
Submitted:
21 January 2026
Posted:
21 January 2026
You are already at the latest version
Abstract
Wastewater-integrated microalgal cultivation offers a sustainable pathway to reduce biofuel production costs while simultaneously addressing nutrient-rich effluent management. In this study, matured compost leachate was systematically evaluated as a sole cultivation medium for Desmodesmus sp. under different dilution regimes, with emphasis on growth kinetics, wastewater remediation efficiency, lipid accumulation behavior, and biodiesel quality. Desmodesmus sp. successfully acclimatized to 100% undiluted matured compost leachate within four days and maintained stable mixotrophic growth without dilution or pre-treatment. Cultivation in undiluted leachate achieved a maximum biomass concentration of 2.69 ± 0.09 g L⁻¹, representing an approximately fourfold increase compared to Bold’s Basal Medium. Concurrently, high treatment efficiencies were obtained, with chemical oxygen demand removal of 82.6%, total nitrogen reduction of 60–72%, and total phosphorus removal of 65–66%, confirming effective integration of biomass production with wastewater remediation. Lipid biosynthesis was strongly governed by nitrogen availability, with lipid concentration increasing from 0.32 g L⁻¹ during exponential growth to 0.72 g L⁻¹ under nitrogen-depleted stationary conditions. Fatty acid methyl ester profiling revealed a stress-induced shift toward saturated and monounsaturated fatty acids, accounting for 75.6% of total fatty acids and dominated by palmitic acid (C16:0). This compositional restructuring resulted in biodiesel properties characterized by a high cetane number of 64.5, low iodine value, and oxidative stability exceeding 30 h, meeting or surpassing international biodiesel quality benchmarks.
Keywords:
Desmodesmus sp.
; matured compost leachate
; mixotrophic cultivation
; biodiesel production
; circular bioeconomy
1. Introduction
The rapid growth of the global population, accelerating urbanization, and expanding industrial activity are exerting unprecedented pressure on global energy systems, freshwater resources, and wastewater management infrastructures. Global population projections indicate an increase to approximately 9.7 billion by 2050, accompanied by an estimated 50% rise in global energy demand (Li & Li, 2023; Dodangodage et al., 2025; Holechek et al., 2022). Despite the increasing deployment of renewable energy technologies, fossil fuels continue to dominate the global energy mix, accounting for nearly 80% of primary energy consumption. This persistent dependence has resulted in severe environmental consequences, with global carbon dioxide (CO₂) emissions exceeding 35.8 Gt in 2023, thereby intensifying climate change, air pollution, and ecosystem degradation (Liu et al., 2024). Among these impacts, fossil fuel derived transportation fuels remain a major contributor to anthropogenic greenhouse gas emissions, underscoring the urgent need for sustainable, low-carbon liquid fuel alternatives (Shahzad & Iqbal Cheema, 2024).
Within this context, biodiesel has emerged as a promising renewable fuel due to its biodegradability, compatibility with existing diesel engines, and potential to reduce net greenhouse gas emissions. Global biodiesel consumption increased from approximately 2.2 million metric tons (MMT) in 2004 to 65.86 MMT in 2023 and is projected to reach nearly 75 MMT by 2030 (Awogbemi & Desai, 2025). However, the long-term sustainability of biodiesel production remains constrained by feedstock availability and cost. First-generation biodiesel derived from edible oils raises critical concerns related to food security, land-use change, and competition for agricultural resources, thereby motivating the development of non-food, third-generation biofuel pathways that decouple fuel production from arable land and conventional food systems (Rulli et al., 2016; Mhetras & Gokhale, 2025; W. Wang & Khanna, 2023).
Microalgae are widely regarded as one of the most promising third-generation biodiesel feedstocks owing to their rapid growth rates, high photosynthetic efficiencies, and superior lipid productivity compared to terrestrial oil crops (Casanova et al., 2023; Adewuyi, 2022; Behera et al., 2015; S. Zhang et al., 2022; Dodangodage et al., 2025; Kanwal et al., 2025). Under nutrient-replete conditions, most microalgal species accumulate approximately 10–30% lipids (w/w), while nutrient stress, particularly nitrogen limitation, can enhance lipid accumulation by two- to three-fold (Maltsev et al., 2023; Udayan et al., 2023; Adams et al., 2013). Several strains, including Desmodesmus, Chlorella, and Chlorococcum, have been reported to achieve lipid contents exceeding 45–60%. In addition to high lipid yields, microalgal biodiesel exhibits favorable physicochemical properties, such as a high cetane number, negligible sulfur content, and improved combustion performance, making it a technically attractive substitute for petroleum diesel (Mallick et al., 2012; Mathimani et al., 2023; Azarpour et al., 2022; Kumar et al., 2023).
Despite these advantages, the commercial deployment of microalgal biodiesel remains economically challenging. The reliance on synthetic growth media, such as BG-11 and Bold’s Basal Medium, substantially increases production costs, with nutrient inputs alone accounting for approximately 30–50% of total cultivation expenses (Gu et al., 2023; Casanova et al., 2023; Kanwal et al., 2025; Maroušek et al., 2023). Techno-economic analyses consistently indicate that microalgal biodiesel production costs remain significantly higher than those of petroleum diesel, primarily due to elevated capital investment and operational expenditures. Consequently, the identification of low-cost, nutrient-rich alternative culture media has become a central research focus for improving the economic feasibility and scalability of microalgal biodiesel systems (Maroušek et al., 2023; Rafa et al., 2021; Patnaik & Mallick, 2021).
In parallel, inadequate wastewater management continues to pose a major global sustainability challenge. Approximately 42–45% of household wastewater worldwide is discharged without safe treatment, while much of the remaining fraction receives only partial treatment, resulting in nutrient pollution, eutrophication, and widespread degradation of aquatic ecosystems (Staff, 2025; UN-Water, 2025). High-strength wastewaters enriched with nitrogen, phosphorus, and biodegradable organic matter therefore represent both an environmental burden and an underutilized resource. From a circular bioeconomy perspective, such waste streams offer substantial opportunities for nutrient recovery and bioresource generation when integrated with biological treatment processes (Dodangodage, Kasturiarachchi, et al., 2025b).
Among these waste streams, matured compost leachate wastewater, generated during the later stages of organic waste composting following prolonged biological stabilization, has received comparatively limited research attention. Unlike fresh compost leachate, which is often characterized by extreme organic loading, high ammoniacal toxicity, and inhibitory effects on microbial growth, matured compost leachate exhibits a more stable chemical composition with reduced toxicity while retaining considerable concentrations of bioavailable nitrogen and phosphorus. These characteristics render matured compost leachate particularly suitable for biological treatment and valorization strategies, including microalgal cultivation (Naveen et al., 2015; Ahamad Sanadi et al., 2021).
The integration of microalgal cultivation with wastewater treatment represents a compelling circular bioeconomy pathway by enabling simultaneous nutrient removal, biomass production, and renewable fuel generation. Wastewater-based microalgal systems have demonstrated effective reductions in chemical oxygen demand (COD), total nitrogen (TN), and total phosphorus (TP) while producing lipid-rich biomass suitable for biodiesel conversion (Kadir et al., 2018; El-Sheekh et al., 2023). However, existing studies on compost-derived wastewaters have predominantly focused on treatment efficiency or biomass production in isolation, with limited emphasis on fuel-oriented lipid quality, fatty acid composition, and biodiesel property estimation. Moreover, systematic evaluations of microalgal performance under the elevated nitrogen and organic loading conditions characteristic of matured compost leachate remain scarce.
In this context, strain selection must prioritize not only high lipid accumulation but also tolerance to nutrient-rich and organically loaded environments. Desmodesmus sp. was selected in this study due to its demonstrated adaptability to wastewater conditions, resilience to high nutrient concentrations, and capacity for lipid accumulation. Previous investigations have reported biomass productivities ranging from 0.2 to 1.8 g L⁻¹ for Desmodesmus spp., alongside removals of 70–99% TN, 60–95% TP, and 40–80% COD, with lipid contents between 20 and 45% (w/w) and fatty acid profiles suitable for biodiesel production (Ji et al., 2014a; Zheng et al., 2023; Ogbonna et al., 2025; Mandotra et al., 2016; C. Jia et al., 2025; Ji et al., 2014b).
Accordingly, this study investigates the sustainable valorization of matured compost leachate wastewater for microalgal biodiesel production using Desmodesmus sp. Optimal leachate dilution ratios were evaluated by simultaneously assessing growth kinetics, biomass productivity, and nutrient uptake performance, with COD, TN, and TP removal treated as primary functional outcomes. Harvested biomass was subsequently analyzed for total lipid content, and fatty acid profiles were determined through fatty acid methyl ester (FAME) analysis. Based on FAME composition, key biodiesel properties, including cetane number, iodine value, saponification value, and higher heating value, were estimated to evaluate fuel suitability in accordance with international biodiesel standards.
To the best of the authors’ knowledge, this is the first study to simultaneously assess biomass productivity, nutrient removal dynamics, fatty acid composition, and biodiesel fuel property estimation for Desmodesmus sp. cultivated in matured compost leachate wastewater. The findings demonstrate an integrated circular bioeconomy approach aligned with the United Nations Sustainable Development Goals SDG 6, SDG 7, and SDG 12 (Sayago et al., 2024; Kanchanamala Delanka-Pedige et al., 2021; Dodangodage, Kasturiarachchi, et al., 2025a; Rajendran et al., 2024).
2. Method
2.1. Wastewater Collection and Pre-Treatment
Matured compost leachate wastewater was collected from the Karadiyana municipal solid waste dumping facility, South Colombo, Sri Lanka. Samples were collected in sterile polyethylene containers, transported to the laboratory under ice-cooled conditions, and processed within 24 h of collection. To remove suspended solids and particulate matter, the leachate was filtered through a 0.45 µm nylon membrane filter (Millipore). The clarified filtrate was subsequently used as the experimental growth medium.
Preliminary screening experiments were conducted to identify the optimal leachate dilution ratio for the cultivation of Desmodesmus sp. Based on observed growth performance, undiluted leachate (100% v/v) was selected for subsequent experiments. The initial pH of the wastewater (less than 3.3) was adjusted to 6.8 using 1 M NaOH to ensure physiological suitability for microalgal growth. Bold’s Basal Medium (BBM) was employed as the control medium. All culture media were sterilized by autoclaving at 121 °C for 20 min prior to inoculation.
2.2. Wastewater Characterization
Initial physicochemical characterization of the matured compost leachate was conducted in accordance with APHA standard methods. All measurements were performed in triplicate (n = 3). The following parameters were analyzed (Association, 1926):
pH: measured using a calibrated benchtop pH meter
Nitrate (NO₃⁻–N): determined by UV absorbance at 220 nm with correction at 275 nm
Phosphate (PO₄³⁻–P): quantified using the molybdenum blue method at 880 nm
COD: Determined by the open reflux titrimetric method with potassium dichromate digestion.
Measured nutrient concentrations were compared with Sri Lankan national discharge standards (Central Environmental Authority, 2022).
2.3. Microalgal Strain and Inoculum Preparation
A pure culture of Desmodesmus sp. was obtained from the Pro Green Laboratory, University of Moratuwa, Sri Lanka. Pre-cultures were maintained in BBM containing standard nitrate concentrations under controlled laboratory conditions (25 ± 2 °C, continuous illumination at 150 μmol photons m⁻² s⁻¹ using cool-white LED lighting, and aeration at 0.5 vvm with sterile-filtered air).
Cells were harvested during the exponential growth phase by centrifugation at 4000 rpm for 20 min and subsequently inoculated into experimental media at an initial biomass concentration of 0.30 g L⁻¹. Cultivation experiments were conducted in 2 L laboratory-scale glass photobioreactors with a working volume of 1.5 L. Each reactor was equipped with three-port GL45 screw caps to facilitate aeration, sampling, and pressure compensation. Aeration ports were fitted with 0.45 µm polytetrafluoroethylene (PTFE) membrane filters to ensure sterility and minimize evaporative losses. Illumination was provided using cool-white LED strips under a 12:12 h light–dark photoperiod (Naseema Rasheed et al., 2023; Mattos et al., 2012; Nicodemou et al., 2024).
2.4. Experimental Setup and Cultivation Conditions
2.4.1. Screening of Wastewater Dilution Factors
Screening experiments were conducted using four dilution ratios of matured compost leachate wastewater (25%, 50%, 75%, and 100% v/v), prepared using distilled water. Cultures were grown in 500 mL Erlenmeyer flasks containing a working volume of 400 mL. BBM (400 mL) served as the control medium. The initial pH of all treatments was adjusted to 6.8 using NaOH.
Cultures were incubated at 25 ± 2 °C under a 12:12 h light–dark cycle, continuously aerated, and illuminated at an intensity of 150 μmol photons m⁻² s⁻¹ using cool-white LED lighting. Biomass growth was monitored at 48 h intervals by measuring optical density at 680 nm (OD₆₈₀) and by dry weight determination.
2.4.2. Main Cultivation Experiment
Based on the screening results, undiluted matured compost leachate (100% v/v) was selected for the main cultivation experiment, as it supported high biomass productivity without the need for freshwater dilution. The selected light intensity (150 μmol photons m⁻² s⁻¹) was retained, as it provided an optimal balance between growth performance and energy input.
Batch cultivation was performed in 2 L sterilized glass photobioreactors with a working volume of 1.5 L using undiluted leachate and BBM as the control. Cultures were maintained at 25 ± 2 °C under a 12:12 h light–dark cycle, aerated with sterile-filtered air (BOYO pumps, 0.45 µm filters), and illuminated using cool-white LED lighting. Light intensity was periodically verified using a quantum sensor (LI-COR LI-250A). The BBM control was cultivated under identical conditions to ensure experimental comparability (Dodangodage, Kasturiarachchi, et al., 2025b).
2.5. Analytical Procedures
2.5.1. Biomass Concentration
Microalgal growth was monitored every 48 h. Optical density at 680 nm (OD₆₈₀) was measured using a UV–Vis spectrophotometer (Shimadzu UV-1800). Biomass concentration was determined gravimetrically by filtering 5 mL culture aliquots through pre-dried and pre-weighed glass microfiber filters (Hyundai GF/C, 47 mm diameter, 1.2 µm pore size)(Dodangodage et al., 2025). Filters were oven-dried at 60 °C for 24 h prior to weighing.
Biomass concentration (DWₙ, g L⁻¹) was calculated as:
where is the final weight of the filter with biomass, is the initial filter weight, and is the volume of the filtered culture sample.
2.5.2. Nutrient and COD Removal
Nitrate, phosphate, COD, and pH were measured at 2-day intervals. Culture samples (5 mL) were centrifuged at 10,000 × g for 5 min, and the supernatant was filtered through 0.22 µm nylon syringe filters. Nitrate and phosphate concentrations were analyzed as described in Section 2.2, while pH was measured using a calibrated pH probe (Hanna HI98194)(Dodangodage et al., 2025).
Removal efficiency (RE, %) was calculated as:
where and represent the initial and final concentrations (mg L⁻¹), respectively.
2.5.3. Analytical Validation
All analytical measurements were conducted in triplicate. UV–Vis analyses were calibrated using standard solutions of potassium nitrate, KH₂PO₄, and potassium hydrogen phthalate. Calibration curves demonstrated strong linearity (R² ≥ 0.996). Instrumental blanks were analyzed with each batch to minimize baseline drift.
2.6. Biomass Harvesting
At the end of the 20-day cultivation period, microalgal biomass was harvested by centrifugation at 4000 rpm for 20 min (Eppendorf 5810R). The biomass was washed twice with sterile distilled water and oven-dried at 60 °C until constant weight was achieved (Potts, 2000; Dodangodage, Kasturiarachchi, et al., 2025b).
2.7. Total Lipid Extraction and Quantification
Microalgal biomass was harvested on the 20th day of cultivation by centrifugation at 4500 × g for 10 min, followed by oven drying at 60 °C for 24 h to obtain constant dry weight. Lipid extraction was performed using a modified Bligh and Dyer protocol [27]. Briefly, 10 mg of dried biomass was homogenized with a chloroform–methanol mixture (1:1, v/v) to facilitate lipid solubilization. Phase separation was subsequently induced by adding 0.8% (w/v) NaCl solution, resulting in a final solvent ratio of chloroform/methanol/0.8% NaCl = 10:10:9 (v/v/v).
The lower chloroform phase containing the extracted lipids was carefully transferred into pre-dried and pre-weighed centrifuge tubes and evaporated under a gentle stream of nitrogen gas. Residual solvent was removed by oven drying at 60 °C until constant mass was achieved. The total lipid content was quantified gravimetrically and expressed as a percentage of dry biomass (Lee et al., 2013; DuBois et al., 1956; J. Jia et al., 2015).
2.8. Fatty Acid Methyl Ester (FAME) Preparation and Fatty Acid Profiling
Total lipids extracted as described in Section 2.7 were dissolved in chloroform, and 0.5 mg of heptadecanoic acid (C17:0) was added to each sample as an internal standard prior to solvent evaporation under nitrogen. Fatty acid transesterification was conducted by adding 5 mL of 1% (v/v) sulfuric acid in methanol, followed by incubation in a 50 °C water bath overnight, as previously reported.
The resulting fatty acid methyl esters (FAMEs) were extracted into n-hexane and analyzed using gas chromatography with flame ionization detection (GC–FID) equipped with a DB-23 capillary column (60 m × 0.25 mm × 0.15 μm). The injector temperature was maintained at 250 °C, with an injection volume of 1 μL and a 1:50 split ratio. Helium was used as the carrier gas at a constant pressure of 230 kPa(Breuer et al., 2013)(Ma et al., 2016).
The oven temperature program consisted of an initial hold at 50 °C for 1 min, followed by an increase at 25 °C min⁻¹ to 175 °C, then ramped at 4 °C min⁻¹ to 230 °C and held for 5 min. The detector temperature was set at 280 °C, with hydrogen, air, and helium flow rates of 40 mL min⁻¹, 450 mL min⁻¹, and 30 mL min⁻¹, respectively. Fatty acids were identified and quantified using a 37-component FAME standard mixture, which was used for calibration.
2.9. Biodiesel Property Estimation Based on Fatty Acid Composition
The physicochemical properties of biodiesel derived from microalgal lipids were estimated theoretically based on the fatty acid methyl ester profile, using established empirical correlations (Islam et al., 2013)(Mondal et al., 2021).
The iodine value (IV) was calculated as:
Iodine value (IV) = ------------------------------------------------(3)
The kinematic viscosity (ν) was determined using:
Kinematic viscosity (v) = --------------------------------------(4)
where: The higher heating value (HHV) was estimated as:
High heating value (HHV) = -----------------------------------------(5)
where:
δi=46.19−(1794/Mi)−0.21N
The density (ρ) of biodiesel was calculated using:
Density (ρ) = -----------------------------------------------------------(6)
where:
ρi=0.8463−(0.49/Mi)+0.0118N
The saponification value (SV) was calculated as:
Saponification value (SV) = -----------------------------------(7)
The cetane number (CN) was estimated using:
Cetane number (CN) = (8)
Finally, oxidation stability was determined as:
Oxidation stability = -------------------------------(9)
where represents the weight percentage of the fatty acid, is its corresponding weight fraction, is the molecular weight, and denotes the number of double bonds. corresponds to the combined weight percentage of linoleic and linolenic acids.
2.10. Statistical Analysis
All experiments were performed in triplicate (n=3). Statistical significance was assessed using one-way analysis of variance (ANOVA) with a 95% confidence interval (p < 0.05), implemented using the Data Analysis Toolpak in Microsoft office 365.
3. Results
3.1. Characterization of Matured Compost Leachate Wastewater
The physicochemical characteristics of the raw matured compost leachate collected from the Karadiyana dump site are summarized in Table 1. The leachate exhibited a slightly acidic to near-neutral pH (6.83 ± 0.15) and a dark brown, turbid appearance, indicating the presence of dissolved humic substances and stabilized organic matter.
The organic load was considerable, with a Chemical Oxygen Demand (COD) of 1365.89 ± 48.19 mg L⁻¹. However, when compared to fresh compost or landfill leachates—which can exhibit COD levels exceeding 10,000–50,000 mg L⁻¹—this value confirms the matured nature of the effluent, reflecting partial biological stabilization during composting.
Nutrient analysis revealed elevated concentrations of nitrogen and phosphorus, classifying the leachate as a high-strength wastewater. Total Nitrogen (TN) and Total Phosphorus (TP) concentrations were 735.29 ± 17.87 mg L⁻¹ and 20.87 ± 1.20 mg L⁻¹, respectively. The resulting N:P ratio (~35:1) deviates substantially from the Redfield ratio (16:1), indicating a phosphorus-limited nutrient balance. Despite this imbalance, the high nutrient content highlights the potential of this waste stream as a nutrient-rich cultivation medium, provided the microalgal strain can tolerate the associated turbidity and ammoniacal stress.
3.2. Effect of Leachate Dilution on Microalgal Growth (Screening Phase)
The growth response of Desmodesmus sp. was evaluated under varying leachate dilutions (25%, 50%, 75%, and 100% v/v) and compared with a BBM control over a 20-day cultivation period. One-way ANOVA revealed statistically significant differences in growth performance among treatments, based on both optical density (F₄,₁₀ = 33.62, p < 0.001) and dry cell weight (F₄,₁₀ = 82.82, p < 0.001).
As shown in Figure 1, cultures grown in higher leachate concentrations (75% and 100%) exhibited a distinct lag phase during the first four days, whereas lower dilutions and the control showed earlier growth initiation. Following this acclimatization phase, rapid exponential growth was observed in all leachate treatments.
By Day 20, the undiluted leachate (100%) achieved the highest optical density (2.269 ± 0.072), which was significantly higher than both the BBM control (1.668 ± 0.070) and the 75% dilution (1.894 ± 0.003) (p < 0.05). Biomass measurements corroborated these trends (Figure 2), with the 75% leachate yielding the highest dry biomass (2.025 ± 0.089 g L⁻¹). Importantly, the undiluted leachate also produced substantially higher biomass (1.567 ± 0.072 g L⁻¹) than the BBM control (1.063 ± 0.048 g L⁻¹).
Based on these results, the 100% leachate condition was selected for subsequent experiments. Although the 75% dilution produced marginally higher biomass, the elimination of freshwater dilution in the 100% condition represents a more sustainable and industrially relevant option, particularly given its ~47% higher biomass yield compared to the control medium.
3.3. Biomass Production in Main Cultivation Experiment
Growth kinetics of Desmodesmus sp. were further evaluated in undiluted matured compost leachate and BBM over a 24-day batch cultivation period. Optical density profiles (Figure 3) revealed distinct growth patterns between the two media. The leachate culture exhibited rapid early growth, reaching a plateau around Day 14 (OD ≈ 3.04), whereas the BBM control showed a more gradual increase, ultimately reaching a higher OD by Day 24 (4.372 ± 0.156).
Given the potential influence of wastewater turbidity and cellular morphology on optical measurements, biomass productivity was assessed using dry cell weight (DCW). As shown in Figure 4, DCW values in the leachate culture were consistently and significantly higher (p < 0.001) than those in the control throughout the cultivation period. The maximum biomass concentration in leachate reached 2.692 ± 0.093 g L⁻¹ on Day 22, compared to only 0.668 ± 0.031 g L⁻¹ in the BBM control.
Overall, cultivation in matured compost leachate resulted in approximately fourfold higher biomass accumulation relative to synthetic media, confirming its suitability as an effective growth substrate for high-density algal cultivation.
3.4. Nutrient Removal Efficiency
3.4.1. Total Nitrogen (TN) Removal
Nitrogen is a critical macronutrient for microalgal protein synthesis and growth. The temporal variation of Total Nitrogen (TN) concentration in the matured compost leachate and the BBM control is presented in Figure 5.
The initial TN concentration in the matured compost leachate was recorded at 735.29 ± 17.87 mg L⁻¹, significantly higher than the synthetic control (139.37 ± 4.42 mg L⁻¹), confirming the nitrogen-rich nature of the compost effluent. Despite this high initial load, Desmodesmus sp. demonstrated efficient nitrogen uptake capability.
A rapid depletion of nitrogen was observed during the first 4 days of cultivation, where the TN concentration in the leachate dropped sharply to 495.20 ± 16.43 mg L⁻¹. Statistical analysis confirms this initial reduction was highly significant (p < 0.01), corresponding to a removal of approximately 32.6% of the initial load. This correlates with the rapid exponential growth phase observed in the biomass data (Section 3.2). Following this initial surge, the removal rate stabilized but continued steadily throughout the cultivation period.
By the end of the experiment (Day 24), the TN concentration in the leachate had decreased to 291.75 ± 12.12 mg L⁻¹. This represents a statistically significant total reduction (p < 0.001) relative to the initial concentration, achieving a removal efficiency of 60.32%. In comparison, the control medium showed a depletion from 139.37 mg L⁻¹ to 10.82 mg L⁻¹, achieving a removal efficiency of 92.2%. Although the percentage removal was higher in the control due to the lower initial starting concentration, the total mass of nitrogen removed in the leachate system (−443.5 mg L⁻¹ removed) was substantially higher than in the control (−128.5 mg L⁻¹ removed). This high nutrient removal capacity highlights the potential of Desmodesmus sp. to bioremediate high-strength nitrogenous wastewaters while converting the excess nutrients into valuable biomass.
3.4.2. Total Phosphorus (TP) Removal
Phosphorus is a vital nutrient for microalgal metabolism, primarily involved in energy transfer (ATP synthesis) and nucleic acid formation. The removal of Total Phosphorus (TP) from the matured compost leachate and BBM control was monitored throughout the cultivation period, as shown in Figure 6.
The matured compost leachate initially contained a TP concentration of 20.87 ± 1.20 mg L⁻¹, which is typical for organic-rich effluents and significantly higher than the control medium (7.37 ± 0.14 mg L⁻¹). The removal trend followed a pattern similar to nitrogen uptake, characterized by a rapid reduction during the initial and rapid growth phases (Days 0–6). During this first week, the TP concentration in the leachate decreased to 12.96 ± 0.69 mg L⁻¹, indicating active assimilation by the algae during cell division.
By the end of the 24-day cultivation period, the TP concentration in the leachate had significantly decreased to 7.18 ± 0.09 mg L⁻¹ (p < 0.001), corresponding to a removal efficiency of 65.57%. In contrast, the BBM control achieved a higher removal efficiency of 91.54%, with the final concentration dropping to 0.62 ± 0.02 mg L⁻¹.
While the percentage removal was superior in the synthetic control, the absolute mass of phosphorus removed was significantly higher in the wastewater system. Desmodesmus sp. removed approximately 13.69 mg L⁻¹ of phosphorus from the leachate, compared to only 6.74 mg L⁻¹ in the control. This robust phosphorus uptake confirms that matured compost leachate provides bioavailable phosphate that can be effectively recovered by microalgal biomass, thereby reducing the eutrophication potential of the effluent before discharge.
3.4.3. Chemical Oxygen Demand (COD) Removal
Chemical Oxygen Demand (COD) serves as a primary indicator of the organic matter content in the wastewater. Unlike the inorganic BBM control, which contains negligible organic carbon, the matured compost leachate presented a high initial organic load, necessitating the evaluation of carbon removal efficiency. The temporal reduction of COD in the leachate culture is illustrated in Figure 7.
The initial COD of the matured compost leachate was recorded at 1365.89 ± 48.19 mg L⁻¹. A precipitous drop in COD was observed within the first 48 hours of cultivation, where the concentration decreased to 611.23 ± 30.48 mg L⁻¹. This corresponds to a removal of nearly 55% of the total organic load in just 2 days. This rapid consumption of organic carbon strongly suggests that Desmodesmus sp. switched to a mixotrophic growth mode, utilizing the dissolved organic compounds as an exogenous carbon source to fuel the rapid biomass production observed in the early growth phase (Section 3.2).
Following this initial rapid assimilation phase, the COD removal rate stabilized, decreasing gradually as the readily biodegradable organic fraction was exhausted. By Day 24, the final COD concentration reached 237.43 ± 10.95 mg L⁻¹, representing a statistically significant reduction (p < 0.001) from the initial value.
The overall COD removal efficiency achieved was 82.62%. This high removal efficiency indicates that Desmodesmus sp. is highly effective at mineralizing the organic pollutants present in matured compost leachate. The final COD levels are well within the discharge limits for many industrial effluents, demonstrating the dual capability of this system for both biomass generation and effective organic wastewater treatment.
3.4.4. pH Evolution During Cultivation
The variation of pH in the culture medium serves as an indirect proxy for microalgal metabolic activity, particularly photosynthetic carbon fixation. The pH profiles of the matured compost leachate and control cultures are presented in Figure 8.
Both media started with a near-neutral pH (6.83 ± 0.15 for leachate and 6.97 ± 0.18 for control). As cultivation progressed, a distinct alkalinization trend was observed in both conditions. In the matured compost leachate, the pH rose significantly, reaching a maximum value of 9.02 ± 0.18 by Day 20. A similar trend was observed in the control, which peaked at 8.88 ± 0.18.
This substantial increase in pH is attributed to the photosynthetic consumption of dissolved inorganic carbon. As Desmodesmus sp. actively assimilates dissolved CO₂ and bicarbonates (HCO₃⁻) during the light period, the equilibrium of the carbonate system shifts, leading to the consumption of hydrogen ions (H⁺) and a subsequent rise in hydroxyl ions (OH⁻). The fact that the pH remained consistently high (>8.5) from Day 6 to Day 20 confirms that the culture was maintaining robust photosynthetic activity throughout the exponential growth phase.
It is worth noting that sustaining a high pH (approx. 9.0) in wastewater treatment systems offers secondary benefits. Alkaline conditions are known to promote ammonia stripping (volatilization of free ammonia) and can induce the precipitation of heavy metals and phosphates. Furthermore, high pH levels create a harsh environment for enteric pathogens and coliforms, potentially enhancing the disinfection quality of the treated effluent. Towards the end of the cultivation (Days 22–24), a slight decline in pH was observed (dropping to 8.13 in leachate), likely signaling the onset of the stationary phase where cellular respiration and organic acid release began to counterbalance photosynthetic carbon uptake.
3.5. Lipid Production and Accumulation
The viability of microalgal biodiesel depends not only on biomass growth but critically on the lipid yield per unit volume of culture. To evaluate this, lipid weight (g) and lipid concentration (g L⁻¹) was quantified during the transition from the exponential to the stationary phase (Days 18–23), as presented in Figure 9 and Figure 10.
Interesting dynamics were observed regarding the timing of lipid accumulation. At Day 18, there was no statistically significant difference (p > 0.05) in lipid concentration between the matured compost leachate (0.320 ± 0.002 g L⁻¹) and the BBM control (0.334 ± 0.009 g L⁻¹). This indicates that during the active growth phase, metabolic energy was primarily directed toward cellular division and biomass synthesis rather than lipid storage.
However, a marked shift occurred as the culture entered the stationary phase. In the matured compost leachate cultures, lipid concentration surged rapidly between Day 18 and Day 22. The lipid yield more than doubled during this 4-day window, peaking at a maximum concentration of 0.715 ± 0.027 g L⁻¹ on Day 22. This accumulation coincides with the depletion of nitrogen observed in Section 3.3.1. It is well-established that under nitrogen-limited conditions, Desmodesmus sp. shifts its metabolic pathway from protein synthesis to triglyceride accumulation as an energy storage mechanism.
In contrast, the control cultures displayed a stagnant lipid profile, with concentrations fluctuating marginally around 0.350–0.360 g L⁻¹ throughout the stationary phase. Consequently, by Day 22, the matured compost leachate yielded approximately 2.03 times more lipid per liter compared to the synthetic control (p < 0.001). This superior volumetric lipid productivity confirms that matured compost leachate not only supports higher biomass densities but effectively triggers lipid accumulation, making it an excellent feedstock for scalable biodiesel production.
3.6. Fatty Acid Profiling and Biodiesel Characterization
3.6.1. Fatty Acid Methyl Ester (FAME) Composition
The quality and performance of microalgal biodiesel are intrinsically governed by the fatty acid profile of the parent oil. The FAME composition of Desmodesmus sp. cultivated in matured compost leachate was determined via GC-MS analysis (Table 3).
The raw chromatograms revealed that the crude lipid extract contained significant quantities of non-saponifiable lipids, primarily stigmasterol (RT 29.24 min), which accounted for approximately 38% of the raw peak area. For biodiesel characterization, the profile was normalized to include only fatty acid methyl esters. The analysis revealed a unique profile dominated by saturated fatty acids (SFAs), specifically medium-chain fatty acids. The most abundant components were Palmitic acid (C16:0) at 26.95% and Lauric acid (C12:0) at 25.78%. Notably, the presence of short-chain fatty acids such as Caprylic (C8:0) and Capric (C10:0) acid was also detected, which significantly influences the fuel's flow properties.
This profile contrasts with previous studies on Desmodesmus sp. cultivated in synthetic media (Pozzobon et al., 2020), which typically report profiles dominated by polyunsaturated C18 fatty acids. The shift towards shorter, saturated chains (C12–C16) in this study is likely a stress response to the complex organic matrix of the compost leachate, favoring the accumulation of energy-dense saturated lipids.
3.6.2. Estimation of Biodiesel Fuel Properties
The theoretical fuel properties were calculated based on the FAME composition and compared against ASTM D6751-12 and EN 14214:2012 standards (Table 4).
The Iodine Value (IV) was calculated to be 25.45 g I2/100 g, which is drastically lower than the EN maximum limit of 120. This indicates exceptional oxidative stability, as confirmed by the predicted Oxidative Stability time of >30 hours (significantly surpassing the ASTM requirement of 3 hours). This stability is attributed to the low content of polyunsaturated fatty acids (only 3.90%), preventing rapid fuel degradation.
The Saponification Value (SV) was calculated to be 228.5 mg KOH g-1. This relatively high value reflects the abundance of low-molecular-weight fatty acids (C8–C12). Consequently, the Cetane Number (CN) was estimated at 64.5. This value significantly exceeds the minimum requirements for both ASTM (>47) and EN (>51) standards, indicating that the fuel possesses superior ignition quality and would result in smoother engine combustion with minimal knocking.
Crucially, the presence of medium-chain fatty acids (C12:0, C14:0) balanced the high viscosity typically associated with saturated palmitic acid. The predicted Kinematic Viscosity was 3.42 mm² s⁻¹, which falls squarely within the ASTM D6751 standard range (1.9–6.0 mm² s⁻¹). Unlike long-chain saturated fats that can cause injector clogging, this viscosity profile suggests excellent atomization properties while maintaining sufficient lubricity for fuel system components. Therefore, the biodiesel derived from this feedstock meets key international standards for use as a standalone fuel or a premium blending stock.
4. Discussion
The ability of Desmodesmus sp. to sustain growth in 100% matured compost leachate without dilution represents a notable advance for wastewater-integrated microalgal cultivation. High-strength organic effluents are typically characterized by elevated ammoniacal nitrogen and organic loads that necessitate dilution or pre-treatment to prevent culture failure. Among these inhibitory constituents, ammonium nitrogen (NH₄⁺-N) is widely recognized as a primary toxicant in anaerobic and compost-derived wastewaters due to its direct interference with cellular metabolism and photosynthetic processes (López-Rosales et al., 2022).
Within this context, the four-day lag phase observed under undiluted leachate conditions should be interpreted as a physiological acclimatization period rather than growth inhibition. Previous kinetic studies have demonstrated a strong correlation between wastewater toxicity intensity and lag-phase duration (λ), with extended lag phases reflecting metabolic reprogramming rather than cell mortality (Khanzada & Övez, 2017) (López-Rosales et al., 2022). During this acclimation phase, free ammonia (NH₃), which readily diffuses across cellular membranes, disrupts photosystem II by uncoupling electron transport and destabilizing thylakoid membranes, resulting in transient suppression of photosynthetic efficiency(Metin & Altınbaş, 2024)(Drath et al., 2008).
In response to ammonium stress, microalgal cells activate detoxification and nitrogen assimilation pathways, particularly the ATP-dependent GS/GOGAT cycle, enabling conversion of toxic ammonium into amino acids and structural biomass. The subsequent recovery and proliferation of Desmodesmus sp. therefore indicate successful metabolic restructuring and restoration of photosynthetic functionality. This adaptive response supports the feasibility of gradual acclimatization strategies for stable microalgal operation in high-strength effluents without dilution, offering clear economic and operational advantages for large-scale wastewater treatment systems (Guo et al., 2025; López-Rosales et al., 2022; Ji et al., 2014a; J. Wang et al., 2019).
Following acclimatization, the rapid biomass accumulation observed confirms the energetic advantage of mixotrophic metabolism under compost leachate conditions. Unlike strict photoautotrophy, which is constrained by CO₂ mass-transfer limitations and photon availability, mixotrophy enables simultaneous utilization of light energy and organic carbon substrates (COD) through respiratory metabolism. This dual metabolic strategy promotes energetic coupling between chloroplasts and mitochondria, facilitating ATP and reducing-equivalent exchange (NADPH) and lowering the energetic cost of biomass synthesis (Yan et al., 2024; Z. Zhang et al., 2021; Bo et al., 2021; López-Rosales et al., 2022; Tossavainen et al., 2017).
Consistent with this framework, the pronounced growth surge recorded aligns with literature reporting 8–12-fold increases in biomass productivity under mixotrophic conditions relative to autotrophic controls. From a kinetic standpoint, the specific growth rate under mixotrophy can be approximated as the additive contribution of autotrophic and heterotrophic metabolic fluxes. This advantage allows Desmodesmus sp. to effectively exploit the biodegradable organic matter present in matured compost leachate, overcoming photosynthetic limitations imposed by the wastewater matrix and explaining the high biomass yields achieved (Eze et al., 2018; Cheng et al., 2013; de Souza et al., 2021; Yun et al., 2021; López-Rosales et al., 2022).
Sustained growth in optically dense leachate is particularly noteworthy, as wastewater systems commonly exhibit elevated background turbidity (K_bg) due to humic substances, suspended solids, and chromophoric organic matter. Such conditions intensify light attenuation, restrict the photic zone, and often induce severe photolimitation, with turbidity levels exceeding 90–100 NTU reported to cause biomass stagnation or collapse in conventional photobioreactors (Segredo-Morales et al., 2023; Otim et al., 2021; Benner et al., 2022; López-Rosales et al., 2022).
Under mixotrophic regimes, however, the organic turbidity characteristic of compost leachates provides a functional advantage. Rather than acting solely as an optical barrier, particulate and dissolved organic compounds serve as metabolizable carbon sources. By reducing reliance on photon penetration, Desmodesmus sp. was able to sustain growth even within light-limited reactor zones, effectively bypassing the classical shadowing effects that inhibit obligate photoautotrophs in turbid wastewater environments (Wu et al., 2025; Licata et al., 2025; López-Rosales et al., 2022).
In addition, the high biomass density achieved during cultivation likely enhanced tolerance to ammonia toxicity. Dense algal cultures are known to buffer toxic shocks more effectively than dilute inocula, as collective assimilation and diffusion gradients reduce intracellular exposure to inhibitory compounds. This density-dependent resistance plausibly contributed to the sustained stability of the culture under prolonged exposure to elevated ammonium concentrations (Collos & Harrison, 2014). Beyond sustaining biomass growth, the same physiological and metabolic mechanisms underpinning mixotrophic proliferation directly governed organic matter attenuation, thereby linking culture stability to wastewater treatment performance.
The high chemical oxygen demand (COD) removal efficiency achieved in this study (82.6%) demonstrates the strong capacity of Desmodesmus sp. to remediate organic-rich matured compost leachate. This value exceeds those commonly reported for Desmodesmus spp. cultivated in high-strength agro-industrial or landfill leachates, where COD reductions typically range from 36% to 70%, depending on wastewater composition and operational conditions (López-Rosales et al., 2022;(Ahmed et al., 2017a). Such performance underscores the suitability of matured compost leachate as an effective substrate for integrated algal wastewater treatment.
COD removal in microalgal systems occurs through multiple concurrent mechanisms, including (i) direct assimilation of dissolved organic carbon under mixotrophic metabolism, (ii) respiratory oxidation of organic substrates, and (iii) adsorption or entrapment of organic matter on algal cell surfaces and extracellular polymeric substances (EPS) (Debeni Devi et al., 2022). Among these pathways, mixotrophic assimilation is widely recognized as the dominant mechanism when readily biodegradable organic compounds are present (Yun et al., 2021). The rapid COD decline observed during the exponential growth phase in this study strongly supports the predominance of this biological assimilation pathway.
The superior COD removal can be further attributed to the distinct composition of matured compost leachate, which differs fundamentally from fresh leachate and many industrial effluents (Doyle et al., 2022). Its organic fraction is dominated by partially stabilized, low-molecular-weight compounds such as volatile fatty acids and humic degradation products, which exhibit higher bioavailability than recalcitrant industrial organics. This compositional characteristic provides a mechanistic explanation for the enhanced COD removal observed relative to untreated landfill leachates (López-Rosales et al., 2022).
Beyond algal assimilation, the observed COD removal efficiency is consistent with functional algal–bacterial cooperation under non-axenic cultivation conditions. In such systems, microalgae supply dissolved oxygen via photosynthesis, supporting aerobic bacterial degradation of organic matter, while heterotrophic bacteria mineralize organics into CO₂ that sustains algal photosynthesis and mixotrophic growth (Ríos et al., 2018). This synergistic carbon exchange has been widely reported to enhance COD removal beyond levels achievable by axenic algal cultures alone; Yun et al., 2021).
In addition, non-sterile conditions promote EPS secretion by both algae and bacteria, enhancing bio-flocculation and physical entrapment of suspended organic particulates. This process contributes further COD reduction through aggregation and sedimentation, particularly under high-strength wastewater conditions (Debeni Devi et al., 2022). Collectively, biological assimilation, microbial synergy, and EPS-mediated physical removal explain the high COD attenuation achieved.
Following organic carbon removal, total nitrogen (TN) removal reached approximately 60–72%, indicating effective nitrogen attenuation despite the high initial concentration (~800 mg L⁻¹). In algal-based systems, nitrogen removal is primarily driven by biological assimilation into proteins, nucleic acids, chlorophyll, and other cellular constituents (Song et al., 2024); (Salbitani & Carfagna, 2021). As biodegradable organic carbon was progressively depleted through mixotrophic assimilation and microbial oxidation, nitrogen transformation emerged as the next dominant regulatory process within the system.
Under ammonium-rich conditions, Desmodesmus sp. preferentially assimilates reduced nitrogen forms due to their lower energetic demand relative to nitrate reduction. This metabolic preference explains the rapid decline in ammoniacal nitrogen following acclimatization and aligns with reports of enhanced ammonium uptake in Desmodesmus cultivated in high-strength wastewaters(Wan et al., 2018)(Olofsson et al., 2019).
However, biological uptake alone does not fully account for nitrogen removal in unbuffered algal systems. The increase in culture pH to 9.02 created favorable conditions for ammonia stripping, whereby ammonium ions (NH₄⁺) shift to volatile free ammonia (NH₃) and are lost to the atmosphere. Experimental and thermodynamic studies indicate that at pH values between 9.0 and 9.5, approximately 10–30% of total ammoniacal nitrogen may be removed via volatilization (Ahmed et al., 2017b);(Liu et al., 2023)). While this abiotic loss does not contribute to biomass nitrogen recovery, it likely reduced free-ammonia toxicity and supported sustained algal growth (López-Rosales et al., 2022).
Phosphorus removal reached approximately 65–66%, consistent with values reported for algal systems operating under alkaline conditions. Phosphorus uptake occurs through assimilation into phospholipids, nucleotides, ATP, and intracellular polyphosphate storage via luxury uptake mechanisms (Martínez et al., 2018). In parallel with nitrogen attenuation, phosphorus dynamics were strongly influenced by the evolving physicochemical environment created by algal metabolism
In addition to biological assimilation, alkaline-induced precipitation contributed significantly to phosphorus removal. Photosynthetic CO₂ consumption increased culture pH, promoting the formation of insoluble phosphate minerals. In compost leachate matrices containing calcium and magnesium, phosphate precipitation may occur as calcium phosphate, magnesium phosphate, or struvite (MgNH₄PO₄·6H₂O) (Shaikh et al., 2024)(Gutiérrez et al., 2015). Previous studies indicate that at pH values above 8.5, physicochemical precipitation can account for 40–60% of total phosphorus removal when Ca²⁺ and Mg²⁺ are sufficient (Tirok & Scharler, 2014). Accordingly, phosphorus reduction in this study resulted from combined metabolic assimilation and mineral precipitation rather than biomass growth alone.
Nitrogen availability is a primary regulator of carbon partitioning in microalgae, governing the balance between biomass formation and lipid accumulation. Under nitrogen-replete conditions, assimilated carbon is preferentially directed toward protein synthesis, chlorophyll production, and active cell division(Fakhry & El Maghraby, 2015;Wang et al., 2019). When nitrogen becomes limiting, however, the synthesis of nitrogen-containing macromolecules is constrained, forcing excess fixed carbon to be redirected toward neutral lipid storage, predominantly as triacylglycerols (TAGs)(Adams et al., 2013;López-Rosales et al., 2022).
This regulatory shift was clearly reflected in the two-stage lipid production pattern observed in the present study. During the exponential growth phase, lipid concentration remained relatively low (0.320 g L⁻¹ at Day 18), coinciding with sufficient nitrogen availability to sustain rapid biomass accumulation. In contrast, lipid content more than doubled during the stationary phase (0.715 g L⁻¹ at Day 22), temporally aligned with near-complete nitrogen depletion from the compost leachate. This decoupling of biomass growth and lipid synthesis provides strong empirical support for the nitrogen starvation hypothesis, wherein lipid biosynthesis functions primarily as a survival-oriented carbon storage strategy rather than a growth-linked process (López-Rosales et al., 2022).
At the mechanistic level, nitrogen starvation suppresses protein and nucleic acid synthesis, redirecting photosynthetically fixed carbon flux toward fatty acid and TAG biosynthesis. Previous studies have shown that under nitrogen-limited conditions, Scenedesmus/Desmodesmus species can channel more than 90% of newly synthesized lipids into neutral lipid pools, forming dense intracellular lipid droplets that enhance survival during prolonged nutrient scarcity (López-Rosales et al., 2022). Accordingly, the stationary phase observed in this system represents a critical “starvation window,” during which metabolic flux was actively diverted toward lipid storage rather than cellular proliferation(Yang et al., 2014;Sulochana & Arumugam, 2020).
The substantially higher lipid yield achieved in compost leachate cultures relative to the BBM control further highlights the importance of carbon availability and the C:N ratio in regulating lipid biosynthesis. Wastewater-grown cultures reached a final lipid concentration of 0.715 g L⁻¹, more than twice that of the BBM control (0.352 g L⁻¹), despite comparable cultivation durations. This enhancement is directly attributable to the presence of biodegradable organic carbon in the matured compost leachate, which supported mixotrophic metabolism (Yun et al., 2021).
Under mixotrophic conditions, microalgae simultaneously utilize inorganic CO₂ and organic carbon substrates, reducing reliance on light-driven carbon fixation while increasing intracellular carbon availability. The high chemical oxygen demand (COD) of the compost leachate therefore created a high C:N ratio environment, which is widely recognized as a key driver of lipid accumulation. Once nitrogen becomes limiting, excess carbon cannot be incorporated into biomass and is instead diverted toward fatty acid and TAG synthesis (López-Rosales et al., 2022).
Consistent experimental evidence demonstrates that organic carbon supplementation significantly enhances lipid productivity compared to purely inorganic media. Carbon-supplemented systems consistently exhibit higher TAG accumulation than nitrogen-rich systems, confirming that lipid induction is not driven by nitrogen limitation alone but is strongly modulated by carbon availability (Yun et al., 2021). In the present study, the biodegradable organic fraction of the matured compost leachate effectively functioned as a heterotrophic substrate, sustaining carbon flux into lipid biosynthesis beyond levels achievable in the BBM control.
Beyond nutrient ratios, the complex chemical matrix of compost leachate imposed multifactorial environmental stress, including elevated ammonium concentrations, residual organic acids, trace metals, salinity, and pH fluctuations. Such stressors are well documented to intensify lipid induction responses in microalgae by activating protective metabolic pathways (Shaari et al., 2021; Alkhamis et al., 2022). Under stress conditions, microalgal cells downregulate protein synthesis and redirect metabolic energy toward fatty acid and TAG formation. Lipid droplets function as inert carbon sinks that reduce intracellular redox pressure, prevent over-reduction of the photosynthetic electron transport chain, and protect cellular components from oxidative damage. This stress-induced metabolic reprogramming explains why wastewater-grown Desmodesmus sp. accumulated substantially more lipids than cultures grown in nutrient-balanced BBM, where minimal physiological stress favored continued growth over carbon storage (Shi et al., 2020; Zienkiewicz et al., 2016; Wu et al., 2012).
The use of matured compost leachate was particularly advantageous in this regard. Compared with fresh leachate, matured leachate exhibits reduced acute toxicity while retaining sufficient nutrient and organic carbon levels to support initial biomass formation. This enabled an optimal cultivation trajectory consisting of an early growth phase supported by available nutrients, followed by rapid nutrient depletion that triggered stress-induced lipid accumulation. Such a naturally phased cultivation strategy mimics two-stage lipid production intrinsically, eliminating the need for artificial stress induction techniques and thereby improving process simplicity and economic feasibility (Apandi et al., 2020; Bomer & Leverett, 2024; Sriram & Seenivasan, 2015).
Qualitatively, this environmental stress induced a pronounced metabolic shift toward saturated fatty acids (SFAs), which accounted for 75.6% of total fatty acids and were dominated by palmitic acid (C16:0) (Vimali et al., 2022). Although this profile contrasts with the PUFA-rich compositions typically reported for algae cultivated in clean, nutrient-replete media, extensive literature confirms that such a shift represents a predictable stress-induced metabolic adaptation. Nutrient limitation and chemical stress inherent to compost leachate environments are well known to trigger lipid reprogramming, where biosynthesis shifts away from membrane-bound polar lipids toward neutral storage triacylglycerols (TAGs) (Arguelles et al., 2019).
This mechanism is further reinforced by the "membrane hardening" hypothesis, whereby microalgae reduce fatty acid unsaturation to decrease membrane fluidity and permeability under hostile environmental conditions. Increased SFA content acts as a defensive mechanism against osmotic and chemical stress by limiting the diffusion of toxic ions into the cell (López-Rosales et al., 2022). Comparable shifts toward SFA-dominated lipid profiles have been reported under salinity and wastewater stress, confirming that the dominance of C16:0 observed in this study reflects a physiologically rational adaptation rather than aberrant metabolism (De Bhowmick et al., 2025; Chen et al., 2022).
From a fuel chemistry perspective, this reduction in PUFA content represents a substantial technical advantage. Polyunsaturated fatty acids are highly susceptible to oxidative degradation, leading to gum formation and reduced fuel stability. Consequently, the high oxidative stability (>30 h) predicted for the biodiesel derived in this study is directly attributable to its elevated SFA fraction and low iodine value, effectively enhancing long-term storage stability without the need for antioxidant additives (Huang et al., 2022; Amran et al., 2022).
The elevated saturation level also translated into superior ignition behavior, as reflected by the high cetane number of 64.5. This value exceeds those reported for most conventional vegetable oil biodiesels, including rapeseed, soybean, and palm oil (Inambao, 2020). Furthermore, compared with biodiesel produced via direct chemical transesterification of landfill-derived oils (Cetane Index ~55.4), the algal biodiesel produced here exhibited markedly superior ignition quality, underscoring the value of microalgal bioconversion for leachate valorization (Mekonnen et al., 2024).
Regarding flow properties, the predicted kinematic viscosity was approximately 3.42 mm² s⁻¹. This value falls squarely within the ASTM D6751 limits (1.9–6.0 mm² s⁻¹), driven by the balancing effect of medium-chain fatty acids (Lauric C12:0) offsetting the heavier Palmitic acid fractions. This indicates that the fuel possesses adequate lubricity for injection systems without requiring viscosity modifiers. While the high saturation level inherently limits cold-flow performance, this leachate-derived biodiesel is optimally positioned for applications in warm climates or stationary energy systems—such as agricultural machinery and backup generators—where oxidative stability is paramount (Calijuri et al., 2025; Velásquez-Orta et al., 2024).
From a sustainability perspective, the ability of Desmodesmus sp. to grow directly in 100% matured compost leachate aligns strongly with circular bioeconomy principles. Nutrient recycling, carbon diversion from mineralization, and the elimination of freshwater dilution reduce environmental impact and improve resource efficiency. Future research should focus on optimizing reactor operation, microbial community dynamics, and downstream biomass valorization to further enhance productivity and techno-environmental benefits.
5. Conclusions
This study provides compelling evidence that matured compost leachate can be directly valorized as both a cultivation medium and nutrient source for microalgal biodiesel production using Desmodesmus sp. Sustained growth in 100% undiluted leachate following a short acclimatization phase demonstrates the robustness of the strain and confirms that dilution, sterilization, or synthetic nutrient supplementation are not prerequisites for stable operation. This represents a significant advancement for wastewater-integrated algal bioprocesses, where high-strength effluents typically impose severe biological constraints.
Stable mixotrophic metabolism enabled simultaneous biomass generation and efficient wastewater remediation, achieving substantial reductions in COD (82.6%), total nitrogen, and total phosphorus. Organic carbon assimilation, algal–bacterial synergy, and physicochemical nutrient removal mechanisms collectively governed treatment performance, underscoring the functional coupling between biological resilience and pollutant attenuation. The naturally evolving cultivation trajectory—comprising acclimation, rapid biomass accumulation, and nutrient-depletion-driven stress—effectively mimicked a two-stage production strategy without external process manipulation.
Nitrogen depletion emerged as the principal trigger for metabolic reprogramming toward lipid accumulation, resulting in significantly enhanced lipid yields relative to the inorganic control medium. The presence of biodegradable organic carbon and a high C:N ratio further intensified lipid biosynthesis under mixotrophic conditions. These findings confirm that lipid induction in wastewater-grown microalgae is governed by the combined effects of nutrient limitation, carbon availability, and environmental stress rather than nitrogen starvation alone.
Fatty acid profiling revealed a pronounced shift toward saturated and monounsaturated fatty acids, yielding biodiesel with high cetane number (64.5), low iodine value, and excellent oxidative stability. The predicted kinematic viscosity (~3.42 mm² s⁻¹) falls well within international biodiesel standards (ASTM D6751), driven by the balancing effect of medium-chain fatty acids. Consequently, the resulting fuel possesses excellent ignition quality and storage stability, making it particularly advantageous for applications in warm climates and stationary energy systems.
Finally, this work establishes matured compost leachate as a low-cost, nutrient-rich feedstock capable of supporting high-performance microalgal cultivation while delivering tangible wastewater treatment benefits. By converting a problematic waste stream into biomass and energy carriers, the proposed system aligns strongly with circular bioeconomy principles and sustainable resource management. Future studies should focus on pilot-scale validation, reactor configuration optimization, microbial community control, and comprehensive life cycle and techno-economic assessments to accelerate translation toward industrial implementation.
References
- Adams, C., Godfrey, V., Wahlen, B., Seefeldt, L., & Bugbee, B. (2013). Understanding precision nitrogen stress to optimize the growth and lipid content tradeoff in oleaginous green microalgae. Bioresource Technology, 131, 188–194. [CrossRef]
- Adewuyi, A. (2022). Production of Biodiesel from Underutilized Algae Oil: Prospects and Current Challenges Encountered in Developing Countries. In Biology (Vol. 11, Issue 10). MDPI. [CrossRef]
- Ahamad Sanadi, N. F. B., Ibrahim, N., Ong, P. Y., Klemeš, J. J., Li, C., & Lee, C. T. (2021). Dilution rate of compost leachate from different biowaste for the fertigation of vegetables. Journal of Environmental Management, 295, 113010. [CrossRef]
- Association, A. P. H. (1926). Standard methods for the examination of water and wastewater (Vol. 6). American public health association.
- Awogbemi, O., & Desai, D. A. (2025). Progress in the conversion of biodiesel-derived crude glycerol into biofuels and other bioproducts. Bioresource Technology Reports, 30, 102106. [CrossRef]
- Adams, C., Godfrey, V., Wahlen, B., Seefeldt, L., & Bugbee, B. (2013). Understanding precision nitrogen stress to optimize the growth and lipid content tradeoff in oleaginous green microalgae. Bioresource Technology, 131, 188–194. [CrossRef]
- Ahmed, A., Jyothi, N., & Ramesh, A. (2017). Improved ammonium removal from industrial wastewater through systematic adaptation of wild type Chlorella pyrenoidosa. Water Science and Technology, 75(1), 182–188. [CrossRef]
- Alkhamis, Y. A., Mathew, R. T., Nagarajan, G., Rahman, S. M., & Rahman, M. M. (2022). pH induced stress enhances lipid accumulation in microalgae grown under mixotrophic and autotrophic condition. Frontiers in Energy Research, 10. [CrossRef]
- Amran, N. A., Bello, U., & Hazwan Ruslan, M. S. (2022). The role of antioxidants in improving biodiesel’s oxidative stability, poor cold flow properties, and the effects of the duo on engine performance: A review. Heliyon, 8(7), e09846. [CrossRef]
- Apandi, N., Maya Saphira Radin Mohamed, R., Ibrahim Abdullah Abuala, A., & Amhimmid, A. A. (2020). Integrated Growth Potential of Scenedesmus sp. using Public Market Wastewater via Phycoremediation Process. INTERNATIONAL JOURNAL OF INTEGRATED ENGINEERING, 12(4), 290–299.
- Arguelles, E., Laurena, A. C., Monsalud, R. G., & Martinez-Goss, M. R. (2019). High lipid and protein-producing epilithic microalga, Desmodesmus sp.(U-AU2): a promising alternative feedstock for biodiesel and animal feed production. Philippine Journal of Crop Science, 44(2), 13–23.
- Bartolacci, C., Andreani, C., El-Gammal, Y., & Scaglioni, P. P. (2021). Lipid Metabolism Regulates Oxidative Stress and Ferroptosis in RAS-Driven Cancers: A Perspective on Cancer Progression and Therapy. In Frontiers in Molecular Biosciences (Vol. 8). Frontiers Media S.A. [CrossRef]
- Bomer, L. K., & Leverett, B. D. (2024). Growth Characteristics of a Desmodesmus Species from the San Antonio Springs and Its Short-Term Impact on Soil Microbial Dynamics. Life, 14(9). [CrossRef]
- Chang, H., Quan, X., Zhong, N., Zhang, Z., Lu, C., Li, G., Cheng, Z., & Yang, L. (2018). High-efficiency nutrients reclamation from landfill leachate by microalgae Chlorella vulgaris in membrane photobioreactor for bio-lipid production. Bioresource Technology, 266, 374–381. [CrossRef]
- Chen, X., He, Y., Liu, L., Zhu, X., Sen, B., & Wang, G. (2022). Nitrogen Starvation Enhances the Production of Saturated and Unsaturated Fatty Acids in Aurantiochytrium sp. PKU#SW8 by Regulating Key Biosynthetic Genes. Marine Drugs, 20(10). [CrossRef]
- Cinq-Mars, M., Bourdeau, N., Marchand, P., Desgagné-Penix, I., & Barnabé, S. (2022). Characterization of two microalgae consortia grown in industrial wastewater for biomass valorization. Algal Research, 62, 102628. [CrossRef]
- De Bhowmick, G., Plouviez, M., Reis, M. G., Guieysse, B., Everett, D. W., Agnew, M. P., Maclean, P., & Thum, C. (2025). Enhancing sn-2 specific fatty acid accumulation in microalgae: Impact of nutrient limitation on structured triacylglycerol profiles. International Journal of Food Science and Technology, 60(2). [CrossRef]
- Debeni Devi, N., Sun, X., Ding, L., Goud, V. V, & Hu, B. (2022). Mixotrophic growth regime of novel strain Scenedesmus sp. DDVG I in municipal wastewater for concomitant bioremediation and valorization of biomass Word count: 8427 including references.
- Doyle, M. G., Odenkirk, M. T., Stewart, A. K., Nelson, J. P., Baker, E. S., & De La Cruz, F. (2022). Assessing the Fate of Dissolved Organic Compounds in Landfill Leachate and Wastewater Treatment Systems. ACS ES&T Water, 2(12), 2502–2509. [CrossRef]
- Fakhry, E. M., & El Maghraby, D. M. (2015). Lipid accumulation in response to nitrogen limitation and variation of temperature in nannochloropsis salina. Botanical Studies, 56. [CrossRef]
- Gutiérrez, R., Passos, F., Ferrer, I., Uggetti, E., & García, J. (2015). Harvesting microalgae from wastewater treatment systems with natural flocculants: Effect on biomass settling and biogas production. Algal Research, 9, 204–211. [CrossRef]
- Howlett, N. G., & Avery, S. V. (1997). Induction of Lipid Peroxidation during Heavy Metal Stress in Saccharomyces cerevisiae and Influence of Plasma Membrane Fatty Acid Unsaturation. In APPLIED AND ENVIRONMENTAL MICROBIOLOGY (Vol. 63, Issue 8). https://journals.asm.org/journal/aem.
- Huang, Y., Li, F., Bao, G., Li, M., & Wang, H. (2022). Qualitative and quantitative analysis of the influence of biodiesel fatty acid methyl esters on iodine value. Environmental Science and Pollution Research, 29(2), 2432–2447. [CrossRef]
- Knothe, G., & Razon, L. F. (2017). Biodiesel fuels. Progress in Energy and Combustion Science, 58, 36–59. [CrossRef]
- Lee, J. C., Moon, K., Lee, N., Ryu, S., Song, S. H., Kim, Y. J., Lee, S. M., Kim, H. W., & Joo, J. H. (2023). Biodiesel production and simultaneous treatment of domestic and livestock wastewater using indigenous microalgae, Chlorella sorokiniana JD1-1. Scientific Reports, 13(1). [CrossRef]
- Liew, L. W., Bashir, M. J., Toh, P. Y., Alazaiza, M. Y. D., Amr, S. S. A., Khoo, K. S., Raksasat, R., & Lim, J. W. (2025). Microalgae cultivation in stabilized landfill leachate for simultaneous treatment and biomass production. Journal of the Taiwan Institute of Chemical Engineers, 166, 105068. [CrossRef]
- Liu, J., Pemberton, B., Scales, P. J., & Martin, G. J. O. (2023). Ammonia tolerance of filamentous algae Oedogonium, Spirogyra, Tribonema and Cladophora, and its implications on wastewater treatment processes. Algal Research, 72. [CrossRef]
- Maneechote, W., Pathom-Aree, W., Kamngoen, A., Wichaphian, A., Cheirsilp, B., Khoo, K. S., Huo, S., Srimongkol, P., & Srinuanpan, S. (2025). Sustainable PUFA-Rich Lipid-Accumulating Biomass Production via Dual Waste Valorization Using Heterotrophic Microalgae Cultivated on Anaerobic Effluent and Molasses. Journal of Microbiology and Biotechnology, 35, e2506037. [CrossRef]
- Martínez, C., Mairet, F., & Bernard, O. (2018). Theory of turbid microalgae cultures. Journal of Theoretical Biology, 456, 190–200. [CrossRef]
- Mekonnen, K. D., Endris, Y. A., & Abdu, K. Y. (2024). Alternative Methods for Biodiesel Cetane Number Valuation: A Technical Note. In ACS Omega (Vol. 9, Issue 6, pp. 6296–6304). American Chemical Society. [CrossRef]
- Miraboutalebi, S. M., Kazemi, P., & Bahrami, P. (2016). Fatty Acid Methyl Ester (FAME) composition used for estimation of biodiesel cetane number employing random forest and artificial neural networks: A new approach. Fuel, 166, 143–151. [CrossRef]
- Moreno-Garcia, L., Gariépy, Y., Barnabé, S., & Raghavan, V. (2021a). Factors affecting the fatty acid profile of wastewater-grown-algae oil as feedstock for biodiesel. Fuel, 304, 121367. [CrossRef]
- Moreno-Garcia, L., Gariépy, Y., Barnabé, S., & Raghavan, V. (2021b). Factors affecting the fatty acid profile of wastewater-grown-algae oil as feedstock for biodiesel. Fuel, 304, 121367. [CrossRef]
- Mortensen, M. S., Ruiz, J., & Watts, J. L. (2023). Polyunsaturated Fatty Acids Drive Lipid Peroxidation during Ferroptosis. In Cells (Vol. 12, Issue 5). MDPI. [CrossRef]
- Olofsson, M., Robertson, E. K., Edler, L., Arneborg, L., Whitehouse, M. J., & Ploug, H. (2019). Nitrate and ammonium fluxes to diatoms and dinoflagellates at a single cell level in mixed field communities in the sea. Scientific Reports, 9(1). [CrossRef]
- Ríos, L. F., Martinez, A., Klein, B. C., Maciel, M. R. W., & Filho, R. M. (2018). Comparison of Growth and Lipid Accumulation at Three Different Growth Regimes with Desmodesmus sp. Waste and Biomass Valorization, 9(3), 421–427. [CrossRef]
- Shaari, A. L., Sa, S. N. C., Surif, M., Zolkarnain, N., & Ghazali, R. (2021). Growth of marine microalgae in landfill leachate and their ability as pollutants removal. Tropical Life Sciences Research, 32(2), 133–146. [CrossRef]
- Shaikh, S. M. R., Quadir, M. A., Nasser, M. S., Rekik, H., Hassan, M. K., Ayesh, A. I., & Sayadi, S. (2024). Investigation of flocculation and rheological properties of microalgae suspensions cultivated in industrial process wastewater. Separation and Purification Technology, 328. [CrossRef]
- Shi, T. Q., Wang, L. R., Zhang, Z. X., Sun, X. M., & Huang, H. (2020). Stresses as First-Line Tools for Enhancing Lipid and Carotenoid Production in Microalgae. In Frontiers in Bioengineering and Biotechnology (Vol. 8). Frontiers Media S.A. [CrossRef]
- Song, H., Li, J., Su, Q., Li, H., Guo, X., Shao, S., Fan, L., Xu, P., Zhou, W., & Qian, J. (2024). Insight into the mechanism of nitrogen sufficiency conversion strategy for microalgae-based ammonium-rich wastewater treatment. Chemosphere, 349, 140904. [CrossRef]
- Sriram, S., & Seenivasan, R. (2015). Biophotonic perception on Desmodesmus sp. VIT growth, lipid and carbohydrate content. Bioresource Technology, 198, 626–633. [CrossRef]
- Sulochana, S. B., & Arumugam, M. (2020). Targeted Metabolomic and Biochemical Changes During Nitrogen Stress Mediated Lipid Accumulation in Scenedesmus quadricauda CASA CC202. Frontiers in Bioengineering and Biotechnology, 8. [CrossRef]
- Tirok, K., & Scharler, U. M. (2014). Influence of variable water depth and turbidity on microalgae production in a shallow estuarine lake system - A modelling study. Estuarine, Coastal and Shelf Science, 146, 111–127. [CrossRef]
- Velásquez-Orta, S. B., Yáñez-Noguez, I., Ramírez, I. M., & Ledesma, M. T. O. (2024). Pilot-scale microalgae cultivation and wastewater treatment using high-rate ponds: a meta-analysis. Environmental Science and Pollution Research, 31(34), 46994–47021. [CrossRef]
- Vimali, E., Senthil Kumar, A., Sakthi Vignesh, N., Ashokkumar, B., Dhakshinamoorthy, A., Udayan, A., Arumugam, M., Pugazhendhi, A., & Varalakshmi, P. (2022). RETRACTED: Enhancement of lipid accumulation in microalga Desmodesmus sp. VV2: Response Surface Methodology and Artificial Neural Network modeling for biodiesel production. Chemosphere, 293, 133477. [CrossRef]
- Wan, X. S., Sheng, H. X., Dai, M., Zhang, Y., Shi, D., Trull, T. W., Zhu, Y., Lomas, M. W., & Kao, S. J. (2018). Ambient nitrate switches the ammonium consumption pathway in the euphotic ocean. Nature Communications, 9(1). [CrossRef]
- Wang, X., Fosse, H. K., Li, K., Chauton, M. S., Vadstein, O., & Reitan, K. I. (2019). Influence of nitrogen limitation on lipid accumulation and EPA and DHA content in four marine microalgae for possible use in aquafeed. Frontiers in Marine Science, 6(MAR). [CrossRef]
- Wu, L. F., Chen, P. C., Huang, A. P., & Lee, C. M. (2012). The feasibility of biodiesel production by microalgae using industrial wastewater. Bioresource Technology, 113, 14–18. [CrossRef]
- Yang, Z. K., Ma, Y. H., Zheng, J. W., Yang, W. D., Liu, J. S., & Li, H. Y. (2014). Proteomics to reveal metabolic network shifts towards lipid accumulation following nitrogen deprivation in the diatom Phaeodactylum tricornutum. Journal of Applied Phycology, 26(1), 73–82. [CrossRef]
- Yun, H. S., Kim, Y. S., & Yoon, H. S. (2021). Effect of Different Cultivation Modes (Photoautotrophic, Mixotrophic, and Heterotrophic) on the Growth of Chlorella sp. and Biocompositions. Frontiers in Bioengineering and Biotechnology, 9. [CrossRef]
- Zienkiewicz, K., Du, Z. Y., Ma, W., Vollheyde, K., & Benning, C. (2016). Stress-induced neutral lipid biosynthesis in microalgae — Molecular, cellular and physiological insights. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids, 1861(9), 1269–1281. [CrossRef]
- Azarpour, A., Zendehboudi, S., Mohammadzadeh, O., Rajabzadeh, A. R., & Chatzis, I. (2022). A review on microalgal biomass and biodiesel production through Co-cultivation strategy. Energy Conversion and Management, 267, 115757. [CrossRef]
- Behera, S., Singh, R., Arora, R., Sharma, N. K., Shukla, M., & Kumar, S. (2015). Scope of Algae as Third Generation Biofuels. In Frontiers in Bioengineering and Biotechnology (Vol. 2). Frontiers Media S.A. [CrossRef]
- Benner, P., Meier, L., Pfeffer, A., Krüger, K., Oropeza Vargas, J. E., & Weuster-Botz, D. (2022). Lab-scale photobioreactor systems: principles, applications, and scalability. Bioprocess and Biosystems Engineering, 45(5), 791–813. [CrossRef]
- Bo, D. D., Magneschi, L., Bedhomme, M., Billey, E., Deragon, E., Storti, M., Menneteau, M., Richard, C., Rak, C., Lapeyre, M., Lembrouk, M., Conte, M., Gros, V., Tourcier, G., Giustini, C., Falconet, D., Curien, G., Allorent, G., Petroutsos, D., … Collin, S. (2021). Consequences of Mixotrophy on Cell Energetic Metabolism in Microchloropsis gaditana Revealed by Genetic Engineering and Metabolic Approaches. Frontiers in Plant Science, 12. [CrossRef]
- Breuer, G., Evers, W. A. C., de Vree, J. H., Kleinegris, D. M. M., Martens, D. E., Wijffels, R. H., & Lamers, P. P. (2013). Analysis of Fatty Acid Content and Composition in Microalgae. JoVE, 80, e50628.
- Casanova, L. M., Mendes, L. B. B., Corrêa, T. de S., da Silva, R. B., Joao, R. R., Macrae, A., & Vermelho, A. B. (2023). Development of Microalgae Biodiesel: Current Status and Perspectives. In Microorganisms (Vol. 11, Issue 1). MDPI. [CrossRef]
- Cheng, H., Tian, G., & Liu, J. (2013). Enhancement of biomass productivity and nutrients removal from pretreated piggery wastewater by mixotrophic cultivation of Desmodesmus sp. CHX1. Desalination and Water Treatment, 51(37–39), 7004–7011. [CrossRef]
- Collos, Y., & Harrison, P. J. (2014). Acclimation and toxicity of high ammonium concentrations to unicellular algae. Marine Pollution Bulletin, 80(1–2), 8–23. [CrossRef]
- de Souza, L., Lima, A. S., Matos, Â. P., Wheeler, R. M., Bork, J. A., Vieira Cubas, A. L., & Moecke, E. H. S. (2021). Biopolishing sanitary landfill leachate via cultivation of lipid-rich Scenedesmus microalgae. Journal of Cleaner Production, 303, 127094. [CrossRef]
- Dodangodage, C. A., Gamage, G. N., Fernando, K. V., Kasturiarachchi, J. C., Perera, T. A., Rajapakshe, S. D., & Halwatura, R. U. (2025). Production of Carbohydrate-Rich Chlorella sp. Biomass Using Clarified Aquaponics Effluent for Bioethanol Feedstock Applications.
- Dodangodage, C. A., Kasturiarachchi, J., Perera, T., Rajapakshe, D., & Halwatura, R. (2025a). Integrated Microalgal-Aquaponic Systems for Enhanced Water Treatment and Food Security: A Critical Review of Recent Advances in Process Integration and Resource Recovery.
- Dodangodage, C. A., Kasturiarachchi, J., Perera, T., Rajapakshe, D., & Halwatura, R. (2025b). Valorization of Canteen Wastewater Through Optimized Spirulina platensis Cultivation for Enhanced Carotenoid Production and Nutrient Removal. [CrossRef]
- Dodangodage, C. A., Premarathne, H., Kasturiarachchi, J. C., Perera, T. A., Rajapakshe, D., & Halwatura, R. U. (2025). Algae-Based Protective Coatings for Sustainable Infrastructure: A Novel Framework Linking Material Chemistry, Techno-Economics, and Environmental Functionality. Phycology, 5(4), 84. [CrossRef]
- DuBois, Michel., Gilles, K. A., Hamilton, J. K., Rebers, P. A., & Smith, Fred. (1956). Colorimetric Method for Determination of Sugars and Related Substances. Analytical Chemistry, 28(3), 350–356. [CrossRef]
- El-Sheekh, M. M., Galal, H. R., Mousa, A. S. H., & Farghl, A. A. M. (2023). Coupling wastewater treatment, biomass, lipids, and biodiesel production of some green microalgae. Environmental Science and Pollution Research, 30(12), 35492–35504. [CrossRef]
- Eze, V. C., Velasquez-Orta, S. B., Hernández-García, A., Monje-Ramírez, I., & Orta-Ledesma, M. T. (2018). Kinetic modelling of microalgae cultivation for wastewater treatment and carbon dioxide sequestration. Algal Research, 32, 131–141. [CrossRef]
- Gu, D., Xiao, Q., Zhao, Y., & Yu, X. (2023). A low-cost technique for biodiesel production in Ankistrodesmus sp. EHY by using harvested microalgal effluent. Science of The Total Environment, 857, 159461. [CrossRef]
- Guo, X., Chen, Z., He, Y., Zhou, Y., Chen, B., & Wang, M. (2025). Study on high ammonium tolerance and nitrogen utilization mechanisms of Chlamydomonas sp. YC in rare earth elements wastewater treatment. Algal Research, 91, 104231. [CrossRef]
- Holechek, J. L., Geli, H. M. E., Sawalhah, M. N., & Valdez, R. (2022). A Global Assessment: Can Renewable Energy Replace Fossil Fuels by 2050? Sustainability (Switzerland), 14(8). [CrossRef]
- Islam, M. A., Magnusson, M., Brown, R. J., Ayoko, G. A., Nabi, M. N., & Heimann, K. (2013). Microalgal species selection for biodiesel production based on fuel properties derived from fatty acid profiles. Energies, 6(11), 5676–5702. [CrossRef]
- Ji, F., Liu, Y., Hao, R., Li, G., Zhou, Y., & Dong, R. (2014a). Biomass production and nutrients removal by a new microalgae strain Desmodesmus sp. in anaerobic digestion wastewater. Bioresource Technology, 161, 200–207. [CrossRef]
- Ji, F., Liu, Y., Hao, R., Li, G., Zhou, Y., & Dong, R. (2014b). Biomass production and nutrients removal by a new microalgae strain Desmodesmus sp. in anaerobic digestion wastewater. Bioresource Technology, 161, 200–207. [CrossRef]
- Jia, C., Liu, N., Chen, L., Yan, W., Zhao, Q., Li, J., Yu, Z., Song, M., Xu, C., & Song, X. (2025). The growth and nutrient removal properties of heterotrophic microalgae using volatile fatty acids under different nitrogen forms and its application in actual acid-producing wastewater. Journal of Environmental Chemical Engineering, 13(1), 115347. [CrossRef]
- Jia, J., Han, D., Gerken, H. G., Li, Y., Sommerfeld, M., Hu, Q., & Xu, J. (2015). Molecular mechanisms for photosynthetic carbon partitioning into storage neutral lipids in Nannochloropsis oceanica under nitrogen-depletion conditions. Algal Research, 7, 66–77. [CrossRef]
- Kadir, W. N. A., Lam, M. K., Uemura, Y., Lim, J. W., & Lee, K. T. (2018). Harvesting and pre-treatment of microalgae cultivated in wastewater for biodiesel production: A review. Energy Conversion and Management, 171, 1416–1429. [CrossRef]
- Kanchanamala Delanka-Pedige, H. M., Munasinghe-Arachchige, S. P., Abeysiriwardana-Arachchige, I. S. A., & Nirmalakhandan, N. (2021). Evaluating wastewater treatment infrastructure systems based on UN Sustainable Development Goals and targets. Journal of Cleaner Production, 298, 126795. [CrossRef]
- Kanwal, F., Aslam, A., & Torriero, A. A. J. (2025). Microalgae-based biodiesel: integrating AI, CRISPR and nanotechnology for sustainable biofuel development. In Emerging Topics in Life Sciences (Vol. 8, Issue 3, pp. 131–143). Portland Press Ltd. [CrossRef]
- Kumar, M., Mandal, B. K., Ganguly, A., Ravi, R., Alam, T., Siddiqui, M. I. H., & Eldin, S. M. (2023). Performance evaluation of a diesel engine fueled with Chlorella Protothecoides microalgal biodiesel. Case Studies in Thermal Engineering, 51, 103609. [CrossRef]
- Lee, Y., Chen, W., Shen, H., Han, D., Li, Y., Jones, H. D. T., Timlin, J. A., & Hu, Q. (2013). Basic culturing and analytical measurement techniques. Handbook of Microalgal Culture: Applied Phycology and Biotechnology, 37–68.
- Li, J., & Li, G. (2023). What drives resource sustainability in Asia? Discovering the moderating role of financial development and industrialization. Resources Policy, 85, 103650. [CrossRef]
- Licata, G., Galasso, C., Palma Esposito, F., Palumbo Piccionello, A., & Villanova, V. (2025). Mixotrophy in Marine Microalgae to Enhance Their Bioactivity. Microorganisms, 13(2). [CrossRef]
- Liu, Z., Deng, Z., Davis, S. J., & Ciais, P. (2024). Global carbon emissions in 2023. Nature Reviews Earth & Environment, 5(4), 253–254. [CrossRef]
- López-Rosales, L., López-García, P., Benyachou, M. A., Molina-Miras, A., Gallardo-Rodríguez, J. J., Cerón-García, M. C., Sánchez Mirón, A., & García-Camacho, F. (2022). Treatment of secondary urban wastewater with a low ammonium-tolerant marine microalga using zeolite-based adsorption. Bioresource Technology, 359. [CrossRef]
- Ma, X., Liu, J., Liu, B., Chen, T., Yang, B., & Chen, F. (2016). Physiological and biochemical changes reveal stress-associated photosynthetic carbon partitioning into triacylglycerol in the oleaginous marine alga Nannochloropsis oculata. Algal Research, 16, 28–35. [CrossRef]
- Mallick, N., Mandal, S., Singh, A. K., Bishai, M., & Dash, A. (2012). Green microalga Chlorella vulgaris as a potential feedstock for biodiesel. Journal of Chemical Technology & Biotechnology, 87(1), 137–145.
- Maltsev, Y., Kulikovskiy, M., & Maltseva, S. (2023). Nitrogen and phosphorus stress as a tool to induce lipid production in microalgae. In Microbial Cell Factories (Vol. 22, Issue 1). BioMed Central Ltd. [CrossRef]
- Mandotra, S. K., Kumar, P., Suseela, M. R., Nayaka, S., & Ramteke, P. W. (2016). Evaluation of fatty acid profile and biodiesel properties of microalga Scenedesmus abundans under the influence of phosphorus, pH and light intensities. Bioresource Technology, 201, 222–229. [CrossRef]
- Maroušek, J., Gavurová, B., Strunecký, O., Maroušková, A., Sekar, M., & Marek, V. (2023). Techno-economic identification of production factors threatening the competitiveness of algae biodiesel. Fuel, 344, 128056. [CrossRef]
- Mathimani, T., Rene, E. R., Devanesan, S., AlSalhi, M. S., & Shanmuganathan, R. (2023). Assessment of taxonomically diverse Chlorococcum species and Chroococcus species for cell density, pigments, biochemical components, and fatty acid composition for fuel/food applications. Algal Research, 74, 103228. [CrossRef]
- Mattos, E. R., Singh, M., Cabrera, M. L., Das, K. C., & Program, B. and C. C. (2012). Effects of Inoculum Physiological Stage on the Growth Characteristics of Chlorella sorokiniana Cultivated Under Different CO2 Concentrations. Applied Biochemistry and Biotechnology, 168(3), 519–530. [CrossRef]
- Mhetras, N., & Gokhale, D. (2025). Sustainable biodiesel production: importance of feedstock resources and production methods. In RSC Advances (Vol. 15, Issue 33, pp. 26739–26754). Royal Society of Chemistry. [CrossRef]
- Mondal, M., Khan, A. A., & Halder, G. (2021). Estimation of biodiesel properties based on fatty acid profiles of Chlamydomonas sp. BTA 9032 and Chlorella sp. BTA 9031 obtained under mixotrophic cultivation conditions. Biofuels.
- Naseema Rasheed, R., Pourbakhtiar, A., Mehdizadeh Allaf, M., Baharlooeian, M., Rafiei, N., Alishah Aratboni, H., Morones-Ramirez, J. R., & Winck, F. V. (2023). Microalgal co-cultivation-recent methods, trends in omic-studies, applications, and future challenges. Frontiers in Bioengineering and Biotechnology, 11, 1193424.
- Naveen, B. P., Sivapullaiah, P. V., & Sitharami, T. G. (2015). Effect of aging on the leachate characteristics from municipal solid waste landfill. 15th Asian Regional Conference on Soil Mechanics and Geotechnical Engineering, ARC 2015: New Innovations and Sustainability, 1940–1945. [CrossRef]
- Nicodemou, A., Konstantinou, D., & Koutinas, M. (2024). Enhanced biomass and lipid production from olive processing wastewater using Scenedesmus obliquus in a two-stage cultivation strategy under salt stress. Biochemical Engineering Journal, 205, 109290.
- Ogbonna, K. E., Ogbonna, J. C., Njoku, O. U., Yamada, K., & Suzuki, I. (2025). Growth characteristics and biodiesel properties of fatty acids methyl esters (FAMEs) of Scenedesmus dimorphus (Turpin) Kützing. Biofuels, 1–12.
- Otim, F. N., Chen, I. R., & Otim, O. (2021). Indirect assessment of biomass accumulation in a wastewater-based Chlorella vulgaris photobioreactor by pH variation. Scientific Reports, 11(1). [CrossRef]
- Patnaik, R., & Mallick, N. (2021). Microalgal Biodiesel Production: Realizing the Sustainability Index. In Frontiers in Bioengineering and Biotechnology (Vol. 9). Frontiers Media S.A. [CrossRef]
- Potts, M. (2000). Nostoc. The Ecology of Cyanobacteria (Whitton BA & Potts M, eds). Kluwer Academic Publishers, Dordrecht, The Netherlands.
- Rafa, N., Ahmed, S. F., Badruddin, I. A., Mofijur, M., & Kamangar, S. (2021). Strategies to Produce Cost-Effective Third-Generation Biofuel From Microalgae. In Frontiers in Energy Research (Vol. 9). Frontiers Media SA. [CrossRef]
- Rajendran, D. S., Venkatraman, S., Rahul, R., Afrrin, M., Karthik, P., & Vaidyanathan, V. K. (2024). Manifesting Sustainability Toward Food Waste into Bioenergy: Biorefinery in a Circular Economic Approach. In G. Baskar, V. Ashokkumar, S. L. Rokhum, & V. S. Moholkar (Eds.), Circular Bioeconomy Perspectives in Sustainable Bioenergy Production (pp. 431–460). Springer Nature Singapore. [CrossRef]
- Rulli, M. C., Bellomi, D., Cazzoli, A., De Carolis, G., & D’Odorico, P. (2016). The water-land-food nexus of first-generation biofuels. Scientific Reports, 6. [CrossRef]
- Sayago, U. F. C., Gómez-Caicedo, M. I., & Mercado Suárez, Á. L. (2024). Design of a sustainable system for wastewater treatment and generation of biofuels based on the biomass of the aquatic plant Eichhornia Crassipes. Scientific Reports, 14(1). [CrossRef]
- Segredo-Morales, E., González-Martín, C., Vera, L., & González, E. (2023). Performance of a novel rotating membrane photobioreactor based on indigenous microalgae-bacteria consortia for wastewater reclamation. Journal of Industrial and Engineering Chemistry, 119, 586–597. [CrossRef]
- Shahzad, K., & Iqbal Cheema, I. (2024). Low-carbon technologies in automotive industry and decarbonizing transport. Journal of Power Sources, 591, 233888. [CrossRef]
- Tossavainen, M., Nykänen, A., Valkonen, K., Ojala, A., Kostia, S., & Romantschuk, M. (2017). Culturing of Selenastrum on diluted composting fluids; conversion of waste to valuable algal biomass in presence of bacteria. Bioresource Technology, 238, 205–213. [CrossRef]
- Staff, B. P. (2025, August 7). Global momentum in wastewater treatment sparks new opportunities for innovation. BCC Research LLC. https://www.bccresearch.com/pressroom/env/global-momentum-in-wastewater-treatment.
- Un-Water, & Un-Water. (2025, June 16). Progress on wastewater treatment – 2024 Update UN-Water. UN-Water. https://www.unwater.org/publications/progress-wastewater-treatment-2024-update.
- Udayan, A., Pandey, A. K., Sirohi, R., Sreekumar, N., Sang, B.-I., Sim, S. J., Kim, S. H., & Pandey, A. (2023). Production of microalgae with high lipid content and their potential as sources of nutraceuticals. Phytochemistry Reviews, 22(4), 833–860. [CrossRef]
- Wang, J., Zhou, W., Chen, H., Zhan, J., He, C., & Wang, Q. (2019). Ammonium nitrogen tolerant Chlorella strain screening and its damaging effects on photosynthesis. Frontiers in Microbiology, 10(JAN). [CrossRef]
- Wang, W., & Khanna, M. (2023). Land use effects of biofuel production in the US. Environmental Research Communications, 5(5). [CrossRef]
- Wu, X., Jin, C., Zhang, C., Li, P., Huang, J. J., Wu, J., Wu, J., & Hu, Z. (2025). Mixotrophic Chlorella pyrenoidosa biofilm with enhanced biomass production, microalgal activity, and nutrient removal from nutrient-rich wastewater. Journal of Environmental Sciences, 157, 366–377. [CrossRef]
- Yan, X., Shan, S., Li, X., Xu, Q., Yan, X., Ruan, R., & Cheng, P. (2024). Carbon and energy metabolism for the mixotrophic culture of Chlorella vulgaris using sodium acetate as a carbon source. Frontiers in Microbiology, 15. [CrossRef]
- Yun, H. S., Kim, Y. S., & Yoon, H. S. (2021). Effect of Different Cultivation Modes (Photoautotrophic, Mixotrophic, and Heterotrophic) on the Growth of Chlorella sp. and Biocompositions. Frontiers in Bioengineering and Biotechnology, 9. [CrossRef]
- Zhang, S., Zhang, L., Xu, G., Li, F., & Li, X. (2022). A review on biodiesel production from microalgae: Influencing parameters and recent advanced technologies. In Frontiers in Microbiology (Vol. 13). Frontiers Media S.A. [CrossRef]
- Zhang, Z., Sun, D., Cheng, K. W., & Chen, F. (2021). Investigation of carbon and energy metabolic mechanism of mixotrophy in Chromochloris zofingiensis. Biotechnology for Biofuels, 14(1). [CrossRef]
- Zheng, M., Li, H., Guo, X., Chen, B., & Wang, M. (2023). A semi-continuous efficient strategy for removing phosphorus and nitrogen from eel aquaculture wastewater using the self-flocculating microalga Desmodesmus sp. PW1. Journal of Environmental Management, 346, 118970. [CrossRef]
- Staff, B. P. (2025, August 7). Global momentum in wastewater treatment sparks new opportunities for innovation. BCC Research LLC. https://www.bccresearch.com/pressroom/env/global-momentum-in-wastewater-treatment.
- Un-Water, & Un-Water. (2025, June 16). Progress on wastewater treatment – 2024 Update UN-Water. UN-Water. https://www.unwater.org/publications/progress-wastewater-treatment-2024-update.
Figure 1.
Growth profiles of Desmodesmus sp. under varying leachate dilutions (25%, 50%, 75%, 100% v/v) compared with BBM control over 20 days, measured as optical density.
Figure 1.
Growth profiles of Desmodesmus sp. under varying leachate dilutions (25%, 50%, 75%, 100% v/v) compared with BBM control over 20 days, measured as optical density.
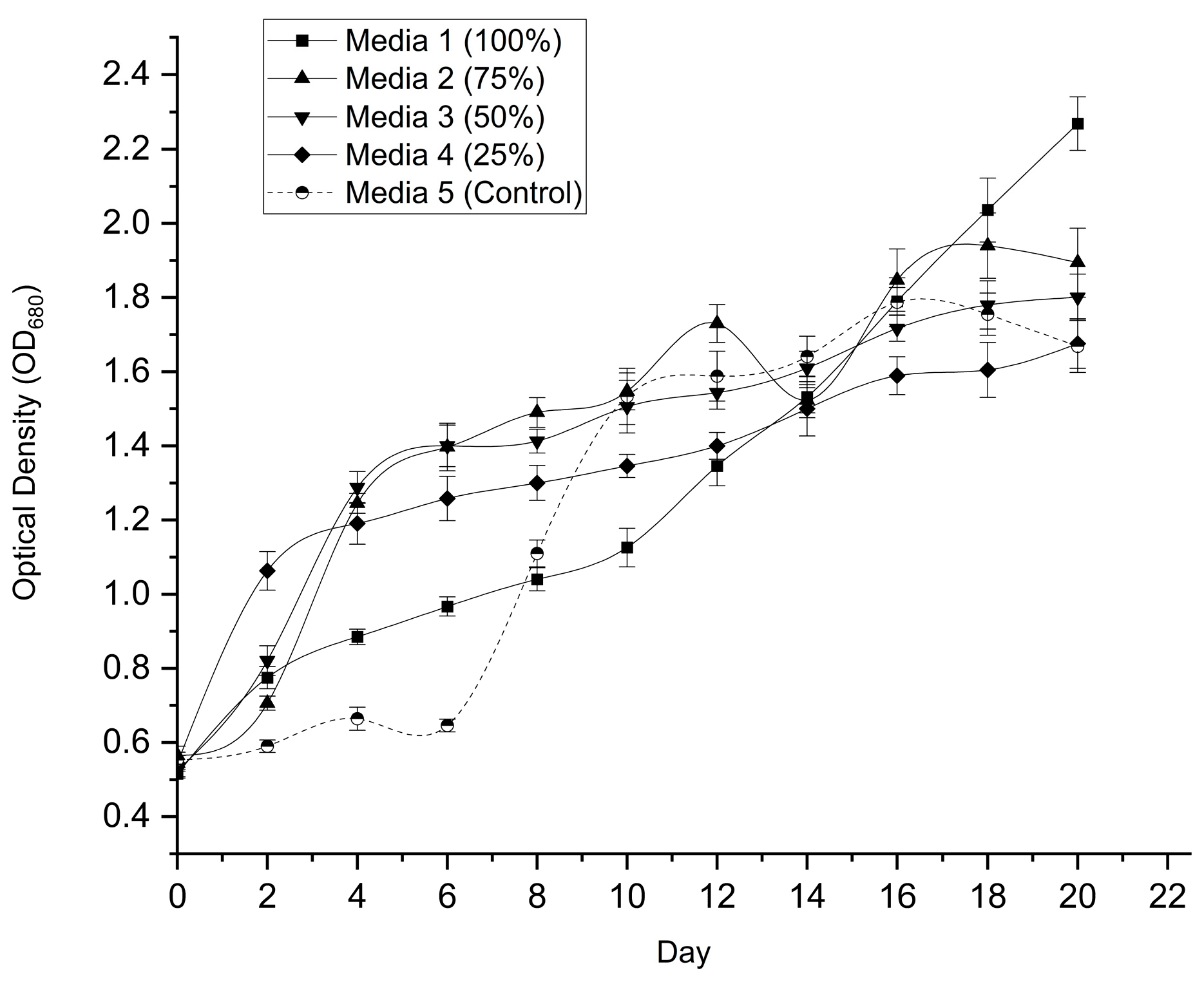
Figure 2.
Dry biomass accumulation of Desmodesmus sp. across different leachate dilutions and BBM control after 20 days of cultivation.
Figure 2.
Dry biomass accumulation of Desmodesmus sp. across different leachate dilutions and BBM control after 20 days of cultivation.
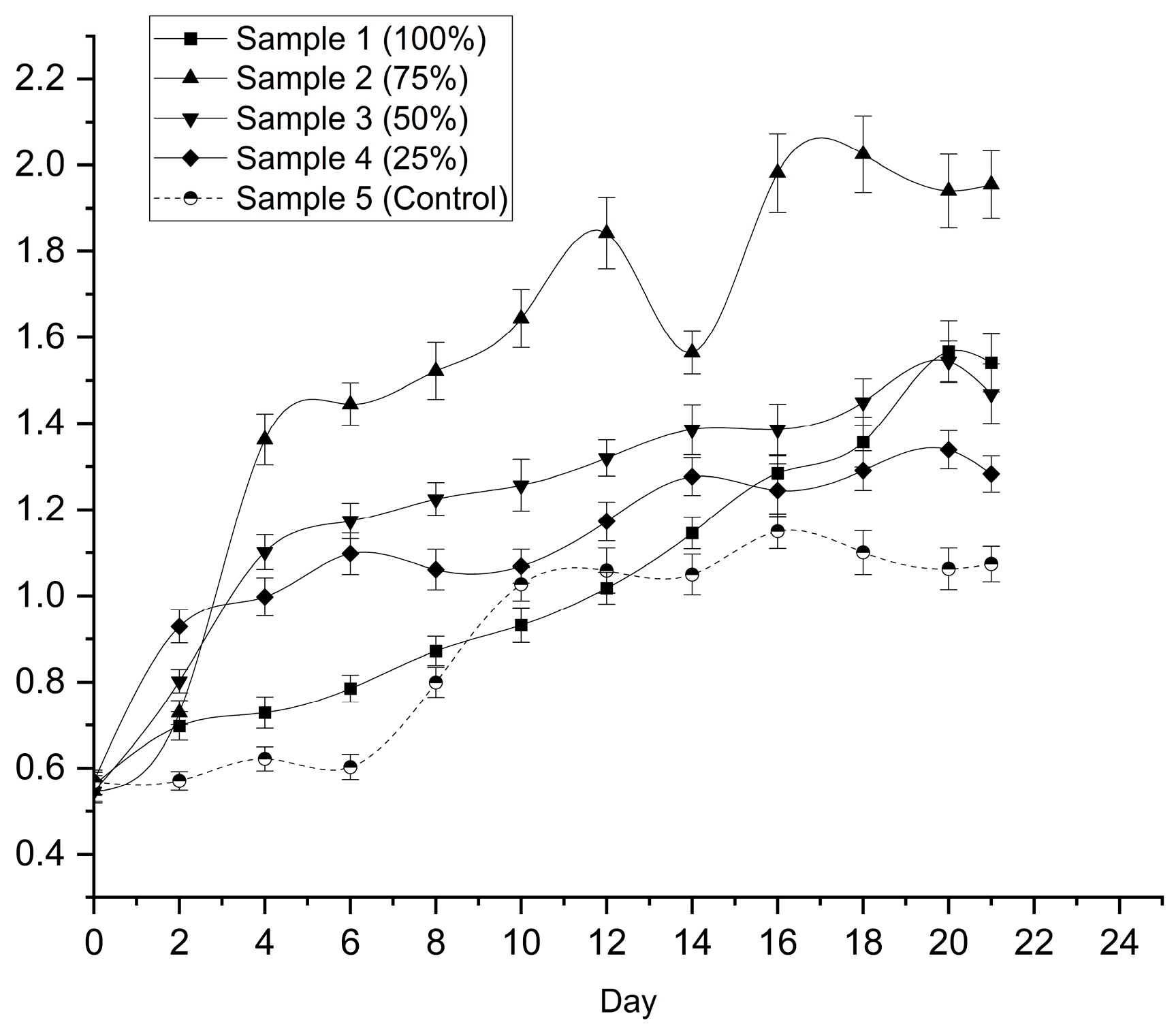
Figure 3.
Optical density-based growth kinetics of Desmodesmus sp. in undiluted matured compost leachate and BBM control over a 24-day batch cultivation period.
Figure 3.
Optical density-based growth kinetics of Desmodesmus sp. in undiluted matured compost leachate and BBM control over a 24-day batch cultivation period.
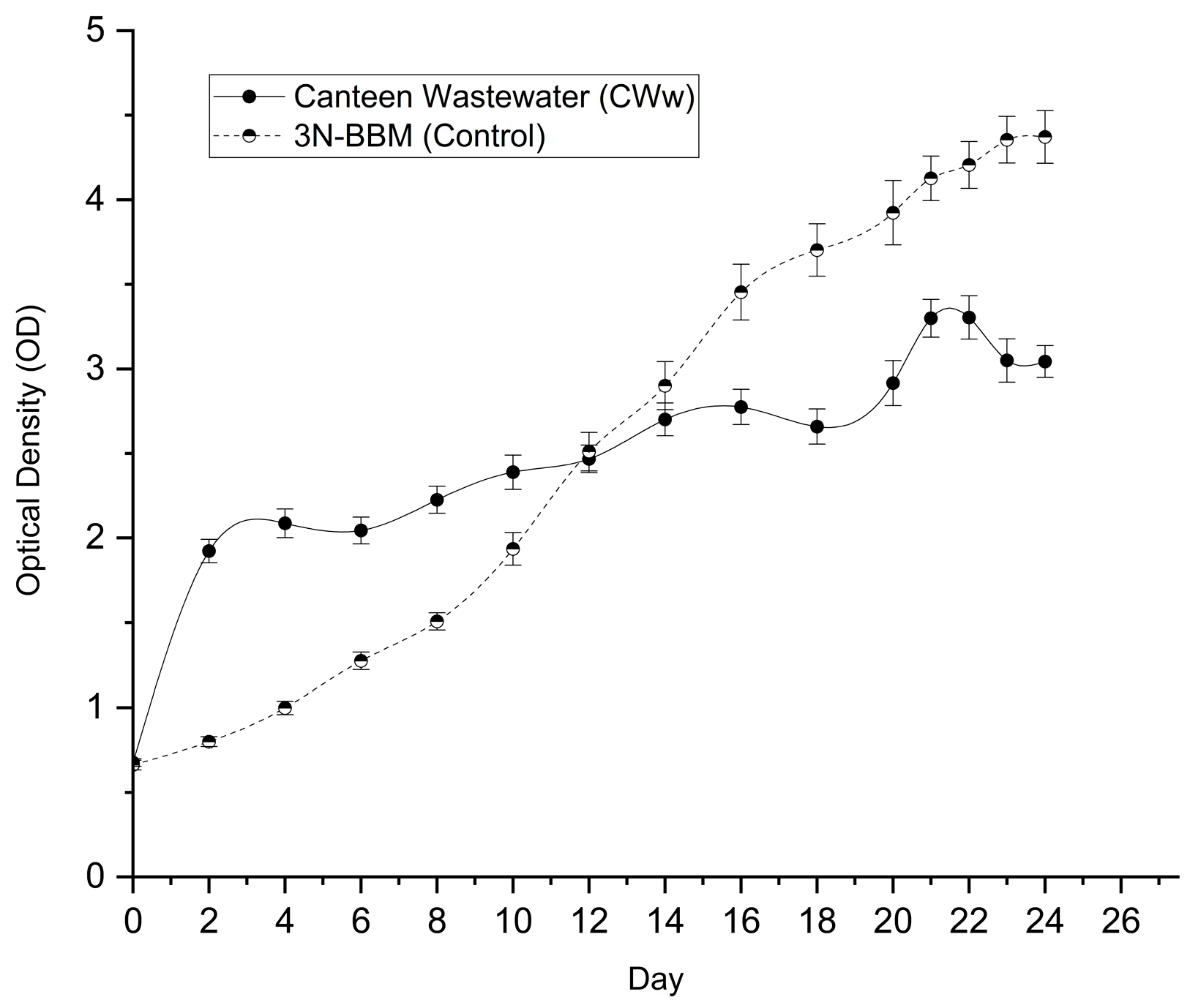
Figure 4.
Dry cell weight (DCW) of Desmodesmus sp. in matured compost leachate versus BBM control during 24 days of batch cultivation.
Figure 4.
Dry cell weight (DCW) of Desmodesmus sp. in matured compost leachate versus BBM control during 24 days of batch cultivation.
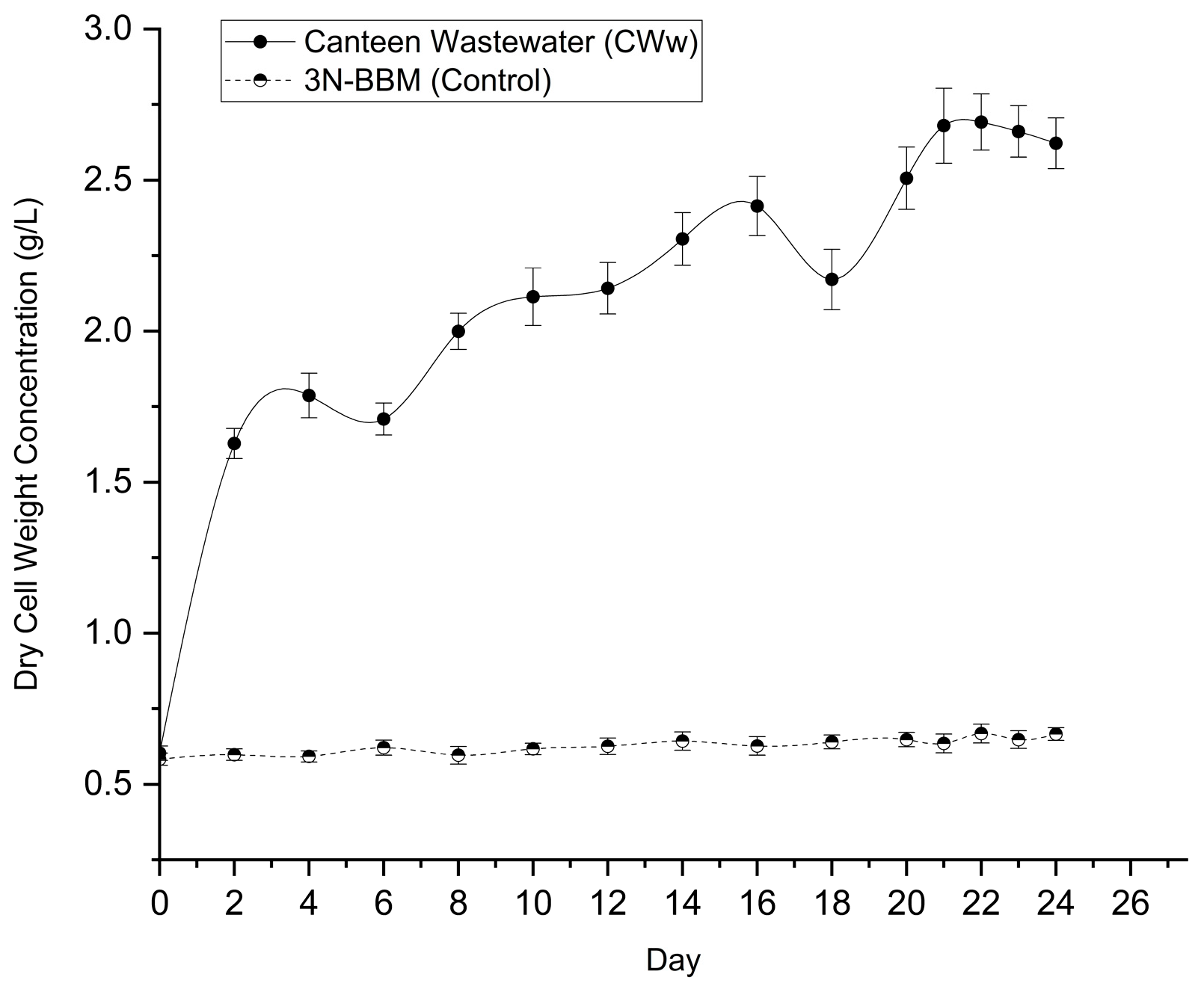
Figure 5.
Total nitrogen (TN) removal by Desmodesmus sp. from matured compost leachate and BBM control over the cultivation period.
Figure 5.
Total nitrogen (TN) removal by Desmodesmus sp. from matured compost leachate and BBM control over the cultivation period.
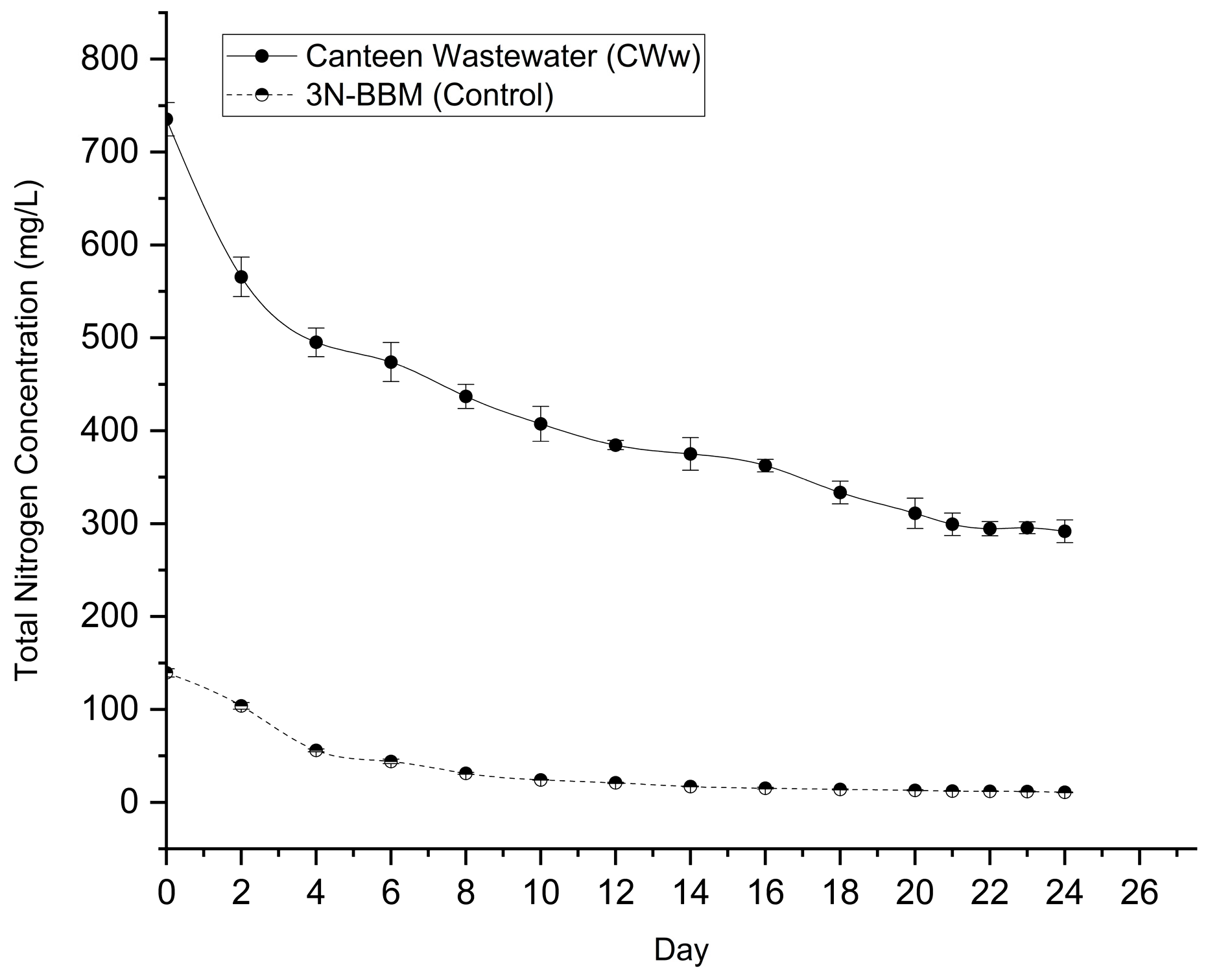
Figure 6.
Total phosphorus (TP) removal by Desmodesmus sp. from matured compost leachate and BBM control during 24-day cultivation.
Figure 6.
Total phosphorus (TP) removal by Desmodesmus sp. from matured compost leachate and BBM control during 24-day cultivation.
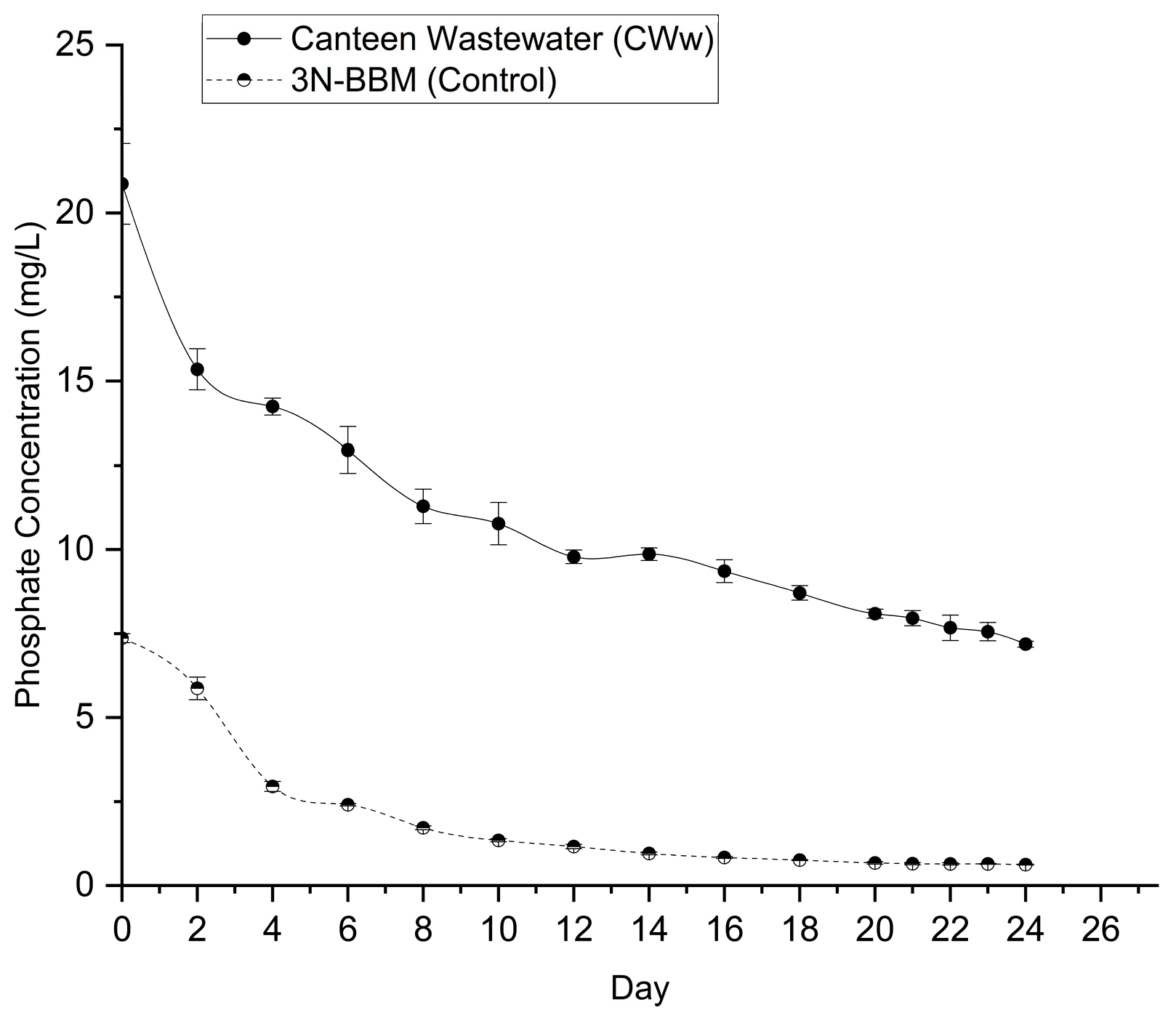
Figure 7.
Chemical Oxygen Demand (COD) reduction in matured compost leachate during cultivation of Desmodesmus sp.
Figure 7.
Chemical Oxygen Demand (COD) reduction in matured compost leachate during cultivation of Desmodesmus sp.
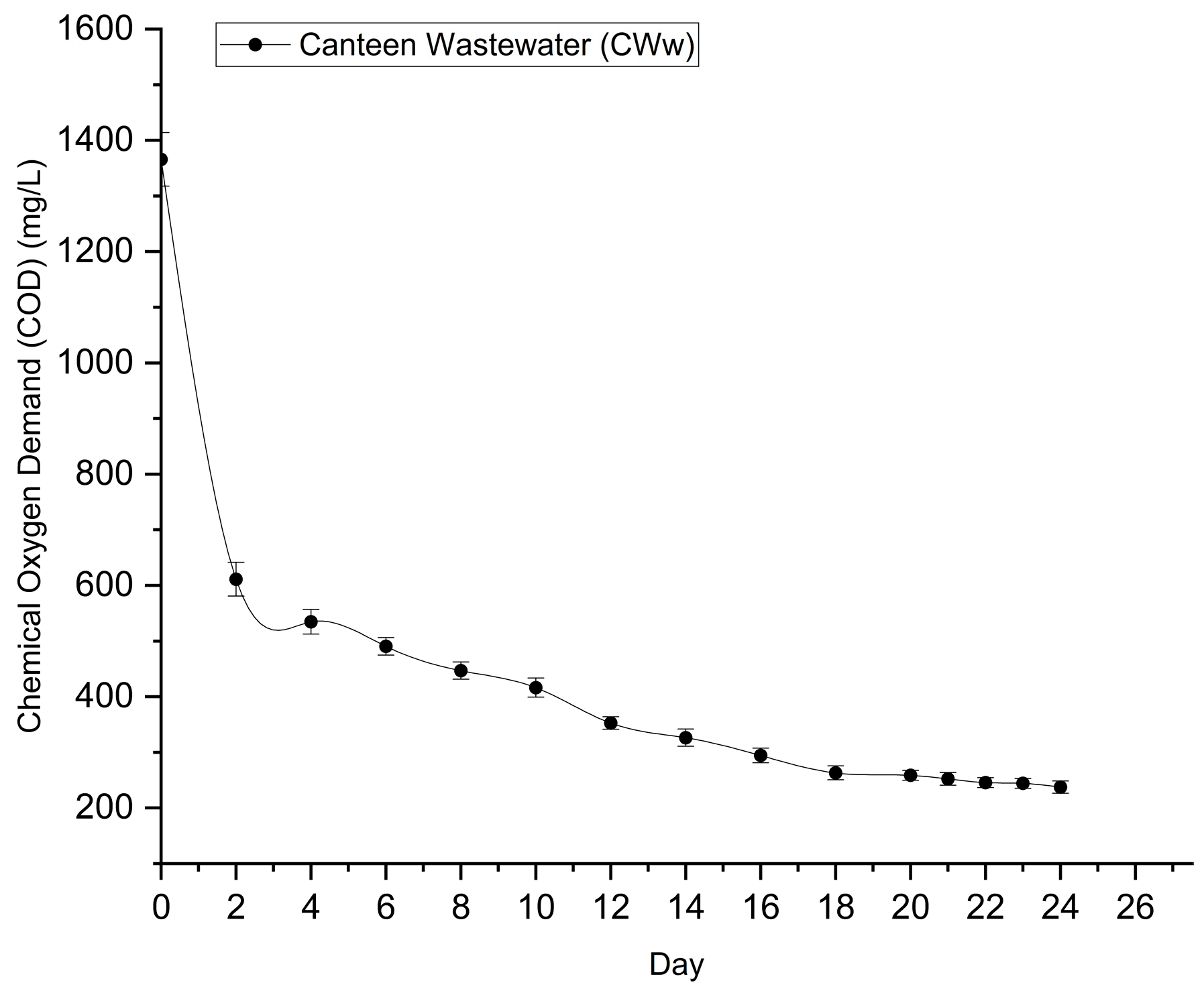
Figure 8.
pH evolution of matured compost leachate and BBM control during the cultivation of Desmodesmus sp.
Figure 8.
pH evolution of matured compost leachate and BBM control during the cultivation of Desmodesmus sp.
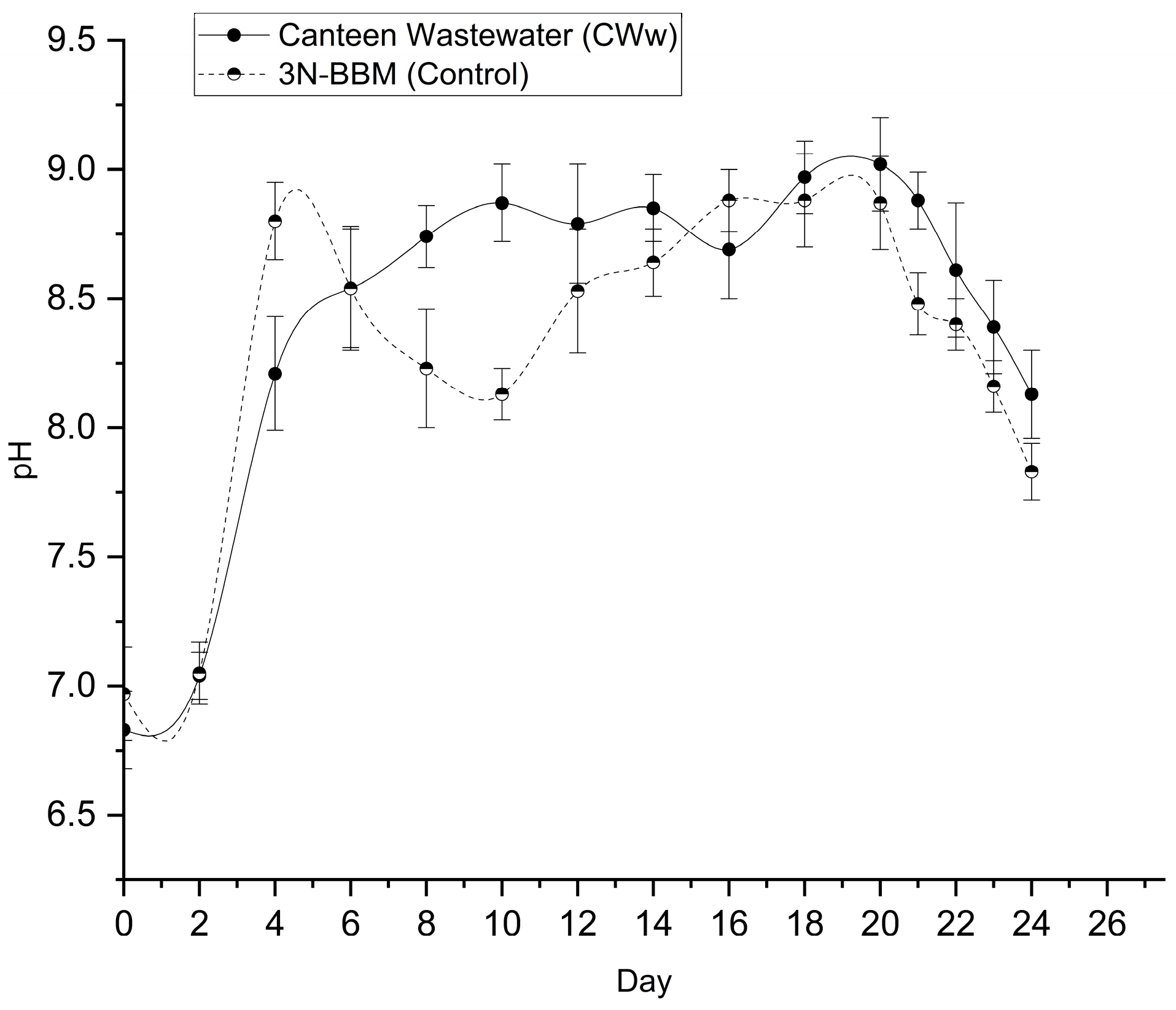
Figure 9.
Lipid accumulation (g) of Desmodesmus sp. in matured compost leachate and BBM control from Day 18 to Day 23.
Figure 9.
Lipid accumulation (g) of Desmodesmus sp. in matured compost leachate and BBM control from Day 18 to Day 23.
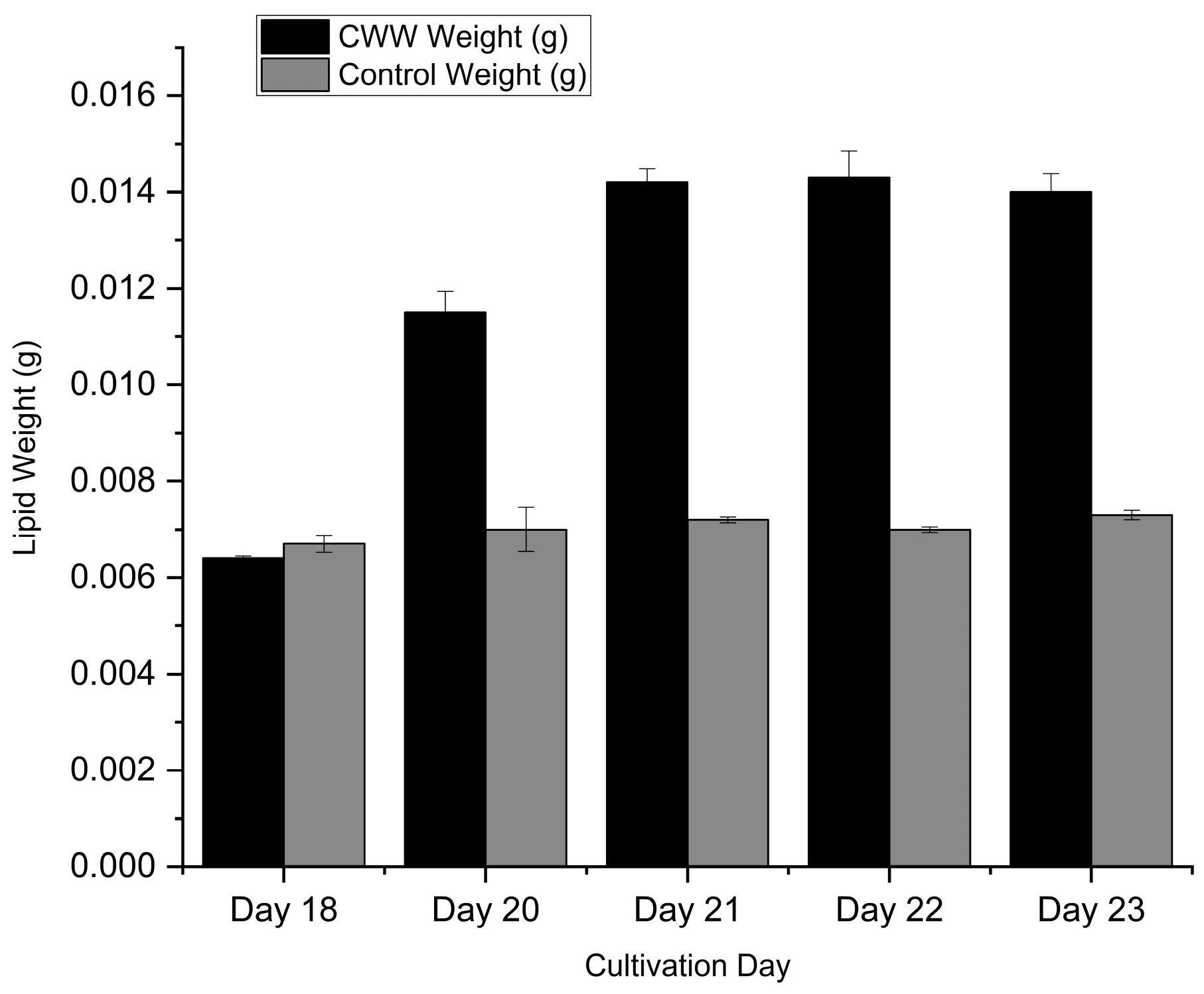
Figure 10.
Lipid concentration (g L⁻¹) of Desmodesmus sp. in matured compost leachate versus BBM control during stationary phase (Days 18–23).
Figure 10.
Lipid concentration (g L⁻¹) of Desmodesmus sp. in matured compost leachate versus BBM control during stationary phase (Days 18–23).
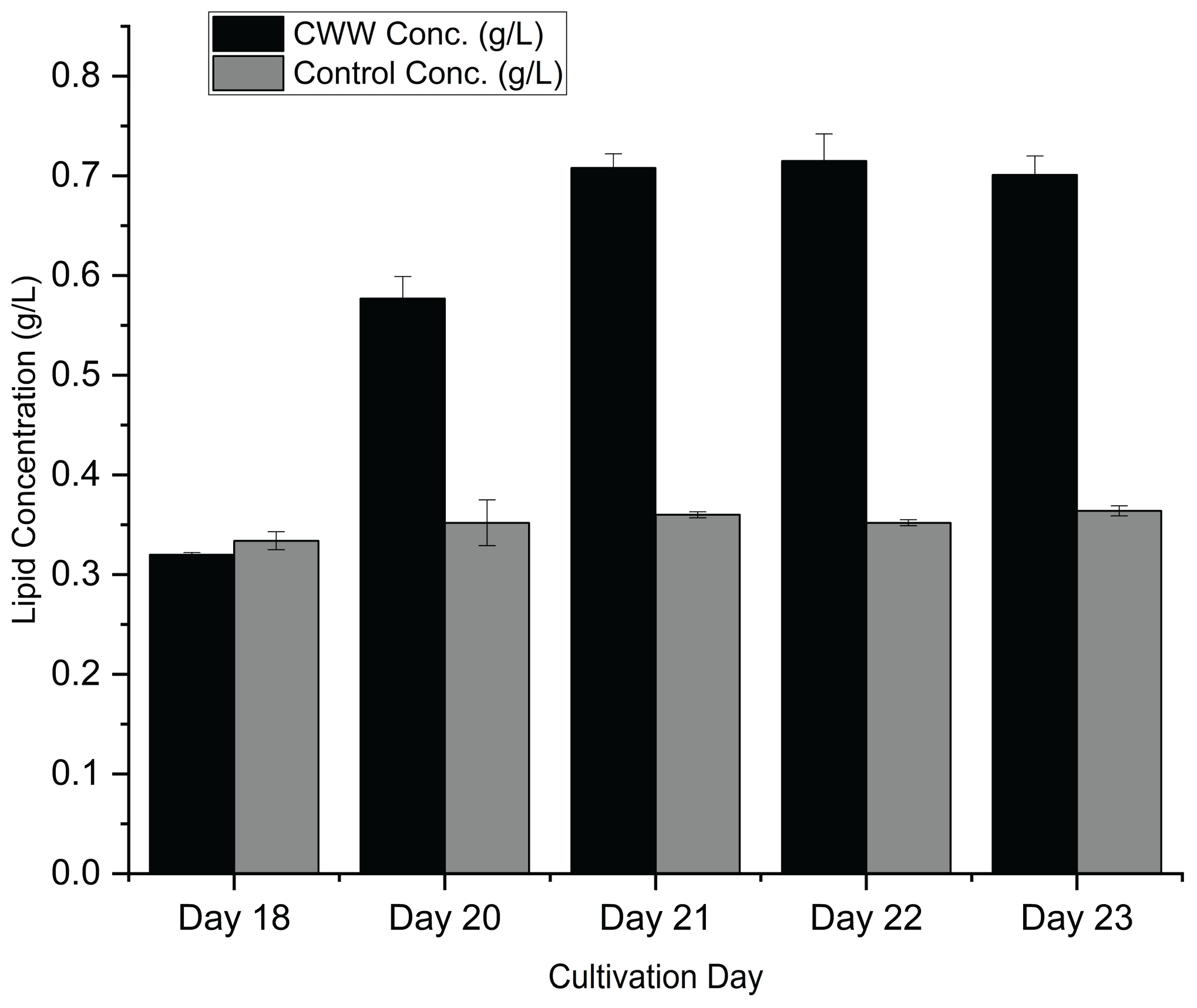

Table 1.
Physicochemical characterization of raw matured compost leachate wastewater used in the study (n = 3).
Table 1.
Physicochemical characterization of raw matured compost leachate wastewater used in the study (n = 3).
| Parameter | Unit | Value (Mean ± SD) |
|---|---|---|
| pH | - | 6.83 ± 0.15 |
| Chemical Oxygen Demand (COD) | mg L⁻¹ | 1365.89 ± 48.19 |
| Total Nitrogen (TN) | mg L⁻¹ | 735.29 ± 17.87 |
| Total Phosphorus (TP) | mg L⁻¹ | 20.87 ± 1.20 |
| Appearance | - | Dark brown, Turbid |
Table 3.
Fatty acid profile of Desmodesmus sp. biomass cultivated in matured compost leachate (normalized % of total FAMEs).
Table 3.
Fatty acid profile of Desmodesmus sp. biomass cultivated in matured compost leachate (normalized % of total FAMEs).
| Fatty Acid | Carbon Structure | Relative Abundance (%) |
|---|---|---|
| Caprylic acid | C8:0 | 4.55 |
| Capric acid | C10:0 | 3.86 |
| Lauric acid | C12:0 | 25.78 |
| Myristic acid | C14:0 | 9.39 |
| Palmitic acid | C16:0 | 26.95 |
| Stearic acid | C18:0 | 5.11 |
| Oleic acid | C18:1 | 20.46 |
| Linoleic acid | C18:2 | 3.90 |
| Total Saturated | - | 75.64 |
| Total Unsaturated | - | 24.36 |
Table 4.
Predicted fuel properties of Desmodesmus sp. biodiesel compared to international standards.
Table 4.
Predicted fuel properties of Desmodesmus sp. biodiesel compared to international standards.
| Biodiesel Property | Unit | This Study (Leachate) | ASTM D6751 | EN 14214 |
|---|---|---|---|---|
| Iodine Value | g I₂/100g | 25.45 | - | < 120 |
| Kinematic Viscosity | mm²/s | 3.42 | 1.9–6.0 | 3.5–5.0 |
| Higher Heating Value | MJ/kg | 38.36 | - | - |
| Density | g/cm³ | 0.85 | - | 0.86–0.90 |
| Saponification Value | mg KOH/g | 228.5 | - | - |
| Cetane Number | - | 64.5 | > 47 | > 51 |
| Oxidative Stability | h | 32.86 | > 3 | > 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



