Submitted:
29 January 2026
Posted:
02 February 2026
You are already at the latest version
Abstract
Rates of Parkinson’s disease (PD) and dementia have risen globally to near-epidemic proportions. Conventional explanations including aging, toxins, genetics, and protein misfolding describe aspects of pathology but do not fully account for the synchrony of recent increases across regions. This paper explores whether chronic microbial persistence in modern food systems, particularly high-speed poultry-processing environments, may represent an underrecognized upstream contributor to PD and dementia risk. Publicly available data from the Global Burden of Disease study and FAOSTAT meat-consumption records indicate that PD incidence has increased alongside poultry consumption, especially in regions using high-throughput, chlorine-based processing such as the United States and China. In the United States, PD and dementia trends rise in near-parallel with poultry consumption, with a multi-year lag consistent with long prodromal intervals. In contrast, Israel shows stable or declining PD rates despite substantial poultry consumption, coinciding with kosher processing practices and long-standing post-BSE feed restrictions. These patterns support a microbial-ecological hypothesis in which biofilm-forming spirochetes persist through chemical sanitation, enter the food chain, and chronically stimulate gut-brain inflammation. This hypothesis paper is intended to stimulate empirical testing; all associations described are ecological and do not imply individual-level causation.
Keywords:
Parkinson’s disease
; dementia
; spirochete
; biofilm
; microbial persistence
; poultry processing
; gut–brain axis
Introduction
Parkinson’s disease (PD) and dementia together represent a rapidly growing global public health burden, with rising incidence, prevalence, and societal cost documented across multiple regions over recent decades[1,2,3,4,5]. In the United States alone, dementia costs approximately $360 billion annually, while Parkinson’s disease adds an estimated $52 billion, together exceeding $420 billion in annual economic burden[6],[7]. Despite decades of research, the fundamental cause of these disorders remains uncertain. The simultaneous acceleration of PD and dementia incidence since the late 1990s[1] suggests that modern environmental factors may be acting across populations.
Limitations of Current Theories
Prevailing frameworks—genetic susceptibility, aging, pesticide exposure [8], and spontaneous protein misfolding [9]—each explain portions of the pathology but do not yet account for the synchronized global timing or the striking regional differences in disease trajectories. The classical prion hypothesis attributes neurodegeneration to self-propagating misfolded proteins [9]; however, similar aggregates are seen in chronic inflammatory and microbial conditions [10,11,12], suggesting that they may represent a downstream manifestation rather than the initiating cause. Current explanatory frameworks for Parkinson’s disease and dementia do not yet fully account for certain emerging epidemiologic patterns—patterns that are examined in subsequent sections of this paper.
The Prion Paradigm Revisited
The prion theory—recognized with the 1997 Nobel Prize for proposing a protein-templating mechanism to explain rare transmissible spongiform encephalopathies (TSEs) such as BSE/vCJD—asserts that misfolded proteins propagate by conformational templating rather than replication [9]. Yet this mechanism cannot account for the modern, non-transmissible rise in Parkinson’s disease and dementia. Multiple studies suggest that prion-like α-synuclein and β-amyloid aggregates may arise downstream of chronic inflammation rather than act as initiating causes [10,11,12,13]. By contrast, spirochetes—ubiquitous, biofilm-forming, gut-colonizing pathogens—were largely missed by the limited microbiologic techniques available in the 1980s–1990s, when the prion paradigm took hold [11].
Modern PCR and immunohistochemical studies have identified Treponema DNA in Alzheimer’s brain tissue [14], and spirochetes have been visualized within β-amyloid plaques in multiple neuropathologic studies [11], [12]. Brachyspira has been detected in poultry gut [15], underscoring its relevance as a food-chain exposure. Oral bacteria may induce α-synuclein pathology through molecular mimicry. Unlike prions, spirochetes multiply, persist, and chronically activate neuroinflammatory pathways—providing a biologically coherent upstream trigger for misfolding cascades.
A New Ecological Hypothesis
This paper explores whether microbial persistence in industrial food systems may represent the missing ecological link between environmental change and neurodegeneration. Biofilm-forming spirochetes and other anaerobes in processing environments could act as latent infectious stimuli driving neurodegenerative disease [15,16,17,18]. These organisms can survive chemical disinfection, colonize the gut, and elicit sustained inflammation via the gut–brain axis [19,20,21].
Spirochetes have been observed in patterns consistent with Koch’s postulates and Hill’s criteria, though definitive proof remains to be shown [11]. Miklossy’s studies [11] demonstrated Treponema within β-amyloid plaques and cortical tissue in neurodegenerative brains [12]—histopathologic evidence suggesting a microbial component that warrants further investigation. Experimental models further show that intestinal bacteria can initiate α-synuclein pathology and motor impairment, supporting biologically plausible gut–brain mechanisms [19].
Industrial Context and Global Timing
The late-20th-century industrialization of poultry processing created ecological conditions favoring microbial persistence—accelerated line speeds, organic-rich environments, and reliance on chlorine-based antimicrobials [22,23,24]. Beginning in the late 1990s and early 2000s, poultry-processing systems—particularly in the United States—expanded production volumes and adopted progressively faster processing speeds in response to rising demand [22], [23]. Although specific throughput limits differ by country, the overall trend toward higher line speeds and greater mechanization is well documented across FAO production statistics [2] and U.S. federal inspection reports [24]. This global acceleration provides ecological context for interpreting the parallel rise in neurodegenerative-disease burden, consistent with long-term trends reported in GBD 2016 analyses [25].
Regional contrasts further reinforce this pattern. Israel reports declining or stable Parkinson’s-disease incidence despite substantial poultry consumption [26], while Germany likewise demonstrates modestly declining PD incidence and prevalence in recent administrative studies [27], [28]. Dementia surveillance in Germany shows comparable downward trends over recent years [29], and Israel’s dementia studies similarly indicate relatively low or stable age-standardized rates compared with many Western nations [30]. Global Alzheimer’s projections also show heterogeneous regional trajectories that may reflect differences in environmental or food-system exposures rather than aging alone [31].
In Germany, stringent feed sterilization requirements, rendering controls, and non-chlorine sanitation approaches—codified in national poultry-hygiene standards—further limit microbial persistence [32]. These differences provide a useful contrast to regions such as the United States and China, where chlorine-based systems and high-throughput processing remain dominant [22,23,24].
The United States remains one of the few major poultry-producing regions that continues to use predominantly chlorine-based antimicrobial systems, whereas countries with stable or declining neurodegenerative trends—such as Israel and Germany—employ salt-, heat-, or peracetic-acid–based sanitation methods [24,25,26,27,28,29,30,31,32]. These differences suggest that sanitation chemistry—rather than consumption volume alone—may influence long-term microbial-exposure risk [16,17,18].
Objectives of This Paper
- Present global and regional evidence describing how PD and dementia incidence align with changes in poultry production and processing systems.
- Integrate mechanistic pathways illustrating how microbial persistence could contribute to neuroinflammatory and protein-misfolding cascades.
- Compare sanitation strategies across regions (e.g., chlorine versus ozone, UV, or salt-based methods) to identify potentially modifiable environmental factors.
- Propose public-health actions for microbial surveillance and clinical screening that could support hypothesis testing.
Together, these aims outline a testable framework that unites multiple theories under a single ecological model: infection first, protein misfolding second, disease last.
Observational Evidence
Global Patterns
Global surveillance data show that Parkinson’s-disease incidence and poultry consumption have risen in close synchrony since the early 1990s [1]. Earlier Global Burden of Disease (GBD) 2016 analyses also reported a worldwide rise in age-standardized PD incidence and prevalence across nearly all regions, confirming that the upward trajectory pre-dates the 2021 update [25]. Globally, PD age-standardized prevalence increased roughly 39% from 1990 to 2021, [1] with GBD 2016 reporting a similarly steep rise (incidence +39%, prevalence +118%, 1990–2016) [25]. This period coincides with a worldwide acceleration in poultry-processing throughput—including higher line speeds, expanded industrial slaughter capacity, and increased chemical-based sanitation—which multiple countries adopted between the mid-1990s and early 2000s [23,24,25,26]. Germany and Israel have partially decoupled from this pattern despite high-quality epidemiologic data and divergent trends [26,27,28,29,30,31,32], contrasting with the broader global rise.
Where national incidence data are unavailable or unstable, prevalence or modeled burden estimates are used for longitudinal comparison; trends are interpreted directionally rather than as absolute risk measures.
Initial exploratory analyses examined multiple animal-protein categories, including beef and pork, given longstanding hypotheses linking red meat or saturated fat to neurodegenerative risk. However, no comparable temporal alignment was observed for beef or pork consumption. Poultry emerged as the clearest and most internally consistent ecological signal, prompting further examination of poultry production and processing history.
Figure 1 illustrates long-term U.S. trends in Parkinson’s-disease incidence using age-standardized rates from GBD 2021 [1] compared with per-capita chicken, beef, and pork consumption from FAOSTAT (2024) [2]. PD incidence and poultry intake follow a broadly parallel upward trajectory from 1990–2020, whereas beef and pork consumption remain stable or declining.
As with all ecological data, these population-level trends may reflect demographic shifts, diagnostic sensitivity, environmental exposures, or food-system factors acting in combination. Individual-level studies are needed to clarify underlying mechanisms. Ecological comparisons can sharpen hypotheses but cannot determine individual-level risk without laboratory, environmental, or patient-level confirmation.
Geographic comparisons further show that long-term Parkinson’s-disease trends differ across countries with distinct processing speeds, sanitation chemistries, and regulatory approaches (see Figure 2 in Section 2.2). These contrasts underscore the importance of examining regional hygiene standards, processing environments, and microbial-control strategies when interpreting international variation in neurodegenerative-disease patterns.
Data source: IHME Global Burden of Disease 2021 [1]. Values represent age-standardized incidence per 100 000 population.
Recent German administrative data show a modest decline in age-standardized prevalence of Parkinson’s disease (0.38% → 0.29%, 2017–2022) and dementia (≈3.4% → 2.8% among adults ≥40 years) [32]. These values reflect insurance-claims prevalence rather than incidence, but the directional pattern aligns with Germany’s broader stability in neurodegenerative-disease indicators and with global trends reported in GBD 2021 [1]. Although ecological and non-causal, these observations highlight the potential relevance of regional differences in poultry-processing speed, sanitation methods, and feed regulations (Section 2.2).
Germany’s long-term pattern closely parallels Israel’s and offers an additional example of a stable neurodegenerative-disease trajectory within a high-income setting. Germany stands out within Western Europe for its relative stability despite broad diagnostic coverage [28], [29], and its performance compares favorably with most Western EU member states, many of which show steadily rising long-term trajectories. This parallel to Israel [26], [30] has raised interest in whether regional sanitation and processing approaches may contribute to these ecological differences. Germany relies predominantly on peracetic-acid or hot-water sanitation across most poultry facilities, with minimal use of chlorine-wash systems [32] and operates under EU-regulated throughput speeds that remain substantially lower than the highest U.S. line rates [24], [32]. Germany additionally maintains strict antibiotic-stewardship programs and comprehensive feed and veterinary-control regulations that exceed EU baselines.
These characteristics contrast with the high-throughput, predominantly chlorine-based systems used in the United States and China, where GBD 2021 data show steeper long-term increases in PD and dementia [1]. Although these ecological contrasts cannot identify causal relationships, they highlight meaningful differences in processing environments. Israel’s use of kosher salting [3,4,5] and Germany’s reliance on peracetic-acid and hot-water sanitation represent differing but similarly rigorous microbial-control frameworks. These convergent approaches may help explain their relatively stable neurodegenerative-disease patterns and warrant further comparative study.
Collectively, these contrasts suggest that cross-national differences in sanitation philosophy, processing environments, and microbial-control strategies may be as relevant as consumption volume when interpreting long-term epidemiologic patterns. The United States and China rely heavily on high-throughput poultry-processing systems, with U.S. facilities operating under NPIS line-speed allowances and chlorinated antimicrobial rinses as described in federal sanitation rules [23], [24] and FAOSTAT production reports [2].
By contrast, Japan maintains more moderate processing throughput under MAFF hygiene standards and relies primarily on physical or thermal sanitation rather than chemical-immersion systems—an approach consistent with its precautionary philosophy toward poultry disinfection. These regulatory and hygiene differences occur against the backdrop of Japan’s comparatively low and stable neurodegenerative-disease trends [30], [31]. Several Northern European countries that employ peracetic-acid or hot-water sanitation show similarly stable or only gradually changing PD trajectories in GBD 2021 estimates [1]. Together, these regional contrasts highlight how sanitation chemistry, processing environments, feed regulations, and broader environmental exposures can create distinct microbial ecologies that warrant deeper mechanistic investigation.
Because Parkinson’s disease is strongly age-related, cumulative lifetime exposures—dietary or environmental—may interact synergistically with aging rather than operate independently, a possibility raised in multiple ecological comparisons. In the United States, chlorine-based disinfection remains the predominant processing method, with only limited adoption of oxidative or peracetic-acid systems since 2020 in selected facilities. Israel’s long-standing reliance on kosher salting [3,4,5] represents a distinct hygiene strategy that merits comparative evaluation.
Regional Contrasts

Table 1.
Poultry-System Hygiene and Parkinson’s-Disease Incidence Trends (1990–2021).
| Country / Region | Hygiene Strategy | PD Trend (EAPC / ASIR) | Dementia Trend (Prevalence / EAPC) | Data Source |
|---|---|---|---|---|
| Germany | Predominantly peracetic-acid / hot-water sanitation; strict feed controls; low antibiotic reliance | Modest decline in claims-based PD prevalence (0.38% → 0.29%, 2017–2022) [28] | Decline in claims-based dementia prevalence (~3.4% → 2.8% among adults ≥40, 2017–2022) [29] | [28,29,32] |
| Israel | Kosher salting and rinsing; feed prohibitions (post-BSE reforms) | Stable to slightly declining PD incidence in national surveillance reports [26] | Stable to modestly changing dementia incidence in EHR-based national studies [30] | [3,4,5,26,30] |
| United States / China | Chlorine-based disinfection; high-throughput processing lines | Long-term increase in PD incidence (significant positive ASIR slope, 1990–2021) [1] | Increasing dementia burden (substantial modeled rise, 1990–2019) [1] | [1] |
Data source: IHME Global Burden of Disease 2021 [1]; RKI Journal of Health Monitoring (2025) [29,32]; national agricultural reports [21]. Values represent age-standardized incidence or prevalence per 100 000 population. Israel’s high-throughput but kosher-regulated poultry system differs from the chlorine-based or antibiotic-intensive processing approaches used in countries with larger PD increases; these regional contrasts are ecological and should be interpreted within that context [3,4,5,26,30].
Unified Neurodegenerative Trends
Projections for 2023–2025 are short-term extrapolations drawn from FAOSTAT [2], GBD 2021 [1], and National Chicken Council trend datasets [22]. These values are illustrative only and will be refined as new surveillance data emerge.
In the United States, both Parkinson’s disease and dementia prevalence have risen in near-lockstep with per-capita chicken consumption over the past three decades (Figure 3). Data from the National Chicken Council [23], the Alzheimer’s Association⁶, and the Parkinson’s Foundation⁷ show dementia prevalence climbing from 3.7 to 6.9 million and Parkinson’s prevalence from 0.53 to 1.0 million while broiler-meat intake increased from approximately 61 to 101 lb per person per year [23]. The United States now produces more than 9.3 billion broilers annually, with production heavily concentrated in southeastern and Midwestern states [33]. These parallel trajectories align with the long prodromal phase typical of neurodegenerative disorders and therefore merit targeted laboratory confirmation.
A notable contrast emerges in feed-chain policy. In the United States and China, intra-species recycling of animal by-products remains legal: rendered poultry and fish meals may still be fed to poultry, and feather meal is permitted in aquaculture [34]. U.S. regulations (21 CFR 589.2000; 2008 revision) prohibit only ruminant-to-ruminant protein, leaving non-ruminant loops intact [35]. Such practices preserve potential pathways for microbial persistence and amplification within the broader food-chain ecology—pathways that Israel permanently severed in 1996 through its comprehensive mammalian-protein feed ban [3,4,5,17].
These population-level patterns, taken together, reinforce the hypothesis that modern poultry-production systems may contribute an under-recognized environmental exposure relevant to the rising burden of Parkinson’s disease and dementia. Although the ecological signal is strongest and most coherent for poultry—owing to its uniquely rapid global industrialization, highest per-capita consumption growth, and most complete production datasets—this does not preclude similar microbial-persistence dynamics in other high-volume meat systems (including pork, beef, or aquaculture). Poultry simply provides the clearest natural experiment available at present.
Statistical Concordance—Across multi-year national aggregates, poultry consumption correlates strongly with PD and dementia burden. Pearson correlations range from r ≈ 0.9–0.95, reflecting population-level associations rather than individual-level causation. PD and dementia values represent modeled U.S. prevalence from GBD 2021 [1], and poultry supply reflects FAOSTAT carcass-weight availability converted to lb/person/year [2]. Beef and pork consumption remained stable or declined during the same period [2]. These correlations are descriptive but may help prioritize hypotheses for further mechanistic and environmental research.
External Epidemiologic Support—Independent epidemiologic studies report similar directional relationships between meat intake and neurodegenerative outcomes, providing context for the U.S. temporal patterns shown in Table 2. For example, cohort and meta-analytic studies have linked higher consumption of processed or animal-derived meats with increased dementia risk [37,38].
U.S. consumption trends compiled by the USDA Economic Research Service (ERS) show long-term increases in poultry availability per capita over the same interval in which national Parkinson’s-disease and dementia burdens have risen [36]. These national production and availability trends offer a population-level backdrop for interpreting the correlation analyses presented in Table 2.
Tong et al. conducted a systematic review and meta-analysis across 204 countries and found a positive ecological correlation between global meat supply—including poultry and processed meats—and dementia prevalence (r = 0.45; p < 0.001) [37]. In a large prospective cohort from the UK Biobank, Zhang et al. reported that higher processed meat consumption was associated with significantly increased risk of incident dementia, demonstrating a robust longitudinal dose–response pattern (hazard ratio 1.44, 95% CI 1.24–1.67) [38].
Figure 3 illustrates long-term U.S. trends in dementia prevalence, Parkinson’s-disease prevalence (scaled ×10 for visualization), and per-capita chicken consumption from 1990–2024. All three measures show sustained upward trajectories with notable accelerations after the mid-2000s. The temporal alignment provides relevant context for evaluating food-system and processing-environment hypotheses discussed in Section 2.2.
Blue line: dementia prevalent cases (millions). Green line: Parkinson’s-disease prevalence (millions), multiplied by 10 for visual comparison. Orange line: U.S. chicken consumption (lb/person/year, right axis).
Data sources: Global Burden of Disease Study 2021 (PD prevalence) [1]; Alzheimer’s Association 2024 (dementia cases) [6]; FAOSTAT 2024/National Chicken Council (chicken consumption) [2,22]. Values for 2022–2024 represent linear extrapolations. This visualization illustrates converging upward U.S. trajectories but does not imply causation.
Interpretation—The parallel rise of U.S. Parkinson’s-disease prevalence, dementia prevalence, and per-capita chicken consumption from 1990–2024 is compatible with—but does not imply—the possibility of a shared long-latency environment exposure. Notably, the acceleration of Parkinson’s disease and dementia prevalence occurs approximately one to two decades after major late-1990s changes in U.S. poultry-processing systems, a temporal offset that is consistent with long-latency exposure models rather than contemporaneous effects. Both neurodegenerative curves show a clear upward inflection after the late 1990s—timing that coincides with major changes in U.S. poultry-processing throughput, including the 1996 HACCP rule and its 1997 pilot implementation and subsequent adoption of higher line speeds in young-chicken plants [23]. These changes expanded total volume processed per facility and increased reliance on chemical antimicrobial systems, conditions that may promote survival of biofilm-forming organisms under certain high-load environments [16,17,18].
Although Figure 3 presents U.S. data only, international contrasts offer useful context. While many U.S. facilities have incorporated peracetic-acid–based antimicrobial blends since roughly 2016, chlorine-based immersion and spray systems remain widely authorized and used, particularly in high-throughput plants. Most high-income countries report rising PD and dementia burdens over the same period [1,6], whereas Israel’s age-standardized trends remained relatively stable following its comprehensive 2001 mammalian-protein feed ban and continued use of kosher salting and rinsing as primary sanitation methods [26,30].
While socioeconomic factors, aging, and improved diagnostic surveillance influence all neurodegenerative-disease estimates, the near-synchronous U.S. rise of Parkinson’s disease and dementia suggests the possibility of overlapping gut–brain inflammatory pathways (e.g., microbially driven α-synuclein induction or neuroinflammatory priming) [19,20,21]. The temporal alignment and latency-compatible trajectories highlight the importance of examining whether industrial-scale poultry-processing environments—particularly those operating at accelerated throughputs—may contribute to persistent microbial exposures reflected in long-term disease trends.
Regional Overlap: The “Chicken Belt” and Neurodegenerative Hotspots
The proposed “chicken belt”—a region encompassing major poultry-producing states such as Georgia, Alabama, Mississippi, Arkansas, and North Carolina—accounts for a substantial share of U.S. broiler production according to USDA National Agricultural Statistics Service data [33]. These states together produce roughly half of all U.S. broilers, placing them among the highest-volume poultry-processing regions in the country.
Patterns of Parkinson’s disease (PD) and Alzheimer’s disease (AD) burden appear elevated in parts of this region relative to national baselines, based on available incidence and mortality estimates [1,6]. Several of these high-production states—along with portions of the Midwest, including Ohio and Indiana within the historically recognized “PD belt”—show PD rates exceeding the U.S. age-standardized national average of 15.6 per 100,000 based on GBD 2021 estimates [1].
Importantly, if exposure arises through centralized, high-throughput processing and modern food-distribution systems, disease incidence would not be expected to cluster tightly around production or processing regions. Instead, exposure would be distributed diffusely across downstream consumer populations, producing a geographically “peppered” pattern that does not map cleanly onto agricultural or industrial locations. Such diffusion effects are well documented in food-system–mediated exposures and are compatible with the heterogeneous regional patterns observed for Parkinson’s disease and dementia.
While these spatial associations do not establish causation, the overlap between regions of high-volume, chemically intensive poultry processing and elevated neurodegenerative-disease burdens supports the hypothesis that microbial persistence in food-production environments may contribute to regional variation in disease trajectories. This convergence underscores the need for targeted investigation of environmental and clinical microbial exposures.
Following the BSE-related feed-chain failures of the 1990s—which demonstrated how feedback loops in industrial animal-protein systems can silently amplify pathogens before detection—Israel enacted one of the world’s strictest prohibitions on feeding mammalian meat-and-bone meal (MBM) to livestock, poultry, or fish. The nationwide ban, implemented in 1996 and reinforced thereafter, remains in force today [5].
Under this 1996 reform, all mammalian MBM—including bovine and ovine sources—was barred from use in feed for every animal sector to prevent cross-species transmission of prion or microbial agents [5]. A 2001 amendment further extended the restriction by prohibiting poultry- and fish-derived MBM in ruminant feed [5]. These combined measures effectively eliminated animal-protein recycling in Israel’s feed system and are widely credited with reducing opportunities for microbial adaptation and prion-related disease risk.
In contrast, the United States and China continue to permit intra-species recycling of animal by-products, particularly poultry and fish meals in non-ruminant feed [34,35,39]. U.S. regulations (21 CFR 589.2000; 2008 revision) restrict only ruminant-to-ruminant feeding, allowing rendered poultry and swine proteins in poultry diets as well as feather meal in aquaculture [34,35].
These divergent feed-chain policies illustrate how Israel’s post-BSE reforms fundamentally disrupted protein-recycling pathways that can sustain cross-species transmissible-agent risk. By comparison, systems that continue to allow intra- and inter-species recycling may preserve microbial or prion feedback loops within the broader food-chain ecology.
Whereas many countries—including the United Kingdom and the United States—reported high Brachyspira colonization rates in poultry flocks from the mid-1990s through the 2010s (UK surveys documenting 46–71% flock prevalence [15]), Israel has maintained stricter feed-composition controls since the late 1990s. Although Brachyspira remains detectable worldwide, Israel’s comprehensive post-BSE prohibition on mammalian meat-and-bone meal in all animal feed (implemented in 1996 and fully enforced through subsequent amendments) [3,4,5,17], together with the continued use of kosher salting and rinsing as primary sanitation methods, coincides with relatively stable age-standardized Parkinson’s-disease and dementia trends compared with most high-income nations [26,30]. While these ecological contrasts do not establish causation, the alignment of stricter feed-chain policies and sanitation practices with more favorable neurodegenerative-disease trajectories warrants further investigation into potential food-system contributions to long-term population health.
According to Global Burden of Disease 2021 estimates, Israel’s age-standardized Parkinson’s-disease incidence rate (~21.9 per 100 000; 95% UI 18.5–25.7) is slightly lower than rates reported in the United States (~20.5 per 100 000) and moderately lower than those observed in China (~24.3 per 100 000) [1]. Israel also reports relatively low, stable age-standardized dementia prevalence compared with many high-income nations, based on national EHR-based surveillance [30]. These patterns coincide with Israel’s earlier and more comprehensive interruption of animal-protein-recycling pathways—reforms enacted beginning in 1996 and strengthened through subsequent feed-control amendments—which may reduce opportunities for persistent microbial exposure within food-chain environments [3,4,5].
Taken together, these regional contrasts outline an ecological framework suggesting that long-term neurodegenerative trends may be shaped not only by demographic and diagnostic factors but also by variation in sanitation practices, processing environments, and feed-system microbial-persistence dynamics.
In contrast, dietary patterns may offer further insight into this ecological framework. Vegetarian and low-poultry-consuming populations provide useful comparison points: participants in the Adventist Health Study-2, who consume little or no poultry, demonstrate substantially lower Parkinson’s-disease risk (approximately 30–50% lower incidence in cohort analyses) [40]. India—where per-capita poultry intake remains among the lowest globally—also reports age-standardized PD rates markedly lower than those observed in many Western nations [25]. This pattern is consistent with large cohort evidence from Zhang et al. [38] showing that higher processed-meat consumption predicts increased dementia risk, providing an additional dietary contrast relevant to poultry-associated exposures.
These observations cannot identify mechanisms, but they are directionally consistent with international sanitation and processing differences and suggest that both dietary patterns and environmental exposures may contribute to regional variation in long-term neurodegenerative trends.
Mechanistic Hypotheses Bridging PD and Dementia
Microbial Persistence and Biofilm Adaptation
Many bacterial species that thrive in food-processing environments—particularly spirochetes and Brachyspira—form resilient biofilms that protect them from heat, desiccation, and commonly used disinfectants. Brachyspira spp., a key group of biofilm-forming spirochetes, expanded markedly in poultry flocks during the 1990s–2000s, with United Kingdom surveys reporting 46–71% prevalence across 222 flocks [15]. Several poultry-associated Brachyspira species—including B. pilosicoli—are established zoonotic pathogens capable of colonizing humans and forming mucosal biofilms, underscoring their relevance for evaluating long-latency microbial exposures in food-chain environments.
Under sublethal chemical exposure, these organisms exhibit hormetic adaptation, up-regulating stress-response pathways and increasing extracellular-matrix production [18]. Repeated exposure to chlorine or peroxide can therefore select for strains with enhanced environmental tolerance [16,17]. Once established, such biofilms can persist through sanitation cycles and continually re-seed downstream surfaces, equipment, and waste streams [16,17]. This dynamic is illustrated conceptually in Figure 4.
Biofilm-forming spirochetes exposed to repeated sublethal stressors (e.g., chlorine, peroxide, heat, desiccation) may undergo hormetic adaptation, strengthening survival within processing and environmental reservoirs. Green arrows denote potential intervention points—antibiotic avoidance, hygienic litter and wastewater management, and high-tech environmental-control systems—that interrupt the microbial-persistence loop and reduce downstream exposure risk.
Supporting evidence for post-processing persistence comes from a Belgian retail study that recovered viable Brachyspira species—including B. pilosicoli from hundreds of spent laying-hen carcasses purchased directly from supermarkets, confirming survival beyond industrial processing and consumer distribution [45]. Ingested resilient microbes can colonize the gut, alter vagal signaling, and chronically activate immunity—key elements of the gut–brain axis [19,20,21]. Over time, this inflammatory feedback can promote misfolding of α-synuclein and, in some models, β-amyloid, thereby providing a plausible biological link between persistent gastrointestinal microbes and neurodegenerative pathology [19,20,21].
Gut–Brain Axis and Protein Misfolding
The gut–brain axis provides a biological conduit through which environmental microbes may influence the central nervous system. Experimental models show that intestinal dysbiosis and microbial metabolites can trigger α-synuclein aggregation within the enteric nervous system and promote its propagation along vagal pathways [19]. Similar inflammatory mechanisms have been implicated in broader neurodegenerative processes, including amyloid and tau pathology, linking persistent gut inflammation with cortical protein misfolding [20,21].
Within this integrated framework, persistent microbial signaling from the gut is hypothesized to act as an early or contributing biological stimulus that can initiate or amplify α-synuclein and β-amyloid misfolding. This perspective is compatible with prion-like propagation theory by positing that misfolded proteins function as downstream amplifiers of a chronic inflammatory cascade originating in the gastrointestinal tract, rather than solely as autonomous initiating agents [13,19,20,21]. It also aligns with established toxin- and oxidative-stress pathways, because sustained microbial activity can drive cytokine release, oxidative stress, and neurotoxic metabolite production—mechanisms that are consistent with the long-latency progression observed in both Parkinson’s disease and dementia [19,20,21].
Revisiting BSE and vCJD Archives
Classical prion theory was developed to explain the transmissible spongiform encephalopathies, including bovine spongiform encephalopathy (BSE) and its human counterpart, variant Creutzfeldt–Jakob disease (vCJD)⁹. Within the spirochetal-persistence framework, these conditions warrant re-examination because the neuropathologic features long attributed solely to prion activity—spongiform change, gliosis, and amyloid deposition—also occur in chronic inflammatory and microbial diseases, including spirochetal infections [10,11,12].
This overlap raises the possibility that microbial persistence may contribute to prion-like neurodegeneration, either as an initiating event or as a parallel inflammatory process that amplifies protein misfolding. Historical BSE and vCJD tissues therefore represent a valuable archive for modern molecular investigation.
Archived formalin-fixed, paraffin-embedded brain and lymphoreticular specimens from vCJD cases and UK prevalence surveys are available for re-examination using contemporary molecular or histologic methods [11,14]. Although sensitivity may be reduced by DNA fragmentation and cross-reactivity among spiral-shaped taxa, even limited recovery of microbial DNA or ultrastructural signatures consistent with spirochetes would justify further targeted studies using freshly collected samples.
Direct molecular screening of archived vCJD tissues, along with comparable analyses of Alzheimer’s disease specimens, could help determine whether persistent bacteria contribute to prion-like neurodegeneration. Such work would provide an empirical test of the hypothesis that microbial persistence may underlie or interact with protein-misfolding pathways traditionally attributed to prions alone.
Integrative Perspective
Taken together, these mechanisms frame PD and dementia as differential outcomes of a shared microbe–inflammation–misfolding axis. The proposed model bridges four principal theories.
This table outlines how leading mechanistic theories of Parkinson’s disease and dementia align with the microbial-persistence model. Rather than competing explanations, these frameworks operate along a shared continuum linking microbial ecology, host immunity, and protein misfolding.
Other microbial or environmental factors—such as viral agents, air pollution, or metal exposure—may also contribute to neurodegenerative risk. The present hypothesis does not exclude these variables but proposes that microbial persistence in food systems offers an upstream ecological context that can incorporate and interact with them.
The convergence of these frameworks suggests that interrupting microbial adaptation within the food chain could simultaneously modulate multiple mechanistic layers. While age, genetics, and toxin exposures each contribute measurably to risk, the marked temporal and geographic alignment between the expansion of industrial poultry systems and rising neurodegenerative burdens [1,2,22] warrants renewed evaluation of microbial-persistence pathways—particularly spirochetes—as a potential upstream factor.
Approximate OR and r values derived from representative meta-analyses and trend correlations cited in text. These figures illustrate relative association strength, not causation. The spirochetal-persistence hypothesis remains a testable unifying framework linking chronic infection, inflammation, and neurodegeneration.
Discussion and Public-Health Implications
Synthesis of Findings
Table 3 and Table 4 indicate that diverse risk frameworks—aging, genetics, environmental toxins, and gut–brain signaling—can be accommodated within a unified ecological model centered on microbial persistence. The marked temporal and geographic alignment between the global expansion of industrial poultry production and rising crude burdens of Parkinson’s disease and dementia [1,2,22], together with the contrasting stability observed in regions with stricter feed-chain controls, supports prioritizing direct investigation of microbial-persistence pathways.
The convergence of these data suggests that persistent, biofilm-forming spirochetes may represent an upstream variable capable of linking multiple mechanistic theories along a single continuum: exposure → persistence → chronic inflammation → protein misfolding → neurodegeneration. This integrative framework unifies ecological and mechanistic evidence and sets the stage for the policy and research priorities outlined in Section 4.
Across global and national datasets from the 1990s onward, Parkinson’s disease (PD) and dementia prevalence trends parallel the expansion of industrial poultry production [2,22,33]. The observed 10–15-year lag between major poultry-processing reforms (e.g., the 1996 HACCP rule and the 1997 high-line-speed pilot programmes²³) and the steepening of PD and dementia incidence curves aligns with the known prodromal phases of these disorders [13,39,40]. These patterns represent population-level temporal alignments that inform, but do not determine, underlying biological mechanisms.
Both disorders exhibit progressive neuroinflammation, mitochondrial impairment, and misfolded protein pathology, all of which could be triggered or amplified by persistent infection. Spirochetal and other biofilm-forming bacteria could act at this upstream stage by sustaining low-grade inflammation and promoting misfolded-protein propagation.
Ecological analyses remain constrained by confounders such as reporting variability, aging demographics, and dietary differences. Temporal alignment alone cannot establish causation. Poultry-consumption figures for 2023–2025 also include projections. Even so, contrasts such as Israel’s stricter feed controls and salt-based sanitation practices [3,4,5,26,30] highlight how differences in sanitation ecology may modulate microbial-exposure risk and reinforce the need for direct microbial testing in food-processing and clinical settings.
Policy and Research Implications
If spirochetal persistence proves contributory, prevention should focus on microbial survival dynamics rather than intensifying chemical sanitation. Excessive reliance on chlorine-based disinfectants can paradoxically induce bacterial hormesis and select for more resistant biofilms [16,18]. In contrast, non-chemical or oxidative approaches—kosher salting and rinsing practices [4], ozone, UV-C, and cold atmospheric plasma—have been shown to inactivate both planktonic cells and established biofilms without promoting adaptive resistance [37,38]. Shifting poultry-processing sanitation toward these methods may therefore offer a more effective strategy for reducing long-term microbial-exposure risk.
Cold plasma, a non-thermal ionized gas that generates reactive oxygen and nitrogen species, effectively destroys microbial biofilms without leaving chemical residues [37,38]. Given the motility and cooperative biofilm behavior of spirochetes, these technologies merit priority evaluation in poultry-processing settings.
Equally critical is judicious antibiotic use in flocks. Broad-spectrum antimicrobial prophylaxis can disrupt normal microbiota, select for resistant strains, and favor biofilm-forming species able to survive sub-lethal exposure. Israel’s long-standing emphasis on feed hygiene and sanitation—rather than routine antibiotic administration—offers a useful real-world contrast [3,4,5,26]. Shifting global poultry practice toward prevention and microbial-ecology management, rather than end-stage treatment, may simultaneously reduce antimicrobial resistance and the persistence of pathogenic spirochetes within the food chain.
Epidemiologic and Laboratory Priorities
Although upgrading disinfection infrastructure entails cost, the long-term public-health and economic benefits are substantial. A cooperative public–private approach offers a win–win opportunity: stronger food safety, reduced disease burden, and increased industry resilience. Federal agencies such as HHS and USDA could support modernization through grants or pilot bioecology audits for food-processing facilities, positioning microbial-control reform as both a health safeguard and an innovation pathway rather than a regulatory burden.
Immediate priorities include targeted environmental sampling within processing plants to test for spirochetes and other resilient taxa that may persist despite current disinfection practices [16,17]. Culture-independent sequencing, combined with electron or confocal microscopy, could determine whether these organisms form mixed-species biofilms capable of resisting chlorine-based sanitation. Given industrial ecology, the most plausible candidates are poultry-associated spirochetes rather than tick-borne Borrelia, which provides a focused and testable starting point. In parallel, clinical investigations should screen neurodegenerative cohorts for latent spirochetal infection using polymerase chain reaction (PCR) and metagenomic assays on cerebrospinal fluid, intestinal biopsies, or olfactory mucosa [9,10,11].
Because Parkinson’s disease, dementia, and historical prion disorders share overlapping neuropathologic features, archived variant Creutzfeldt–Jakob disease (vCJD) and bovine spongiform encephalopathy (BSE) tissues provide a logical reference point for retrospective comparison. Re-examining these materials with contemporary molecular and histopathologic methods could clarify whether microbial cofactors were historically present but unrecognized. Such context strengthens the rationale for exploring whether persistent food-chain exposures may contribute to long-term neurodegenerative patterns.
Broader Significance
The convergence of industrial hygiene, chronic infection, and neurodegeneration reflects the One Health principle linking animal and human well-being [16,17,35].
If spirochetes such as B. pilosicoli constitute a persistent reservoir within the food chain, interventions aimed at reducing their adaptation and prevalence would potentially simultaneously support antimicrobial stewardship, food-safety sustainability, and contribute to strategies for neurological-disease prevention.
This framework underscores the need for coordinated action among government, industry, and research sectors to foster a cleaner microbial ecology and promote long-term neurological health.
Conclusions and Future Directions
Summary of Evidence
Archived variant Creutzfeldt–Jakob disease (vCJD) and bovine spongiform encephalopathy (BSE) tissues offer a rare opportunity to revisit historical neurodegenerative pathology with contemporary tools. Prior analyses have reported tubulofilamentous structures in vCJD brains resembling spiroplasma-like forms observed in Alzheimer’s disease and experimental infection models [42], raising the possibility that microbial cofactors may have been present but overlooked. Applying molecular and immunohistochemical assays to these archived specimens could determine whether Borrelia, Brachyspira, or morphologically similar organisms co-existed in those cases [11,42]. Even limited detection of such agents would justify targeted surveillance in modern neurodegenerative disorders.
Research Priorities
- Environmental Verification. Comprehensive sampling of poultry-processing environments should determine whether spirochetes or other biofilm-forming taxa persist despite current disinfection protocols [16,17]. Metagenomic and microscopic approaches can clarify survival niches and guide improved hygiene design [11,12].
- Clinical Correlation. Prospective studies comparing neurodegenerative patients with matched controls for serologic or PCR evidence of spirochetes in intestinal or cerebrospinal samples are needed to test the causal link.
Limitations and Future Work
Ecological inference—The observed associations between poultry-system variables and neurodegenerative-disease trends are population-level and cannot establish causation. Individual-level exposure data are currently lacking, and the correlations should therefore be interpreted as hypothesis-generating rather than confirmatory. Evident parallel trends may reflect shared time-dependent factors such as aging demographics, diagnostic expansion, changes in health-system reporting, or unrelated environmental exposures. Without individual-level data, these ecological patterns cannot determine causality. Direct molecular evidence is required to establish whether persistent spirochetes constitute an upstream driver rather than a correlated marker.
Sampling biases—Existing datasets rely on national reporting that varies in diagnostic sensitivity, age distribution, and dietary record accuracy, which may introduce systematic bias.
Uncontrolled confounders—Other microbial or environmental factors—such as viral, fungal, metal, or pollutant exposures—may influence the same neurodegenerative trajectories and have not been excluded.
Molecular validation gap—Direct molecular confirmation of spirochetes in post-sanitation poultry carcasses, processing environments, or patient gut and cerebrospinal fluid samples is currently absent. Although ecological correlations and historical neuropathology provide circumstantial support, empirical detection using 16S rRNA PCR or shotgun metagenomics in contemporary samples is essential to verify microbial persistence as a contributing factor [11,14]. This evidence gap must be addressed before causal inferences can be made.
Future work—Next-generation studies should integrate environmental metagenomics, case–control microbiome profiling, and controlled exposure models to determine whether biofilm-forming spirochetes act as initiators or amplifiers of neurodegenerative pathology [11,12,19].
Recommendations for Confirmation and Intervention
Confirmatory testing methods—Prioritize high-reliability tools to establish the spirochetal link: shotgun metagenomic sequencing of stool from PD and dementia patients versus controls to detect Brachyspira and dysbiosis patterns; fluorescence in situ hybridization (FISH) to visualize spirochetes in tissues; targeted PCR amplification of spirochetal 16S rRNA in clinical and environmental samples; and environmental PCR or metagenomics in processing facilities [11,12,14].
For scalability, targeted PCR of spirochetal 16S rRNA in stool or serum—combined with serologic ELISA for anti-Brachyspira IgG—could enable broad population-level screening and efficient case–control comparison. Environmental PCR or metagenomic testing in poultry-processing plants can trace biofilm sources to human exposure [11,14].
If fresh post-sanitation poultry samples and neurodegenerative-patient tissues show no detectable spirochetal DNA using modern metagenomics, the microbial-persistence hypothesis would be weakened or potentially falsified.
Applied sanitation lessons—Lessons from Germany and Israel demonstrate distinct but convergent paths to microbial control. Germany’s oxidative and thermal systems—dominated by peracetic-acid and hot-water processing, slower line speeds, low antibiotic reliance, and mandatory microbiologic monitoring—have coincided with stable or declining PD and dementia trends [28,29].
Israel achieves similar outcomes through kosher salting and rinsing, which mechanically reduce microbial loads without heavy chemical use [4]. Together, these approaches illustrate that preventive sanitation design—not consumption volume alone—can interrupt microbial adaptation and reduce long-term exposure risk.
Antibiotic stewardship—Flock-level overuse disrupts microbial ecology, selects for resistant taxa, and perpetuates biofilm persistence. Shifting focus toward flock health and feed hygiene—rather than routine antimicrobial prophylaxis—would strengthen resilience at the source [4,26].
Feed reform and monitoring—Permanent feed reform eliminating rendered animal proteins, as demonstrated in Israel’s 1996 post-BSE policy, remains vital for breaking cross-species microbial cycles [4,5,26]. Mandatory biofilm and antimicrobial-resistance surveillance—guided by international food-safety frameworks and justified by the known persistence of biofilm-forming taxa—would enable ongoing monitoring and early detection of microbial adaptation [17].
Implementation strategy—Phased implementation—pilot testing (6 months), validation (12 months), and regulatory reform—offers a high return on modest investment against the > US $420 billion annual neurodegenerative burden [1,2]. The BSE disaster demonstrated how microscopic feedback loops within the food chain—driven by the recycling of animal proteins—can silently amplify pathogens before detection [4,5]. Prevention at the microbial-ecological level is far less costly than crisis response after systemic contamination. Disregarding these lessons risks repeating them in new and more complex forms.
The opportunity to close this microbial feedback loop—before another cross-species crisis emerges—remains within reach. Testing this hypothesis would cost far less than enduring the disease burden it aims to prevent—approximately $45,000–$65,000 for a comparative metagenomic study capable of confirming or refuting the model.
A single comparative test could resolve the question. Sample 100 post-sanitation broiler carcasses from U.S. high-speed processing lines and 100 from Israeli kosher facilities. Apply 16S rRNA PCR targeting Brachyspira and Treponema sequences [11,14]. If spirochetal DNA is detected more frequently on U.S. carcasses—despite chlorine disinfection—while Israeli samples remain near zero, the microbial-persistence hypothesis gains support; if not, it can be rejected. Estimated cost: < US $50,000. Estimated duration: six months.
The prion model has dominated for four decades; this empirical test could settle the question within half a year. Because verification requires only a modest comparative microbial assay, the public-health cost of delay now far exceeds the cost of testing.
Closing Perspective
The persistence-infection hypothesis calls for renewed vigilance and collaboration. With emerging evidence linking microbial adaptation to chronic disease, food-system, hospital, and community-hygiene practices should be re-examined through the lens of microbial ecology. As outlined earlier, preventing microbial persistence requires precision rather than potency: chemical intensification can provoke hormesis, strengthen biofilm defenses, and foster antimicrobial resistance. Reform will depend on coordinated efforts across government, industry, and science, guided by a shared commitment to sustainable disinfection and public health.
Miklossy identified spirochetes in multiple neural and ocular tissues—including brain, spinal cord, and retinal regions—in patients with Alzheimer’s disease, multiple sclerosis, and amyotrophic lateral sclerosis [11,12]. Riviere and colleagues detected oral Treponema DNA within Alzheimer’s brain tissue in 87.5% of examined cases, supporting a potential hematogenous or mucosal route of neuroinvasion [14]. The most coherent ecological signal, however, emerges for Parkinson’s disease and dementia, which show broadly parallel rising trajectories from the late 1990s onward that mirror the intensification of poultry-processing systems [2,22,23].
These convergent findings suggest that upstream food-system microbial exposures warrant serious consideration as modifiable contributors to long-latency neurodegenerative disease. Prioritizing sanitation strategies that limit microbial persistence—rather than relying on increasingly aggressive chemical disinfection—may offer a pragmatic avenue for disease prevention, public-health cost reduction, and future empirical testing of the proposed ecological model.
Conflicts
None.
Funding
None.
References
- Steinmetz, JD; et al. Global, regional, and national burden of disorders affecting the nervous system, 1990-2021: a systematic analysis for the Global Burden of Disease Study 2021. Lancet Neurol 2024, 23(4), 344–381. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT: Food Balance Sheets. 2024. Available online: https://www.fao.org/faostat/en/#data/FBS (accessed on 12 Nov 2025).
- World Organization for Animal Health (WOAH; formerly OIE). Recognition of Israel as having negligible BSE risk under the Terrestrial Animal Health Code.; OIE: Paris, 2011; Available online: https://www.woah.org (accessed on 26 January 2026).
- Israeli Ministry of Health; Ministry of Health. Food safety and kosher poultry processing standards. Jerusalem. Available online: https://www.gov.il/en/departments/ministry_of_health (accessed on 26 January 2026).
- Yakobson, BA; Perl, S; Edery, N; et al. Recognition of Israel’s BSE status: review of surveillance and feed-ban regulations. Israel Journal of Veterinary Medicine 2014, 69(4), 173–182. Available online: https://www.ijvm.org.il/sites/default/files/negligible_bse_status.pdf (accessed on 12 Nov 2025).
- Alzheimer’s Association. Alzheimer’s disease facts and figures. Alzheimers Dement. 2024, 20(3), 1011–1080. [Google Scholar] [CrossRef]
- Parkinson’s Foundation. Parkinson’s disease: statistics and facts. 2024. Available online: https://www.parkinson.org/understanding-parkinsons/statistics (accessed on 12 Nov 2025).
- Van Maele-Fabry, G; Hoet, P; Lison, D. Occupational exposure to pesticides and Parkinson’s disease: a systematic review and meta-analysis of cohort studies. Environ Res. 2012, 117, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Prusiner, SB. Prions. Proc Natl Acad Sci U S A 1998, 95(23), 13363–13383. [Google Scholar] [CrossRef] [PubMed]
- Allen, HB; Morales, D; Jones, K; Joshi, S. Alzheimer’s disease: a spirochetosis. J Alzheimers Dis. 2016, 54(3), 859–872. [Google Scholar] [CrossRef]
- Miklossy, J. Alzheimer’s disease—a neurospirochetosis. J Neuroinflammation 2011, 8, 90. [Google Scholar] [CrossRef]
- Miklossy, J. Historic evidence to support a causal relationship between spirochetal infections and Alzheimer’s disease. Front Aging Neurosci. 2015, 7, 46. [Google Scholar] [CrossRef]
- Brundin, P; Melki, R; Kopito, R. Prion-like transmission of protein aggregates in neurodegenerative diseases. Nat Rev Mol Cell Biol. 2010, 11(4), 301–307. [Google Scholar] [CrossRef]
- Riviere, GR; Riviere, KH; Smith, KS. Molecular and immunological evidence of oral Treponema in the human brain. J Alzheimers Dis. 2002, 4(5), 337–346. [Google Scholar] [CrossRef]
- Burch, DGS; Harding, C; Newell, DG. A survey of 222 commercial broiler flocks for Brachyspira infection in Great Britain. Avian Pathol. 2006, 35(3), 213–217. [Google Scholar] [CrossRef]
- Bridier, A; Briandet, R; Thomas, V; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: a review. Biofouling 2011, 27(9), 1017–1032. [Google Scholar] [CrossRef]
- Giaouris, E; Heir, E; Hébraud, M; et al. Attachment and biofilm formation by foodborne bacteria in meat processing environments: a review. Meat Sci. 2014, 97(3), 298–309. [Google Scholar] [CrossRef]
- Calabrese, EJ; Mattson, MP. How does hormesis impact biology, toxicology, and medicine? NPJ Aging Mech Dis 2017, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Sampson, TR; Debelius, JW; Thron, T; et al. Gut microbiota regulate motor deficits and neuroinflammation in a model of Parkinson’s disease. Cell. 2016, 167(6), 1469–1480.e12. [Google Scholar] [CrossRef]
- Foster, JA; McVey Neufeld, KA. Gut-brain axis: how the microbiome influences anxiety and depression. Trends Neurosci. 2013, 36(5), 305–312. [Google Scholar] [CrossRef]
- Cryan, JF; Dinan, TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci. 2012, 13(10), 701–712. [Google Scholar] [CrossRef] [PubMed]
- National Chicken Council. Per capita consumption of poultry and livestock, 1960–forecast 2026 (lb/person/year). Available online: https://www.nationalchickencouncil.org/industry/statistics (accessed on 26 January 2026).
- USDA Food Safety and Inspection Service. Pathogen Reduction; Hazard Analysis and Critical Control Point (HACCP) Systems; Final Rule. Federal Register 1996, 61(144), 38806–38889. [Google Scholar]
- USDA Food Safety and Inspection Service. New Poultry Inspection System (NPIS). 2023. Available online: https://www.fsis.usda.gov/policy/federal-register-rulemaking/federal-register-rules/2014-18526 (accessed on 12 November 2025).
- GBD 2016 Parkinson’s Disease Collaborators. Global, regional, and national burden of Parkinson’s disease, 1990–2016: a systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2018, 17(11), 939–953. [Google Scholar] [CrossRef] [PubMed]
- Giladi, N; Gurevich, T; Shabtai, H; et al. Declining incidence of Parkinson’s disease in Israel. J Neural Transm (Vienna) 2025, 132(7), 987–995. [Google Scholar] [CrossRef]
- Fink, A; Pavlou, MAS; Roomp, K; Schneider, JG. Declining trends in the incidence of Parkinson’s disease. J Parkinsons Dis. 2025, 15(1), 182–188. [Google Scholar] [CrossRef]
- Rommel, A; Deuschl, G; Dodel, R; Yessimova, D; Neuhauser, H; Brückner, G; Schröder, H; Schüssel, K; Porst, M. Parkinson disease – Prevalence, trends and regional patterns in Germany: an analysis based on routine data from the statutory health insurance. J Health Monit. Accessed. 2025, 10(1), e13070. (accessed on 26 January 2026). [Google Scholar] [CrossRef] [PubMed]
- Rommel, A; Gaertner, B; Neuhauser, H; Yessimova, D; Schröder, H; Brückner, G; Schüssel, K; Porst, M. Dementia – prevalence, trends and regional patterns in Germany: an analysis based on routine data from the statutory health insurance. J Health Monit. Accessed. 2025, 10(1), e13079. (accessed on 26 January 2026). [Google Scholar] [CrossRef] [PubMed]
- Lutski, M; Rasooli, I; Sternberg, S; et al. Prevalence and incidence rates of dementia in Israel: a national study using electronic health records. J Alzheimers Dis. 2022, 85(3), 1153–1161. [Google Scholar] [CrossRef]
- Hou, M; Yang, L. Trends and forecasts of the prevalence and mortality of Alzheimer’s disease in China from 1990 to 2050: a systematic analysis and comparison. Front Public Health 2025, 13, 1616232. [Google Scholar] [CrossRef]
- Bourassa, DV. Antimicrobial use in poultry processing. Food Safety Magazine. Jan 2017. Available online: https://www.food-safety.com/articles/5581-antimicrobial-use-in-poultry-processing (accessed on 26 January 2026).
- USDA National Agricultural Statistics Service. Poultry – Production and Value 2024 Summary. United States Department of Agriculture: Washington, DC, April 2025; Available online: https://usda.library.cornell.edu/usda-esmis/ (accessed on 12 November 2025).
- FDA Center for Veterinary Medicine. Animal Food: Ingredients Permitted in Feed. 2024. Available online: https://www.fda.gov/animal-veterinary/animal-food-feeds/animal-food-ingredients-permitted-feed (accessed on 12 November 2025).
- U.S. Food and Drug Administration. 21 CFR 589.2000—Animal proteins prohibited in ruminant feed. 2008. Available online: https://www.ecfr.gov/current/title-21/chapter-I/subchapter-E/part-589/section-589.2000 (accessed on 12 November 2025).
- USDA Economic Research Service. Livestock and Meat Domestic Data. 2024. Available online: https://www.ers.usda.gov/data-products/livestock-meat-domestic-data/ (accessed on 12 November 2025).
- Tong, TYN; Imamura, F; Brauer, P; Griffin, SJ; Wareham, NJ; Finer, N; et al. Meat consumption, cognitive function and disorders: a systematic review with narrative synthesis and meta-analysis. Nutrients 2020, 12(6), 1650. [Google Scholar] [CrossRef]
- Zhang, H; Greenwood, DC; Risch, HA; Bunce, D; Hardie, LJ; Cade, JE. Meat consumption and risk of incident dementia: cohort study of 493,888 UK Biobank participants. Am J Clin Nutr. 2021, 114(1), 175–184. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture Animal and Plant Health Inspection Service. Export to China: Non-Ruminant Feeds, Feed Ingredients and Fats. April 2024. Available online: https://www.aphis.usda.gov/regulations/vs/iregs/animals/downloads/ch_feeds.pdf (accessed on 12 November 2025).
- Fraser, GE; Cosgrove, CM; Mashchak, AD; et al. Lower rates of Parkinson’s disease in the Adventist Health Study-2 cohort. Mov Disord. 2020, 35(12), 2269–2277. [Google Scholar] [CrossRef]
- Chen, SG; Stribinskis, V; Rane, MJ; et al. Exposure to bacterial amyloids enhances α-synuclein aggregation via cross-seeding. eLife 2020, 9, e53111. [Google Scholar] [CrossRef]
- Liberski, PP; Sikorska, B; Hauw, JJ; et al. Tubulofilamentous structures in variant Creutzfeldt–Jakob disease resemble spiroplasma-like forms described in Alzheimer disease and experimental spiroplasma infection. Prion 2010, 4(3), 175–188. [Google Scholar] [CrossRef]
- Ozone treatment of meat and meat products: a review. Front Food Sci Technol. 2024, 4, 1351801. [CrossRef]
- Application of cold plasma and ozone technology for decontamination of Escherichia coli in foods—A review. Food Control 2021, 127, 108106. [CrossRef]
- Verlinden, M; Pasmans, F; Garmyn, A; De Zutter, L; Haesebrouck, F; Martel, A. Occurrence of viable Brachyspira spp. on carcasses of spent laying hens from supermarkets. Food Microbiol. 2012, 32(2), 321–324. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
United States trends in Parkinson’s-disease age-standardized incidence rate (ASIR), dementia age-standardized death rate (ASDR), and per-capita chicken, beef, and pork consumption, 1990–2020 (5-year intervals). Disease rates are drawn from GBD 2021 [1], and meat-supply data (kg/person/year) from FAOSTAT 2024². PD ASIR rose by roughly 60% over this interval, closely paralleling long-term increases in chicken consumption, while beef and pork intake remained comparatively stable or declined. Lines for PD and dementia are plotted on the left axis (scaled for visibility), and meat consumption on the right axis.
Figure 1.
United States trends in Parkinson’s-disease age-standardized incidence rate (ASIR), dementia age-standardized death rate (ASDR), and per-capita chicken, beef, and pork consumption, 1990–2020 (5-year intervals). Disease rates are drawn from GBD 2021 [1], and meat-supply data (kg/person/year) from FAOSTAT 2024². PD ASIR rose by roughly 60% over this interval, closely paralleling long-term increases in chicken consumption, while beef and pork intake remained comparatively stable or declined. Lines for PD and dementia are plotted on the left axis (scaled for visibility), and meat consumption on the right axis.
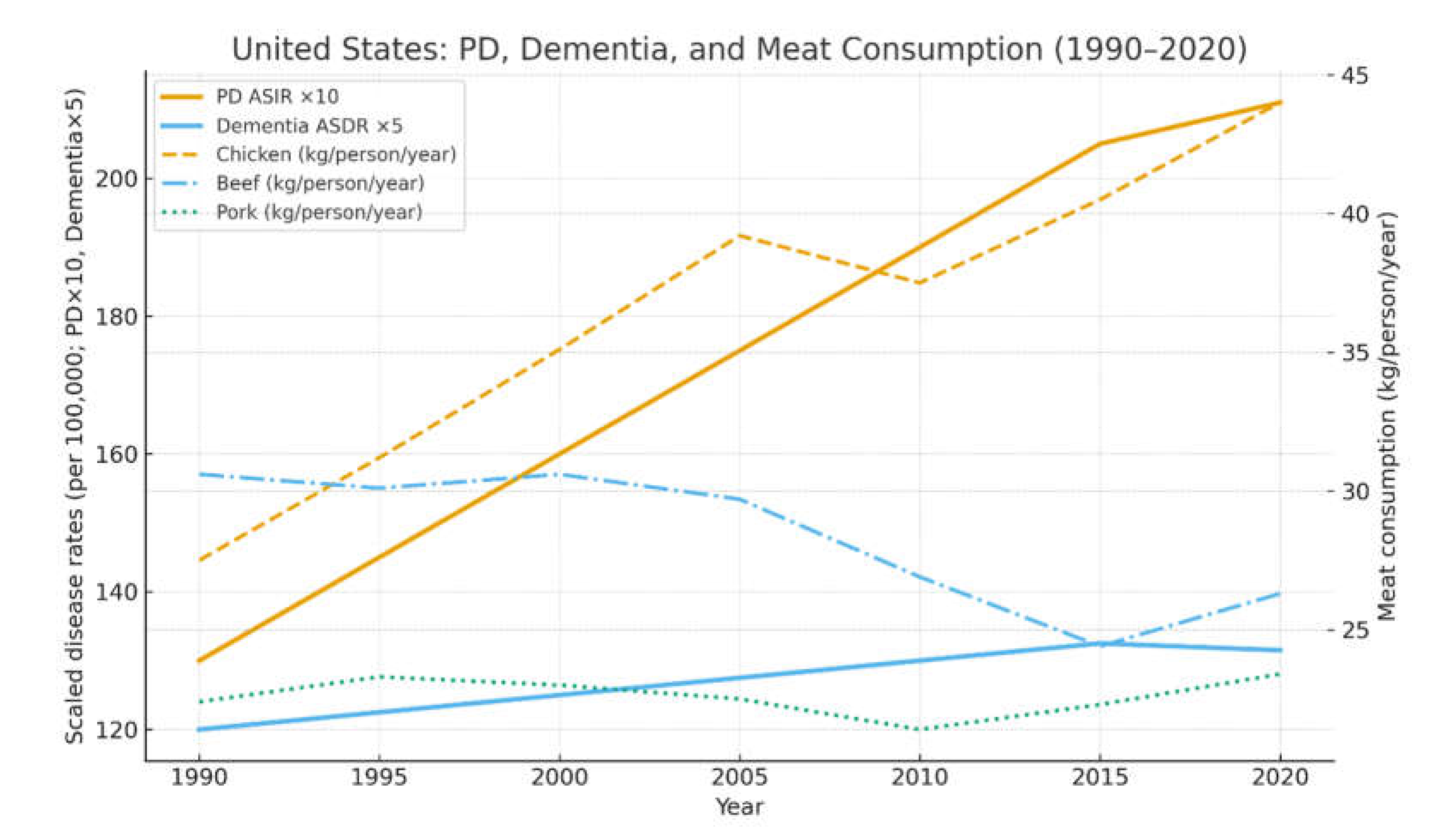
Figure 2.
compares age-standardized Parkinson’s-disease incidence rates (ASIR) for the United States, China, Israel, and global estimates using GBD 2021 data [1]. Since the late 1990s, PD incidence has risen much more sharply in the United States and China, whereas Israel and the global series show comparatively stable or slightly declining long-term trajectories [29,30,31,32].
Figure 2.
compares age-standardized Parkinson’s-disease incidence rates (ASIR) for the United States, China, Israel, and global estimates using GBD 2021 data [1]. Since the late 1990s, PD incidence has risen much more sharply in the United States and China, whereas Israel and the global series show comparatively stable or slightly declining long-term trajectories [29,30,31,32].
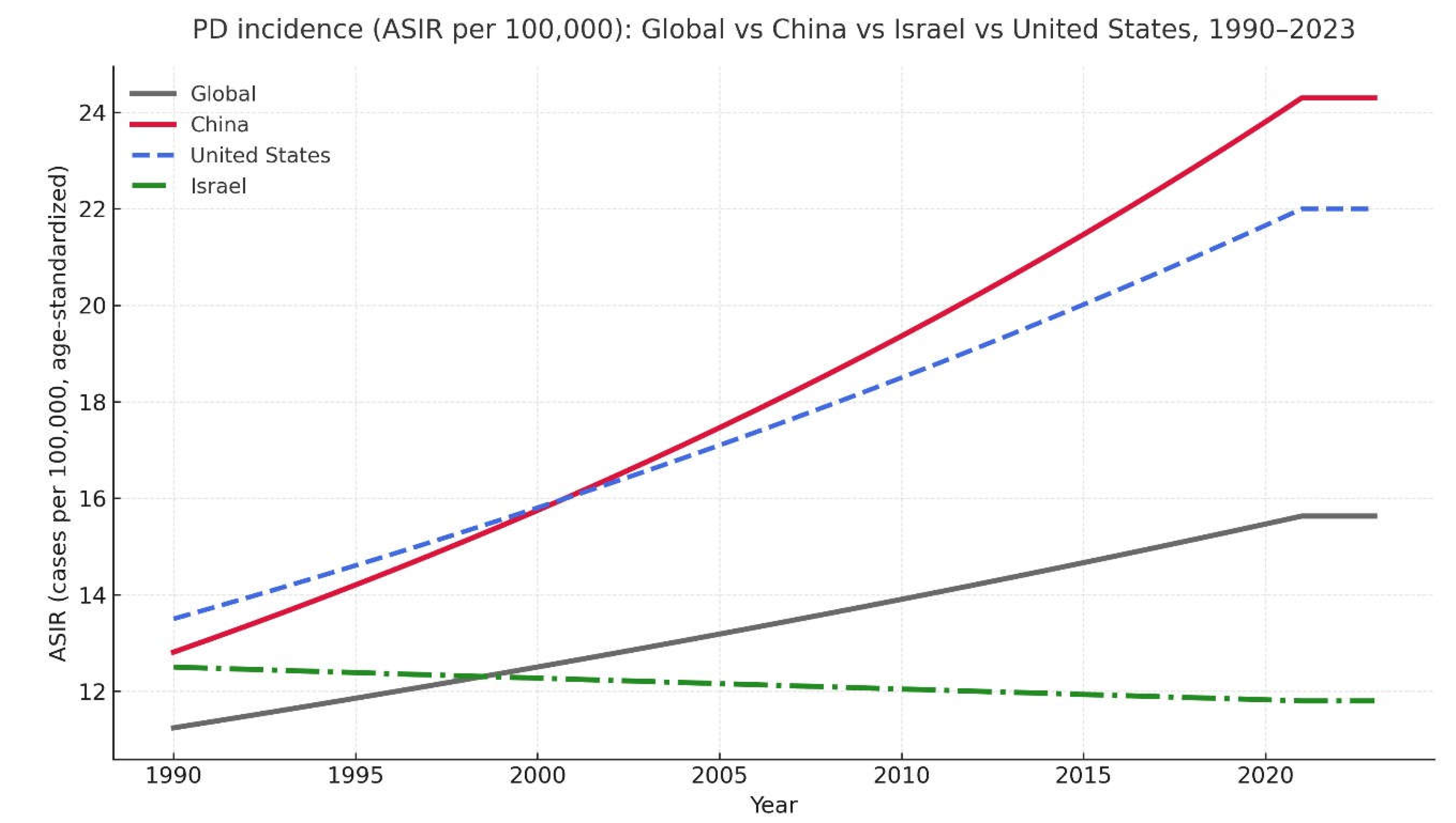
Figure 3.
U.S. trends in dementia incidence, Parkinson’s disease prevalence (scaled ×10), and per-capita chicken consumption, 1990–2024.
Figure 3.
U.S. trends in dementia incidence, Parkinson’s disease prevalence (scaled ×10), and per-capita chicken consumption, 1990–2024.
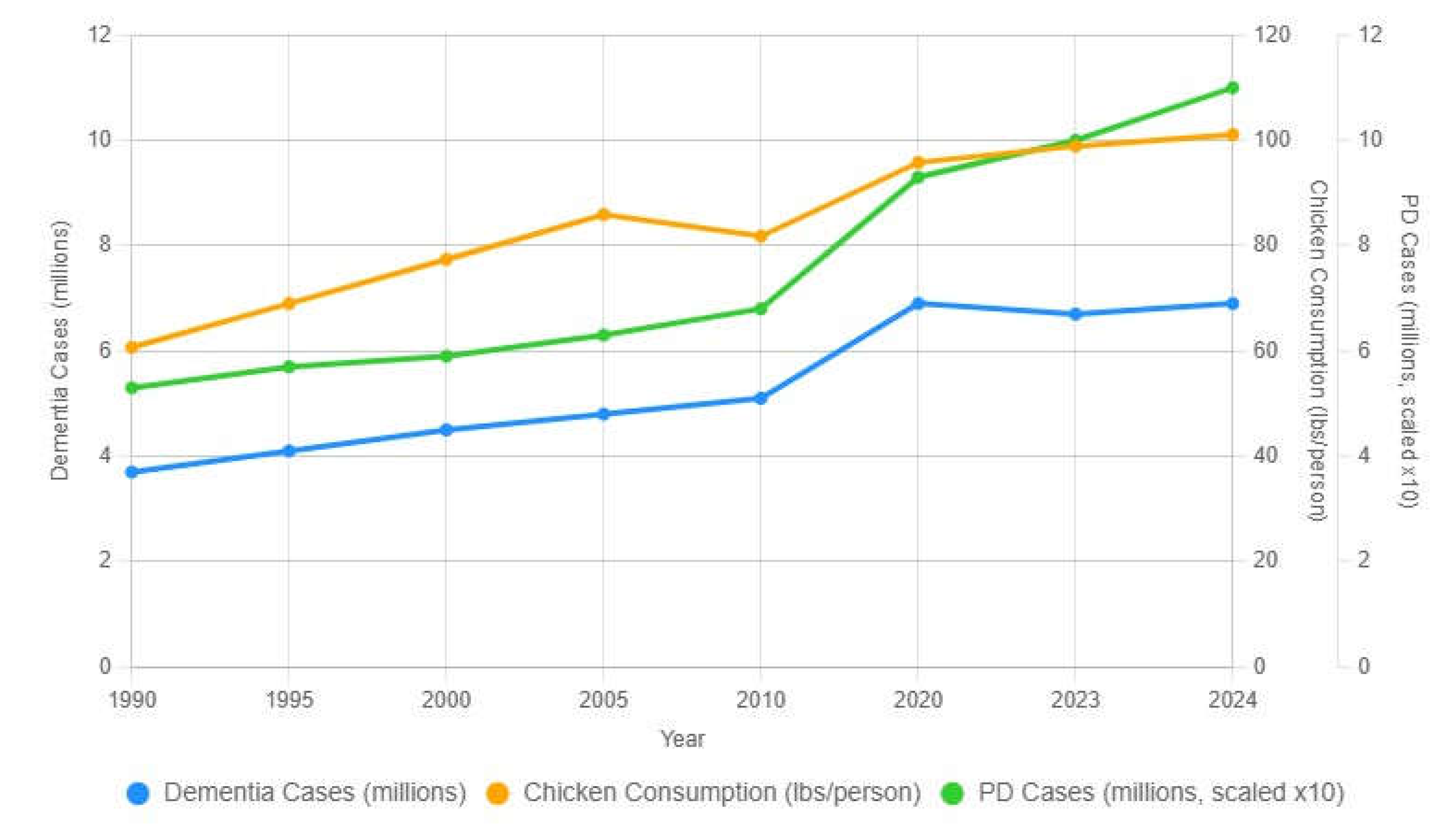
Figure 4.
Spirochete feedback cycle illustrating how hormetic pressure can sustain microbial persistence across poultry-production and human-exposure pathways.
Figure 4.
Spirochete feedback cycle illustrating how hormetic pressure can sustain microbial persistence across poultry-production and human-exposure pathways.
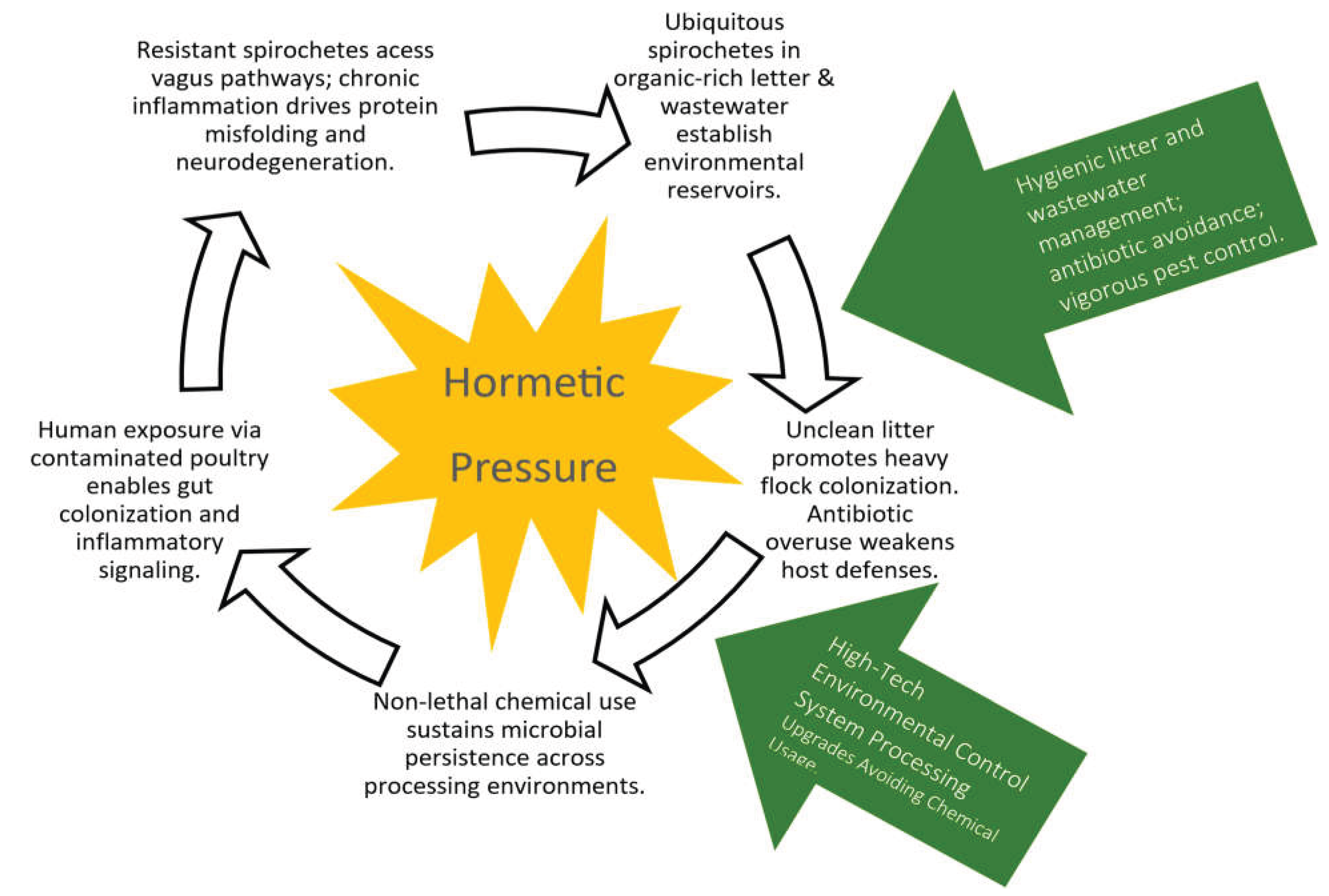
Table 2.
Correlation coefficients (Pearson r and Spearman ρ) for U.S. poultry-related meat supply and modeled Parkinson’s-disease and dementia prevalence, 1990–2024.
Table 2.
Correlation coefficients (Pearson r and Spearman ρ) for U.S. poultry-related meat supply and modeled Parkinson’s-disease and dementia prevalence, 1990–2024.
| COMPARISON | PEARSON R | SPEARMAN ρ |
| PD VS MEAT (CHICKEN + PROCESSED) | 0.994 | 1.000 |
| DEMENTIA VS MEAT | 0.992 | 1.000 |
| PD VS DEMENTIA | 0.979 | 1.000 |
Note. Spearman ρ = 1.000 indicates a perfect monotonic rank-order alignment across all seven multi-year intervals (p < 0.001). Values for PD, dementia, and poultry-related meat supply were indexed to 1990 = 100 for comparability. Data sources: GBD 2021 [1]; FAOSTAT [2]; USDA Economic Research Service (ERS) [36].
Table 3.
Principal Theories of Neurodegeneration and Their Relationship to the Microbial Persistence Model.
Table 3.
Principal Theories of Neurodegeneration and Their Relationship to the Microbial Persistence Model.
| Major Theory | Core Premise | Relationship to Microbial-Persistence Model |
|---|---|---|
| Infection Hypothesis | Chronic pathogens (including spirochetes) initiate sustained inflammation | Serves as the initiating driver of this framework |
| Gut–Brain Axis | Enteric microbiota and immune signaling influence brain pathology | Provides the physiologic conduit linking infection to neurodegeneration |
| Prion-Like Propagation | Misfolded proteins spread pathology cell-to-cell | Represents downstream amplification of pathology that may be initiated by inflammatory or microbial triggers |
| Toxin / Oxidative Stress | Chemical or metabolic stress causes neuronal injury | Considered a secondary effect of microbial metabolism or chemical hormesis |
Table 4.
Comparative Strength of Evidence for Major Neurodegenerative Triggers.
| Proposed Primary Trigger | Mechanistic Focus | Representative Association (Approx.) | Key Supporting Sources | Relationship to Spirochetal-Persistence Model |
|---|---|---|---|---|
| Aging | Cellular senescence, mitochondrial decline, oxidative stress | OR increases steadily with age (≈1.07–1.09 per year after 60) | Postuma 2015 [39]; Dorsey 2007 [40] | Age increases vulnerability to persistent microbial infection and reduces clearance capacity |
| Genetic Susceptibility | Mutations affecting protein handling or inflammation (LRRK2, SNCA, APOE-ε4) | OR ≈ 2–5 | Brundin 2010 [13] | Genetic variation modulates inflammatory response to chronic infection |
| Environmental Toxins | Pesticides, solvents, metals | OR ≈ 1.5–2.0 | Van Maele-Fabry 2012 [8] | Toxins amplify inflammation triggered by microbes; hormesis selects for resistant organisms |
| Gut–Brain Axis | Microbiota–brain signaling, vagal pathways | Supported by experimental and human observational evidence | Sampson 2016 [19]; Foster 2013 [20] | Spirochetes fit within this pathway as chronic gut colonizers initiating inflammation |
| Prion-Like Propagation | Cell-to-cell spread of misfolded α-synuclein or β-amyloid | Supported by mechanistic studies and pathology | Brundin 2010 [13] | Chronic microbial inflammation may initiate misfolding events that then propagate |
| Viral or Other Infections | HSV-1, H. pylori, gut pathogens | OR ≈ 1.3–1.7 (meta-analytic ranges) | Allen 2016 [10]; Miklossy 2011 [11] | Spirochetes meet infection criteria including latency, neurotropism, and persistence |
| Spirochetal Persistence (Proposed) | Chronic biofilm-forming infection (Brachyspira, Borrelia, Treponema) | Temporal and geographic alignment; ecological correlations in multiple datasets (r ≈ 0.6–0.8) | Miklossy 2011 [11]; Riviere 2002 [14]; Tong 2020 [37] | Integrates microbial, inflammatory, gut–brain, toxin, and prion-like pathways into a unified ecological model |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



