
Submitted:
17 January 2026
Posted:
20 January 2026
You are already at the latest version
Abstract
The “Great Filter” hypothesis proposes that the apparent absence of extraterrestrial civilizations, despite the vastnumber of potentially habitable planets, is the result of one or more extremely improbable steps, or “filters,” in theevolution of intelligent life. These filters may lie either in humanity’s future, predicting our eventual self-destruction, or in our evolutionary past, implying that intelligent civilizations are rare. Based on empirical evidenceand evolutionary analysis, this study argues that no probable future filter exists with sufficient destructive capabilityto account for the absence of evidence of extraterrestrial intelligent life. Instead, several improbable and sequentiallydependent events in Earth’s geological and biological history have been hypothesized as effective filters, includingthe Theia collision initiating plate tectonics, tectonic activity providing sufficient sustained chemical energy toenable abiogenesis, and the evolutionary dominance of mammals. By estimating the probabilities of these unlikelyevents, the probability of a civilization arising was calculated to be less than one per ~1040 star systems—while thereare estimated to be only ~1030 stars in the observable universe. These findings suggest that Earth may host the onlytechnological civilization within its cosmic horizon.
Keywords:
great filter
; Fermi paradox
; abiogenesis
; evolution
; Rare Earth hypothesis
1. Introduction to the Great Filter and Fermi Paradox
Since Marconi transmitted the first radio waves in 1895, radio broadcasts and other signals emitted by our technology have been escaping Earth’s atmosphere, forming a “radiosphere” that is propagating into space. It has been estimated that these signals from Earth have travelled approximately 100 light-years into space (Kaltenegger and Faherty, 2021), intersecting thousands of star systems that are appropriately located to detect signals of life from Earth. Over time, the radiosphere of human civilization has become more concentrated with the development and use of advanced technologies. Considering the total number of stars in the observable universe (3 × 1023), there are approximately 3 × 1019 stars within a 1 billion light-year radius of the solar system (Marov, 2015), with numerous habitable planets (~1022) that could potentially sustain life (Cassan et al., 2012) and develop advanced civilizations. Despite the high probability of intelligent life elsewhere in the Universe, we have not yet detected conclusive evidence for extraterrestrial life. This contradiction is known as the Fermi Paradox (Wandel, 2022).
Numerous hypotheses have been proposed to explain the phenomenon. For example, intelligent life is extremely rare or non-existent elsewhere in the universe; the development of advanced technology by intelligent species is rare; technological development takes too long, and technological civilizations invariably destroy themselves or are driven to extinction by natural disasters or cosmic events before leaving evidence of their existence; alien civilizations may be too far away, may transmit signals incomprehensible to humans (like neutrino signals), or may choose to hide their existence. Another possible explanation for the Fermi paradox is the “Great Filter” hypothesis, which proposes that there are certain evolutionary or developmental barriers that prevent civilizations from advancing, thus making intelligent life exceedingly rare (Aldous, 2010; Hanson, 1998; Verendel and Häggström, 2015). In the case of human civilization, the Great Filter event may lie in our past (a rare evolutionary event) or future (a destructive event that prevents other civilizations from observing our existence). The hypothesis that this filter occurs in the future is largely speculative. Assuming that similar filters are the reason why we do not observe evidence of extraterrestrial life, and considering the immense number of stars and habitable planets in the observable universe, a Great Filter would need to be destructive enough to eliminate the vast majority of civilizations, a scenario that seems implausible given the difficulty of completely eradicating a civilization. Natural disasters such as volcanoes or cosmic impacts are unlikely to be future filters for humanity because they are infrequent, and human civilization has the potential to become multi-planetary in the future, thereby reducing such risks. Therefore, the author posits that the most plausible future filter is self-destruction.
Three commonly discussed self-destruction scenarios include nuclear war, excessive global warming, and conflict with or domination by artificial intelligence (AI). While all three scenarios are theoretically possible, there is little evidence to suggest that any one is probable enough to qualify as a Great Filter. Although nuclear war could destroy human civilization, the principle of mutually assured destruction makes such an event unlikely. Moreover, even if a nuclear conflict occurs, underground survivors could potentially rebuild their civilization over time. The main risk of climate change is the runaway greenhouse effect, in which warming melts ice caps, reduces polar albedo, and accelerates further warming. However, this is unlikely as a Great Filter, as global warming is reversible in the long term using existing technologies, such as carbon capture, and the Earth’s natural systems have great capacity for self-repair. Temporary mitigation is also possible through geoengineering methods such as releasing reflective aerosols into the upper atmosphere to cool the planet. Although it is unlikely that humans will intentionally create AI capable of destroying our civilization, the unintentional development of AI technology that escapes our control is considered a real concern by many (Bostrom, 2014). However, AI-induced extinction does not appear sufficient to serve as a universal filter.
The author argues that a future Great Filter appears less likely than an improbable event in Earth’s evolutionary history that made intelligent life possible; that is, the Great Filter has already occurred in the past. This view is supported by multiple events and by many of the characteristics of the Solar System, as discussed in detail in the book “Rare Earth: Why Complex Life Might Be Uncommon in the Universe” (Ward and Brownlee, 2000). This book is also the source of the Rare Earth Hypothesis, which states that complex life is exceedingly rare in the universe.
Building on this background, this study analyzes the potential filters in the Earth’s evolutionary past and uses statistical methods to show that Earth may be the only planet in the observable universe to host a civilization. The Great Filter hypothesis has been studied via statistical analysis (Aldous, 2010). Furthermore, Bayesian analysis of the Great Filter hypothesis compared the outcomes of a single linear history and branching pathway of intelligence during evolution (Verendel and Häggström, 2015), showing that the statistical evidence supporting it depends greatly on the selection of the prior. Other studies have mathematically evaluated the Rare Earth hypothesis (Forgan and Rice, 2010; Sandberg et al., 2018). Many prior studies have briefly discussed possible filters, including the well-known evolutionary events proposed by Hanson as possible filters (Hanson, 1998). In contrast, this study proposes various filters in the Earth’s past and attempts to quantify their relative probabilities of being the Great Filter event.
2. Earth is Likely to Host the Only Civilization in the Observable Universe
2.1. Statistical Analysis: The 30% Rule and Principle of Mediocrity
It took at least 3.8 billion years for life on Earth to evolve from single-celled organisms to civilization. Current estimates suggest that the Earth has approximately one billion years remaining before the sun’s increasing luminosity renders it uninhabitable (Ozaki and Reinhard, 2021). Comparing this timescale to the duration of Earth’s evolutionary history, it can be argued that life fortuitously evolved fast enough to develop an advanced civilization. As shown in Figure 1, if evolution proceeded 30% slower, the Sun would likely have destroyed life on Earth before intelligent life could emerge. This is hereafter referred to as the “30% rule.”
The principle of mediocrity states that “if an item is drawn at random from one of several sets or categories, it's likelier to come from the most numerous category than from any one of the less numerous categories” (Kukla, 2009). This principle aligns closely with basic probability theory, considering random sampling from a population divided into categories, where the probability of selecting an item from a particular category depends on the category’s size (i.e., frequency). For n categories (C1, C2, …, Cn), the total number of items is N = N1 + N2 + … + Nn and the probability that a randomly drawn item belongs to Ci is P = Ni/N. Therefore, it is reasonable to assume that human civilization is a typical ordinary civilization, meaning that it is most likely one of the most numerous types of civilizations (i.e., a mammal-dominant civilization). Any of the conditions that allowed human civilization to become mammal-dominant (30% or more) likely arose in the most common (ordinary) way possible. To clarify, mammal dominance follows the 30% rule because mammals are necessary for civilization as explained in Section 2.2. Therefore, the probability of human existence can serve as a general probability for the existence of other intelligent species, which is calculated by estimating the combined probability of all 30% rule aspects.
2.2. Probability of Dinosaur Extinction
The dinosaur extinction event was particularly notable because it killed all non-avian dinosaurs, whereas mammals survived and thrived because of the lack of larger predators. If the dinosaur extinction event followed the 30% rule, this implies that the following three conditions must be true:
- Egg-laying tetrapods are naturally more ecologically dominant than mammals.
- Mammals naturally create civilization if they are dominant in an ecosystem.
- Reptile-dominated ecosystems cannot evolve into civilizations, given any amount of time.
Therefore, it is reasonable to assume that mammals do not naturally outcompete egg-laying tetrapods. Because fish lay eggs, it is reasonable to assume that the first tetrapods that moved onto land also laid eggs. Egg-laying offers many advantages, such as the ability to grow large and produce many offspring, which causes evolution to accelerate. Giving birth to free-living offspring has become increasingly difficult owing to increasing birth sizes (Werner and Griebeler, 2011). Assuming the mediocrity principle, egg-laying species are expected to be more dominant than mammals.
Mammals are more likely to evolve into civilization-building species, whereas reptiles tend to evolve into physically dominant organisms (such as dinosaurs). This difference is attributed to their distinct reproductive strategies: mammals give birth to free-living offspring, whereas reptiles lay eggs. Species with live birth have an incentive to nurture and care for their young because the offspring are typically born smaller and weaker than is necessary for independent survival. This evolutionary adaptation shortens the gestation period and reduces maternal risks. Numerous post-Cretaceous mammalian species (e.g., orcas, chimpanzees, and humans) have evolved complex social structures rooted in parenting (Royle et al., 2016). In contrast, egg-laying animals have fewer incentives to parent their young. Egg-laying creatures probably do not interact with young (this applies especially to dinosaurs). The lack of dinosaur eggshells suggests that most dinosaurs laid soft-shelled eggs that were buried to provide a protective environment for incubation, rather than being brooded and hatched by a parent (Chapelle et al., 2025). Although there is evidence that some dinosaurs parented their young (Horner, 2000), as with many modern reptiles, dinosaur young were likely born highly self-sufficient, and social interactions were minimal. Dinosaurs were the dominant animals on Earth for 165 MY, whereas it only 66 MY were required for mammals to evolve from rats to modern-day humans with global civilization.
Assuming that a dinosaur extinction event follows the 30% rule, this set of events would constitute one of the most common ways to eliminate the dinosaurs while allowing mammals to survive. Four major cosmic events appear in the fossil record (Figure 2), three of which were significant in driving the extinction of dinosaurs. The first event that occurred was the eruption of the Deccan Traps, approximately three million years ago (Pande, 2002). Approximately 150 KY later, the Chicxulub meteor, with an estimated diameter of 10 km, impacted sulfur deposits in the Yucatán Peninsula (Schulte et al., 2010a). This meteor impact is thought to have triggered a magnitude 11 earthquake that would have reached the Deccan Traps and magnified their volcanic activity, while the sulfur dioxide released from the Yucatán Peninsula would have caused a 10-year extreme-cooling effect (Gertsch et al., 2011). The next significant event was the Shiva impact, in which a meteor ~40 km in diameter hit the Deccan Traps (Chatterjee et al., 2006), enhancing volcanic eruptions more significantly than the Chicxulub meteor. In contrast, the event that produced the Boltysh crater does not seem significant because it is only 1 km in diameter, and the resulting meteorological event probably did not impact a geologically important area (although is suspected to have a role, owing to its proximity in time).
Therefore, a reasonable lower-bound probability for a dinosaur extinction event was determined by calculating the probabilities of these three significant cosmic events occurring within the observed timeframe. For simplicity, and because of the uncertainty in pinpointing the age of the Shiva event, the probabilities of these events occurring within a 2 MY timeframe were determined. Table 1 lists the event rates and probabilities of at least one event occurring within this timeframe.
2.3. Probability of End-Devonian Tetrapod Transition
The evolutionary transition from fish to tetrapods occurred during the end-Devonian extinction, punctuated by two separate events. The Kellwasser event (372 Mya) was a period of ocean anoxia (Carmichael et al., 2019), whereas the Hangenberg event (359 Mya) was a period of both ozone depletion and ocean anoxia (Lyu et al., 2025). The precise cause of the Kellwasser event is unknown, but is likely to be submarine volcanism. The Hangenberg Event may have been caused by a supernova (Fields et al., 2020). Assuming that the end-Devonian tetrapod transition followed the 30% rule, these events aided the transition from fish to land animals. This may also imply that civilization tends to occur on land.
The oldest known example of a tetrapod transition is Tiktaalik, a genus of lungfish that lived 375 Mya (Daeschler et al., 2006). Anoxia during the Kellwasser and Hangenberg periods provided favorable conditions for this genus and its evolutionary lineage because anoxia weakened its non-lunged competitors. The anoxia incentivized it to venture into shallow water while keeping its head above the water to obtain oxygen from the air. This would have driven its ancestors to evolve more efficient legs for walking, as illustrated in Figure 3 (Shubin et al. 2006).
The effects of the supernova that occurred 359 Mya can be analyzed in terms of its specific timing within the fossil record. Acanthostega, which lived 365 Mya, is the closest transitional fossil to this event (Sanchez et al., 2016). While its limbs have leg-like qualities, their bone structure suggests that they were used primarily as fins (Figure 4) (Clack, 2002). All fossils of this lineage dating from before the supernova 359 Mya show evidence that they were primarily aquatic animals, with the first non-aquatic tetrapods occurring after this supernova. After this supernova, all lineages had five digits and asymmetrical metatarsals, illustrating completion of the transition.
The principle of mediocrity assumes that the transition from lungfish to non-aquatic tetrapods occurred in the most ordinary manner possible. Therefore, because the Kellwasser and Hangenberg events were the major factors influencing the environmental and geological conditions during this timeframe, it is reasonable to assume that they were the driving forces of this transition. Recent in-depth studies on the causes of catastrophic Kellwasser events have highlighted kimberlite-like eruptions and/or orbital modulation (Racki, 2020), ocean anoxia (Carmichael et al., 2014), and volcanic activity (Carmichael et al., 2019) as the major factors. Ocean anoxia tends to be the leading explanation; however, it is unclear whether other unlikely factors are also involved. Harsh environmental conditions that lead to mass extinction events are known to clear ecological niches and reduce competition, thus allowing the surviving lineages to diversify rapidly. Environmental stressors place strong selective pressure on evolutionary processes. For example, ocean anoxia played a significant role in the Permian extinction, and the post-extinction recovery saw accelerated speciation in fish and mollusks driven by the novel environmental conditions (Aldous, 2010). Moreover, rapid genetic adaptation to toxic pollution has been reported for present-day fish (Reid et al., 2016).
Assuming that the Hangenberg event was caused by a supernova (Fields et al., 2020), the supernova would have emitted ionizing gamma and UV rays for 100,000 years. In addition to the ocean anoxia present during this period, gamma radiation may have been important in causing evolution to accelerate. Ionizing gamma rays damage DNA and increase the mutation rate, resulting in epigenetic changes that enhance variation between offspring, which could significantly accelerate the transition to land-based tetrapods. Accelerated mutation rates were observed in birds and mammals exposed to ionizing radiation from the Chernobyl accident (Møller and Mousseau, 2015). Therefore, the combined effects of various extreme environmental conditions during this period resulted in major differences in species before and after significant events in the fossil record.
Table 2 presents the results of the probability calculations for the events (discussed in this section) that are considered important for the transition to land-dwelling tetrapods. The timeframe used for the calculations is 15 million years, considering that the transition to tetrapods is thought to have occurred over approximately 15–20 million years (Wood and Nakamura, 2018). For simplicity, the Kellwasser and Hangenberg periods are considered together as a single Late Devonian extinction event. The 30% ozone depletion during the Hangenberg period (Fields et al., 2020) requires the presence of a supernova ca. 212 light years from Earth (Gehrels et al., 2003). Our galaxy experiences one to three supernovas every century (Adams et al., 2013). However, at a distance of 33 light-years from the Earth, 0.05–10 supernova occur every billion years (Gehrels et al., 2003). In Table 2, this rate has been scaled in terms of spherical volume to obtain the λ for a supernova 212 light-years from Earth. The passage of the solar system through the spiral arms of the galaxy increases the likelihood of supernovas. The Hangenberg event occurred during this period (Fields et al., 2020), and the actual probability of a supernova at this time may have been even higher than the probability provided here.
The evolution of fish (specifically, of the tetrapod ancestors), their survival during mass extinction events, and their ability to develop tetrapod traits were considered when estimating the probability of tetrapod evolution. The NHPP cannot be used for this purpose, because these are not discrete events. The Kellwasser event resulted in the loss of 20–25% of marine species, while ~50% of lobed fish survived as they were better adapted to breathing anoxic water (McGhee, 2012). Subsequently, the Hangenberg event eradicated approximately 20% of genera (Marshall et al., 2020), with the adaptability of lobed fish and their newly evolved strong fins aiding their survival. The appearance of tetrapodomorphs (such as Tiktaalik) and tetrapods (such as Acanthostega) in the fossil record suggests a high survival rate of their ancestors. Therefore, the probability of surviving these extinction events (estimated using Eq. 8) and of the development of tetrapod traits, is likely to be low (estimated at P = 0.1), owing to the specificity of the ecological and genetic factors and the fact that only a few known lineages among the surviving sarcopterygians (e.g., tetrapodomorphs) exhibit these traits.
2.4. Theia Collision
The favored model for the formation of the Moon describes a collision between a protoplanet, Theia, and proto-Earth during the last stage of planetary formation. The Theia collision is considered the most transformative event in the Earth’s history, enabling it to develop into a habitable world with life rather than an average rocky planet (Meier et al., 2014). The Earth’s stabilized tilt and spin, and the Moon’s tidal forces are direct results of the collision. The Theia collision model is used because it is the only model that properly explains why the Moon orbits at an unusual angle of 5° (Ćuk et al., 2016). This model proposes that, before the collision, Earth’s obliquity and Moon’s orbit angle were 75°. As the Moon moved further from the Earth, the Sun’s gravity pulled the Moon’s orbit angle down to 0° (relative to the solar system disk), which in turn dragged the Earth’s obliquity with it (approaching but never reaching 0°, because the Moon moved too far away). There is significant evidence that the Theia collision deposited necessary volatiles such as carbon, nitrogen, and water onto the Earth (Budde et al., 2019; Grewal et al., 2019; Li et al., 2016).
Although overwhelming evidence indicates that the Theia collision was critical for ensuring Earth’s habitability, there is as yet no way to determine which specific properties of the collision and its effects were the most important in determining the rate of evolution on Earth. For example, if the Earth’s tilt were 1° higher, would this collision have caused the evolution of life on Earth to slow by more than 30%? Rather than estimating the probability of each factor contributing to the evolutionary rate, it was conservatively assumed that the Theia collision was more likely than the dinosaur extinction event. The probability estimate for Theia collision was assumed to be 1 × 10−12, based on the following factors:
- Theia’s impact angle and location must have resulted in ideal obliquity.
- The impact velocity of Theia did not destabilize Earth’s orbit and resulted in an ideal day length (spin). Excessively high velocity would have resulted in increased tectonic activity.
- The relative sizes of Theia and proto-Earth.
- Theia’s elemental composition was volatile and water-rich.
- Earth’s spin before the collision.
- The Moon’s mass and orbit angles. A smaller Moon would not have produced stable obliquity, and a larger Moon would have produced obliquity closer to 0°.
- Stable plate tectonics (the hypothesis that the Theia collision resulted in plate tectonics is discussed later)
2.5. Solar System
Considering the 30% rule and the principle of mediocrity, it is possible to obtain a reasonable estimate of the aspects of the solar system necessary for our existence. Red dwarf stars are more numerous and last longer than more massive stars such as the Sun (a yellow dwarf).
The Kroupa initial mass function (Kroupa, 2001) (describing the distribution of stellar masses at the formation of the Milky Way) was used to estimate the probability of a randomly selected star having approximately the mass of our sun (0.95–1.05 solar masses), giving a probability of 0.1. It has been estimated that 10–20% of stars in our galaxy are within the galactic habitable zone (GHZ) (Lineweaver et al., 2004). The Earth’s position (8 kpc from the galactic center) is considered ideal for the development of life, as metallicity is sufficient to form rocky planets and supernova rates and radiation are low enough to avoid frequent sterilization. The habitable zone is estimated to be at 4–10 kpc from the galactic center (Gonzalez et al., 2001; Lineweaver et al., 2004), placing Earth within this zone. Considering the fraction of stars within the GHZ compared to the total number in the star-forming disk (2–13 kpc from the center) and assuming that 15% of stars have Earth-like planets (Petigura et al., 2013), the probability of a star such as the sun being located in the GHZ is approximately 0.2.
It has been proposed that the presence of a large number of planets in our solar system may have been necessary for the Theia collision to occur. Sun-like stars typically have <1 planets (Burke et al., 2015). Considering a Poisson distribution for the number of planets per star (Zhu, 2019), the probability of 8 planets around a sun-like star is 1×10−5 (assuming a mean of 1 planet per star). Furthermore, the asteroid belt between Jupiter and Mars is an important factor. Maintaining an asteroid belt at an appropriate location for dinosaur extinction is critical. Jupiter is extremely massive and close to Earth, thus protecting the inner planets from the outer solar system.

Table 3.
Rates and probabilities of the characteristics of the solar system thought to be necessary for life.
Table 3.
Rates and probabilities of the characteristics of the solar system thought to be necessary for life.
| Characteristic | Generalized characteristic | P | Data source |
| Sun’s mass | Star with a solar mass of ~1 | 0.1 | (Kroupa, 2001) |
| Earth in the GHZ | Earth-like planet located in the GHZ | 0.2 | (Lineweaver et al., 2004; Petigura et al., 2013) |
| 8 planets in our solar system | G-type star with 8 planets | 1×10−5 | (Zhu, 2019) |
| Jupiter/Mars/asteroid belt formation | 1×10−3 | ||
| Total probability | 2×10−10 |
3. Results and Discussion
The total probability of the dinosaur extinction, the end-Devonian tetrapod transition, the Theia collision, and the solar system conditions was calculated based on their respective probabilities, as follows: [6.5×10−15] × [3.6×10−4] × [1×10−12] × [2×10−10], resulting in a probability of (4.68×10−40). However, because many unknown factors also contribute to the rate of evolution on Earth, civilization is likely orders of magnitude less common than this estimate.
3.1. Other Possible Filters
This section presents other potential filters that may have contributed to the rate of evolution of life on Earth and helped to provide the conditions necessary for the improbable development of civilization.
- The End-Permian extinction was likely necessary for the later evolution of dinosaurs and mammals.
- The End-Ediacaran extinction 541 Mya may have been necessary for the evolution of vertebrates (this extinction event significantly reduced the diversity of existing invertebrates).
- Snowball Earth, which began at 720 Mya and ended at 635 Mya, may have been necessary for the first widespread distribution of multicellular life.
- The first entry of mitochondria into cells may have been the result of an uncommon event (e.g., an event leading to the evolution of eukaryotes).
- The Late Heavy Bombardment may have deposited the volatile elements necessary for rapid evolution or altered the plate tectonics beneficially.
- Protein folding may be probabilistic. Is it possible that the development of all life on Earth was enabled by the favorable structure of the first proteins? Is the complex folding of proteins a natural result of angiogenesis?
- Although Nuclear War might be a typical event for civilizations, it has been mitigated on Earth to date. Geographically, the fact that the Earth has several continents, rather than a single Pangea-like continent, may have promoted democratic systems, making nuclear war less likely.
3.2. Effect of the Theia Collision on Plate Tectonics
After a rocky planet finishes accretion, its crust hardens into a solid plate that is similar to an eggshell. This is the normal state of the crust of a rocky planet such as Venus, Mars, or Mercury; in contrast, Earth is the only planet known to have plate tectonics. Although the origin of plate tectonics remains unclear, recent studies have provided the earliest evidence of subduction (a critical process in plate tectonics) 4.3 billion years ago, very soon after the Theia impact (Canup and Asphaug, 2001; Yuan et al., 2024). The core of Theia was absorbed by the proto-Earth, increasing the core-mantle boundary temperature and providing heat-producing elements, thereby providing the conditions necessary to drive the first subduction events (Yuan et al., 2024). The Theia impact provided sufficient energy to remelt the Earth’s upper crust, resulting in the violent mixing of molten elements. Crust hardening occurred during the mixing of elements from proto-Earth and Theia, resulting in a heterogeneous crust with subduction zones and divergent boundaries. In addition, the moon, formed from the material ejected by the impact, is thought to play a role in the Earth’s tectonic system. The moon was formed from an accretion disk of debris from the collision, and its primary orbit was originally orders of magnitude closer to Earth than it currently is. The gravitational pull of the moon on the softened Earth caused the Earth to become highly oblate, resulting in variation in crustal thickness and density during the gradual despinning process; such variation is critical for maintaining tectonic activity (Lock and Asimow, 2020). In addition, the tidal heating of the young Earth by the close-orbiting moon was a major source of energy for the development of magma oceans (Zahnle et al., 2007).
3.3. Role of Plate Tectonics in Abiogenesis
Abiogenesis, the development of life from inert materials, requires chemical energy and appropriate environmental conditions. Before the evolution of photosynthesis, the necessary chemical energy may have originated from plate tectonics. When elements are subducted into the mantle, they are exposed to heat and pressure and eventually resurface near divergent boundaries or hydrothermal vents, thus providing a constant source of chemical energy. The various mixtures of chemical species ejected into the oceans by plate tectonics would have enabled a wide range of chemical reactions, which may have produced the necessary precursor molecules for abiogenesis. When the incremental process of increasing the chemical and molecular complexity became sufficient to produce self-replicating systems, the first true life forms evolved. Constant recycling and abundant chemical energy are ideal conditions for abiogenesis, and it is hypothesized that, apart from tectonic activity, there were no other mechanisms on early Earth that could have provided the appropriate conditions for abiogenesis. Volcanoes can deliver chemicals to the surface, and weather can create complex chemistry; however, these isolated events do not provide the long-term consistent chemical conditions necessary to achieve sufficient complexity.
The famous experiments performed by Miller and Urey in the early 1950s replicated the “primordial soup” conditions thought at the time to be present on early Earth (Miller, 1953; Miller and Urey, 1959). Starting with water and common gases, they applied an electric arc to simulate lightning and demonstrated the first synthesis of organic molecules from inorganic materials. Subsequent experiments have shown that all 20 amino acids required for life can be produced in vitro under simulated early-Earth conditions (Criado-Reyes et al., 2021). Ocean hydrothermal vents caused by plate tectonics are prime candidates for the origin of first life because they provide thermal and chemical energy and a wide range of elements and molecules. The constant replenishment of chemicals via hydrothermal vents (due to plate tectonics) supports persistent chemical reactions. Additionally, some of the earliest fossils originate from vent environments (Dodd et al., 2017). Recently, it has been demonstrated that vesicle formation is favored in hydrothermal vent environments with high temperature, salinity, and alkalinity. Furthermore, weather conditions such as lightning can produce the molecules necessary for membrane formation (Hess et al., 2021). Further experiments are required to examine the mechanisms whereby simple proto-cells develop into complex cells in hydrothermal environments, where the necessary precursor materials are constantly replenished and react with nearby molecules.
3.4. Role of Quantum Fluctuations in the Big Bang
To understand this hypothesis, the quantum uncertainty principle can be applied to infinite space–time (visualizing space–time as a large wavefunction/probability curve where everything is happening simultaneously). In this model, anything that is physically possible has a 100% probability of eventually occurring and hypothetically occurring simultaneously within infinite space-time (Carroll, 2019). Therefore, it is necessary to prove that a series of quantum fluctuations can physically cause the Big Bang. Quantum fluctuations are purely random, which indicates that there is no theoretical limit to the number of quantum fluctuations that can occur simultaneously. Therefore, if numerous quantum fluctuations were annihilated at the same time and place, the energy would temporarily create a cosmic singularity and a large band of finite size. The existence of a highly improbable civilization in an extremely rare solar system implies that the Universe is extraordinarily large, which is likely necessary to provide sufficient variation and randomness to enable the development of intelligent life. The entire universe is therefore likely to be larger than the observable universe.
One common counterargument to the hypothesis that quantum fluctuations caused the Big Bang is that our physical models are not compatible with the state of the Universe before this event, in which time and space (as we understand them) may not have existed. Assuming that the Big Bang occurred at least once, it is reasonable to assume that this was neither the first nor the last such event (based on a many-world interpretation). For example, assuming that another universe existed before the current one, it is hypothesized that all matter in that universe will eventually decay, leaving only space-time, as there is no known mechanism by which space decays. It would then be reasonable to assume that the multiverse mostly comprises one endless pit of infinite spacetime with some larger-than-average exploding dust specks, as in our universe.
3.5. Study Limitations
The statistical analysis of highly improbable events is notoriously complex, particularly with a sample size of one (i.e., the Earth) (Ward et al., 2000). Despite our lack of knowledge and the difficulties in estimating probabilities on a universal scale, researchers have proposed various methods to calculate the probability that we are alone in the universe. An important assumption in the Fermi Paradox is that alien life and civilization could evolve in an enormous number of locations. Therefore, for any nonzero probability of evolution, some intelligent life would have emerged, and it is reasonable to expect that we will encounter evidence of this. In the Drake equation (Drake et al., 2015), the number of detectable/contactable civilizations in the Milky Way is expressed as the product of the rate of star formation per year, the fraction of stars with planets, the number of habitable planets in systems with planets, the fraction of such planets with life, the fraction of life that develops intelligence, the fraction of intelligent civilizations that are detectable/contactable, and the average lifetime of detectable civilizations. It has been shown that the estimates yielded by the Drake equation vary by eight orders of magnitude, depending on the values selected by scientists (Sandberg et al., 2018). To improve the scientific rigor of estimating the probabilities of extraterrestrial life, a recent study incorporating uncertainty ranges provided a strong statistical argument that it is highly probable that we are alone in the universe (Sandberg et al., 2018). This conclusion was also reached in a study using Monte Carlo simulations of galaxies, stars, life, and interactions between civilizations to verify the Rare Earth Hypothesis (Forgan and Rice, 2010). Therefore, although it remains difficult to verify the accuracy of the Great Filter event probabilities predicted here, the general conclusion of this study supporting the Rare Earth Hypothesis is consistent with the findings of other studies.
Author Contributions
Jobe Soffa Clarke: Conceptualization, data curation, formal analysis, investigation, methodology.
Data Availability Statement
No new data were generated or analyzed in support of this research.
Acknowledgments
Editorial support, in the form of writing, collating author comments, copyediting, fact checking, and referencing, was provided by Editage, Cactus Communications.
Conflicts of Interest
None.
References
- Adams, S.M.; Kochanek, C.S.; Beacom, J.F.; Vagins, M.R.; Stanek, K.Z. Observing the next galactic supernova. Astrophysical Journal 2013, 778(2), 164. [Google Scholar] [CrossRef]
- Aldous, D.J. The great filter, branching histories and unlikely events. Mathematical Scientist 2010, 37, 55–64. [Google Scholar]
- Bostrom, N. Superintelligence: Paths, Dangers, Strategies, Superintelligence: Paths, Dangers, Strategies; Oxford University Press; New York, NY, US, 2014. [Google Scholar]
- Budde, G.; Burkhardt, C.; Kleine, T. Molybdenum isotopic evidence for the late accretion of outer Solar System material to Earth. Nature Astronomy 2019, 3(8), 736–741. [Google Scholar] [CrossRef]
- Burke, C.J.; Christiansen, J.L.; Mullally, F.; Seader, S.; Huber, D.; Rowe, J.F.; Coughlin, J.L.; Thompson, S.E.; Catanzarite, J.; Clarke, B.D.; Morton, T.D.; Caldwell, D.A.; Bryson, S.T.; Haas, M.R.; Batalha, N.M.; Jenkins, J.M.; Tenenbaum, P.; Twicken, J.D.; Li, J.; Quintana, E.; Barclay, T.; Henze, C.E.; Borucki, W.J.; Howell, S.B.; Still, M. Terrestrial planet occurrence rates for the Kepler GK dwarf sample. Astrophysical Journal 2015, 809(1), 8. [Google Scholar] [CrossRef]
- Canup, R.M.; Asphaug, E. Origin of the Moon in a giant impact near the end of the Earth’s formation. Nature 2001, 412(6848), 708–712. [Google Scholar] [CrossRef]
- Carmichael, S.K.; Waters, J.A.; Königshof, P.; Suttner, T.J.; Kido, E. Paleogeography and paleoenvironments of the Late Devonian Kellwasser event: a review of its sedimentological and geochemical expression. Global & Planetary Change 2019, 183, 102984. [Google Scholar]
- Carmichael, S.K.; Waters, J.A.; Suttner, T.J.; Kido, E.; DeReuil, A.A. A new model for the Kellwasser Anoxia Events (Late Devonian): shallow water anoxia in an open oceanic setting in the Central Asian Orogenic Belt. Palaeogeography, Palaeoclimatology, Palaeoecology 2014, 399, 394–403. [Google Scholar] [CrossRef]
- Carroll, S. Something Deeply Hidden: Quantum Worlds and the Emergence of Spacetime; Penguin Publishing Group, 2019. [Google Scholar]
- Cassan, A.; Kubas, D.; Beaulieu, J.-P.; Dominik, M.; Horne, K.; Greenhill, J.; Wambsganss, J.; Menzies, J.; Williams, A.; Jørgensen, U.G.; Udalski, A.; Bennett, D.P.; Albrow, M.D.; Batista, V.; Brillant, S.; Caldwell, J. a. R.; Cole, A.; Coutures, C.; Cook, K.H.; Dieters, S.; Prester, D.D.; Donatowicz, J.; Fouqué, P.; Hill, K.; Kains, N.; Kane, S.; Marquette, J.-B.; Martin, R.; Pollard, K.R.; Sahu, K.C.; Vinter, C.; Warren, D.; Watson, B.; Zub, M.; Sumi, T.; Szymański, M.K.; Kubiak, M.; Poleski, R.; Soszynski, I.; Ulaczyk, K.; Pietrzyński, G.; Wyrzykowski, Ł. One or more bound planets per Milky Way star from microlensing observations. Nature 2012, 481, 167–169. [Google Scholar] [CrossRef]
- Chapelle, K.E.J.; Griffin, C.T.; Pol, D. Growing with dinosaurs: a review of dinosaur reproduction and ontogeny. Biology Letters 2025, 21(1), 20240474. [Google Scholar] [CrossRef]
- Chapman, C.R.; Morrison, D. Impacts on the Earth by asteroids and comets: assessing the hazard. Nature 1994, 367(6458), 33–40. [Google Scholar] [CrossRef]
- Chatterjee, S.; Necip, G.; Yoshinobu, A.; Donofrio, R. Shiva Structure: a Possible KT Boundary Impact Crater on the Western Shelf of India; Museum of Texas Tech University. Special Publications, 2006; Vol. 50. [Google Scholar]
- Clack, J.A. An early tetrapod from ‘Romer’s Gap. Nature 2002, 418(6893), 72–76. [Google Scholar] [CrossRef]
- Collins, G.S.; Patel, N.; Davison, T.M.; Rae, A.S.P.; Morgan, J.V.; Gulick, S.P.S. IODP-ICDP Expedition 364 Science Party, Third-Party Scientists, 2020. A steeply-inclined trajectory for the Chicxulub impact. Nature Communications 11(1), 1480. [CrossRef]
- Criado-Reyes, J.; Bizzarri, B.M.; García-Ruiz, J.M.; Saladino, R.; Di Mauro, E. The role of borosilicate glass in Miller–Urey experiment. Scientific Reports 2021, 11(1), 21009. [Google Scholar] [CrossRef]
- Ćuk, M.; Hamilton, D.P.; Lock, S.J.; Stewart, S.T. Tidal evolution of the Moon from a high-obliquity, high-angular-momentum earth. Nature 2016, 539(7629), 402–406. [Google Scholar] [CrossRef] [PubMed]
- Daeschler, E.B.; Shubin, N.H.; Jenkins, F.A. A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 2006, 440(7085), 757–763. [Google Scholar] [CrossRef]
- Dodd, M.S.; Papineau, D.; Grenne, T.; Slack, J.F.; Rittner, M.; Pirajno, F.; O’Neil, J.; Little, C.T.S. Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nature 2017, 543(7643), 60–64. [Google Scholar] [CrossRef]
- dos Reis, M.; Thawornwattana, Y.; Angelis, K.; Telford, M.J.; Donoghue, P.C.J.; Yang, Z. Uncertainty in the timing of origin of animals and the limits of precision in molecular timescales. Current Biology: CB 2015, 25(22), 2939–2950. [Google Scholar] [CrossRef] [PubMed]
- Drake, F.; Vacoch, D.A.; Dowd, M.F. The Drake Equation: Estimating the Prevalence of Extraterrestrial Life through the Ages; Cambridge University Press, 2015. [Google Scholar]
- Ernst, R.E.; Youbi, N. How Large Igneous Provinces affect global climate, sometimes cause mass extinctions, and represent natural markers in the geological record. Palaeogeography, Palaeoclimatology, Palaeoecology 2017, 478, 30–52. [Google Scholar] [CrossRef]
- Fields, B.D.; Melott, A.L.; Ellis, J.; Ertel, A.F.; Fry, B.J.; Lieberman, B.S.; Liu, Z.; Miller, J.A.; Thomas, B.C. Supernova triggers for end-Devonian extinctions. Proceedings of the National Academy of Sciences of the United States of America 2020, 117(35), 21008–21010. [Google Scholar] [CrossRef] [PubMed]
- Forgan, D.H.; Rice, K. Numerical testing of the rare earth hypothesis using Monte Carlo realization techniques. International Journal of Astrobiology 2010, 9(2), 73–80. [Google Scholar] [CrossRef]
- Gehrels, N.; Laird, C.M.; Jackman, C.H.; Cannizzo, J.K.; Mattson, B.J.; Chen, W. Ozone depletion from nearby supernovae. Astrophysical Journal 2003, 585(2), 1169–1176. [Google Scholar] [CrossRef]
- Gertsch, B.; Keller, G.; Adatte, T.; Garg, R.; Prasad, V.; Berner, Z.; Fleitmann, D. Environmental effects of Deccan volcanism across the Cretaceous–Tertiary transition in Meghalaya, India. Earth & Planetary Science Letters 2011, 310(3–4), 272–285. [Google Scholar]
- Gonzalez, G.; Brownlee, D.; Ward, P. The galactic habitable zone: galactic chemical evolution. Icarus 2001, 152(1), 185–200. [Google Scholar] [CrossRef]
- Grewal, D.S.; Dasgupta, R.; Sun, C.; Tsuno, K.; Costin, G. Delivery of carbon, nitrogen, and sulfur to the silicate earth by a giant impact. Science Advances 2019, 5(1), eaau3669. [Google Scholar] [CrossRef]
- Hanson, R. The great filter, are we almost past it? 1998. Available online: https://mason.gmu.edu/~rhanson/greatfilter.html (accessed on 6.3.25).
- Hess, B.L.; Piazolo, S.; Harvey, J. Lightning strikes as a major facilitator of prebiotic phosphorus reduction on early Earth. Nature Communications 2021, 12(1), 1535. [Google Scholar] [CrossRef] [PubMed]
- Horner, J.R. Dinosaur reproduction and parenting. Annual Reviews. Annual Review of Earth & Planetary Sciences 2000, 28(1), 19–45. [Google Scholar]
- Kaltenegger, L.; Faherty, J.K. Past, present and future stars that can see Earth as a transiting exoplanet. Nature 2021, 594(7864), 505–507. [Google Scholar] [CrossRef] [PubMed]
- Kroupa, P. On the variation of the initial mass function. Monthly Notices of the Royal Astronomical Society 2001, 322(2), 231–246. [Google Scholar] [CrossRef]
- Kukla, A. Extraterrestrials: A Philosophical Perspective; Lexington Books, 2009; p. 20. [Google Scholar]
- Li, Y.; Dasgupta, R.; Tsuno, K.; Monteleone, B.; Shimizu, N. Carbon and sulfur budget of the silicate Earth explained by accretion of differentiated planetary embryos. Nature Geoscience 2016, 9(10), 781–785. [Google Scholar] [CrossRef]
- Lineweaver, C.H.; Fenner, Y.; Gibson, B.K. The galactic habitable zone and the age distribution of complex life in the milky way. Science 2004, 303(5654), 59–62. [Google Scholar] [CrossRef]
- Lock, S.J.; Asimow, P.D. A tectonically active early Earth driven by the tidal recession of the Moon. Presented at the 51st Lunar and Planetary Science Conference, 2020; p, p. 1848. [Google Scholar]
- Lyu, Y.J.; Zhou, M.-F.; Cui, Y.; Hu, R.-Z.; Liu, Z.R.; Li, X. Decoding the end-Devonian Hangenberg oceanic anoxia event: insights from ironstones in South China. Earth & Planetary Science Letters 2025, 653, 119203. [Google Scholar]
- Marov, M.Y. The structure of the universe, in: The Fundamentals of Modern Astrophysics; Springer: New York, New York, NY, 2015; pp. 279–294. [Google Scholar]
- Marshall, J.E.A.; Lakin, J.; Troth, I.; Wallace-Johnson, S.M. UV-B radiation was the Devonian-Carboniferous boundary terrestrial extinction kill mechanism. Science Advances 2020, 6(22), eaba0768. [Google Scholar] [CrossRef]
- McGhee, G.R. Extinction: Late Devonian mass extinction. In Encyclopedia of Life Sciences; John Wiley & Sons, 2012. [Google Scholar]
- Meier, M.M.M.; Reufer, A.; Wieler, R. On the origin and composition of Theia: constraints from new models of the Giant Impact. Icarus 2014, 242, 316–328. [Google Scholar] [CrossRef]
- Miller, S.L. A production of amino acids under possible primitive earth conditions. Science 1953, 117(3046), 528–529. [Google Scholar] [CrossRef]
- Miller, S.L.; Urey, H.C. Organic compound synthesis on the primitive earth. Science 1959, 130(3370), 245–251. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.P.; Mousseau, T.A. Strong effects of ionizing radiation from Chernobyl on mutation rates. Scientific Reports 2015, 5, 8363. [Google Scholar] [CrossRef] [PubMed]
- Olsen, P.E.; Kent, D.V.; Sues, H.D.; Koeberl, C.; Huber, H.; Montanari, A.; Rainforth, E.C.; Fowell, S.J.; Szajna, M.J.; Hartline, B.W. Ascent of dinosaurs linked to an iridium anomaly at the Triassic-Jurassic boundary. Science 2002, 296(5571), 1305–1307. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, K.; Reinhard, C.T. The future lifespan of Earth’s oxygenated atmosphere. Nature Geoscience 2021, 14(3), 138–142. [Google Scholar] [CrossRef]
- Pande, K. Age and duration of the Deccan Traps, India: a review of radiometric and paleomagnetic constraints. Proceedings of the Indian Academy of Science Earth & Planetary Sciences 2002, 111(2), 115–123. [Google Scholar]
- Petigura, E.A.; Howard, A.W.; Marcy, G.W. Prevalence of Earth-size planets orbiting Sun-like stars. Proceedings of the National Academy of Sciences of the United States of America 2013, 110(48), 19273–19278. [Google Scholar] [CrossRef]
- Racki, G. A volcanic scenario for the Frasnian–Famennian major biotic crisis and other Late Devonian global changes: more answers than questions? Global & Planetary Change 2020, 189, 103174. [Google Scholar]
- Raup, D.M.; Sepkoski, J.J. Mass extinctions in the marine fossil record. Science 1982, 215(4539), 1501–1503. [Google Scholar] [CrossRef]
- Reid, N.M.; Proestou, D.A.; Clark, B.W.; Warren, W.C.; Colbourne, J.K.; Shaw, J.R.; Karchner, S.I.; Hahn, M.E.; Nacci, D.; Oleksiak, M.F.; Crawford, D.L.; Whitehead, A. The genomic landscape of rapid repeated evolutionary adaptation to toxic pollution in wild fish. Science 2016, 354(6317), 1305–1308. [Google Scholar] [CrossRef]
- Royle, N.J.; Alonzo, S.H.; Moore, A.J. Co-evolution, conflict and complexity: what have we learned about the evolution of parental care behaviours? Current Opinion in Behavioral Sciences 2016, 12, 30–36. [Google Scholar] [CrossRef]
- Sahney, S.; Benton, M.J. Recovery from the most profound mass extinction of all time. Proceedings. Biological Sciences 2008, 275(1636), 759–765. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.; Tafforeau, P.; Clack, J.A.; Ahlberg, P.E. Life history of the stem tetrapod Acanthostega revealed by synchrotron microtomography. Nature 2016, 537(7620), 408–411. [Google Scholar] [CrossRef] [PubMed]
- Sandberg, A.; Drexler, E.; Ord, T. Dissolving the Fermi Paradox. 2018. Available online: https://arxiv.org/abs/1806.02404.
- Schulte, P.; Alegret, L.; Arenillas, I.; Arz, J.A.; Barton, P.J.; Bown, P.R.; Bralower, T.J.; Christeson, G.L.; Claeys, P.; Cockell, C.S.; Collins, G.S.; Deutsch, A.; Goldin, T.J.; Goto, K.; Grajales-Nishimura, J.M.; Grieve, R.A.F.; Gulick, S.P.S.; Johnson, K.R.; Kiessling, W.; Koeberl, C.; Kring, D.A.; MacLeod, K.G.; Matsui, T.; Melosh, J.; Montanari, A.; Morgan, J.V.; Neal, C.R.; Nichols, D.J.; Norris, R.D.; Pierazzo, E.; Ravizza, G.; Rebolledo-Vieyra, M.; Reimold, W.U.; Robin, E.; Salge, T.; Speijer, R.P.; Sweet, A.R.; Urrutia-Fucugauchi, J.; Vajda, V.; Whalen, M.T.; Willumsen, P.S. The Chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary. Science 2010a, 327(5970), 1214–1218. [Google Scholar] [CrossRef]
- Shubin, N.; Daeschler, E.; Jenkins, F. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature 2006, 440, 764–771. [Google Scholar] [CrossRef]
- Verendel, V.; Häggström, O. Fermi’s Paradox, Extraterrestrial Life and the Future of Humanity: a Bayesian Analysis; Cambridge University Press, 2015. [Google Scholar]
- Wandel, A. The Fermi paradox revisited: Technosignatures and the contact era. Astrophysical Journal 2022, 941(2), 184. [Google Scholar] [CrossRef]
- Ward, P.D.; Brownlee, D. Rare Earth: Why Complex Life Is Uncommon in the Universe; Springer New York; New York, NY, 2000. [Google Scholar]
- Ward, P.D.; Brownlee, D.; Krauss, L. Rare earth: why complex life is uncommon in the universe. Physics Today 2000, 53(9), 62–63. [Google Scholar] [CrossRef]
- Werner, J.; Griebeler, E.M. Reproductive biology and its impact on body size: comparative analysis of mammalian, avian and dinosaurian reproduction. PLOS One 2011, 6(12), e28442. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.W.P.; Nakamura, T. Problems in fish-to-tetrapod transition: genetic expeditions into old specimens. Frontiers in Cell & Developmental Biology 2018, 6, 70. [Google Scholar]
- Yuan, Q.; Gurnis, M.; Asimow, P.D.; Li, Y. A giant impact origin for the first subduction on earth. Geophysical Research Letters 2024, 51(9), GL106723, e2023. [Google Scholar] [CrossRef]
- Zahnle, K.; Arndt, N.; Cockell, C.; Halliday, A.; Nisbet, E.; Selsis, F.; Sleep, N.H. Emergence of a habitable planet. Space Science Reviews 2007, 129(1–3), 35–78. [Google Scholar] [CrossRef]
- Zhu, W. Influence of stellar metallicity on occurrence rates of planets and planetary systems. Astrophysical Journal 2019, 873(1), 8. [Google Scholar] [CrossRef]
Figure 1.
Schematic explaining the proposed 30% rule.

Figure 2.
Geological evidence for events occurring during dinosaur extinction. Reproduced from Chatterjee et al. (2006) with the permission of the Museum of Texas Technical University.
Figure 2.
Geological evidence for events occurring during dinosaur extinction. Reproduced from Chatterjee et al. (2006) with the permission of the Museum of Texas Technical University.
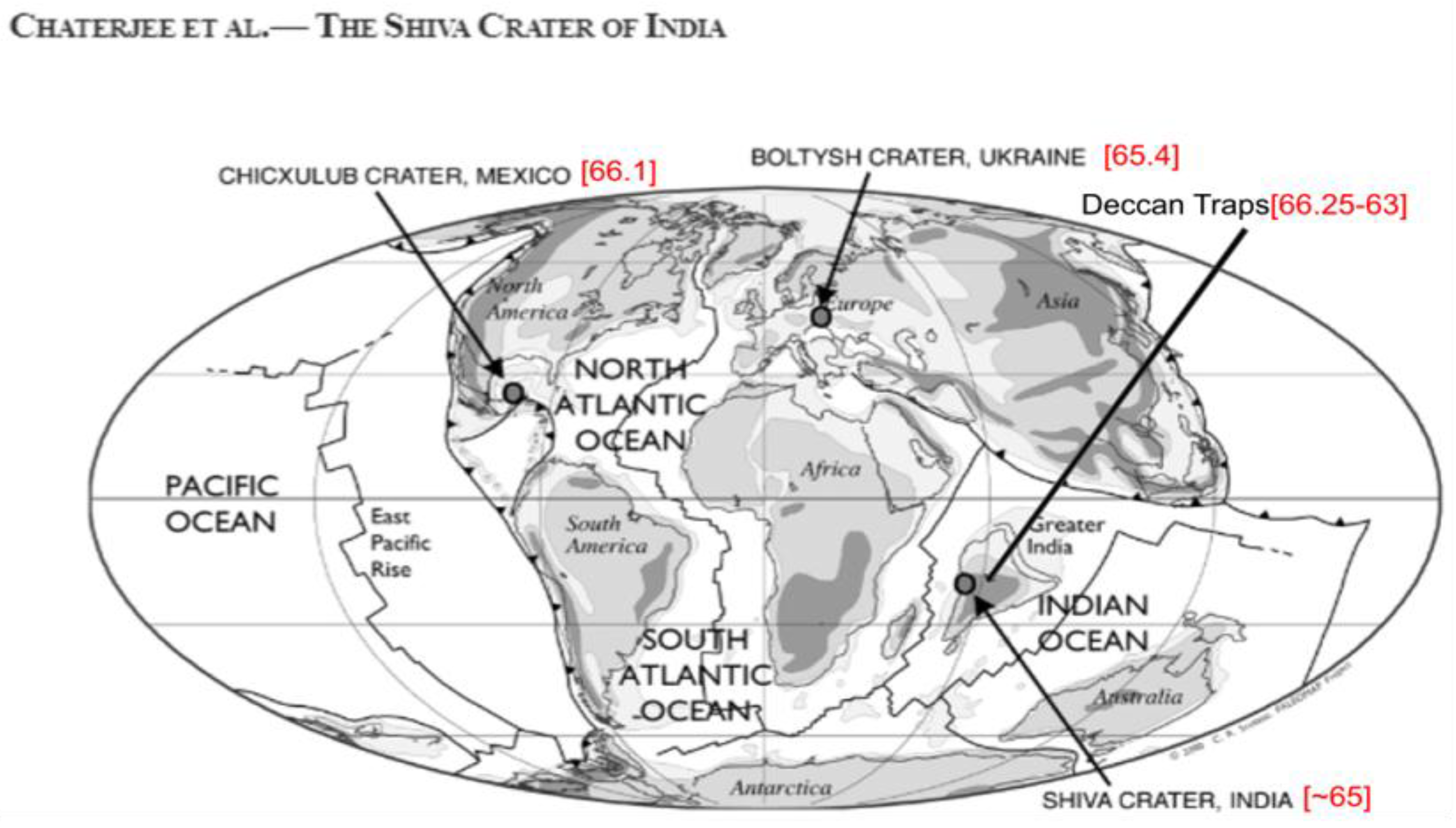
Figure 3.
Tetrapod transition before and after the Kellwasser event. Reproduced from (Shubin et al. 2006) with permission from N. Shubin.
Figure 3.
Tetrapod transition before and after the Kellwasser event. Reproduced from (Shubin et al. 2006) with permission from N. Shubin.
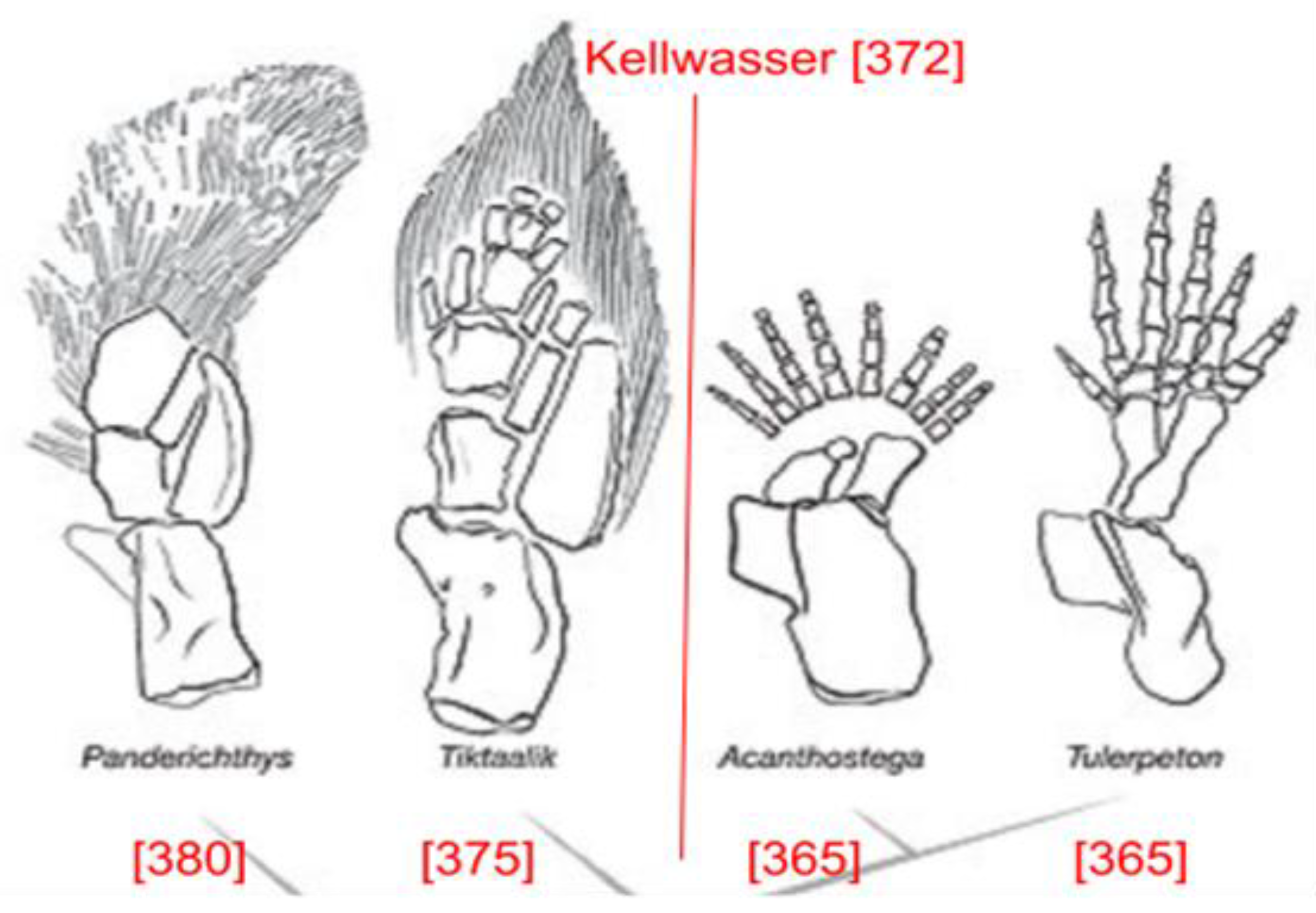
Figure 4.
Illustration showing that before the Hangenberg event, all members of this lineage were primarily aquatic, while after this event, the first non-aquatic tetrapods appeared in the fossil record. Reproduced from Clack (2002) with the permission of Nature.
Figure 4.
Illustration showing that before the Hangenberg event, all members of this lineage were primarily aquatic, while after this event, the first non-aquatic tetrapods appeared in the fossil record. Reproduced from Clack (2002) with the permission of Nature.
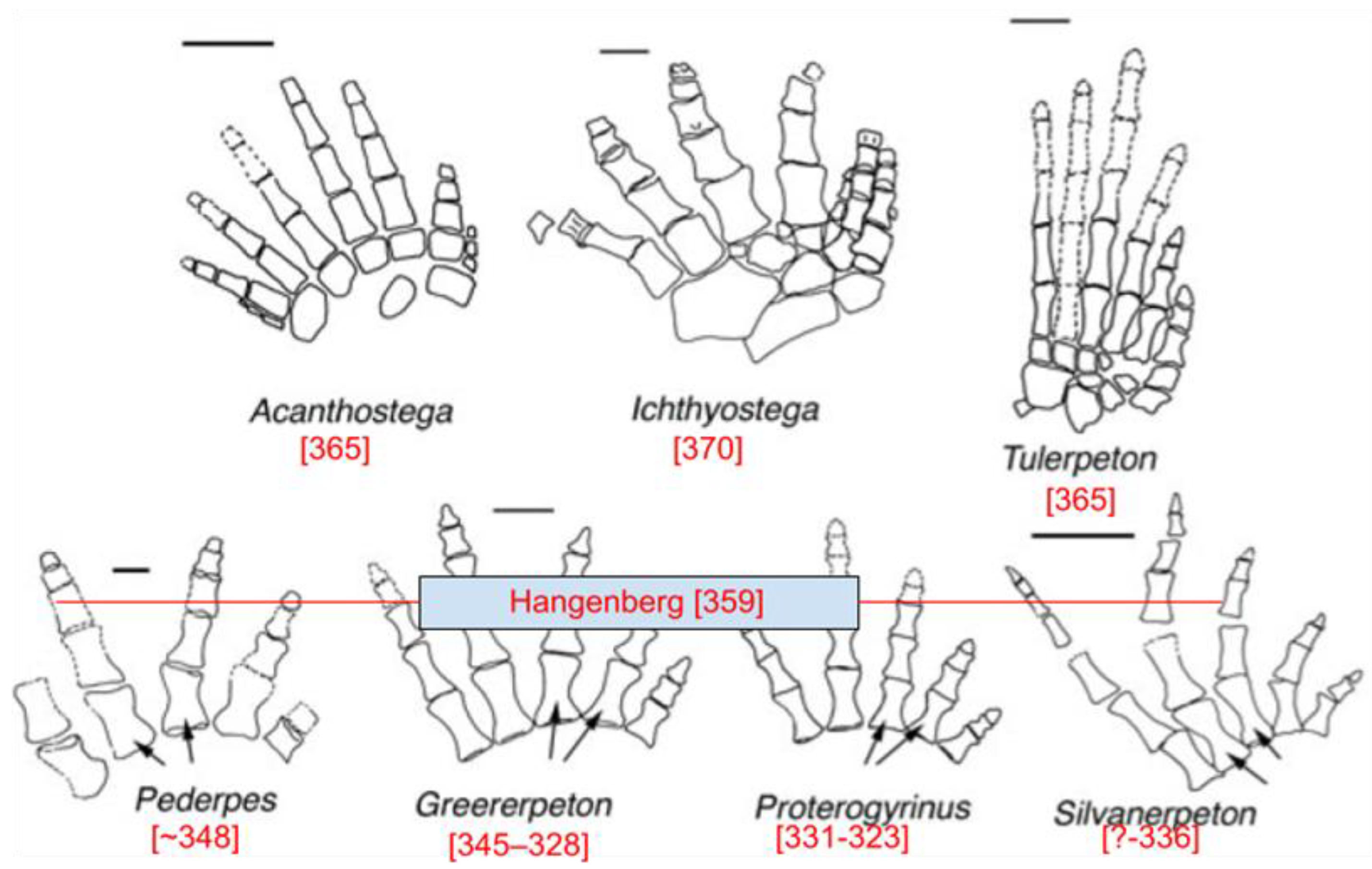
Table 1.
Rates and probabilities of the occurrence of the rare events contributing to dinosaur extinction.
Table 1.
Rates and probabilities of the occurrence of the rare events contributing to dinosaur extinction.
| Event | Generalized event | λ (yr−1) | P (yr−2MY) | Data source | Notes |
| Deccan Traps eruption | Large igneous province (LIP) events | 5×10−9 | 1×10−2 | (Ernst and Youbi, 2017) | 15 major events in 3.5 billion years |
| Chicxulub meteor hits the Yucatán sulfur deposit at a 60° angle | Impact of >10 km diameter asteroid | 2×10−9 | 4×10−3 | (Chapman and Morrison, 1994) | |
| Surface area ratio of the sulfur field to Earth | 2.5×10−4 | 2.5×10−4 | (Schulte et al., 2010b) | 200-km estimated radius of sulfur field | |
| Shiva meteor impacts India | 40-km wide meteor | 5×10−10 | 1×10−3 | (Chapman and Morrison, 1994) | Data for >10 km impactors scaled using a power law |
| Surface area ratio of India to Earth | 6.5×10−3 | 6.5×10−3 | India’s area is 0.65% of Earth | ||
| Dinosaur dominance and mammals exist | 0.1 | (dos Reis et al., 2015; Olsen et al., 2002; Sahney and Benton, 2008) | |||
| Total probability | 6.5×10−15 |
Table 2.
Rates and probabilities of the occurrence of the rare events contributing to tetrapod transition.
Table 2.
Rates and probabilities of the occurrence of the rare events contributing to tetrapod transition.
| Event | Generalized event | λ (yr−1) | P (yr−15MY) | Data source |
| Late-Devonian extinction | Extinction event with ≥ 20% species loss | 2×10−8 | 0.3 | (Raup and Sepkoski, 1982) |
| 30% ozone depletion | Supernova exploding 212 light years from Earth | 1×10−9 | 1.5×10−2 | (Fields et al., 2020; Gehrels et al., 2003) |
| Tetrapod ancestors survived long enough to enable the transition | Fish survived extinction events | 0.8 | (McGhee, 2012) | |
| Tetrapod traits evolved | 0.1 | (Wood and Nakamura, 2018) | ||
| Total probability | 3.6×10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



