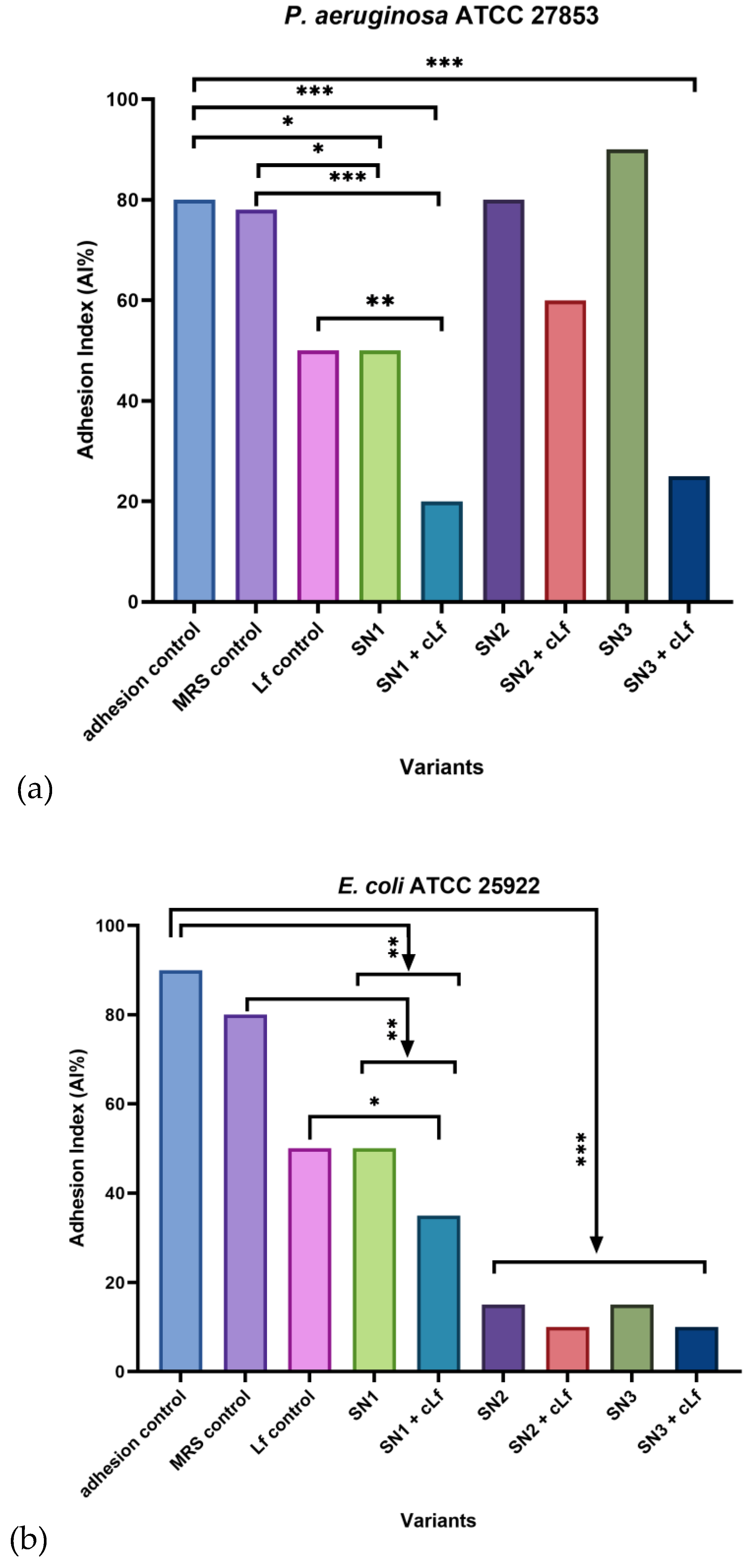
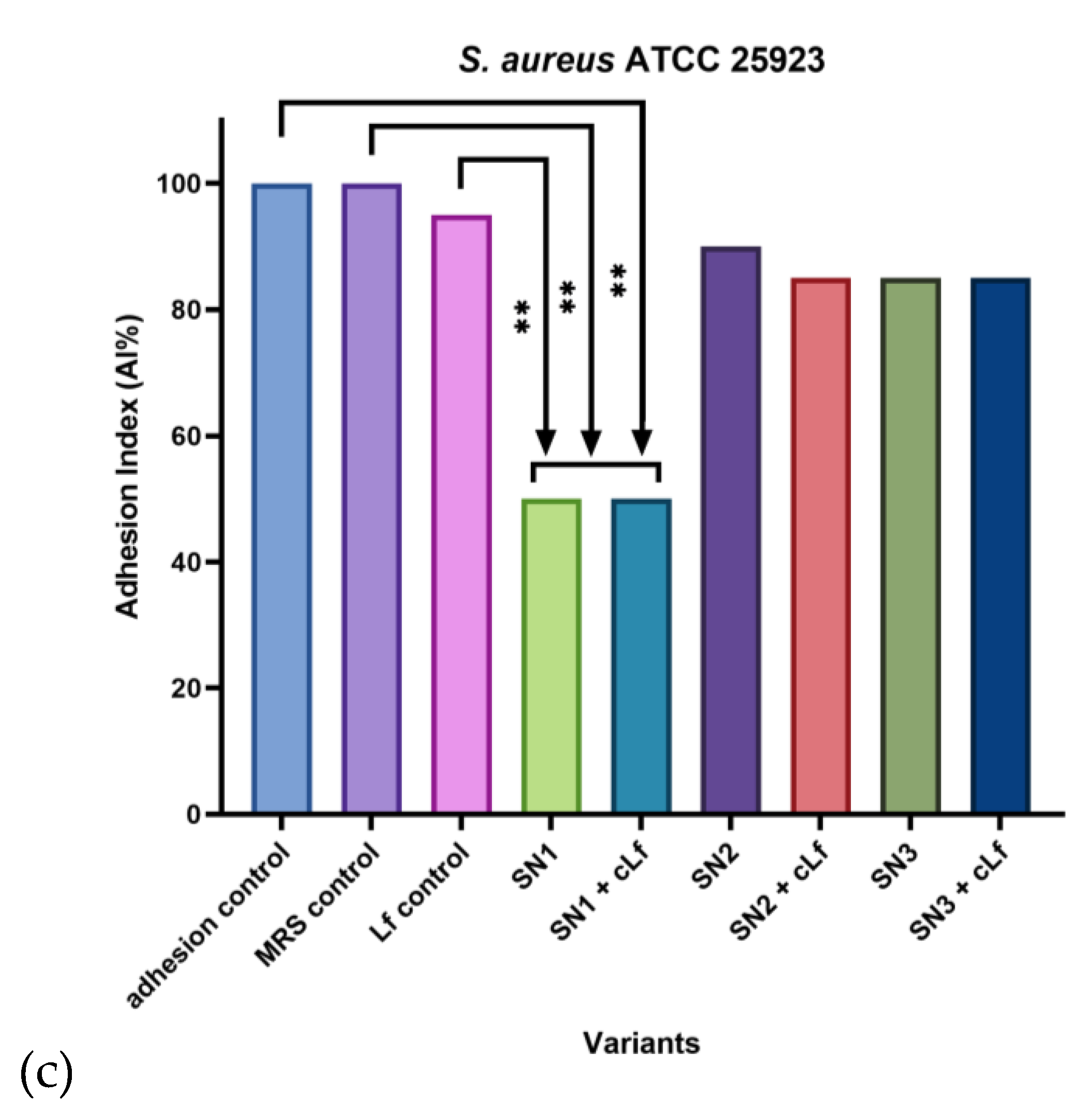
Figure 13.
Adhesion patterns of P. aeruginosa ATCC 27853 to cellular substrates in the presence of tested combinations and in controls (Giemsa staining, 1000x).
Figure 13.
Adhesion patterns of P. aeruginosa ATCC 27853 to cellular substrates in the presence of tested combinations and in controls (Giemsa staining, 1000x).
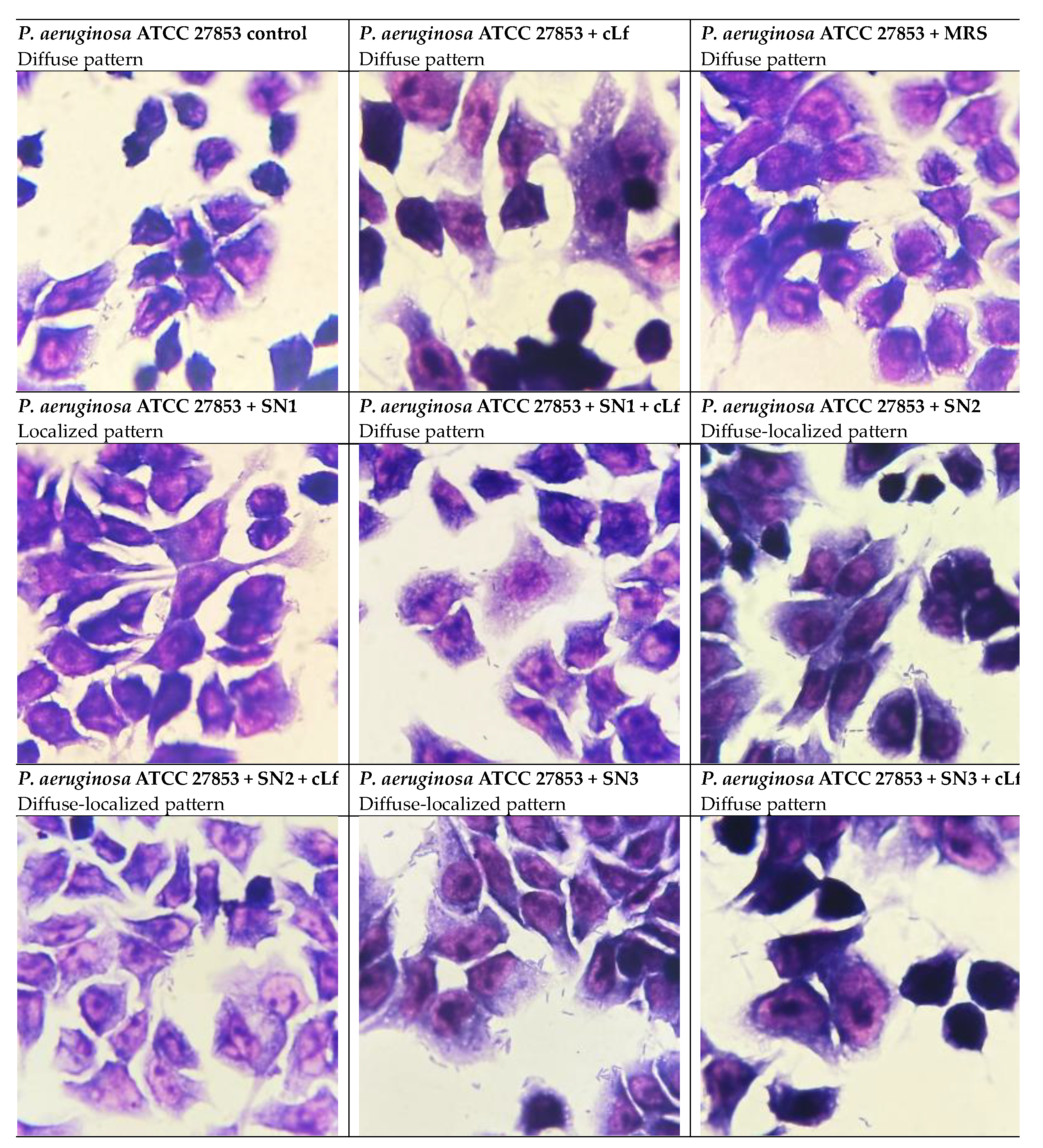

Table 1.
Procedures to obtain the tested compounds and their stock solutions.
| Compound | Stock solution(s) | Procedure |
|---|---|---|
| bLf 10 mg (Sigma-Aldrich, St. Louis, Missouri, USA) |
(1) 1 mg/mL in distilled water; (2) 2 mg/mL in nutrient broth. |
- |
| commercial Lf 250 mg (Jarrow Formulas, Los Angeles, California, USA) |
(1) 5 mg/mL in nutrient broth; (2) 25 mg/mL in Phosphate Buffered Saline (PBS); (3) 25 mg/mL in liquid Man-Rogosa-Sharpe (MRS) media; (4) 25 mg/mL in LAB supernatants. |
- |
| gentamicin-sulphate 1 g (Sigma-Aldrich, St. Louis, Missouri, USA) | 1 mg/mL in distilled water | - |
|
Cell-Free Supernatants (CFS) derived from LAB cultures (Lactobacillus rhamnosus MF9 from newborn feces; Lb. plantarum F, Lb. brevis 10 from fermented foods) |
- |
(1) obtaining fresh LAB cultures by transferring 100 µL from previously cultured pro-biotic strains into 10 mL of liquid MRS media in Eppendorf falcons (Eppendorf, Hamburg, Germany), followed by incubation at 37oC, for 24h. (2) placing of 2 mL of each Lactobacillus culture in two Eppendorf tubes, pursued by centrifugation for 5 min at 6000 rpm. (3) filtration of LAB supernatants with 5 mL syringes Injekt®® (B. Braun, Melsungen, Germany) and ReliaPrepTM 0.2 µm filters (Ahlstrom-Munksjö, Helsinki, Finland) in order to remove any contaminants and bacterial cells. (4) acidity buffering of LAB supernatants (using a NaOH 10% solution), bringing the pH to a neutral value: 7-7,5 [36] |
Table 2.
MIC and MBEC values for bLf versus commercial Lf (cLf) compared to gentamicin (Ge), for the studied Gram-positive strains (S. aureus ATCC 25923 and E. faecium).
Table 2.
MIC and MBEC values for bLf versus commercial Lf (cLf) compared to gentamicin (Ge), for the studied Gram-positive strains (S. aureus ATCC 25923 and E. faecium).
| Tested variant | MIC (µg/mL) | MBEC (µg/mL) |
|---|---|---|
| S. aureus ATCC 25923 + cLf | 5000 | 1250 |
| S. aureus ATCC 25923 + bLf | 2000 | 500 |
| S. aureus ATCC 25923 + Ge | 4 | 4 |
| E. faecium + cLf | 5000 | 1250 |
| E. faecium + bLf | 500 | 250 |
| E. faecium + Ge | 25 | 12.5 |
Table 3.
MIC and MBEC values for bLf versus commercial Lf (cLf) compared to gentamicin (Ge), for the studied Gram-negative strains (E. coli ATCC 25922 and P. aeruginosa ATCC 27853).
Table 3.
MIC and MBEC values for bLf versus commercial Lf (cLf) compared to gentamicin (Ge), for the studied Gram-negative strains (E. coli ATCC 25922 and P. aeruginosa ATCC 27853).
| Tested variant | MIC (µg/mL) | MBEC (µg/mL) |
|---|---|---|
| E. coli ATCC 25922+ cLf | 5000 | 5000 |
| E. coli ATCC 25922+ bLf | 2000 | 2000 |
| E. coli ATCC 25922+ Ge | 4 | 4 |
| P. aeruginosa ATCC 27853 + cLf | 5000 | 1250 |
| P. aeruginosa ATCC 27853 + bLf | 2000 | 62.5 |
| P. aeruginosa ATCC 27853 + Ge | 25 | 25 |
Table 4.
MBEC values of LAB supernatants (SN), alone and associated with commercial Lf (cLf) obtained for the S. aureus ATCC 25923 strain.
Table 4.
MBEC values of LAB supernatants (SN), alone and associated with commercial Lf (cLf) obtained for the S. aureus ATCC 25923 strain.
| Tested variant | SN (%) | cLf (µg/mL) |
|---|---|---|
| SN2 | 50 | - |
| MRS control | 50 | - |
| SN1 + cLf | 6.25 | 0.31 |
| SN2 + cLf | 6.25 | 0.31 |
| SN3 + cLf | 1.56 | 0.08 |
| MRS + cLf control | 50 | 2.5 |
Table 5.
MBEC values of LAB supernatants (SN), alone and associated with commercial Lf (cLf) obtained for the E. faecium strain.
Table 5.
MBEC values of LAB supernatants (SN), alone and associated with commercial Lf (cLf) obtained for the E. faecium strain.
| Tested variant | SN (%) | cLf (µg/mL) |
|---|---|---|
| SN1 | 0.2 | - |
| SN2 | 0.2 | - |
| SN3 | 0.39 | - |
| MRS control | 0.39 | - |
| SN1 + cLf | 1.56 | 0.08 |
| SN2 + cLf | 1.56 | 0.08 |
| SN3 + cLf | 1.56 | 0.08 |
| MRS + cLf control | 6.25 | 0.31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



