
Submitted:
20 January 2026
Posted:
21 January 2026
You are already at the latest version
Abstract
The Qinghai–Tibet Plateau and surrounding montane valleys comprise one of the world’s most pronounced freshwater environmental gradients, where cold, intense UV radiation, variable dissolved oxygen, and heterogeneous hydrodynamics interact with drainage reorganization and connectivity constraints. Schizothoracinae are among the most representative endemic lineages in these systems, combining exceptional lineage diversity with pervasive polyploid genomic backgrounds. Here we synthesize schizothoracine research through an “environment–evolution–conservation” framework, linking (i) taxonomic and phylogenetic foundations under pervasive convergence, cryptic diversity, and hybridization; (ii) geologic history and drainage evolution as drivers of radiation and gene exchange; (iii) polyploidy, post-WGD structural remodeling, and early rediploidization as a testable process rather than a static ploidy fact; and (iv) omics resources and analytical pipelines that enable verifiable evidence chains across comparative genomics, population genomics, and tissue-level stress responses. Across major stressors, recurrent molecular themes emerge for cold-associated metabolic remodeling and UV-associated DNA repair and genome maintenance, whereas hypoxia-related signals are often inconsistent, plausibly reflecting strong spatiotemporal heterogeneity and multi-route physiological accommodation. We further connect molecular candidates to functional outcomes using reusable phenotypic evidence streams, including geometric morphometrics, high-throughput phenotyping, otolith microchemistry, and age–growth life-history syndromes of slow growth and longevity. Finally, we translate population structure into operational MU/ESU delineation and propose an auditable, iterative management checklist centered on MU-aligned stocking, connectivity performance metrics during critical seasonal windows, and quantified habitat restoration targets. We conclude by outlining priorities to raise evidentiary strength across basins: chromosome-level genomes across lineages, systematic SV/TE comparisons, standardized stressor–phenotype–transcriptome designs, and intensified sampling in geomorphic transition zones and putative hybrid regions to enable cumulative, decision-ready synthesis.
Keywords:
Schizothoracinae
; Qinghai–Tibet plateau
; drainage evolution
; polyploidy
; rediploidization
; population genomics
; management units/evolutionarily significant unit (MU/ESU)
; conservation management
1. Introduction
The Qinghai–Tibet Plateau and its surrounding mountain valleys constitute one of the world’s most representative freshwater environmental gradients. This gradient spans low temperature, reduced dissolved oxygen, and intense ultraviolet radiation associated with increasing elevation [1], as well as torrential plateau rivers characterized by strong hydrodynamics, pronounced spatial heterogeneity in flow, and marked seasonal fluctuations. In addition, some semi-enclosed lakes or atypical water bodies may experience salinization/alkalinization, shifts in ionic composition, and nutrient limitation [2,3]. Together, these factors impose long-term “combined stresses” on aquatic organisms [4]. Unlike terrestrial systems, adaptation in freshwater fishes is not determined by a single driver (e.g., hypoxia) alone; it is simultaneously constrained by the physical structure of water bodies, hydrochemical properties, and changes in river-network connectivity and drainage configuration [5,6]. Consequently, plateau river–lake systems provide a natural framework for linking environmental gradients to ecological function and, ultimately, to genetic mechanisms, making the taxa inhabiting these waters ideal models for understanding adaptation and diversification under extreme conditions.
In such plateau–montane freshwater environments, the subfamily Schizothoracinae is among the most representative endemic lineages. It is distributed mainly across the Qinghai–Tibet Plateau and adjacent river–lake systems [7], and has generated rich species and lineage diversity in multiple drainages of Sichuan [8], Yunnan [9], and Tibet [1]. This group not only exhibits a continuous morphological gradient from “primitive” to “highly specialized” forms [10,11], but—more importantly—shows pervasive polyploid backgrounds at the genomic level, making it a natural system for investigating the coupling among environmental pressures, genome evolution, and phenotypic adaptation [12]. In other words, schizothoracines are not merely “fishes living at high elevation”; rather, they represent a subfamily that has diversified under long-term, multifactor constraints in aquatic environments and evolved distinctive adaptive features. Accordingly, they provide a key entry point for elucidating the origins of biodiversity in plateau waters, freshwater adaptation to extreme environments, and the evolutionary trajectories of polyploid genomes.
In recent years, research on schizothoracines has shifted from primarily descriptive accounts toward more mechanistic explanations, and three interlocking lines of inquiry have gradually taken shape. First, episodic uplift of the Qinghai–Tibet Plateau and its surrounding regions, Quaternary glacial–interglacial fluctuations, and drainage reorganizations provide a clear historical context for lineage divergence and geographic patterning in schizothoracines, making them a classic aquatic model for examining how geological events shape biodiversity [13,14]. Second, their polyploid and repeat-rich genomic architecture confers unique value for investigating post-WGD structural remodeling and early re-diploidization processes [1,15]. Third, under the joint influence of multidimensional environmental gradients in plateau waters, schizothoracines exhibit a suite of testable adaptive themes that ultimately manifest as functional outcomes at the levels of morphology and behavioral ecology [16,17,18]. Meanwhile, hydropower development, overfishing, and habitat fragmentation are accelerating population declines in some systems, inevitably coupling schizothoracine research with conservation biology and management practice [19]. Thus, beyond asking how these fishes evolved and adapted, we must also identify which populations are most vulnerable and what interventions are most likely to be effective. This context motivates a synthetic framework that links historical drivers to genomic mechanisms, and further to phenotypic function and conservation applications.
Across the current literature, schizothoracine research is increasingly assembling a continuous chain spanning data generation, mechanistic inference, phenotypic interpretation, and application. Omics resources have expanded from mitochondrial markers to reference genomes, transcriptomes, and population genomics datasets [10,20]. Mechanistic hypotheses are commonly derived from signals of selection and functional enrichment that highlight candidate genes or pathways [16]. Phenotypic and ecological evidence encompasses functional levels such as morphological differentiation, life-history strategies, and behavioral ecology [18]. Finally, escalating human pressures are pushing these insights toward practical outputs, including the delineation of conservation units and the design of engineering and management actions [21].
Building on the above context and key knowledge gaps, this review adopts an “environment–evolution–conservation” framework to synthesize evidence in sequence, from plateau environmental gradients to geological history and drainage evolution, taxonomic and phylogenetic foundations, polyploidy and early re-diploidization, and the omics resources and methodological advances that underpin mechanistic inference. We further link shared molecular themes to functional outcomes in morphology, life history, and behavioral ecology, and—within a population genetic and conservation-unit (MU/ESU) perspective—evaluate anthropogenic pressures and management practices to propose priorities for future research and conservation (Figure 1).
2. Taxonomy and Phylogenetic Framework: Species Delimitation, Lineage Structure, and Geographic Patterns
2.1. Traditional Morphological Classification and Its Controversies
For decades, the subfamily Schizothoracinae has been classified and grouped primarily using external morphology and osteological characters, including oral morphology, barbels, pharyngeal teeth, squamation, and skeletal structures; on this basis, a continuum ranging from “primitive” to “specialized” and “highly specialized” forms has been proposed [27,28,29]. However, this morphology-based approach faces two systematic challenges. First, convergent and parallel adaptation is widespread in plateau river–valley and lake environments, and many traits associated with hydrodynamics, substrate use, and feeding ecology recur across different lineages, meaning that “morphological similarity” does not necessarily imply close relatedness [11,30]. Second, cryptic species, species complexes, and natural hybridization commonly coexist in schizothoracines, producing continuous or intermediate phenotypic variation that weakens the stability of diagnostic boundaries and key identification traits [31]. Consequently, morphology alone often fails to meet the requirements for delimitation that is simultaneously diagnosable, repeatable, and comparable, and it further amplifies the dependence of subsequent biogeographic and adaptive inferences on a stable taxonomic framework.
2.2. Molecular Phylogenetics and DNA Barcoding
To improve the repeatability of species identification and phylogenetic inference, mitochondrial markers such as COI and Cytb have been widely used in schizothoracines for species recognition, lineage delimitation, and the description of geographic patterns, providing an efficient solution when sample sizes are large and budgets are limited [32,33]. Building on this foundation, complete mitochondrial genomes offer higher resolution by adding many more informative sites and enabling standardized comparisons of genome organization, thereby refining inferences on intrageneric relationships, subclade divergence, and biogeographic scenarios. For example, a study of snow trout (the genus Schizothorax) used mitogenomes together with barcode-distance evidence to identify putative independent species and suggested that historical gene flow or mitochondrial capture may have occurred among closely related taxa [34]. In addition, the publication of mitogenomes for species such as S. macropogon [35] and S. kozlovi [36] has helped establish a more comprehensive mtDNA reference framework for subsequent work.
Despite these advantages, mitogenomes still represent a single maternally inherited linkage unit and are therefore susceptible to introgression, mitochondrial replacement, and incomplete lineage sorting (ILS). Discordance between mtDNA trees and species trees is not uncommon, particularly under rapid radiations or persistent gene flow [22,37,38]. Accordingly, mtDNA in schizothoracine research more often serves as a tool for rapid identification and preliminary lineage assignment, whereas robust species delimitation and species-tree inference ultimately require evidence from nuclear genomic data.
2.3. Multi-Locus Data, Coalescent Models, and Gene Flow
To address biases inherent to mtDNA and the uncertainty of single-gene trees, schizothoracine studies have increasingly adopted multilocus frameworks that combine mitochondrial and nuclear loci, and have introduced coalescent-based models to infer divergence history and gene exchange [39]. In the Nujiang (Salween) schizothoracine species complex, for instance, researchers complemented Cytb with nuclear markers (RAG-1/RAG-2) and used haplotype networks, population differentiation statistics, and phylogenetic reconstructions to characterize lineage structure. They further applied coalescent approaches such as MDIV and IMa2 to estimate divergence times and migration rates, and to evaluate the relative contributions of incomplete lineage sorting (ILS) versus recent gene flow to observed gene-tree discordance [9]. In parallel, demographic reconstruction methods such as the extended Bayesian skyline plot (EBSP) can link lineage differentiation with population history, providing temporal context for hypotheses about divergence driven by geological and climatic events [40].
Overall, multilocus and coalescent frameworks have shifted schizothoracine phylogenetic research from merely “building trees” to “explaining trees”: beyond comparing topological differences, they allow explicit estimation of gene flow and divergence parameters, thereby yielding a more testable evidence chain for species delimitation, interpretation of distributional patterns, and the definition of conservation units.
2.4. Natural Hybridization and Species Boundaries
Species boundaries in schizothoracines are not static lines; instead, they often lie within a dynamic continuum of “divergence–secondary contact–hybridization/introgression.” A well-documented case from the middle Yarlung Tsangpo River involving natural hybridization between Schizothorax o’connori and S. waltoni shows that morphometric measurements, clustering/network-based approaches, and multivariate discriminant analyses can distinguish parental and hybrid individuals with relatively high accuracy. Importantly, this performance is typically achieved under specific conditions where parental sources are well defined, the hybrid zone is known, and sympatric samples provide an internal reference for comparison. In such settings, hybrids frequently exhibit intermediate phenotypes between the parental forms, with certain traits biased toward one parent, suggesting that hybridization may be associated with local niche differentiation (e.g., resource use and spatial habitat partitioning) [31].
These cases also highlight a practical boundary of morphology- or mtDNA-based inference. When hybrid risk is high or divergence is shallow—especially in river reaches with ongoing contact—morphology and mtDNA alone often cannot reliably separate parental individuals from backcrosses or introgressed populations. Consequently, high-resolution nuclear genomic data (e.g., SNP panels, RAD/GBS, or whole-genome resequencing) become essential for inferring the directionality and extent of introgression and for assessing the strength of genome-wide reproductive isolation. Such evidence is directly consequential for taxonomic revision and for making MU/ESU delineation operational and defensible in management contexts [41,42].
3. Geologic History and Drainage Evolution as Drivers of Radiation: From “Plateau Uplift” to “Lineage Divergence”
3.1. How Episodic Uplift, Glaciations, and Canyon Formation Shape Schizothoracine Divergence
The geological evolution of the Qinghai–Tibet Plateau and surrounding mountain systems provides a relatively clear external framework for schizothoracine lineage diversification. Episodic tectonic uplift and river incision continuously modify divide elevations, valley gradients, and hydrodynamic regimes, creating a shifting spatial mosaic of “isolation versus connectivity” across the landscape [43]. On the one hand, canyon incision and deep channel downcutting can fragment formerly continuous habitats into functionally semi-independent units, reducing gene exchange and promoting allopatric divergence [44]. On the other hand, connectivity may be re-established intermittently in geomorphic transition zones or near tributary confluences, facilitating secondary contact and introgression [45].
Meanwhile, repeated Quaternary glacial–interglacial cycles altered glacial and hydrological conditions—such as discharge, water temperature, and the duration of ice cover—driving range contractions and expansions that leave composite genetic signatures of historical isolation and subsequent secondary contact [13,39]. In addition, ancient drainage reorganizations, including river capture, can reshape historical connectivity so that phylogeographic structure does not always align with modern basin boundaries: some inter-basin populations may be genetically closer to each other, whereas deeper divergence can occur among populations within the same present-day drainage [21].
3.2. A Representative Case: Geomorphic Development of the Yarlung Tsangpo–Parlung Tsangpo System and Patterns of Gene Exchange
Along the southern margin of the Qinghai–Tibet Plateau, the Yarlung Tsangpo River flows eastward from the plateau interior and forms deeply incised gorges at the Eastern Himalayan syntaxis. The evolution of this drainage network and associated geomorphic restructuring provides a clear geographic context for changes in basin connectivity. Schizothoracine fishes in this system show lineage structure broadly consistent with geomorphic barriers, making the Yarlung Tsangpo basin a useful natural setting to test the hypothesis that geomorphic reorganization alters connectivity and thereby shapes phylogeographic structure [39,46].
Mechanistically, tectonic activity and valley incision can reshape river passability by changing channel gradients and generating geomorphic elements such as knickpoints and gorges, which in turn reorganize hydrodynamic regimes [47]. These bottlenecks and high-velocity barriers can constrain longitudinal movement and effective dispersal distances, amplifying spatial differentiation at the river-reach scale [48]. In schizothoracines from the Yarlung Tsangpo system, pronounced lineage divergence can correspond to areas upstream versus downstream of the Grand Canyon, yet signatures consistent with connectivity or gene exchange can still be detected in some reaches, producing a mixed pattern of “strengthened divergence with localized exchange retained” [39].
Transitional zones—such as the middle reaches of the Yarlung Tsangpo—are more likely to host lineage contact and secondary contact events [49]. Records of sympatry and natural hybridization among closely related taxa in this region, including hybridization between Schizothorax o’connori and S. waltoni [31], suggest that species boundaries may be dynamic under episodically shifting connectivity. Isolation may be reinforced in certain periods or reaches, whereas introgression and hybridization windows may persist where tributary inputs and valley configuration maintain dispersal corridors [41,50]. Thus, within a single drainage, “lineage divergence” and “gene exchange” are not mutually exclusive; their relative prominence is jointly modulated by barrier strength, the location of transition zones, and the temporal scale considered.
Available evidence also supports a zoned interpretation that better reflects the complexity of this basin: headwater regions may retain older lineages and deeper divergence, transitional zones may more readily become lineage-intersection and hybrid-zone windows, and reaches with intensified canyon incision may more strongly enforce isolation and promote localized differentiation [31,51,52]. Under this framework, biogeographic inference for the Yarlung Tsangpo system is best constrained by jointly evaluating divergence times, dispersal directions, and the magnitude of gene flow, and cross-validating these inferences against reach-scale passability and the distribution of barriers—rather than relying on phylogenetic topology alone as a single-track explanation.
3.3. A Representative Case: The Tianshan–Pamir Corridor, Westward Dispersal, and Rapid Diversification
Relative to the major drainages of the Qinghai–Tibet Plateau, snow trouts in the Tianshan region provide a useful contrasting montane freshwater system. Geological evidence indicates pronounced uplift of the Tianshan since the Late Miocene, with subsequent acceleration, providing an external forcing background for regional landscape reorganization and drainage rearrangement [52].
Based on analyses of complete mitochondrial genomes together with COI barcodes, snow trouts can be divided into two major lineages, which appear to have undergone multiple episodes of stepwise divergence since the Late Miocene. In particular, S. eurystomus has been proposed to have dispersed westward via a “Pamir–Tianshan corridor,” contributing to the present-day distribution pattern [34]. The same study also suggests that potential interspecific gene exchange may occur among some lineages and emphasizes that mitochondria-only evidence can yield non-unique interpretations. Where gene flow is plausible, the direction of dispersal and the magnitude of introgression therefore require additional constraints and quantification using nuclear genomic and population-scale data [34].
4. Polyploidy and Genome Evolution: From Ploidy Patterns to Early Rediploidization
4.1. Ploidy Landscape and Core Questions
Against the broader backdrop of polyploidization in Cypriniformes, schizothoracines exhibit pronounced ploidy complexity, with tetraploid and even higher-ploidy lineages reported across multiple drainage systems [39,53,54]. Accordingly, the central aim of this chapter is not to re-establish whether polyploidization occurred, but to clarify how the timing and mode of genome doubling may differ among lineages, and to summarize the available evidence—together with its limits—on how post-duplication genomes move toward long-term stability through rediploidization and structural remodeling. At present, direct evidence in schizothoracines is still anchored in a small number of high-quality reference genomes and population datasets; many mechanistic propositions therefore remain testable hypotheses that require broader lineage sampling and higher-resolution data to substantiate.
4.2. “Young Tetraploids” and Early Rediploidization
Within current studies of polyploidy in schizothoracines, the tetraploid reference genome of Schizothorax o’connori serves as a key anchor. Using a single analytical framework, it not only provides an estimate of the time scale of whole-genome duplication (WGD), but also links post-duplication genome remodeling to early rediploidization, thereby yielding a traceable chain of evidence from genome doubling to restructuring and subsequent rediploidization [12,55]. Compared with inferences based on a small number of markers or transcriptomes, chromosome-scale assemblies allow self-alignment and synteny analyses to identify homeologous chromosomes and syntenic blocks, offering structurally testable evidence for “paired correspondences” expected under tetraploidy [56,57].
Importantly, “young tetraploid” does not mean “not yet rediploidized.” Within this temporal window, many homeologous gene pairs may still retain high sequence similarity, yet genomic divergence often proceeds in a regionally heterogeneous manner. Certain regions may show earlier restriction of recombination or accumulation of structural differences, and regulatory divergence can occur even earlier, manifesting as condition- or tissue-specific expression differences [58,59]. The particular value of S. o’connori lies in the fact that, among the few schizothoracine lineages with chromosome-scale evidence, early rediploidization can be followed more explicitly through convergent signals of structural correspondence, regionally uneven divergence, and regulatory partitioning—helping to move the discussion from the existence of polyploidy to how rediploidization unfolds in practice.
4.3. Evidence Framework for Polyploid Origins and Post-WGD Processes
Debates over the origin of polyploidy in schizothoracines typically center on two alternative hypotheses: autopolyploidization versus allopolyploidization [60]. At the genomic level, commonly used lines of evidence include (i) clustering patterns of homeologous genes in phylogenetic trees, (ii) the degree and symmetry of subgenome divergence, and (iii) lineage consistency in repeat-element families and their landscapes [15,61,62]. However, such inferences are sensitive to incomplete lineage sampling, gene flow, and incomplete lineage sorting (ILS). It is therefore more appropriate to frame this issue as an “evidence framework plus key gaps”: current genomic signals can provide directional clues, but decisive nodes still require joint constraints from cytogenetic evidence (e.g., karyotypes and pairing behavior) and population-scale data [63].
Similarly, processes such as transposable-element activity and structural variation, the evolutionary fates of duplicated genes, and meiotic stability are often regarded as closely tied to the progression of rediploidization [55]. In schizothoracines, however, systematic evidence that can be directly anchored to specific lineages and clearly defined gene sets remains concentrated in a small number of species and datasets. At present, what is most broadly reusable are the diagnostic criteria and analytical frameworks for interrogating these processes, whereas fine-grained functional interpretation will depend on denser and more phylogenetically comprehensive data in future work.
4.4. Comparison and Synthesis
Studies of other polyploid cyprinids show that rediploidization often proceeds in a regionally heterogeneous, “mosaic” manner: some chromosomal segments are more prone to structural rearrangements or homeologous exchange; transposable-element (TE) landscapes and homeolog divergence can be used to trace separation histories; and the retention and expression divergence of duplicated genes frequently exhibit functional biases [57,64]. What distinguishes schizothoracines is the combination of complex ploidy histories with strong environmental gradients and dynamic drainage reorganization. A key question therefore becomes whether the spatiotemporal trajectory of rediploidization has been jointly shaped by geological history, changes in basin connectivity, and persistent environmental pressures. Moving this proposition from a plausible inference to testable conclusions will require filling a central gap: chromosome-level genomes across multiple lineages paired with comparable population sampling, ideally allowing structural rearrangements and TE landscapes to be evaluated in the same coordinate framework as population divergence and gene-flow signals.
5. Omics Resources and Methodological Advances: From Data Accumulation to Testable Evidence Chains
5.1. Inventory of Available Resources
Growth in omics resources for schizothoracines has been driven first by the gradual formation of cross-species, cross-tissue data systems. The establishment of multi-species, multi-tissue transcriptome datasets has made it routine to compare expression patterns and functional annotations across lineages and tissues within a unified analytical framework, reducing dependence on any single dataset and improving reproducibility [10]. Building on this foundation, full-length transcriptomes (long-read sequencing) provide more reliable references for alternative splicing, lncRNAs, transcript structure, and SSR discovery, and are particularly useful for improving annotation and interpreting homeolog-specific expression under polyploid backgrounds [65]. Meanwhile, chromosome-level reference genomes (e.g., long reads + Hi-C + RNA-seq) have begun to emerge in schizothoracines and are being incorporated into downstream comparative analyses [12,66]. Their primary value is not simply “more genes,” but the ability to place synteny, homeologous correspondence, and repetitive/structural variation into a shared coordinate system, making structural inferences more directly verifiable.
At the population scale, whole-genome resequencing (WGS) and reduced-representation approaches (GBS/RAD) are also increasing, enabling questions that align closely with “progress-style” synthesis—such as population structure across river reaches and basins, genetic signatures of historical bottlenecks and connectivity shifts, and candidate adaptive signals associated with environmental gradients [16]. A persistent limitation, however, is the uneven distribution of available data: high-quality genomes and dense population sampling remain concentrated in a few representative species and a small number of basins, whereas several key lineages and geomorphology–drainage transition zones are still clearly under-sampled [7,10,67]. Accordingly, the focus of this chapter is not to be method-exhaustive, but to clarify which data types currently support which classes of conclusions, and to highlight the critical gaps that remain to be tested.
5.2. Key Analytical Pipelines
Chromosome-level genome studies in schizothoracines typically construct pseudochromosomes using long reads plus Hi-C, with transcript evidence used to refine gene annotation. On this basis, synteny analyses and duplication-aware metrics (e.g., Ks or 4dTv distributions) are used to anchor WGD events and homeologous correspondence at the chromosome scale, providing a reference for side-by-side comparisons of structure, expression, and selection signals within the same coordinate system [71]. Studies often then combine self-alignment and redundancy filtering (to identify duplicated or redundant segments) with synteny tools such as MCScan to extract syntenic blocks, thereby identifying paired duplicated regions and chromosome correspondences and generating reproducible structural evidence for polyploidization [20,71]. This “structure-first” framework moves inference beyond ploidy estimates and a handful of gene trees to chromosome-scale comparisons, enabling conserved regions, structural variation, and repeat landscape differences to be evaluated in a unified reference and tested for spatial correspondence with expression shifts or candidate adaptive signals [12,52].
At the cross-species comparative level, a common approach is to integrate transcriptome resources with a phylogenetic framework. Comparable ortholog sets and gene families are built across species, and gene family expansion/contraction, positive selection, or accelerated evolution are evaluated under an explicit species tree [68]. Candidate signals are then organized into relatively stable functional modules using GO/KEGG enrichment, generating “candidate sets” that support the stressor-organized synthesis in Chapter 6 [69]. The reusability of such conclusions often depends less on any single algorithm than on improved data coverage: multi-species, multi-tissue datasets that can be repeatedly queried reduce sensitivity to a particular sampling design or tissue choice.
In population-scale studies, the dominant workflow resembles a combined evidence chain from “geographic structure-historical processes-candidate signals.” Researchers typically begin by identifying population structure and divergence patterns using PCA, clustering, or phylogenetic networks, then interpret these patterns using F_ST/AMOVA, demographic reconstructions, and coalescent frameworks. Candidate regions are subsequently scanned using divergence statistics and haplotype-based metrics, and candidate genes are evaluated against environmental gradients or phenotypic contrasts [9,16]. In schizothoracines, WGS resequencing studies have already produced relatively complete narratives linking population structure, demographic history, and adaptive candidates, providing direct nuclear-genomic entry points for later discussion of management units and connectivity [16]. Meanwhile, transcriptomic analyses often serve to map candidate signals back onto tissues and stress responses: differential expression, co-expression networks (e.g., WGCNA), and enrichment analyses are used to distill modules associated with tissue functions or environmental responses, and to cross-validate signals inferred from comparative genomics or population scans. In this way, “candidate genes” are not only statistically significant, but also re-embedded in physiologically interpretable contexts [12,16,70].
Transposable elements/structural variation, duplicate gene fates, and heterogeneous rediploidization are recognized as key background mechanisms. However, in schizothoracines, systematic evidence that can be pinpointed simultaneously to a specific lineage, genomic region, and gene set remains concentrated in a small number of species and datasets. Accordingly, this chapter uses existing studies as anchors to summarize reusable analytical chains and to delineate the steps that still require validation; functional interpretations and their alignment with major stressors are developed in the next chapter using more specific evidence. In practice, the most effective ways to raise the evidentiary level tend to come from filling three types of data gaps: systematic SV/repeat comparisons, standardized transcriptome–phenotype joint measurements under key stressors, and broad lineage-level population genomic sampling that covers transition zones and putative hybrid regions.
6. Molecular Mechanisms of High-Altitude Adaptation: Shared Themes and What Makes “Aquatic High-Altitude Adaptation” Distinct
This chapter organizes molecular evidence for high-altitude adaptation in schizothoracines by major stressors, prioritizing topics for which existing data can support a closed loop of “comparative omics-population divergence-tissue expression/stress responses.” Key tissues frequently used for mechanistic inference and stress-response profiling are summarized in Figure 2. For modules where evidence is fragmented or debated, we specify the scope of applicability and the key steps that remain to be tested.
6.1. Cold Stress and Metabolic Remodeling
High-altitude lakes and rivers typically experience long ice-covered periods and short ice-free seasons. Persistently low temperatures constrain aquatic primary productivity, which becomes concentrated in the ice-free window and shows pronounced seasonality [71,72]. Under such conditions, cold adaptation in schizothoracines is often better captured as a system-level remodeling of energy balance rather than as a small set of “cold genes”: comparative genomic analyses, population-based selection scans, and transcriptomic studies repeatedly converge on pathways involved in energy metabolism [12,73]. Across-species comparisons in schizothoracines and closely related high-altitude cyprinids frequently report enrichment of candidate genes in mitochondrial energy pathways and in modules related to carbohydrate and lipid metabolism. At the population scale, whole-genome resequencing (WGS)–based selection scans often recover these metabolic modules again, suggesting that they represent cross-study “shared themes” rather than idiosyncratic signals from individual datasets [16,69,74]. In population genomic workflows, studies commonly resolve population structure first, then combine divergence statistics with selection scans to localize candidate regions, and finally cross-check these candidates against transcriptomic responses in metabolically relevant tissues (e.g., brain, gill, hepatopancreas/liver, and muscle) under cold stress or additional stressors such as salinity–alkalinity challenges [17,75].
6.2. Intense UV Radiation and DNA Damage Repair
UV (Ultraviolet Radiation) exposure in high-altitude waters is jointly modulated by optical properties—especially CDOM, suspended particles/turbidity, and water clarity—and by organisms’ vertical distribution and depth use. However, in clear, oligotrophic plateau lakes and river reaches, UV can penetrate deeper and may impose chronic DNA damage and genome-maintenance pressure on fishes [76]. In comparative genomic analyses and population WGS studies of schizothoracines and closely related high-altitude cyprinids, DNA repair–related pathways recur in enrichment results and can be regarded as a relatively distinctive candidate axis of adaptation [12,16].
Current evidence broadly falls into two complementary layers. At the genomic level, chromosome-scale assemblies and population resequencing studies repeatedly highlight DNA repair and genome-stability maintenance processes among candidate genes or enriched categories, with signals consistent with population differentiation and local adaptation [12,16]. At the transcriptomic level, multi-tissue or organ-focused RNA-seq under hypoxia, cold exposure, or engineering-related stressors often detects differential regulation of gene networks involved in oxidative damage responses, cell-cycle control/apoptosis, and repair regulation; the response tends to be tissue-specific and dependent on exposure duration and intensity [75,77].
6.3. Hypoxia: Inconsistent Signals
In some schizothoracine studies, the HIF axis or canonical hypoxia pathways do not show the strong and consistent enrichment of positive selection or convergent selection that might be expected. However, targeted comparative analyses and functional assays focusing on the HIF-α family suggest that adaptive changes are indeed present in schizothoracines’ hypoxia responses [78]. This discrepancy likely reflects the pronounced spatiotemporal heterogeneity of dissolved oxygen (DO) in plateau rivers and the re-oxygenation effects of hydrodynamics: steep gradients and turbulence can markedly enhance gas exchange and reaeration, while discharge and temperature fluctuations further drive DO dynamics [79]. In addition, behavioral selection for flow conditions and microhabitats may buffer the ecological experience of “chronic hypoxia,” thereby weakening strong, single-pathway selection signatures at the sequence level [80].
At present, beyond potential “signal underestimation” caused by sampling a single tissue or time point, or by limited sample sizes, another plausible interpretation is that adaptation to DO variability can be achieved through multiple routes in fishes. In addition to classical hypoxia regulation, shifts in gill structure/oxygen transport capacity and metabolic regulation of energy use may contribute, with a substantial component of plasticity and reversibility [81].
6.4. Osmoregulation, Ion Homeostasis, and Water Chemistry
Lakes and rivers across the Qinghai–Tibetan Plateau show pronounced heterogeneity in water chemistry, including salinity, pH, hardness, and the composition of major ions; many plateau lakes are characterized by distinctly saline–alkaline conditions [82]. Accordingly, adaptations associated with water chemistry are most likely to be concentrated in functional modules related to ion transport, osmoregulatory control, and acid–base homeostasis. Comparative genomic and multi-omics studies in schizothoracines and closely related plateau cyprinids have repeatedly detected signals of accelerated evolution and/or differential expression in genes involved in ion balance and osmoregulation, suggesting that this module can be treated as a “water-specific divergence module” and retained as a priority target for cross-system comparisons under contrasting hydrochemical regimes [1,17,83].
A more operational next step is to integrate basin-wide population genomic datasets with environmental covariates (e.g., salinity, pH, and major-ion composition) using genotype–environment association analyses jointly modeled with river-network connectivity. Such a framework would allow the relationships among population structure, environmental gradients, and candidate pathways to be evaluated within a unified statistical model and tested in a reproducible manner across catchments.
6.5. Immunity and Stress Responses
Immune- and stress-related processes are frequently reported in studies of schizothoracines (and closely related plateau cyprinids). In cross-species comparative and phylogenetic frameworks, analyses based on dN/dS, tests of positive selection, or accelerated evolution often highlight immune-related GO categories (e.g., “immune system process/immune response”) alongside candidate themes such as metabolism, DNA repair, and cardiovascular development as components of putative high-altitude adaptation [69].
At the level of tissue transcriptomics and stress-exposure experiments (e.g., hypoxia challenges or engineering-related stressors), RNA-seq studies commonly reveal coordinated shifts in immune genes together with molecular chaperones (including heat shock proteins), oxidative-damage responses, and cell-cycle/apoptosis regulatory networks. These responses are typically tissue dependent and strongly contingent on exposure duration and intensity [77]. Consequently, immune signals in schizothoracines are more appropriately interpreted as part of an integrated response network coupled to energy metabolism and oxidative stress, rather than as isolated pathway readouts.
Importantly, both the direction and magnitude of immune-related signals vary with tissue type, stress severity, and sampling time window. A more conservative synthesis is therefore that schizothoracines often exhibit coupled immune–metabolic–oxidative stress responses under multiple stress contexts, whereas the relative contributions of long-term genetic adaptation versus short-term plasticity remain to be disentangled through standardized exposure designs and coordinated multi-tissue phenotyping and transcriptomic profiling.
7. Phenotypic, Morphological, and Life-History Adaptation: Linking “Molecular Modules” to Ecological Function
This chapter synthesizes testable evidence at the phenotypic and life-history levels. It highlights reusable evidence chains in morphometrics, otolith-based approaches, and age–growth analyses in schizothoracines, providing support for subsequent MU/ESU delineation and the pressure–management sections.
7.1. Morphological Differentiation and Niche Association
Morphological variation in schizothoracines has long served as a primary entry point for taxonomic identification and studies of ecological adaptation. Across plateau river–lake systems, hydrodynamic regimes and food-resource gradients often impose consistent functional constraints on body shape, fin morphology, and feeding-related traits such as head and mouth structures [28]. However, the limitations of morphology-based inference are also well recognized: traits associated with feeding or flow conditions may evolve repeatedly across lineages, weakening the robustness of morphology-only grouping and classification [11]. In addition, natural hybridization and introgression can produce intermediates and continuous trait distributions, contributing to species complexes and blurred boundaries in some taxa [22]. Current studies therefore tend to rely on two complementary strategies. First, traditional linear measurements are increasingly extended to landmark-based geometric morphometrics coupled with multivariate statistics; after correcting for body size and allometric effects, this framework can more reliably separate closely related species or geographically structured groups within a shared “morphospace” [84,85]. Second, morphological differences are interpreted explicitly within environmental and functional contexts by linking body and fin traits to flow regimes, and mouth morphology to feeding resources or substrate use, thereby placing observed variation within habitat gradients and functional constraints [11,86]. In questions of species boundaries and hybridization, morphology is most informative for indication and localization (e.g., identifying candidate contact zones or suspect individuals), but it is generally insufficient as a standalone basis for definitive inference. It should also be noted that morphological divergence in plateau fishes is shaped by biological state (including ontogeny and reproductive condition) and can be strongly plastic in response to environmental drivers such as temperature, flow velocity, and food availability [87,88]. As a result, within-species variation across habitats or sampling periods may be large enough to confound morphology-only assignment [89]. For reproducible inference, studies should therefore report sampling season, body-size ranges, and key environmental background variables, and evaluate morphological differentiation jointly with genetic divergence and/or life-history evidence where possible.
7.2. High-Throughput Phenotyping
With the expanding use of imaging and computer-vision approaches in fish research, schizothoracines have begun to see studies centered on automated phenotyping and high-throughput acquisition of morphological data. For example, multi-view image datasets can be combined with deep-learning–based keypoint detection and 2D/3D keypoint reconstruction to generate large numbers of quantitative traits—such as body proportions and metrics of the head and caudal peduncle—at scale, enabling comparisons of morphological differentiation and aiding taxonomic discrimination across habitat types including rivers, lakes, and transition zones [91].
The main contribution of this line of work is that it converts conventional “low-throughput, labor-intensive” measurement steps into a more efficient and reproducible workflow, thereby providing practical tools for resource surveys and long-term monitoring. Nevertheless, transferring these models across river basins or among species typically requires external validation and calibration, because differences in imaging conditions, body posture, ontogenetic stage, and lineage-specific morphology can reduce generalization performance without targeted re-training or standardization.
7.3. Otolith Microchemistry and Life-History Reconstruction
Otolith microchemistry provides an independent line of evidence for reconstructing schizothoracine life histories, complementary to genomics and morphology. In Schizothorax grahami, for instance, variation in Sr:Ca and Mg:Ca along the otolith growth axis can help differentiate natal river reaches or spawning-ground sources, supporting early-life history reconstruction and identification of population units [90]. Methodologically, using elemental ratios such as Sr, Ba, and Mg relative to Ca—and interpreting them against hydrochemical and geological “baselines”—is a common framework in otolith microchemistry for schizothoracines and other freshwater fishes. When integrated with annulus information, “core-to-edge” chemical transects can be mapped onto ontogenetic stages, allowing reconstruction of the timing of movements and residency over an individual’s lifetime [92,93].
For schizothoracine syntheses, within-basin differences in spawning-ground water chemistry can be reflected in otolith elemental ratios and their trajectories along the growth axis, enabling inference of early origin and key life-history transitions [90]. Under damming or river-network fragmentation, otolith microchemistry can be used to evaluate whether individuals have moved across reaches, whereas population genetic structure more directly reflects intergenerational gene flow and longer-term isolation [94]. Combining these two evidence streams places contemporary ecological processes and historical connectivity into the same evaluative framework, strengthening the interpretation of management-relevant questions [95].
Interpretation of otolith microchemistry depends critically on hydrochemical baselines: spatiotemporal variability in water chemistry, as well as temperature and salinity effects, can influence elemental incorporation, meaning chemical signatures are not always in one-to-one correspondence with habitats [96]. Accordingly, otolith-based inferences should be grounded in well-characterized water-chemistry baselines and supported by multi-element approaches and spatiotemporal replicate sampling to improve the reliability of provenance and life-history interpretation.
7.4. Age and Growth
Across many schizothoracine lineages, the plateau environment—characterized by low temperatures and a short growing season—is associated with a life-history syndrome of slow growth, longevity, and late maturation. These traits represent a key functional endpoint for understanding high-elevation adaptation and also provide parameters directly relevant to fisheries assessment and management. In Schizothorax o’connori, otolith-section–based age and growth analyses indicate that maximum age can reach ~50 years in females, and that von Bertalanffy growth coefficients are low (k ≈ 0.081–0.095), consistent with a classic long-lived, slow-growing strategy [97]. Subsequent age-validation work further shows that annual periodicity of annulus formation in S. o’connori can be supported using approaches such as daily increment analysis and marginal increment ratios, and that annulus formation occurs within a distinct seasonal window [98].
From a management perspective, stock assessment and per-recruit analyses for Oxygymnocypris stewartii have compared strategies such as minimum-age limits and seasonal closures. Results suggest that slow growth, longevity, and late maturation can amplify vulnerability to human disturbance; delayed female maturation also implies that large, old females contribute disproportionately to reproductive output, and thus warrant higher protection priority in management designs [99]. Similarly, population-level work on S. waltoni emphasizes that its life-history profile—longevity, slow growth, late maturation, and low fecundity—can increase sensitivity to pressures such as overfishing [13].
In schizothoracine age-and-growth studies, progress is not only about deriving age-reading metrics, but also about clearly documenting quality control procedures and sources of uncertainty. Age estimation typically relies on annuli in hard structures such as otoliths and requires seasonal validation of annulus formation and replicated readings. In older individuals, otolith-based age can show systematic discrepancies, making structural comparisons and explicit validation procedures essential for error control [98,100].
7.5. Reproductive Ecology and Trophic Ecology
Compared with the literature on age and growth, evidence on reproductive and trophic ecology in Schizothoracinae is more uneven across species and drainage basins. Nevertheless, these lines of evidence map more directly onto management because they determine where spawning grounds are located, how spawning microhabitats are assembled, and how reproductive activity is synchronized with water temperature and discharge regimes—factors that directly affect the effectiveness of reserve placement, ecological flow regulation, and habitat restoration [101]. In recent years, studies have begun to quantify spawning-habitat preferences using tools such as acoustic telemetry and hydrodynamic–substrate surveys, providing operational evidence for identifying “key reaches” and “key seasonal windows” [18].
On the trophic side, gut microbiome profiling and metabolic/physiological indicators are increasingly used to characterize health status and infer signals of environmental accommodation in different schizothoracine fishes. These studies suggest that diet and habitat differences can strongly shape microbial community structure [102,103]. However, such signals show pronounced seasonal and spatial variability; a more cautious interpretation is therefore to treat them as auxiliary indicators of health and habitat quality. Comparable conclusions will require standardized sampling across seasons and habitats, together with appropriate environmental baselines, to support robust cross-study comparisons.
8. Population Genetic Structure and Conservation Units: From Genetic Diversity to Management Units (MU/ESU)
Conservation genetics in schizothoracine fishes is strongly shaped by river-network spatial structure. Population differentiation within a single species across reaches and tributary systems is often jointly molded by canyon topography, the legacy of river capture, modern engineering barriers, and habitat fragmentation, and thus manifests as restricted contemporary gene flow and increasingly pronounced genetic structure. Accordingly, this chapter synthesizes existing population genetic and population genomic studies of Schizothoracinae, prioritizing evidence that can close the loop from spatial differentiation to historical processes and then to management implications, and translating genetic inferences into operational delineation of management units and actionable recommendations.
8.1. Operationalizing MU/ESU in Schizothoracine Research
In the conservation-genetic context of schizothoracines, a management unit (MU) is commonly used to describe population units at contemporary or near-contemporary timescales in which gene flow is insufficient to maintain demographic connectivity. Evidence typically comes from clear genetic clustering, significant differentiation, and indications of reduced migration rates [104,105]. By contrast, an evolutionarily significant unit (ESU) emphasizes longer-term evolutionary independence, generally requiring stable lineage-level evidence and, where available, additional support from signals of adaptive differentiation associated with environmental gradients [106].
In plateau river networks, dendritic topology and reach-scale barriers constrain and channel dispersal, and—together with asymmetric gene flow—often produce spatial genetic structure along “mainstem–tributary–key node” axes. Under these conditions, MU delineation is frequently closer to the scale of implementable basin and reach management. ESUs, in turn, are more likely to correspond to deeper lineage partitions shaped by longer-term geological history [107]. For example, whole-genome resequencing studies have resolved clear reach-level population differentiation in schizothoracines and proposed MU boundaries accordingly [108].
Therefore, MU/ESU delineation in schizothoracines should rarely be reduced to a single statistic or a fixed threshold. Instead, it depends on the completeness of the evidence chain—whether sampling covers key transition zones and potential contact areas, whether analyses can distinguish barrier effects from isolation by distance, and whether potential hybrid zones can be detected and evaluated [104,105].
8.2. Upgrading Data Types and Elevating Evidence Strength
Early population genetic studies of Schizothoracinae largely relied on low-cost markers—typically mitochondrial sequences combined with microsatellites—which were often sufficient to resolve clear geographic differentiation and population structure across multiple basins and reach scales, thereby providing a first-pass framework for putative isolated units and management zoning [39]. In recent years, genome-wide SNP datasets have become increasingly available in schizothoracines. For example, GBS data have been used in Schizothorax kozlovi to characterize population structure and differentiation and to further evaluate how basin isolation and environmental gradients contribute to genetic divergence—effectively extending the question of “whether populations cluster” from a limited number of loci to much denser genome-wide evidence [109].
Meanwhile, population studies based on whole-genome resequencing (WGS resequencing) have begun to emerge in schizothoracines, enabling genetic diversity, effective population size, and candidate-region mapping to be assessed jointly within a single analytical framework. This provides a more direct route for moving MU/ESU inference from reliance on a single metric to evidence grounded in multi-criterion concordance [10]. Such data upgrades also make it easier to extend MU/ESU delineation beyond “structure and clustering” toward integrated risk assessment and management translation.
8.3. Multi-Scale Evidence Chains and Management Translation
Schizothorax o’connori (a representative schizothoracine in the Yarlung Tsangpo River system) is among the taxa for which a relatively complete “evidence loop” can be assembled in conservation genetics. The availability of a chromosome-level reference genome provides a unified coordinate system for variant discovery and population comparisons, allowing population divergence and candidate regions to be anchored to a verifiable genomic background. In addition, whole-genome resequencing (WGS) population studies have integrated population structure with signals potentially related to adaptation within a single analytical framework, making inferences about connectivity differences, spatial patterns of divergence, genetic diversity and risk, and proposed management units (MUs) more operational and testable [12].
At the lineage and population-structure level, populations from different reaches of the Yarlung Tsangpo River are not fully admixed. WGS resequencing can resolve spatial differentiation at higher resolution, quantify levels of genetic diversity, and detect signatures consistent with bottlenecks or connectivity differences, thereby providing a data basis for linking divergence patterns to canyon geomorphology, changes in river-network connectivity, and potential barrier effects [12,110]. Where genetic clustering aligns closely with geographic units, a precautionary management implication is to minimize translocation and stocking across units to reduce the risk of disrupting existing genetic structure, while prioritizing monitoring on both sides of transition zones to support adaptive, data-driven updates of MU boundaries.
At the population-genomic level, WGS studies commonly evaluate population structure together with genetic diversity, demographic history, and putative selection signals, bringing “genetic health risks” and “potential adaptive hints” into the same inferential framework [16,111]. For S. o’connori, existing WGS resequencing work has described nuclear-genomic relationships and divergence across multiple geographic groups/sites, reported diversity levels and the strength of among-population differentiation, and identified candidate signals potentially associated with environmental adaptation. On this basis, these studies further discuss conservation priorities and management implications across geographic units, providing an evidential interface in which population structure, genetic risk, and candidate signals can corroborate one another [16]. When low diversity, bottleneck/inbreeding indicators, or strong geographic concordance in divergence are observed, MU boundaries are no longer merely “statistical clusters” but can be directly translated into prioritization schemes, broodstock sourcing rules, and risk controls for cross-reach translocation.
Based on currently available population genetic and population genomic evidence, MU delineation for S. o’connori can be hypothesized to follow major river reaches and tributary systems, with intensified sampling in geomorphic transition zones and potential contact areas to test boundary stability. In enhancement stocking and broodstock conservation, a conservative principle is “source within the same unit, prioritize the same reach,” while assigning higher protection weight to populations with low diversity or signals consistent with bottlenecks/inbreeding risk. Coupling these practices with long-term monitoring of connectivity and use of key habitats will also allow MU boundaries to be updated dynamically as new evidence accumulates [16,112].
9. Anthropogenic Pressures and Management Responses: Translating Evidence into an Actionable Checklist
A central challenge in schizothoracine conservation is that multiple pressures often accumulate over the long term within the same river-network unit. Population recovery therefore tends to depend disproportionately on a limited number of key reaches, seasonal windows, and high-value breeders, making it necessary to translate evidence into management actions that are verifiable, auditable, and adaptable.
9.1. Minimum Actionable Response Framework for Three Major Pressure Types
Because many schizothoracine taxa exhibit slow growth, late maturation, and long lifespans, recovery is often more sensitive to size/age structure and recruitment bottlenecks than to interannual fluctuations in total catch [13,113]. Accordingly, harvest regulation should shift from “controlling total yield only” to “protecting population structure.” A practical core is to enforce minimum legal size and seasonal closures during the spawning period, narrowing enforcement to two readily checkable violations—fishing during the closed season and harvesting undersized fish—while using rebounds in body-length/age structure and improved post-spawning juvenile recruitment as more sensitive performance indicators. In severely depleted reaches, additional restrictions on large-bodied (high-fecundity) individuals can further reduce recovery dependence on a small number of breeders [114].
Under engineering disturbance and reach fragmentation, management and engineering should share explicit, auditable connectivity targets to avoid situations where “infrastructure is built but function remains unclear.” A more operational approach is to translate connectivity into acceptance criteria: prioritize passability during key migration periods, and evaluate performance using arrival efficiency and passage efficiency, the number of passable days, and passage outcomes under standardized discharge conditions [115,116]. Meanwhile, enhancement stocking and cross-reach translocations should align with MU boundaries. When genetic clustering corresponds closely to geographic units, a conservative default is “source within the same MU and release within the same MU,” coupled with intensified monitoring on both sides of transition zones or suspected barriers to reduce risks of genetic swamping or outbreeding via introduced genotypes and to provide evidence for subsequent, adaptive updates of MU delineation and management measures [117].
Habitat degradation and altered hydrological regimes commonly weaken juvenile recruitment via two pathways: (i) phenological mismatch in the flow–temperature regime that supports spawning and early development, and (ii) shrinkage of suitable microhabitat area for spawning and nursery functions. Restoration and ecological flow regulation are therefore best organized around a “key reach–key time window–key microhabitat” unit, prioritizing interventions in the spatial and temporal windows that contribute most to recruitment [118,119]. In practice, this can begin with functional zoning of river reaches, treating spawning grounds, nursery shallows, and overwintering deep pools as priority functional units; then defining critical windows for reproduction and early development based on the coupled temperature–discharge regime, and targeting ecological regulation toward relative stability of discharge and hydraulics within those windows to minimize disturbance to spawning and early life stages [119,120]. Microhabitat restoration should be implemented with a quantified and auditable parameter set (e.g., depth, velocity, substrate, cover structures, and their effective areas), enabling engineering acceptance, cross-site comparison, and closed-loop integration with post-restoration monitoring [121].
9.2. One-Page Action Checklist and Evaluation Metrics
Management actions can be condensed into a tiered “one-page” checklist: implement a closed season and minimum legal size; establish fixed monitoring stations in transition zones; register broodstock and juveniles used for stocking and, as a default, match sources to MUs; complete functional zoning of key river reaches; and, in one to two representative reaches, quantify passage performance during critical seasonal windows while running pilot demonstrations of ecological flow regulation and microhabitat restoration [99,122,123]. A tiered one-page checklist and corresponding evaluation metrics are summarized in Table 1. To keep implementation costs manageable, evaluation can be limited to three indicator groups: connectivity (arrival/passage efficiency during key windows or number of passable days), population status (juvenile recruitment and body-length/age structure), and management risk (compliance of stocking sources and early-warning signals of structural anomalies in transition zones). If core indicators show no improvement for two consecutive years, management should adjust flow-regulation rules, relocate restoration interventions, or tighten harvest/stocking policies [124].
Overall, MU boundaries, critical seasonal windows, and quantified connectivity/habitat metrics can serve as an integrating framework that places genetic structure, life-history vulnerability, and engineering interventions into a common evaluation context, enabling a closed-loop system of adaptive management.
10. Integrated Framework and Outlook
Research on schizothoracine fishes can be summarized as a transferable chain: river-network history and environmental gradients shape the context for population isolation and selection, and—on the basis of polyploidy and genome remodeling—give rise to key molecular modules; these modules then project onto differences in phenotype, life history, and ecological function, and are ultimately translated into actionable conservation units and management rules. The next priority is not to further expand a “phenomena checklist”, but to ensure that this evidence chain and its indicator system remain verifiable, comparable, and accumulative across more basins and more species.
Looking ahead, debated topics should be reorganized into testable questions. For stressors with inconsistent signals (e.g., hypoxia), the key is not to add more “significant or not” results, but to produce stable, comparable evidence under a unified framework so that disagreements become interpretable differences. In parallel, emphasis should shift from stacking enrichment outputs to convergent mechanistic explanation, focusing on modules that recur across studies and show relatively consistent directions, thereby improving transferability and reproducibility. Cross-scale linkage is equally critical: only when population divergence, key phenotypes, and ecological processes can be observed and interpreted within the same spatial unit and time window can inferences “from divergence to function to risk” approach a testable causal narrative. Finally, outputs should better match governance needs by compressing scientific conclusions into a small set of shared indicators and rule-based recommendations that are traceable, auditable, and iterative, reducing the gap between “correct conclusions” and “hard-to-implement actions”.
To enable cross-basin accumulation of evidence, future work should adopt sampling designs that cover key transition zones, reproducible metadata standards, and a core set of cross-study comparable indicators as minimum requirements. With these foundations, schizothoracine research can shift more reliably from case-driven findings to framework-driven synthesis and provide more replicable knowledge and tools for conserving biodiversity in plateau river networks.
Author Contributions
Conceptualization, Y.C., X.Z. and H.Q.; Literature search, Y.C.; Data curation, Y.C.; Formal analysis, Y.C., X.Z., H.Q.; Writing – original draft, Y.C.; Writing – review & editing, Y.C., N.W., X.Z. and H.Q.; Supervision, X.Z. and H.Q.; Project administration, X.Z.; Funding acquisition, X.Z.; Validation, Y.C. and H.Q. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Wunonglong–Lidi Hydropower Plants 2025–2027 Fish Lift and Fish Collection/Transport System Operation and Maintenance Management Contract (Grant Number LDDC2025/P11).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article, as no new data was generated.
Conflicts of Interest
The authors state no conflicts of interest.
References
- Tian, F.; Liu, S.; Zhou, B.; Tang, Y.; Zhang, Y.; Zhang, C.; Zhao, K. Chromosome-level genome of Tibetan naked carp (Gymnocypris przewalskii) provides insights into Tibetan highland adaptation. DNA Res. 2022, 29, dsac025. [CrossRef]
- Xie, Y. G.; Qi, Y. L.; Luo, Z. H.; Qu, Y. N.; Yang, J.; Liu, S. Q.; Yang, H. L.; Xie, D. W.; Wang, Z.; Jiang, H. C.; et al. Lake salinization on the Qinghai-Tibetan Plateau alters viral community composition and lifestyles. Commun. Earth Environ. 2025, 6, 49. [CrossRef]
- Ren, W.; Gao, Y.; Qian, H.; Qu, W.; Shi, X.; Ma, Y.; Su, Z.; Ma, W. The Evolution and Drivers of Hydrochemistry in Nam Co Lake, the Third Largest Lake on the Tibetan Plateau, over the Last 20 Years. Sustainability 2025, 17, 2180. [CrossRef]
- Grabner, D.; Rothe, L. E.; Sures, B. Parasites and Pollutants: Effects of Multiple Stressors on Aquatic Organisms. Environ. Toxicol. Chem. 2023, 42, 1946-1959. [CrossRef]
- Hughes, S. J.; Santos, J.; Ferreira, T.; Mendes, A. Evaluating the response of biological assemblages as potential indicators for restoration measures in an intermittent Mediterranean river. Environ. Manage. 2010, 46, 285-301. [CrossRef]
- Vieira, T. B.; Tejerina-Garro, F. L. Relationships Between Environmental Conditions And Fish Assemblages InTropical Savanna Headwater Streams. Sci. Rep. 2020, 10, 2174. [CrossRef]
- Du, T.; Ding, C.; Yang, K.; Chen, J.; Liu, X.; Lv, W.; Ding, L.; He, D.; Tao, J. A global dataset on species occurrences and functional traits of Schizothoracinae fish. Sci. Data 2024, 11, 272. [CrossRef]
- He, Z.; Gao, K.; Chen, H.; Yang, D.; Pu, Y.; Zheng, L.; Jiao, Y.; Xiong, J.; Chen, Q.; Lai, B.; et al. Comparative Population Dynamics of Schizothorax wangchiachii (Cyprinidae: Schizothoracinae) in the Middle Reaches of the Yalong River and the Upper Reaches of the Jinsha River, China. Animals 2023, 13, 2209. [CrossRef]
- Chen, W.; Yue, X.; He, S. Genetic differentiation of the Schizothorax species complex (Cyprinidae) in the Nujiang River (upper Salween). Sci. Rep. 2017, 7, 5944. [CrossRef]
- Zhou, C.; Xiao, S.; Liu, Y.; Mou, Z.; Zhou, J.; Pan, Y.; Zhang, C.; Wang, J.; Deng, X.; Zou, M.; et al. Comprehensive transcriptome data for endemic Schizothoracinae fish in the Tibetan Plateau. Sci. Data 2020, 7, 28. [CrossRef]
- Tang, Y.; Li, C.; Wanghe, K.; Feng, C.; Tong, C.; Tian, F.; Zhao, K. Convergent evolution misled taxonomy in schizothoracine fishes (Cypriniformes: Cyprinidae). Mol. Phylogenet. Evol. 2019, 134, 323-337. [CrossRef]
- Xiao, S.; Mou, Z.; Fan, D.; Zhou, H.; Zou, M.; Zou, Y.; Zhou, C.; Yang, R.; Liu, J.; Zhu, S.; et al. Genome of Tetraploid Fish Schizothorax o'connori Provides Insights into Early Re-diploidization and High-Altitude Adaptation. iScience 2020, 23, 101497. [CrossRef]
- Guo, X. Z.; Zhang, G. R.; Wei, K. J.; Ji, W.; Yan, R. J.; Wei, Q. W.; Gardner, J. P. A. Phylogeography of the threatened tetraploid fish, Schizothorax waltoni, in the YarlungTsangpo River on the southern Qinghai-Tibet Plateau: implications for conservation. Sci. Rep. 2019, 9, 2704. [CrossRef]
- Regmi, B.; Douglas, M. R.; Wangchuk, K.; Zbinden, Z. D.; Edds, D. R.; Tshering, S.; Douglas, M. E. The Himalayan uplift and evolution of aquatic biodiversity across Asia: Snowtrout (Cyprininae: Schizothorax) as a test case. PLoS One 2023, 18, e0289736. [CrossRef]
- Xu, M. R.; Liao, Z. Y.; Brock, J. R.; Du, K.; Li, G. Y.; Chen, Z. Q.; Wang, Y. H.; Gao, Z. N.; Agarwal, G.; Wei, K. H.; et al. Maternal dominance contributes to subgenome differentiation in allopolyploid fishes. Nat. Commun. 2023, 14, 8357. [CrossRef]
- Gao, K.; He, Z.; Xiong, J.; Chen, Q.; Lai, B.; Liu, F.; Chen, P.; Chen, M.; Luo, W.; Huang, J.; et al. Population structure and adaptability analysis of Schizothorax o'connori based on whole-genome resequencing. BMC Genomics 2024, 25, 145. [CrossRef]
- Zhou, B.; Sui, R.; Yu, L.; Qi, D.; Fu, S.; Luo, Y.; Qi, H.; Li, X.; Zhao, K.; Liu, S.; et al. Transcriptomics and proteomics provide insights into the adaptative strategies of Tibetan naked carps (Gymnocypris przewalskii) to saline-alkaline variations. BMC Genomics 2025, 26, 162. [CrossRef]
- Li, B.; Hu, F.; Li, W.; Su, W.; Zhu, J.; Jiang, W. Spawning habitat selection in Schizothorax wangchiachii using acoustic tagging and tracking. Front. Ecol. Evol. 2025, 13, 1615081. [CrossRef]
- Lu, Y.; Zhu, W. Y.; Liu, Q. Y.; Li, Y.; Tian, H. W.; Cheng, B. X.; Zhang, Z. Y.; Wu, Z. H.; Qing, J.; Sun, G.; et al. Impact of Low-Head Dam Removal on River Morphology and Habitat Suitability in Mountainous Rivers. Int. J. Environ. Res. Public Health 2022, 19, 11743. [CrossRef]
- Niu, J.; Zhang, R.; Hu, J.; Zhang, T.; Liu, H.; Minavar, M.; Zhang, H.; Xian, W. Chromosomal-scale genome assembly of the near-extinction big-head schizothorcin (Aspiorhynchus laticeps). Sci. Data 2022, 9, 556. [CrossRef]
- Yuan, Q.; Li, K.; Wang, Y.; Ji, Q.; Liang, R. Tolerance and Growth of Adult Schizothorax prenanti Exposed and Re-Exposed to Supersaturated Gas Downstream of a Dam. Water 2022, 14, 2501. [CrossRef]
- Cheng, L.; Song, D.; Yu, X.; Du, X.; Huo, T. Endangered Schizothoracin Fish in the Tarim River Basin Are Threatened by Introgressive Hybridization. Biology 2022, 11, 981. [CrossRef]
- He, L.; Bi, Y.; Weese, D.; Wu, J.; Xu, S.; Ren, H.; Zhang, F.; Liu, X.; Chen, L.; Zhang, J. Genetic Signature of River Capture Imprinted in Schizopygopsis Fish from the Eastern Tibetan Plateau. Genes 2024, 15, 1148. [CrossRef]
- Li, M.; An, R.; Chen, M.; Li, J. Evaluation of Volitional Swimming Behavior of Schizothorax prenanti Using an Open-Channel Flume with Spatially Heterogeneous Turbulent Flow. Animals 2022, 12, 752. [CrossRef]
- Hoelzel, A. R. Where to now with the evolutionarily significant unit? Trends Ecol. Evol. 2023, 38, 1134-1142. [CrossRef]
- Alves, F.; Banks, S. C.; Edworthy, M.; Stojanovic, D.; Langmore, N. E.; Heinsohn, R. Using conservation genetics to prioritise management options for an endangered songbird. Heredity 2023, 130, 289-301. [CrossRef]
- Cao, W. X.; Chen, Y. Y.; Wu, Y. F.; Zhu, S. Q. Origin and evolution of schizothoracine fishes in relation to the upheaval of the Xizang Plateau. In Studies on the Period, Amplitude and Type of the Uplift of the Qinghai–Xizang Plateau; Tibetan Expedition Team of the Chinese Academy of Sciences, Ed.; Science Press: Beijing, China, 1981; pp. 118–130. https://www.cabidigitallibrary.org/doi/full/10.5555/19812608810.
- Qi, D.; Chao, Y.; Guo, S.; Zhao, L.; Li, T.; Wei, F.; Zhao, X. Convergent, parallel and correlated evolution of trophic morphologies in the subfamily schizothoracinae from the Qinghai-Tibetan plateau. PLoS One 2012, 7, e34070. [CrossRef]
- Zhang, C.; Tong, C.; Ludwig, A.; Tang, Y.; Liu, S.; Zhang, R.; Feng, C.; Li, G.; Peng, Z.; Zhao, K. Adaptive Evolution of the Eda Gene and Scales Loss in Schizothoracine Fishes in Response to Uplift of the Tibetan Plateau. Int. J. Mol. Sci. 2018, 19, 2953. [CrossRef]
- Li, Y.; Luo, Y.; Lv, Y.; Ou, Y.; Zhang, R.; Zhang, R. A Novel Mitochondrial Genome Resource for the Endemic Fish Gymnodiptychus integrigymnatus and Insights into the Phylogenetic Relationships of Schizothoracinae. Biology 2025, 14, 1760. [CrossRef]
- Ma, B.; Zhao, T.; Xu, B.; Zhong, L.; Wu, X.; Wei, K.; Zhang, Z.; Li, Y. Morphological variation in Schizothorax oconnori, Schizothorax waltoni (Teleostei: Cyprinidae: Schizothoracinae), and their natural hybrids from the middle Yarlung Zangbo River, Tibet. Ecol. Evol. 2024, 14, e11342. [CrossRef]
- Wang, T.; Qi, D.; Sun, S.; Liu, Z.; Du, Y.; Guo, S.; Ma, J. DNA barcodes and their characteristic diagnostic sites analysis of Schizothoracinae fishes in Qinghai province. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2019, 30, 592-601. [CrossRef]
- Antil, S.; Abraham, J. S.; Sripoorna, S.; Maurya, S.; Dagar, J.; Makhija, S.; Bhagat, P.; Gupta, R.; Sood, U.; Lal, R.; et al. DNA barcoding, an effective tool for species identification: a review. Mol. Biol. Rep. 2023, 50, 761-775. [CrossRef]
- Rozimov, A.; Wang, Y.; Wang, M.; Zou, M.; Sobirov, J.; Karimov, E.; Kholmatov, B.; Freyhof, J.; Namozov, S.; Wang, C.; et al. Mitochondrial genome insights into the phylogenetics and biogeographic evolution of snow trout (Cyprinidae, Schizothorax) in the Tien Shan Mountains. Zoosystematics and Evolution 2025, 101, 91-102. [CrossRef]
- Yue, X.; Shi, J.; Zou, Y.; Wang, B. The complete mitochondrial genome sequence of Schizothorax lissolabiatus (Cypriniformes: Cyprinidae). Mitochondrial DNA A DNA Mapp. Seq. Anal. 2016, 27, 2450-2. [CrossRef]
- Qin, Q.; Chen, L.; Zhang, F.; Xu, J.; Zeng, Y. Characterization of the Complete Mitochondrial Genome of Schizothorax kozlovi (Cypriniformes, Cyprinidae, Schizothorax) and Insights into the Phylogenetic Relationships of Schizothorax. Animals 2024, 14, 721. [CrossRef]
- Andersen, M. M.; Balding, D. J. How many individuals share a mitochondrial genome? PLoS Genet. 2018, 14, e1007774. [CrossRef]
- DeRaad, D. A.; McCullough, J. M.; DeCicco, L. H.; Hime, P. M.; Joseph, L.; Andersen, M. J.; Moyle, R. G. Mitonuclear discordance results from incomplete lineage sorting, with no detectable evidence for gene flow, in a rapid radiation of Todiramphus kingfishers. Mol. Ecol. 2023, 32, 4844-4862. [CrossRef]
- Guo, X. Z.; Zhang, G. R.; Wei, K. J.; Yan, R. J.; Ji, W.; Yang, R. B.; Wei, Q. W.; Gardner, J. P. Phylogeography and population genetics of Schizothorax o'connori: strong subdivision in the Yarlung Tsangpo River inferred from mtDNA and microsatellite markers. Sci. Rep. 2016, 6, 29821. [CrossRef]
- Heled, J.; Drummond, A. J. Bayesian inference of population size history from multiple loci. BMC Evol. Biol. 2008, 8, 289. [CrossRef]
- Payseur, B. A.; Rieseberg, L. H. A genomic perspective on hybridization and speciation. Mol. Ecol. 2016, 25, 2337-60. [CrossRef]
- Hohenlohe, P. A.; Funk, W. C.; Rajora, O. P. Population genomics for wildlife conservation and management. Mol. Ecol. 2021, 30, 62-82. [CrossRef]
- Cai, D.; Wang, X.; Li, G.; Jiao, R.; Kohn, B.; Zhu, W.; De Grave, J.; Lu, H. Fault systems impede incision of the Yarlung river into the Tibetan plateau. Commun. Earth Environ. 2023, 4, 200. [CrossRef]
- Stokes, M. F.; Kim, D.; Gallen, S. F.; Benavides, E.; Keck, B. P.; Wood, J.; Goldberg, S. L.; Larsen, I. J.; Mollish, J. M.; Simmons, J. W.; et al. Erosion of heterogeneous rock drives diversification of Appalachian fishes. Science 2023, 380, 855-859. [CrossRef]
- Swartz, E. R.; Chakona, A.; Skelton, P. H.; Bloomer, P. The genetic legacy of lower sea levels: does the confluence of rivers during the last glacial maximum explain the contemporary distribution of a primary freshwater fish (Pseudobarbus burchelli, Cyprinidae) across isolated river systems? Hydrobiologia 2014, 726, 109-121. [CrossRef]
- Robinson, R. A. J.; Brezina, C. A.; Parrish, R. R.; Horstwood, M. S. A.; Oo, N. W.; Bird, M. I.; Thein, M.; Walters, A. S.; Oliver, G. J. H.; Zaw, K. Large rivers and orogens: The evolution of the Yarlung Tsangpo–Irrawaddy system and the eastern Himalayan syntaxis. Gondwana Res. 2014, 26, 112-121. [CrossRef]
- Schmidt, J. L.; Zeitler, P. K.; Pazzaglia, F. J.; Tremblay, M. M.; Shuster, D. L.; Fox, M. Knickpoint evolution on the Yarlung river: Evidence for late Cenozoic uplift of the southeastern Tibetan plateau margin. Earth Planet. Sci. Lett. 2015, 430, 448-457. [CrossRef]
- Ledger, K. J.; Su, Y.; Jeon, J. Y.; Fullerton, A. H.; Kuligowski, D.; Bennett, T.; Denton, K.; McHenry, M.; McMillan, J. H.; et al. Using riverscape genetics to investigate the genetic response of two species and their life-history forms to dam removal. Front. Ecol. Evol. 2023, 11, 1225229. [CrossRef]
- Li, K.; Wang, J.; Wang, X.; Wang, M.; He, R.; Wang, M. Distribution pattern of fish richness in the Yarlung Zangbo River Basin. Diversity 2022, 14, 1142. [CrossRef]
- Barreto, S. B.; Knowles, L. L.; Affonso, P. R. A. M.; Batalha-Filho, H. Riverscape properties contribute to the origin and structure of a hybrid zone in a Neotropical freshwater fish. J. Evol. Biol. 2020, 33, 1530-1542. [CrossRef]
- Payseur, B. A.; Rieseberg, L. H. A genomic perspective on hybridization and speciation. Mol. Ecol. 2016, 25, 2337-60. [CrossRef]
- Liang, Y.; He, D.; Jia, Y.; Sun, H.; Chen, Y. Phylogeographic studies of schizothoracine fishes on the central Qinghai-Tibet Plateau reveal the highest known glacial microrefugia. Sci. Rep. 2017, 7, 10983. [CrossRef]
- Caves, J. K.; Bayshashov, B. U.; Zhamangara, A.; Ritch, A. J.; Ibarra, D. E.; Sjostrom, D. J.; Mix, H. T.; Winnick, M. J.; Chamberlain, C. P. Late Miocene uplift of the Tian Shan and Altai and reorganization of Central Asia climate. GSA Today 2017, 27, 20-26. [CrossRef]
- Dai, Y.; Han, H. Karyological analysis of two species in the subfamily Schizothoracinae (Cypriniformes: Cyprinidae) from China, with notes on karyotype evolution in Schizothoracinae. Turk. J. Fish. Aquat. Sci. 2018, 18, 175-186. [CrossRef]
- Chen, C. N.; Huang, Y. Y.; Li, H.; Long, Z. H.; Lai, J. S.; Liu, G. X.; Zhao, G. The complete mitochondrial genome of Schizothorax prenanti (Tchang) (Teleostei, Cyprinidae, Schizothoracinae). Mitochondrial DNA A DNA Mapp. Seq. Anal. 2016, 27, 253-254. [CrossRef]
- Gundappa, M. K.; To, T. H.; Grønvold, L.; Martin, S. A. M.; Lien, S.; Geist, J.; Hazlerigg, D.; Sandve, S. R.; Macqueen, D. J. Genome-Wide Reconstruction of Rediploidization Following Autopolyploidization across One Hundred Million Years of Salmonid Evolution. Mol. Biol. Evol. 2022, 39, msab310. [CrossRef]
- Krabbenhoft, T. J.; MacGuigan, D. J.; Backenstose, N. J. C.; Hannah Waterman H.; Lan T. Y.; Pelosi J. A.; Tan M.; Sandve S. R. Chromosome-level genome assembly of Chinese sucker (Myxocyprinus asiaticus) reveals strongly conserved synteny following a catostomid-specific whole-genome duplication. Genome Biol. Evol. 2021, 13, evab190. [CrossRef]
- Li, J. T.; Wang, Q.; Huang Yang, M. D.; Li, Q. S.; Cui, M. S.; Dong, Z. J.; Wang, H. W.; Yu, J. H.; Zhao, Y. J.; Yang, C. R.; et al. Parallel subgenome structure and divergent expression evolution of allo-tetraploid common carp and goldfish. Nat. Genet. 2021, 53, 1493-1503. [CrossRef]
- Redmond, A. K.; Casey, D.; Gundappa, M. K.; Macqueen, D. J.; McLysaght, A. Independent rediploidization masks shared whole genome duplication in the sturgeon-paddlefish ancestor. Nat. Commun. 2023, 14, 2879. [CrossRef]
- Lien, S.; Koop, B. F.; Sandve, S. R.; Miller, J. R.; Kent, M. P.; Nome, T.; Hvidsten, T. R.; Leong, J. S.; Minkley, D. R.; Zimin, A.; et al. The Atlantic salmon genome provides insights into rediploidization. Nature 2016, 533, 200-205. [CrossRef]
- Yang, L.; Mayden, R. L.; Naylor, G. J. P. Origin of Polyploidy, Phylogenetic Relationships, and Biogeography of Botiid Fishes (Teleostei: Cypriniformes). Biology 2025, 14, 531. [CrossRef]
- Glover, N. M.; Redestig, H.; Dessimoz, C. Homoeologs: What Are They and How Do We Infer Them? Trends Plant Sci. 2016, 21, 609-621. [CrossRef]
- Sancho, R.; Inda, L. A.; Díaz-Pérez, A.; Des Marais, D. L.; Gordon, S.; Vogel, J. P.; Lusinska, J.; Hasterok, R.; Contreras-Moreira, B.; Catalán, P. Tracking the ancestry of known and 'ghost' homeologous subgenomes in model grass Brachypodium polyploids. Plant J. 2022, 109, 1535-1558. [CrossRef]
- Soares, N. R.; Mollinari, M.; Oliveira, G. K.; Pereira, G. S.; Vieira, M. L. C. Meiosis in Polyploids and Implications for Genetic Mapping: A Review. Genes 2021, 12, 1517. [CrossRef]
- Xu, P.; Xu, J.; Liu, G.; Chen, L.; Zhou, Z.; Peng, W.; Jiang, Y.; Zhao, Z.; Jia, Z.; Sun, Y.; et al. The allotetraploid origin and asymmetrical genome evolution of the common carp Cyprinus carpio. Nat. Commun. 2019, 10, 4625. [CrossRef]
- Wang, L.; Zhu, P.; Mo, Q.; Luo, W.; Du, Z.; Jiang, J.; Yang, S.; Zhao, L.; Gong, Q.; Wang, Y. Comprehensive analysis of full-length transcriptomes of Schizothorax prenanti by single-molecule long-read sequencing. Genomics 2022, 114, 456-464. [CrossRef]
- Liang, X.; Wang, W.; Huang, J.; Luo, M.; Wangdui, N.; Sun, C.; Lu, J. A chromosome-level genome assembly of big-barbel schizothorcin, Schizothorax macropogon. Sci. Data 2024, 11, 1402. [CrossRef]
- Zhang, J.; Chen, Z.; Zhou, C.; Kong, X. Molecular phylogeny of the subfamily Schizothoracinae (Teleostei: Cypriniformes: Cyprinidae) inferred from complete mitochondrial genomes. Biochem. Syst. Ecol. 2016, 64, 6-13. [CrossRef]
- Tong, C.; Fei, T.; Zhang, C.; Zhao, K. Comprehensive transcriptomic analysis of Tibetan Schizothoracinae fish Gymnocypris przewalskii reveals how it adapts to a high altitude aquatic life. BMC Evol. Biol. 2017, 17, 74. [CrossRef]
- Zhang, D.; Yu, M.; Hu, P.; Peng, S.; Liu, Y.; Li, W.; Wang, C.; He, S.; Zhai, W.; Xu, Q.; et al. Genetic Adaptation of Schizothoracine Fish to the Phased Uplifting of the Qinghai-Tibetan Plateau. G3 2017, 7, 1267-1276. [CrossRef]
- Lei, L.; Deng, X.; Liu, F.; Gao, H.; Duan, Y.; Li, J.; Fu, S.; Li, H.; Zhou, Y.; Liao, R.; et al. Exploitation of Key Regulatory Modules and Genes for High-Salt Adaptation in Schizothoracine by Weighted Gene Co-Expression Network Analysis. Animals 2024, 15, 56. [CrossRef]
- Kirillin, G. B.; Shatwell, T.; Wen, L. Ice-covered lakes of Tibetan Plateau as solar heat collectors. Geophys. Res. Lett. 2021, 48, e2021GL093429. [CrossRef]
- Ban, X.; Dang, Y.; Shu, P.; Qi, H.; Luo, Y.; Xiao, F.; Feng, Q.; Zhou, Y. Estimation of phytoplankton primary productivity in Qinghai Lake using ocean color satellite data: Seasonal and interannual variations. Water 2024, 16, 1433. [CrossRef]
- Tong, C.; Tian, F.; Zhao, K. Genomic signature of highland adaptation in fish: a case study in Tibetan Schizothoracinae species. BMC Genomics 2017, 18, 948. [CrossRef]
- Xu, W.; Zhu, F.; Wang, D.; Chen, D.; Duan, X.; Liu, M.; Li, D. Comparative analysis of metabolites between different altitude Schizothorax nukiangensis (Cyprinidae, Schizothoracine) on the Qinghai-Tibet Plateau in Nujiang River. Water 2023, 15, 284. [CrossRef]
- Liu, S.; Tian, F.; Qi, D.; Qi, H.; Wang, Y.; Xu, S.; Zhao, K. Physiological, metabolomic, and transcriptomic reveal metabolic pathway alterations in Gymnocypris przewalskii due to cold exposure. BMC Genomics 2023, 24, 545. [CrossRef]
- Watanabe, S.; Overholt, E. P.; Schladow, S. G.; Vincent, W. F.; Williamson, C. E. Climate change and underwater light: Large-scale changes in ultraviolet radiation transparency associated with intensifying wet–dry cycles. Limnol. Oceanogr. Lett. 2025, 10, 485-494. [CrossRef]
- Qi, D.; Chao, Y.; Wu, R.; Xia, M.; Chen, Q.; Zheng, Z. Transcriptome Analysis Provides Insights Into the Adaptive Responses to Hypoxia of a Schizothoracine Fish (Gymnocypris eckloni). Front. Physiol. 2018, 9, 1326. [CrossRef]
- Guan, L.; Chi, W.; Xiao, W.; Chen, L.; He, S. Analysis of hypoxia-inducible factor alpha polyploidization reveals adaptation to Tibetan Plateau in the evolution of schizothoracine fish. BMC Evol. Biol. 2014, 14, 192. [CrossRef]
- Benson, A.; Zane, M.; Becker, T. E.; et al. Quantifying reaeration rates in alpine streams using deliberate gas tracer experiments. Water 2014, 6, 1013-1027. [CrossRef]
- Liang, Y.; Hou, Y.; Hu, W.; et al. Flow velocity preference of Schizothorax oconnori Lloyd swimming upstream. Glob. Ecol. Conserv. 2021, 32, e01902. [CrossRef]
- Mucha, S.; Chapman, L. J.; Krahe, R. Normoxia exposure reduces hemoglobin concentration and gill size in a hypoxia-tolerant tropical freshwater fish. Environ. Biol. Fishes 2023, 106, 1405-1423. [CrossRef]
- Jin, Y.; Zhu, B.; Wang, F.; et al. Analysis of water chemistry characteristics and main ion controlling factors of lakes in the Nagqu area of the Qinghai–Tibet plateau in summer. Water 2023, 15, 2900. [CrossRef]
- Tong, C.; Li, M. Genomic signature of accelerated evolution in a saline-alkaline lake-dwelling Schizothoracine fish. Int. J. Biol. Macromol. 2020, 149, 341-347. [CrossRef]
- Kousar, Y.; Singh, M.; Singh, D. Stock assessment of Schizothorax richardsonii (Gray, 1832) using geometric morphometrics and mitochondrial marker COX1 from tributaries of the Chenab River, India. J. Appl. Nat. Sci. 2025, 17, 600-613. [CrossRef]
- Gul, S.; Rashid, I. Morphological variations among Schizothorax species from Kashmir Himalayas. Zool. Anz. 2025, 316, 130-139. [CrossRef]
- Rajput, V.; Johnson, J. A.; Sivakumar, K. Environmental effects on the morphology of the snow trout Schizothorax richardsonii (Gray, 1832). TAPROBANICA: J. Asian Biodivers. 2013, 5, 102-110. [CrossRef]
- Martinez-Leiva, L.; Landeira, J. M.; Fatira, E.; Díaz-Pérez, J.; Hernández-León, S.; Roo, J.; Tuset, V. M. Energetic Implications of Morphological Changes between Fish Larval and Juvenile Stages Using Geometric Morphometrics of Body Shape. Animals 2023, 13, 370. [CrossRef]
- Reyes Corral, W. D.; Aguirre, W. E. Effects of temperature and water turbulence on vertebral number and body shape in Astyanax mexicanus (Teleostei: Characidae). PLoS One 2019, 14, e0219677. [CrossRef]
- Hetzel, C.; Forsythe, P. Phenotypic plasticity of a generalist fish species resident to lotic environments: Insights from the Great Lakes region. Ecol. Evol. 2023, 13, e10715. [CrossRef]
- Zhou, Y.; He, Z.; Cui, W.; Lu, Q.; Qin, J.; Han, Z.; Liu, J.; He, T. Strontium and Magnesium in Otoliths Can Trace Schizothorax grahami (Regan, 1904) Life History. Animals 2025, 15, 3170. [CrossRef]
- Gao, H.; Fu, S.; Xing, M.; et al. A high-throughput phenome-based analysis of morphological variation and environmental adaptation in extremely high-altitude schizothoracine fishes. Water Biol. Secur. 2025, 4, 100381. [CrossRef]
- Zhou, L.; Jin, Z. D.; Li, F. C. Mineralogy of the otoliths of naked carp Gymnocypris przewalskii (Kessler) from Lake Qinghai and its Sr/Ca potential implications for migratory pattern. Sci. China Earth Sci. 2012, 55, 983–990. [CrossRef]
- Belay, T. H.; Mengist, A. B. The application of otolith chemistry in fish life history assessment. Fish. Aquat. Sci. 2025, 28, 583-595. [CrossRef]
- Kratina, G. J.; DeVries, D. R.; Wright, R. A.; Peatman, E.; Rider, S. J.; Zhao, H. Using fish hard-part microchemistry and genetics to quantify population impacts of low-use lock-and-dam structures on the Alabama River. Trans. Am. Fish. Soc. 2023, 152, 490-512. [CrossRef]
- Wang, G.; Tang, Q.; Chen, Z.; Guo, D.; Zhou, L.; Lai, H.; Li, G. Otolith microchemistry and demographic history provide new insight into the migratory behavior and heterogeneous genetic divergence of coilia grayii in the pearl river. Fishes 2022, 7, 23. [CrossRef]
- Sturrock, A. M.; Hunter, E.; Milton, J. A.; Eimf; Johnson, R. C.; Waring, C. P.; Trueman, C. N. Quantifying physiological influences on otolith microchemistry. Methods Ecol. Evol. 2015, 6, 806-816. [CrossRef]
- Ma, B.; Xie, C.; Huo, B.; Yang, X.; Huang, H. Age and growth of a long-lived fish Schizothorax o'connori in the Yarlung Tsangpo River, Tibet. Zool. Stud. 2010, 49, 749-759. [CrossRef]
- Ma, B.; Xie, C.; Huo, B.; Yang, X.; Li, P. Age validation, and comparison of otolith, vertebra and opercular bone for estimating age of Schizothorax o’connori in the Yarlung Tsangpo River, Tibet. Environ. Biol. Fishes 2011, 90, 159-169. [CrossRef]
- Huo, B.; Ma, B. S.; Xie, C. X.; Duan, Y. J.; Yang, X. F.; Huang, H. P. Stock assessment and management implications of an endemic fish, Oxygymnocypris stewartii, in the Yarlung Zangbo River in Tibet, China. Zool. Stud. 2015, 54, 53. [CrossRef]
- Pfennig, M. B.; Crane, D. P.; Smith, N. G.; Buckmeier, D. L. Age estimation and validation in otoliths, spines, and fin rays from four central Texas fishes. N. Am. J. Fish. Manag. 2024, 44, 590-606. [CrossRef]
- Xu, L.; Zhou, Y.; Cui, W.; Lu, Q.; Liu, J.; Duan, C.; He, T. Characteristics of surface flow field and substrate in the spawning ground of Schizothorax grahami. Isr. J. Aquac. 2024, 76, 1-12. [CrossRef]
- Pan, H.; Liu, H.; Liu, F.; Xie, J.; Zhou, Y.; Zheng, Q.; Guo, M. Gut microbiota: a new frontier in understanding and protecting endangered plateau schizothorax fish. Front Microbiol. 2025, 16, 1592312. https://pubmed.ncbi.nlm.nih.gov/40584040/. [CrossRef]
- Zhong, C.; Chen, L.; Huang, Z.; Hu, Y.; Jiang, Y.; Zhou, J.; Long, X. Comparison of metabolism, gut histology, and microbiota between Schizothorax lissolabiatus and Schizothorax griseus under identical farming conditions. Front. Mar. Sci. 2024, 11, 1510780. [CrossRef]
- Mamoozadeh, N. R.; Wade, M. J.; Reid, B. N.; Bardwell, E.; Collins, E. E.; Hugentobler, S. A.; Jackson, S. A.; Kline, B. C.; Rothkopf, H. E.; Zhang, A.; Meek, M. H. A practical introduction to effective population size for fisheries management. Trans. Am. Fish. Soc. 2025, 154, 352-371. [CrossRef]
- Funk, W. C.; McKay, J. K.; Hohenlohe, P. A.; Allendorf, F. W. Harnessing genomics for delineating conservation units. Trends Ecol. Evol. 2012, 27, 489-496. [CrossRef]
- Semlitsch, R. D.; Hotz, H.; Guex, G. D. Competition among tadpoles of coexisting hemiclones of hybridogenetic Rana esculenta: support for the frozen niche variation model. Evolution 1997, 51, 1249-1261. [CrossRef]
- Liao, J.; Chen, S.; Liu, P.; Fontaneto, D.; Han, B. P. Environmental selection and gene flow jointly determine the population genetic diversity and structure of Diaphanosoma dubium along a watershed elevation. Glob. Ecol. Conserv. 2024, 49, e02773. [CrossRef]
- Yan, T.; Gao, K.; He, L.; Pu, Y.; Tang, Z.; Xiong, J.; Chen, Q.; Lai, B.; Liu, F.; Chen, P.; Chen, M.; Luo, W.; Huang, J.; Ding, W.; Yang, D.; He, Z. Genomic-based revelation of genetic structure and adaptive characterization of Schizopygopsis malacanthus in the Jinsha River and Yalong River. BMC Genomics 2025, 26, 870. [CrossRef]
- He, J.; He, Z.; Yang, D.; Ma, Z.; Chen, H.; Zhang, Q.; Deng, F.; Ye, L.; Pu, Y.; Zhang, M.; Yang, S.; Yang, S.; Yan, T. Genetic Variation in Schizothorax kozlovi Nikolsky in the Upper Reaches of the Chinese Yangtze River Based on Genotyping for Simplified Genome Sequencing. Animals 2022, 12, 2181. [CrossRef]
- Wang, F.; Wang, L.; Liu, D.; Gao, Q.; Nie, M.; Zhu, S.; Chao, Y.; Yang, C.; Zhang, C.; Yi, R.; et al. Chromosome-level assembly of Gymnocypris eckloni genome. Sci. Data 2022, 9, 464. [CrossRef]
- Wang, Q.; Lan, T.; Li, H.; Sahu, S. K.; Shi, M.; Zhu, Y.; Han, L.; Yang, S.; Li, Q.; Zhang, L.; Deng, Z.; Liu, H.; Hua, Y. Whole-genome resequencing of Chinese pangolins reveals a population structure and provides insights into their conservation. Commun. Biol. 2022, 5, 821. [CrossRef]
- Wedekind, C. Sexual selection and life-history decisions: implications for supportive breeding and the management of captive populations. Conserv. Biol. 2002, 16, 1204-1211. http://www.jstor.org/stable/3095316.
- Han, H.; Wang, L.; Zhang, C.; Li, H.; Ma, B. Population Structure, Growth Characteristics, Resource Dynamics, and Management Strategies of Schizopygopsis younghusbandi in Four Tributaries of the Yarlung Zangbo River, Tibet. Biology 2025, 14, 707. [CrossRef]
- Hixon, M. A.; Johnson, D. W.; Sogard, S. M. BOFFFFs: on the importance of conserving old-growth age structure in fishery populations. ICES J. Mar. Sci. 2014, 71, 2171-2185. [CrossRef]
- Cooke, S. J.; Hinch, S. G. Improving the reliability of fishway attraction and passage efficiency estimates to inform fishway engineering, science, and practice. Ecol. Eng. 2013, 58, 123-132. [CrossRef]
- Bunt, C. M.; Castro-Santos, T.; Haro, A. Performance of fish passage structures at upstream barriers to migration. River Res. Appl. 2012, 28, 457-478. [CrossRef]
- Frankham, R.; Ballou, J. D.; Eldridge, M. D.; Lacy, R. C.; Ralls, K.; Dudash, M. R.; Fenster, C. B. Predicting the probability of outbreeding depression. Conserv. Biol. 2011, 25, 465-475. https://10.1111/j.1523-1739.2011.01662.x.
- Humphries, P.; King, A.; McCasker, N.; Kopf, R. K.; Stoffels, R.; Zampatti, B.; Price, A. Riverscape recruitment: a conceptual synthesis of drivers of fish recruitment in rivers. Can. J. Fish. Aquat. Sci. 2020, 77, 213-225. [CrossRef]
- King, A. J.; Gwinn, D. C.; Tonkin, Z.; Mahoney, J.; Raymond, S.; Beesley, L. Using abiotic drivers of fish spawning to inform environmental flow management. J. Appl. Ecol. 2016, 53, 34-43. [CrossRef]
- Krellwitz, E. M.; Gido, K. B.; Mehl, H. E.; Totten, L. A.; Jones, T. C. Abiotic Drivers of Spawning and Early Life Stage Assemblage Dynamics of Great Plains Fishes. River Res. Appl. 2026, 42, 3-18. [CrossRef]
- Liu, Q. Y.; Li, J.; An, R. D.; Li, Y. Ecohydraulogical characteristic index system of Schizopygopsis younghusbandi during spawning periods in the Yarlung Tsangpo River. Int. J. Environ. Res. Public. Health 2018, 15, 1949. [CrossRef]
- Laikre, L.; Schwartz, M. K.; Waples, R. S.; Ryman, N.; GeM Working Group. Compromising genetic diversity in the wild: unmonitored large-scale release of plants and animals. Trends Ecol. Evol. 2010, 25, 520-529. https://doi: 10.1016/j.tree.2010.06.013.
- Palmé, A. Assessing and monitoring genetic patterns for conservation purposes with special emphasis on Scandinavia. Department of Zoology, Stockholm University, 2010. https://www.diva-portal.org/smash/get/diva2:353156/FULLTEXT01.pdf.
- Nie, M. A.; Schultz, C. A. Decision-making triggers in adaptive management. Conserv. Biol. 2012, 26, 1137-1144. http://www.jstor.org/stable/23360129.
Figure 1.
An environment–evolution–conservation framework for Schizothoracine fishes in plateau river networks. Single-headed arrows indicate causality or a process flow; double-headed arrows indicate dynamic change or feedback; grey arrows indicate feedback effects acting back on upstream components. UV = Ultraviolet radiation, DO = Dissolved oxygen, WGD = Whole-genome duplication, TE = Transposable element, SV = Structural variation, MU = Management unit, ESU = Evolutionarily significant unit.
Figure 1.
An environment–evolution–conservation framework for Schizothoracine fishes in plateau river networks. Single-headed arrows indicate causality or a process flow; double-headed arrows indicate dynamic change or feedback; grey arrows indicate feedback effects acting back on upstream components. UV = Ultraviolet radiation, DO = Dissolved oxygen, WGD = Whole-genome duplication, TE = Transposable element, SV = Structural variation, MU = Management unit, ESU = Evolutionarily significant unit.
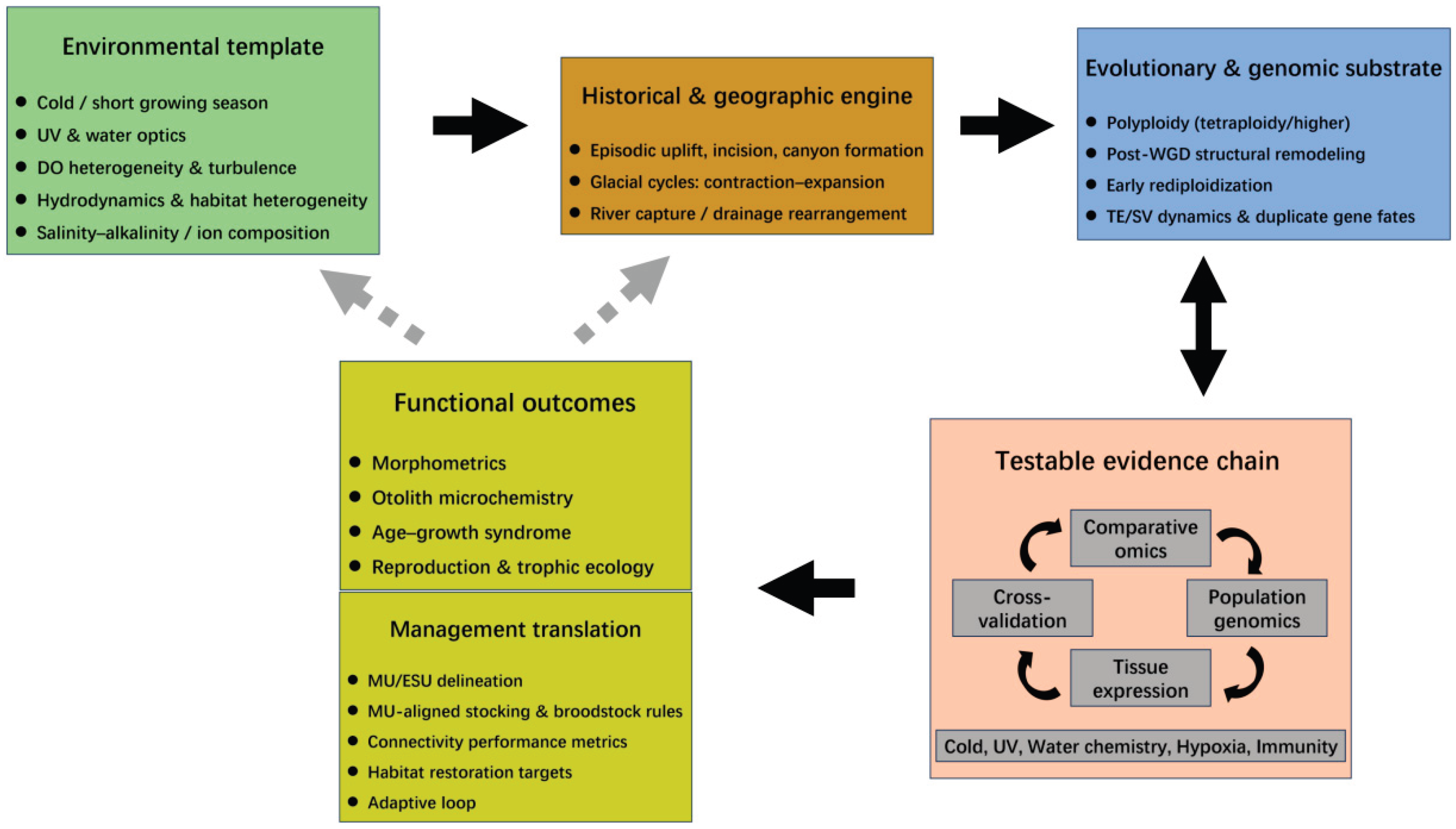
Figure 2.
Key tissues for mechanistic inference and stress-response profiling in Schizothoracine fishes. UV = Ultraviolet radiation, DO = Dissolved oxygen, OXPHOS = Oxidative phosphorylation, NER = Nucleotide excision repair, SLC = Solute carrier family, HSPs = Heat shock proteins, ROS = Reactive oxygen species.
Figure 2.
Key tissues for mechanistic inference and stress-response profiling in Schizothoracine fishes. UV = Ultraviolet radiation, DO = Dissolved oxygen, OXPHOS = Oxidative phosphorylation, NER = Nucleotide excision repair, SLC = Solute carrier family, HSPs = Heat shock proteins, ROS = Reactive oxygen species.
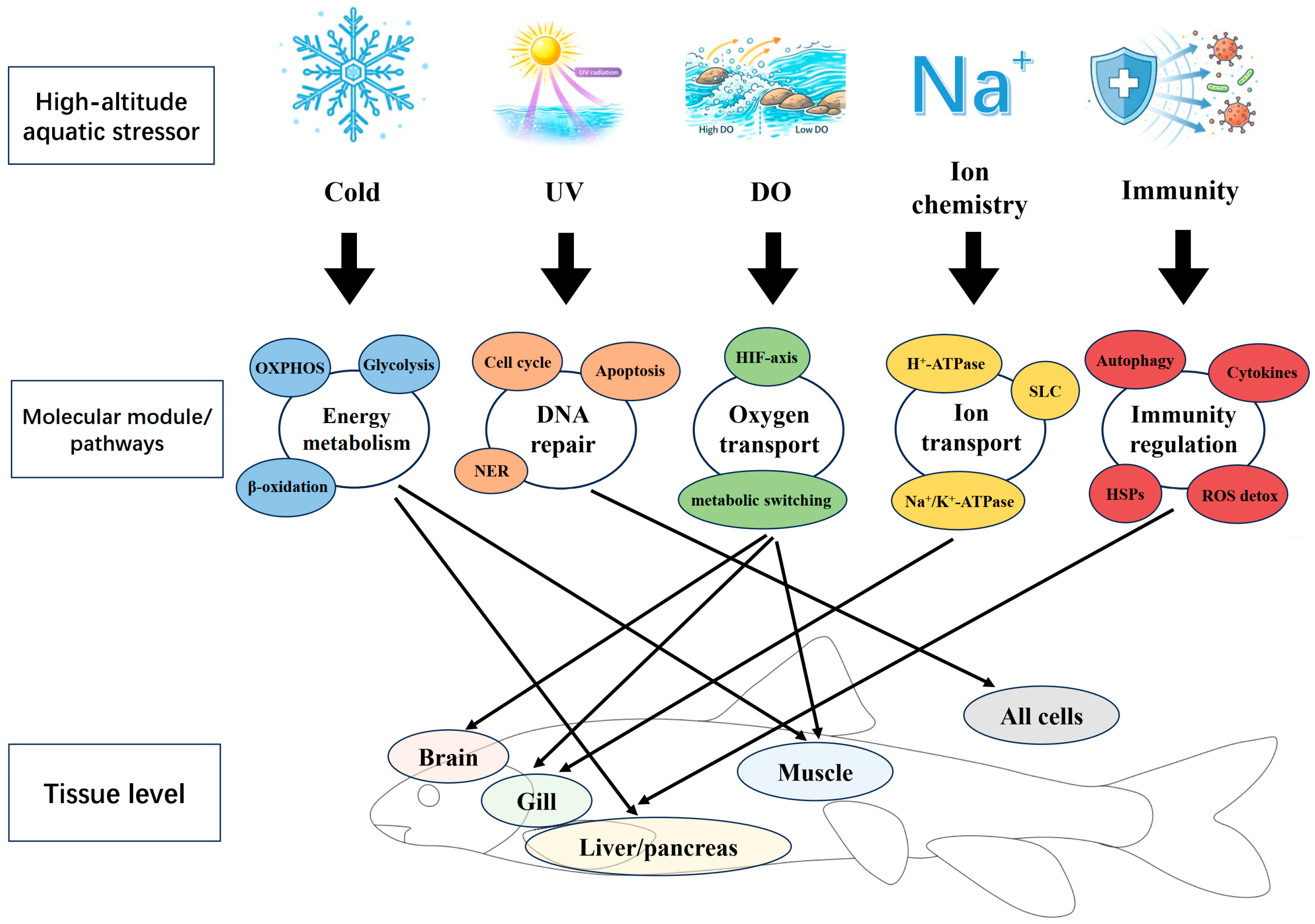

Table 1.
Translating MU/ESU evidence into actionable management measures and quantified performance indicators.
Table 1.
Translating MU/ESU evidence into actionable management measures and quantified performance indicators.
| Objective | Action (minimum) | Metric (quantified) | Focus | Data | Rule | Reference |
| Structure protection | Closed season + Min size | Size/age rebound; recruitment ↑ | Key reaches; spawning season | P | No improv. ×2 yrs → tighten | [13,113,114,124] |
| Connectivity | Set passage target | Arrival efficiency; passage efficiency; number of passable days |
Migration season; representative reaches | C | No improv. ×2 yrs → redesign/adjust | [99,115,116,122,123,124] |
| Stocking risk control | MU-aligned source & release | Source compliance; anomaly warning | Transition zone | G + R | Violation → stop/rewire | [104,105,117] |
| Habitat restoration | Microhabitat restoration | Depth/ velocity/substrate/cover area | Key reach | H | No improv. ×2 yrs → relocate/retune | [118,119,120,121,124] |
This table summarizes a minimum set of implementable management actions and auditable metrics for Schizothoracine conservation, based on the evidence chain synthesized in Chapters 7–9. Data codes: G, genetics/population genomics (SNP/GBS/WGS; MU delineation and risk assessment); P, population status (juvenile recruitment; size/age structure); C, connectivity monitoring (arrival/passage efficiency; passable days); H, habitat survey (depth, velocity, substrate, cover structures, and effective area); R, stocking registry (broodstock/juvenile origin records).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



