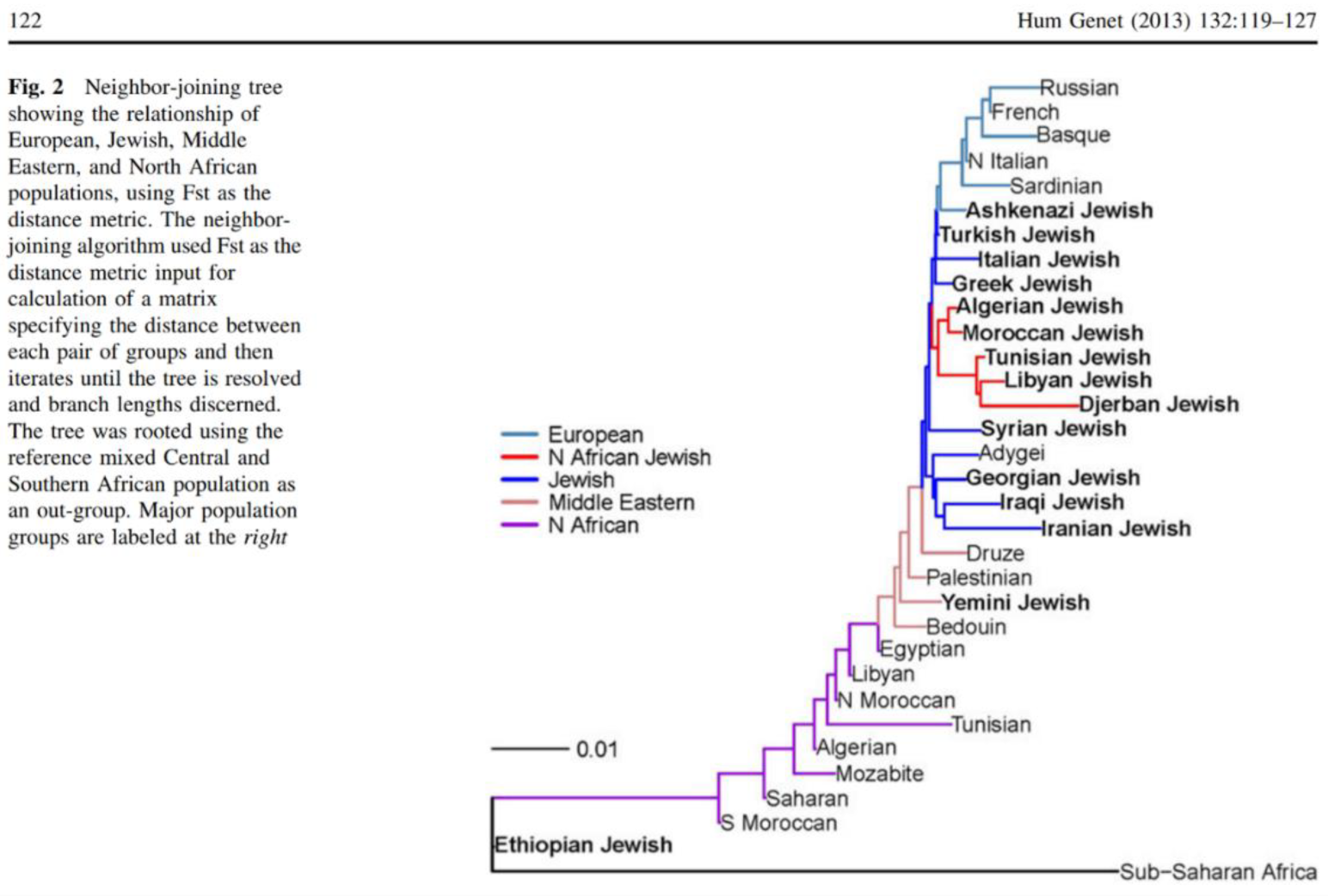
Uniparental Misinterpretation and the Illusion of a Levantine Paternal Origin
Claims that Ashkenazi Jews possess a predominantly Levantine paternal ancestry have relied heavily on a small set of Y-chromosomal studies published between approximately 2001 and 2009. These studies are frequently cited as having resolved the question of Ashkenazi origins, particularly through the presence of haplogroups J1, J2, and E, which are routinely labeled as Near Eastern or Levantine. However, closer examination shows that this conclusion is not anchored in robust comparative inference. Instead, it reflects a combination of reference population omission, low phylogenetic resolution, and population pooling that together limit what these datasets can actually support about Ashkenazi-specific paternal origins.
With the exception of Semino et al. (2004), the major Ashkenazi Y-DNA analyses from this period did not include Southern Italian populations as comparators, despite their central relevance to Mediterranean genetic structure and their well-documented Jewish presence since antiquity. Instead, these studies typically contrasted Ashkenazi Jews with Middle Eastern populations on one side and Northern or Central European populations on the other. This reproduces, in uniparental form, the same asymmetric reference framework later observed in autosomal studies. Under such a design, haplogroups shared broadly across the eastern Mediterranean and Southern Europe are implicitly reclassified as Levantine by default, not because their geographic specificity has been demonstrated, but because Southern European baselines are absent.
Hammer et al. (2009) illustrates the problem clearly. Although frequently cited as evidence for a Levantine paternal origin of Ashkenazi Jews, the study explicitly pools Ashkenazi and non-Ashkenazi Jewish samples and does not report haplogroup frequencies stratified by Jewish subgroup. The authors describe their dataset as 1,575 Jews representative of the Diaspora, without providing Ashkenazi-specific proportions. As a result, the reported haplogroup distributions cannot be used to infer Ashkenazi paternal structure, yet they are routinely cited as if they do. In addition, haplogroups are often reported at coarse resolution, such as J or E-M35, which obscures subclade-level geographic structure and increases the risk of treating pan-Mediterranean lineages as region-specific markers.
Other frequently cited studies show similar limitations. Behar et al. (2004) and Nebel et al. (2001) report Ashkenazi Y-DNA frequencies but do so using broad haplogroup bins and without Southern European comparators. Shen et al. (2004) is based on a very small Ashkenazi sample and lacks the resolution required to distinguish Levantine-specific from pan-Mediterranean lineages. In all cases, haplogroups such as J2 and E-M35 are interpreted as Near Eastern in origin despite their well-established prevalence in Southern Italy, Greece, and Anatolia.
Semino et al. (2004) is distinct because it includes multiple Southern Italian populations alongside Ashkenazi Jews and reports downstream resolution for haplogroups E and J. When that comparison is made explicitly, the supposed Levantine signal collapses. Ashkenazi Jews exhibit frequencies of J2 and E-M123 that fall within Southern Italian ranges, while Levantine populations often show markedly different distributions, including elevated J1 and reduced E-M123. Notably, E-M123, frequently cited as evidence of Levantine ancestry in Ashkenazim, reaches comparable or higher frequencies in Southern Italy and Anatolia, while being relatively uncommon in several Levantine populations.
Thus, the interpretation of Ashkenazi Y-chromosomal lineages as predominantly Levantine is not supported by the totality of the evidence. Rather, it emerges from a methodological framework in which Southern Europe is excluded, haplogroup resolution is insufficient, and population pooling obscures Ashkenazi-specific structure. When appropriate Mediterranean comparators are included, Ashkenazi paternal lineages align closely with Southern Italian and Aegean populations, consistent with a Mediterranean origin shaped by founder effects and endogamy rather than a Levantine paternal majority model.
Table note. In all tables in this manuscript, missing source values are coded as NR (not reported in the cited study). Values prefixed by ~ are approximate as reported. All haplogroup frequencies are percent.
As summarized in
Table 1, the early Y-chromosomal literature does not provide consistent or properly contextualized evidence for a predominantly Levantine paternal origin of Ashkenazi Jews. Resolving this ambiguity requires focusing on the only dataset from this period that places Ashkenazi Jews and Southern Italian populations side by side at subclade resolution.
The only early Y-chromosomal study to directly compare Ashkenazi Jews with Southern Italian regional populations at subclade resolution is Semino et al. (2004). Unlike later syntheses that rely on pooled Jewish samples or coarse haplogroup bins, this study reports haplogroups E and J for Ashkenazi Jews alongside multiple Southern Italian regional samples, including Apulia, Calabria, and Sicily. This design enables direct evaluation of whether Ashkenazi paternal lineages commonly labeled as Levantine are exceptional relative to Southern Europe.
At this resolution, Ashkenazi Y-chromosomal structure is dominated by haplogroups E and J2, both of which fall within Southern Italian variation. Ashkenazi Jews show E-M123 at 11.7 percent and E-M78 at 5.2 percent. These values closely match Southern Italian regions such as Apulia (E-M123 = 11.6 percent), Calabria 2 (E-M123 = 13.2 percent), and Sicily (E-M78 = 12.7 percent). Calabria 1 shows the inverse pattern, with high E-M78 (16.3 percent) and low E-M123 (2.5 percent), underscoring strong regional heterogeneity within Southern Italy itself. This heterogeneity shows that neither E-M123 nor E-M78 is geographically diagnostic of the Levant.
Haplogroup J shows the same pattern of Mediterranean continuity. In Semino et al. (2004), Ashkenazi Jews exhibit J2 (J-M172) at 23.2 percent, making it the dominant J lineage. Comparable or higher J2 frequencies are observed in Apulia (29.1 percent), Calabria 1 (22.8 percent), and Calabria 2 (20.0 percent), while Sicily shows moderately lower J2 (16.7 percent). In both Ashkenazim and Southern Italians, J2 exceeds J1, a structure that contrasts sharply with Levantine populations.
J1 (J-M267) is present in Ashkenazi Jews at 14.6 percent, elevated relative to some Southern Italian regions but well within a Mediterranean range. Southern Italy itself exhibits substantial regional variation in J1, ranging from very low values in Apulia (2.3 percent) and Calabria 1 (1.8 percent) to higher values in Sicily (7.1 percent). This intra-regional spread shows that J1 is not absent from Southern Europe and that moderate J1 frequencies cannot be treated as evidence of Levantine origin in isolation.
Crucially, the Ashkenazi J1 frequency remains far below Levantine paternal structures, in which J1 typically dominates the Y-chromosomal pool and often exceeds J2. The Ashkenazi pattern instead mirrors Southern European structure, with J2 dominant and J1 secondary. Given the long-documented founder effects and endogamy in Ashkenazi populations, modest elevation of J1 relative to some Southern Italian regions is parsimoniously explained by genetic drift acting on a small number of founding lineages rather than by descent from a Levantine paternal majority.
When Southern Italian comparators are included, the supposed Levantine signal in Ashkenazi Y-DNA disappears. Ashkenazi Jews align with a Mediterranean continuum characterized by substantial E-M123 and J2, moderate and variable J1, and strong regional heterogeneity rather than with the J1-dominant structure typical of Levantine populations. The interpretation of Ashkenazi paternal ancestry as predominantly Levantine arises primarily when Southern Europe is excluded and when drift-sensitive lineages are over-interpreted without comparative context.
Table 2 summarizes the Semino et al. (2004) data directly.
In contrast, Levantine populations are characterized by a fundamentally different Y-chromosomal structure, with J1 typically exceeding J2 and E-M123 occurring at substantially lower frequencies than those observed in both Ashkenazi Jews and Southern Italians. This inversion of haplogroup dominance is incompatible with a Levantine paternal majority model for Ashkenazim and instead supports a Mediterranean paternal structure shaped by founder effects and long-term endogamy.
Demonstration of Ashkenazi Divergence from Levantine Paternal Structure
If Ashkenazi paternal ancestry were predominantly Levantine, Ashkenazi Y-chromosomal structure would be expected to resemble Levantine populations when evaluated using the same haplogroups and subclades. Levantine populations are commonly characterized by high frequencies of J1-M267, reduced representation of J2-M172 relative to J1, and comparatively low frequencies of E-M123. This pattern reflects demographic expansions that differ markedly from Southern European and Mediterranean Y-chromosomal structure.
When Ashkenazi Jews are compared against Levantine populations using these same markers, a clear divergence is observed. Ashkenazim show E-M123 and J2 at levels comparable to Southern Italian and Aegean populations, while J1 remains secondary. In contrast, Levantine populations show a reversal of this structure, with J1 dominating the paternal pool and E-M123 occurring at lower frequencies.
This contrast is summarized below.
Table 3.
Y-DNA haplogroup structure in Ashkenazi Jews compared with Levantine populations (E, J1, J2 only).
Table 3.
Y-DNA haplogroup structure in Ashkenazi Jews compared with Levantine populations (E, J1, J2 only).
| Population |
E-M78 |
E-M123 |
J1-M267 |
J2-M172 |
| Ashkenazi Jews |
5.2 |
11.7 |
14.6 |
23.2 |
| Lebanese |
~2 to 5 |
~4 to 5 |
~25 to 35 |
~10 to 15 |
| Palestinians |
~1 to 3 |
~2 to 4 |
~35 to 45 |
~10 to 15 |
| Iraqis |
~1 to 3 |
~2 to 3 |
~40 to 50 |
~10 to 15 |
In this comparison, Ashkenazi Jews do not align with Levantine populations on the paternal axis. Instead, they occupy a Mediterranean position characterized by elevated J2 and E-M123 alongside a secondary J1 component. The Levantine profile is J1-dominant with lower J2 and reduced E-M123. This inversion rules out a Levantine paternal majority model for Ashkenazi Jews.
Importantly, moderate elevation of J1 in Ashkenazim relative to some Southern Italian regions does not imply Levantine origin. Southern Italy itself exhibits substantial J1 heterogeneity, and Ashkenazi populations experienced strong founder effects and long-term endogamy that can amplify specific lineages. Drift can elevate J1 frequencies without implying descent from a population whose overall structure is J1-dominant. By contrast, Levantine J1 dominance reflects sustained demographic expansion that is not mirrored by the Ashkenazi paternal profile.
Thus, when Ashkenazi Jews are compared directly to both Southern European and Levantine populations using the same Y-chromosomal markers, their paternal structure aligns with a Mediterranean continuum and diverges sharply from Levantine patterns. Claims that Ashkenazi Y-DNA confirms a predominantly Levantine origin arise primarily when Southern Europe is excluded and when drift-sensitive lineages such as J1 are interpreted without comparative context.
Fine-Scale Southern European J1 Variation and Upper-Range Overlap
Fine-scale surveys of Southern European Y-chromosomal variation show that the range of haplogroup J1 observed in Southern Italy by Semino et al. (2004) does not represent an upper bound. When Southern Europe is sampled at micro-regional resolution (Di Giacomo et al., 2003; Boattini et al., 2013), J1-M267 frequencies are highly heterogeneous and locally elevated across the broader Greco-Roman Mediterranean continuum. Province-level Italian datasets document multiple Southern Italian localities with J1 frequencies exceeding those reported in Semino et al., particularly in Sicily and parts of Campania. A later synthesis of Sicilian and Southern Italian genetic data (Sarno et al., 2017), incorporating these underlying regional surveys, confirms the presence of elevated local Southern Italian values while showing that these populations remain J2-dominant overall. These upper-range J1 values approach or overlap those observed in Ashkenazi Jews, demonstrating that moderate J1 enrichment in Ashkenazim does not exceed plausible Southern European source variation once regional structure and founder effects are taken into account.
Comparable J1 levels are also observed in parts of Central Italy, including Tuscany, indicating that elevated J1 is not restricted to the southern peninsula. Elevated values are likewise reported in the Aegean. Fine-scale sampling on Crete reports J1-M267 frequencies overlapping Southern Italian and Ashkenazi levels, including a value of approximately 10.5 percent at Nea Nikomedeia (King et al., 2008).
Table 4.
Upper-range regional J1-M267 frequencies in Southern Italy and the Aegean.
Table 4.
Upper-range regional J1-M267 frequencies in Southern Italy and the Aegean.
| Region |
Population |
J1-M267 |
| Italy |
Sicily, Agrigento |
11.1 |
| Italy |
Western Sicily |
10.13 |
| Greece |
Crete, Nea Nikomedeia |
10.5 |
| Italy |
Trapani, Sicily |
8.82 |
| Italy |
Campania, Benevento |
8.3 |
| Greece |
Crete |
8.3 |
| Italy |
Sicily, Southwest |
7.0 |
| Italy |
Calabria, Tyrrhenian Calabria |
6.8 |
Frequencies shown represent high reported regional J1-M267 values in Southern Italy and the Aegean. Agrigento and other synthesized values are reported in Sarno et al. (2017) and derive from underlying province-level datasets, including Boattini et al. (2013). Western Sicily and Trapani values are taken directly from Boattini et al. (2013). The Nea Nikomedeia value is reported in King et al. (2008). All listed populations retain a J2-dominant Mediterranean paternal structure.