
Submitted:
19 January 2026
Posted:
21 January 2026
You are already at the latest version
Abstract
The nutritional and functional quality of eggplants is heavily influenced by cultivar selection and fertilization techniques. This study evaluated the effects of cultivar, grafting, and fertilization regime on the nutritional and functional quality of eggplant (Solanum melongena L.) cultivated under plastic tunnel conditions. Two commercial hybrids (Mirval and Black Pearl) were assessed under two grafting treatments (Solanum sylvatica and Rezistar F1) and three fertilization regimes: biological (Micoseed®), organic (Orgevit®), and chemical (Nutrispore®), with ungrafted and unfertilized plants used as controls. Fruit samples were analyzed for total polyphenol content (TPC), antioxidant capacity (ABTS and DPPH assays), chlorophyll a and b, lycopene, and tannin concentrations. Mirval exhibited significantly higher levels of TPC, antioxidant activity, photosynthetic pigments, and lycopene than Black Pearl. Ungrafted plants and those grafted onto S. sylvatica showed superior fruit quality, particularly when combined with organic or chemical fertilization. The highest concentrations of polyphenols (3.61 mg·100 g⁻¹ fresh weight), lycopene (3.18 mg·100 g⁻¹ fresh weight), and total chlorophylls were recorded in Mirval fruits from ungrafted or S. sylvatica-grafted plants under organic or chemical fertilization. These findings indicate that optimizing cultivar–rootstock combinations and fertilization management can significantly enhance the functional quality of eggplant fruits, and support the use of organic fertilization as a sustainable strategy in protected vegetable production systems.
Keywords:
eggplant cultivars
; functional quality
; bioactive compounds
; antioxidant activity
; polyphenol content
; lycopene
; organic farming
; plastic tunnel cultivation
The nutritional and functional quality of eggplants is heavily influenced by cultivar selection and fertilization techniques. This study evaluated the effects of cultivar, grafting, and fertilization regime on the nutritional and functional quality of eggplant (Solanum melongena L.) cultivated under plastic tunnel conditions. Two commercial hybrids (Mirval and Black Pearl) were assessed under two grafting treatments (Solanum sylvatica and Rezistar F1) and three fertilization regimes: biological (Micoseed®), organic (Orgevit®), and chemical (Nutrispore®), with ungrafted and unfertilized plants used as controls. Fruit samples were analyzed for total polyphenol content (TPC), antioxidant capacity (ABTS and DPPH assays), chlorophyll a and b, lycopene, and tannin concentrations. Mirval exhibited significantly higher levels of TPC, antioxidant activity, photosynthetic pigments, and lycopene than Black Pearl. Ungrafted plants and those grafted onto S. sylvatica showed superior fruit quality, particularly when combined with organic or chemical fertilization. The highest concentrations of polyphenols (3.61 mg·100 g⁻¹ fresh weight), lycopene (3.18 mg·100 g⁻¹ fresh weight), and total chlorophylls were recorded in Mirval fruits from ungrafted or S. sylvatica-grafted plants under organic or chemical fertilization. These findings indicate that optimizing cultivar–rootstock combinations and fertilization management can significantly enhance the functional quality of eggplant fruits, and support the use of organic fertilization as a sustainable strategy in protected vegetable production systems.
1. Introduction
Eggplant (Solanum melongena L.) is cultivated predominantly in subtropical regions, which account for approximately 94% of global production. Due to its wide popularity and economic relevance, it is often referred to as the “king of vegetables”. According to the Food and Agriculture Organization of the United Nations (FAO) [1], China and India are the world’s leading producers, with annual outputs of approximately 28 and 13 million tons, respectively. In Europe, production is mainly concentrated in Turkey (827,000 t), Italy (220,000 t), Spain (206,000 t), and Romania (123,000 t per year) [1]. Globally, eggplant ranks sixth among vegetable crops in total production, after tomato, watermelon, onion, cucumber, and cabbage, with 52.3 million tons produced in 2017 (FAO, 2019) [2]. Furthermore, eggplant is included among the 35 most important food crops for global food security and is listed in Annex I of the International Treaty on Plant Genetic Resources for Food and Agriculture [3].
Eggplant has high gastronomic and economic value due to its broad ecological adaptability, consumer acceptance, and suitability for diverse cultivation systems [4]. The fruit contains carbohydrates, sugars, proteins, and vitamin C, contributing to both palatability and nutritional value. Approximately 65–70% of global production is processed by the canning industry, while consumption also includes cooked, stuffed, marinated, and fried products [5]. Owing to its relatively low cost, eggplant constitutes an important dietary component for low-income populations [6]. In recent years, interest in eggplant has increased because of its high content of antioxidants, particularly phenolic compounds, which are associated with beneficial effects on human health [7]. In addition, traditional medicinal uses of eggplant and related Solanum species have been reported, including treatments for skin disorders, digestive problems, and urinary conditions, as well as applications as purgatives and tonics in several Asian regions [8].
Plant nutrition plays a central role in eggplant growth, productivity, and fruit quality. Over the past three decades, numerous cultivars and hybrids have been introduced into European and Romanian markets; however, many show limited adaptation to changing climatic conditions and shifting consumer preferences. Recent studies indicate that cultivar selection, fertilization strategy, and irrigation regime are key factors affecting both yield and quality in organic and conventional systems [4]. Lycopene and other carotenoids present in eggplant fruits are associated with reduced risks of cancer and cardiovascular diseases, largely due to their antioxidant properties [9]. Tannins, also known as proanthocyanidins, represent another important group of bioactive compounds, exhibiting antioxidant, anti-inflammatory, anticancer, anti-atherosclerotic, and cardioprotective effects [10].
Modern agricultural systems often prioritize yield maximization, sometimes at the expense of environmental sustainability and long-term soil fertility [11]. Increasing consumer demand for healthier foods and stricter regulations on environmental protection have accelerated the transition toward sustainable agricultural practices, including organic farming [12]. As a result, the global area under organic cultivation and the organic food market have expanded steadily, with a further increase in consumer demand observed during the COVID-19 pandemic [13]. Organic production systems are considered beneficial due to reduced chemical inputs and improved soil health, contributing to enhanced ecosystem resilience and potentially improved nutritional quality of produce [14].
Fertilization strongly influences soil properties, plant physiological processes, photosynthetic efficiency, and the accumulation of bioactive compounds such as chlorophylls and flavonoids in fruits [15]. Organic fertilizers and biological amendments improve soil structure and microbial activity, while chemical fertilizers provide rapid nutrient availability and high yields [16,17,18]. Therefore, selecting appropriate fertilization strategies is critical for optimizing both productivity and nutritional quality. Increasing research efforts have focused on identifying cultivation practices that enhance fruit functional properties while maintaining sustainable production systems [19,20].
Protected cultivation systems, including greenhouses and plastic tunnels, are increasingly adopted to mitigate climatic variability and extend production seasons, particularly in regions with cooler climates such as Romania [21]. Previous studies have demonstrated that suitable combinations of cultivar selection and fertilization management can significantly enhance the accumulation of bioactive compounds under varying climatic conditions [21,22]. However, comparative studies evaluating the combined effects of fertilization regime and cultivation system on the nutritional quality of eggplant under protected conditions remain limited.
Therefore, this study aimed to assess the effects of fertilization regime and agricultural system (organic versus conventional) on the nutritional and functional quality of two eggplant hybrids, Mirval and Black Pearl, selected for their adaptability to Romanian environmental conditions, pest resistance, high productivity, and fruit quality. The experiment was conducted under greenhouse conditions at the “V. Adamachi” Farm of the Iași University of Life Sciences, while biochemical analyses were performed at the Andalusian Institute of Agricultural and Fisheries Research and Training (IFAPA), Córdoba, Spain. Antioxidant activity, total polyphenol content, chlorophyll a and b, lycopene, and total carotenoids, as well as their ratios, were evaluated to determine the influence of fertilization strategies on eggplant fruit nutritional quality under protected cultivation.
2. Materials and Methods
2.1. Experimental Site and Design
The experiment was carried out under plastic tunnel conditions at the “V. Adamachi” Farm of the Iași University of Life Sciences, Romania (47°19′25″ N, 27°54′99″ E; 150 m a.s.l.), during the 2021–2022 growing seasons. The study included three experimental factors: cultivar, grafting, and fertilization regime. Two eggplant (Solanum melongena L.) hybrids, Mirval F1 and Black Pearl F1, were used as scions. Grafting treatments included two rootstocks (Solanum sylvatica and Rezistar F1), as well as ungrafted plants. Fertilization treatments consisted of three regimes: biological (Micoseed®), organic (Orgevit®), and chemical (Nutrispore®), compared with unfertilized control.
The experiment was organized in a split-plot design with three replicates (n = 3). Cultivars constituted factor A, while fertilization regimes represented factor B: B1—unfertilized control (Ct), B2—Micoseed® (Mo), B3—Orgevit® (O), and B4—Nutrispore® (Ch). Each experimental unit consisted of equal numbers of plants, and border plants were excluded from sampling.
The soil was classified as silty-clay chernozem, with the following characteristics: pH 7.20, electrical conductivity (EC) 482 μS cm⁻¹, CaCO₃ 0.42%, organic matter 28.32 mg kg⁻¹, C/N ratio 5.87, total nitrogen 2.8 g kg⁻¹, and available phosphorus 34 mg kg⁻¹.
2.2. Plant Production and Crop Management
Seedlings were produced in multicellular trays under organic-compliant conditions and transplanted into the plastic tunnel at 55 days after sowing, in mid-April, at a density of 2.5 plants m⁻². Standard cultural practices were applied throughout the growing period, including plant-training, pruning, and integrated pest and disease management, in accordance with recommended horticultural practices [2,4]. The cultivation cycle ended on 31 October.
2.3. Fertilization Treatments
Chemical fertilization (Ch) was applied using Nutrispore® NPK 20–20–20 at 200 kg ha⁻¹ prior to transplanting, followed by two applications of Nutrispore® NPK 9–18–27 + 2 MgO at 300 kg ha⁻¹ each during the growing season.
Organic fertilization (O) consisted of commercial chicken manure (Orgevit®), applied at a total rate of 2000 kg ha⁻¹, and with 1250 kg ha⁻¹ incorporated before transplanting and the remainder applied 30 days after planting. Nutrient equivalence between organic and chemical treatments was adjusted, considering that approximately 70% of nutrients from organic fertilizers are available to plants in the first year.
Biological fertilization (Mo) was applied using Micoseed® at 30 kg ha⁻¹, split into two equal applications before transplanting and 30 days after planting, combined with 5 L ha⁻¹ Nutryaction®, following the manufacturer’s recommendations. The biological formulation contains arbuscular mycorrhizal fungi (Claroideoglomus etunicatum, Funneliformis mosseae, Glomus aggregatum, Rhizophagus intraradices), along with beneficial microorganisms from the genera Trichoderma, Streptomyces, Bacillus, and Pseudomonas.
2.4. Sample Collection and Preparation
At physiological maturity, five fruits were collected from each experimental unit, resulting 120 fruit samples from 24 treatment combinations. Fruits were cut into approximately 1 cm pieces, homogenized, and freeze-dried using an ECO EVO freeze dryer (Tred Technology S.R.L., Ripalimosani, Italy). The dried samples were ground into powder and stored at −80 °C until chemical analysis.
2.5. Chemicals and Reagents
HPLC-grade methanol, formic acid, hydrochloric acid, acetone, hexane, and water were obtained from Panreac (Barcelona, Spain). Sodium carbonate, Folin–Ciocalteu reagent, gallic acid, Trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid), DPPH (2,2-diphenyl-1-picrylhydrazyl), and ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)) were supplied by Sigma-Aldrich (Steinheim, Germany). Potassium persulfate, (+)-catechin, and vanillin were also obtained from Sigma-Aldrich.
2.6. Hydrophilic Extraction
Hydrophilic compounds were extracted using deionized water–methanol (20:80, v/v) containing 1% formic acid. Lyophilized samples (0.2 g) were extracted with 1 mL solvent, sonicated for 10 min, and centrifuged at 15,000 rpm for 15 min. The supernatant was collected, and the residue was re-extracted under identical conditions. Combined extracts were stored at −80 °C until analysis of antioxidant activity, total polyphenols, and condensed tannins.
2.7. Determination of Total Polyphenol Content
Total polyphenol content (TPC) was determined using the Folin–Ciocalteu method as described by Slinkard and Singleton [23]. Briefly, 10 μL of extract was mixed with 175 μL distilled water and 12 μL Folin–Ciocalteu reagent. After 3 min, 30 μL of 20% sodium carbonate solution was added. Samples were incubated for 1 h, and absorbance was measured at 765 nm. Results were expressed as mg gallic acid equivalents per 100 g dry weight (mg GAE·100 g⁻¹ DW).
2.8. Antioxidant Activity
Antioxidant capacity was evaluated using ABTS and DPPH assays with a Synergy HTX multimode microplate reader (Biotek Instruments, Winooski, VT, USA), following the method described in [24]. Results were expressed as mmol Trolox equivalents per 100 g dry weight (mmol TE·100 g⁻¹ DW).
2.9. Determination of Condensed Tannins
Condensed tannins were quantified using the vanillin–HCl method described by Deepa et al. [25], with minor modifications. Hydrophilic extracts were mixed with vanillin–methanol solution (4% w/v) and concentrated hydrochloric acid and incubated for 20 min in the dark at room temperature. Absorbance was measured at 500 nm, and results were expressed as mg catechin equivalents per 100 g dry weight.
2.10. Pigment Extraction and Analysis
Lipophilic pigments were extracted following the method described by [26]. Lyophilized samples (0.2 g) were extracted with 1 mL hexane–acetone (4:6, v/v) in the dark, centrifuged at 15,000 rpm for 15 min, and re-extracted. Absorbance was measured at 453, 505, 645, and 663 nm using a Synergy HTX microplate reader. Concentrations of chlorophyll a, chlorophyll b, β-carotene, and lycopene were calculated using published equations [27,28].
2.11. Statistical Analysis
Data were analyzed using analysis of variance (ANOVA). Mean comparisons were performed using Tukey’s post hoc test at a significance level of p < 0.05. Statistical analyses were conducted using SPSS and Statistix v.9.0 software.
3. Results
3.1. Effect of Cultivar on Fruit Quality
The majority of quality parameters showed significant varietal differences (Table 1). Most quality traits differed significantly between the two hybrids (Table 1). Overall, Mirval F1 showed higher values than Black Pearl F1 for total polyphenol content (TPC), DPPH radical scavenging activity, chlorophyll a, chlorophyll b, lycopene, and tannins (p ≤ 0.05). Mirval accumulated 38% more TPC than Black Pearl (2.97 vs. 2.15 mg GAE·100 g⁻¹ DW). Similarly, Mirval exhibited higher DPPH activity (1.23 vs. 1.20 mmol TE·100 g⁻¹ DW) and markedly higher pigment concentrations, including chlorophyll a (3.44 vs. 2.19 mg·100 g⁻¹ DW), chlorophyll b (4.74 vs. 2.82 mg·100 g⁻¹ DW), and lycopene (1.58 vs. 0.93 mg·100 g⁻¹ DW). Tannin concentration followed the same direction, although the absolute difference was small (0.10 vs. 0.10 mg CE·100 g⁻¹ DW; p ≤ 0.05). In contrast, ABTS activity did not differ between cultivars (p ≥ 0.05). These results indicate that Mirval F1 possesses a stronger antioxidant and pigment profile, suggesting greater potential for improving nutritional quality under protected cultivation.
3.2. Effect of Grafting on Fruit Quality.
Grafting significantly affected all measured parameters (Table 2). In general, ungrafted plants produced fruits with the highest pigment concentrations and lycopene, whereas Rezistar F1 resulted in the lowest pigment values. TPC was highest in ungrafted plants (2.70 mg GAE·100 g⁻¹ DW), significantly exceeding Rezistar F1 (2.51 mg GAE·100 g⁻¹ DW) and S. sylvatica (2.52 mg GAE·100 g⁻¹ DW; p ≤ 0.05). In contrast, antioxidant capacity was highest in fruits grafted onto S. sylvatica, which showed the greatest ABTS (1.03 mmol TE·100 g⁻¹ DW) and DPPH (1.24 mmol TE·100 g⁻¹ DW) activity. Rootstock also strongly influenced pigments: ungrafted plants had the highest chlorophyll a (3.63 mg·100 g⁻¹ DW) and chlorophyll b (4.97 mg·100 g⁻¹ DW), whereas Rezistar F1 had the lowest values (2.10 and 2.66 mg·100 g⁻¹ DW, respectively). Lycopene was highest in ungrafted plants (1.67 mg·100 g⁻¹ DW) and lowest in Rezistar F1 (0.89 mg·100 g⁻¹ DW). Tannins varied modestly across grafting treatments but differed significantly (p ≤ 0.05). Collectively, these findings suggest that ungrafted plants favor pigment and lycopene accumulation, while S. sylvatica enhances radical scavenging activity.
3.3. Effect of Fertilization on Fruit Quality.
Fertilization significantly influenced most traits, except TPC (Table 3). TPC values were not significantly different among fertilization regimes (p > 0.05). However, antioxidant activity, pigments, lycopene, and tannins responded to fertilization. ABTS activity was highest under biological fertilization (1.02 mmol TE·100 g⁻¹ DW) and lowest under chemical fertilization (0.96 mmol TE·100 g⁻¹ DW; p ≤ 0.05). For DPPH, the unfertilized control showed the highest activity (1.25 mmol TE·100 g⁻¹ DW), significantly exceeding the biological treatment (1.19 mmol TE·100 g⁻¹ DW). Chlorophyll concentrations were also affected: the unfertilized control produced the highest chlorophyll a (3.30 mg·100 g⁻¹ DW) and chlorophyll b (4.44 mg·100 g⁻¹ DW), whereas biological fertilization resulted in the lowest pigment values. Lycopene followed a similar pattern, with the highest level in the unfertilized control (1.47 mg·100 g⁻¹ DW) and the lowest in the biological regime (1.03 mg·100 g⁻¹ DW). Tannins were highest in the unfertilized control (0.11 mg CE·100 g⁻¹ DW) and lowest under chemical fertilization (0.10 mg CE·100 g⁻¹ DW; p ≤ 0.05). Overall, the unfertilized treatment favored pigment, lycopene, and tannin accumulation, whereas biological fertilization enhanced ABTS activity.
3.4. Interaction Effects of Cultivar × Grafting on Fruit Quality
The cultivar × grafting interaction significantly affected all traits (Table 4). In general, Mirval combined with either ungrafted plants or S. sylvatica produced superior outcomes compared with other combinations, whereas Black Pearl grafted onto Rezistar F1 or S. sylvatica tended to show lower values for several parameters. Mirval × S. sylvatica had the highest TPC (3.03 mg GAE·100 g⁻¹ DW), while Black Pearl × S. sylvatica showed the lowest (2.01 mg GAE·100 g⁻¹ DW; p ≤ 0.05). Antioxidant activity also varied significantly: Mirval × S. sylvatica exhibited the highest ABTS activity (1.05 mmol TE·100 g⁻¹ DW), whereas Mirval × ungrafted had the lowest (0.93 mmol TE·100 g⁻¹ DW). For DPPH, Mirval × Rezistar F1 and Black Pearl × S. sylvatica showed the highest values (p ≤ 0.05).
Pigment accumulation was strongly interaction-dependent. Mirval × ungrafted showed the highest chlorophyll a (4.70 mg·100 g⁻¹ DW), chlorophyll b (6.65 mg·100 g⁻¹ DW), and lycopene (2.24 mg·100 g⁻¹ DW). The lowest pigment values were observed in combinations involving Rezistar F1. These results indicate that Mirval exhibits greater compatibility with ungrafted and S. sylvatica treatments, promoting higher pigment and lycopene accumulation compared with Black Pearl.
3.5. Interaction Effects of Cultivar × Fertilization on Fruit Quality
The cultivar × fertilization interaction significantly affected all measured parameters (Table 5). Mirval generally exhibited stronger responses than Black Pearl across fertilization regimes, particularly under chemical and organic fertilization. Mirval × chemical showed the highest TPC (3.11 mg GAE·100 g⁻¹ DW), followed by Mirval × biological and Mirval × organic (≥3.00 mg GAE·100 g⁻¹ DW), whereas Black Pearl under organic or chemical fertilization had the lowest values (≤2.10 mg GAE·100 g⁻¹ DW; p ≤ 0.05). ABTS activity was highest in Mirval × organic (1.06 mmol TE·100 g⁻¹ DW) and Black Pearl × unfertilized (1.07 mmol TE·100 g⁻¹ DW), while Black Pearl × organic and Black Pearl × chemical showed the lowest ABTS values (≤0.89 mmol TE·100 g⁻¹ DW). Pigment and lycopene values were markedly higher in Mirval × chemical compared with Black Pearl × chemical (chlorophyll a: 4.22 vs. 1.85 mg·100 g⁻¹ DW; chlorophyll b: 6.11 vs. 2.09 mg·100 g⁻¹ DW; lycopene: 2.04 vs. 0.69 mg·100 g⁻¹ DW). Tannins varied less, but Black Pearl × unfertilized had the highest value (0.12 mg CE·100 g⁻¹ DW), significantly exceeding Black Pearl × chemical (0.09 mg CE·100 g⁻¹ DW). Overall, Mirval benefited most from chemical and organic fertilization for pigment and lycopene accumulation, whereas Black Pearl responded more weakly to fertilization.
3.6. Interaction Effects of Grafting × Fertilization on Fruit Quality
The grafting × fertilization interaction significantly affected all traits (Table 6). TPC was highest in S. sylvatica × unfertilized (3.11 mg GAE·100 g⁻¹ DW) and ungrafted × organic (2.98 mg GAE·100 g⁻¹ DW), while the lowest value occurred in S. sylvatica × organic (1.92 mg GAE·100 g⁻¹ DW; p ≤ 0.05). ABTS activity reached its maximum under Rezistar F1 × biological (1.12 mmol TE·100 g⁻¹ DW) and S. sylvatica × unfertilized (1.08 mmol TE·100 g⁻¹ DW), whereas Rezistar F1 × chemical and Rezistar F1 × unfertilized showed the lowest ABTS values (≤0.90 mmol TE·100 g⁻¹ DW). DPPH was highest in S. sylvatica × unfertilized (1.37 mmol TE·100 g⁻¹ DW) and S. sylvatica × chemical (1.36 mmol TE·100 g⁻¹ DW).
Pigment concentrations also differed substantially among treatment combinations. The highest chlorophyll a and b values were obtained in the ungrafted × unfertilized control (4.41 and 5.94 mg·100 g⁻¹ DW, respectively) and ungrafted × organic. In contrast, Rezistar F1 combined with organic or chemical fertilization resulted in the lowest pigment concentrations (chlorophyll a ≤ 1.65 mg·100 g⁻¹ DW; chlorophyll b ≤ 1.96 mg·100 g⁻¹ DW). Lycopene was highest in ungrafted × organic (2.08 mg·100 g⁻¹ DW) and S. sylvatica × chemical (1.95 mg·100 g⁻¹ DW), while the lowest level occurred in Rezistar F1 × organic (0.62 mg·100 g⁻¹ DW). Tannins were highest in ungrafted × organic (0.12 mg CE·100 g⁻¹ DW) and lowest in S. sylvatica × organic and S. sylvatica × chemical (0.08 mg CE·100 g⁻¹ DW). Overall, ungrafted plants combined with organic fertilization enhanced phenolics, lycopene, and tannins, whereas S. sylvatica under unfertilized conditions promoted higher antioxidant activity; Rezistar F1, particularly with organic or chemical fertilization, was associated with lower pigment and lycopene accumulation.
3.7. Interaction Effects of Cultivar × Grafting × Fertilization on Fruit Quality
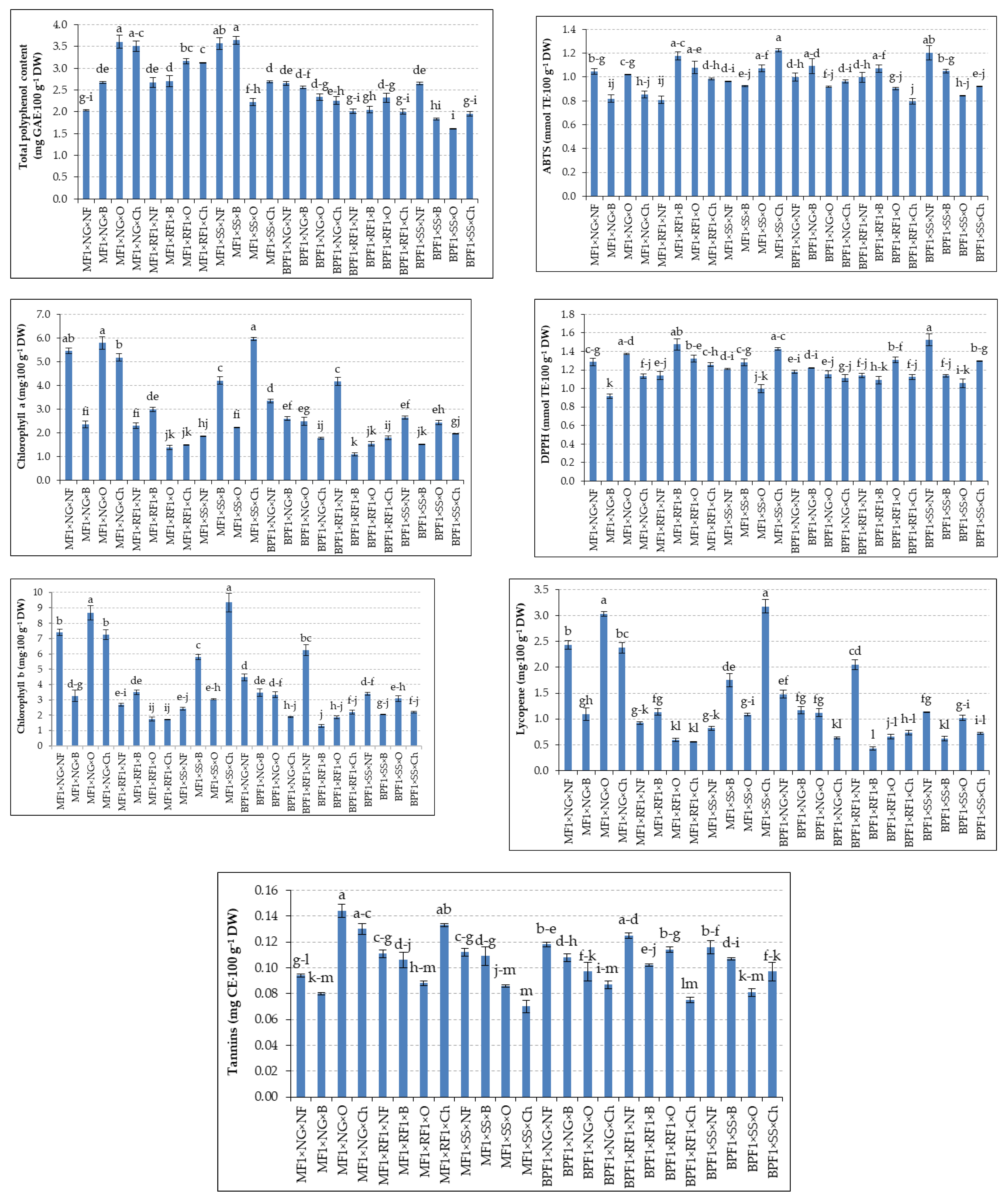
The cultivar × grafting × fertilization interaction significantly affected all evaluated traits (Figure 1). The highest polyphenol concentration (3.61 mg·100 g⁻¹ DW) occurred under Mirval × ungrafted × organic fertilization, whereas the lowest values were observed in Black Pearl combinations involving S. sylvatica × organic fertilization. ABTS activity was maximal in Mirval × S. sylvatica × chemical (1.22 mmol TE·100 g⁻¹ DW) and Mirval × Rezistar F1 × biological (1.18 mmol TE·100 g⁻¹ DW), while Black Pearl × Rezistar F1 × chemical showed the lowest value (0.80 mmol TE·100 g⁻¹ DW). DPPH activity peaked in Black Pearl × S. sylvatica × unfertilized (1.52 mmol TE·100 g⁻¹ DW) and Mirval × Rezistar F1 × biological (1.48 mmol TE·100 g⁻¹ DW), whereas Mirval × ungrafted × biological had the lowest value (0.92 mmol TE·100 g⁻¹ DW).
Pigment accumulation exhibited pronounced interaction effects. The highest chlorophyll a and chlorophyll b were recorded for Mirval × S. sylvatica × chemical (5.96 and 9.34 mg·100 g⁻¹ DW, respectively) and Mirval × ungrafted × organic (5.80 and 8.69 mg·100 g⁻¹ DW). The lowest pigment levels occurred primarily in treatments involving Rezistar F1 under organic or biological fertilization. Lycopene content was highest in Mirval × S. sylvatica × chemical (3.18 mg·100 g⁻¹ DW) and Mirval × ungrafted × organic (3.04 mg·100 g⁻¹ DW), whereas Black Pearl × Rezistar F1 × biological showed the minimum concentration (0.43 mg·100 g⁻¹ DW). Tannins also differed significantly, with the maximum value in Mirval × ungrafted × organic (0.14 mg CE·100 g⁻¹ DW) and the minimum in Mirval × S. sylvatica × chemical (0.07 mg CE·100 g⁻¹ DW). Collectively, the three-way interaction indicates that Mirval achieves the highest nutritional and functional quality when combined with ungrafted or S. sylvatica treatments and organic or chemical fertilization, whereas Black Pearl combinations—particularly with Rezistar F1—tended to produce lower-quality fruit under the tested conditions.
Figure 1.
Effect of cultivar × grafting × fertilization on eggplant quality. MF1—Mirval F1; BPF1—Black Pearl F1; NG—non-grafted; RF1—Rezistar F1; SS—Solanum sylvatica. NF—unfertilized, B—biological, O—organic, Ch—chemical. Bars represent means ± SE (n = 3). For each parameter, the letter ”a” corresponds to the highest value. Different letters indicate statistically significant differences according to Tukey’s test (p ≤ 0.05).
Figure 1.
Effect of cultivar × grafting × fertilization on eggplant quality. MF1—Mirval F1; BPF1—Black Pearl F1; NG—non-grafted; RF1—Rezistar F1; SS—Solanum sylvatica. NF—unfertilized, B—biological, O—organic, Ch—chemical. Bars represent means ± SE (n = 3). For each parameter, the letter ”a” corresponds to the highest value. Different letters indicate statistically significant differences according to Tukey’s test (p ≤ 0.05).
4. Discussion
The present study demonstrates that cultivar, grafting treatment, fertilization regime, and their interactions significantly influenced the nutritional and functional quality of eggplant fruits. These outcomes underscore the complex interplay between genotype and agronomic management in determining the accumulation of bioactive compounds and pigments under protected cultivation.
4.1. Influence of Cultivar on Bioactive and Pigment Profiles
Mirval F1 consistently exhibited higher levels of total polyphenols, DPPH radical scavenging activity, chlorophylls, lycopene, and tannins compared to Black Pearl. This aligns with previous research showing that genotype strongly determines phenolic and carotenoid profiles in eggplant and other Solanaceae, with important effects on antioxidant capacity and health-related traits [29,30,36]. Phenolic and carotenoid compounds are associated with protection against oxidative stress and confer potential health benefits linked to reduced risk of chronic disease [37,38]. For example, cultivars with higher phenolic and carotenoid contents exhibit increased antioxidant capacity in both eggplant and pepper [31].
4.2. Grafting Effects on Nutrient Allocation and Stress Response
Grafting significantly modulated fruit biochemical traits, particularly pigment content and antioxidant activity. Ungrafted plants produced fruits with higher chlorophyll and lycopene concentrations, whereas grafting onto S. sylvatica enhanced antioxidant activity. These results are consistent with studies showing that rootstock selection can influence nutrient uptake, stress responses, and secondary metabolism in grafted vegetables [32,39]. Increased pigment concentrations in ungrafted plants may indicate more effective nutrient assimilation and source–sink relationships under our conditions. Enhanced antioxidant responses in S. sylvatica grafts might reflect rootstock-induced biochemical regulation via improved water relations or phytohormone signaling [40].
4.3. Fertilization Regime and Secondary Metabolism
Fertilization regimes elicited contrasting responses in secondary metabolites and pigments. While TPC did not differ significantly across fertilization treatments, antioxidant activity, pigment content, lycopene, and tannins showed significant variation. Higher pigment and tannin levels in unfertilized controls support the theory that mild nutrient limitation can activate the phenylpropanoid pathway and increase secondary metabolite production [33,41]. Microbial-based fertilization enhanced ABTS activity, suggesting that biological inputs can positively influence plant nutrient efficiency and antioxidant pathways [43,44,45,50]. In contrast, high levels of mineral fertilization favored primary growth processes, often at the expense of secondary metabolite accumulation [34,46,48].
4.4. Interaction Effects and Optimized Management Strategies
Strong two-way and three-way interactions among cultivar, grafting, and fertilization emphasize that fruit quality outcomes are best achieved through integrative management. The combinations Mirval × ungrafted × organic and Mirval × S. sylvatica × chemical produced the most favorable profiles for antioxidants, pigments, and lycopene, indicating synergistic effects of genetic and agronomic factors. Similar interactive effects have been observed in tomato and pepper, where specific scion–rootstock and nutrient combinations improved fruit quality and stress resilience [35,47,49].
Conversely, Black Pearl under certain treatments (e.g., grafted onto Rezistar F1 with organic or chemical fertilization) tended to produce lower biochemical values, suggesting that genotype × management compatibility is critical when optimizing fruit quality under protected cultivation.
4.5. Future Research
Despite providing valuable insights, this study has limitations: (1) it was carried out in a single greenhouse environment; (2) only a subset of fertilization regimes and doses were tested; (3) yield vs. quality trade-offs were not quantitatively assessed.
Future research should address multi-season, multi-site validation, dose–response fertilization trials, systems biology analyses of metabolic regulation, and integration of quality with yield outcomes.
The results highlight that combining high-performing cultivars (e.g., Mirval), compatible rootstocks (ungrafted or S. sylvatica), and targeted fertilization strategies (particularly organic/biological) can enhance eggplant functional quality, aligning with consumer demand for nutritious foods and sustainable production systems.
5. Conclusions
Cultivar differences were observed, with the Mirval hybrid consistently outperforming Black Pearl in total polyphenol content, antioxidant activity, chlorophylls, lycopene, and tannins, confirming the dominant role of genotype in determining eggplant fruit nutritional quality. Grafting also significantly influenced fruit biochemical composition: ungrafted plants promoted higher pigment and lycopene accumulation, whereas grafting onto S. sylvatica enhanced antioxidant activity, demonstrating that rootstock selection can be tailored to target specific functional quality attributes. Fertilization regimes further modulated fruit quality. The unfertilized treatment resulted in the highest accumulation of chlorophylls, lycopene, and tannins, suggesting stimulation of secondary metabolism under moderate nutrient limitation, while biological and organic fertilization improved antioxidant capacity. In contrast, chemical fertilization tended to reduce the concentration of several bioactive compounds, with negative consequences on the functional and nutritional quality of eggplant fruits.
Strong interaction effects indicated that optimal nutritional outcomes were achieved through integrated management rather than single-factor approaches. In particular, the combinations Mirval × ungrafted × organic and Mirval × S. sylvatica × chemical were the most effective for enhancing polyphenols, pigments, lycopene, and tannins.
Overall, these findings demonstrate that combining high-performing cultivars with compatible rootstocks and sustainable fertilization strategies can substantially improve eggplant functional quality under protected cultivation. Such integrated approaches support the production of nutritionally enriched vegetables while aligning with environmentally sustainable horticultural practices.
Author Contributions
Conceptualization, V.S., S.M.A.E., J.M.M.-R.; methodology, V.S., J.L.O.-D., S.N., J.M.M.-R.; software, V.S., G.-C.T., S.N., S.M.A.E.; validation, V.S., J.L.O.-D., J.M.M.-R.; formal analysis, S.M.A.E., A.C.; investigation, V.S., G.-C.T., A.C., S.M.A.E., A.C.; resources, V.S., J.L.O.-D., J.M.M.-R.; data curation, V.S., J.M.M.-R.; writing—original draft preparation, V.S.; writing—review and editing, V.S., G.-C.T., J.M.M.-R.; visualization, V.S., S.N., G.-C.T., S.M.A.E., A.C.; bibliography, S.M.A.E., G.-C.T., A.C.; supervision, V.S., S.N., J.L.O.-D., J.M.M.-R. All authors have read and agreed to the published version of the manuscript.
Funding
The authors wish to thank “Ion Ionescu de la Brad” Iasi University of Life Sciences, Universidad de Córdoba and Department of Agricultural Industry and Food Quality, Andalusian Institute of Agricultural and Fisheries Research and Training (IFAPA), for financial support.
Data Availability Statement
Data are included in the article and supplementary materials.
Acknowledgments
The authors wish to thank “Ion Ionescu de la Brad” Iasi University of Life Sciences, Universidad de Córdoba and Department of Agricultural Industry and Food Quality, Andalusian Institute of Agricultural and Fisheries Research and Training (IFAPA), for financial support.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Caruso, G.; Pokluda, R.; Sękara, A.; Kalisz, A.; Jezdinský, A.; Kopta, T.; Grabowska, A. Agricultural practices, biology and quality of eggplant cultivated in Central Europe: A review. Hortic. Sci. 2017, 44, 201–222. [Google Scholar] [CrossRef]
- Elhawary, S.M.A.; et al. Quality attributes of eggplant varieties grown under different fertilization regimes. Sci. Pap. Ser. B Hortic. 2024, 67, 103–111. [Google Scholar]
- Plazas, M.; Nguyen, H.T.; González-Orenga, S.; Fita, A.; Vicente, O.; Prohens, J.; Boscaiu, M. Comparative analysis of the responses to water stress in eggplant (Solanum melongena) cultivars. Plant Physiol. Biochem. 2019, 143, 72–82. [Google Scholar] [CrossRef]
- Ciubotăriță, A.; Stoleru, V.; Teliban, G.C.; Cojocaru, A.; Horaicu, A. Effect of fertilization regime on eggplant crops under greenhouse conditions. Sci. Pap. Ser. B Hortic. 2022, 66, 438–443. [Google Scholar]
- Kostadinov, K.; Filipov, S.; Valcheva, V.; Kuneva, V. Influence of biological fertilization on vegetative behaviour and productivity of greenhouse eggplant. Sci. Pap. Ser. B Hortic. 2019, 63, 297–305. [Google Scholar]
- Gürbüz, N.; Uluişik, S.; Frary, A.; Frary, A.; Doğanlar, S. Health benefits and bioactive compounds of eggplant. Food Chem. 2018, 268, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Gajewski, M.; Kaczynska, K.; Bajer, M. Influence of postharvest storage on quality characteristics of eggplant cultivars. Not. Bot. Horti Agrobot. Cluj-Napoca 2009, 37, 200–205. [Google Scholar] [CrossRef]
- Meyer, R.S.; Bamshad, M.; Fuller, D.Q.; Litt, A. Comparing medicinal uses of eggplant and related Solanaceae in Asia suggests independent development of uses and recent species substitutions. Econ. Bot. 2014, 68, 137–152. [Google Scholar] [CrossRef]
- Rashmi, H.B.; Negi, P.S. Phenolic acids from vegetables: A review on processing stability and health benefits. Food Res. Int. 2020, 136, 109584. [Google Scholar] [CrossRef] [PubMed]
- Christova-Bagdassarian, V.; Atanassova, M.; Bagdassarian, V.C. Determination of tannins content by titrimetric method for comparison of different plant species. J. Univ. Chem. Technol. Metall. 2009, 44, 354–362. [Google Scholar]
- Guilherme, R.; Reboredo, F.; Guerra, M.; Ressurreição, S.; Alvarenga, N. Elemental composition and nutritional parameters of sweet pepper from organic and conventional agriculture. Plants 2020, 9, 863. [Google Scholar] [CrossRef]
- Elhawary, S.M.A.; Ordóñez-Díaz, J.L.; Nicolaie, F.; Montenegro, J.C.; Teliban, G.C.; Cojocaru, A.; Moreno-Rojas, J.M.; Stoleru, V. Quality responses of sweet pepper varieties under irrigation and fertilization regimes. Horticulturae 2025, 11, 128. [Google Scholar] [CrossRef]
- Fernández, J.A.; Ayastuy, M.E.; Belladonna, D.P.; Comezaña, M.M.; Contreras, J.; de Mourão, I.M.; Orden, L.; Rodríguez, R.A. Current trends in organic vegetable crop production: Practices and techniques. Horticulturae 2022, 8, 1103. [Google Scholar] [CrossRef]
- Fess, T.L.; Benedito, V.A. Organic versus conventional cropping sustainability: A comparative systems analysis. Sustainability 2018, 10, 272. [Google Scholar] [CrossRef]
- Ye, S.; Peng, B.; Liu, T. Effects of organic fertilizers on growth characteristics and fruit quality of pear-jujube in the Loess Plateau. Sci. Rep. 2022, 12, 13345. [Google Scholar] [CrossRef] [PubMed]
- Canatoy, R.C.; Daquiado, N.P. Fertilization influence on biomass yield and nutrient uptake of sweet corn under no-tillage. Bull. Natl. Res. Cent. 2021, 45, 66. [Google Scholar] [CrossRef]
- Stamenković, S.; Beškoski, V.; Karabegović, I.; Lazić, M.; Nikolić, N. Microbial fertilizers: A comprehensive review. Span. J. Agric. Res. 2018, 16, e10R01. [Google Scholar] [CrossRef]
- Ammar, E.E.; Aioub, A.A.A.; Elesawy, A.E.; Karkour, A.M.; Mouhamed, M.S.; Amer, A.A.; El-Shershaby, N.A. Algae as bio-fertilizers: Current status and future prospects. Saudi J. Biol. Sci. 2022, 29, 3083–3096. [Google Scholar] [CrossRef]
- Inculet, C.S.; Mihalache, G.; Sellitto, V.M.; Hlihor, R.M.; Stoleru, V. Effects of microorganisms-based products on morphological and biochemical traits of tomato under different water regimes. Microorganisms 2019, 7, 706. [Google Scholar] [CrossRef]
- Stoleru, V.; Munteanu, N.; Istrate, A. Perception towards organic vs. conventional products in Romania. Sustainability 2019, 11, 2394. [Google Scholar] [CrossRef]
- Rusu, O.R.; Mangalagiu, I.; Amăriucăi-Mantu, D.; Teliban, G.C.; Cojocaru, A.; Burducea, M.; Mihalache, G.; Roșca, M.; Caruso, G.; Sękara, A. Interaction effects of cultivars and nutrition on tomato quality and yield. Horticulturae 2023, 9, 541. [Google Scholar] [CrossRef]
- Stoleru, V.; Munteanu, N.; Hura, C.; Sellitto, V.M.; Gavrilescu, M. Organochlorine pesticide residues in soil and edible vegetables. Environ. Eng. Manag. J. 2016, 15, 527–535. [Google Scholar] [CrossRef]
- Moreno-Rojas, J.M.; Moreno-Ortega, A.; Ordóñez, J.L.; Moreno-Rojas, R.; Pérez-Aparicio, J.; Pereira-Caro, G. UHPLC-HRMS determination of flavonoids and organosulfur compounds in black onion. LWT 2018, 97, 376–383. [Google Scholar] [CrossRef]
- Ordóñez-Díaz, J.L.; Hervalejo, A.; Pereira-Caro, G.; Muñoz-Redondo, J.M.; Romero-Rodríguez, E.; Arenas-Arenas, F.J.; Moreno-Rojas, J.M. Effect of rootstock and harvest period on bioactive compounds in orange. Processes 2020, 8, 1212. [Google Scholar] [CrossRef]
- Deepa, N.; Kaur, C.; George, B.; Singh, B.; Kapoor, H.C. Antioxidant constituents in sweet pepper during maturity. LWT 2007, 40, 121–129. [Google Scholar] [CrossRef]
- Nagata, M.; Yamashita, I. Simple method for simultaneous determination of chlorophyll and carotenoids in tomato fruit. Nippon Shokuhin Kogyo Gakkaishi 1992, 39, 925–928. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y.; Colla, G.; Zrenner, R.; Schwarz, D. Vegetable grafting: Implications for fruit quality and nutritive value. Front. Plant Sci. 2017, 8, 741. [Google Scholar] [CrossRef]
- Serrano, M.; Zapata, P.J.; Castillo, S.; Guillén, F.; Martínez-Romero, D.; Valero, D. Antioxidant and nutritive constituents of sweet pepper during development. Food Chem. 2010, 118, 497–503. [Google Scholar] [CrossRef]
- Stommel, J.; Whitaker, B.D. Phenolic acid content and composition of eggplant fruit in a germplasm collection. J. Agric. Food Chem. 2003, 51, 6018–6024. [Google Scholar] [CrossRef]
- Barros, L.; Dueñas, M.; Carvalho, A.M.; Ferreira, I.C.F.R. Phenolic profiles of cultivated, in vitro cultured and commercial samples of Solanum melongena. Food Chem. 2010, 122, 79–89. [Google Scholar] [CrossRef]
- Pék, Z.; et al. Antioxidant capacity and polyphenol content in different pepper (Capsicum annuum L.) cultivars during ripening. J. Agric. Food Chem. 2009, 57, 7784–7791. [Google Scholar] [CrossRef]
- Ruiz, J.M.; et al. Influence of grafting on tomato and eggplant fruit quality: A review. Sci. Hortic. 2018, 235, 389–403. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; et al. The effect of irrigation and fertilization on phenolic content and antioxidant capacity in eggplant. Food Chem. 2012, 135, 55–66. [Google Scholar] [CrossRef]
- Yang, R.Y.; et al. Effect of nitrogen fertilization on flavonoids and antioxidant activity in tomatoes (Solanum lycopersicum). J. Sci. Food Agric. 2008, 88, 2036–2044. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Influence of grafting on product quality of fruit vegetables. Sci. Hortic. 2005, 127, 172–179. [Google Scholar] [CrossRef]
- Caruso, G.; et al. Phenolic composition and antioxidant capacity of eggplant wraps and flesh. Food Chem. 2014, 142, 317–324. [Google Scholar] [CrossRef]
- Liu, R.H. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr. 2003, 78, 517S–520S. [Google Scholar] [CrossRef]
- Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef]
- Lee, J.M.; et al. Grafting effects on vegetable quality. HortScience 2010, 45, 1710–1716. [Google Scholar] [CrossRef]
- Martínez-Ballesta, M.C.; et al. Rootstock effects on antioxidant metabolism in grafted tomatoes under salinity. Plant Physiol. Biochem. 2010, 48, 140–146. [Google Scholar] [CrossRef]
- Chinnici, F.; et al. Effect of fertilization on antioxidant content in apple fruit. J. Agric. Food Chem. 2004, 52, 1248–1252. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Gupta, A.; et al. Impact of arbuscular mycorrhizal fungi on plant nutrition and secondary metabolites. Agronomy 2020, 10, 1675. [Google Scholar] [CrossRef]
- Nacoon, S.; et al. Inoculation with beneficial bacteria improves antioxidant responses in vegetables. J. Plant Nutr. Soil Sci. 2014, 177, 389–395. [Google Scholar] [CrossRef]
- Ruzzi, M.; Aroca, R. Plant-microbe mutualistic interactions: A review. Plant Signal. Behav. 2015, 10, e1036212. [Google Scholar] [CrossRef]
- Zandonadi, D.B.; et al. Nitrogen fertilization effects on phenolic metabolism in leafy vegetables. Plant Sci. 2011, 180, 350–357. [Google Scholar] [CrossRef]
- Colla, G.; et al. Graµfting in vegetable crops: From production to quality improvement. Adv. Agron. 2010, 108, 109–147. [Google Scholar] [CrossRef]
- Kaur, C.; et al. Health benefits and safety of functional vegetables. Food Rev. Int. 2017, 33, 1–23. [Google Scholar] [CrossRef]
- Willer, H.; Lernoud, J. The World of Organic Agriculture 2023; FiBL & IFOAM — Organic International, 2023. [Google Scholar]
- Islam, S.; et al. Environmental drivers of secondary metabolite profiles in vegetables. Environ. Exp. Bot. 2019, 158, 240–252. [Google Scholar] [CrossRef]

Table 1.
Biochemical quality parameters of eggplant fruits across cultivars.
| Cultivar | TPC (mg GAE·100 g⁻¹ DW) |
ABTS (mmol TE·100 g⁻¹ DW) |
DPPH (mmol TE·100 g⁻¹ DW) |
Chl. a (mg·100 g⁻¹ DW) |
Chl. b (mg·100 g⁻¹ DW) |
Lycopene (mg·100 g⁻¹ DW) |
Tannins (mg CE·100 g⁻¹ DW) |
|---|---|---|---|---|---|---|---|
| MF1 | 2.97 ± 0.01 | 1.00 ± 0.00 | 1.23 ± 0.00 | 3.44 ± 0.07 | 4.74 ± 0.18 | 1.58 ± 0.05 | 0.10 ± 0.00 |
| BPF1 | 2.15 ± 0.04 | 0.98 ± 0.02 | 1.20 ± 0.00 | 2.19 ± 0.02 | 2.82 ± 0.02 | 0.93 ± 0.01 | 0.10 ± 0.00 |
| Sign. | * | ns | * | * | * | * | * |
MF1—Mirval F1; BPF1—Black Pearl F1. The values are presented as mean ± standard error of individual replicates. Within each column, n.s.—no statistically significant difference, *—significant difference, according to Tukey’s test at p < 0.05.
Table 2.
Fruit quality attributes of grafted and non-grafted eggplant plants.
| Graft | TPC (mg GAE·100 g⁻¹ DW) | ABTS (mmol TE·100 g⁻¹ DW) | DPPH (mmol TE·100 g⁻¹ DW) | Chl. a (mg·100 g⁻¹ DW) | Chl. b (mg·100 g⁻¹ DW) | Lycopene (mg·100 g⁻¹ DW) | Tannins (mg CE·100 g⁻¹ DW) |
|---|---|---|---|---|---|---|---|
| NG | 2.70 ± 0.04 a | 0.96 ± 0.03 b | 1.17 ± 0.01 b | 3.63 ± 0.04 a | 4.97 ± 0.08 a | 1.67 ± 0.01 a | 0.11 ± 0.00 a |
| RF1 | 2.51 ± 0.04 b | 0.98 ± 0.01 ab | 1.23 ± 0.01 a | 2.10 ± 0.02 c | 2.66 ± 0.01 c | 0.89 ± 0.01 c | 0.11 ± 0.00 a |
| SS | 2.52 ± 0.01 b | 1.03 ± 0.01 a | 1.24 ± 0.01 a | 2.86 ± 0.05 b | 3.92 ± 0.14 b | 1.29 ± 0.04 b | 0.10 ± 0.00 b |
| Sign. | * | * | * | * | * | * | * |
NG—non-grafted; RF1—Rezistar F1; SS—Solanum sylvatica. The values are presented as mean ± standard error of individual replicates. Within each column, n.s.—no statistically significant difference, *—significant difference. For each parameter, the letter ”a” corresponds to the highest value. Different letters indicate statistically significant differences according to Tukey’s test (p ≤ 0.05).
Table 3.
Biochemical parameters of eggplant fruits under different fertilization regimes.
| Fertili-zation | TPC (mg GAE·100 g⁻¹ DW) | ABTS (mmol TE·100 g⁻¹ DW) | DPPH (mmol TE·100 g⁻¹ DW) | Chl. a (mg·100 g⁻¹ DW) | Chl. b (mg·100 g⁻¹ DW) | Lycopene (mg·100 g⁻¹ DW) | Tannins (mg CE·100 g⁻¹ DW) |
|---|---|---|---|---|---|---|---|
| NF | 2.60 ± 0.02 | 1.00 ± 0.02 ab | 1.25 ± 0.02 a | 3.30 ± 0.03 a | 4.44 ± 0.08 a | 1.47 ± 0.02 a | 0.11 ± 0.00 a |
| B | 2.58 ± 0.03 | 1.02 ± 0.02 a | 1.19 ± 0.02 b | 2.46 ± 0.01 d | 3.23 ± 0.01 c | 1.03 ± 0.02 d | 0.10 ± 0.00 b |
| O | 2.54 ± 0.05 | 0.97 ± 0.01 ab | 1.20 ± 0.01 ab | 2.65 ± 0.03 c | 3.62 ± 0.09 b | 1.25 ± 0.00 c | 0.10 ± 0.00 b |
| Ch | 2.59 ± 0.01 | 0.96 ± 0.01 b | 1.22 ± 0.00 ab | 3.04 ± 0.01 b | 4.10 ± 0.11 a | 1.37 ± 0.03 b | 0.10 ± 0.00 b |
| Sign. | ns | * | * | * | * | * | * |
NF—unfertilized, B—biological, O—organic, Ch—chemical. The values are presented as mean ± standard error of individual replicates. Within each column, n.s.—no statistically significant difference, *—significant difference. For each parameter, the letter ”a” corresponds to the highest value. Different letters indicate statistically significant differences according to Tukey’s test (p ≤ 0.05).
Table 4.
Biochemical quality parameters of eggplant fruits under cultivar × grafting interaction.
| Cultivar x Graft | TPC (mg GAE· 100 g⁻¹ DW) | ABTS (mmol TE·100 g⁻¹ DW) | DPPH (mmol TE·100 g⁻¹ DW) | Chl. a (mg·100 g⁻¹ DW) | Chl. b (mg·100 g⁻¹ DW) | Lycopene (mg·100 g⁻¹ DW) | Tannins (mg CE·100 g⁻¹ DW) |
|---|---|---|---|---|---|---|---|
| MF1xNG | 2.96 ± 0.02 a | 0.93 ± 0.02 b | 1.18 ± 0.03 b | 4.70 ± 0.17 a | 6.65 ± 0.34 a | 2.24 ± 0.09 a | 0.11 ± 0.00 a |
| MF1xRF1 | 2.92 ± 0.02 a | 1.01 ± 0.03 ab | 1.30 ± 0.01 a | 2.05 ± 0.03 d | 2.42 ± 0.03 d | 0.80 ± 0.02 d | 0.11 ± 0.00 ab |
| MF1xSS | 3.03 ± 0.01 a | 1.05 ± 0.01 a | 1.23 ± 0.00 ab | 3.57 ± 0.06 b | 5.15 ± 0.22 b | 1.71 ± 0.07 b | 0.09 ± 0.00 c |
| BPF1xNG | 2.45 ± 0.06 b | 0.99 ± 0.03 ab | 1.17 ± 0.01 b | 2.56 ± 0.09 c | 3.29 ± 0.18 c | 1.10 ± 0.06 c | 0.10 ± 0.00 abc |
| BPF1xRF1 | 2.10 ± 0.07 c | 0.94 ± 0.01 b | 1.17 ± 0.01 b | 2.15 ± 0.02 cd | 2.90 ± 0.01 cd | 0.97 ± 0.01 cd | 0.10 ± 0 abc |
| BPF1xSS | 2.01 ± 0.00 c | 1.00 ± 0.02 ab | 1.25 ± 0.02 a | 2.15 ± 0.04 d | 2.69 ± 0.06 cd | 0.87 ± 0.02 cd | 0.10 ± 0.00 bc |
| Sign. | * | * | * | * | * | * | * |
MF1—Mirval F1; BPF1—Black Pearl F1; NG—non-grafted; RF1—Rezistar F1; SS—Solanum sylvatica. The values are presented as mean ± standard error of individual replicates. Within each column, n.s.—no statistically significant difference, *—significant difference. For each parameter, the letter ”a” corresponds to the highest value. Different letters indicate statistically significant differences according to Tukey’s test (p ≤ 0.05).
Table 5.
Biochemical quality parameters of eggplant fruits under cultivar × fertilization interaction.
Table 5.
Biochemical quality parameters of eggplant fruits under cultivar × fertilization interaction.
| Cultivar x Fertilization | TPC (mg GAE·100 g⁻¹ DW) | ABTS (mmol TE·100 g⁻¹ DW) | DPPH (mmol TE·100 g⁻¹ DW) | Chl. a (mg·100 g⁻¹ DW) | Chl. b (mg·100 g⁻¹ DW) | Lycopene (mg·100 g⁻¹ DW) | Tannins (mg CE·100 g⁻¹ DW) |
|---|---|---|---|---|---|---|---|
| MF1xNF | 2.76 ± 0.01 b | 0.94 ± 0.00 bc | 1.21 ± 0.00 abc | 3.21 ± 0.01 b | 4.17 ± 0.08 b | 1.39 ± 0.03 bc | 0.11 ± 0.00 bc |
| MF1xB | 3.01 ± 0.02 a | 0.97 ± 0.00 abc | 1.22 ± 0.03 abc | 3.19 ± 0.07 b | 4.18 ± 0.12 b | 1.32 ± 0.06 c | 0.10 ± 0 cd |
| MF1xO | 3.00 ± 0.04 a | 1.06 ± 0.03 a | 1.23 ± 0.03 abc | 3.14 ± 0.12 b | 4.49 ± 0.22 b | 1.57 ± 0.02 b | 0.11 ± 0.00 bc |
| MF1xCh | 3.11 ± 0.05 a | 1.02 ± 0.02 ab | 1.27 ± 0.01 ab | 4.22 ± 0.07 a | 6.11 ± 0.29 a | 2.04 ± 0.07 a | 0.11 ± 0.00 b |
| BPF1xNF | 2.44 ± 0.02 c | 1.07 ± 0.05 a | 1.28 ± 0.04 a | 3.39 ± 0.05 b | 4.71 ± 0.08 b | 1.55 ± 0.01 b | 0.12 ± 0.00 a |
| BPF1xB | 2.15 ± 0.03 d | 1.07 ± 0.04 a | 1.15 ± 0.02 c | 1.74 ± 0.05 d | 2.27 ± 0.11 c | 0.74 ± 0.02 e | 0.11 ± 0.00 bc |
| BPF1xO | 2.09 ± 0.06 d | 0.89 ± 0.00 c | 1.17 ± 0.01 bc | 2.16 ± 0.05 c | 2.76 ± 0.04 c | 0.93 ± 0.02 d | 0.10 ± 0.00 d |
| BPF1xCh | 2.07 ± 0.06 d | 0.89 ± 0.00 c | 1.18 ± 0.01 bc | 1.85 ± 0.05 cd | 2.09 ± 0.07 c | 0.69 ± 0.02 e | 0.09 ± 0.00 e |
| Sign. | * | * | * | * | * | * | * |
MF1—Mirval F1; BPF1—Black Pearl F1; NF—unfertilized, B—biological, O—organic, Ch—chemical. The values are presented as mean ± standard error of individual replicates. Within each column, n.s.—no statistically significant difference, *—significant difference. For each parameter, the letter ”a” corresponds to the highest value. Different letters indicate statistically significant differences according to Tukey’s test (p ≤ 0.05).
Table 6.
Biochemical quality parameters of eggplant fruits under grafting × fertilization interaction.
Table 6.
Biochemical quality parameters of eggplant fruits under grafting × fertilization interaction.
| Graft x Fertilization | TPC (mg GAE·100 g⁻¹ DW) | ABTS (mmol TE·100 g⁻¹ DW) | DPPH (mmol TE·100 g⁻¹ DW) | Chl. a (mg·100 g⁻¹ DW) | Chl. b (mg·100 g⁻¹ DW) | Lycopene (mg·100 g⁻¹ DW) | Tannins (mg CE·100 g⁻¹ DW) |
|---|---|---|---|---|---|---|---|
| NGxNF | 2.35 ± 0.03 e | 1.02 ± 0.03 abcd | 1.23 ± 0.03 bcde | 4.41 ± 0.02 a | 5.94 ± 0.01 a | 1.96 ± 0.00 a | 0.11 ± 0.00 bcd |
| NGxB | 2.61 ± 0.03 cde | 0.95 ± 0.05 de | 1.07 ± 0.01 fg | 2.48 ± 0.04 f | 3.37 ± 0.06 de | 1.13 ± 0.03 cd | 0.09 ± 0.00 de |
| NGxO | 2.98 ± 0.11 ab | 0.97 ± 0.00 cde | 1.26 ± 0.02 abcd | 4.15 ± 0.05 b | 6.01 ± 0.14 a | 2.08 ± 0.01 a | 0.12 ± 0.01 a |
| NGxCh | 2.88 ± 0.02 abc | 0.91 ± 0.02 e | 1.12 ± 0.01 fg | 3.48 ± 0.06 c | 4.57 ± 0.13 b | 1.50 ± 0.04 b | 0.11 ± 0 abc |
| RF1xNF | 2.34 ± 0.03 e | 0.90 ± 0.01 e | 1.14 ± 0.01 defg | 3.24 ± 0.03 d | 4.47 ± 0.14 bc | 1.49 ± 0.03 b | 0.12 ± 0.00 ab |
| RF1xB | 2.38 ± 0.10 e | 1.12 ± 0.00 a | 1.28 ± 0.05 abc | 2.04 ± 0.07 g | 2.40 ± 0.12 fg | 0.78 ± 0.04 e | 0.10 ± 0.00 bcd |
| RF1xO | 2.74 ± 0.08 bcd | 0.99 ± 0.03 bcde | 1.32 ± 0.00 abc | 1.47 ± 0.00 h | 1.80 ± 0.02 g | 0.62 ± 0.01 e | 0.10 ± 0.00 cd |
| RF1xCh | 2.57 ± 0.03 de | 0.89 ± 0.01 e | 1.19 ± 0.02 cdef | 1.65 ± 0.04 h | 1.96 ± 0.07 g | 0.65 ± 0.02 e | 0.10 ± 0.00 cd |
| SSxNF | 3.11 ± 0.06 a | 1.08 ± 0.03 ab | 1.37 ± 0.03 a | 2.26 ± 0.04 fg | 2.92 ± 0.10 ef | 0.98 ± 0.02 d | 0.11 ± 0.00 abc |
| SSxB | 2.74 ± 0.05 bcd | 0.99 ± 0.00 bcde | 1.21 ± 0.03 cde | 2.87 ± 0.07 e | 3.92 ± 0.09 cd | 1.18 ± 0.09 c | 0.11 ± 0.00 abc |
| SSxO | 1.92 ± 0.04 f | 0.96 ± 0.01 de | 1.02 ± 0.05 g | 2.33 ± 0.05 f | 3.06 ± 0.11 e | 1.05 ± 0.02 cd | 0.08 ± 0.00 e |
| SSxCh | 2.32 ± 0.02 e | 1.07 ± 0.01 abc | 1.36 ± 0.01 ab | 3.97 ± 0.02 b | 5.77 ± 0.27 a | 1.95 ± 0.06 a | 0.08 ± 0.00 e |
| Sign. | * | * | * | * | * | * | * |
NG—non-grafted, RF1—Rezistar F1; SS—Solanum sylvatica; NF—unfertilized, B—biological, O—organic, Ch—chemical. The values are presented as mean ± standard error of individual replicates. Within each column, n.s.—no statistically significant difference, *—significant difference. For each parameter, the letter ”a” corresponds to the highest value. Different letters indicate statistically significant differences according to Tukey’s test (p ≤ 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



