
Submitted:
19 January 2026
Posted:
21 January 2026
You are already at the latest version
Abstract
Potato tubers can be dormant for 1–15 weeks after harvest, and the germination of tuber eye-buds is suppressed by cold. Genetic mechanisms regulating bud dormancy at low temperatures are not completely understood. We performed RNA-seq to compare gene expression in tubers after 0.5, 3.5, and 6.5 months of cold storage. Differentially expressed genes (DEGs) in non-dormant (3.5/6.5 months) vs dormant (0.5 months) tubers were associated with transcriptional and translational activation, cell growth, metabolism, hormonal signaling, meristem development, dormancy break, and reproduction, confirming the non-dormant state of the meristem in the middle of storage. DEGs encoding transcription factors (TFs) (CBF1, FLC, SVP, HY5, GI, CO, FT, SOC1, CDF1, POTM1) were associated with the regulation of dormancy, flowering, and tuberization. TF DEGs upregulated (78) or downregulated (224) over four times in non-dormant vs dormant tubers are considered potential coordinators of the endo- to non-dormancy transition and upcoming tuber sprouting during cold storage. RT-qPCR analysis of non-dormant and germinating buds revealed the upregulation of tuberigens (StSP3D, StFTL1-1, StFTL1-2), anti-tuberigens (StSP5G-like), and TF genes positively associated with tuberization or flowering/germination (StFDL1, StFDL, StCDF1, StCO-like). Our results should further investigation of the mechanisms underlying tuber meristem dormancy release and sprouting during long-term cold storage.
Keywords:
potato
; Solanum tuberosum L.
; tuber dormancy
; long-term cold storage
; transcriptome
; gene expression
; transcription factor
1. Introduction
Potato (Solanum tuberosum L.) is the most important non-cereal food crop in the world. Potato tubers, which are formed at the ends of modified stem shoots (stolons), are rich in starch, proteins, and vitamins [1]. Commercial cultivars are propagated vegetatively through tuberous reproduction, whereas in breeding practice seeds are used to contribute to the morphophysiological variability of the species [2].
Potato flowering and tuberization occur simultaneously depending on endogenous cues and seasonal changes in temperature and photoperiod. Thus, in response to day length phloem-mobile signals are transmitted from the leaves to the shoot apical meristem and the tips of stolons, where flower and tuber formation is stimulated through similar regulatory mechanisms called florigen- and tuberigen-activating complexes (FAC and TAC, respectively) [3].
Both FAC and TAC include transcription factors (TFs). The FLOWERING LOCUS T (FT) gene is a component of FAC; its mRNA and protein (TF of the PEBP family) are transported from the leaves to the shoot meristem and trigger flowering [3]. Tuber formation in potato is induced in a similar manner through the activity of FT-like genes. The potato genome contains seven FT-like genes: florigen SELF-PRUNING 3D (StSP3D), main tuberigen StSP6A, additional tuberigens StSP3D and StFTL1, and anti-tuberigens StSP5G-A/B and StSP5G-like which suppress tuber formation [4].
Besides FT factors, FAC and TAC include bZIP-TF FLOWERING LOCUS D (FD; StFDL1 in potato), which is bound to FT through a 14-3-3 protein [3].
Tuberization is negatively regulated by BBX-TF CONSTANS (CO) and CO-like TFs StCOL1 and StCOL2, which activate the transcription of anti-tuberigen StSP5G [4,5,6]. Under inductive conditions, StCOL expression is suppressed by CYCLIC DOF FACTOR 1 (CDF1), which otherwise is degraded by the joint activity of TF GIGANTEA (GI) and blue light receptor FLAVIN-BINDING KELCH-REPEAT F-BOX 1 (FKF1) [5]. The stability of StCOL is positively regulated by phytochromes (PHYs) StPHYB and StPHYF [4]. The expression of StCDF1, StSP6A, and some StSP6A target genes is induced by BELL-TF BELLRINGER-1 LIKE 5 (StBEL5) and KNOX-TF POTH1 [4,6]. In the stolon, TAC activates the AGL8-like MADS-box TF gene, StMADS1 (POTM1), which stimulates StSP6A transcription in a feedback manner [7].
Tuber formation is negatively regulated by competitive replacement of the FT factor in TAC by FT homologue PEBP-TF TERMINAL FLOWER 1 (TFL1 or CENTRORADIALIS [StCEN]) [8]. Besides, tuberization depends upon the activity of microRNAs, which can stimulate (miR172) or suppress (miR156) the process [9].
Under inductive conditions, stolons swell and turn into tubers. Mature tubers enter a state of dormancy, which can be of three possible types: endodormancy or true dormancy regulated by endogenous signals, ecodormancy determined by environmental signals, and paradormancy or apical dominance (suppression of lateral bud outgrowth) when the bud meristem is in a non-dormant state but can grow only at favorable conditions [1,10]. The duration of endodormancy depends on the cultivar and can range from 1 to 15 weeks under growth-promoting conditions (15–20°C and humidity of about 90%). Harvested tubers are usually stored at low temperatures (2–4°C) for the maintenance of the dormant state to prevent germination until seasonal sowing as well as desiccation and disease development [11,12].
The onset and maintenance of tuber dormancy is accompanied by the accumulation and activation of phytohormones such as abscisic acid (ABA), jasmonic acid (JA), strigolactones (SLs), and salicylic acid (SA), whereas the exit from dormancy is promoted by cytokinins (CKs), gibberellins (GAs), and auxins. Ethylene (ETH) signaling has been shown to be associated with both tuber states, and brassinosteroid (BR) signaling modulate tuber sprouting [12,13,14,15].
The morphological sign of the tuber dormancy release is the germination of buds [12]. The process is accompanied by an increase in the content of amino acids and their derivatives, alkaloids, flavonoids, reducing sugars, a decrease in sucrose and starch, and significant changes in zeatin and phenylpropanoid biosynthesis, glutathione metabolism, and pentose phosphate pathway [15,16].
The regulation of potato tuber dormancy depends on multiple genes. Dormancy release is associated with the activation of genes involved in the cell cycle, metabolism of phytohormones and carbohydrates, stress response, DNA methylation, and histone acetylation [11,13]. Visible tuber germination is preceded by changes in the expression of genes encoding deoxyuridine 5'-triphosphate nucleotidohydrolase (dUTPase) [17] and ADP-ribosylation factor (ARF1) [18] and those regulating the cell cycle, carbohydrate metabolism, phytohormone synthesis and signaling, and vesicular transport [11]. RNA-seq analysis of dormant and germinating tubers has identified 7,728 differentially expressed genes (DEGs) and revealed changes in the expression of stu-miR319c, miRNA8047, and miRNA482a genes [19,20], indicating the complexity of the dormant state control in potato.
Previous studies have addressed molecular mechanisms of tuber dormancy and dormancy release focusing mainly on the cell cycle and metabolic pathways; however, the intricate regulatory network behind dormancy exit is still not understood. In our study, we performed comparative transcriptomics of mature potato tubers at the endodormancy and ecodormancy states (0.5–6.5 months at 3°C) and after induction of sprouting (20°C), focusing on potential molecular triggers of dormancy break.
2. Materials and Methods
2.1. Plant Material
In this study, we used potato (S. tuberosum L.) cultivar ‘Lady Clair’. In September 2023, one-plant tubers of a similar size (55–60 mm in diameter) were harvested, defined as fully matured tubers, and stored at 3°C in the dark for 0.5 month (putative dormant tubers) and 3.5 and 6.5 months (putative non-dormant tubers). At each of three time points, the tubers were sampled by cutting around the apical eye-bud to include periderm, cortex, vascular ring, and perimedulla; two biological replicates were used. Samples were frozen in liquid nitrogen and stored at -80°C until analysis.
Tubers stored in cold for 6.5 months were also used to study the expression of genes putatively associated with dormancy release. For this, apical eye-buds were cut from tubers at 0 time (just taken from cold) and after exposure to room temperature (20°C) for 1, 7, and 9 days; the two latter time points corresponded to sprouted (0.2–1 and 1–2 mm, respectively) eye-buds.
2.2. mRNA Library Sequencing
Total RNA was extracted from tuber samples using the RNeasy Plant Mini Kit and RNase free DNase set (QIAGEN, Hilden, Germany). RNA was quantified (Qubit® RNA Assay Kit and Qubit® 2.0 Fluorometer, Life Technologies, Carlsbad, CA, USA) and qualified using agarose gel electrophoresis and the RNA integrity number (RIN) determination (Agilent 2100 Bioanalyzer, Agilent Technologies Inc., Santa Clara, CA, USA). According to the analyses, all six RNA preparations were of good quality for RNA-seq (RIN ∼ 8.5–9.6).
A NEBNext® mRNA Library Prep Reagent Set for Illumina® (New England BioLabs Inc., Ipswich, MA, USA) was used to prepare mRNA libraries. Six libraries were obtained and sequenced by MyGene Co. (Moscow, Russia) using the Illumina HiSeq 2500 (Illumina, San Diego, CA, USA).
2.3. Mapping Reads to the Reference Genome
The S. tuberosum cv. DM 1-3 516 R44 genome (SolTub_3.0; GCF_000226075.1) was used for mapping of the obtained transcript reads. After removing adapter-containing (cutadapt v.1.17 [21]) and low quality (sickle v.1.33 [22]) sequences, paired overlapping reads were merged using FLASH v.1.2.11 (https://ccb.jhu.edu/software/FLASH/; accessed on 10 November 2024) and aligned to the annotated genes in the potato genome. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were performed with ShinyGO 0.82 ([23,24]; https://bioinformatics.sdstate.edu/go/; accessed on 18 April 2025).
2.4. Differentially Expressed Genes
Relative transcription levels of the identified genes were calculated as numbers of reads per kilobase of gene exon region per million mapped reads (RPKM). Differentially expressed genes (DEGs) were identified using the DESeq2 software; DEGs were considered if the expression change was not less than two-fold (absolute log2FoldChange [FC] ≥ 1) and padj (P-value adjusted, or false discovery rate) was < 0.05. To visualize DEGs, Venn diagrams, Heatmaps, Volcano plots, and PCR vs RNA-seq plots were used ([25]).
2.5. Gene Expression Analysis by Quantitative Real-Time PCR
Based on the RNA-seq data, a number of DEGs were selected and analyzed for expression by quantitative (q) real-time (RT) PCR. Total RNA extracted from tuber samples (section 2.2) was used for cDNA synthesis (Reverse Transcription System; Promega, Madison, WI, USA). RT-qPCR was performed with SYBR Green and ROX RT-qPCR mixture (Syntol, Moscow, Russia), 3 ng cDNA, and 10 μM gene-specific primers (Supplementary Table S1) in a CFX96 Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA, USA) at the following conditions: pre-denaturation at 95°C for 5 min and 40 cycles at 95°C for 15 s and 62°C for 50 s. The data were normalized to the expression of two S. tuberosum genes, elongation factor 1-alpha (ef1, LOC102600998) and exocyst complex component (SEC3A, LOC102599118), which are stably expressed under abiotic stress conditions and can be used in pairs to improve the efficiency of normalization of target gene expression [26]. One-way ANOVA (multiple comparisons corrected with Bonferroni test) in GraphPad Prism v.9 software (https://www.graphpad.com, San Diego, CA, USA) was used to test for significant differences (P < 0.05) between mean gene expression values in each sample (two biological and three technical replicates were used).
3. Results
3.1. Transcriptome Sequencing, Mapping, and Primary Analysis
Eye-buds of mature potato tubers (0.5 months of cold storage) were considered dormant. The time point of 3.5 months corresponded to the maximum known period for endodormancy release of the tuber bud meristem (about 15 weeks [10]) and that of 6.5 months corresponded to the approaching end of storage.
To analyze changes in gene expression between tuber dormancy and dormancy release states, we performed RNA-seq of 0.5-, 3.5-, and 6.5-month tuber transcriptomes. Reads were aligned to the potato reference genome. Sequencing yielded 19,436,490 (0.5 months), 21,677,357 (3.5 months), and 17,269,387 (6.5 months) good quality reads, representing approximately 4 GB of sequence data for each time point and approximately 4.7 times the size of the potato genome (0.84 Gb).
A total of 23,425 (0.5 months), 22,601 (3.5 months), and 22,576 (6.5 months) expressed genes were identified, which covered 24,957 of 32,873 annotated potato genes (Supplementary Table S2). The distribution of the expressed genes in the three stages is summarized in Venn diagrams (Figure 1). In total, 20,962 genes were expressed across all three stages and approximately 500–600 genes were co-expressed between stage pairs. The highest number of genes (1,333) were specifically expressed in mature dormant tubers (0.5 months) (Figure 1a).
Among the expressed genes, 26.6%, 27.6%, and 4.8% were identified as DEGs (at least a two-fold change) at 3.5 vs 0.5 months (2,393/4,242 up/downregulated), 6.5 vs 0.5 months (2,633/4,207), and 6.5 vs 3.5 months (722/470), respectively (Figure 1b, c, Supplementary Tables S3-5). Compared with 0.5-mo, 3.5- and 6.5-mo tubers had 1,721 upregulated and 3,433 downregulated common DEGS (Figure 1b, c). The changes were quantitatively similar between 3.5- vs 0.5-mo and 6.5- vs 0.5-mo samples and were notably higher than those for 6.5- vs 3.5-mo tubers, indicating that the most significant transcriptional shifts occured in tubers during the 0.5–3.5-month period.
DEGs with the most significant expression changes (Volcano plots in Figure 2) may serve as key biomarkers of the transition between tuber dormancy and dormancy release without visually detectable morphological changes. In particular, the DEGs significantly upregulated in 3.5- and 6.5- vs 0.5-mo tubers included dUTPase (LOC102589306) (Supplementary Table S3-5) considered as a marker of tuber dormancy release [17]. Another such marker is ADP-ribosylation factor (ARF1) [18], and we found that one of the ARF1-like genes (LOC102577686) was significantly downregulated in 3.5- vs 0.5-mo tubers (Supplementary Table S3-5).
3.2. Enriched GO and KEGG Terms
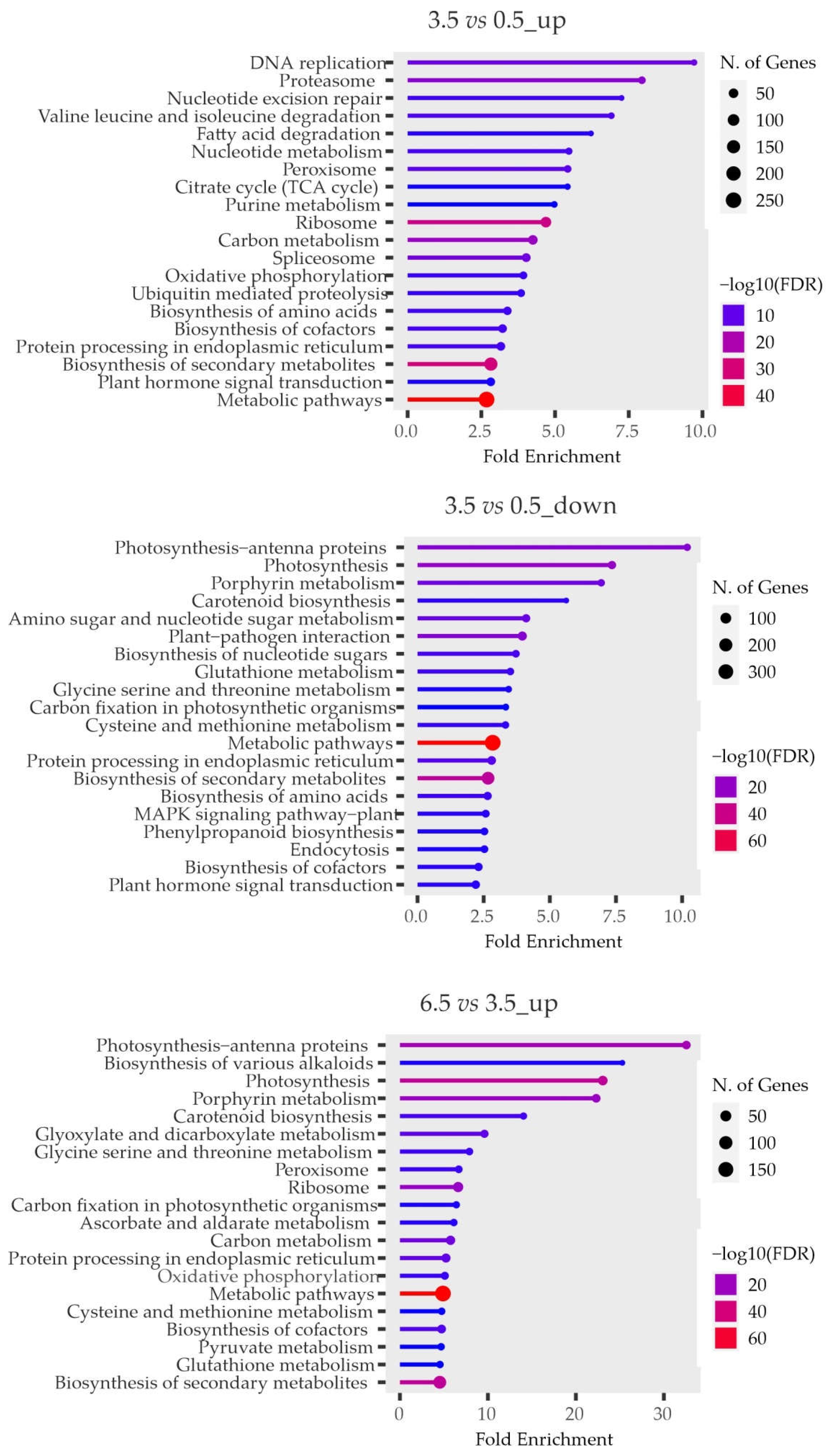
To reveal the functions of the overrepresented DEGs, we characterized them in GO and KEGG terms. It was found that in 3.5- and 6.5- vs 0.5-mo samples, the most enriched genes were associated with transcriptional and translational activation, post-translational modifications, cell cycle/differentiation, phytohormone metabolism and signaling, and other metabolic pathways (carbohydrate, lipid, etc.), as well as with the suppression of photosynthesis and biosynthesis of secondary metabolites (carotenoids, phenolic compounds, and porphyrins). In 6.5- vs 3.5-mo tubers, the most enriched genes were related to photosynthesis and secondary metabolism (Figure 3, Supplementary Tables S6-11), indicating the upcoming germination of eye-bud meristems.
As expected, storage of tubers at low temperature affected the expression of stress response- and lipid metabolism-related genes. In both 3.5- and 6.5- vs 0.5-mo tubers, the activity of ~15% genes in the categories “response to cold”(GO:0009409) and “response to temperature stimulus”( GO:0009266) was increased, whereas that of 15% was decreased. Also, 15% of the genes belonging to the categories “response to water deprivation” (GO:0009414) and “response to osmotic stress” (GO:0006970) were upregulated and 15% were downregulated (Supplementary Tables S6-9), which is consistent with the tendency of tubers to desiccate during long-term storage.
At 3.5 vs 0.5 months, many DEGs were found to be associated with main phytohormone signaling pathways. The activity of genes involved in SA-, JA-, and BR-related processes was rather suppressed (corresponding categories were enriched only in the downregulated DEGs), whereas some genes associated with ETH, CK, and ABA pathways were stimulated and the other such genes – inhibited. The auxin-mediated pathway was activated, but no DEGs related to the GA pathway were found (Supplementary Tables S6, 7). These data are somewhat inconsistent with the results of previous studies showing that dormancy release is associated with the upregulation of CK, GA, auxin, and BR signaling and downregulation of ABA, ETH, JA, SL, and SA pathways [12,13,14,15]. Thus, the status of phytohormones after 3.5 months of cold storage could indicate that the tubers differed in terms of the time of exit from endodormancy and ecodormancy.
Figure 2.
DEGs with the most significant expression changes between the 0.5-, 3.5-, and 6.5-mo tuber transcriptomes.
Figure 2.
DEGs with the most significant expression changes between the 0.5-, 3.5-, and 6.5-mo tuber transcriptomes.
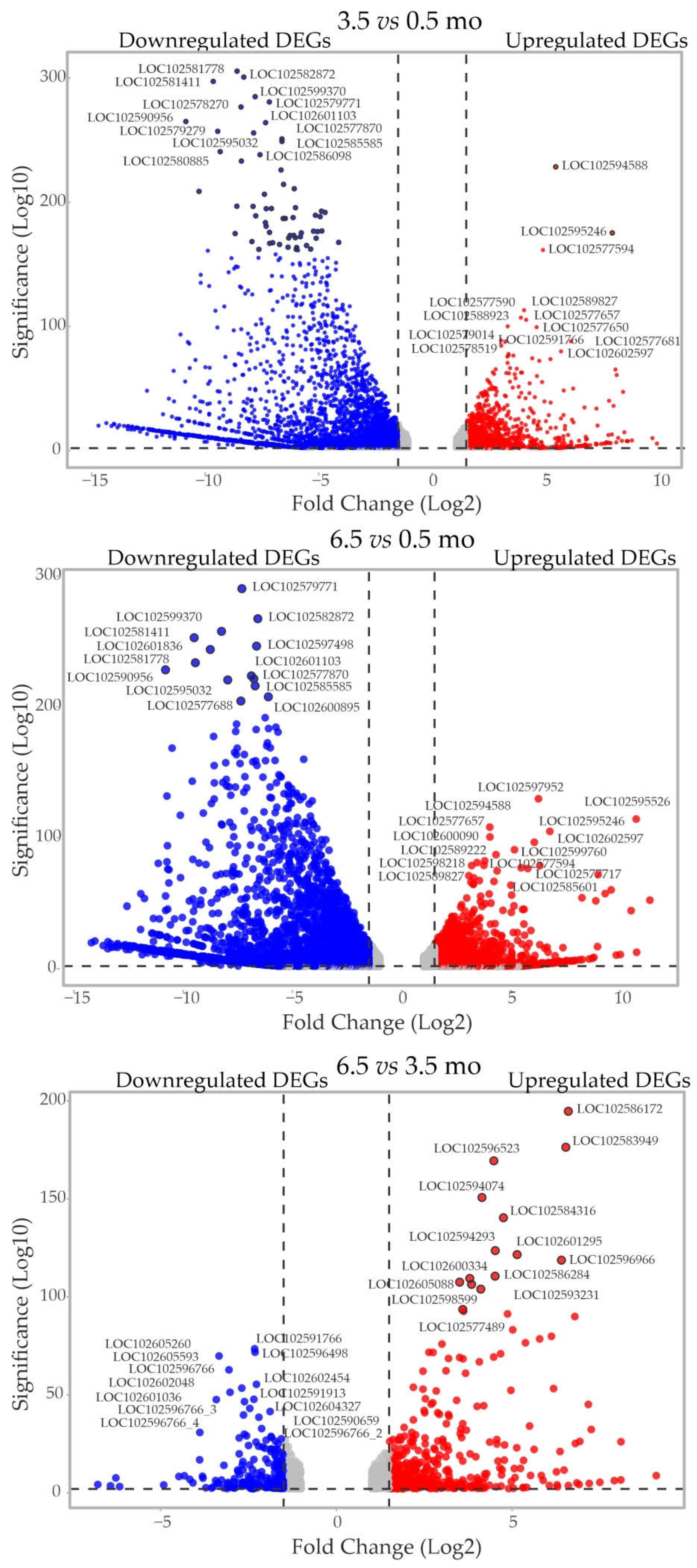
Figure 3.
The most enriched functional categories (KEGG terms) of DEGs between the 0.5-, 3.5-, and 6.5-mo transcriptomes.
Figure 3.
The most enriched functional categories (KEGG terms) of DEGs between the 0.5-, 3.5-, and 6.5-mo transcriptomes.
Both 3.5- and 6.5- vs 0.5-mo tubers were characterized by the enrichment of DEGs related to the development of tuber embryonic meristems (e.g. “embryo development” GO:0009790, “embryo development ending in seed dormancy” GO:0009793, and “meristem development” GO:0048507) (Table 1, Supplementary Tables S6–9). The enrichment in DEGs belonging to the category “postembryonic development” (GO:0009791) and “plant organ development” (GO:0099402) was even more pronounced in 6.5-mo vs 3.5-mo samples. Taken together, these results may indicate the activation of the eye-bud meristems in potato tubers at 3.5 months and their progressive development and upcoming sprouting at 6.5 months of cold storage.
Analysis of DEGs associated with plant reproductive development produced interesting results. Thus, genes belonging to the category “vegetative to reproductive phase transition of meristem” (GO:0010228) were upregulated at 3.5 months , which may indicate the involvement of flowering initiation-related genes in the determination of the non-dormant state of the bud. This result was supported by the finding that DEGs (both up- and downregulated) associated with the initiation of flowering and flower development were enriched in the corresponding categories (“flower development” GO:0009908, “regulation of photoperiodism flowering” GO:2000028, “inflorescence development” GO:0010229, and “reproductive shoot system development” GO:0090567) (Table 1, Supplementary Tables S6,7). These genes may be related to the regulation of tuber dormancy release, which causes transcriptomic/metabolomic changes previously detected by comparative analysis of dormant and non-dormant tubers ([12,13]).
3.3. Detailed Analysis of the DEGs
3.3.1. Storage Proteins
Patatins are the major potato tuber storage proteins, which are degraded during tuber sprouting due to suppression of protease inhibitors; accordingly, the amino acids necessary for the initial sprouting are released [27]. Our results indicate that DEGs of most protease inhibitors were upregulated in 3.5- vs 0.5-mo tubers (dormancy release) and then downregulated (many of them sharply) in 6.5- vs 3.5-mo samples (upcoming sprouting state). Under stimulating conditions (20°C), the expression of patatin genes decreases when tubers exit from dormancy and increases during their germination [12]. In our study, the profile of patatin genes was different, probably because of cold storage: five patatin DEGs were upregulated at 3.5 vs 0.5 months and downregulated at 6.5 vs 3.5 months, whereas seven such DEGs were downregulated at both 3.5 and 6.5 vs 0.5 months (Figure 4a, Supplementary Table S12).
Analysis of lipoxygenase genes showed that 2 and 3 of them were upregulated and 9 and 6 – downregulated in 3.5- and 6.5- vs 0.5-mo samples, respectively (Figure 4a, Supplementary Table S12). These data are consistent with the finding that lipoxygenases are involved in the formation of secondary metabolites that regulate shoot development [27].
Thus, under low-temperature conditions, when bud endodormancy breaks (presumably at 3.5 months or earlier), the expression of most protease inhibitor genes increased, whereas that of about half of the patatin genes and most lipoxygenase genes decreased, suggesting maintenance of the ecodormancy state with the suppression of eye-bud germination. These tendencies continued in 6.5-mo tubers (upcoming sprouting), in which the expression of most genes encoding patatins, protease inhibitors, and lipoxygenases was decreased.
3.3.2. Phytohormones
Potato tuber dormancy is maintained by the ABA, JA, SL, and SA pathways and is broken by the upregulation of auxin, CK, and GA signaling, whereas tuber sprouting is regulated by BRs [14]. ETH is positively associated with both tuber states but more with dormancy maintenance; in dormant tubers, ETH accumulation is increased, which supports the dormancy state through effects on carbohydrate and phenylpropanoid metabolism [14] and on the inhibition of cell division, DNA synthesis, meristem activation, and sprouting [28]. In contrast, short-term treatment with exogenous ETH breaks dormancy by stimulating the expression of the ABA 8'-hydroxylase gene StCYP707A2 associated with ABA catabolism, which leads to a decrease in ABA levels [14]. Dormancy-to-dormancy release transition in tubers is accompanied by an increase in the expression of ETH signaling-related genes [15].
Table 2 and Figure 4, Figure 5 and Figure 6 summarize the data on the expression change of some genes associated with phytohormone signaling in tubers at 3.5 vs 0.5 months; more detailed information on the relevant DEGs can be found in Supplementary Table S12.
Comparison of DEG profiles at different time points (Figure 4, Figure 5 and Figure 6) revealed that the expression of hormonal signaling-related genes had a turning point at 3.5 months of tuber cold storage. The patterns of DEGs at 3.5/6.5 vs 0.5 months may indicate a decrease in the content of storage proteins and carotenoids and a turn to the opposite mode of phytohormone signaling pathways.
3.3.3. Stress Response
To prevent diseases and sprouting, field-grown tubers are stored at low temperature, which, however, also has negative effects such as cold stress, desiccation, and cold-induced sweetening (CIS) [39].
Master regulators of cold acclimation Dehydration-Response Element-Binding Protein 1/C-Repeat Binding Factors (DREB1/CBF1-3) are induced by cold stress and upregulate COLD-REGULATED (COR) and RESPONSIVE TO DEHYDRATION (RD) genes. DREB1 expression is activated by Inducer of CBF Expression 1 (ICE1), Calmodulin-Binding Transcription Activator 3 (CAMTA3), CAMTA5, REVEILLE 4 (RVE4), and RVE8, and negatively regulated by Late Elongated Hypocotyl (LHY) and Phytochrome-Interacting Factors (PIFs). Basic Transcription Factor 3 (BTF3) and BTF3-like (BTF3L) promote DREB1 stability. BZR and Zinc Finger of Arabidopsis thaliana 12 (ZAT12) TFs also positively regulate cold-stress tolerance. Low temperature promotes the gene expression and protein stability of Elongated Hypocotyl 5 (HY5) TF, causing upregulation of its downstream targets such as anthocyanin biosynthesis enzymes [40]. Prolonged low temperature storage induces genes encoding heat shock proteins (Hsps), which may prevent tuber cold damage [41].
At 3.5 and 6.5 vs 0.5 months, the expression of four DREB1-like, RVE8, LHY, CAMTA3, CAMTA4, PIF1-like, and two BZR-like genes was downregulated and that of two COR-like, HY5, and EARLY RESPONSIVE TO DEHYDRATION 15 (ERD15) genes was upregulated. Among 23 Hsp DEGs, two (Hsp21 LOC102604571 and Hsp70 LOC102579387) were upregulated at 3.5 vs 0.5 months and then downregulated at 6.5 vs 3.5 months, whereas 14 other Hsp genes showed an upregulation trend (Supplementary Table S12, Figure 7a).
The HY5 gene and some genes encoding flavonoid/anthocyanin biosynthesis enzymes, such as CHS (chalcone synthase), CHI (chalcone isomerase), F3H (flavanone 3-hydroxylase), FLS (flavonol synthase), IFR (isoflavone reductase), DFR (dihydroflavonol-4-reductase), ANR (anthocyanidin reductase) were upregulated, whereas the other genes such as F3’5’H (flavonoid 3',5'-hydroxylase-like), DFR, UFGT2 (anthocyanidin 3-O-glucosyltransferase 2), UFGT5, and UFGT6 were downregulated. DEGs encoding TFs that target the anthocyanin pathway (ANTHOCYANINLESS 2, GLABRA 2, GLABRA 1, MYB113, MYBA1, StAN2, and StAN1) were downregulated (Supplementary Table S12, Figure 7b).
CIS is characterized by the accumulation of soluble sugars, which is mostly regulated by the combined activity of granule-bound starch synthase 1 (GBSS1), beta-amylases (BAM), invertase inhibitors (INH), and fructokinases stimulating disaccharide and polyglucan breakdown [41]. At 3.5 vs 0.5 months, BAM7, inhibitor of fructosidase 1 (LOC102586828), and fructokinase (LOC102577816) DEGs were upregulated and BAM1 and BAM3 – downregulated, whereas two GBSS-like and two soluble starch synthase-encoding genes were slightly upregulated (by 1.3–1.6-fold) (Supplementary Table S12, Figure 7c).
Some DEGs related to disaccharide hydrolysis were downregulated (sucrose-phosphate synthase SPS4, alkaline/neutral invertase A, beta-fructosidase PAIN-1, cell wall invertase CWINV1, beta-galactosidases 3 and 4, alpha-galactosidase) and some – upregulated (sucrose synthase SS16, alkaline/neutral invertase D, beta-galactosidase 16, and alpha-galactosidase 3). The following DEGs encoding enzymes involved in polyglucan breakdown (including that of starch) were downregulated (nine glucan endo-1,3-beta-D-glucosidases, beta-glucosidases 11 and 40, alpha-amylase, and alpha-1,4 glucan phosphorylase L-1 isozyme) and the other such genes were upregulated (beta-glucosidases 18, 42 and 46, phosphoglucan phosphatase DSP4, and alpha-glucan phosphorylase type H isozyme) (Supplementary Table S12, Figure 7c).
The DEGs strongly upregulated at 6.5 vs 3.5 months encoded BAM1, PAIN-1, beta-galactosidase 8, and beta-glucosidase 11 and those downregulated encoded two GBSS-like, soluble starch synthase LOC102577674, fructokinase, inhibitor of fructosidase 1, alpha-glucan phosphorylase type H isozyme, alpha-1,4 glucan phosphorylase L-1 isozyme, and sucrose synthase (Supplementary Table S12, Figure 7c).
The resistance of long-term stored tubers to desiccation depends on the formation of hydrophobic barriers at the periderm level to reduce water loss, including suberization which directly depends on ABA concentration. The enzymes of the suberin biosynthesis pathway in potato are β-ketoacyl-coenzyme A (CoA) synthase StKCS6, cytochrome CYP86A33, and feruloyl transferase (FHT) [42]. In addition, late embryogenesis abundant proteins (LEA) play an important role in the adaptation of tubers to dehydration [43]. ERD15 negatively affects ABA sensitivity [44], and aquaporin membrane channels maintain water balance [45].
At 3.5 vs 0.5 months, three LEA-like DEGs were upregulated and two – downregulated. The expression of the FHT suberization gene was upregulated and that of StKCS6, StKCS4, and StKCS11 was downregulated. ERD15 and three aquaporin genes (TIP2-1 and two TIP3-2) were upregulated, whereas eight aquaporin DEGs were downregulated. At 6.5 vs 3.5 months, five aquaporin DEGs (PIP1-1, PIP2-1, TIP1-1, and two TIP3-2 genes) were strongly upregulated (Supplementary Table S12, Figure 7d). It could be suggested that these DEGs are involved in the regulation of water balance during long-term cold storage and dormancy release of potato tubers.
It should also be noted that at 3.5 vs 0.5 months, most DEGs encoding pathogenesis-related proteins (chitinases, aspartic proteases, glucosidases, thaumatin-like proteins, peroxidases, etc. [46]), as well as most DEGs coding for WRKY-TFs important for plant defense responses [47] were downregulated, except for WRKY3, WRKY5, WRKY12, WRKY41, and WRKY65 related to cold and/or osmotic stresses (drought, salt), which were upregulated (Supplementary Table S3). Three of four detected DEGs for universal stress proteins A-like (USPAs), which are vital for plant survival under stressful conditions [48], were upregulated by 3–5-fold (Supplementary Table S3).
Overall, these results suggest that the detected changes in the expression of genes associated with adaptive responses to low temperature (including CIS) and desiccation are associated not only with stress conditions but also with the stage of tuber development – dormancy release (3.5 months) and upcoming sprouting (6.5 months).
3.3.4. Transcription Factors
In search of possible regulators initiating tuber exit from dormancy, we analyzed the DEGs encoding TFs.
At 3.5 vs 0.5 months, some TF families (BZIP, ARF, NF-YA, GRF, C3H, TALE, LSD, Dof, HB-other, HD-ZIP, TCP, and WOX) were enriched with the upregulated DEGs and the other (ERF, GRAS, WRKY, C2H2, AP2, CO-like, DBB, SBP, BES1, MYB, MYB-related, HB-other, Nin-like, TCP, BZIP, BHLH, Trihelix, and ZF-HD) with the downregulated DEGs (Supplementary Table S13).
At 6.5 vs 0.5 months, the ARF, C3H, G2-like, and HSF families contained downregulated DEGs. At 6.5 vs 3.5 mo, the CO-like and DBB TF families were enriched with the upregulated DEGs and families such as C2H2, Trihelix, BHLH, BZIP, HB-other, ARF, TALE, DBB, C3H, GRAS, HD-ZIP, and WOX – with the downregulated DEGs (Supplementary Table S13).
Heatmap analysis revealed distinct TF DEG profiles in the samples, pointing on the differences in the regulation of gene expression between the dormant (0.5 months) and dormancy release (3.5 and 6.5 months) states (Supplementary Table S2, Figure 8).
At 3.5 vs 0.5 months, the expression of 78 TF genes was activated and that of 224 was inhibited by more than 4 times. Both DEG groups were mainly related to abiotic (cold, drought, desiccation) and/or biotic stress responses, flavonoid/anthocyanin biosynthesis, phytohormone signaling, shoot/root development, and meristem/leaf patterning. More detailed information on the functional activity of the identified TF DEGs is presented in Supplementary Tables S14 and S15. These genes can be the subject of future studies on the regulation of dormancy release and sprouting suppression during long-term cold storage of potato tubers.
Next, we focused on TF DEGs associated with the processes related to dormancy, germination of seeds/buds, flowering, tuber sprouting, and meristem development.
The pool of the meristem-related upregulated (> 4-fold) DEGs included WUS homologs which maintain stem cell niche in the shoot apical meristem [49], ZPR2 which regulates shoot meristem activity [50], TSJT1 involved in shoot/stem development [51], and LET6 (POTH15) and HAT3 important for shoot meristem maintenance and leaf development [52].
Examples of the upregulated TF DEGs that may be related to dormancy/release/sprouting include MFT homologs playing a role in seed dormancy regulation [53], NAC25 and bZIP53 involved in the seed-to-seedling transition regulation [54,55], AGL21-like StMADS23 controlling seed germination and early post-germination growth [56], StGRF1 involved in potato tuber dormancy release and sprouting, and StGRF4 and StGRF9 playing a role in tuber dormancy maintenance [57].
Among the downregulated (> 4-fold) DEGs associated with dormancy/germination, there are WRKY2 homologs which mediate seed germination [58], WRKY71 which may be involved in dormancy release in lily bulbs [59], bHLH92 which reduces seed dormancy [60], and bZIP53, ERF2, ERF4, and ERF12 which regulate dormancy release and seed germination [55,61,62,63].
In addition, the genes upregulated more than 4-fold included COL2 which, together with COL1, is one of the most important factors in potato tuberization process, GI involved in tuber initiation [6], and an AGL27 homolog which belongs to the FLOWERING LOCUS C (FLC) clade of cold-sensitive floral repressors [64]. Considering the similarity of pathways regulating flowering initiation and tuberization in potato [3], we hypothesized that the eye-bud dormancy release may involve related mechanisms.
We made a list of flowering/tuberization-associated TF DEGs which demonstrated significant expression changes (by more than 2-fold or at least ≥ 1.4-fold for particularly important genes) at 3.5 and/or 6.5 vs 0.5 months (Table 3). Some DEGs in this list were associated with the FLC-mediated plant response to vernalization (FLC-clade StAGL27) and FLC expression regulation (FRI, FRL3, FRI-ESSENTIAL 1, and FLK), whereas the other DEGs were related to FLC-independent flowering/tuberization and regulation of FT expression (AGL19, NF-YB3, APL, BBX19, and COL). The expression of TAC components StBEL5 and FD did not change significantly during tuber storage. The list also included dormancy-associated DEGs (StMADS11, StMADS16, StMADS23, StSOC1, MFT, GI, LHY, HY5, AGL62, ELF3, and ELF4) (Table 3).
3.3.4. MiRNAs
Four microRNA genes were found to be differentially expressed at 3.5 vs 0.5 months: MIR172B, MIR6023, and MIR8047 were upregulated and MIR482A – downregulated (Supplementary Table S13).
MiRNA482a belongs to the miRNA482/2118 superfamily; it is specifically involved in GA signaling via phasiRNA-mediated targeting of GID1 genes and, as such, in seed development [20]. The expression of two DEGs encoding GA receptor GID1B-like proteins was upregulated by 5-fold (LOC102604924) or downregulated by 2.2-fold (LOC102579350) (Supplementary Table S13), suggesting them as possible targets of miRNA482a during tuber dormancy maintenance or release.
MiRNA172 targets AP2/ERF-TF genes. In tree peony, miRNA172 targeting of the ETH-responsive TOE3 gene was shown to suppress bud dormancy release [89]. Among potato DEGs, the expression of the TOE3-like gene (LOC102582479) was increased by 2.5-fold at 3.5 vs 0.5 months (indirectly confirming tuber dormancy release), whereas the expression of other AP2/ERF family genes mostly decreased (29 of 39 genes), suggesting their involvement in dormancy maintenance (Supplementary Table S13).
MiRNA6023 mediates potato cold resistance by targeting pectinesterases [90]. In our transcriptomes, the expression of pectinesterase DEGs was either downregulated by 1.4–37.5-fold (or even to zero) (LOC102588428, LOC102584217, LOC102583978, and LOC102605006) or upregulated by 6–7-fold (LOC102598671 and LOC102598327) at 3.5 vs 0.5 months (their expression level at 0.5 months was very low [~0.2 RPKM]) (Supplementary Table S13).
MiRNA8047 targets ARF genes and is associated with tuber dormancy release [19] which should be accompanied by the activation of auxin signaling [14]. The upregulation of this microRNA at 3.5 vs 0.5 months was accompanied by a 2.4- to 9.7-fold increase in the expression of four ARF DEGs (ARF5 LOC102590158, ARF17 LOC102588218, ARF18 LOC102586871, and ARF18-like LOC102599257) which are likely responsible for the induction of auxin signaling, and by a 1.4–1.8-fold decrease in the expression of the other four ARF genes (ARF1 LOC102604408, ARF2 LOC102582348, ARF7 LOC102594021, and ARF18-like LOC102592252) (Supplementary Table S13).
3.4. RT-qPCR Analysis of Selected DEGs
3.4.1. Validation of Transcriptome Data on Selected DEGs
To validate the RNA-seq results, the expression of eight genes in tubers was analyzed by RT-qPCR. Candidate genes were randomly selected among those putatively associated with dormancy release. As shown in Figure 9, the expression dynamics of these genes generally corresponded to the transcriptome data.
3.4.2. Expression of Genes Presumably Related to Dormancy Release in Tuber Buds
The expression of 17 selected genes (StCO, StCOL, StCOL2, StCOL16, StPHYB, StCDF1, POTM1, StSP3D, StFTL1-1, StFTL1-2, StSP5GA, StSP5GB, StGPT2, StBLH8, FDL1, and StFDL) putatively associated with dormancy release or tuberization was compared in the eye-buds of 6.5-mo tubers (non-dormant, 6.5) and 6.5-mo tubers kept at 20°C for 1 day (6.5-0), 7 days (0.2–1 mm shoots, 6.5-1), and 9 days (1–2 mm shoots, 6.5-2).
All analyzed FT-like genes, including tuberigens (StSP3D, StFTL1-1, and StFTL1-2) and anti-tuberigens (StSP5GA and StSP5GB) [4] were upregulated in 6.5-0 vs 6.5 samples. The expression of the main tuberigen StSP6A [4] was not analyzed, since the RPKM of the gene in the transcriptomes was less than 1 for all three time points and did not change significantly between them. The two other possible TAC components [3] were either upregulated (StFDL) or did not change (StFDL1). In 6.5-1 vs 6.5-0 samples, the expression of tuberigens and anti-tuberigens continued to increase along with that of StFDL1 and StFDL but then all these genes were downregulated in 6.5-2 vs 6.5-1 samples (Figure 10).
In 6.5-0 vs 6.5 samples, one of potential positive regulators of tuberigens [4,5,7] (StBLH8) was activated, whereas the other were either repressed (StCDF1 and POTM1) or did not change (StPHYB). At later points, POTM1 continued to be repressed, StBLH8 did not change, and StCDF1 and StPHYB were slightly activated and then repressed (Figure 10).
The expression of possible CO-like regulators of tuberization/flowering [4,5,6] in 6.5-0 vs 6.5 samples was increased (StCO and StCOL2) or did not change (StCOL and StCOL16). In 6.5-1 vs 6.5-0 samples, the expression of StCO continued to increase but then decreased in 6.5-2 samples, whereas that of StCOL2 decreased in 6.5-2 vs 6.5-1 samples, and that of StCOL and StCOL16 did not change (Figure 10).
4. Discussion
In potato tubers, the endodormancy state of eye-bud meristems persists from 1 to 15 weeks after harvesting, depending on the genotype and environmental conditions, and is regulated by endogenous signals [10]. Long-term storage of tubers at 2–4°C prevents bud sprouting, disease development, and weight loss but leads to sweetening [12,18,28,41]. An analogue of such storage in the wild is seasonal cold, when tuber endodormancy is maintained, among other things, through molecular pathways triggered by low temperatures [10]. Moreover, prolonged vernalization is necessary for the efficient exit of buds, seeds, or storage organs from dormancy under favorable conditions, as demonstrated for many plant species [10,67,79,80].
If the duration of cold exposure exceeds the tuber genotype-specific dormancy breaking time, endodormancy is likely to be lost during storage, but germination should nevertheless be inhibited by low temperature and darkness. Although the dormancy break is not visually detectable, it can still be monitored at the molecular level owing to significant changes in gene transcription and metabolic indicators [12,15,18,57]. Despite of that, RNA-seq analysis of cold-stored tubers has only been used to study the CIS phenomenon [41], whereas the dormancy release mechanism has been mainly investigated under dormancy exit-promoting conditions, i.e., at the temperature about 20°C [12,15,18,57] or after treatment of tubers with chemical agents [13,15].
In the present study, we attempted to analyze the transcriptomes of potato tubers during storage in the dark at low temperature for 6.5 months in order to clarify the regulatory mechanism(s) of eye-bud dormancy release, especially considering vernalization-mediated pathways. Gene expression patterns were compared at three time points: 0.5 months (endodormancy), 3.5 months (mid-storage), and 6.5 months (end of storage). The mid-point was chosen as the longest possible endodormancy release period (15 weeks [10]).
The considerable changes in gene expression, which we found at 3.5- vs 0.5 months (Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8), including a significant increase in the expression of StdUTPase considered as a marker of tuber bud exit from dormancy [17], confirmed that bud endodormancy had already been broken by the middle of the cold storage period (or even earlier), whereas ecodormancy (suppression of sprouting) had been maintained.
These findings were supported by the results of GO/KEGG analysis, which showed that in 3.5- vs 0.5-mo tubers, the most enriched functional categories of the DEGs were related to transcriptional and translational activation, cell cycle/differentiation, post-translational modifications, phytohormone signaling, and metabolic pathways. (Figure 3, Supplementary Tables S6–11). In 6.5- vs 3.5-mo samples, the DEGs in the enriched GO/KEGG categories were associated with the activation of photosynthesis (Figure 3), which may indicate the upcoming germination of the buds exposed to cold for 6.5 months. This notion is supported by previous results demonstrating the induction of photosynthetic activity in germinating buds [12]. Therefore, the 6.5-mo tubers were used to obtain eye-buds at different stages of sprouting induced at 20°C (6.5-0, 6.5-1 and 6.5-2 samples) for further expression analysis of individual regulatory genes.
The activation of eye-buds after 3.5 months of cold storage was also confirmed by the enrichment of DEGs in functional categories associated with seed dormancy termination and embryo/meristem and reproductive system development (Table 1, Supplementary Tables S6–9). Multiple changes at 3.5 vs 0.5 months in the expression of genes related to hormonal signaling, cell division and cell cycle activity, storage proteins, carbohydrate, carotenoid/flavonoid metabolism, and stress response (Figure 4, Figure 5, Figure 6 and Figure 7, Table 2, Supplementary Table S12) were consistent with previous studies on dormancy-released potato tubers [12,14,15,18,57].
Analysis of ETH-associated genes revealed transcriptional repression at 3.5 vs 0.5 months (Figure 4b, Table 2, Supplementary Table S12), indicating a decrease in ETH-mediated signaling, which is characteristic for non-dormant tuber buds [14].
Most DEGs related to the biosynthesis of carotenoids and their by-products ABA/SLs [14] were downregulated, suggesting a decrease in the ABA content. At the same time, only some genes of the ABA signaling pathways were downregulated, whereas the others were upregulated (Figure 4a, Table 2, Supplementary Table S12). Nevertheless, given that carotenoid biosynthesis is positively correlated with the ABA content [28], there could be a decrease of the ABA level in 3.5-mo tubers, suggesting broken dormancy. In this case, the decrease in the expression of genes involved in JA synthesis and signaling (Figure 5c, Table 2, Supplementary Table S12) suggests negative association between JA content and dormancy release, which is consistent with the observation that JAs inhibit potato tuber sprouting and MeJA maintains dormancy [14].
The reduction of ABA levels activates hormones that promote dormancy break, such as GA; thus, a high ABA/GA ratio corresponds to dormancy and low – to sprouting [14]. Given that transcriptional changes in the genes encoding enzymes of GA biosynthesis and its positive regulators were both positive and negative (Figure 5b, Table 2, Supplementary Table S12), it is unclear whether GA production was activated in tubers at 3.5 vs 0.5 months. However, the suggested decrease in the ABA content should in turn decrease the ABA/GA ratio and promote dormancy break.
The expression of genes involved in the metabolic and signaling pathways of CK, another dormancy-breaking hormone [14,33], changed insignificantly or was repressed (Figure 5d, Table 2, Supplementary Table S12). One of the few exceptions was the upregulation of the CK-responsive WUS gene at 3.5 vs 0.5 months (Table 2), which may correspond to the activation of meristem development in tuber buds.
Among the DEGs of auxin-mediated signaling, which is positively associated with tuber dormancy release [14], two ARF6-like genes were slightly upregulated (Figure 6a, Table 2, Supplementary Table S12). This finding is consistent with a previous study showing ARF6 activation in tuber buds during dormancy release [35].
Considering the role of BRs as key modulators of potato tuber sprouting [14], the decreased expression of BR-related DEGs at both 3.5 and 6.5 months vs 0.5 months (Figure 6b, Table 2, Supplementary Table S12) suggests suppression of BR signaling, probably until bud germination. The repression of BR pathway-related genes during tuber dormancy release has also been reported earlier [12,36].
The proximity of the 3.5-month time point to the moment of tuber transition from the dormant to non-dormant state was also confirmed by the sharply changed dynamics of the expression of regulatory TF genes (Figure 8). Given the similarity of the mechanisms controlling flowering initiation and tuber induction [3], we hypothesized that the tuber exit from dormancy could be regulated in a similar way but with the adjustment for the vernalization effect.
Previous transcriptomic studies of tuber dormancy break under break-promoting conditions have identified several TF genes possibly involved in the regulation of this process: GRF3, GRF4, GRF10 (GRF4-like), and StMADS16 [12], POTM1 (AGL8-like MADS-box gene PGSC0003DMT400010451) [13], and SVP, ASYMMETRIC LEAVES 1 (AS1), SHOOT MERISTEMLESS (STM, POTH15), and MONOPTEROS (ARF5) [36]. We found that at 3.5/6.5 vs 0.5 months, the expression of POTM1, POTH15, ARF5, GRF4, and GRF10 was upregulated and that of AS1 – downregulated, whereas that of GRF3, SVP-like StMADS11, and StMADS16 did not change (Supplementary Table S13). Our results confirm possible involvement of these TFs in the regulation of dormancy release in potato tubers.
In addition to the TF genes mentioned above, HY5 and AGL21-like StMADS23 were upregulated at 3.5 vs 0.5 months. These genes are positively associated with dormancy breaking and sprouting in several plant species through transcriptional control of the corresponding targets [56,88]. Other significant genes found included upregulated MFT and downregulated two SOC1-like genes which promote dormancy [53,82], upregulated GRF1 and GRF9 involved in dormancy break and maintenance [57], induced GI genes positively associated with potato flowering/tuber initiation [4,6] and A. thaliana seed dormancy release [77], downregulated StCO-like genes, which repress potato tuberization [4,6] and promote A. thaliana seed dormancy break [77], upregulated NAC25, downregulated bZIP53-like, and three differentially regulated miRNA genes (MIR482A, MIR172, and MIR8047) regulating bud dormancy/sprouting [19,20,54,55,89]. Considering that these genes are associated (positively or negatively) with dormancy break, their expression dynamics at 3.5 vs 0.5 months (Table 3, Supplementary Table S13) provide evidence of the tuber exit from endodormancy.
The effect of vernalization on the regulation of the tuber eye-bud state may be associated with CBF1 and DAM/SVP genes, whose expression is rapidly and sequentially activated by short-term chilling, thus promoting dormancy. After prolonged exposure to cold, DAM/SVP expression is suppressed, leading to the upregulation of CO and FT genes and, consequently, dormancy exit. This effect has been demonstrated in perennial tree buds in response to seasonal cold [10].
Another group of TF genes, which constitute the FLC clade, downregulate the transcription of FT and SOC1 genes at low temperatures and may play a role in inhibiting the germination of non-dormant meristems [64,91].
In view of the data described above, our finding that CBF1 and SVP-like genes (StMADS11 and StMADS16) were downregulated at 3.5 vs 0.5 months may indicate a non-dormant state of the eye-buds, whereas the simultaneous increase in the expression of FLC-like gene StAGL27 may be consistent with the lack of sprouting of non-dormant meristems (Table 3, Supplementary Tables S13–S15).
Among the SVP and FLC target genes, most StCO-like and two StSOC1 genes were downregulated, FT-like anti-tuberigens upregulated, and FT-like tuberigens remained unchanged at 3.5 vs 0.5 month (Table 3, Supplementary Tables S13–S15). This transcriptional profile suggests that the changes in the expression of potato CO and SOC1 genes and FT tuberigens are associated not with the exit of buds from dormancy but rather with the induction of germination in non-dormant buds. At the same time, the increase in the expression of FT-like anti-tuberigenes StSP5GA and StSP5GB may suggest their participation in the suppression of premature germination of meristems in cold conditions.
Given that in Arabidopsis, FT plays a role in temperature responses and promotes seed germination [77], it is reasonable to suggest that in potato, which has several FT homologs [4], the eye-bud germination may be finely regulated through joint activity of tuberigens (stimulators) and anti-tuberigens (repressors). This notion is supported by the RT-qPCR analysis showing that under favorable conditions (20°C), not only anti-tuberigens but also other FT-like potato genes were activated in non-dormant buds (Figure 10).
More TF genes involved in the control of flowering/tuberization timing [3,4,6] were also differentially expressed at 3.5 vs 0.5 months (Table 3, Supplementary Tables S13–S15); they can be considered to play a role in the initiation of tuber bud dormancy release as secondary regulators directed by CBF-SVP-FLC.
Other two important genes, including StBEL5 which together with POTH1 upregulates StCDF1 expression, and StFDL1 which is a component of FAC and TAC [3,4,5,6], were expressed at a relatively constant level throughout storage (0.5–6.5 months) (Table 3, Supplementary Tables S13–S15). Thus, despite the supposed importance of these genes in the control of tuber dormancy release, they may not exert a decisive effect but may coordinate eye-bud germination. For example, in 6.5-0 and 6.5-1 vs 6.5-mo samples, the expression of StCDF1 (CO suppressor [5]), and POTM1 (StCDF1 activator [7]) was decreased, whereas that of StFDL1, StFDL, StCO, StCOL2, and StCOL16, potential positive regulators of germination, was increased (Figure 10). Furthermore, the upregulated expression of StBLH8 (Figure 10), a POUND-FOOLISH homolog involved in the maintenance of the shoot apical meristem [92], may indicate progression of the eye-bud meristem development during germination.
5. Conclusions
RNA-seq of potato tubers during long-term cold storage (0.5–6.5 months) revealed that the gene expression profiles at 3.5/6.5 months differed significantly from that at 0.5 months, suggesting possible transcriptional and translational activation and stimulation of cell growth and development, metabolism, and hormonal signaling. These expression changes may reflect the release from dormancy and meristem activation and at the same time, the suppression of bud sprouting due to low temperature. Among the DEGs, a number of TF genes associated with dormancy were identified, including those regulating the vernalization pathway (StCBF1, FLC-clade StAGL27, and SVP-like StMADS11 and StMADS16), initiation of flowering and/or tuber formation (CO-like StCO and StCOLs, and FT-like StSP3D, StFTL1-1, StFTL1-2, StSP5GA, and StSP5GB), flowering, and/or dormancy (two SOC1-like genes, AGL8-like POTM1, StMFT, and STM-like POTH15). Four miRNA genes were also differentially expressed at 3.5 vs 0.5 months: miRNA172b, miRNA482a, and miRNA8047 associated with dormancy and miRNA6023 associated with cold stress response. Changes in the expression of StCO, StCOL, StCOL2, StCOL16, StPHYB, StCDF1, POTM1, StSP3D, StFTL1-1, StFTL1-2, StSP5GA, StSP5GB, StGPT2, StBLH8, FDL1, and StFDL genes in sprouting versus non-dormant tuber eye-buds suggest that these genes, jointly with those of the vernalization pathway, may play an important role in the finely tuned regulation of bud dormancy exit and sprouting during long-term cold storage. The set of differentially expressed TF genes (78 activated and 224 repressed by more than 4 times) which were identified in non-dormant vs dormant buds can be considered as regulators of the dormancy break and upcoming germination of potato tuber eye-buds during long-term cold storage and could form a basis for future research.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1: List of primers for RT-qPCR analysis of Solanum tuberosum gene expression. Table S2: List of genes expressed in 0.5, 3.5 and 6.5 mo potato tubers. Table S3: List of genes differentially expressed at 3.5 vs 0.5 mo. Table S4: List of genes differentially expressed at 6.5 vs 0.5 mo. Table S5: List of genes differentially expressed at 6.5 vs 3.5 mo. Table S6: GO categories enriched in upregulated DEGs at 3.5 vs 0.5 mo. Table S7: GO categories enriched in downregulated DEGs at 3.5 vs 0.5 mo. Table S8: GO categories enriched in upregulated DEGs at 6.5 vs 0.5 mo. Table S9: GO categories enriched in downregulated DEGs at 6.5 vs 0.5 mo. Table S10: GO categories enriched in upregulated DEGs at 6.5 vs 3.5 mo. Table S11: GO categories enriched in downregulated DEGs at 6.5 vs 3.5 mo. Table S12: Selected DEGs related to the pathways of storage proteins, phytohormones, cold response, flavonoids, carbohydrates and desiccation response. Table S13: List of DEGs of transcription factors. Table S14: DEGs of transcription factors upregulated more than 4 times. Table S15: DEGs of transcription factors downregulated more than 4 times.
Author Contributions
Investigation, A.V.B., E.Z.K. and A.V.S.; formal analysis: E.Z.K., and A.V.S.; writing, A.V.S., and E.Z.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science and Higher Education of the Russian Federation.
Data Availability Statement
All data mentioned in the text are provided in the tables, figures and supplementary materials.
Acknowledgments
We would like to thank Dr. Marina Chuenkova for English language editing.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Zierer, W.; Ruscher, D.; Sonnewald, U.; Sonnewald, S. Tuber and tuberous root development. Annu. Rev. Plant Biol. 2021, 72, 551–580. [CrossRef]
- Huang, B.; Spooner, D.M.; Liang, Q. Genome diversity of the potato. Proc. Natl. Acad. Sci. U. S. A. 2018, 115(28), E6392-E6393. [CrossRef]
- Colleoni, P.E.; van Es, S.W.; Winkelmolen, T.; Immink, R.G.H.; van Esse, G.W. Flowering time genes branching out. J. Exp. Bot. 2024, 75(14), 4195-4209. [CrossRef]
- Abelenda, J.A.; Cruz-Oro, E.; Franco-Zorrilla, J.M.; Prat, S. Potato StCONSTANS-like 1 suppresses storage organ formation by directly activating the FT-like StSP5G repressor. Curr. Biol. 2016, 26, 872–881. [CrossRef]
- Sawa, M.; Nusinow, D.A.; Kay, S.A.; Imaizumi, T. FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis. Science. 2007, 318, 261–265.
- Kondhare, K.R.; Natarajan, B.; Banerjee, A.K. Molecular signals that govern tuber development in potato. Int. J. Dev. Biol. 2020, 64(1-2-3), 133-140. [CrossRef]
- Wang, E.; Zhou, T.; Jing, S.; Dong, L.; Sun, X.; Fan, Y.; Shen, Y.; Liu, T.; Song, B. Leaves and stolons transcriptomic analysis provide insight into the role of phytochrome F in potato flowering and tuberization. Plant J. 2023, 113(2), 402-415. [CrossRef]
- Zhang, X.; Campbell, R.; Ducreux, L.J.M.; Morris, J.; Hedley, P.E.; Mellado-Ortega, E.; Roberts, A.G.; Stephens, J.; Bryan, G.J.; Torrance, L.; Chapman, S.N.; Prat, S.; Taylor, M.A. TERMINAL FLOWER-1/CENTRORADIALIS inhibits tuberisation via protein interaction with the tuberigen activation complex. Plant J. 2020, 103, 2263–2278. [CrossRef]
- Bhogale, S.; Mahajan, A.S.; Natarajan, B.; Rajabhoj, M.; Thulasiram, H.V.; Banerjee, A.K. MicroRNA156: a potential graft-transmissible microRNA that modulates plant architecture and tuberization in Solanum tuberosum ssp. andigena. Plant Physiol. 2014, 164, 1011–1027.
- Roitman, M.; Eshel, D. Similar chilling response of dormant buds in potato tuber and woody perennials. J. Exp. Bot. 2024, 75(19), 6076-6092. [CrossRef]
- Dogramaci, M.; Dobry, E.P.; Fortini, E.A.; Sarkar, D.; Eshel, D.; Campbell, M.A. Physiological and molecular mechanisms associated with potato tuber dormancy. J. Exp. Bot. 2024, 75(19), 6093-6109. [CrossRef]
- Liu, B.; Zhang, N.; Wen, Y.; Jin, X.; Yang, J.; Si, H.; Wang, D. Transcriptomic changes during tuber dormancy release process revealed by RNA sequencing in potato. J. Biotechnol. 2015, 198, 17-30. [CrossRef]
- Campbell, M.; Suttle, J.; Douches, D.S.; Buell, C.R. Treatment of potato tubers with the synthetic cytokinin 1-(α-ethylbenzyl)-3-nitroguanidine results in rapid termination of endodormancy and induction of transcripts associated with cell proliferation and growth. Funct. Integr. Genomics. 2014, 14(4), 789-799.
- Di, X.; Wang, Q.; Zhang, F.; Feng, H.; Wang, X.; Cai, C. Advances in the modulation of potato tuber dormancy and sprouting. Int. J. Mol. Sci. 2024, 25(10), 5078. [CrossRef]
- Zheng, X.; Li, M.; Zhang, X.; Chen, J.; Ge, X.; Li, S.; Tian, J.; Tian, S. Unraveling the mechanism of potato (Solanum tuberosum L.) tuber sprouting using transcriptome and metabolome analyses. Front. Plant Sci. 2024, 14, 1300067.
- Liu, H.; Wang, H.; Feng, Y.; Yang, Y.; Feng, C.; Li, J.; Zaman, Q.U.; Kong, Y.; Fahad, S.; Deng, G. Integrated physiological, transcriptomic and metabolomic analyses reveal potential mechanisms of potato tuber dormancy release. Physiol. Plant. 2025, 177(1), e70081.
- Senning, M.; Sonnewald, U.; Sonnewald, S. Deoxyuridine triphosphatase expression defines the transition from dormant to sprouting potato tuber buds. Molecular breeding. 2010, 26, 525-531. [CrossRef]
- Liu, B.; Zhang, N.; Wen, Y.; Si, H.; Wang, D. Identification of differentially expressed genes in potato associated with tuber dormancy release. Mol. Biol. Rep. 2012, 39(12), 11277-11287.
- Liu, S.; Yang, J.; Zhang, N.; Si, H. Genome-wide analysis of non-coding RNA reveals the role of a novel miR319c for tuber dormancy release process in potato. Hortic. Res. 2024, 12(2), uhae303.
- Zhang, Y.; Zeng, Z.; Hu, H.; Zhao, M.; Chen, C.; Ma, X.; Li, G.; Li, J.; Liu, Y.; Hao, Y.; Xu, J.; Xia, R. MicroRNA482/2118 is lineage-specifically involved in gibberellin signalling via the regulation of GID1 expression by targeting noncoding PHAS genes and subsequently instigated phasiRNAs. Plant Biotechnol. J. 2024, 22(4), 819-832.
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17(1), 10-12.
- Joshi, N.A.; Fass, J.N. Sickle: A sliding-window, adaptive, quality-based trimming tool for FastQ files (Version 1.33) [Software]. 2011, Available at https://github.com/najoshi/sickle.
- Ge, S.X.; Jung, D.; Yao, R.; ShinyGO: a graphical gene-set enrichment tool for animals and plants. Bioinformatics. 2020, 36(8), 2628-2629.
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49(D1), D545-D551. [CrossRef]
- Tang, D.; Chen, M.; Huang, X.; Zhang, G.; Zeng, L.; Zhang, G.; Wu, S.; Wang, Y. SRplot: A free online platform for data visualization and graphing. PLoS One. 2023, 18(11), e0294236.
- Slugina, M.A.; Meleshin, A.A.; Kochieva, E.Z.; Shchennikova, A.V. The opposite effect of low temperature on the Pho1a starch phosphorylase gene expression in Solanum tuberosum L. tubers and Petota species leaves. American Journal of Potato Research. 2020, 97(1), 78-87.
- Jørgensen, M.; Stensballe, A.; Welinder, K.G. Extensive post-translational processing of potato tuber storage proteins and vacuolar targeting. FEBS J. 2011, 278(21), 4070-4087.
- Sonnewald, S.; Sonnewald, U. Regulation of potato tuber sprouting. Planta. 2014, 239(1), 27-38. [CrossRef]
- Nakashima, K.; Yamaguchi-Shinozaki, K. ABA signaling in stress-response and seed development. Plant Cell Rep. 2013, 32(7), 959-970.
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic acid signaling pathway in plants. Int. J. Mol. Sci. 2019, 20(10), 2479.
- Wang, X.; Wei, H.; Zhang, N.; Li, S.; Si, H. StNF-YA8-YB20-YC5 module regulates potato tuber dormancy by modulating gibberellin and abscisic acid pathways. Plant J. 2025, 121(6), e70106.
- Hartweck, L.M. Gibberellin signaling. Planta. 2008, 229(1), 1-13. [CrossRef]
- Argueso, C.T.; Raines, T.; Kieber, J.J. Cytokinin signaling and transcriptional networks. Curr. Opin. Plant Biol. 2010, 13(5), 533-539.
- Gomes, G.L.B.; Scortecci, K.C. Auxin and its role in plant development: structure, signalling, regulation and response mechanisms. Plant Biol. (Stuttg). 2021, 23(6), 894-904. [CrossRef]
- Faivre-Rampant, O.; Cardle, L.; Marshall, D.; Viola, R.; Taylor, M.A. Changes in gene expression during meristem activation processes in Solanum tuberosum with a focus on the regulation of an auxin response factor gene. J. Exp. Bot. 2004, 55(397), 613-622.
- Liu, T.; Wu, Q.; Zhou, S.; Xia, J.; Yin, W.; Deng, L.; Song, B.; He, T. Molecular insights into the accelerated sprouting of and apical dominance release in potato tubers subjected to post-harvest heat stress. Int. J. Mol. Sci. 2024, 25(3), 1699.
- Tian, X.; He, M.; Mei, E.; Zhang, B.; Tang, J.; Xu, M.; Liu, J.; Li, X.; Wang, Z.; Tang, W.; Guan, Q.; Bu, Q. WRKY53 integrates classic brassinosteroid signaling and the mitogen-activated protein kinase pathway to regulate rice architecture and seed size. Plant Cell. 2021, 33(8), 2753-2775. [CrossRef]
- Coll-Garcia, D.; Mazuch, J.; Altmann, T.; Müssig, C. EXORDIUM regulates brassinosteroid-responsive genes. FEBS Lett. 2004, 563(1-3), 82-86. [CrossRef]
- Alamar, M.C.; Tosetti, R.; Landahl, S.; Bermejo, A.; Terry, L.A. Assuring potato tuber quality during storage: A future perspective. Front. Plant Sci. 2017, 8, 2034.
- Kidokoro, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulatory network of plant cold-stress responses. Trends Plant Sci. 2022, 27(9), 922-935. [CrossRef]
- Lin, Q.; Xie, Y.; Guan, W.; Duan, Y.; Wang, Z.; Sun, C. Combined transcriptomic and proteomic analysis of cold stress induced sugar accumulation and heat shock proteins expression during postharvest potato tuber storage. Food Chem. 2019, 297, 124991.
- Keren-Keiserman, A.; Baghel, R.S.; Fogelman, E.; Faingold, I.; Zig, U.; Yermiyahu, U.; Ginzberg, I. Effects of polyhalite fertilization on skin quality of potato tuber. Front. Plant Sci. 2019, 10, 1379. [CrossRef]
- Wang, G.; Su, H.; Abou-Elwafa, S.F.; Zhang, P.; Cao, L.; Fu, J.; Xie, X.; Ku, L.; Wen, P.; Wang, T.; Wei, L. Functional analysis of a late embryogenesis abundant protein ZmNHL1 in maize under drought stress. J. Plant Physiol. 2023, 280, 153883.
- Alves, M.S.; Fontes, E.P.; Fietto, L.G. EARLY RESPONSIVE to DEHYDRATION 15, a new transcription factor that integrates stress signaling pathways. Plant Signal Behav. 2011, 6(12), 1993-1996.
- Wang, L.; Liu, Y.; Feng, S.; Yang, J.; Li, D.; Zhang, J. Roles of plasmalemma aquaporin gene StPIP1 in enhancing drought tolerance in potato. Front. Plant Sci. 2017, 8, 616.
- Zribi, I.; Ghorbel, M.; Brini, F. Pathogenesis Related proteins (PRs): From cellular mechanisms to plant defense. Curr. Protein Pept. Sci. 2021, 22(5), 396-412. [CrossRef]
- Javed, T.; Gao, S.J. WRKY transcription factors in plant defense. Trends Genet. 2023, 39(10), 787-801.
- Luo, D.; Wu, Z.; Bai, Q.; Zhang, Y.; Huang, M.; Huang, Y.; Li, X. Universal stress proteins: From gene to function. Int. J. Mol. Sci. 2023, 24(5), 4725.
- Cruz, J.O.; San Martin, J.A.B.; Lubini, G.; Strini, E.J.; Sobral, R.; Pinoti, V.F.; Ferreira, P.B.; Thomé, V.; Quiapim, A.C.; Dornelas, M.C.; Pranchevicius, M.C.S.; Madueño, F.; Costa, M.M.R.; Goldman, M.H.S. SCI1 is a direct target of AGAMOUS and WUSCHEL and is specifically expressed in the floral meristematic cells. Front. Plant Sci. 2021, 12, 642879.
- Weits, D.A.; Kunkowska, A.B.; Kamps, N.C.W.; Portz, K.M.S.; Packbier, N.K.; Nemec Venza, Z.; Gaillochet, C.; Lohmann, J.U.; Pedersen, O.; van Dongen, J.T.; Licausi, F. An apical hypoxic niche sets the pace of shoot meristem activity. Nature. 2019, 569(7758), 714-717.
- Sah, S.K.; Jumaa, S.; Li, J.; Reddy, K.R. Proteomic analysis response of rice (Oryza sativa) leaves to ultraviolet-B radiation stress. Front. Plant Sci. 2022, 13, 871331. [CrossRef]
- Turchi, L.; Carabelli, M.; Ruzza, V.; Possenti, M.; Sassi, M.; Peñalosa, A.; Sessa, G.; Salvi, S.; Forte, V.; Morelli, G.; Ruberti, I. Arabidopsis HD-Zip II transcription factors control apical embryo development and meristem function. Development. 2013, 140(10), 2118-2129.
- Dave, A.; Vaistij, F.E.; Gilday, A.D.; Penfield, S.D.; Graham, I.A. Regulation of Arabidopsis thaliana seed dormancy and germination by 12-oxo-phytodienoic acid. J. Exp. Bot. 2016, 67(8), 2277-2284.
- Sánchez-Montesino, R.; Bouza-Morcillo, L.; Marquez, J.; Ghita, M.; Duran-Nebreda, S.; Gómez, L.; Holdsworth, M.J.; Bassel, G.; Oñate-Sánchez, L. A regulatory module controlling GA-mediated endosperm cell expansion is critical for seed germination in Arabidopsis. Mol. Plant. 2019, 12(1), 71-85.
- Restovic, F.; Espinoza-Corral, R.; Gómez, I.; Vicente-Carbajosa, J.; Jordana, X. An active mitochondrial complex II present in mature seeds contains an embryo-specific iron-sulfur subunit regulated by ABA and bZIP53 and is involved in germination and seedling establishment. Front. Plant Sci. 2017, 8, 277.
- Yu, L.H.; Wu, J.; Zhang, Z.S.; Miao, Z.Q.; Zhao, P.X.; Wang, Z.; Xiang, C.B. Arabidopsis MADS-Box transcription factor AGL21 acts as environmental surveillance of seed germination by regulating ABI5 expression. Mol. Plant. 2017, 10(6), 834-845.
- Cui, D.; Song, Y.; Jiang, W.; Ye, H.; Wang, S.; Yuan, L.; Liu, B. Genome-wide characterization of the GRF transcription factors in potato (Solanum tuberosum L.) and expression analysis of StGRF genes during potato tuber dormancy and sprouting. Front. Plant Sci. 2024, 15, 1417204.
- Jiang, W.; Yu, D. Arabidopsis WRKY2 transcription factor mediates seed germination and postgermination arrest of development by abscisic acid. BMC Plant Biol. 2009, 9, 96. [CrossRef]
- Fan, X.; Yang, Y.; Li, M.; Fu, L.; Zang, Y.; Wang, C.; Hao, T.; Sun, H. Transcriptomics and targeted metabolomics reveal the regulatory network of Lilium davidii var. unicolor during bulb dormancy release. Planta. 2021, 254(3), 59.
- Zhao, P.; Li, X.; Jia, J.; Yuan, G.; Chen, S.; Qi, D.; Cheng, L.; Liu, G. bHLH92 from sheepgrass acts as a negative regulator of anthocyanin/proanthocyandin accumulation and influences seed dormancy. J. Exp. Bot. 2019, 70(1), 269-284.
- Pirrello, J.; Jaimes-Miranda, F.; Sanchez-Ballesta, M.T.; Tournier, B.; Khalil-Ahmad, Q.; Regad, F.; Latché, A.; Pech, J.C.; Bouzayen, M. Sl-ERF2, a tomato ethylene response factor involved in ethylene response and seed germination. Plant Cell Physiol. 2006, 47(9), 1195-1205. [CrossRef]
- Chen, X.; Li, Q.; Ding, L.; Zhang, S.; Shan, S.; Xiong, X.; Jiang, W.; Zhao, B.; Zhang, L.; Luo, Y.; Lian, Y.; Kong, X.; Ding, X.; Zhang, J.; Li, C.; Soppe, W.J.J.; Xiangm Y. The MKK3-MPK7 cascade phosphorylates ERF4 and promotes its rapid degradation to release seed dormancy in Arabidopsis. Mol. Plant. 2023, 16(11), 1743-1758.
- Li, X.; Chen, T.; Li, Y.; Wang, Z.; Cao, H.; Chen, F.; Li, Y.; Soppe, W.J.J.; Li, W.; Liu, Y. ETR1/RDO3 regulates seed dormancy by relieving the inhibitory effect of the ERF12-TPL complex on DELAY OF GERMINATION1 expression. Plant Cell. 2019, 31(4), 832-847. [CrossRef]
- Nasim, Z.; Susila, H.; Jin, S.; Youn, G.; Ahn, J.H. Polymerase II-associated factor 1 complex-regulated FLOWERING LOCUS C-clade genes repress flowering in response to chilling. Front. Plant Sci. 2022, 13, 817356.
- Lim, M.H.; Kim, J.; Kim, Y.S.; Chung, K.S.; Seo, Y.H.; Lee, I.; Kim, J.; Hong, C.B.; Kim, H.J.; Park, C.M. A new Arabidopsis gene, FLK, encodes an RNA binding protein with K homology motifs and regulates flowering time via FLOWERING LOCUS C. Plant Cell. 2004, 16(3), 731-740.
- Adkar-Purushothama, C.R.; Sano, T.; Perreault, J.P. Viroid-derived small RNA induces early flowering in tomato plants by RNA silencing. Mol. Plant Pathol. 2018, 19(11), 2446-2458.
- Schmitz, R.J.; Hong, L.; Michaels, S.; Amasino, R.M. FRIGIDA-ESSENTIAL 1 interacts genetically with FRIGIDA and FRIGIDA-LIKE 1 to promote the winter-annual habit of Arabidopsis thaliana. Development. 2005, 132(24), 5471-5478.
- Sung, S.; Schmitz, R.J.; Amasino, R.M. A PHD finger protein involved in both the vernalization and photoperiod pathways in Arabidopsis. Genes Dev. 2006, 20(23), 3244-3248. [CrossRef]
- Schönrock, N.; Bouveret, R.; Leroy, O.; Borghi, L.; Köhler, C.; Gruissem, W.; Hennig, L. Polycomb-group proteins repress the floral activator AGL19 in the FLC-independent vernalization pathway. Genes Dev. 2006, 20(12), 1667-1678.
- Schiessl, S.; Williams, N.; Specht, P.; Staiger, D.; Johansson, M. Different copies of SENSITIVITY TO RED LIGHT REDUCED 1 show strong subfunctionalization in Brassica napus. BMC Plant Biol. 2019, 19(1), 372. [CrossRef]
- Wang, X.; Xu, X.; Mo, X.; Zhong, L.; Zhang, J.; Mo, B.; Kuai, B. Overexpression of TCP8 delays Arabidopsis flowering through a FLOWERING LOCUS C-dependent pathway. BMC Plant Biol. 2019, 19(1), 534.
- Siriwardana, C.L.; Risinger, J.R.; Carpenter, E.M.; Holt, B.F. 3rd. Analysis of gene duplication within the Arabidopsis NUCLEAR FACTOR Y, subunit B (NF-YB) protein family reveals domains under both purifying and diversifying selection. PLoS One. 2023, 18(8), e0289332.
- Abe, M.; Kaya, H.; Watanabe-Taneda, A.; Shibuta, M.; Yamaguchi, A.; Sakamoto, T.; Kurata, T.; Ausín, I.; Araki, T.; Alonso-Blanco, C. FE, a phloem-specific Myb-related protein, promotes flowering through transcriptional activation of FLOWERING LOCUS T and FLOWERING LOCUS T INTERACTING PROTEIN 1. Plant J. 2015, 83(6), 1059-1068. [CrossRef]
- Paffendorf, B.A.M.; Qassrawi, R.; Meys, A.M.; Trimborn, L.; Schrader, A. TRANSPARENT TESTA GLABRA 1 participates in flowering time regulation in Arabidopsis thaliana. PeerJ. 2020, 8, e8303.
- Bai, M.; Sun, J.; Liu, J.; Ren, H.; Wang, K.; Wang, Y.; Wang, C.; Dehesh, K. The B-box protein BBX19 suppresses seed germination via induction of ABI5. Plant J. 2019, 99(6), 1192-1202.
- Wang, C.Q.; Guthrie, C.; Sarmast, M.K.; Dehesh, K. BBX19 interacts with CONSTANS to repress FLOWERING LOCUS T transcription, defining a flowering time checkpoint in Arabidopsis. Plant Cell. 2014, 26(9), 3589-3602. [CrossRef]
- Chen, M.; MacGregor, D.R.; Dave, A.; Florance, H.; Moore, K.; Paszkiewicz, K.; Smirnoff, N.; Graham, I.A.; Penfield, S. Maternal temperature history activates Flowering Locus T in fruits to control progeny dormancy according to time of year. Proc. Natl. Acad. Sci. U. S. A. 2014, 111(52), 18787-18792.
- Matilla, A.J. Auxin: Hormonal signal required for seed development and dormancy. Plants (Basel). 2020, 9(6), 705. [CrossRef]
- Canton, M.; Forestan, C.; Bonghi, C.; Varotto, S. Meta-analysis of RNA-Seq studies reveals genes with dominant functions during flower bud endo- to eco-dormancy transition in Prunus species. Sci. Rep. 2021, 11(1), 13173.
- Sato, H.; Yamane, H. Histone modifications affecting plant dormancy and dormancy release: common regulatory effects on hormone metabolism. J. Exp. Bot. 2024, 75(19), 6142-6158.
- Hu, J.Y.; Zhou, Y.; He, F.; Dong, X.; Liu, L.Y.; Coupland, G.; Turck, F.; de Meaux, J. miR824-regulated AGAMOUS-LIKE16 contributes to flowering time repression in Arabidopsis. Plant Cell. 2014, 26(5), 2024-2037.
- Huang, Q.; Chen, X.; Zhong, S.; Wu, S.; Guo, J.; Wang, Q.; Li, J.; Li, D.; Xia, Y.; Zhang, J.; Wang, X. MIKC-type MADS-box gene analysis reveals the role of PlSOC1 in bud dormancy transition in herbaceous peony. Plants (Basel). 2025, 14(6), 928.
- Zhao, H.; Xu, D.; Tian, T.; Kong, F.; Lin, K.; Gan, S.; Zhang, H.; Li, G. Molecular and functional dissection of EARLY-FLOWERING 3 (ELF3) and ELF4 in Arabidopsis. Plant Sci. 2021, 303, 110786. [CrossRef]
- Zha, P.; Liu, S.; Li, Y.; Ma, T.; Yang, L.; Jing, Y.; Lin, R. The evening complex and the chromatin-remodeling factor PICKLE coordinately control seed dormancy by directly repressing DOG1 in Arabidopsis. Plant Commun. 2019, 1(2), 100011.
- Steinbach, Y. The Arabidopsis thaliana CONSTANS-LIKE 4 (COL4) – A modulator of flowering time. Front. Plant Sci. 2019, 10, 651.
- Hassidim, M.; Harir, Y.; Yakir, E.; Kron, I.; Green, R.M. Over-expression of CONSTANS-LIKE 5 can induce flowering in short-day grown Arabidopsis. Planta. 2009, 230(3), 481-491.
- Liu, B.; Long, H.; Yan, J.; Ye, L.; Zhang, Q.; Chen, H.; Gao, S.; Wang, Y.; Wang, X.; Sun, S. A HY5-COL3-COL13 regulatory chain for controlling hypocotyl elongation in Arabidopsis. Plant Cell Environ. 2021, 44(1), 130-142. [CrossRef]
- Flynn, N. During long days, HY5a keeps dormancy away. Plant Cell. 2024, 36(5), 1596-1597.
- Zhang, Y.; Gao, L.; Wang, Y.; Niu, D.; Yuan, Y.; Liu, C.; Zhan, X.; Gai, S. Dual functions of PsmiR172b-PsTOE3 module in dormancy release and flowering in tree peony (Paeonia suffruticosa). Hortic. Res. 2023, 10(4), uhad033.
- Li, H.; Dong, Y.; Chang, J.; He, J.; Chen, H.; Liu, Q.; Wei, C.; Ma, J.; Zhang, Y.; Yang, J.; Zhang, X. High-Throughput microRNA and mRNA sequencing reveals that micrornas may be involved in melatonin-mediated cold tolerance in Citrullus lanatus L. Front. Plant Sci. 2016, 7, 1231. [CrossRef]
- Nishiyama, S.; Matsushita, M.C.; Yamane, H.; Honda, C.; Okada, K.; Tamada, Y.; Moriya, S.; Tao, R. Functional and expressional analyses of apple FLC-like in relation to dormancy progress and flower bud development. Tree Physiol. 2021, 41(4), 562-570.
- Ung, N.; Lal, S.; Smith, H.M. The role of PENNYWISE and POUND-FOOLISH in the maintenance of the shoot apical meristem in Arabidopsis. Plant Physiol. 2011, 156(2), 605-614. [CrossRef]
Figure 1.
Venn diagrams of the expressed genes (a) and DEGs upregulated (b) and downregulated (c) between the 0.5-, 3.5-. and 6.5- month transcriptomes.
Figure 1.
Venn diagrams of the expressed genes (a) and DEGs upregulated (b) and downregulated (c) between the 0.5-, 3.5-. and 6.5- month transcriptomes.
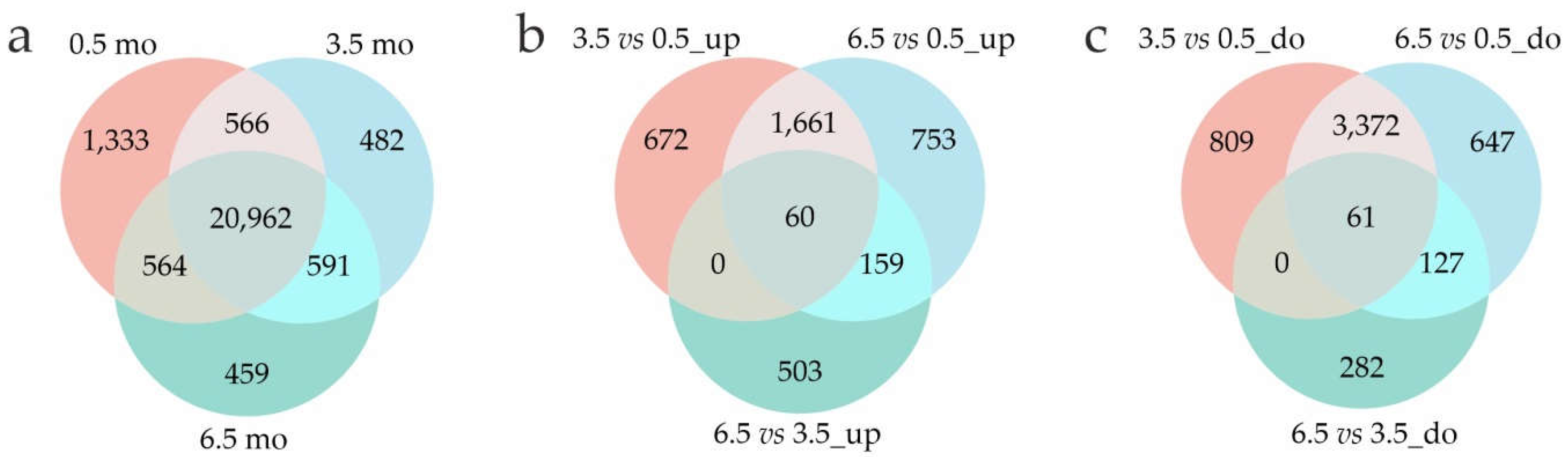
Figure 4.
Heatmap of DEGs related to storage proteins (a) and ETH signaling (b) in 0.5-mo (green), 3.5-mo (pink), and 6.5-mo (blue) S. tuberosum tubers based on the transcriptome data (RPKM). The color gradient indicates changes in gene expression from low (blue) to high (red); the names of the proteins corresponding to the LOC numbers are given in the Supplementary Table S12.
Figure 4.
Heatmap of DEGs related to storage proteins (a) and ETH signaling (b) in 0.5-mo (green), 3.5-mo (pink), and 6.5-mo (blue) S. tuberosum tubers based on the transcriptome data (RPKM). The color gradient indicates changes in gene expression from low (blue) to high (red); the names of the proteins corresponding to the LOC numbers are given in the Supplementary Table S12.
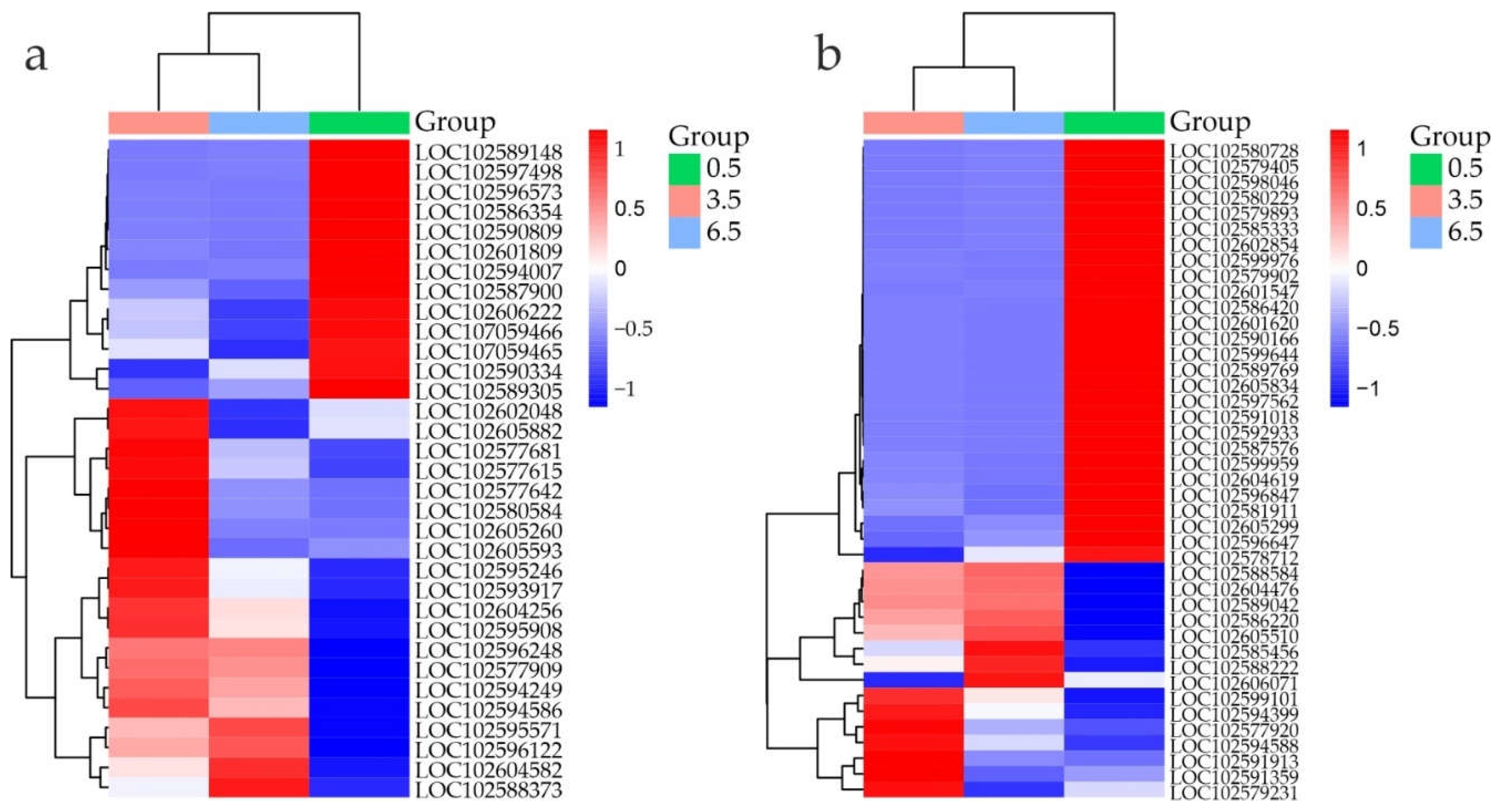
Figure 5.
Heatmap of DEGs related to the carotenoid/ABA/SL pathway (a) and GA (b), JA (c), and CK (d) signaling in S. tuberosum tubers at 0.5 (green), 3.5 (pink), and 6.5 (blue) months based on transcriptome data (RPKM). The color gradient indicates changes in gene expression from low (blue) to high (red); the names of the proteins corresponding to the LOC numbers are given in the Supplementary Table S12.
Figure 5.
Heatmap of DEGs related to the carotenoid/ABA/SL pathway (a) and GA (b), JA (c), and CK (d) signaling in S. tuberosum tubers at 0.5 (green), 3.5 (pink), and 6.5 (blue) months based on transcriptome data (RPKM). The color gradient indicates changes in gene expression from low (blue) to high (red); the names of the proteins corresponding to the LOC numbers are given in the Supplementary Table S12.
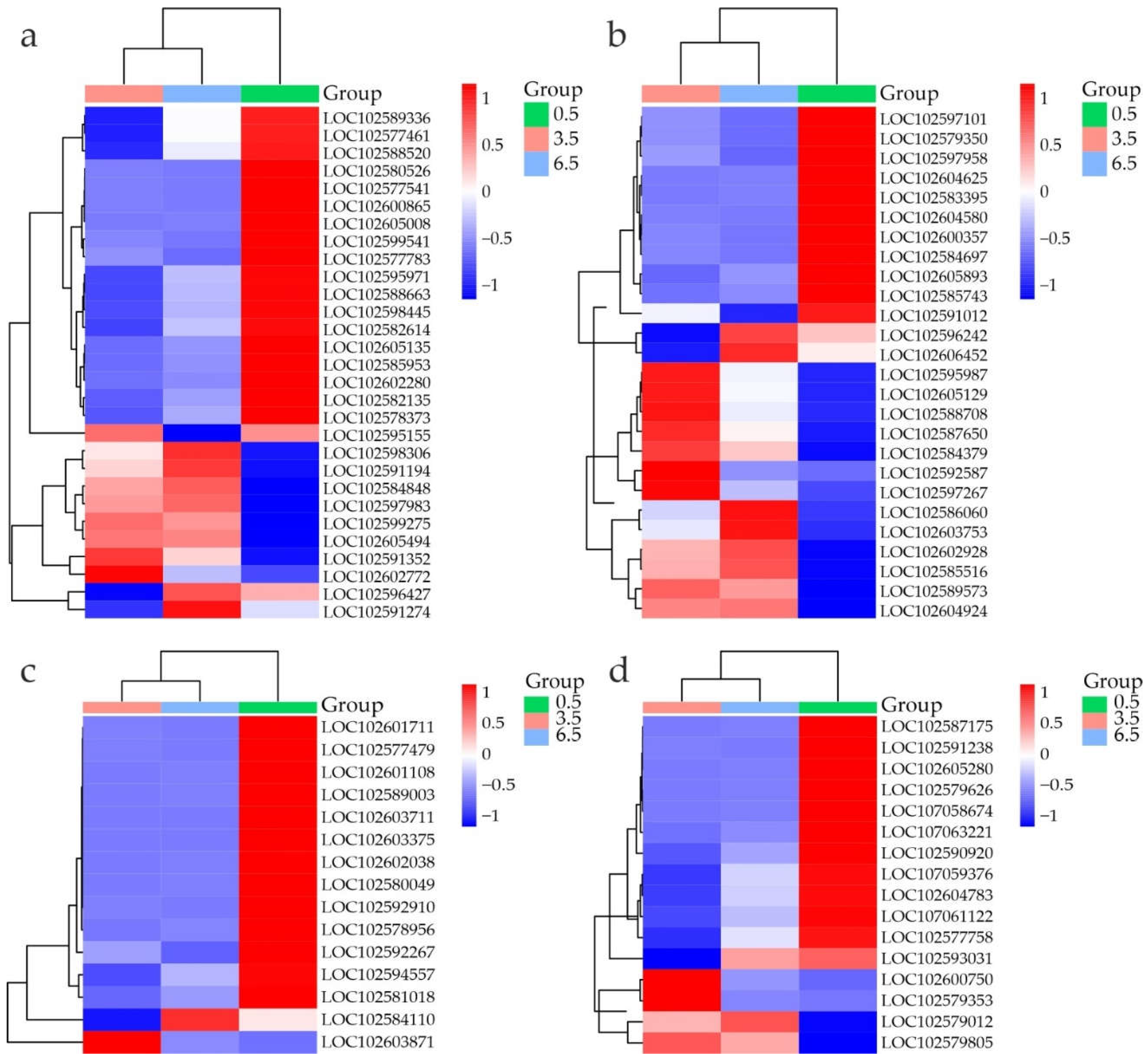
Figure 6.
Heatmap of DEGs related to auxin (a) and BR (b) signaling, in S. tuberosum tubers at 0.5 (green), 3.5 (pink), and 6.5 (blue) months based on transcriptome data (RPKM). The color gradient indicates gene expression changes from low (blue) to high (red); the names of the proteins corresponding to the LOC numbers are given in the Supplementary Table S12.
Figure 6.
Heatmap of DEGs related to auxin (a) and BR (b) signaling, in S. tuberosum tubers at 0.5 (green), 3.5 (pink), and 6.5 (blue) months based on transcriptome data (RPKM). The color gradient indicates gene expression changes from low (blue) to high (red); the names of the proteins corresponding to the LOC numbers are given in the Supplementary Table S12.
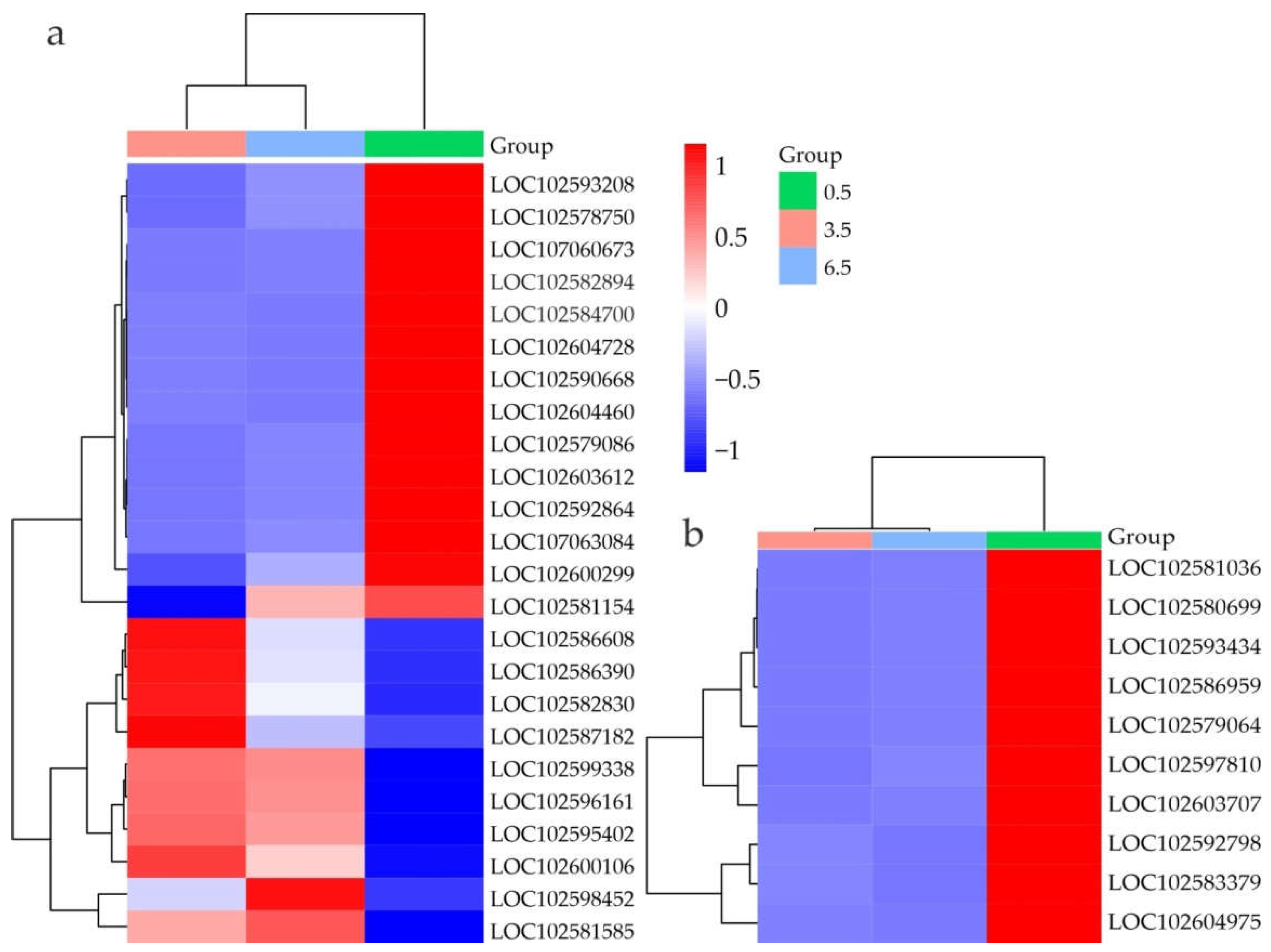
Figure 7.
Heatmap of DEGs related to cold response (a), flavonoid/anthocyanin synthesis (b), carbohydrate metabolism (c), and desiccation response (d) in S. tuberosum tubers at 0.5 (green), 3.5 (pink), and 6.5 (blue) months based on transcriptome data (RPKM). The color gradient indicates gene expression changes from low (blue) to high (red); the names of the proteins corresponding to the LOC numbers are given in the Supplementary Table S12.
Figure 7.
Heatmap of DEGs related to cold response (a), flavonoid/anthocyanin synthesis (b), carbohydrate metabolism (c), and desiccation response (d) in S. tuberosum tubers at 0.5 (green), 3.5 (pink), and 6.5 (blue) months based on transcriptome data (RPKM). The color gradient indicates gene expression changes from low (blue) to high (red); the names of the proteins corresponding to the LOC numbers are given in the Supplementary Table S12.
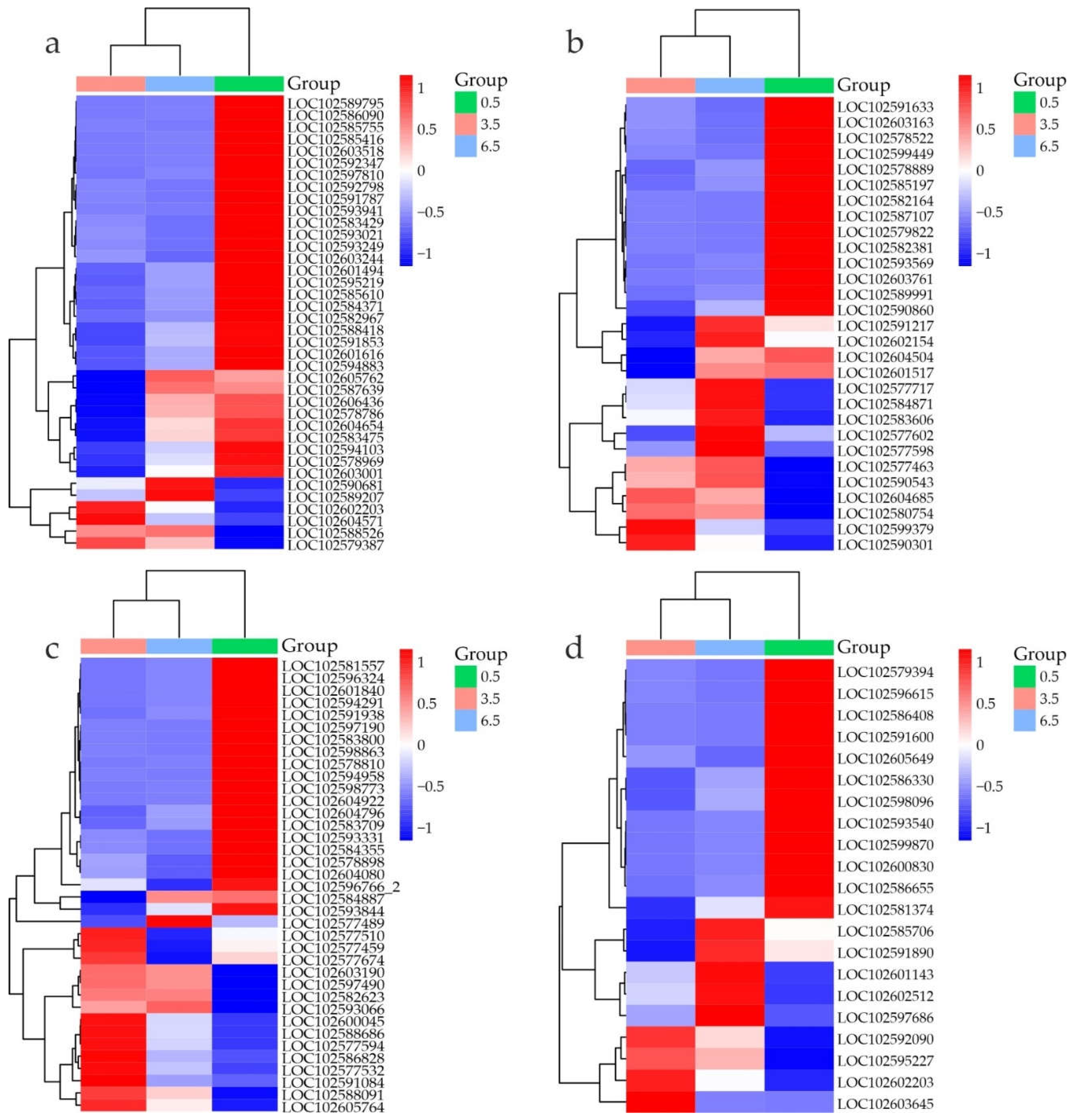
Figure 8.
Heatmap of TF DEG expression in S. tuberosum tubers at 0.5 (green), 3.5 (pink), and 6.5 (blue) months based on transcriptome data (RPKM). The color gradient indicates gene expression changes from low (blue) to high (red).
Figure 8.
Heatmap of TF DEG expression in S. tuberosum tubers at 0.5 (green), 3.5 (pink), and 6.5 (blue) months based on transcriptome data (RPKM). The color gradient indicates gene expression changes from low (blue) to high (red).
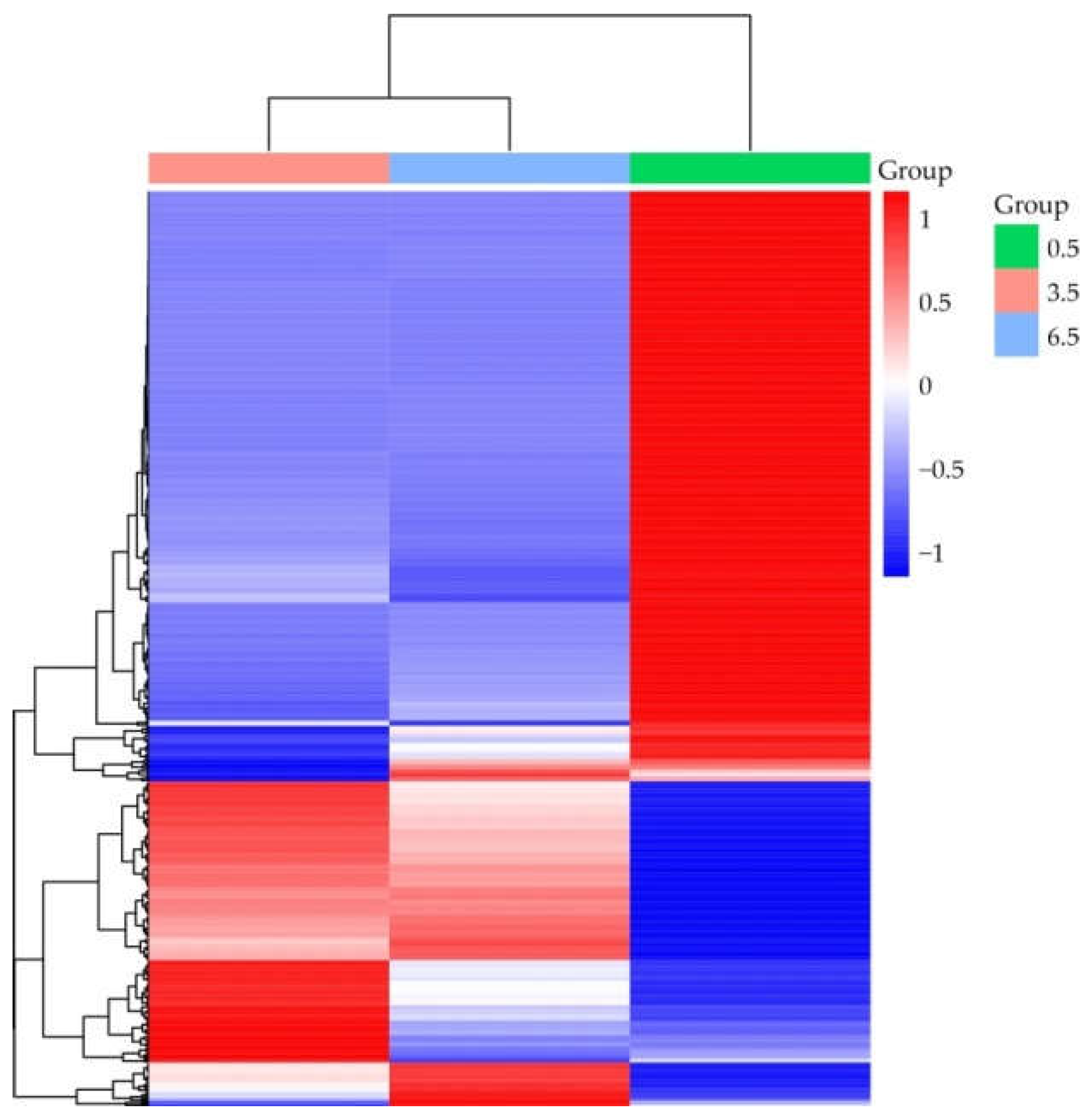
Figure 9.
Comparison of RT-qPCR and RNA-seq results on the expression of StBLH8, StSP5GB, POTM1, StSP5GA, StFDL1, StPHYB, StCOL16, and StGPT2 presented as RT-qPCR vs RNA-seq dual Y axis plots. a–cP < 0.05 indicates significant differences in RT-qPCR-data between 0.5-, 3.5-, and 6.5-mo samples.
Figure 9.
Comparison of RT-qPCR and RNA-seq results on the expression of StBLH8, StSP5GB, POTM1, StSP5GA, StFDL1, StPHYB, StCOL16, and StGPT2 presented as RT-qPCR vs RNA-seq dual Y axis plots. a–cP < 0.05 indicates significant differences in RT-qPCR-data between 0.5-, 3.5-, and 6.5-mo samples.
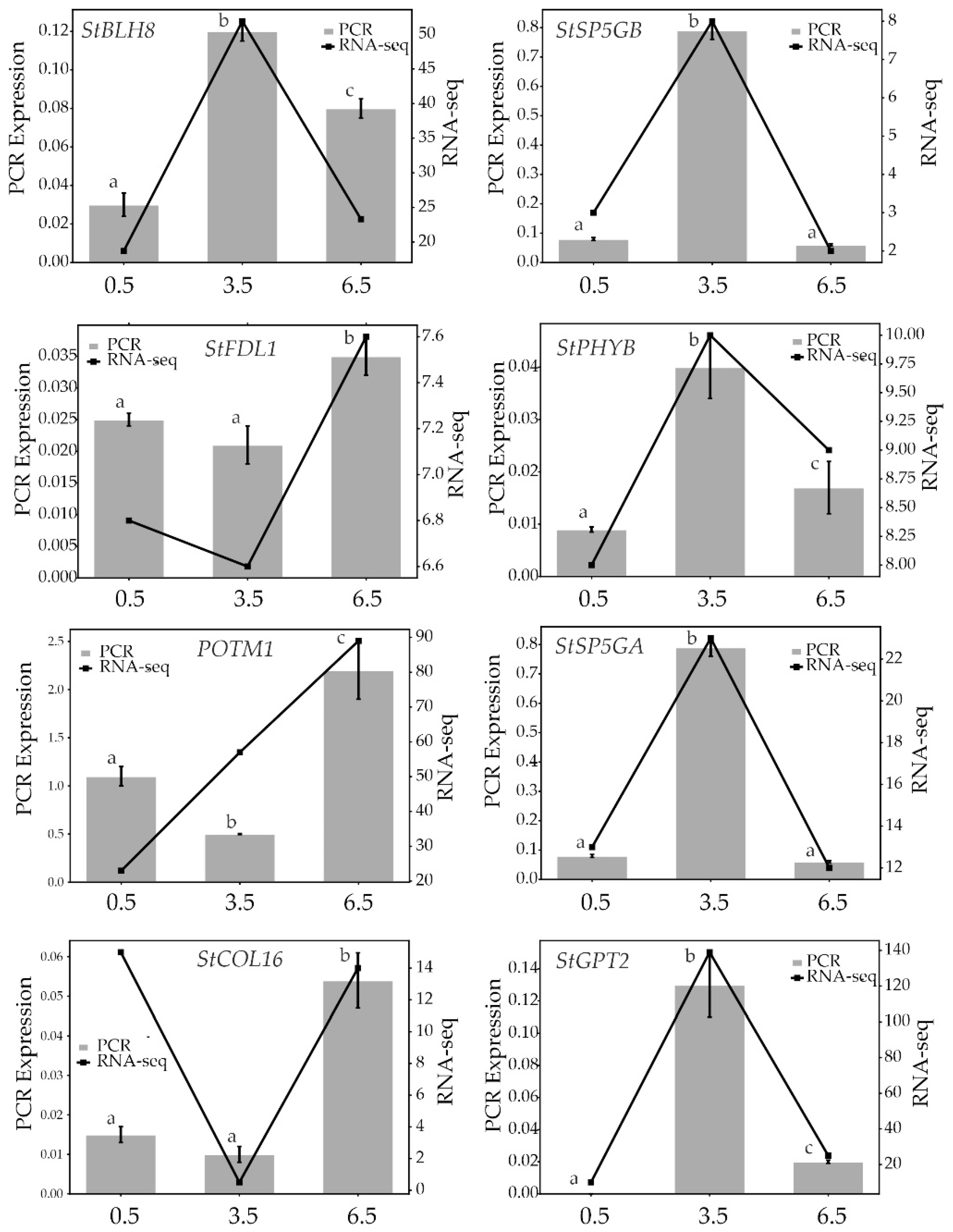
Figure 10.
Expression of the StBLH8, StSP3D, StFTL1-1, StFTL1-2, StSP5GA, StSP5GB, FDL1, StFDL, StCO, StCOL, StCOL2, StCOL16, StCDF1, StPHYB, and POTM1 genes in S. tuberosum non-dormant (6.5) tuber eye-buds and buds in stimulating conditions (6.5-mo tubers after 1 d (6.5-0), 7 d (6.5-1), and 9 d (6.5-2) at 20°C); the letters above the columns indicate the difference in gene expression between the corresponding eye-bud states (a–dP < 0.05).
Figure 10.
Expression of the StBLH8, StSP3D, StFTL1-1, StFTL1-2, StSP5GA, StSP5GB, FDL1, StFDL, StCO, StCOL, StCOL2, StCOL16, StCDF1, StPHYB, and POTM1 genes in S. tuberosum non-dormant (6.5) tuber eye-buds and buds in stimulating conditions (6.5-mo tubers after 1 d (6.5-0), 7 d (6.5-1), and 9 d (6.5-2) at 20°C); the letters above the columns indicate the difference in gene expression between the corresponding eye-bud states (a–dP < 0.05).
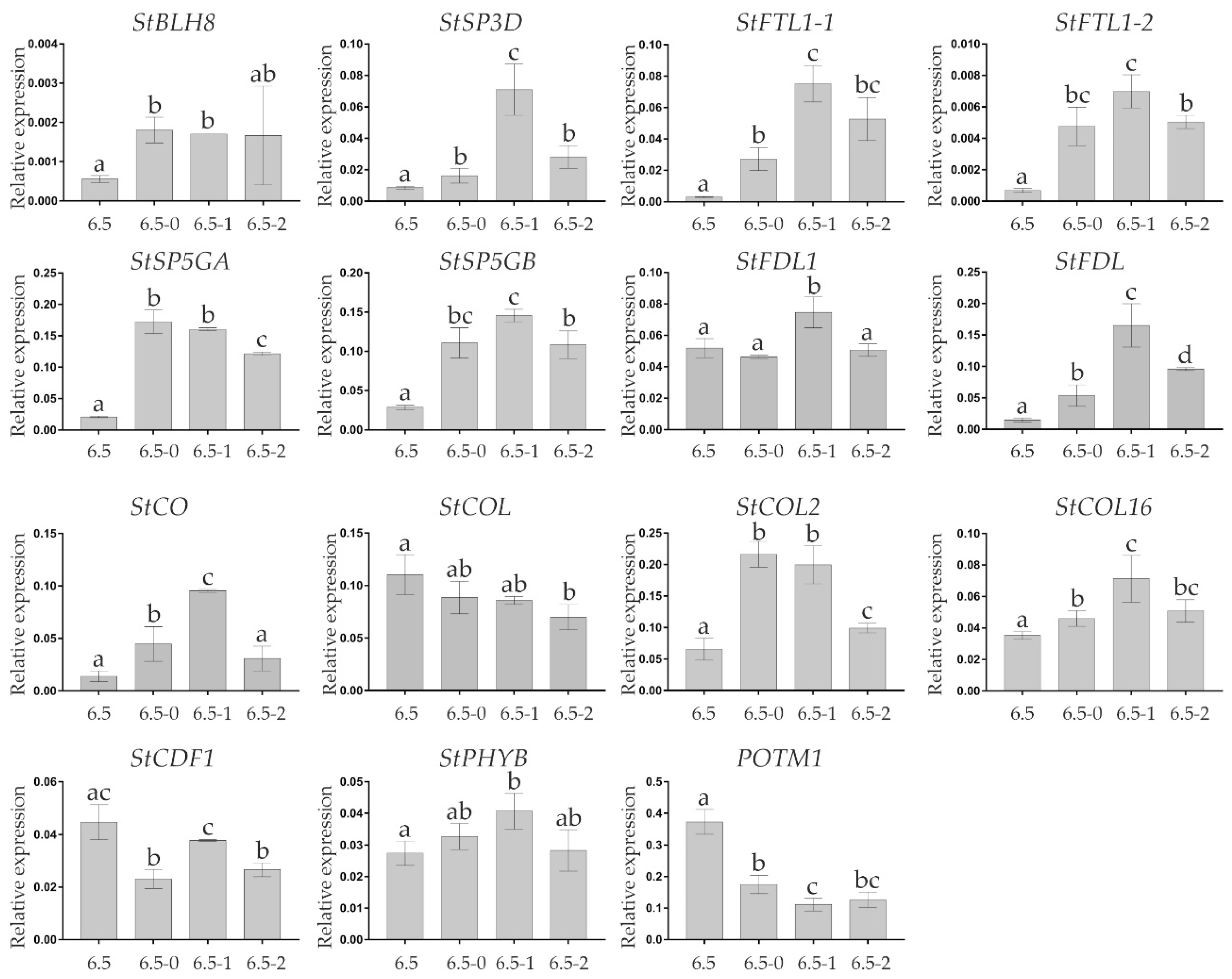

Table 1.
Enriched GO categories related to biological processes (BPs) and associated with the meristem and reproductive development.
Table 1.
Enriched GO categories related to biological processes (BPs) and associated with the meristem and reproductive development.
| GO: BP term | Pathway (number of genes in the category) | Upregulated DEGs | Downregulated DEGs | ||||
| 3.5 vs 0.5 | 6.5 vs 0.5 | 6.5 vs 3.5 | 3.5 vs 0.5 | 6.5 vs 0.5 | 6.5 vs 3.5 | ||
| GO:0009790 | Embryo development (103) | 20 | 17 | 0 | 0 | 0 | 0 |
| GO:0009793 | Embryo development ending in seed dormancy (99) | 19 | 17 | 0 | 0 | 0 | 0 |
| GO:0009791 | Post-embryonic development (431) | 63 | 59 | 22 | 68 | 62 | 25 |
| GO:0048507 | Meristem development (85) | 15 | 0 | 0 | 0 | 0 | 0 |
| GO:0010073 | Meristem maintenance (54) | 10 | 8 | 0 | 0 | 0 | 0 |
| GO:0099402 | Plant organ development (267) | 29 | 30 | 0 | 41 | 40 | 11 |
| GO:0010228 | Vegetative to reproductive phase transition of meristem (54) | 7 | 0 | 0 | 0 | 9 | 6 |
| GO:2000028 | Regulation of photoperiodism flowering (22) | 4 | 0 | 0 | 5 | 6 | 3 |
| GO:0003006 | Developmental process involved in reproduction (370) | 55 | 48 | 0 | 56 | 55 | 18 |
| GO:0022414 | Reproductive process (571) | 68 | 58 | 0 | 73 | 73 | 22 |
| GO:0009908 | Flower development (130) | 17 | 16 | 0 | 26 | 23 | 0 |
| GO:0010154 | Fruit development (154) | 23 | 21 | 0 | 0 | 0 | 7 |
Table 2.
Phytohormone signaling-releated DEGs at 3.5 vs 0.5 months.
| Phytohormone | DEGs and their functions |
|---|---|
| ETH | 43 DEGs associated with ETH signaling were identified. The upregulated DEGs encode ETHYLENE-OVERPRODUCTION PROTEIN 1 (ETO1), ETHYLENE INSENSITIVE 3-like (EIN3), 13 ETH-responsive TFs (ERFs), and three ETH receptors homologous to Arabidopsis thaliana ETHYLENE RESPONSE 1 (ETR1), ETR2, and ETHYLENE RESPONSE SENSOR 1 (ERS1); the most upregulated genes were ETH-responsive ERF114-like and RAP2-3 (Figure 4b). |
| ABA, SLs | 30 DEGs associated with carotenoid/ABA/SL synthesis/signaling were identified. All DEGs related to carotenoid metabolism, such as NCED1 and 3 (9-cis-epoxycarotenoid dioxygenase), AAO (abscisic-aldehyde oxidase), and ABA 8'-hydroxylase genes were downregulated (Figure 5a). CCD8 (carotenoid cleavage dioxygenase) associated with SL synthesis [14] was upregulated (Figure 5a), supporting the notion about ABA content decrease. DEGs encoding ABA signaling-related proteins, including ABA receptors (PYL4-like LOC102602772 and LOC102580526, PYL9-like, and PYL8-like), ABA-responsive element-binding factors (ABF), ABA-INSENSITIVE 5-like protein 4 (ABI5-like 4), and sucrose non-fermenting-1-related protein kinases 2 (SnRK2) [29] were either upregulated (PYL4-like LOC102602772, PYL9-like, ABI5-like 4, four SnRK2 genes) or downregulated (PYL4-like LOC102580526, PYL8-like, ABF-binding bZIP-TFs, one SnRK2). |
| JAs | 16 DEGs associated with JA synthesis/signaling were identified. DEGs related to JA signaling, such as TIFY (JA ZIM-domain protein), TPL (TOPLESS), and AOS (allene oxide synthase, the first enzyme in JA synthesis), were downregulated, except for one of the nine TIFY genes, TIFY8, which was activated by 2.6-fold (Figure 5c). In response to JA, the F-box protein CORONATINE-INSENSITIVE 1 (COI1) enters the SCFCOI1 E3 ubiquitin ligase complex involved in the degradation of JA signaling negative regulators TIFY and represses the JA cascade through interaction with corepressor TPL [30]. The expression of the COI1 gene, the only positive regulator of JA signaling among the listed, did not change at 3.5 vs 0.5 months (although it was increased by 1.5 times at 6.5 vs 0.5/3.5 months (Figure 5c), which may be related to the inhibition of non-dormant bud sprouting). Thus, JA signaling was likely downregulated in 3.5-mo tubers because of TIFY8 activation and JA synthesis suppression. |
| GAs | 27 DEGs associated with GA synthesis/signaling were identified. The expression of the DEGs encoding GA synthesis enzymes copalyl pyrophosphate synthase (CPS), Ent-kaurene synthase (KS), Ent-kaurene oxidase (KO), GA20 oxidase (GA20ox), GA3 oxidase (GA3ox), and GA2 oxidase (GA2ox) changed in both directions: some (CPS, two GA20ox, one GA3ox, and three GA2ox) were repressed, whereas the other (KS, KO, one GA20ox, and one GA3ox) – activated. The GAI- and RGL1-like genes of DELLA TFs, which negatively regulate GA signaling but activate GA receptor (GID1) [31,32], were downregulated as well as one of the GID1B-like genes, whereas the other gene was upregulated. The expression of most genes that belong to the nuclear factor Y (NF-Y) TF complex and postively regulate GA-mediated dormancy [14,31], including StNF-YA8 and StNF-YB20, was increased and that of StNF-YC5 – decreased (Figure 5b). |
| CKs | 17 DEGs associated with CK synthesis/signaling were identified. The expression of CK metabolism/signaling genes [14,33] changed insignificantly or was repressed, with the exception of IPT (isopentenyl transferase regulating CK accumulation), two ABCG14-like (regulating CK efflux across the plasma membrane), and ARR17-like (type-A response regulator negatively affecting CK signaling) genes which were activated. One of the CK-responsive genes WUS (WUSCHEL TF maintaining the stem cell niche in the shoot meristem) was strongly upregulated, whereas WUS-related homeobox 8-like (WOX8) was downregulated (Figure 5d). |
| Auxins | 25 DEGs associated with Auxin signaling were identified. Auxin target genes [34] such as two SAUR (SMALL AUXIN UP-REGULATED RNA) and GH3 (GRETCHEN HAGEN 3 indole-3-acetic acid-amido synthetase) DEGs were upregulated, whereas four SAUR and five GH3 DEGs were downregulated (Figure 6a). ARF5 (auxin response factor) and four AUX/IAA (AUXIN RESISTANT/INDOLE-3-ACETIC ACID INDUCIBLE repressor) genes were upregulated, and two IAA genes were downregulated (Figure 6a). Two ARF6-like genes were slightly upregulated (Figure 6a); the upregulation of the ARF6 gene in tuber buds during dormancy release has also been shown earlier [35]. |
| BRs | 12 DEGs associated with BR synthesis/signaling were identified. Of the five genes encoding important enzymes in the GA biosynthetic pathway (Ent-kaurenoic acid oxidase 2 (KAO2), deetiolated2 (det2), dwarf1 (DWF1), and DWF5 [36]), only DWF5 expression was significantly changed (upregulated) at 3.5 vs 0.5 months. BIN2 (BR receptor BRASSINOSTEROID-INSENSITIVE 2 kinase [34]), three WRKY53 and two BZR1 (BRASSINAZOLE-RESISTANT 1) TF-encoding genes (positive regulators of BR-related processes [37]), and five EXO (EXORDIUM mediator of BR-promoted growth [38]) genes were downregulated at 3.5 and 6.5 vs 0.5 months (Figure 6b). |
Table 3.
Selected TF-encoding DEGs related to flowering/tuberization.
| Gene ID | Protein | 3.5 vs 0.5 | 6.5 vs 0.5 | Comments |
|---|---|---|---|---|
| LOC102585059 | Agamous-like MADS-box protein AGL27 | 5.3 up | 3.1 up | Belongs to the FLC subfamily consisting of FLC, MADS AFFECTING FLOWERING 1 (MAF1, AGL27, or FLOWERING LOCUS M (FLM)), MAF2, MAF3, MAF4, and MAF5; some interact with SHORT VEGETATIVE PHASE (SVP) to inhibit flowering (cold dependent) by directly repressing FT and SOC1 transcription [64]. |
| LOC102603965 | Flowering locus K homology domain-like (FLK) | 3.0 up | 3.5 up | FLK and FPA promote flowering by suppressing FLC. [65]. |
| LOC102593682 | Flowering time control protein (FPA) | 1.6 up | 2.6 up | |
| LOC102598110 | FRIGIDA-like 3 | 2.6 up | 3.7 up | FRI induces FLC transcription, which is epigenetically repressesed by vernalization [64]; FRI-like protein 3 (FRL3) functions in flowering suppression [66]; FRI-ESSENTIAL 1 genetically interacts with FRI and FRI-LIKE 1 to promote FLC expression [67]. |
| LOC102588299 | FRIGIDA-like 4a | 2.2 up | 2.3 up | |
| LOC102597500 | FRIGIDA-ESSENTIAL 1-like | 2.1 do | 3.7 do | |
| LOC102589537 | FRIGIDA-like (FRI) | 2.1 up | 2.3 up | |
| LOC102593387 | VIN3-like protein 1 (VIL1) | 2.3 do | 2.0 do | VIL1 participates in both photoperiod and vernalization pathways by regulating FLC and FLM expression. In the vernalization pathway, VIL1, along with VIN3, is necessary for the modifications of FLC and FLM chromatin associated with the epigenetically silenced state and acquisition of competence to flower [68]. |
| LOC102599642 | Agamous-like MADS-box protein AGL19 | 4.9 do | 4.3 do | AGL19 acts in the FLC-independent vernalization pathway; elevated AGL19 levels result in the activation of LEAFY and APETALA1 and flowering [69]. |
| LOC102601949 LOC102603864 |
SENSITIVITY TO RED LIGHT REDUCED 1-like | 4.8 do 4.2 do |
4.3 do - |
SRR1 delays flowering by controlling FT repression [70]. |
| LOC102589240 | Transcription factor TCP8-like | 7.9 do | 7.2 do | TCP8 upregulation leads to FLC increase [71]. |
| LOC102588708 | Nuclear transcription factor Y subunit B-3-like | 8.2 do | 4.1 do | NF-YB3 is essential for flowering induction through regulation of FT expression [72]. |
| LOC102597323 | MYB family transcription factor APL | 136 do | 91.7 do | APL promotes flowering through FT activation [73]. |
| LOC102593569 LOC102603163 |
TRANSPARENT TESTA GLABRA 1-like | 22.7 do 9.7 do |
13.8 do 30.1 do |
TTG1 can repress the transcription of floral integrators FT and SOC1 and activate that of FLC [74]. |
| LOC102606359 | B-box zinc finger protein 19-like | 4.5 do | - | BBX19 suppresses seed germination via ABI5 induction [75] and interacts with CO to repress FT transcription, thus defining flowering time [76]. |
| LOC102580467 | MOTHER of FT and TFL1 | 50 up | 15 up | MOTHER-OF-FT-AND-TFL1 (MFT) is seed dormancy-promoting factor [53]. |
| LOC102606292 | FLOWERING LOCUS T-like (StSP5GB) | 2.4 up | ˗ | StSP5G genes are repressors of tuber formation [4]; FT is involved in temperature responses and has an uncharacterized role in the promotion of seed germination [77]. |
| LOC102579194 | FLOWERING LOCUS T-like (StSP5GA) | 1.7 up | ˗ | |
| LOC102583457 | GIGANTEA-like | 11 up | 5.8 up | A. thaliana accessions with gigantea (gi) mutations have significantly increased seed dormancy; the phenotype is shared by constans (co) and flowering locus t (ft) mutants [77]. In potato, GI is involved in flowering and tuber initiation [6]. |
| LOC102578719 | GIGANTEA-like | 2.2 up | ˗ | |
| LOC102577795 | Agamous-like MADS-box protein AGL8 homolog (POTM1) | 2.4 up | 3.8 up | POTM1 (StMADS1) stimulates the transcription of tuberigen StSP6A in a feedback manner [7]. |
| LOC102591375 | Agamous-like MADS-box protein AGL62 | 3.4 do | 4.2 do | AGL62 regulates the transport of auxin (positively associated with dormancy release) from endosperm [78]. |
| LOC102596767 | Floral homeotic protein AGAMOUS-like | 3.7 do | 9.2 do | A homolog of AG-clade gene SHATTERPROOF 1 (SHP1, AGL1) is upregulated during flower bud endodormancy to ecodormancy transition in Prunus species during chilling [79]. |
| LOC102577664 | MADS transcriptional factor STMADS11 | 6.5 do | 12.6 do | STMADS11 and its homolog STMADS16 belong to the SVP/AGL24-clade MADS-box genes, which include DORMANCY-ASSOCIATED MADS-box (DAM); DAM/SVP represses the bud dormancy break; DAM expression is decreased during dormancy release in response to prolonged chilling [80]. |
| LOC102577849 | MADS16 protein | 1.4 up | ˗ | |
| LOC102593715 | MADS-box TF 23-like | 2.0 do | 2.7 do | AGL16 homolog directly interacting with SVP and indirectly with FLC; reduces flowering time [81]. |
| LOC102601650 | MADS-box protein SOC1-like | 3.8 do | 1.8 do | SOC1 is linked to flowering regulation and plays a key role in bud dormancy transition [82]. |
| LOC102586019 | MADS-box protein SOC1-like | 2.1 do | ˗ | |
| LOC102582884 | MADS-box TF 23-like | 8.6 up | 3.9 up | AGL21 homolog invoved in seed germination and early post-germination growth [56]. |
| LOC102597046 | EARLY FLOWERING 3 (ELF3) | 2.1 do | 2.1 do | ELF3 and ELF4 are negative regulators of flowering time [83] and may have a role in seed dormancy control [84]. |
| LOC102586641 | EARLY FLOWERING 3-like | 15.6 do | 5.9 do | |
| LOC102587775 | ELF4-LIKE 3 | 1.9 up | 2.2 up | |
| LOC102577671 | CONSTANS | 12.4 do | 3.6 do | In potato, CO, COL2, and COL1 activate anti-tuberigen expression and function as regulators of the tuberization pathway [6]. |
| LOC102587352 | zinc finger protein CONSTANS-like | 4.7 do | 5.9 do | |
| LOC102587905 | CONSTANS-LIKE 2 | 14.0 do | 10.1 do | |
| LOC102598089 | CONSTANS-LIKE 2-like | 2.9 do | 6.4 do | |
| LOC102585080 | CONSTANS-LIKE 4-like | 3.9 do | 1.5 do | COL4 is a flowering repressor [85]. |
| LOC102578495 | CONSTANS-LIKE 4-like | 3.1 do | 1.4 do | |
| LOC102582832 | CONSTANS-LIKE 5-like | 3.5 do | 3.2 do | COL5 acts as a flowering inductor [86]. |
| LOC102600093 | CONSTANS-LIKE 3-like | 3.9 do | ˗ | LONG HYPOCOTYL 5 (HY5)-COL3-COL13 regulatory chain functions in hypocotyl elongation [87]. |
| LOC102582339 | CONSTANS-LIKE 13 | 3.2 do | 3.7 do | |
| LOC102588526 | transcription factor HY5 | 5.1 up | 5.4 up | HY5 is positively associated with dormancy release [88]. |
| LOC102603307 | cyclic dof factor 1 | - | 1.9 up | StCDF1 represses CO expression, stimulating tuberization [5]. |
| LOC102597615 | cyclic dof factor 1-like | 13–15 RPKM* | ||
| LOC102577460 | BEL1-related homeotic protein 5 |
29–65 RPKM* | StBEL5 together with POTH1 positively regulates the expression of StCDF1, StSP6A, and some StSP6A target genes [4,6]. | |
| LOC102602563 | FLOWERING LOCUS D (FD) | 7–8 RPKM* 13–20 RPKM* |
FD is included in both FAC and TAC [3]. | |
| LOC102593535 | Agamous-like MADS-box protein AGL27 | Flowering repressors [64]. |
||
| LOC102605812 | Truncated TF CAULIFLOWER A-like (MAF4 homolog) | 4–6 RPKM* | ||
| LOC102605245 | MADS AFFECTING FLOWERING 5-like | 14–21 RPKM* | ||
Note: * indicates insignificant gene expression changes between 0.5-, 3.5-, and 6.5-mo tubers; “up” or “do” – upregulated or downregulated, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



