
Submitted:
21 January 2026
Posted:
22 January 2026
You are already at the latest version
Abstract
The oral-gut microbiome axis represents a critical, interdependent ecosystem whose stability is essential for systemic immune homeostasis and barrier defense. In the intensive care unit (ICU), this axis undergoes a profound and targeted assault from life-sustaining interventions. Mechanical ventilation, broad-spectrum antibiotics, and proton pump inhibitors (PPIs) act synergistically to dismantle the physiological barriers separating the oral and gut microbiomes, driving dysbiosis at both sites and facilitating the ectopic translocation and colonization of oral pathobionts in the gastrointestinal tract. This process transcends passive microbial spillover, actively fueling intestinal inflammation, compromising epithelial integrity, and exacerbating systemic immune dysregulation—key pathways in the pathogenesis of ventilator-associated pneumonia (VAP), enterogenic sepsis, and multiple organ dysfunction. This review synthesizes contemporary evidence to posit that the integrity of the oral-gut axis is a modifiable determinant of outcome in critical illness. Consequently, we advocate for a paradigm shift in infection prevention from external disinfection towards the active stewardship of endogenous microbial ecology. We critically evaluate evolving oral care strategies, deconstruct the limitations of non-selective antiseptics like chlorhexidine, and highlight the evidence supporting ecological approaches such as mechanical hygiene, physiological irrigation, and selective oropharyngeal decontamination. Future research must prioritize the standardization of axis-preserving protocols and rigorously evaluate microbiome-targeted interventions, including the refinement of selective decontamination strategies for targeted populations, to restore ecological resilience and improve patient survival.
Keywords:
oral-gut microbiome axis
; mechanical ventilation
; proton pump inhibitors
; oral care
; ICU
1. Introduction
The human body is an integrated ecosystem where microbial communities maintain health and prevent disease. The oral cavity and gastrointestinal tract host the body’s most complex microbial reservoirs, in continuous dialogue with host physiology. The digestive system mediates the interface between internal and external environments, performing nutrient assimilation and barrier defense [1].
Anatomically contiguous, the mouth and gut form the oral-gut microbiome axis—a sophisticated network involving microbial translocation, components, metabolites, and immune-metabolic signaling [2,3]. The oral cavity, as entry point to respiratory and digestive tracts with rich vascular drainage, influences distal colonization and systemic dissemination [2,4,5,6,7,8]. Growing evidence links oral dysbiosis to systemic diseases including atherosclerosis, rheumatoid arthritis, adverse pregnancy outcomes, liver diseases, stroke, and cardiometabolic and neurodegenerative disorders [2,3,4,5,6,7,8]. Periodontal pathogens like Porphyromonas gingivalis, Fusobacterium nucleatum, and Tannerella forsythia are strongly associated with extra-oral conditions, often via the oral-gut axis [3,9,10].
In the intensive care unit (ICU), critically ill patients experience a synergistic assault on microbial homeostasis from endotracheal intubation, broad-spectrum antibiotics, and proton pump inhibitors (PPIs). These interventions dismantle barriers, induce dysbiosis, and facilitate ectopic colonization of oral bacteria in the gut [11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34]. This active process drives intestinal inflammation, barrier dysfunction, and systemic immune dysregulation, central to ventilator-associated pneumonia (VAP), enterogenic sepsis, and multiple organ dysfunction [35,36,37,38,39].
This review synthesizes evidence that oral-gut axis integrity is a modifiable determinant of critical illness outcomes. We deconstruct ICU-induced disruption, detail consequences, and trace oral care evolution from non-selective disinfection to ecological stewardship, evaluating chlorhexidine limitations and highlighting mechanical hygiene, physiological irrigation, and selective decontamination strategies.
2. The Oral-Gut Microbiome Axis: Fundamentals of a Critical Connection
2.1. The Distinct Yet Interdependent Ecosystems
A nuanced understanding of the oral-gut axis begins with appreciating the unique environments it connects. The intestinal microbiota is arguably the body’s most complex microbial organ, comprising trillions of bacteria, archaea, fungi, and viruses. Its assembly and function are shaped by a lifelong interplay of host genetics and modifiable environmental factors. These include diet—particularly the intake of fiber, fats, and refined sugars—antibiotic exposure, lifestyle elements like sleep and stress, and broader influences such as urbanization and hygiene practices [35,36,37,38,39]. This community is metabolically and immunologically indispensable. It is pivotal for the education and regulation of the host immune system, the maintenance of intestinal epithelial barrier integrity, the synthesis of essential vitamins (e.g., vitamin K, B vitamins), and the provision of colonization resistance. This latter function, a cornerstone of host defense, is achieved through mechanisms like nutrient competition, spatial occupation, and the production of antimicrobial substances such as short-chain fatty acids (SCFAs) [42]. Disruption of this delicate equilibrium, termed gut dysbiosis, is not merely associative but is increasingly understood to be causally implicated in a vast spectrum of chronic conditions. These range from gastrointestinal disorders like inflammatory bowel disease (IBD) and irritable bowel syndrome (IBS) to colorectal cancer, obesity, type 2 diabetes, cardiovascular and neurodegenerative diseases (e.g., Alzheimer’s and Parkinson’s), and even respiratory ailments including asthma and chronic obstructive pulmonary disease (COPD) [43,44,45].
In parallel, the oral cavity harbors an exceptionally diverse and dense microbiome, second in scale only to the gut. It is home to over 770 identified bacterial species, which are not randomly distributed but are organized into distinct biogeographical niches shaped by local variations in oxygen tension, pH, salivary flow, and surface topography [46,47,48]. These niches include mucosal surfaces (e.g., buccal mucosa, keratinized gingiva), shedding surfaces (saliva and tongue dorsum), and the highly structured biofilms of dental plaque—both supragingival (above the gumline) and subgingival (below the gumline), the latter being a notorious reservoir for periodontal pathogens [47,49]. The oral microbiome is inherently dynamic, constantly reshaped by substrate availability from diet, oral hygiene behaviors, habits like smoking, medication use (notably antibiotics and antiseptics), and the host’s local and systemic immune responses. Its composition is dominated by members of the phyla Firmicutes, Proteobacteria, and Bacteroidetes, with key genera including Streptococcus, Neisseria, Porphyromonas, and Fusobacterium [50].
2.2. The Physiological Barrier and Basal Microbial Exchange
Despite their direct anatomical connection via the digestive tract—a conduit for the daily swallowing of approximately 1.5 × 1012 oral bacteria [51]—the oral and gut microbiomes maintain distinct profiles in health. This separation is enforced by a defense is gastric acid, with a pH typically between 1.5 and 3.5, which acts as a sterilizing filter for the vast majority of ingested microbes [52]. Bile acids add another layer of chemical defense, with some possessing direct antibacterial properties through membrane disruption [52]. Within the intestinal lumen, a chemical and immunological shield comprised of antimicrobial peptides (e.g., α-defensins), secretory IgA, and the mucous gel layer prevents pathogen adhesion and proliferation [53]. Perhaps the most sophisticated component is biological: the resident gut microbiota itself provides colonization resistance. By outcompeting incoming microbes for nutrients and adhesion sites and by producing inhibitory metabolites like SCFAs, it effectively guards its territory [54,55]. Consequently, in healthy individuals, most swallowed oral bacteria are eliminated, and survivors typically fail to establish stable, expanding populations, rendering them nearly undetectable in standard fecal microbiome analyses [56].
However, this separation is not absolute, and a basal level of exchange occurs. Landmark research utilizing strain-level metagenomic analysis of paired saliva and fecal samples from hundreds of individuals demonstrated that oral bacteria do frequently translocate to the lower gastrointestinal tract and can successfully colonize under permissive conditions. Commonly transferred taxa include genera highly prevalent in the mouth, such as Streptococcus, Veillonella, Actinomyces, and Haemophilus [57]. This suggests the oral-gut axis facilitates a low-level, natural process of microbial exchange, as further evidenced by the regular detection of salivary Prevotella species in stool [58]. Under homeostasis, this exchange is likely inconsequential or even potentially functional. The critical shift occurs when barrier function is compromised, transforming this conduit into a pathway for disease.
2.3. Pathological Translocation: When Oral Commensals Become Gut Pathobionts
The collapse of the oral-intestinal barrier escalates microbial exchange from a background process to a central driver of pathology. Periodontitis, a chronic inflammatory oral disease, is a key instigator. Salivary microbiota associated with periodontitis, including members of the Porphyromonadaceae family, Tannerella, and Treponema, can be transferred to the gut, where they induce intestinal flora imbalance and inflammation [59]. This pathological translocation is markedly amplified in specific disease states. In patients with colorectal cancer, rheumatoid arthritis, and IBD, DNA and viable cells of specific oral commensals are found enriched in intestinal tissues and tumor microenvironments. Fusobacterium nucleatum, Campylobacter rectus, and Peptostreptococcus stomatis are prominent examples, and their presence frequently correlates with increased disease severity and poorer clinical outcomes [57,60].
The mechanisms by which these translocated oral bacteria exert harm are diverse and impactful (Figure 1). Intestinal inflammation, as seen in Crohn’s disease, alters the gut environment in ways that favor colonization by oral bacteria like Veillonella. Once established, it can suppress the expression of the apical sodium-dependent bile acid transporter, leading to abnormal luminal accumulation of bile acids that subsequently promote Clostridium difficile infection [61]. Hepatic dysfunction can dysregulate bile acid synthesis, depleting intestinal levels of antibacterial bile acids such as 12-KetoLCA. This weakening of the chemical barrier undermines colonization resistance and permits pro-inflammatory, oral-derived Streptococcus species to expand ectopically in the intestine [62]. Fusobacterium nucleatum, a classic oral periodontopathogen, can promote the progression of ulcerative colitis by activating the pro-inflammatory IL-17F/NF-κB signaling cascade, thereby enhancing the expression of key inflammatory cytokines including IL-1β, IL-6, IL-17F, and TNF-α [60,63].
Porphyromonas gingivalis serves as a paradigm for an oral pathobiont with systemic reach. Upon reaching the intestine, it directly assaults the epithelial barrier by downregulating the expression of critical tight junction proteins such as ZO-1, occludin, and Tjp1, thereby increasing gut permeability [64]. It also elevates systemic levels of lipopolysaccharide (LPS), which drives the upregulation of hepatic flavin-containing monooxygenase 3 (FMO3), increases plasma concentrations of the pro-atherogenic metabolite trimethylamine N-oxide (TMAO), and induces both gut dysbiosis and mucosal inflammation [65]. Notably, P. gingivalis has been detected in extra-intestinal pathological sites like atherosclerotic plaques and pancreatic tumors, powerfully illustrating the systemic dissemination potential of orally derived microbes [60,66,67,68,69,70]. Importantly, this relationship is bidirectional; inflammatory intestinal conditions such as IBD can themselves exacerbate oral microbial dysbiosis, creating a self-perpetuating vicious cycle of inflammation [71].
2.4. Bacterial Adaptations and the Immune-Mediated “Oral-Immune-Gut Axis”
Certain oral bacteria possess inherent traits that enhance their survival during gastrointestinal transit. Acid tolerance is paramount. Streptococcus mutans, a primary etiological agent of dental caries, is remarkably aciduric, maintaining viability at pH levels as low as 4.0, which aids its survival through the stomach [72]. Similarly, planktonic cells of Porphyromonas gingivalis can endure exposure to artificial gastric juice at pH 5.0, likely due to adaptive stress responses and protective capsule formation [73]. This trait becomes clinically significant when gastric acidity is pharmacologically reduced, such as during PPI therapy. Other bacteria exhibit metabolic flexibility. For instance, Veillonella parvula can shift its metabolic strategy from fermentation to anaerobic respiration during intestinal inflammation, utilizing amino acids and peptides as carbon sources to facilitate successful colonization [74].
Beyond the direct effects of translocated bacteria, periodontitis can exacerbate gut inflammation through sophisticated, immune-mediated mechanisms, defining a distinct “oral-immune-gut axis.” Systemic effects of oral infection include the reduction of small intestinal Th17 cell populations, a shift in macrophage polarization towards a pro-inflammatory M1 phenotype, a decrease in fecal IgA levels, and disturbances in microbial metabolic profiles (e.g., elevated lactate, reduced succinate and butyrate), all of which worsen gut dysbiosis [75]. Oral pathobionts like Klebsiella pneumoniae can, upon gut colonization, induce IL-1β, which promotes the accumulation of pro-inflammatory innate lymphoid cells and Th17 cells and acts as a factor driving the differentiation of pathogenic Th1/Th17 hybrid cells [76].
The migration and pathogenicity of immune cells are central to this axis. Research using advanced imaging has revealed unexpectedly broad leukocyte trafficking between the gut and other sites [77]. Within the gut, the interplay between periodontal inflammation and intestinal immunity is complex, with Th17 cells playing a crucial role [77]. These Th17 cells can give rise to Th1-like cells that are required for the pathogenesis of colitis [79], a process driven by cytokines like IL-23 [80]. A seminal study elegantly delineated a precise cellular mechanism linking oral and gut inflammation [80]. Periodontitis leads to an increased oral abundance of pathobionts like Klebsiella and Enterobacter. When swallowed, these bacteria can reach the gut and activate colonic mononuclear phagocytes via inflammasome pathways (e.g., NLRP3). Concurrently, the oral infection drives the differentiation of pathogen-specific Th17 effector memory (TEM) cells in the cervical lymph nodes. These cells enter the systemic circulation, home to the intestinal lamina propria, and upon re-encountering their cognate oral bacterial antigens, become reactivated. They then secrete potent pro-inflammatory cytokines such as IL-17A and IFN-γ, causing localized mucosal damage. Crucially, these T cells do not react to the resident commensal gut flora, establishing a direct, antigen-specific immunological link between a remote oral inflammatory site and intestinal pathology. Other oral bacteria like F. nucleatum can modulate intestinal Th17 responses in a metabolite receptor (FFAR2)-dependent manner to promote their own colonization [82], while orally administered P. gingivalis can be sampled by M cells in Peyer’s patches and presented by dendritic cells to drive local Th17 differentiation with potential for systemic migration [83]. Furthermore, specific oral bacteria like Prevotella intermedia can synergistically exacerbate inflammation induced by other oral streptococci, highlighting complex inter-bacterial interactions [84].
3. The ICU Crucible: Synergistic Disruption of the Axis by Standard Interventions
The critically ill patient represents the epitome of a host in which the homeostatic mechanisms guarding the oral-gut axis are overwhelmed. A confluence of severe illness and iatrogenic interventions creates a perfect storm for catastrophic dysbiosis and its sequelae (Figure 2).
3.1. Endotracheal Intubation: Engineering a Dysbiotic Oral Reservoir
Endotracheal intubation, while lifesaving for airway management and mechanical ventilation, catastrophically disrupts normal oropharyngeal physiology. It functionally abolishes the natural self-cleaning mechanisms of the mouth: speech, mastication, and swallowing are suppressed [11]. These actions are not trivial; they facilitate the mechanical clearance of food debris and microbes, while swallowing stimulates salivary flow. Saliva itself is a critical defense fluid, delivering a cocktail of antimicrobial peptides (e.g., lysozyme, lactoferrin, secretory IgA), buffers, and enzymes [12]. Intubated patients frequently experience significant reductions in salivary secretion (xerostomia) due to diminished pharyngeal stimulation, systemic dehydration, and the pharmacological effects of sedatives, opioids, and anticholinergic drugs. The resulting environment—stagnant, moist, and often nutrient-rich from enteral feeding residuals—is ideal for uncontrolled microbial overgrowth and the formation of tenacious biofilms on teeth, mucosal surfaces, and the endotracheal tube itself. This ecological shift favors the proliferation of classic ICU pathogens, including Staphylococcus aureus, Pseudomonas aeruginosa, and Candida albicans [13,85]. The process of bacterial adhesion to oral tissues is a critical first step in colonization and biofilm formation [86,87].
A 2025 longitudinal study provided direct, dynamic evidence of this shift, showing that high oral bacterial load at admission independently predicted the development of VAP and delirium [88]. In mechanically ventilated patients receiving standardized oral care, bacterial load on the tongue was high and the α-diversity of the tongue microbiome was significantly lower after extubation compared to the pre-intubation state, indicating a loss of microbial diversity and a shift towards a dysbiotic state enriched in opportunistic pathogens [14]. The observation of partial microbial recovery after extubation suggests the process is dynamic and at least partly driven by the presence of the tube and associated physiological impairments. A critical limitation of such studies, however, is their frequent reliance on sampling accessible sites like the tongue. This approach may grossly underestimate dysbiosis in high-risk, biofilm-dense niches like subgingival plaque pockets, which are major reservoirs for pathogens like Porphyromonas gingivalis and Fusobacterium nucleatum and are directly implicated in systemic dissemination [15,16]. Thus, endotracheal intubation fundamentally and rapidly engineers the oral cavity into a persistent, enriched reservoir of pathobionts, primed for both pulmonary aspiration and gastrointestinal translocation.
3.2. Broad-Spectrum Antibiotics: Depleting the Gut’s Indigenous Defense Army
Antibiotic therapy is a non-negotiable yet ecologically devastating mainstay of ICU practice. While targeting life-threatening infections, broad-spectrum agents cause collateral damage by indiscriminately decimating the commensal gut microbiota, the very foundation of colonization resistance. The perturbation of the gut microbiome by antibiotics is profound and multi-faceted [17,18]. A crucial refinement in understanding this process is the “Marker Hypothesis” [19,20]. Research demonstrates that the commonly observed increase in the relative abundance of oral bacterial taxa in the fecal microbiome of critically ill or antibiotic-treated patients often does not primarily reflect an active, numerical expansion of these oral bacteria within the gut. Instead, it is largely a mathematical artifact signaling a catastrophic collapse in the density and diversity of the indigenous gut microbiota. A pivotal mouse experiment made this clear: after one week of broad-spectrum antibiotic treatment, total fecal bacterial load plummeted by over 1000-fold, while the absolute abundance of orally derived bacteria remained unchanged. Their relative proportion, therefore, appeared dramatically inflated [19,20].
This conceptual distinction is vital for accurate interpretation but in no way diminishes the clinical danger. The antibiotic-induced creation of “ecological vacancies” or “niche opportunities” in the gut allows even small, previously insignificant populations of surviving or newly arriving oral bacteria to establish stable colonization in the absence of competition. Normally benign oral commensals like Klebsiella pneumoniae and Enterococcus faecalis can become dominant gut inhabitants following antibiotic treatment and trigger robust inflammatory responses. Ectopic colonization by these oral strains activates innate immune pathways, including the NLRP3 inflammasome, and potently induces pro-inflammatory T helper 1 (Th1) cell responses, leading to significant colonic inflammation and tissue damage [21]. Furthermore, the overall degree of gut microbial depletion correlates with a broader impairment of mucosal immunity, including reduced fecal IgA levels and decreased numbers of anti-inflammatory regulatory T cells (Tregs) in the intestinal lamina propria, populations essential for maintaining tolerance [22,23]. Oral Porphyromonas gingivalis can exploit this dysregulated, inflamed environment, exacerbating colitis by skewing the gut microbiota and its metabolic output, specifically through disrupting linoleic acid metabolism and unbalancing the Th17/Treg axis [24]. Interestingly, this depleted, low-diversity state influences the success of microbiome-based therapies. In fecal microbiota transplantation (FMT), recipients with a pre-treatment gut microbiome characterized by low diversity and high relative abundance of oral taxa—a signature of prior antibiotic use—show higher engraftment success of donor microbiota. This suggests that antibiotic-induced ecological voids enhance niche availability, and some oral commensals like Streptococcus salivarius may even play a facilitatory role in donor community establishment [25].
3.3. Proton Pump Inhibitors: Disabling the Gastric Acid Gatekeeper
PPIs are among the most frequently prescribed medications in hospitalized patients, routinely administered for stress ulcer prophylaxis in the critically ill. By potently and persistently suppressing gastric acid secretion, they effectively dismantle a primary chemical barrier of the oral-intestinal axis. However, long-term PPI use is associated with multiple adverse outcomes, including increased risk of gastrointestinal infections (e.g., C. difficile), community-acquired pneumonia, nutrient deficiencies, and potentially cardiovascular and renal events [26,27,28]. Large-scale observational microbiome studies provide consistent evidence: PPI use is associated with reduced gut microbial α-diversity and a compositional shift towards a community structure that resembles dysbiotic states. There is a marked increase in facultative anaerobes typically enriched in the upper gastrointestinal tract and oral cavity, such as Enterococcus, Streptococcus, Staphylococcus, and Escherichia coli [29,30,31,32,33]. Metagenomic sequencing studies have confirmed and detailed this phenomenon, identifying a consistent enrichment of oral commensals, with genera like Streptococcus and Lactobacillus being particularly prominent in the gut microbiomes of PPI users [29,30,31,32,33]. These effects appear to be a class-wide, dose-dependent phenomenon [29,30,31,32,33].
A well-designed 2024 randomized controlled trial provided direct causal evidence in humans, contrasting PPIs with histamine H2-receptor antagonists (H2RAs) [29,30,31,32,33]. In healthy volunteers, just seven days of PPI use (but not H2RA) significantly increased the transmission of oral microbes to the gut and promoted the intestinal colonization of specific oral bacteria linked to disease, namely Fusobacterium nucleatum and Streptococcus anginosus. Crucially, the composition of the oral microbiome itself was not altered by PPI therapy, pinpointing the weakened gastric acid barrier—not an oral source shift—as the primary culprit. In vitro validation confirmed that S. anginosus could not survive at a pH below 5.0, directly linking its survival through the stomach to pharmacological acid suppression. An instructive secondary finding was that the concurrent use of chlorhexidine mouthwash reduced the PPI-driven rise in gut S. anginosus, demonstrating that interventions targeting the oral reservoir can modulate downstream consequences in the gut.
3.4. The Synergistic Cascade: A Trajectory of Escalating Dysbiosis
The clinical reality in the ICU is that these three factors rarely act in isolation; they interact synergistically and sequentially, creating an escalatory cascade of dysbiosis.
Step 1: Reservoir Creation. Endotracheal intubation and impaired consciousness promote oral dysbiosis and the overgrowth of pathogens, creating a loaded “gun”. Step 2: Gate Opening. Concurrent PPI use reduces gastric acidity, disabling a key safety mechanism and allowing these acid-sensitive oral pathobionts to survive transit, effectively “pulling the trigger”. Step 3: Ground Preparation. Broad-spectrum antibiotic therapy devastates the indigenous gut microbiota, destroying colonization resistance and creating vacant ecological niches in the intestine, “preparing the battlefield”. The translocated oral bacteria then colonize the vulnerable gut ecosystem, where they can disrupt barrier function, activate local and systemic immune pathways, and fuel inflammation. This cascade is a fundamental, underappreciated driver of ICU-acquired complications, conceptually linking routine oral care directly to outcomes far beyond the prevention of pneumonia.
4. Oral Care Reimagined: From Chemical Antisepsis to Ecological Stewardship in the ICU
Given the oral cavity’s demonstrated role as a nexus for systemic infection and inflammation, meticulous oral care is a non-negotiable component of high-quality critical care. Its implementation, however, is fraught with practical challenges: the physical impediment of endotracheal and other tubes, limited patient cooperation due to sedation or delirium, variability in nursing knowledge and practice, and a frequent lack of standardized, protocol-driven approaches [29,30,31,32,33]. A systematic review of practices confirms this variability and underscores the need for evidence-based protocols [92]. The traditional and primary goal of oral care has been the prevention of ventilator-associated pneumonia (VAP), justified by the well-established pathogenesis of microaspiration of oropharyngeal secretions into the lower airways [93,94]. Microbial sequencing studies consistently demonstrate shared bacterial strains between the oral niche and the lungs in patients with VAP [95,96,97], and a high oral bacterial load at ICU admission has been identified as an independent predictor for the subsequent development of both VAP and delirium [98].
The colonization of dental plaque by respiratory pathogens is a key event preceding infection [99,100], and genetic studies confirm the relatedness of isolates from plaque and the lungs in VAP patients [99,100]. Even commensal oral communities can interfere with the integration of pathogens like P. aeruginosa [102], but this balance is lost in dysbiosis. The etiological agents of VAP often originate from this oral reservoir [103], with pathogens such as P. gingivalis implicated in worsening aspiration pneumonia [103]. Early trials demonstrated that oral care could reduce pneumonia incidence and modify mortality risk [105,106], laying the foundation for modern protocols.
However, in light of the oral-gut axis, the purpose and potential impact of oral care must be expansively redefined. It is not solely a pulmonary protection strategy but a fundamental intervention to protect the integrity of the oral-gut axis and mitigate the systemic inflammatory cascade that can originate from it [35,36,37,38,39].
4.1. The Fall of a Former Standard: Re-Evaluating Chlorhexidine
For decades, chlorhexidine gluconate (CHG) mouthwash or gel was the cornerstone of ICU oral care protocols, prized for its broad-spectrum bactericidal and fungicidal activity. This practice was bundled into VAP prevention guidelines worldwide. However, accumulating evidence from high-quality meta-analyses has fundamentally challenged this doctrine. While CHG reliably reduces quantitative oral bacterial colonization, it has not been shown to significantly reduce the incidence of VAP in general, mixed medical-surgical ICU populations [35,36,37,38,39]. More alarmingly, several meta-analyses have reported a signal of increased mortality among patients receiving routine CHG, prompting serious safety concerns [107,109,110,111]. The evidence for combining toothbrushing with CHG also requires careful interpretation, as benefits may stem from the mechanical action rather than the antiseptic [112].
The mechanisms underlying this potential for harm are multifactorial and align with principles of microbial ecology. First, mucosal toxicity: Frequent application, particularly of the 2% concentration, is associated with a significant incidence of oral mucosal injury, including ulcerations, erythema, and desquamation [113]. These lesions compromise the epithelial barrier, potentially increasing pain, discouraging adequate care, and facilitating the systemic translocation of bacteria. Second, emerging resistance: There is growing documentation of reduced susceptibility to CHG among relevant ICU pathogens such as Pseudomonas aeruginosa, methicillin-resistant Staphylococcus aureus (MRSA), and Enterobacter species [114,115]. This resistance may develop under the selective pressure of long-term, sub-lethal exposure. Third, and perhaps most critically in the context of the microbiome axis, ecological disruption: CHG’s non-selective, “scorched-earth” antimicrobial approach disrupts the balance of the oral microbiome. By indiscriminately suppressing commensal Streptococci and other early colonizers that contribute to a healthy microbial landscape, it creates ecological vacancies. These vacancies can then be exploited by more virulent, often multidrug-resistant, opportunistic pathogens, potentially paradoxically increasing the risk of subsequent infection. Furthermore, a damaged mucosa combined with an altered microbiome may paradoxically enhance biofilm formation on endotracheal tubes. A multi-center stepped-wedge cluster RCT (the CHORAL trial) found that discontinuing chlorhexidine and implementing a bundled oral care protocol did not worsen mortality or other key outcomes, supporting the safety of de-adopting this agent [116].
4.2. The Rise of Evidence-Based, Ecological Alternatives
This compelling evidence has catalyzed a paradigm shift in ICU oral care, moving away from reliance on non-selective chemical agents towards strategies that are more physiological, targeted, and focused on mechanical disruption (Figure 3).
4.2.1. Mechanical Debridement: Re-Establishing Tooth Brushing as the Cornerstone
The physical removal of dental plaque biofilm using a soft-bristled toothbrush is now unequivocally recognized as the foundational practice. A landmark 2024 systematic review and meta-analysis encompassing 15 randomized trials and over 10,000 patients, provided robust, high-level evidence [117]. The analysis concluded that daily tooth brushing significantly reduced the risk of hospital-acquired pneumonia (HAP) and ICU mortality among mechanically ventilated patients. It was also associated with clinically meaningful reductions in the duration of mechanical ventilation (by approximately 1.24 days) and ICU length of stay (by about 1.78 days). Importantly, no additional benefit was observed with brushing more than twice daily, suggesting a ceiling effect for frequency. This mechanical action directly targets the structured, adherent biofilm reservoir—a sanctuary for pathogens—without causing the collateral ecological damage associated with antiseptics. Reflecting this evidence, guidelines from the Society for Healthcare Epidemiology of America (SHEA) now strongly recommend tooth brushing as a core component of oral care for ventilated patients and explicitly advise against the routine use of chlorhexidine [118].
4.2.2. Physiological Irrigation: The Re-Emergence of Normal Saline
In response to the concerns surrounding antiseptics, normal saline (0.9% sodium chloride) has re-emerged as a safe and effective irrigation solution. A network meta-analysis comparing various oral care solutions found that while antiseptic agentslike chlorhexidine were associated with higher mortality risks, normal saline was associated with a significant reduction in mortality [119]. Saline works primarily through mechanical action: it loosens, dilutes, and flushes away debris, thick secretions, and loosely adherent biofilm. Its advantages are its safety profile—it is isotonic, non-irritating, and poses no risk of mucosal injury or antimicrobial resistance—and its ecological neutrality. It cleanses without attempting to sterilize, thereby preserving the existing microbiome structure. When combined with regular tooth brushing and suctioning, saline-based care offers a gentle, effective, and low-risk approach to maintaining oral hygiene, particularly suitable for patients with fragile oral tissues or where microbiome preservation is a priority [120,121].
4.2.3. Selective Oropharyngeal Decontamination (SOD): A Paradigm of Precision Targeting
SOD represents a sophisticated “middle path” between non-selective antiseptics and purely mechanical care. It involves the topical application of a paste or gel containing non-absorbable antibiotics (typically polymyxin E/colistin and tobramycin) and an antifungal agent (nystatin) directly to the oropharyngeal mucosa. The philosophy is one of precision: unlike chlorhexidine, SOD aims to selectively suppress known, high-threat pathogenic Gram-negative bacilli (e.g., Pseudomonas aeruginosa, Klebsiella pneumoniae) and Candida species without causing broad disruption to the commensal oral and gut flora [120,121].
Large-scale, cluster-randomized controlled trials, most notably the Dutch series of studies, have consistently demonstrated that SOD—and its more comprehensive counterpart, Selective Digestive Decontamination (SDD), which adds enteral and short-term intravenous antibiotics—reduces mortality in critically ill, mechanically ventilated patients compared to standard care, and is superior to chlorhexidine-based care [109,122]. A recent multinational SuDDICU trial provides the largest and most contemporary assessment of SDD’s impact in a broader ICU context [123]. In over 9,000 mechanically ventilated patients, SDD did not achieve a statistically significant reduction in the primary endpoint of 90-day in-hospital mortality. However, it delivered significant and clinically important reductions in key secondary infectious outcomes: the incidence of new bloodstream infections (4.9% vs. 6.8%) and the acquisition of new antibiotic-resistant organisms (16.8% vs. 26.8%). This dissociation between mortality and infection endpoints underscores the multifactorial nature of death in critical illness but firmly establishes SDD’s efficacy in mitigating specific complications linked to oral-gut axis disruption. A critical and counterintuitive finding from these long-term studies is that neither SOD nor SDD leads to an increase in the prevalence of antibiotic-resistant Gram-negative bacteria in the ICU. In fact, they are associated with a decreased carriage and infection rate of bacteria resistant to third-generation cephalosporins and carbapenems [122,124,125]. This is likely because by preventing heavy colonization and overgrowth of these pathogens, the interventions reduce the overall bacterial burden and patient-to-patient transmission, offsetting any local selective pressure from the topical agents. A five-year case-control study further reinforced these benefits, showing that SOD implementation was associated with a lower incidence of infections with extended-spectrum β-lactamase (ESBL)-producing Klebsiella pneumoniae, as well as reduced rates of VAP and ICU mortality [126]. SOD thus stands as a powerful example of how a localized, microbiome-sparing intervention can yield significant systemic benefits, though its adoption requires careful consideration of local resistance patterns and institutional protocols.
5. Conclusions and Future Prospects
The integrity of the oral-gut microbiome axis—a bidirectional ecosystem essential for systemic immune homeostasis—is systematically disrupted in the ICU by life-sustaining interventions. This synergy drives dysbiosis and pathogenic microbial translocation, contributing to ICU-acquired infections and organ dysfunction. Consequently, evidence-based oral care must evolve from a focus on non-selective antisepsis toward an ecological stewardship model centered on mechanical plaque control, physiological cleansing, and, in appropriate settings, selective decontamination. Preserving this axis represents a tangible strategy to enhance microbial and immune resilience, attenuate systemic inflammation, and potentially improve outcomes in critical illness.
5.1. Expanding the Research Horizon: From Pulmonary to Systemic Outcomes
Research must extend beyond VAP to investigate the systemic implications of the oral-gut axis in critical illness. Prospective, multicenter mechanistic studies employing multi-omics technologies are needed to trace the real-time effects of oral care interventions along the axis and identify biomarkers of efficacy.
5.2. Exploring Novel Microbiome-Targeted Therapies
Future work should evaluate adjunctive therapies such as next-generation probiotics, prebiotics, and postbiotics, as well as the role of FMT in specific patient subsets [126]. A holistic “microbiome stewardship” approach should also consider the ecological impact of systemic medications.
5.3. Refining SDD/SOD Strategies: Towards Precision Application
The heterogeneous response to SDD/SOD calls for precision application. Notably, prespecified subgroup analysis of the SuDDICU trial revealed a significant survival benefit in patients with acute brain injury [123]. This finding underscores the need for RCTs targeted at well-defined high-risk subgroups, such as patients with acute brain injury, burns, immunocompromising conditions, or those with prolonged expected mechanical ventilation duration. Efficacy and safety of these strategies in settings with high baseline antimicrobial resistance remain unproven and require urgent study. Research should also explore simplified or “light” protocols to preserve benefits while minimizing ecological risk.

Table 1.
Factors Affecting the Oral-Gut Microbiome Axis in the ICU and Their Mechanisms.
| Factor | Mechanism of Disruption | Clinical Implications |
|---|---|---|
| Endotracheal Intubation | Impairs salivary flow, reduces swallowing, suppresses speech and mastication, decreases natural oral clearance, promotes biofilm formation, favors pathogenic overgrowth (e.g., Staphylococcus aureus, Pseudomonas aeruginosa, Candida albicans). | Increased oral dysbiosis, higher risk of aspiration, ventilator-associated pneumonia (VAP), gut colonization by oral pathogens, potential progression to enteric sepsis and multiple organ dysfunction. |
| Antibiotics | Depletes commensal gut microbiota, reduces colonization resistance, promotes relative enrichment of oral bacteria in the gut (e.g., Klebsiella, Enterococcus), disrupts immune homeostasis (↓ IgA, ↓ Tregs). | Intestinal dysbiosis, immune dysfunction, increased risk of Clostridioides difficile infection, systemic inflammation, and impaired mucosal immunity. |
| Proton Pump Inhibitors (PPIs) | Reduces gastric acidity (pH ↑), weakens chemical barrier, enhances survival and colonization of acid-sensitive oral bacteria (e.g., Streptococcus anginosus, Fusobacterium nucleatum), increases oral-to-gut transmission. | Increased risk of gastrointestinal infections (e.g., C. difficile), colorectal cancer, systemic inflammation, altered gut microbiota composition (↑ Streptococcus, ↑ Lactobacillus). |
| Sedation & Immobility | Decreases oral and gut motility, impairs secretion clearance, reduces cough reflex and mucociliary clearance, promotes stasis and bacterial overgrowth. | Facilitates microbial translocation, exacerbates dysbiosis, increases risk of aspiration, VAP, and gut-derived sepsis. |
Abbreviations: ICU, intensive care unit; VAP, ventilator-associated pneumonia; PPIs, proton pump inhibitors; Tregs, regulatory T cells; IgA, immunoglobulin A.
Table 2.
Comparison of Oral Care Strategies in the ICU.
| Oral Care Method | Key Features/Mechanism | Clinical Outcomes/Evidence | Practical Considerations |
|---|---|---|---|
| Chlorhexidine-based Care | Broad-spectrum antiseptic; reduces bacterial load; easy to apply. | Associated with mucosal injury, microbial resistance, ecological disruption, potential ↑ mortality; no significant reduction in VAP incidence. | Use discouraged in recent guidelines; avoid routine use in mixed ICU populations. |
| Standardized Oral Care without Chlorhexidine | Bundled care: oral assessment, tooth brushing, moisture maintenance, secretion aspiration; no antiseptic. | No significant mortality benefit; may improve oral health; feasible and acceptable in clinical practice. | Emphasizes mechanical cleaning and mucosal protection; suitable for most ventilated patients. |
| Saline-based Oral Care | Isotonic saline irrigation; mechanical flushing; preserves microbiome integrity. | Safe, non-toxic; associated with reduced mortality in network meta-analysis; does not promote resistance. | Ideal for patients with mucosal compromise, renal dysfunction, or where microbiome preservation is prioritized. |
| Tooth Brushing | Physically disrupts dental plaque and biofilms; reduces microbial burden; uses soft-bristled brush. | Reduces HAP/VAP incidence, shortens ICU stay and mechanical ventilation duration; recommended by SHEA. | Should be done ≥ twice daily; no additional benefit from more frequent brushing; compatible with other oral care methods. |
| Selective Oropharyngeal Decontamination (SOD) | Topical non-absorbable antibiotic paste (e.g., polymyxin, tobramycin, nystatin) applied to oropharynx. | Reduces ICU-acquired bacteremia, VAP, and 28-day mortality; does not increase antimicrobial resistance in low/moderate resistance settings. | Best suited for high-risk patients in controlled resistance environments; requires monitoring. |
| Saliva Substitutes/Moisturizers | Artificial saliva or moisturizing gel; maintains mucosal integrity and hydration. | Prevents xerostomia, supports natural antimicrobial peptides and mucosal barrier function. | Often used as adjunct to brushing or saline care; especially useful in sedated or dehydrated patients. |
Abbreviations: ICU, intensive care unit; VAP, ventilator-associated pneumonia; HAP, hospital-acquired pneumonia; SHEA, Society for Healthcare Epidemiology of America; SOD, selective oropharyngeal decontamination.
Author Contributions
All authors contributed to the literature search and acquisition of data, drafting of the initial manuscript and critical revision of the final manuscript and approved the final version of the manuscript for publication.
Funding
The study was funded by the National Natural Science Foundation of China (Grant No. 82472223 [to JCZ]), Noncommunicable Chronic Diseases-National Science and Technology Major Project (Grant No. 2024ZD0529004 [to JCZ]).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
All authors have provided consent for publication.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no competing interests.
Abbreviations
ICU, intensive care unit;
PPIs, proton pump inhibitors;
FMT, fecal microbiota transplantation;
IBD, inflammatory bowel disease;
IBS, irritable bowel syndrome;
COPD, chronic obstructive pulmonary disease;
LPS, lipopolysaccharide;
DCs, dendritic cells;
NLRP3, NOD-, LRR-, and pyrin domain-containing protein 3;
VAP, ventilator-associated pneumonia;
Tregs, regulatory T cells;
RCT, randomized controlled trial;
HAP, hospital-acquired pneumonia;
SOD, selective oropharyngeal decontamination;
SDD, selective digestive tract decontamination.
References
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Leonov, G.; Salikhova, D.; Starodubova, A.; Vasilyev, A.; Makhnach, O.; Fatkhudinov, T.; Goldshtein, D. Oral microbiome dysbiosis as a risk factor for stroke: A comprehensive review. Microorganisms 2024, 12, 1732. [Google Scholar] [CrossRef]
- Xu, Q.; Wang, W.; Li, Y.; Cui, J.; Zhu, M.; Liu, Y.; Liu, Y. The oral-gut microbiota axis: A link in cardiometabolic diseases. Npj Biofilms Microbiomes 2025, 11, 11. [Google Scholar] [CrossRef]
- Dodson, B.; Suner, T.; Haar, E.L.V.; Han, Y.W. Oral microbiome and adverse pregnancy outcomes. Am. J. Reprod. Immunol. 2025, 93, e70107. [Google Scholar] [CrossRef] [PubMed]
- Clasen, F.; Yildirim, S.; Arıkan, M.; Garcia-Guevara, F.; Hanoğlu, L.; Yılmaz, N.H.; şen, A.; Celik, H.K.; Neslihan, A.A.; Demir, T.K.; et al. Microbiome signatures of virulence in the oral-gut-brain axis influence Parkinson’s disease and cognitive decline pathophysiology. Gut Microbes 2025, 17, 2506843. [Google Scholar] [CrossRef]
- Kuraji, R.; Shiba, T.; Dong, T.S.; Numabe, Y.; Kapila, Y.L. Periodontal treatment and microbiome-targeted therapy in management of periodontitis-related nonalcoholic fatty liver disease with oral and gut dysbiosis. World J. Gastroenterol. 2023, 29, 967–996. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, F.; Gtif, I.; Alfadhli, S.; Charfeddine, S.; Ghorbel, W.; Abdelhédi, R.; Benmarzoug, R.; Abid, L.; Bouayed Abdelmoula, N.; Elloumi, I.; et al. A potential oral microbiome signature associated with coronary artery disease in Tunisia. Biosci. Rep. 2022, 42, R20220583. [Google Scholar] [CrossRef]
- González-Febles, J.; Sanz, M. Periodontitis and rheumatoid arthritis: What have we learned about their connection and their treatment? Periodontology 2000 2021, 87, 181–203. [Google Scholar] [CrossRef]
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Chavakis, T. Local and systemic mechanisms linking periodontal disease and inflammatory comorbidities. Nat. Rev. Immunol. 2021, 21, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, M.; Ready, D.; Brealey, D.; Ryu, J.; Bercades, G.; Nagle, J.; Borja-Boluda, S.; Agudo, E.; Petrie, A.; Suvan, J.; et al. Changes in dental plaque following hospitalisation in a critical care unit: An observational study. Crit. Care 2013, 17, R189. [Google Scholar] [CrossRef] [PubMed]
- Lagerlöf, F.; Dawes, C. The effect of swallowing frequency on oral sugar clearance and pH changes by Streptococcus mitior in vivo after sucrose ingestion. J. Dent. Res. 1985, 64, 1229–1232. [Google Scholar] [CrossRef]
- Chevalier, M.; Medioni, E.; Prêcheur, I. Inhibition of Candida albicans yeast-hyphal transition and biofilm formation by Solidago virgaurea water extracts. J. Med. Microbiol. 2012, 61, 1016–1022. [Google Scholar] [CrossRef]
- Kanamori, D.; Fujii, T.; Yoshida, M.; Ito, R.; Sakai, A.; Takahashi, H.; Kuramitsu, K.; Funasaka, K.; Ohno, E.; Hirooka, Y.; et al. Oral care for intubated patients in the intensive care unit: Examination of bacterial count and microbiota. Crit. Care 2025, 29, 320. [Google Scholar] [CrossRef]
- Shi, T.; Wang, J.; Dong, J.; Hu, P.; Guo, Q. Periodontopathogens Porphyromonas gingivalis and Fusobacterium nucleatum and their Roles in the Progression of Respiratory Diseases. Pathogens 2023, 12, 1110. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wang, B.; Gao, H.; He, C.; Hua, R.; Liang, C.; Xin, S.; Wang, Y.; Xu, J. Insight into the Relationship between Oral Microbiota and the Inflammatory Bowel Disease. Microorganisms 2022, 10, 1868. [Google Scholar] [CrossRef]
- Maier, L.; Goemans, C.V.; Wirbel, J.; Kuhn, M.; Eberl, C.; Pruteanu, M.; Müller, P.; Garcia-Santamarina, S.; Cacace, E.; Zhang, B.; et al. Unravelling the collateral damage of antibiotics on gut bacteria. Nature 2021, 599, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Fishbein, S.R.S.; Mahmud, B.; Dantas, G. Antibiotic perturbations to the gut microbiome. Nat. Rev. Microbiol. 2023, 21, 772–788. [Google Scholar] [CrossRef]
- Liao, C.; Rolling, T.; Djukovic, A.; Fei, T.; Mishra, V.; Liu, H.; Lindberg, C.; Dai, L.; Zhai, B.; Peled, J.U.; et al. Oral bacteria relative abundance in faeces increases due to gut microbiota depletion and is linked with patient outcomes. Nat. Microbiol. 2024, 9, 1555–1565. [Google Scholar] [CrossRef]
- Jin, S.; Wetzel, D.; Schirmer, M. Deciphering mechanisms and implications of bacterial translocation in human health and disease. Curr. Opin. Microbiol. 2022, 67, 102147. [Google Scholar] [CrossRef]
- Atarashi, K.; Suda, W.; Luo, C.; Kawaguchi, T.; Motoo, I.; Narushima, S.; Kiguchi, Y.; Yasuma, K.; Watanabe, E.; Tanoue, T.; et al. Ectopic colonization of oral bacteria in the intestine drives T(H)1 cell induction and inflammation. Science 2017, 358, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Contijoch, E.J.; Britton, G.J.; Yang, C.; Mogno, I.; Li, Z.; Ng, R.; Llewellyn, S.R.; Hira, S.; Johnson, C.; Rabinowitz, K.M.; et al. Gut microbiota density influences host physiology and is shaped by host and microbial factors. eLife 2019, 8, e40553. [Google Scholar] [CrossRef]
- Britton, G.J.; Contijoch, E.J.; Spindler, M.P.; Aggarwala, V.; Dogan, B.; Bongers, G.; San Mateo, L.; Baltus, A.; Das, A.; Gevers, D.; et al. Defined microbiota transplant restores Th17/RORγt(+) regulatory T cell balance in mice colonized with inflammatory bowel disease microbiotas. Proc. Natl. Acad. Sci. USA 2020, 117, 21536–21545. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Jiang, Y.; Wu, L.; Fu, J.; Du, J.; Luo, Z.; Guo, L.; Xu, J.; Liu, Y. Porphyromonas gingivalis aggravates colitis via a gut microbiota-linoleic acid metabolism-Th17/Treg cell balance axis. Nat. Commun. 2024, 15, 1617. [Google Scholar] [CrossRef]
- Schmidt, T.S.B.; Li, S.S.; Maistrenko, O.M.; Akanni, W.; Coelho, L.P.; Dolai, S.; Fullam, A.; Glazek, A.M.; Hercog, R.; Herrema, H.; et al. Drivers and determinants of strain dynamics following fecal microbiota transplantation. Nat. Med. 2022, 28, 1902–1912. [Google Scholar] [CrossRef] [PubMed]
- Willems, R.P.J.; Schut, M.C.; Kaiser, A.M.; Groot, T.H.; Abu-Hanna, A.; Twisk, J.W.R.; van Dijk, K.; Vandenbroucke-Grauls, C.M.J.E. Association of proton pump inhibitor use with risk of acquiring Drug-Resistant enterobacterales. JAMA Netw. Open 2023, 6, e230470. [Google Scholar] [CrossRef]
- de Jager, C.P.C.; Wever, P.C.; Gemen, E.F.A.; van Oijen, M.G.H.; van Gageldonk-Lafeber, A.B.; Siersema, P.D.; Kusters, G.C.M.; Laheij, R.J.F. Proton pump inhibitor therapy predisposes to community-acquired Streptococcus pneumoniae pneumonia. Aliment. Pharmacol. Ther. 2012, 36, 941–949. [Google Scholar] [CrossRef]
- Haastrup, P.F.; Thompson, W.; Søndergaard, J.; Jarbøl, D.E. Side effects of Long-Term proton pump inhibitor use: A review. Basic Clin. Pharmacol. Toxicol. 2018, 123, 114–121. [Google Scholar] [CrossRef]
- Jackson, M.A.; Verdi, S.; Maxan, M.; Shin, C.M.; Zierer, J.; Bowyer, R.C.E.; Martin, T.; Williams, F.M.K.; Menni, C.; Bell, J.T.; et al. Gut microbiota associations with common diseases and prescription medications in a population-based cohort. Nat. Commun. 2018, 9, 2655. [Google Scholar] [CrossRef]
- Imhann, F.; Bonder, M.J.; Vich Vila, A.; Fu, J.; Mujagic, Z.; Vork, L.; Tigchelaar, E.F.; Jankipersadsing, S.A.; Cenit, M.C.; Harmsen, H.J.M.; et al. Proton pump inhibitors affect the gut microbiome. Gut 2016, 65, 740–748. [Google Scholar] [CrossRef]
- Forslund, S.K.; Chakaroun, R.; Zimmermann-Kogadeeva, M.; Markó, L.; Aron-Wisnewsky, J.; Nielsen, T.; Moitinho-Silva, L.; Schmidt, T.S.B.; Falony, G.; Vieira-Silva, S.; et al. Combinatorial, additive and dose-dependent drug-microbiome associations. Nature 2021, 600, 500–505. [Google Scholar] [CrossRef]
- Nagata, N.; Nishijima, S.; Miyoshi-Akiyama, T.; Kojima, Y.; Kimura, M.; Aoki, R.; Ohsugi, M.; Ueki, K.; Miki, K.; Iwata, E.; et al. Population-level metagenomics uncovers distinct effects of multiple medications on the human gut microbiome. Gastroenterology 2022, 163, 1038–1052. [Google Scholar] [CrossRef]
- Vich Vila, A.; Collij, V.; Sanna, S.; Sinha, T.; Imhann, F.; Bourgonje, A.R.; Mujagic, Z.; Jonkers, D.M.A.E.; Masclee, A.A.M.; Fu, J.; et al. Impact of commonly used drugs on the composition and metabolic function of the gut microbiota. Nat. Commun. 2020, 11, 362. [Google Scholar] [CrossRef]
- Zhu, J.; Sun, C.; Li, M.; Hu, G.; Zhao, X.; Chen, W. Compared to histamine-2 receptor antagonist, proton pump inhibitor induces stronger oral-to-gut microbial transmission and gut microbiome alterations: A randomised controlled trial. Gut 2024, 73, 1087–1097. [Google Scholar] [CrossRef]
- Patel, J.J.; Barash, M. The gut in critical illness. Curr. Gastroenterol. Rep. 2025, 27, 11. [Google Scholar] [CrossRef] [PubMed]
- Violi, F.; Cammisotto, V.; Bartimoccia, S.; Pignatelli, P.; Carnevale, R.; Nocella, C. Gut-derived low-grade endotoxaemia, atherothrombosis and cardiovascular disease. Nat. Rev. Cardiol. 2023, 20, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, H.; Beckmann, T.S.; Fröhlich, L.; Soccorsi, T.; Le Terrier, C.; de Watteville, A.; Schrenzel, J.; Heidegger, C. The central and biodynamic role of gut microbiota in critically ill patients. Crit. Care 2022, 26, 250. [Google Scholar] [CrossRef] [PubMed]
- Xie, B.; Wang, M.; Zhang, X.; Zhang, Y.; Qi, H.; Liu, H.; Wu, Y.; Wen, X.; Chen, X.; Han, M.; et al. Gut-derived memory γδ T17 cells exacerbate sepsis-induced acute lung injury in mice. Nat. Commun. 2024, 15, 6737. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Y.; Yu, Y.; Wang, X.; Zhen, Z.; Yuan, Y.; Xie, B.; Han, M.; Wang, M.; Zhang, X.; et al. Small intestinal γδ T17 cells promote SAE through STING/C1q-induced microglial synaptic pruning in male mice. Nat. Commun. 2025, 16, 6779. [Google Scholar] [CrossRef]
- Cho, N.A.; Strayer, K.; Dobson, B.; Mcdonald, B. Pathogenesis and therapeutic opportunities of gut microbiome dysbiosis in critical illness. Gut Microbes 2024, 16, 2351478. [Google Scholar] [CrossRef]
- Van Hul, M.; Cani, P.D.; Petitfils, C.; De Vos, W.M.; Tilg, H.; El-Omar, E.M. What defines a healthy gut microbiome? Gut 2024, 73, 1893–1908. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.; Chen, X.; Wang, J.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in health and diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Ding, T.; Li, H. Oral microbiome in human health and diseases. mLife 2024, 3, 367–383. [Google Scholar] [CrossRef]
- Willis, J.R.; Gabaldón, T. The human oral microbiome in health and disease: From sequences to ecosystems. Microorganisms 2020, 8, 308. [Google Scholar] [CrossRef]
- Baker, J.L.; Mark Welch, J.L.; Kauffman, K.M.; Mclean, J.S.; He, X. The oral microbiome: Diversity, biogeography and human health. Nat. Rev. Microbiol. 2024, 22, 89–104. [Google Scholar] [CrossRef]
- Kitamoto, S.; Nagao-Kitamoto, H.; Hein, R.; Schmidt, T.M.; Kamada, N. The bacterial connection between the oral cavity and the gut diseases. J. Dent. Res. 2020, 99, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Zhou, F.; Wang, H.; Xu, X.; Xu, S.; Zhang, C.; Zhang, Y.; Lu, M.; Zhang, Y.; Zhou, M.; et al. Systemic antibiotics increase microbiota pathogenicity and oral bone loss. Int. J. Oral Sci. 2023, 15, 4. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.K.; Tong, S.L.Y.; Wong, M.C.S.; Chan, J.Y.K.; Ip, M.; Hui, M.; Lai, C.K.C.; Ng, R.W.Y.; Ho, W.C.S.; Yeung, A.C.M.; et al. Extent of Oral-Gut transmission of bacterial and fungal microbiota in healthy chinese adults. Microbiol. Spectr. 2023, 11, e281422. [Google Scholar] [CrossRef]
- Murgolo, N.J.; Cerami, A.; Henderson, G.B. Biomedical science and the third world. Under the volcano. Trypanothione reductase. Ann. N. Y. Acad. Sci. 1989, 569, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Nuccio, S.; Mohanty, I.; Hagey, L.R.; Dorrestein, P.C.; Chu, H.; Raffatellu, M. How bile acids and the microbiota interact to shape host immunity. Nat. Rev. Immunol. 2024, 24, 798–809. [Google Scholar] [CrossRef]
- Neurath, M.F.; Artis, D.; Becker, C. The intestinal barrier: A pivotal role in health, inflammation, and cancer. Lancet Gastroenterol. Hepatol. 2025, 10, 573–592. [Google Scholar] [CrossRef]
- Woelfel, S.; Silva, M.S.; Stecher, B. Intestinal colonization resistance in the context of environmental, host, and microbial determinants. Cell Host Microbe 2024, 32, 820–836. [Google Scholar] [CrossRef] [PubMed]
- Iliev, I.D.; Ananthakrishnan, A.N.; Guo, C. Microbiota in inflammatory bowel disease: Mechanisms of disease and therapeutic opportunities. Nat. Rev. Microbiol. 2025, 23, 509–524. [Google Scholar] [CrossRef]
- Rashidi, A.; Ebadi, M.; Weisdorf, D.J.; Costalonga, M.; Staley, C. No evidence for colonization of oral bacteria in the distal gut in healthy adults. Proc. Natl. Acad. Sci. USA 2021, 118, e2114152118. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.S.; Hayward, M.R.; Coelho, L.P.; Li, S.S.; Costea, P.I.; Voigt, A.Y.; Wirbel, J.; Maistrenko, O.M.; Alves, R.J.; Bergsten, E.; et al. Extensive transmission of microbes along the gastrointestinal tract. eLife 2019, 8, e42693. [Google Scholar] [CrossRef]
- Könönen, E.; Gursoy, U.K. Oral prevotella species and their connection to events of clinical relevance in gastrointestinal and respiratory tracts. Front. Microbiol. 2022, 12, 798763. [Google Scholar] [CrossRef]
- Bao, J.; Li, L.; Zhang, Y.; Wang, M.; Chen, F.; Ge, S.; Chen, B.; Yan, F. Periodontitis may induce gut microbiota dysbiosis via salivary microbiota. Int. J. Oral Sci. 2022, 14, 32. [Google Scholar] [CrossRef]
- Kunath, B.J.; De Rudder, C.; Laczny, C.C.; Letellier, E.; Wilmes, P. The oral-gut microbiome axis in health and disease. Nat. Rev. Microbiol. 2024, 22, 791–805. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, J.; Chen, Y.; Chen, T.; Shen, Z.; Wang, Y.; Jian, Y.; Xiang, G.; Ma, X.; Zhao, N.; et al. Veillonella intestinal colonization promotes C. Difficile infection in Crohn’s disease. Cell Host Microbe 2025, 33, 1518–1534. [Google Scholar] [CrossRef]
- Wang, Y.; Mu, W.; Guan, J.; Ma, P.; Li, Y.; Zhang, Y.; Zhu, W.; Zhou, Y.; Zou, Y.; Zeng, T.; et al. Bile acid synthesis dysregulation in liver diseases promotes ectopic expansion of oral streptococci in the intestine. Cell Rep. 2025, 44, 116374. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Cao, P.; Su, W.; Zhan, N.; Dong, W. Fusobacterium nucleatum facilitates ulcerative colitis through activating IL-17F signaling to NF-κB via the upregulation of CARD3 expression. J. Pathol. 2020, 250, 170–182. [Google Scholar] [CrossRef]
- Arimatsu, K.; Yamada, H.; Miyazawa, H.; Minagawa, T.; Nakajima, M.; Ryder, M.I.; Gotoh, K.; Motooka, D.; Nakamura, S.; Iida, T.; et al. Oral pathobiont induces systemic inflammation and metabolic changes associated with alteration of gut microbiota. Sci. Rep. 2014, 4, 4828. [Google Scholar] [CrossRef]
- Xiao, L.; Huang, L.; Zhou, X.; Zhao, D.; Wang, Y.; Min, H.; Song, S.; Sun, W.; Gao, Q.; Hu, Q.; et al. Experimental periodontitis deteriorated atherosclerosis associated with trimethylamine N-Oxide metabolism in mice. Front. Cell. Infect. Microbiol. 2022, 11, 820535. [Google Scholar] [CrossRef]
- Gimbrone, M.A., Jr.; García-Cardeña, G. Endothelial cell dysfunction and the pathobiology of atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, M.; Huang, X.; Chen, G.; Yin, Y.; Lu, X.; Feng, G.; Yu, R.; Chen, L. The Effects of Porphyromonas gingivalis on Atherosclerosis-Related Cells. Front. Immunol. 2021, 12, 766560. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Ma, X.; Yang, B.; Liu, Y.; Xie, Y.; Wang, X.; Yuan, W.; Ma, J. Periodontitis pathogen Porphyromonas gingivalis promotes pancreatic tumorigenesis via neutrophil elastase from tumor-associated neutrophils. Gut Microbes 2022, 14, 2073785. [Google Scholar] [CrossRef] [PubMed]
- Saba, E.; Farhat, M.; Daoud, A.; Khashan, A.; Forkush, E.; Menahem, N.H.; Makkawi, H.; Pandi, K.; Angabo, S.; Kawasaki, H.; et al. Oral bacteria accelerate pancreatic cancer development in mice. Gut 2024, 73, 770–786. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Wu, F.; Kwak, S.; Wang, C.; Usyk, M.; Freedman, N.D.; Huang, W.; Um, C.Y.; Gonda, T.A.; Oberstein, P.E.; et al. Oral bacterial and fungal microbiome and subsequent risk for pancreatic cancer. JAMA Oncol. 2025, 11, 1331–1340. [Google Scholar] [CrossRef]
- Zilberstein, N.F.; Engen, P.A.; Swanson, G.R.; Naqib, A.; Post, Z.; Alutto, J.; Green, S.J.; Shaikh, M.; Lawrence, K.; Adnan, D.; et al. The bidirectional effects of periodontal disease and oral dysbiosis on gut inflammation in inflammatory bowel disease. J. Crohn’s Colitis 2025, 19, e162. [Google Scholar] [CrossRef] [PubMed]
- Mcneill, K.; Hamilton, I.R. Acid tolerance response of biofilm cells of Streptococcus mutans. FEMS Microbiol. Lett. 2003, 221, 25–30. [Google Scholar] [CrossRef]
- Sato, K.; Takahashi, N.; Kato, T.; Matsuda, Y.; Yokoji, M.; Yamada, M.; Nakajima, T.; Kondo, N.; Endo, N.; Yamamoto, R.; et al. Aggravation of collagen-induced arthritis by orally administered Porphyromonas gingivalis through modulation of the gut microbiota and gut immune system. Sci. Rep. 2017, 7, 6955. [Google Scholar] [CrossRef]
- Rojas-Tapias, D.F.; Brown, E.M.; Temple, E.R.; Onyekaba, M.A.; Mohamed, A.M.T.; Duncan, K.; Schirmer, M.; Walker, R.L.; Mayassi, T.; Pierce, K.A.; et al. Inflammation-associated nitrate facilitates ectopic colonization of oral bacterium Veillonella parvula in the intestine. Nat. Microbiol. 2022, 7, 1673–1685. [Google Scholar] [CrossRef]
- Kobayashi, R.; Ogawa, Y.; Hashizume-Takizawa, T.; Kurita-Ochiai, T. Oral bacteria affect the gut microbiome and intestinal immunity. Pathog. Dis. 2020, 78, a24. [Google Scholar] [CrossRef]
- Coccia, M.; Harrison, O.J.; Schiering, C.; Asquith, M.J.; Becher, B.; Powrie, F.; Maloy, K.J. IL-1β mediates chronic intestinal inflammation by promoting the accumulation of IL-17A secreting innate lymphoid cells and CD4(+) Th17 cells. J. Exp. Med. 2012, 209, 1595–1609. [Google Scholar] [CrossRef] [PubMed]
- Morton, A.M.; Sefik, E.; Upadhyay, R.; Weissleder, R.; Benoist, C.; Mathis, D. Endoscopic photoconversion reveals unexpectedly broad leukocyte trafficking to and from the gut. Proc. Natl. Acad. Sci. USA 2014, 111, 6696–6701. [Google Scholar] [CrossRef]
- Kitamoto, S.; Kamada, N. Periodontal connection with intestinal inflammation: Microbiological and immunological mechanisms. Periodontology 2000 2022, 89, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Harbour, S.N.; Maynard, C.L.; Zindl, C.L.; Schoeb, T.R.; Weaver, C.T. Th17 cells give rise to Th1 cells that are required for the pathogenesis of colitis. Proc. Natl. Acad. Sci. USA 2015, 112, 7061–7066. [Google Scholar] [CrossRef]
- Ahern, P.P.; Schiering, C.; Buonocore, S.; Mcgeachy, M.J.; Cua, D.J.; Maloy, K.J.; Powrie, F. Interleukin-23 drives intestinal inflammation through direct activity on T cells. Immunity 2010, 33, 279–288. [Google Scholar] [CrossRef]
- Kitamoto, S.; Nagao-Kitamoto, H.; Jiao, Y.; Gillilland, M.G.R.; Hayashi, A.; Imai, J.; Sugihara, K.; Miyoshi, M.; Brazil, J.C.; Kuffa, P.; et al. The intermucosal connection between the mouth and gut in commensal Pathobiont-Driven colitis. Cell 2020, 182, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.A.; Clay, S.L.; Lavoie, S.L.; Bae, S.; Lang, J.K.; Fonseca-Pereira, D.; Rosinski, K.G.; Ou, N.; Glickman, J.N.; Garrett, W.S. Fusobacterium nucleatum drives a pro-inflammatory intestinal microenvironment through metabolite receptor-dependent modulation of IL-17 expression. Gut Microbes 2021, 13, 1987780. [Google Scholar] [CrossRef] [PubMed]
- Nagao, J.; Kishikawa, S.; Tanaka, H.; Toyonaga, K.; Narita, Y.; Negoro-Yasumatsu, K.; Tasaki, S.; Arita-Morioka, K.; Nakayama, J.; Tanaka, Y. Pathobiont-responsive Th17 cells in gut-mouth axis provoke inflammatory oral disease and are modulated by intestinal microbiome. Cell Rep. 2022, 40, 111314. [Google Scholar] [CrossRef]
- Ashizawa, H.; Iwanaga, N.; Nemoto, K.; Hirayama, T.; Yoshida, M.; Takeda, K.; Ide, S.; Tashiro, M.; Hosogaya, N.; Takazono, T.; et al. Prevotella intermedia Synergistically Exacerbates Pneumonia Induced by Oral Streptococci. J. Infect. Dis. 2025, 232, e280–e289. [Google Scholar] [CrossRef]
- Colombo, A.V.; Barbosa, G.M.; Higashi, D.; di Micheli, G.; Rodrigues, P.H.; Simionato, M.R.L. Quantitative detection of Staphylococcus aureus, Enterococcus faecalis and Pseudomonas aeruginosa in human oral epithelial cells from subjects with periodontitis and periodontal health. J. Med. Microbiol. 2013, 62, 1592–1600. [Google Scholar] [CrossRef]
- Gibbons, R.J. Bacterial adhesion to oral tissues: A model for infectious diseases. J. Dent. Res. 1989, 68, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, R.J.; Hay, D.I.; Childs, W.C.R.; Davis, G. Role of cryptic receptors (cryptitopes) in bacterial adhesion to oral surfaces. Arch. Oral Biol. 1990, 35, 107S–114S. [Google Scholar] [CrossRef]
- Arimizu, C.; Akahoshi, T.; Jinno, T.; Furuta, M.; Ohashi, A.; Takamori, S.; Wada, N. Association of number of oral bacteria with Ventilator-Associated pneumonia and delirium in patients in the intensive care unit. J. Intensive Care Med. 2025, 40, 779–788. [Google Scholar] [CrossRef]
- Dale, C.M.; Angus, J.E.; Sutherland, S.; Dev, S.; Rose, L. Exploration of difficulty accessing the mouths of intubated and mechanically ventilated adults for oral care: A video and photographic elicitation study. J. Clin. Nurs. 2020, 29, 1920–1932. [Google Scholar] [CrossRef]
- Aktan, G.G.; Coşkun Palaz, S. Oral care practices, attitudes and challenges faced by intensive care unit nurses in the care of intubated patients: A descriptive study. Nurs. Crit. Care 2025, 30, e70095. [Google Scholar] [CrossRef]
- Alqaissi, N.; Qtait, M. Knowledge, attitudes, and practices of intensive care unit nurses regarding oral care for intubated patients in hebron hospitals, palestine. SAGE Open Nurs. 2025, 11, 2126021225. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, A.A.; Silva, L.D.D.A.; Santos, C.C.D.O.; Fonseca-Silva, T. Oral care practices for patients in intensive care unit: A systematic review. Int. J. Dent. Hyg. 2025, 23, 80–88. [Google Scholar] [CrossRef]
- Tuon, F.F.; Gavrilko, O.; Almeida, S.D.; Sumi, E.R.; Alberto, T.; Rocha, J.L.; Rosa, E.A. Prospective, randomised, controlled study evaluating early modification of oral microbiota following admission to the intensive care unit and oral hygiene with chlorhexidine. J. Glob. Antimicrob. Resist. 2017, 8, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Sands, K.M.; Wilson, M.J.; Lewis, M.A.O.; Wise, M.P.; Palmer, N.; Hayes, A.J.; Barnes, R.A.; Williams, D.W. Respiratory pathogen colonization of dental plaque, the lower airways, and endotracheal tube biofilms during mechanical ventilation. J. Crit. Care 2017, 37, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Bahrani-Mougeot, F.K.; Paster, B.J.; Coleman, S.; Barbuto, S.; Brennan, M.T.; Noll, J.; Kennedy, T.; Fox, P.C.; Lockhart, P.B. Molecular analysis of oral and respiratory bacterial species associated with ventilator-associated pneumonia. J. Clin. Microbiol. 2007, 45, 1588–1593. [Google Scholar] [CrossRef]
- Talbert, S.; Bourgault, A.M.; Rathbun, K.P.; Abomoelak, B.; Deb, C.; Mehta, D.; Sole, M.L. Pepsin a in tracheal secretions from patients receiving mechanical ventilation. Am. J. Crit. Care: Off. Publ. Am. Assoc. Crit.-Care Nurses 2021, 30, 443–450. [Google Scholar] [CrossRef]
- Jaillette, E.; Girault, C.; Brunin, G.; Zerimech, F.; Behal, H.; Chiche, A.; Broucqsault-Dedrie, C.; Fayolle, C.; Minacori, F.; Alves, I.; et al. Impact of tapered-cuff tracheal tube on microaspiration of gastric contents in intubated critically ill patients: A multicenter cluster-randomized cross-over controlled trial. Intensive Care Med. 2017, 43, 1562–1571. [Google Scholar] [CrossRef]
- Arimizu, C.; Akahoshi, T.; Jinno, T.; Furuta, M.; Ohashi, A.; Takamori, S.; Wada, N. Association of number of oral bacteria with Ventilator-Associated pneumonia and delirium in patients in the intensive care unit. J. Intensive Care Med. 2025, 40, 779–788. [Google Scholar] [CrossRef]
- Sumi, Y.; Miura, H.; Michiwaki, Y.; Nagaosa, S.; Nagaya, M. Colonization of dental plaque by respiratory pathogens in dependent elderly. Arch. Gerontol. Geriatr. 2007, 44, 119–124. [Google Scholar] [CrossRef]
- Tada, A.; Hanada, N. Opportunistic respiratory pathogens in the oral cavity of the elderly. FEMS Immunol. Med. Microbiol. 2010, 60, 1–17. [Google Scholar] [CrossRef]
- Heo, S.; Haase, E.M.; Lesse, A.J.; Gill, S.R.; Scannapieco, F.A. Genetic relationships between respiratory pathogens isolated from dental plaque and bronchoalveolar lavage fluid from patients in the intensive care unit undergoing mechanical ventilation. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2008, 47, 1562–1570. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Hu, W.; He, J.; Guo, L.; Lux, R.; Shi, W. Community-based interference against integration of Pseudomonas aeruginosa into human salivary microbial biofilm. Mol. Oral Microbiol. 2011, 26, 337–352. [Google Scholar] [CrossRef]
- Charles, M.P.; Easow, J.M.; Joseph, N.M.; Ravishankar, M.; Kumar, S.; Sivaraman, U. Aetiological agents of ventilator-associated pneumonia and its resistance pattern—A threat for treatment. Australas. Med. J. 2013, 6, 430–434. [Google Scholar] [CrossRef]
- Kawamura, S.; Goto, H.; Kikuchi, T.; Okabe, T.; Hasegawa, Y.; Sugita, Y.; Fujitsuka, H.; Kataoka, R.; Katsumata, K.; Goto, R.; et al. IL-35 may prevent the exacerbation of aspiration pneumonia involving porphyromonas gingivalis by suppressing IL-17 production. Am. J. Pathol. 2025, 195, 652–662. [Google Scholar] [CrossRef]
- Bassim, C.W.; Gibson, G.; Ward, T.; Paphides, B.M.; Denucci, D.J. Modification of the risk of mortality from pneumonia with oral hygiene care. J. Am. Geriatr. Soc. 2008, 56, 1601–1607. [Google Scholar] [CrossRef]
- Yoneyama, T.; Yoshida, M.; Matsui, T.; Sasaki, H. Oral care and pneumonia. Oral Care Working Group. Lancet 1999, 354, 515. [Google Scholar] [CrossRef]
- Klompas, M.; Speck, K.; Howell, M.D.; Greene, L.R.; Berenholtz, S.M. Reappraisal of routine oral care with chlorhexidine gluconate for patients receiving mechanical ventilation: Systematic review and meta-analysis. JAMA Intern. Med. 2014, 174, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Wu, X.; Zhang, Q.; Li, C.; Worthington, H.V.; Hua, F. Oral hygiene care for critically ill patients to prevent ventilator-associated pneumonia. Cochrane Database Syst. Rev. 2020, 12, D8367. [Google Scholar]
- Price, R.; Maclennan, G.; Glen, J.; Suddicu, C. Selective digestive or oropharyngeal decontamination and topical oropharyngeal chlorhexidine for prevention of death in general intensive care: Systematic review and network meta-analysis. BMJ (Clin. Res. Ed.) 2014, 348, g2197. [Google Scholar] [CrossRef]
- Deschepper, M.; Waegeman, W.; Eeckloo, K.; Vogelaers, D.; Blot, S. Effects of chlorhexidine gluconate oral care on hospital mortality: A hospital-wide, observational cohort study. Intensive Care Med. 2018, 44, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Klompas, M.; Li, L.; Kleinman, K.; Szumita, P.M.; Massaro, A.F. Associations between ventilator bundle components and outcomes. JAMA Intern. Med. 2016, 176, 1277–1283. [Google Scholar] [CrossRef]
- Sozkes, S.; Sozkes, S. Use of toothbrushing in conjunction with chlorhexidine for preventing ventilator-associated pneumonia: A random-effect meta-analysis of randomized controlled trials. Int. J. Dent. Hyg. 2023, 21, 389–397. [Google Scholar] [CrossRef]
- Plantinga, N.L.; Wittekamp, B.H.J.; Leleu, K.; Depuydt, P.; Van den Abeele, A.; Brun-Buisson, C.; Bonten, M.J.M. Oral mucosal adverse events with chlorhexidine 2% mouthwash in ICU. Intensive Care Med. 2016, 42, 620–621. [Google Scholar] [CrossRef] [PubMed]
- Cieplik, F.; Jakubovics, N.S.; Buchalla, W.; Maisch, T.; Hellwig, E.; Al-Ahmad, A. Resistance toward chlorhexidine in oral bacteria—Is there cause for concern? Front. Microbiol. 2019, 10, 587. [Google Scholar]
- Kampf, G. Acquired resistance to chlorhexidine—Is it time to establish an ‘antiseptic stewardship’ initiative? J. Hosp. Infect. 2016, 94, 213–227. [Google Scholar] [CrossRef]
- Dale, C.M.; Rose, L.; Carbone, S.; Pinto, R.; Smith, O.M.; Burry, L.; Fan, E.; Amaral, A.C.K.; Mccredie, V.A.; Scales, D.C.; et al. Effect of oral chlorhexidine de-adoption and implementation of an oral care bundle on mortality for mechanically ventilated patients in the intensive care unit (CHORAL): A multi-center stepped wedge cluster-randomized controlled trial. Intensive Care Med. 2021, 47, 1295–1302. [Google Scholar] [CrossRef]
- Ehrenzeller, S.; Klompas, M. Association between daily toothbrushing and Hospital-Acquired pneumonia: A systematic review and Meta-Analysis. JAMA Intern. Med. 2024, 184, 131–142. [Google Scholar] [CrossRef]
- Klompas, M.; Branson, R.; Cawcutt, K.; Crist, M.; Eichenwald, E.C.; Greene, L.R.; Lee, G.; Maragakis, L.L.; Powell, K.; Priebe, G.P.; et al. Strategies to prevent ventilator-associated pneumonia, ventilator-associated events, and nonventilator hospital-acquired pneumonia in acute-care hospitals: 2022 Update. Infect. Control. Hosp. Epidemiol. 2022, 43, 687–713. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Peng, Z.; He, C.; Zhang, C.; Hu, R. Effect of different mouthwashes on ventilator-related outcomes and mortality in intensive care unit patients: A network meta-analysis. Aust. Crit. Care Off. J. Confed. Aust. Crit. Care Nurses 2025, 38, 101095. [Google Scholar] [CrossRef]
- Rathbun, K.P.; Bourgault, A.M.; Sole, M.L. Oral microbes in Hospital-Acquired pneumonia: Practice and research implications. Crit. Care Nurse 2022, 42, 47–54. [Google Scholar] [CrossRef]
- Maciel, J.G.; Gomes, A.C.G.; Sugio, C.Y.; Garcia, A.A.; Zani, I.F.; Fernandes, M.H.; Soares, S.; Neppelenbroek, K.H. Denture biofilm increases respiratory diseases in the elderly. A mini-review. Am. J. Dent. 2024, 37, 288–292. [Google Scholar]
- Oostdijk, E.A.N.; Kesecioglu, J.; Schultz, M.J.; Visser, C.E.; de Jonge, E.; van Essen, E.H.R.; Bernards, A.T.; Purmer, I.; Brimicombe, R.; Bergmans, D.; et al. Effects of decontamination of the oropharynx and intestinal tract on antibiotic resistance in ICUs: A randomized clinical trial. JAMA 2014, 312, 1429–1437. [Google Scholar] [CrossRef]
- Suddicu, I.F.T.A.; Cuthbertson, B.H.; Billot, L.; Campbell, M.K.; Daneman, N.; Davis, J.S.; Delaney, A.; Devaux, A.; Ferguson, N.D.; Finfer, S.R.; et al. Selective decontamination of the digestive tract during ventilation in the ICU. N. Engl. J. Med. 2025. ahead of print. [Google Scholar]
- Wittekamp, B.H.; Plantinga, N.L.; Cooper, B.S.; Lopez-Contreras, J.; Coll, P.; Mancebo, J.; Wise, M.P.; Morgan, M.P.G.; Depuydt, P.; Boelens, J.; et al. Decontamination strategies and bloodstream infections with Antibiotic-Resistant microorganisms in ventilated patients: A randomized clinical trial. JAMA 2018, 320, 2087–2098. [Google Scholar] [CrossRef] [PubMed]
- Plantinga, N.L.; Wittekamp, B.H.J.; Brun-Buisson, C.; Bonten, M.J.M. R-GNOSIS ICU Study Group The effects of topical antibiotics on eradication and acquisition of third-generation cephalosporin and carbapenem-resistant Gram-negative bacteria in ICU patients; A post hoc analysis from a multicentre cluster-randomized trial. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2020, 26, 485–491. [Google Scholar]
- Wang, B.; Briegel, J.; Krueger, W.A.; Draenert, R.; Jung, J.; Weber, A.; Bogner, J.; Schubert, S.; Liebchen, U.; Frank, S.; et al. Ecological effects of selective oral decontamination on multidrug-resistance bacteria acquired in the intensive care unit: A case-control study over 5 years. Intensive Care Med. 2022, 48, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Disruption of the oral-gut microbiome axis by common ICU interventions and its systemic consequences. Schematic illustration showing how mechanical ventilation, broad-spectrum antibiotics, and proton pump inhibitors (PPIs) disrupt microbial homeostasis and compromise epithelial barrier integrity. Endotracheal intubation impairs salivary flow and oral self-cleaning mechanisms, promoting oral dysbiosis and overgrowth of pathogens. PPIs reduce gastric acidity, weakening a key chemical defense and enabling the survival and translocation of acid-sensitive oral bacteria to the gastrointestinal tract. Antibiotics deplete commensal gut microbiota, diminishing colonization resistance and creating an ecological niche that translocated oral pathobionts can occupy. This sequence of events drives intestinal dysbiosis, triggers mucosal inflammation, and increases intestinal permeability.
Figure 1.
Disruption of the oral-gut microbiome axis by common ICU interventions and its systemic consequences. Schematic illustration showing how mechanical ventilation, broad-spectrum antibiotics, and proton pump inhibitors (PPIs) disrupt microbial homeostasis and compromise epithelial barrier integrity. Endotracheal intubation impairs salivary flow and oral self-cleaning mechanisms, promoting oral dysbiosis and overgrowth of pathogens. PPIs reduce gastric acidity, weakening a key chemical defense and enabling the survival and translocation of acid-sensitive oral bacteria to the gastrointestinal tract. Antibiotics deplete commensal gut microbiota, diminishing colonization resistance and creating an ecological niche that translocated oral pathobionts can occupy. This sequence of events drives intestinal dysbiosis, triggers mucosal inflammation, and increases intestinal permeability.
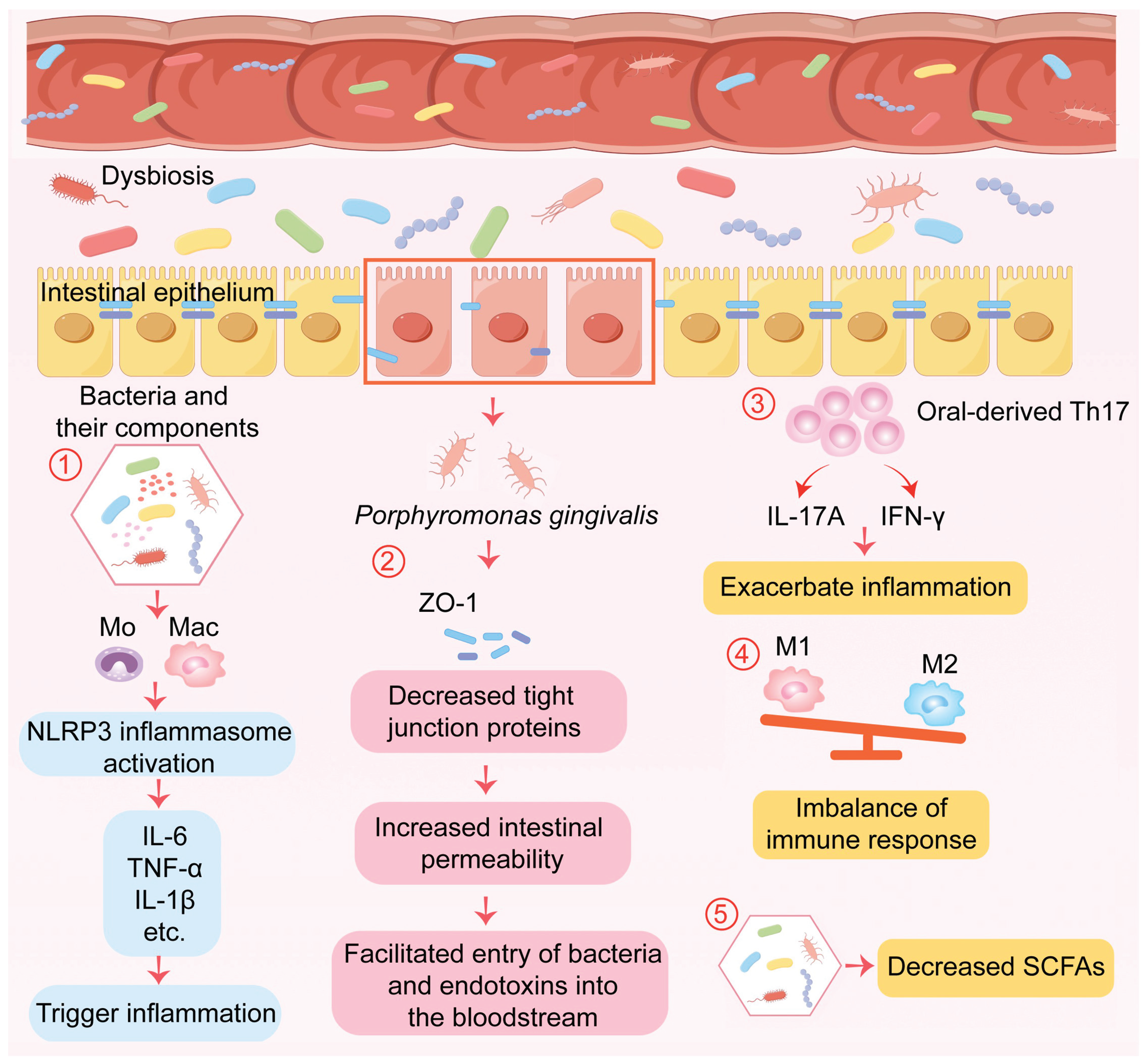
Figure 2.
Molecular mechanisms of gut colonization and inflammation driven by oral pathobionts. Detailed illustration of the mechanisms through which specific oral-derived bacteria induce gut pathology. 1) Certain oral bacteria and their microbial components can activate the NOD-, LRR-, and pyrin domain-containing protein 3 (NLRP3) inflammasome in mononuclear phagocytes, triggering the release of pro-inflammatory cytokines such as IL-6, IL-1β, and TNF-α; 2) Porphyromonas gingivalis compromises intestinal epithelial barrier integrity by downregulating tight junction proteins, including ZO-1 and occludin; 3) Oral-derived pro-inflammatory T-helper 17 (Th17) cells expand in the intestinal mucosa and secrete IL-17 and IFN-γ, contributing to local inflammation; 4) Oral infection promotes a shift in intestinal macrophage polarization toward the pro-inflammatory M1 phenotype, while suppressing the anti-inflammatory M2 phenotype, thereby amplifying inflammatory responses; 5) Additionally, it alters the metabolic profile of the gut microbiota, particularly the levels of short-chain fatty acids (SCFAs), which disrupts immunological homeostasis in the gut.
Figure 2.
Molecular mechanisms of gut colonization and inflammation driven by oral pathobionts. Detailed illustration of the mechanisms through which specific oral-derived bacteria induce gut pathology. 1) Certain oral bacteria and their microbial components can activate the NOD-, LRR-, and pyrin domain-containing protein 3 (NLRP3) inflammasome in mononuclear phagocytes, triggering the release of pro-inflammatory cytokines such as IL-6, IL-1β, and TNF-α; 2) Porphyromonas gingivalis compromises intestinal epithelial barrier integrity by downregulating tight junction proteins, including ZO-1 and occludin; 3) Oral-derived pro-inflammatory T-helper 17 (Th17) cells expand in the intestinal mucosa and secrete IL-17 and IFN-γ, contributing to local inflammation; 4) Oral infection promotes a shift in intestinal macrophage polarization toward the pro-inflammatory M1 phenotype, while suppressing the anti-inflammatory M2 phenotype, thereby amplifying inflammatory responses; 5) Additionally, it alters the metabolic profile of the gut microbiota, particularly the levels of short-chain fatty acids (SCFAs), which disrupts immunological homeostasis in the gut.
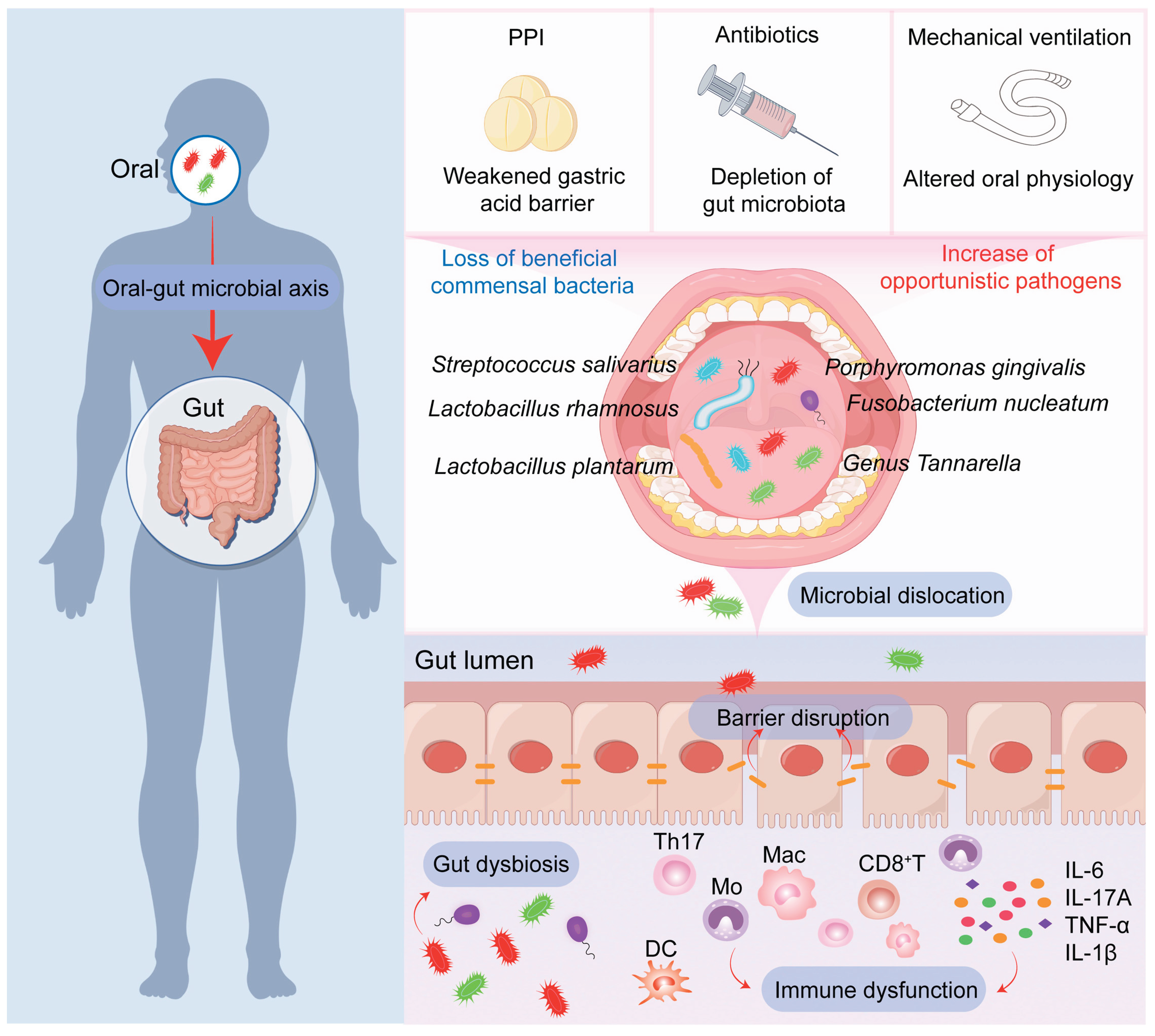
Figure 3.
Evolving strategies for oral care in the ICU: from chemical antiseptics to ecological management. Comparison of oral care paradigms. The traditional approach, centered on chlorhexidine, is discouraged due to risks of mucosal injury, microbial resistance, ecological disruption, and potential associations with increased mortality. The modern, evidence-based paradigm emphasizes a multifaceted strategy: 1) Mechanical debridement through toothbrushing as the cornerstone for physically disrupting dental biofilms; 2) Use of physiological solutions such as normal saline for irrigation, which is safe and preserves the integrity of the microbial ecology; and 3) Selective Oropharyngeal Decontamination (SOD) for high-risk patients, employing a topical, non-absorbable antibiotic paste to target specific pathogens without promoting widespread antimicrobial resistance. This integrated approach aims to reduce the oral bioburden while supporting microbial resilience.
Figure 3.
Evolving strategies for oral care in the ICU: from chemical antiseptics to ecological management. Comparison of oral care paradigms. The traditional approach, centered on chlorhexidine, is discouraged due to risks of mucosal injury, microbial resistance, ecological disruption, and potential associations with increased mortality. The modern, evidence-based paradigm emphasizes a multifaceted strategy: 1) Mechanical debridement through toothbrushing as the cornerstone for physically disrupting dental biofilms; 2) Use of physiological solutions such as normal saline for irrigation, which is safe and preserves the integrity of the microbial ecology; and 3) Selective Oropharyngeal Decontamination (SOD) for high-risk patients, employing a topical, non-absorbable antibiotic paste to target specific pathogens without promoting widespread antimicrobial resistance. This integrated approach aims to reduce the oral bioburden while supporting microbial resilience.
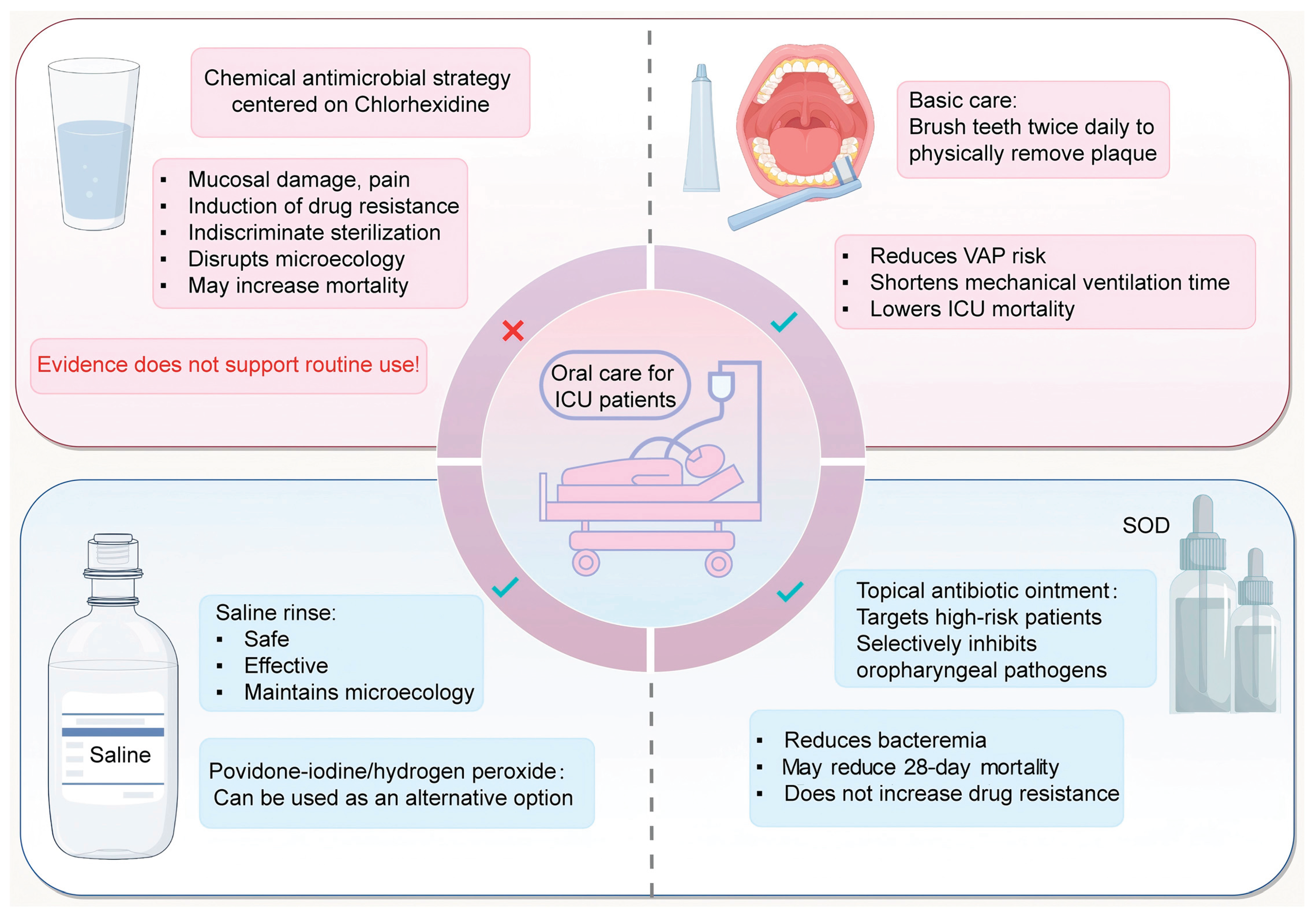
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



