Submitted:
25 January 2026
Posted:
26 January 2026
You are already at the latest version
Abstract
Bacteriophages and phage-derived lytic enzymes are emerging as promising alternatives to antibiotics in aquaculture; however, their effects on non-target microbial communities remain insufficiently characterized. This study evaluated the impact of a lytic phage (CH20) and a phage-derived lysin (LysVp1) on the microbiota associated with seawater, rotifers, and zebrafish larvae challenged with Vibrio alginolyticus GV09. Parallel treatments with phage CH20, lysin LysVp1, or no treatment were independently applied to each biological matrix. Microbial commu-nities were analyzed using 16S rRNA gene sequencing, with DNA- and RNA-derived datasets evaluated separately. Alpha diversity indices were compared using appropriate statistical tests, while beta diversity was assessed using Aitchison distance, PERMANOVA, and dispersion anal-yses, and differential abundance was evaluated using ANCOM-BC2. Alpha diversity metrics (Chao1, Shannon, and Simpson indices) showed no significant differences among treatments across all matrices, indicating preservation of microbial richness and diversity. Beta diversity pat-terns differed according to the nucleic acid source, with RNA-based analyses revealing treat-ment-associated shifts in rotifer and water microbiota that were not consistently detected at the DNA level. In zebrafish larvae, neither phage nor lysin treatment significantly altered overall community structure, although dispersion effects reflected limitations related to sample size. Overall, these results indicate that phage CH20 and lysin LysVp1 exert minimal impact on alpha diversity and limited, context-dependent effects on microbial community structure, supporting their microbiota-safe potential for aquaculture applications.
Keywords:
microbiota
; lysin
; endolysin
; vibrio
; lytic phage
; rotifer
; live feed
1. Introduction
Microbial communities associated with early life stages of fish play a pivotal role in host development, nutrient assimilation, immune maturation, and protection against opportunistic pathogens [1,2,3,4]. In marine hatcheries, the microbiota of rearing water and live feed constitutes the primary source of microbial exposure for fish larvae and strongly influences the establishment of larval-associated microbial assemblages [5,6,7]. Perturbations of these microbial communities during early development can increase disease susceptibility and compromise larval performance, making microbiota management a central challenge in modern aquaculture.
Among live feeds, rotifers (Brachionus spp.) are widely used as the first exogenous food in marine larviculture due to their appropriate size, nutritional flexibility, and ease of large-scale production [8]. However, rotifers harbor dense and metabolically active microbial communities that often exceed those present in the surrounding rearing water [9,10]. These microbiota may include opportunistic and pathogenic bacteria, particularly members of the genus Vibrio, which can be efficiently transferred to fish larvae during feeding [11,12,13]. As a result, rotifers are increasingly recognized as critical microbial vectors in hatchery systems, capable of shaping larval microbiota assembly and influencing the onset of bacterial diseases during sensitive developmental windows.
Vibriosis remains one of the most prevalent and economically damaging diseases in marine aquaculture worldwide [14,15]. While antibiotics have historically been used to control bacterial outbreaks, their extensive application has raised concerns related to antimicrobial resistance, environmental persistence, and unintended disruption of microbial community structure [16,17,18,19]. These limitations have accelerated the search for alternative disease control strategies that combine antimicrobial efficacy with microbiota compatibility.
Bacteriophages are frequently considered microbiota-sparing due to their narrow host specificity; however, recent evidence indicates that phage-induced lysis of target bacteria may indirectly alter microbial community structure through nutrient release and ecological interactions [20,21,22,23]. Similarly, endolysins—phage-derived enzymes that enzymatically degrade bacterial peptidoglycan—have emerged as promising antibacterial agents, particularly against antibiotic-resistant bacteria [24,25]. Despite their increasing biotechnological relevance, the ecological effects of endolysins on complex microbial communities remain poorly characterized, especially in aquatic environments.
Although phage therapy has been successfully applied in aquaculture to reduce pathogen loads and improve host survival, most studies have focused primarily on disease outcomes rather than microbiota-level responses [26,27,28]. Only a limited number of investigations have evaluated how phage treatments influence water or host-associated microbiota, often reporting minimal or context-dependent effects [29,30,31]. In contrast, systematic assessments of endolysin impacts on aquaculture-relevant microbiota are largely absent.
In this study, we examined the effects of a lytic phage and a phage-derived endolysin on the microbiota associated with seawater, rotifers, and fish larvae under controlled experimental conditions. By integrating microbiota profiling with targeted antimicrobial exposure, this work addresses a critical knowledge gap regarding the ecological compatibility of phage-based interventions and contributes to the evidence base supporting their rational implementation in sustainable aquaculture.
2. Results
2.1. Effect of Phage and Lysin Application on Rotifer Microbiota
A total of 453 ASVs were retained after applying filters to the dataset of 54 samples. The rarefaccion curves of all samples reached the plateau, indicating that the sequencing depth used was sufficient to detect the existing diversity (Supplementary Figure S1).
2.1.1. Alpha and Beta Diversity
Alpha diversity indices (Chao1, Shannon, and Simpson) of the rotifer and water microbiota, calculated from DNA- and RNA-derived datasets, showed no significant differences among control, lysin-, and phage-treated groups, as indicated by the shared superscript letters, confirming the absence of treatment-related effects on microbial diversity (Supplementary Table S1).
At the beta diversity level (Aitchison distance), differences were observed depending on whether DNA- or RNA-derived data were analyzed. In the principal coordinate analysis (PCoA) of rotifer samples (Figure 1A,B), DNA-based profiles did not cluster according to treatment groups, whereas RNA-based profiles showed a distinct separation of the lysin-treated (Lys) samples from the Control and Phage groups (Figure 1B). These differences were supported by PERMANOVA, which confirmed that the Lys group was significantly different from both the Control and Phage groups (p < 0.05; Table 1). In contrast, water samples exhibited an opposite pattern, with DNA-based analyses revealing significant differences among groups, while RNA-based analyses did not show significant separation (Table 1).
2.1.2. Microbiota Composition of Rotifer
At the phylum level, the rotifer and water microbiota was mainly composed of Proteobacteria, Actinobacteriota, Firmicutes and Bacteroidota, in descending order (Figure 2), in both DNA and RNA samples. It is noteworthy that none of these phyla showed significant differences between groups in both rotifers and water samples (Supplementary Table S12).
At the genus level, 18 components were identified with average relative abundance greater than 2% in at least one of the groups (Figure 3). In the rotifer samples, a similar structure was observed in the most abundant components in both DNA and RNA samples, with Glutamicibacter, Escherichia-Shigella, Mesoflavibacter y Marinomonas being the most abundant genera, with average relative abundance of at least 4% per group. These results are in agreement with those obtained by ANCOM-BC2 (Supplementary Table S12), where no significant differences were found at the genus level among these groups. These results contrast with the composition of the microbiota in the water samples, where differences in the most abundant components (Figure 3). In both DNA and RNA samples, Glutamicibacter was predominantly found in the Phage group, with an average abundance of 20% and 15%, respectively, while Escherichia-Shigella was the most abundant in the Lys group, with an average abundance of 23% and 13%, respectively.
Full information on the relative abundance of the components of the microbiota at the different taxonomic levels can be found in the supplementary material (Supplementary Tables S2 - S11).
The differences in microbiota composition at the genus level in the water samples described above were statistically significant according to ANCOM-BC2 (Figure 4), adding in the case of DNA samples, two more genera, Pseudoalteromonas and Streptococcus.
2.2. Effect of Phage and Lys Application on Zebrafish Microbiota
A total of 302 ASVs were retained after applying filters to the dataset of 18 samples (DNA only). The rarefaccion curves of all samples reached the plateau (Supplementary Figure S2).
2.2.1. Alpha and Beta Diversity
Alpha diversity indices (Chao1, Shannon, and Simpson) of the zebrafish microbiota showed no significant differences among control, phage-, lysin-, and Vibrio alginolyticus GV09–challenged groups, as indicated by the shared superscript letters, confirming the absence of treatment-related effects on microbial diversity (Supplementary Table S13).
In addition, visualization of the beta diversity of the zebrafish microbiota by PCoA showed no clustering of the samples according to the different treatment groups (Figure 5). These results were supported by PERMANOVA analysis (Table 2), where no significant differences between groups were reported (P value < 0.05). However, it should be noted that the Betadisper analysis revealed significant differences in some comparison such as, Control vs GV09 (P value = 0.028), Control vs GV09_Lys (P value = 0.026), Control vs GV09_Phage (P value = 0.031) and GV09 vs GV09_Lys (P value = 0.032), suggesting a difference in dispersion but not in composition, indicating that the PERMANOVA analysis might not be reliable. These results reflect a limitation of the design in which a low number of replicates per group was used.
2.2.2. Composition of the Larvae Microbiota
The composition of the zebrafish microbiota was represented by the phylum Proteobacteria (ranging from 60 to 83% of average relative abundance), followed by the phyla Firmicutes, Bacteroidota, Actinobacteriota and Planctomycetota (Figure 6) regardless of control or treatment group. These phyla, and also those with less than 1% relative abundance, showed no significant differences between groups according to ANCOM-BC2.
Full information on the relative abundance of the components of the microbiota at the different taxonomic levels can be found in the supplementary material (Supplementary Tables S14 - S18).
At the genus level, the composition of the microbiota presented different profiles according to the control or treatment groups (Figure 7). In the control group, the most abundant genera were Pseudomonas, Aeromonas, while the treatment groups containing the Vibrio strain GV09 presented Vibrio as the most abundant genus, which was lower in the group treated with Lys (GV09_Lys). In the latter, the most abundant genus was Escherichia-Shigella, similar to the Lys group. On the other hand, the group containing only the phage addition (Phage group) presented a profile more similar to the control group. However, no significant differences were detected in any genus according to ANCOM-BC2, which could be associated to the dispersion of the samples and the low number of replicates used per group. But differences were found between groups at higher taxonomic levels such as at the family level (Supplementary Table S19), where Enterobacteriaceae presented significant differences between the Lys and Phage groups (lfc = -2.970, P value = 0.038), highlighting that Escherichia belongs to this family. Another family that presented differences between the groups was Pseudomonadaceae, which was found in the comparison between the Control group and GV09, GV09_Lys and Lys, the genus Pseudomonas belonging to this family be highlighted with higher abundance in the Control group.
Overall, these results suggest that it might be appropriate to include ASVs classified as Escherichia-Shigella as a group to be excluded when evaluating the effect on the microbiota of any overexpressed protein in competent bacteria of the E. coli species.
3. Discussion
A central challenge in microbial management for aquaculture is achieving pathogen control without destabilizing the microbial communities that support host development, immune maturation, and colonization resistance. Antibiotics remain effective for acute outbreak control, but their broad-spectrum activity frequently produces collateral damage to commensal communities, promotes long-term shifts in community structure, and enriches antimicrobial resistance determinants [16,18]. In fish, antibiotic exposure has repeatedly been associated with reductions in intestinal microbiota diversity and compositional rearrangements, including depletion of putatively beneficial taxa and expansion of opportunistic groups [17]. These microbiota-level consequences are particularly concerning in hatchery settings, where early-life microbial assembly can shape later host performance and disease susceptibility [10,32]. Furthermore, Hurtado [33] highlight live feeds as a critical reservoir of antibiotic-resistant bacteria and resistance genes in larval fish culture systems, particularly under sustained antibiotic pressure. The high prevalence of florfenicol-resistant Vibrio spp. and associated resistance determinants underscores the urgent need for alternative, non-antibiotic strategies to control bacterial loads in live feeds and prevent the dissemination of antimicrobial resistance in aquaculture environments.
Phage-based interventions are often positioned as “microbiota-sparing” because of their narrow host range; however, evidence across model systems suggests a more nuanced picture. In murine models, phage administration can modulate both microbiota composition and metabolite profiles, in part through indirect ecological effects following targeted lysis [20]. Importantly, multiple studies have shown that phage therapy can reduce specific pathogens while preserving overall alpha diversity and avoiding large-scale perturbation of community structure, supporting the idea that phages can provide targeted control with limited collateral impact [21,31]. More recently, work in aquaculture-relevant systems indicates that phage treatment can prevent infection while minimally affecting water microbiota, reinforcing their suitability for aquatic environments where microbial stability is desirable [29]. Taken together, these results indicate that phage-based strategies can achieve pathogen control while maintaining overall microbiota stability, with community-level effects largely shaped by ecological context rather than indiscriminate disturbance. Recent work in marine host-associated microbial communities (sponge) highlights that phage-induced shifts are system specific and reflect structured ecological reorganization rather than community collapse [34]. This perspective aligns with the present findings and supports the positioning of phages—and phage-derived endolysins—as microbiota-compatible alternatives to conventional antibiotics.
Endolysins represent a parallel “precision antimicrobial” strategy, but their ecological footprint on complex microbiomes has been much less studied than that of phages. Foundational work describes endolysins as potent enzymatic antibacterials with potential for high specificity and reduced off-target effects, particularly when engineered or formulated to target defined bacterial groups [24,25]. Recent experimental evidence in aquaculture systems demonstrates that recombinant endolysins can rapidly and significantly reduce pathogenic Vibrio loads in both live feed organisms (rotifers) and fish larvae, achieving multi-log reductions within short exposure times without the use of antibiotics [35]. Notably, these effects were observed across several clinically relevant Vibrio species, supporting the applicability of endolysins as targeted, non-antibiotic tools for controlling Vibrio-associated disease risk during early life stages in marine hatchery settings. In line with this view, recent reviews emphasize that advances in bacteriophage and endolysin engineering have enabled highly targeted antimicrobial strategies that enhance specificity against foodborne pathogens while reducing unintended effects on non-target microbial communities, reinforcing their potential as precision biocontrol tools [36]. Recent reviews also explicitly frame next-generation endolysins as precision antimicrobials and highlight the critical gap in empirical studies assessing their microbiome compatibility [36,37]. Where evaluated, endolysins (phage lysins) are widely framed as microbiome-friendly antimicrobials due to their comparatively high specificity, which can—at least in principle—enable targeted depletion of defined bacterial groups while limiting off-target effects [38]. At the same time, this microbiota-compatibility should not be assumed a priori: recent syntheses stress that specificity does not automatically translate into microbiome-friendliness, and therefore lysins should be assessed case-by-case using complementary in vitro, ex vivo, and in vivo microbiome models [38,39]. In this context, the present study adds aquaculture-relevant evidence that lysin exposure is not associated with broad losses of alpha diversity and that any detectable community-level effects are subtle and matrix-dependent, consistent with a precision-antimicrobial paradigm that prioritizes pathogen control while preserving community-level stability [38,39].
A key interpretive dimension raised by recent studies is that “no change in alpha diversity” does not necessarily imply “no ecological effect”. Several phage studies report preserved richness and evenness alongside shifts in beta diversity, reflecting altered relative abundances, niche release, and resource redistribution after lysis [20,40,41]. This framework is particularly useful for interpreting scenarios where compositional separation is detectable without diversity collapse. Moreover, emerging evidence suggests that analytical choice (e.g., profiling total DNA-based communities versus RNA-enriched active fractions) can influence sensitivity to treatment effects. Studies emphasizing functional or activity-linked readouts indicate that active-community shifts can occur without parallel changes in total-community structure, which may explain why RNA-based profiles sometimes reveal treatment-associated separation not evident in DNA-based analyses [42,43]. Consistent with this view, RNA-based analyses have been shown to reveal shifts in the active fraction of microbial communities that remain undetected in DNA-based profiles, despite apparent structural stability [44]. Similar observations in fish-associated microbiota indicate that activity-based approaches can uncover biologically relevant community responses to perturbation without concurrent changes in overall diversity [45]. This concept is especially relevant for live feed and rearing water, where microbial turnover is rapid and activity states can change quickly in response to perturbations. In the present study, this distinction is exemplified by rotifer-associated microbiota, where RNA-based analyses revealed treatment-related clustering not observed in DNA-derived datasets, suggesting modulation of metabolically active fractions rather than restructuring of the total community. In contrast, zebrafish larvae were assessed only at the DNA level, and the absence of detectable beta diversity separation is consistent with either true community stability or limited sensitivity to subtle, activity-level shifts under the experimental conditions used. From a methodological perspective, these results underscore the value of combining DNA- and RNA-based microbiome profiling when assessing targeted antimicrobial interventions. While DNA-based analyses capture overall community structure, RNA-based approaches are more sensitive to shifts in metabolically active fractions, particularly in rapidly turning-over systems such as live feeds and rearing water, thereby improving the detection and interpretation of subtle treatment effects.
Across all matrices, microbial communities were dominated by the phyla Proteobacteria, Actinobacteriota, Firmicutes, and Bacteroidota, consistent with previous reports for marine rearing systems, live feeds, and fish-associated microbiota [46,47,48]. Despite this conserved higher-level structure, compositional analyses revealed treatment-associated shifts at finer taxonomic resolution. Rotifer- and fish larva–associated microbiota remained largely stable at the phylum level, whereas rearing water samples exhibited detectable changes at finer taxonomic resolution. In rearing water, Glutamicibacter showed higher relative abundance under phage treatment, while Pseudoalteromonas and Streptococcus also tended to increase in phage-treated conditions and remained low or more variable following lysin exposure. These differences were confined to a limited number of low- to moderate-abundance genera and did not translate into broad alterations of alpha diversity, indicating selective, treatment-dependent redistribution within an otherwise stable water microbiota.
4. Materials and Methods
4.1. Bacterial Strain, Phage, and Endolysin
The bacterial strain Vibrio alginolyticus GV09, originally isolated from marine aquaculture environments, was used as the target pathogen and has been previously characterized for susceptibility to both lytic phages and phage-derived endolysins [35]. The lytic bacteriophage CH20, specific to V. alginolyticus, was propagated using the double-layer agar method, and high-titer stocks were quantified by plaque assay and stored at 4 °C until use. The recombinant endolysin LysVp1, derived from a Vibrio-infecting bacteriophage, was produced by heterologous expression in Escherichia coli and purified by affinity chromatography as previously described [35].
4.2. Experimental Design
The experimental design was conducted under controlled laboratory conditions and included three biological matrices: rotifer rearing water, rotifers (Brachionus plicatilis), and zebrafish larvae (Danio rerio), following procedure described previously [35]. Each matrix was independently subjected to three experimental conditions: untreated control, phage treatment (CH20), and endolysin treatment (LysVPp1). All assays were performed using independent biological replicates. For rotifer assays, cultures were concentrated by filtration through a 30 µm nylon mesh and resuspended in 1 µm-filtered seawater. Rotifer suspensions were inoculated with Vibrio alginolyticus GV09 to a final concentration of 5 × 10⁶ CFU mL⁻¹ and incubated for 15 min at 22 °C to allow bacterial association. Subsequently, aliquots of 9 mL, containing approximately 350 rotifers per experimental unit, were distributed into sterile plates. Treatments were applied by adding 1 mL of the corresponding solution: sterile seawater for controls, phage CH20 to reach a multiplicity of infection (MOI) of 100, or endolysin LysVPp1 to achieve a final concentration of 1 mg mL⁻¹. Following treatment, samples were incubated for 20 min at 22 °C. Rotifers were then recovered by filtration, washed, and processed for downstream microbiota analyses. For zebrafish larval assays, larvae were maintained in E3 medium and distributed into sterile plates at a density of six larvae per 9 mL well. Larval cultures were challenged with V. alginolyticus GV09 at 5 × 10⁶ CFU mL⁻¹ and incubated for 15 min at 22 °C prior to antimicrobial treatment. Control groups received 1 mL of E3 medium, while treated groups received 1 mL of phage CH20 (MOI = 100) or 1 mL of LysVPp1 (final concentration 1 mg mL⁻¹). Incubation was continued for 20 min, after which larvae and surrounding water were collected for microbiota characterization.
The selected exposure times, antimicrobial concentrations, and organism densities were designed to reflect short-contact bioassays compatible with hatchery practices, as previously validated for phage and endolysin activity in live feed and fish larvae systems. Sample sizes were constrained by larval availability and ethical considerations, and these constraints were accounted for during statistical analysis.
4.3. Nucleic Acid Extraction
For simultaneous DNA and RNA extraction, rotifer and water samples were processed following the protocol described by [43], using a single co-extraction workflow. Rotifer samples were recovered by filtration, thoroughly washed with phosphate-buffered saline to remove loosely associated microorganisms, and transferred to extraction tubes. Water samples were processed by filtration through 0.45 µm membrane filters, and the filters were directly used as input material for nucleic acid extraction. Both rotifer biomass and water filters were subjected to enzymatic pretreatment with lysozyme and proteinase K, followed by mechanical disruption by bead beating [43]. Total DNA and RNA were simultaneously extracted using the AllPrep PowerFecal DNA/RNA Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Nucleic acid concentrations were quantified fluorometrically, and RNA extracts were reverse-transcribed using random primers to generate cDNA. DNA- and cDNA-derived templates were subsequently processed independently for downstream sequencing analyses, as previously described [43].
Genomic DNA from zebrafish larvae was extracted following a washing step analogous to the procedure applied to rotifer samples. Briefly, pooled larvae were rinsed 2 times with sterile phosphate-buffered saline and transferred to 2 mL bead-beating tubes. Samples were subjected to enzymatic pretreatment with lysozyme and proteinase K as above. DNA was extracted using the DNeasy PowerSoil Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions, with minor modifications to enhance inhibitor removal.
DNA concentration and purity were assessed using a NanoDrop spectrophotometer and a Qubit dsDNA High Sensitivity assay (Thermo Fisher Scientific, Waltham, MA, USA), and only high-quality extracts were used for downstream analyses. For bacterial community profiling, the V4 region of the 16S rRNA gene was amplified using the universal primers 515F and 806R. PCR reactions were performed in a final volume of 30 µL containing 1.5 U of GoTaq® G2 Flexi DNA Polymerase (Promega, Madison, WI, USA), 6 µL of 5× reaction buffer, 1.5 mM MgCl₂, 0.25 µM of each primer, 0.5 mM of each dNTP, and 20 ng of template DNA. Negative controls without DNA template were included in all PCR runs. PCR products were purified using the QIAquick PCR Purification Kit (Qiagen, Hilden, Germany) and sequenced using paired-end chemistry (2 × 300 bp) on an Illumina HiSeq PE250 platform at CD Genomics.
4.4. Metagenomic Analysis
Raw paired-end 16S rRNA gene sequences were processed in R (v4.4.0) using the DADA2 pipeline, following the workflow described by [49]. Primer sequences were removed, and reads were quality filtered and truncated based on quality score profiles. Error rates were learned from the data, and amplicon sequence variants (ASVs) were inferred through denoising, merging of paired-end reads, and removal of chimeric sequences. Taxonomic assignment was performed using a naïve Bayesian classifier against the SILVA v138.1 reference database [50]. ASV count tables, taxonomic assignments, and sample metadata were integrated into phyloseq objects for downstream analyses. Alpha diversity indices (Chao1, Shannon, and Simpson) were calculated from untransformed ASV count data. For beta diversity analyses, ASV tables were subjected to a centered log-ratio (clr) transformation after zero handling, and Aitchison distances were calculated as Euclidean distances on clr-transformed data [51,52]. Community dissimilarities were visualized using principal coordinate analysis (PCoA). Differences in bacterial community composition among experimental groups were tested using PERMANOVA with 999 permutations [53]. Homogeneity of multivariate dispersions was assessed using betadisper followed by permutation testing [54]. Differential abundance analyses were conducted using ANCOM-BC2, which corrects for compositional and sampling biases [55]. All analyses were performed in R using the packages phyloseq, vegan, microbiome, and ANCOMBC.
5. Conclusions
This study demonstrates that exposure to a lytic phage (CH20) or a phage-derived endolysin (LysVp1) does not induce broad disruptions of microbial diversity in seawater, rotifers, or zebrafish larvae, supporting their microbiota-compatible profile in aquaculture settings. Across all matrices, alpha diversity remained stable, while detected compositional changes were limited, context-dependent, and largely restricted to low-abundance taxa, with clearer treatment signals emerging only when RNA-based analyses were applied. Together, these findings indicate that phages and endolysins can achieve targeted antimicrobial effects without compromising overall microbial community structure, reinforcing their potential as precision, non-antibiotic tools for sustainable disease management in early-life aquaculture systems.
Supplementary Materials
Doc file: Figure S1: Rarefaction curves of rotifer and water microbiota; Figure S2: Rarefaction curves of zebrafish microbiota; Table S1: alpha diversity indices of rotifer and water microbiota; Table S13: Alpha diversity indices of the zebrafish microbiota. Excel file: Table S2. Relative abundance (%) at phylum level from DNA samples of microbiota; Table S3. Relative abundance (%) at phylum level from RNA samples of microbiota; Table S4. Relative abundance (%) at class level from DNA samples of microbiota; Table S5. Relative abundance (%) at class level from RNA samples of microbiota; Table S6. Relative abundance (%) at order level from DNA samples of microbiota; Table S7. Relative abundance (%) at order level from RNA samples of microbiota; Table S8. Relative abundance (%) at family level from DNA samples of microbiota; Table S9. Relative abundance (%) at family level from RNA samples of microbiota; Table S10. Relative abundance (%) at genus level from RNA samples of microbiota; Table S11. Relative abundance (%) at genus level from DNA samples of microbiota; Table S12. Summary of results of pairwise comparison using ANCOM-BC2 in microbiota samples from rotifers and water exposed to Phage and Lys; Table S14. Relative abundance (%) at phylum level of zebrafish microbiota; Table S15. Relative abundance (%) at class level of zebrafish microbiota; Table S16. Relative abundance (%) at order level of zebrafish microbiota; Table S17. Relative abundance (%) at family level of zebrafish microbiota; Table S18. Relative abundance (%) at genus level of zebrafish microbiota; Table S19. Summary of results of pairwise comparison using ANCOM-BC2 in zebrafish microbiota samples.
Author Contributions
Conceptualization, J.R. , C.D.M. and R.R.; methodology, M.M.; A.P and L.H. investigation, C.R. and M.M.; data curation, C.R.; writing—original draft preparation, C.R. and J.R; writing—review and editing, J.R. and C.D.M; project administration, J.R.; funding acquisition, J.R. , C.D.M.
Funding
This research was funded by ANID, Anillo Tecnológico ACT240045.
Institutional Review Board Statement
CICUA, University of Chile, Ethical code 20364-INTA-UCH.
Data Availability Statement
Data were included in Supplementary Material (Excel file). The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
We would like to express our sincere gratitude to Victoria Urzúa for her tireless dedication and exceptional technical support throughout this work.
Conflicts of Interest
The authors declare no conflicts tof interest.
References
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75.
- Selber-Hnatiw, S.; Rukundo, B.; Ahmadi, M.; Akoubi, H.; Al-Bizri, H.; Aliu, A.F.; Ambeaghen, T.U.; Avetisyan, L.; Bahar, I.; Baird, A.; et al. Human Gut Microbiota: Toward an Ecology of Disease. Front. Microbiol. 2017, 8, 1265–1265. [CrossRef]
- Pérez-Pascual, D.; Vendrell-Fernández, S.; Audrain, B.; Bernal-Bayard, J.; Patiño-Navarrete, R.; Petit, V.; Rigaudeau, D.; Ghigo, J.-M. Gnotobiotic rainbow trout (Oncorhynchus mykiss) model reveals endogenous bacteria that protect against Flavobacterium columnare infection. PLOS Pathog. 2021, 17, e1009302. [CrossRef]
- A Stressmann, F.; Bernal-Bayard, J.; Perez-Pascual, D.; Audrain, B.; Rendueles, O.; Briolat, V.; Bruchmann, S.; Volant, S.; Ghozlane, A.; Häussler, S.; et al. Mining zebrafish microbiota reveals key community-level resistance against fish pathogen infection. ISME J. 2020, 15, 702–719. [CrossRef]
- Bakke, I.; Coward, E.; Andersen, T.; Vadstein, O. Selection in the host structures the microbiota associated with developing cod larvae ( G adus morhua ). Environ. Microbiol. 2015, 17, 3914–3924. [CrossRef]
- I Vestrum, R.; Attramadal, K.J.K.; Vadstein, O.; Gundersen, M.S.; Bakke, I. Bacterial community assembly in Atlantic cod larvae (Gadus morhua): contributions of ecological processes and metacommunity structure. FEMS Microbiol. Ecol. 2020, 96. [CrossRef]
- Bugten, A.V.; et al. Rearing water microbiomes influence gut microbiota of Atlantic salmon parr. Aquaculture 2022, 548, 737661.
- Lubzens, E.; et al. BiotechRotifers in aquaculture. Hydrobiologia 2001, 446/447, 337–353.
- Munro, P.; Henderson, R.; Barbour, A.; Birkbeck, T. Partial decontamination of rotifers with ultraviolet radiation: the effect of changes in the bacterial load and flora of rotifers on mortalities in start-feeding larval turbot. Aquaculture 1999, 170, 229–244. [CrossRef]
- Vadstein, O.; Bergh, Ø.; Gatesoupe, F.; Galindo-Villegas, J.; Mulero, V.; Picchietti, S.; Scapigliati, G.; Makridis, P.; Olsen, Y.; Dierckens, K.; et al. Microbiology and immunology of fish larvae. Rev. Aquac. 2013, 5, S1–S25. [CrossRef]
- Gatesoupe, F.-J. Lactic acid bacteria increase the resistance of turbot larvae,Scophthalmus maximus,against pathogenic vibrio. Aquat. Living Resour. 1994, 7, 277–282. [CrossRef]
- Skjermo, J.; Vadstein, O. Techniques for microbial control in live feed production. Aquaculture 1999, 177, 333–343.
- Attramadal, K.J.; Salvesen, I.; Xue, R.; Øie, G.; Størseth, T.R.; Vadstein, O.; Olsen, Y. Recirculation as a possible microbial control strategy in the production of marine larvae. Aquac. Eng. 2012, 46, 27–39. [CrossRef]
- Ina-Salwany, M.Y.; Al-Saari, N.; Mohamad, A.; Mursidi, F.A.; Mohd-Aris, A.; Amal, M.N.A.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in Fish: A Review on Disease Development and Prevention. J. Aquat. Anim. Health 2019, 31, 3–22. [CrossRef]
- Silva, Y.J.; Costa, L.; Pereira, C.; Mateus, C.; Cunha, Â.; Calado, R.; Gomes, N.C.M.; Pardo, M.A.; Hernandez, I.; Almeida, A. Phage Therapy as an Approach to Prevent Vibrio anguillarum Infections in Fish Larvae Production. PLOS ONE 2014, 9, e114197. [CrossRef]
- Blaser, M.J. Antibiotic use and its consequences for the normal microbiome. Science 2016, 352, 544–545. [CrossRef]
- Navarrete, P.; et al. Oxytetracycline reduces intestinal microbiota diversity in Atlantic salmon. J. Aquat. Anim. Health 2008, 20, 177–183.
- Murray, C.J.L.; et al. Global burden of antimicrobial resistance in 2019. Lancet 2022, 399, 629–655.
- Zhou, L.; et al. Environmental antibiotics impair zebrafish gut health. Environ. Pollut. 2018, 235, 245–254.
- Hsu, B.B.; Gibson, T.E.; Yeliseyev, V.; Liu, Q.; Lyon, L.; Bry, L.; Silver, P.A.; Gerber, G.K. Dynamic Modulation of the Gut Microbiota and Metabolome by Bacteriophages in a Mouse Model. Cell Host Microbe 2019, 25, 803–814.e5. [CrossRef]
- Dissanayake, U.; Ukhanova, M.; Moye, Z.D.; Sulakvelidze, A.; Mai, V. Bacteriophages Reduce Pathogenic Escherichia coli Counts in Mice Without Distorting Gut Microbiota. Front. Microbiol. 2019, 10, 1984. [CrossRef]
- Donati, V.L.; Madsen, L.; Middelboe, M.; Strube, M.L.; Dalsgaard, I. The Gut Microbiota of Healthy and Flavobacterium psychrophilum-Infected Rainbow Trout Fry Is Shaped by Antibiotics and Phage Therapies. Front. Microbiol. 2022, 13, 771296. [CrossRef]
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114. [CrossRef]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [CrossRef]
- Nelson, D.C.; Schmelcher, M.; Rodriguez-Rubio, L.; Klumpp, J.; Pritchard, D.G.; Dong, S.; Donovan, D.M. Endolysins as Antimicrobials. Adv. Virus Res. 2012, 83, 299–365.
- Laanto, E.; Bamford, J.K.H.; Ravantti, J.J.; Sundberg, L.-R. The use of phage FCL-2 as an alternative to chemotherapy against columnaris disease in aquaculture. Front. Microbiol. 2015, 6, 829. [CrossRef]
- Kunttu, H.M.T.; Runtuvuori-Salmela, A.; Middelboe, M.; Clark, J.; Sundberg, L.-R. Comparison of Delivery Methods in Phage Therapy against Flavobacterium columnare Infections in Rainbow Trout. Antibiotics 2021, 10, 914. [CrossRef]
- Liu, R.; Han, G.; Li, Z.; Cun, S.; Hao, B.; Zhang, J.; Liu, X. Bacteriophage therapy in aquaculture: current status and future challenges. Folia Microbiol. 2022, 67, 573–590. [CrossRef]
- Fiedler, A.W.; Gundersen, M.S.; Vo, T.P.; Almaas, E.; Vadstein, O.; Bakke, I. Phage therapy minimally affects the water microbiota in an Atlantic salmon (Salmo salar) rearing system while still preventing infection. Sci. Rep. 2023, 13, 1–13. [CrossRef]
- Feng, C.; Jia, K.; Chi, T.; Chen, S.; Yu, H.; Zhang, L.; Raza, S.H.A.; Alshammari, A.M.; Liang, S.; Zhu, Z.; et al. Lytic Bacteriophage PZL-Ah152 as Biocontrol Measures Against Lethal Aeromonas hydrophila Without Distorting Gut Microbiota. Front. Microbiol. 2022, 13, 898961. [CrossRef]
- Richards, P.J.; Connerton, P.L.; Connerton, I.F. Phage Biocontrol of Campylobacter jejuni in Chickens Does Not Produce Collateral Effects on the Gut Microbiota. Front. Microbiol. 2019, 10, 476. [CrossRef]
- Paralika, V.; Makridis, P. Microbial Interactions in Rearing Systems for Marine Fish Larvae. Microorganisms 2025, 13, 539. [CrossRef]
- Hurtado, L.; Miranda, C.D.; Rojas, R.; Godoy, F.A.; Añazco, M.A.; Romero, J. Live Feeds Used in the Larval Culture of Red Cusk Eel, Genypterus chilensis, Carry High Levels of Antimicrobial-Resistant Bacteria and Antibiotic-Resistance Genes (ARGs). Animals 2020, 10, 505. [CrossRef]
- Steiner, L.X.; Schmittmann, L.; Rahn, T.; Lachnit, T.; Jahn, M.T.; Hentschel, U. Phage-induced disturbance of a marine sponge microbiome. Environ. Microbiome 2024, 19, 1–20. [CrossRef]
- Wang, S.-M.; Wu, J.-X.; Gunawan, H.; Tu, R.-Q. Optimization of Machining Parameters for Corner Accuracy Improvement for WEDM Processing. Appl. Sci. 2023, 12, 10324. [CrossRef]
- Galanakis, C.M. Phenols recovered from olive mill wastewater as additives in meat products. Trends Food Sci. Technol. 2018, 79, 98–105. [CrossRef]
- Cui, X.; Chai, L.; Zhang, Y.; Pan, Y.; Liu, H.; Lei, X.; Le, T. Next-generation antimicrobials: A review of phage lysins as precision weapons against drug-resistant pathogens. Virulence 2025, 16, 2562634. [CrossRef]
- Pottie, J.; De Maesschalck, C.; Van Immerseel, F.; Dewulf, J.; Boyen, F. Phage Lysins as Precision Antimicrobials: Opportunities and Challenges for Microbiome Compatibility. Gut Microbes 2024, 16, 2387144.
- Khan, F. M.; Ahmad, S.; Khan, A.; Ullah, A.; Khan, S. Phage Lysins as Next-Generation Precision Antimicrobials against Drug-Resistant Pathogens. Front. Pharmacol. 2024, 15, 1385261.
- Wang, S.-M.; Wu, J.-X.; Gunawan, H.; Tu, R.-Q. Optimization of Machining Parameters for Corner Accuracy Improvement for WEDM Processing. Appl. Sci. 2023, 12, 10324. [CrossRef]
- Wang, S.-M.; Wu, J.-X.; Gunawan, H.; Tu, R.-Q. Optimization of Machining Parameters for Corner Accuracy Improvement for WEDM Processing. Appl. Sci. 2023, 12, 10324. [CrossRef]
- De Vrieze, J.; Pinto, A.J.; Sloan, W.T.; Ijaz, U.Z. The active microbial community more accurately reflects the anaerobic digestion process: 16S rRNA (gene) sequencing as a predictive tool. Microbiome 2018, 6, 1–13. [CrossRef]
- Blazewicz, S.J.; Barnard, R.L.; A Daly, R.; Firestone, M.K. Evaluating rRNA as an indicator of microbial activity in environmental communities: limitations and uses. ISME J. 2013, 7, 2061–2068. [CrossRef]
- Pardo, A.; Villasante, A.; Romero, J. Skin Microbiota of Salmonids: A Procedure to Examine Active Bacterial Populations Using an RNA-Based Approach. Appl. Microbiol. 2023, 3, 485–492. [CrossRef]
- Jamet, E. An eye-tracking study of cueing effects in multimedia learning. Comput. Human Behav. 2014, 32, 47-53. [CrossRef]
- Walburn, J.W.; Wemheuer, B.; Thomas, T.; Copeland, E.; O'COnnor, W.; Booth, M.; Fielder, S.; Egan, S. Diet and diet-associated bacteria shape early microbiome development in Yellowtail Kingfish ( Seriola lalandi ). Microb. Biotechnol. 2018, 12, 275–288. [CrossRef]
- Najafpour, B.; Pinto, P.I.; Sanz, E.C.; Martinez-Blanch, J.F.; Canario, A.V.; Moutou, K.A.; Power, D.M. Core microbiome profiles and their modification by environmental, biological, and rearing factors in aquaculture hatcheries. Mar. Pollut. Bull. 2023, 193, 115218. [CrossRef]
- Roeselers, G.; Mittge, E.K.; Stephens, W.Z.; Parichy, D.M.; Cavanaugh, C.M.; Guillemin, K.; Rawls, J.F. Evidence for a core gut microbiota in the zebrafish. ISME J. 2011, 5, 1595–1608. [CrossRef]
- Wang, S.-M.; Wu, J.-X.; Gunawan, H.; Tu, R.-Q. Optimization of Machining Parameters for Corner Accuracy Improvement for WEDM Processing. Appl. Sci. 2023, 12, 10324. [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [CrossRef]
- Aitchison, J. The Statistical Analysis of Compositional Data. J. R. Stat. Soc. Ser. B 1986, 44, 139–177.
- Gloor, G.B.; Macklaim, J.M.; Pawlowsky-Glahn, V.; Egozcue, J.J. Microbiome Datasets Are Compositional: And This Is Not Optional. Front. Microbiol. 2017, 8, 2224. [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [CrossRef]
- Anderson, M.J. Distance-Based Tests for Homogeneity of Multivariate Dispersions. Biometrics 2005, 62, 245–253. [CrossRef]
- Lin, H.; Peddada, S. Das Analysis of Compositions of Microbiomes with Bias Correction. Nat. Commun. 2020, 11, 3514.
Figure 1.
Principal coordinate analysis (PCoA) using clr-transformed data in Aitchison distance. Each point represents a sample corresponding to a group highlighted in different colors. A) and B) represent the samples of rotifer microbiota, according to DNA and RNA, respectively; C) and D), similar to the above but in water samples.
Figure 1.
Principal coordinate analysis (PCoA) using clr-transformed data in Aitchison distance. Each point represents a sample corresponding to a group highlighted in different colors. A) and B) represent the samples of rotifer microbiota, according to DNA and RNA, respectively; C) and D), similar to the above but in water samples.
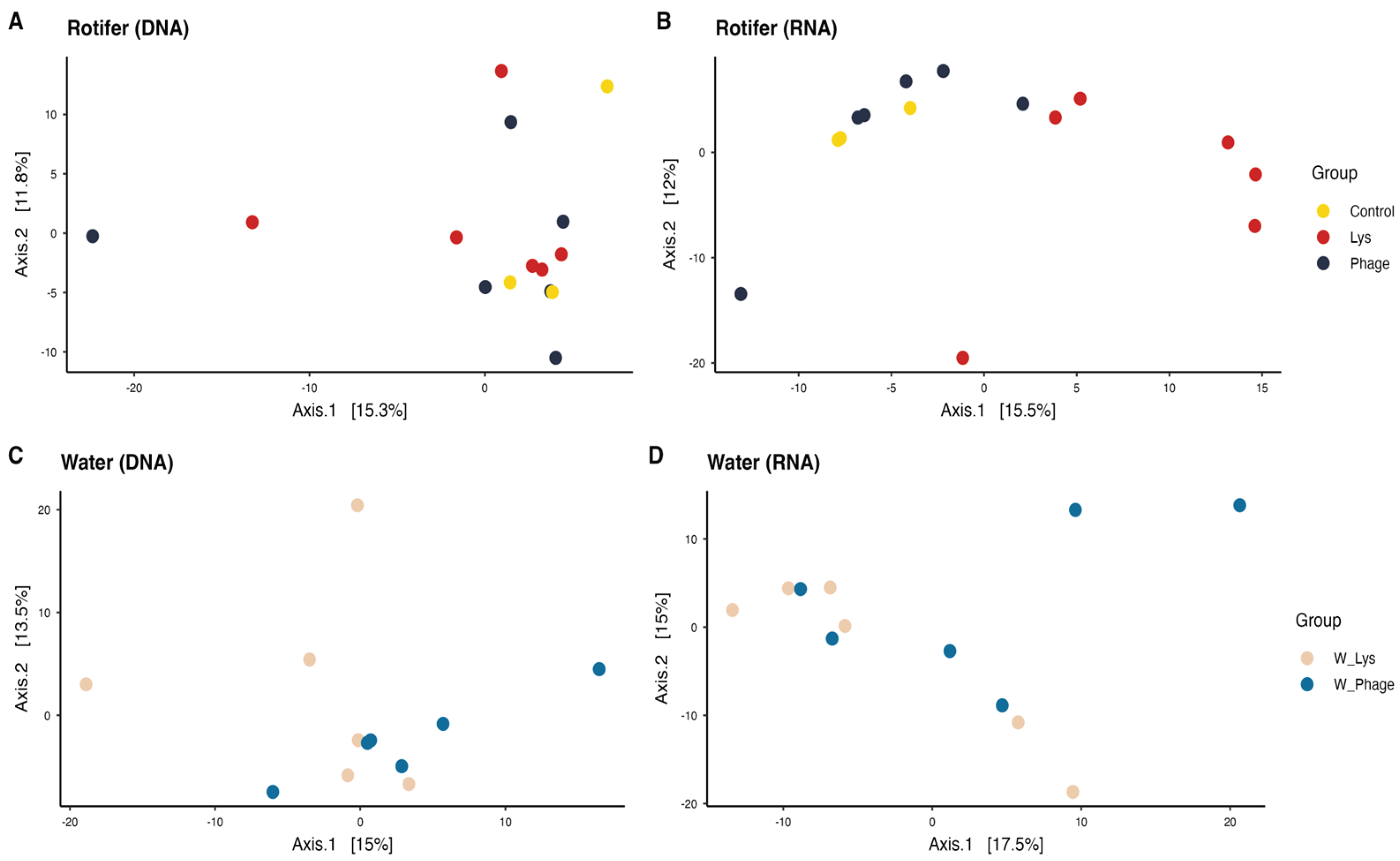
Figure 2.
Average relative abundance of major phyla represented in the rotifer and water microbiota. In this figure, phyla with an average relative abundance per group higher than 1% in at least one group were included.
Figure 2.
Average relative abundance of major phyla represented in the rotifer and water microbiota. In this figure, phyla with an average relative abundance per group higher than 1% in at least one group were included.
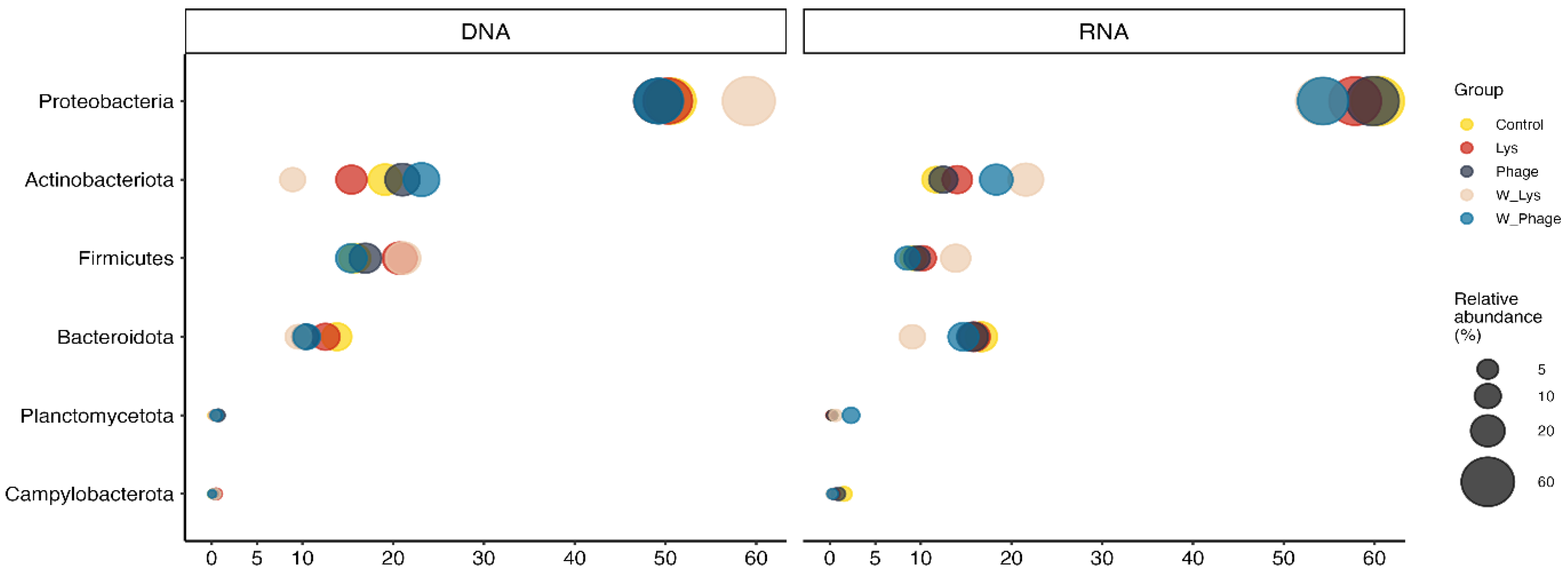
Figure 3.
Average relative abundance of major genera represented in rotifer and water microbiota. In this figure, genera with an average relative abundance per group higher than 2% in at least one group were included.
Figure 3.
Average relative abundance of major genera represented in rotifer and water microbiota. In this figure, genera with an average relative abundance per group higher than 2% in at least one group were included.
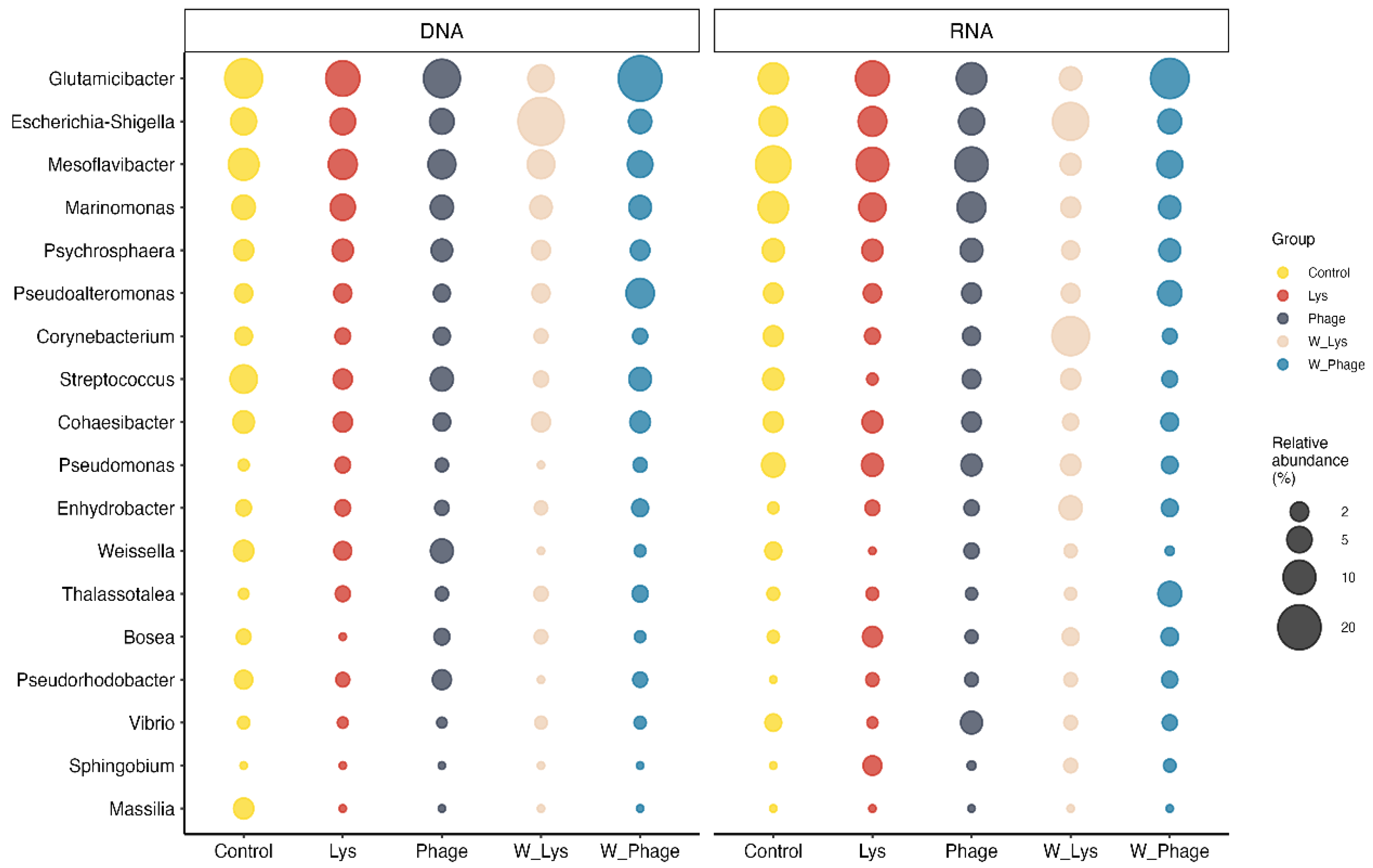
Figure 4.
Statistically significant changes at the genus level in water microbiota after exposure to phage and Lys. A) log fold change (lfc) obtained from pair comparison using ANCOM-BC2; B) relative abundance of genera that showed significant differences according to ANCOM-BC2.
Figure 4.
Statistically significant changes at the genus level in water microbiota after exposure to phage and Lys. A) log fold change (lfc) obtained from pair comparison using ANCOM-BC2; B) relative abundance of genera that showed significant differences according to ANCOM-BC2.
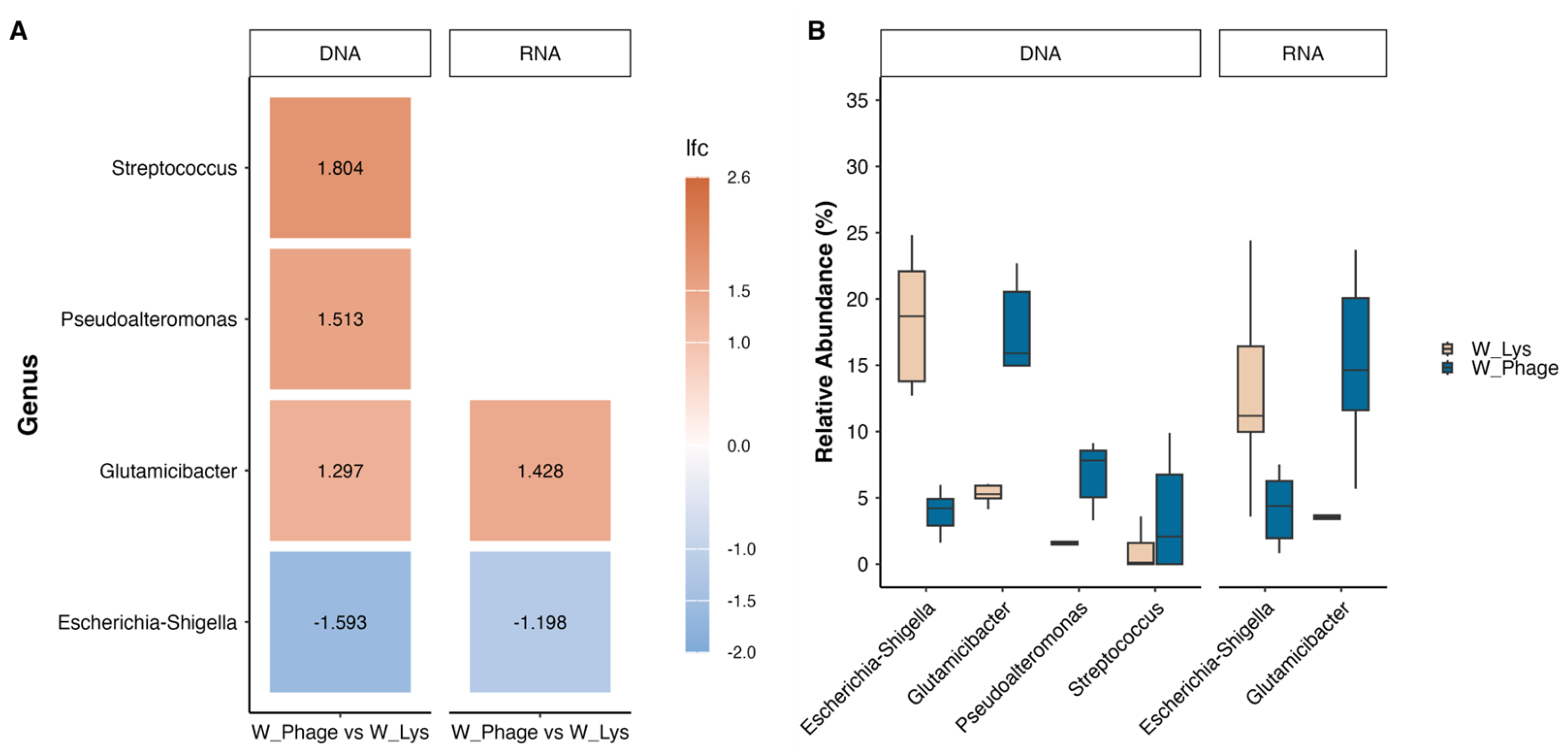
Figure 5.
Principal coordinate analysis (PCoA) using clr-transformed data in Aitchison distance. Each point represents a sample corresponding to a group highlighted in different colors.
Figure 5.
Principal coordinate analysis (PCoA) using clr-transformed data in Aitchison distance. Each point represents a sample corresponding to a group highlighted in different colors.
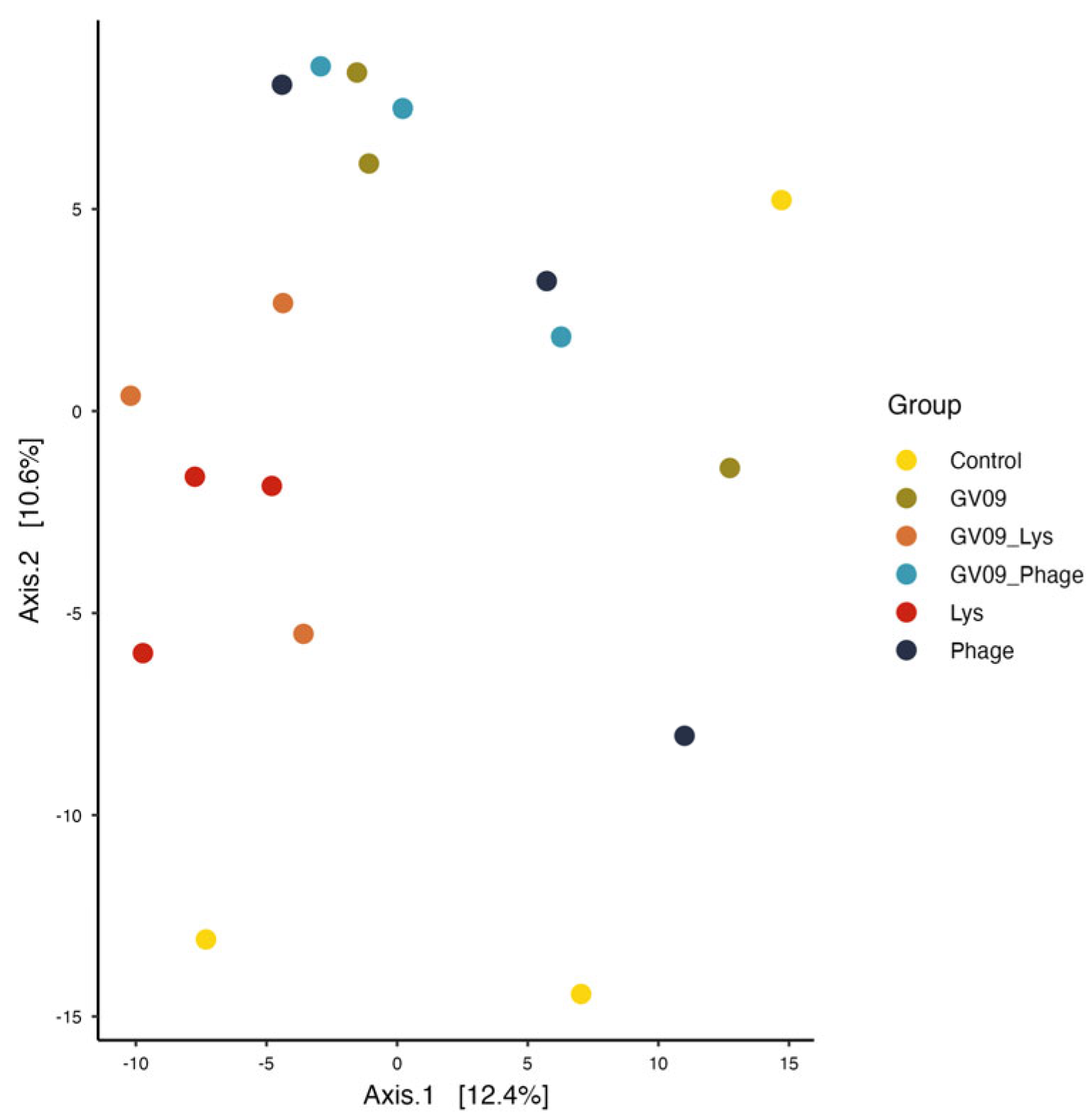
Figure 6.
Average relative abundance of major phyla represented in the zebrafish microbiota. In this figure, phyla with an average relative abundance per group higher than 1% in at least one group were included.
Figure 6.
Average relative abundance of major phyla represented in the zebrafish microbiota. In this figure, phyla with an average relative abundance per group higher than 1% in at least one group were included.
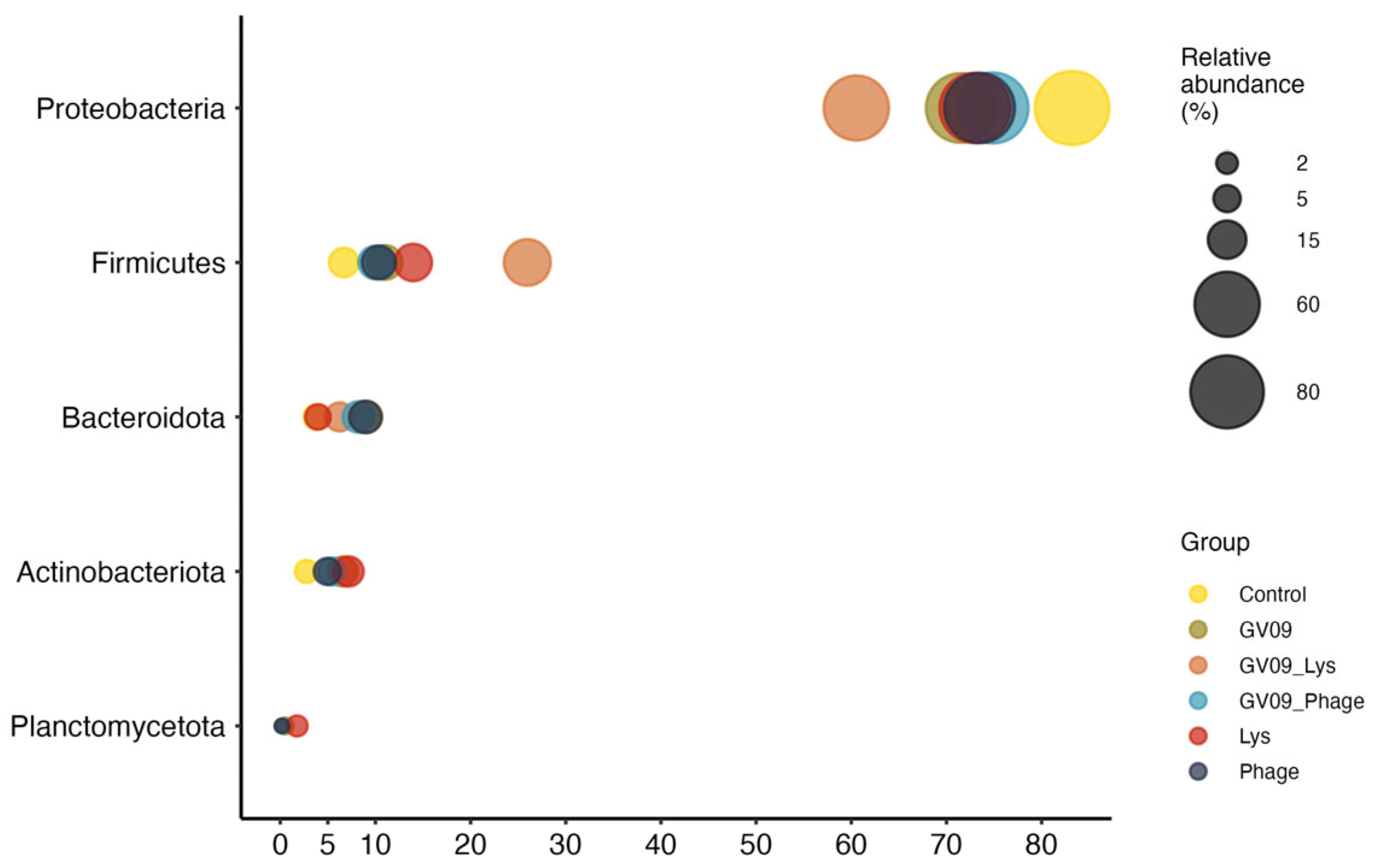
Figure 7.
Average relative abundance of major genera represented in zebrafish microbiota. In this figure, genera with an average relative abundance per group higher than 2% in at least one group were included.
Figure 7.
Average relative abundance of major genera represented in zebrafish microbiota. In this figure, genera with an average relative abundance per group higher than 2% in at least one group were included.
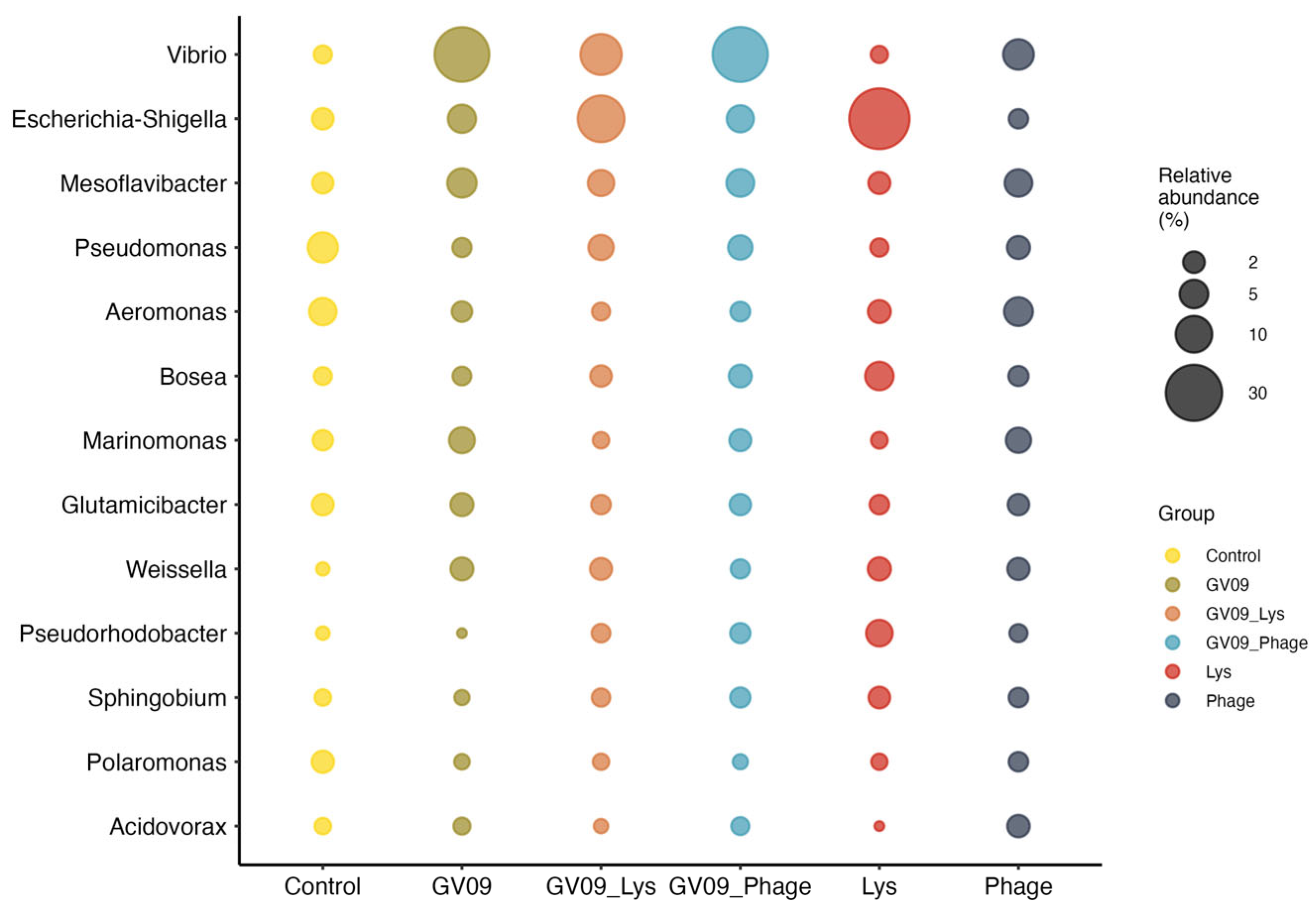

Table 1.
Summary of PERMANOVA and Betadisper results comparing microbial community structure (Aitchison distance) in rotifers and water treated with Phage and Lys.
Table 1.
Summary of PERMANOVA and Betadisper results comparing microbial community structure (Aitchison distance) in rotifers and water treated with Phage and Lys.
| Source | Group | Comparison |
PERMANOVA R2 P value |
Betadisper P value |
|
| DNA | Rotifer Water |
Lys vs Phage Lys vs Control Phage vs Control Lys vs Phage |
0.074 0.121 0.110 0.109 |
0.875 0.591 0.802 0.035 |
0.490 0.951 0.505 0.842 |
| RNA | Rotifer Water |
Lys vs Phage Lys vs Control Phage vs Control Lys vs Phage |
0.138 0.181 0.101 0.099 |
0.004 0.030 0.872 0.287 |
0.786 0.070 0.086 0.584 |
Table 2.
Summary of PERMANOVA and Betadisper results comparing microbial community structure (Aitchison distance) in zebrafish challenged with Vibrio strain GV09 and treated with Phage and Lys.
Table 2.
Summary of PERMANOVA and Betadisper results comparing microbial community structure (Aitchison distance) in zebrafish challenged with Vibrio strain GV09 and treated with Phage and Lys.
| Comparison |
PERMANOVA R2 P value |
Betadisper P value |
|
| All Lys vs Control Phage vs Control GV09 vs Control |
0.323 0.227 0.197 0.220 |
0.058 0.1 0.6 0.3 |
0.066 0.218 0.468 0.028 |
| GV09_Lys vs Control GV09_Phage vs Control Lys vs GV09_Lys Phage vs Lys |
0.245 0.234 0.208 0.240 |
0.1 0.1 0.4 0.1 |
0.026 0.031 0.474 0.496 |
| Lys vs GV09 | 0.263 | 0.1 | 0.812 |
| Lys vs GV09_Phage | 0.249 | 0.1 | 0.717 |
| GV09_Lys vs Phage | 0.241 | 0.1 | 0.064 |
| GV09_Lys vs GV09 | 0.223 | 0.2 | 0.032 |
| GV09_Lys vs GV09_Phage | 0.212 | 0.4 | 0.238 |
| Phage vs GV09 | 0.183 | 0.6 | 0.174 |
| Phage vs GV09_Phage | 0.196 | 0.6 | 0.120 |
| GV09 vs GV09_Phage | 0.189 | 0.8 | 0.546 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



