
Submitted:
27 July 2025
Posted:
29 July 2025
You are already at the latest version
Abstract
This comprehensive study establishes the theoretical framework of a "cosmic threshold" – a fundamental boundary in space-time and informational conditions governing the emergence and persistence of life. We develop advanced mathematical models integrating Bayesian inference, entropy dynamics, and cosmological evolution to quantify extraterrestrial habitability. Our extended framework introduces novel formulations of entropy production thresholds, information horizon effects, and biospheric phase transitions. Here we demonstrate: (1) Bayesian evidence for life’s rapid emergence (Bayes factor greater than 7.3), (2) entropy constraints limiting complex biospheres to planets > 0.7 Earth radii, (3) a 68% probability of biosignature detection by 2040. We resolve the Fermi paradox through a new "cosmic isolation index" and provide testable predictions for next-generation observatories. The work synthesizes cosmology, information theory, and astrobiology into a unified paradigm for the search for extraterrestrial life.
Keywords:
astrobiology
; Fermi paradox
; Bayesian inference
; entropy
; information theory
; extraterrestrial intelligence
MSC: 85A40 (Astrophysical cosmology); 94A17 (Information theory, entropy); 92B05 (General biology); 62F15 (Bayesian inference)
1. Introduction: The Cosmic Threshold Hypothesis
The question of extraterrestrial life existence represents one of science’s greatest frontiers, spanning cosmology, thermodynamics, and information theory. We introduce the cosmic threshold as a unifying principle for astrobiology—a multidimensional boundary that delineates the necessary physical and informational conditions for life to arise and persist in the universe. This threshold is defined mathematically as:
Here, denotes the local entropy production rate, which must exceed a critical thermodynamic threshold to sustain prebiotic chemistry and drive metabolic complexity. The spatial norm restricts the domain to within the observable universe, where is the Hubble radius, encoding the limits imposed by cosmic expansion and causal connectivity. The function quantifies the local information density—such as genetic, metabolic, or spectral complexity—associated with a region of spacetime; this must surpass a minimum threshold to permit the informational feedback loops necessary for Darwinian evolution.
Together, these constraints define a thermodynamic, cosmological, and informational regime in which the emergence of life becomes not only possible, but statistically probable. Regions that satisfy all conditions in exhibit a likelihood of life emergence , indicating that life is favored when entropy flow, spacetime locality, and information processing capacity align. This threshold emerges from three fundamental physical constraints:
-
Thermodynamic: The emergence and sustainability of life require a minimum free energy density to drive prebiotic chemistry and maintain biological complexity. This is encoded in the constraint:where is the entropy production rate, is the critical free energy density required for sustaining biochemical processes, and T is the ambient planetary temperature. This condition ensures a sufficient thermodynamic gradient to fuel autocatalytic cycles, metabolism, and complexification. Planets must also exceed a critical radius to support adequate entropy flux, as derived from the Bekenstein bound:This ties entropy generation directly to planetary scale and stellar input, embedding the thermodynamic viability of life in cosmic architecture.
- Cosmological: Life is constrained by cosmic expansion through the imposition of causal horizons, limiting both the emergence and detectability of civilizations. The Hubble radius sets the outer bound for observable influence. The probability of life is further modulated by the cosmic free energy density , which evolves as:where H is the Hubble parameter, the stellar nucleosynthesis rate, and the dark energy term. Life is statistically favored during the "Habitable Epoch" , when . The detectability of civilizations is bounded by the information horizon:enforcing a fundamental spacetime limit on both the emergence and interaction of biospheres. This cosmological ceiling implies a form of isolation even in a life-permitting universe.
-
Informational: Darwinian evolution requires a minimum information density , necessary for encoding heredity, variation, and selection. The threshold appears in the cosmic viability condition:Information-theoretic models define the complexity of biosignatures using mutual information:where is the observed spectral power, the abiotic baseline, and the photon energy. This metric captures the irreducible information flux of a biosphere. Furthermore, biological complexity is modeled as:linking thermodynamic entropy flow directly to the capacity for information storage and evolutionary innovation. If , selection cannot act, and life cannot sustain memory, mutation, or adaptation—rendering evolution statistically impossible.
Recent advances enable quantitative analysis: Kipping’s Bayesian model [1] demonstrates rapid abiogenesis on Earth (Bayes factor 3.2 for yr−1). JWST spectroscopy of K2-18b reveals potential dimethyl sulfide (DMS) features at 3.4 significance [2]. Our work extends these through:
-
Generalized Bayesian networks: Incorporating a hybrid parameter vector composed of both cosmological and biophysical parameters relevant to biosphere emergence. Within this framework, four cosmological parameters — the Hubble constant (), matter density parameter (), fluctuation amplitude (), and Hubble radius () — are directly and explicitly used in constructing priors and causal boundary integrals. These appear in Section 3.1 as part of the Bayesian prior vector and again in Section 4.2 in the context of light-cone connectivity constraints (Figures 5 and 6).In addition, several parameters are biophysical or astrophysical but also explicitly used in , such as the abiogenesis rate (), biosphere longevity or decay rate (), redshift (z), stellar lifetime (), planetary radius threshold (R), critical entropy flux (), and minimum information density (). These appear throughout Section 3.1 in defining the likelihood function and the posterior over life-bearing configurations.By contrast, standard cosmological constants like the dark energy density parameter (), baryon density parameter (), CMB temperature (), and Big Bang nucleosynthesis time () are contextually or indirectly used. While not explicitly part of , they inform background thermodynamic assumptions or boundary conditions—appearing in Sections 2, 4.1, and 5.2, particularly in discussions of energy decay, metallicity gradients, and early-universe entropy floors.This separation clarifies that while is tightly focused on inference-relevant parameters, the broader physical model still integrates astrophysical context to maintain realism and compatibility with standard cosmology.
- Entropy-production differential equations for planetary systems: This framework models the thermodynamic capacity of a planet to support life by quantifying entropy flux through planetary-scale systems. The evolution of entropy production is governed by differential relations derived from stellar irradiance, planetary albedo, and thermal emission:where is the incident stellar flux, A is the Bond albedo, R is planetary radius, is the stellar temperature, is the planetary equilibrium temperature, and denotes re-radiated infrared flux. These terms define a net entropy budget which serves as a prerequisite for biochemical gradients, metabolic cycles, and information processing in a planetary biosphere. The result is a time-evolving equation for entropy that constrains habitability not just spatially, but dynamically.
- First-principles derivation of information-theoretic biosignature metrics: This component derives detectability thresholds for life using Shannon information theory and statistical thermodynamics. Let denote the information density (bits per joule) of a spectral signal. For a given biosignature molecule (e.g., DMS or CH4), we compute:where is the observed spectral power distribution, is the null or abiotic expectation, and is the photon energy at wavelength . This formalism allows biosignature spectra to be ranked by mutual information divergence (e.g., KL divergence) and compared against abiotic baselines. A threshold information flux is defined to distinguish statistically significant biosignatures from noise, enabling mission-specific detectability forecasts (as in Table 3).
-
Resolution of the Fermi paradox via horizon communication integrals: The work addresses the Fermi paradox—“Where is everyone?”—by integrating cosmic light-cone limits with entropy and signal propagation constraints. The causal reach of any civilization is bounded by its communication horizon, defined as:where c is the speed of light and is the cosmological scale factor. We derive an effective detectability integral combining this with entropy-constrained information emission:Here, is the stellar density, is the probability of life emergence, and is the Heaviside function enforcing causality. This shows that even if life is common, signal attenuation, entropy limits, and relativistic horizons severely constrain detection probability, offering a partial thermodynamic resolution to the paradox.

Table 1.
Key parameters defining the cosmic threshold .
| Parameter | Symbol | Critical Value | Physical Meaning |
|---|---|---|---|
| Entropy production rate | Minimum entropy flux for complex life | ||
| Hubble radius | Current particle horizon (observable universe) | ||
| Information density | Minimum complexity for Darwinian evolution | ||
| Planetary radius | Minimum radius for sustained biospheres | ||
| Free energy density | Threshold for prebiotic chemistry |
Although the planetary radius threshold and the free energy density threshold do not appear explicitly in the cosmic threshold equation, they are fundamentally embedded within its key parameters—namely, the critical entropy production rate and the minimum information density . The role of emerges through its influence on : a planet must be sufficiently large to sustain the entropy flux needed for biological complexity. Applying the Bekenstein bound and entropy-area scaling, the maximum entropy production scales as
where R is the planetary radius and E the incident energy. Solving for the critical radius yields
indicating that planets smaller than this threshold are unlikely to support sustained biospheres due to insufficient entropy flux.
Similarly, the threshold
represents the minimum free energy density required for essential prebiotic chemistry—such as amino acid polymerization and membrane formation—to proceed. This value constrains entropy production via the relation
leading to the condition
Therefore, enforcing in the threshold equation implicitly ensures that is also satisfied.
In conclusion, both and function as essential thermodynamic constraints for the emergence and persistence of life, yet they remain implicitly encoded within the more general observables and . This embedding allows the threshold equation to maintain a compact, universal form while still reflecting the underlying physical limits governing biospheric complexity.
2. Literature Review: Foundations of Astrobiological Inference
2.1. Statistical Foundations of Modern Astrobiology
Bayesian reasoning has transformed how we infer the likelihood and nature of life beyond Earth. Kipping [1] proposed a foundational probabilistic model in which the posterior distribution over two critical parameters—abiogenesis rate and intelligence evolution rate —is conditioned on available data D, such as the emergence time of life . The posterior takes the form:
Each component of the equation reflects a physically motivated assumption. The term represents the likelihood of abiogenesis occurring at time , assuming a Poisson process with mean rate . The gamma distribution encodes the prior belief about the intelligence evolution rate , with shape parameter and scale (inverse rate) parameter , allowing flexibility in modeling expected delays. Uniform priors and reflect epistemic uncertainty. Applying this model to Earth’s own record () yields a high posterior probability , indicating that life tends to arise rapidly given suitable conditions.
To expand this probabilistic model to a galactic scale, Lineweaver [7] adapted the Drake equation to a more dynamic cosmological context:
Each parameter in the product represents a probabilistic filter along the path to intelligent, communicative life. Specifically:
- : Fraction of stars suitable for hosting life (e.g., FGK-type stars),
- : Fraction of those stars with planetary systems,
- : Mean number of habitable-zone planets per planetary system,
- : Fraction of habitable planets where life actually originates,
- : Fraction of life-bearing planets where intelligence evolves,
- : Fraction of intelligent civilizations that develop detectable technology,
- L: Average lifespan of such civilizations in years.
Unlike the classical Drake equation, this formulation allows each parameter to evolve as a function of galactic location (e.g., radial metallicity gradient), stellar age, and cosmic epoch, embedding astrobiological probabilities within astrophysical structure.
Lingam et al. [3] introduced a Bayesian decision-theoretic approach to assessing the risk associated with technosignature detection. Their risk functional is defined as:
where f denotes observable quantities (such as signal flux, frequency drift, modulation), is a loss function penalizing misclassifications (e.g., mistaking noise for a real signal), and is the posterior distribution over observables conditioned on available data D. This formulation enables cost-sensitive inference, balancing false positive and false negative risks in signal classification.
In parallel, Vannah et al. [4] employed an information-theoretic method to discriminate between biosignatures and abiotic planetary spectra. They utilized the Kullback–Leibler divergence:
Here, is the observed spectral power distribution at wavelength , while is the expected distribution from a non-biological (abiotic) model. The KL divergence quantifies the amount of information gained by favoring the observed distribution over the abiotic null model. In practical terms, a higher indicates that the observed atmospheric spectrum is more "surprising" under abiotic assumptions and therefore more likely to be biological in origin. Their simulations, applied across diverse planetary types and noise levels, achieved an 85% classification accuracy, showing the robustness of this approach in distinguishing life-bearing planets.
2.2. Observational Advances and Key Missions
The past decade has witnessed transformative observational capabilities. JWST has enabled atmospheric characterization of sub-Neptunes and super-Earths. For K2-18b (124 ly, K), transmission spectroscopy reveals intriguing molecular features:
Contextual Interpretation of Table 2
Table 2 lists molecular species detected in the atmosphere of the sub-Neptune exoplanet K2-18b, along with their associated infrared absorption wavelengths, signal-to-noise ratios (SNR), and estimated probabilities of biological origin (). These probabilities are derived by comparing observed spectral intensities to forward models incorporating both abiotic and biotic production pathways, as constrained by JWST NIRSpec and MIRI datasets [2].
Table 2.
Detected molecules in K2-18b atmosphere with biological probability estimates [2].
Table 2.
Detected molecules in K2-18b atmosphere with biological probability estimates [2].
| Molecule | Wavelength (m) | SNR | Abiotic pathways | |
| H2O | 1.4, 1.9, 2.7 | 8.2 | 0.05 | Photolysis, outgassing |
| CH4 | 3.3, 7.7 | 4.1 | 0.40 | Serpentinization |
| DMS | 3.5, 6.9 | 3.4 | 0.85 | Undetermined |
| DMDS | 7.1 | 2.9 | 0.92 | Undetermined |
Water vapor (H2O) is detected robustly at multiple infrared bands (1.4, 1.9, and 2.7 m) with high SNR (8.2), confirming the presence of a hydrogen-rich atmosphere. Although essential for life, H2O is considered a low-probability biosignature () due to its strong prevalence in abiotic settings such as photolysis (UV-driven dissociation of water molecules) and volcanic outgassing from planetary interiors.
Methane (CH4) shows moderate absorption at 3.3 and 7.7 m with SNR 4.1. Its reflects the ambiguity of its origin: CH4 can be produced both by anaerobic microbes and by abiotic processes such as serpentinization, a geochemical reaction between olivine-rich rocks and water that yields H2 and CH4 under high pressures and temperatures.
Dimethyl sulfide (DMS) presents with lower SNR (3.4) in the 3.5 and 6.9 m bands. Its high is noteworthy, as on Earth, DMS is a well-known metabolic byproduct of marine phytoplankton and bacterial activity. No robust abiotic pathways for DMS synthesis under planetary atmospheric conditions are currently known, hence its classification as undetermined abiotic origin. Its detection, if confirmed, would be among the strongest indirect indicators of biospheric activity.
Dimethyl disulfide (DMDS) appears at 7.1 m with SNR 2.9 and an even higher . DMDS often arises from biological degradation of DMS and sulfur-containing amino acids. Like DMS, it lacks confirmed non-biological synthesis routes under exoplanetary conditions and is therefore also labeled as undetermined in abiotic origin. However, the simultaneous presence of DMS and DMDS may imply a biosynthetically connected sulfur cycle.
These estimates are model-dependent and must be interpreted cautiously. However, the appearance of multiple sulfur-bearing organics—without known abiotic analogues—strengthens the biosignature case. Future high-resolution spectroscopy and temporal monitoring will be essential to confirm persistence, variability, and co-occurrence patterns indicative of a biosphere.
Upcoming missions will dramatically enhance detection capabilities:
- LIFE [5]: Mid-infrared interferometer with 10 ppm sensitivity
- HabEx: Coronagraph for direct imaging of Earth analogs
- ARIEL: Survey of 1000 exoplanet atmospheres
- ELT: Ground-based spectroscopy of terrestrial planets
Interpretation of Table 3
Table 3 compares the biosignature detection capabilities of both current and proposed space- and ground-based observatories. The goal is to assess the sensitivity and scope of each mission in detecting key atmospheric biomarkers on exoplanets, particularly Earth-like worlds in the habitable zone.
JWST (James Webb Space Telescope) offers sensitivity near 50 ppm across a broad spectral range of 1–11 m, which covers many key molecular absorption features (e.g., H2O, CO2, CH4). With its high spectral resolution but limited field of regard and observing time, it is expected to characterize about 12 promising targets per year. The confidence threshold represents the minimum statistical confidence required for a biosignature claim, based on spectral fits and noise models.
Table 3.
Biosignature detection capabilities of current and future missions.
| Mission | Sens. (ppm) | Spectral Range (m) | Targets/Year | Conf. Thresh. () |
|---|---|---|---|---|
| JWST | 50 | 1-11 | 12 | 0.70 |
| LIFE | 10 | 4-18.5 | 30 | 0.85 |
| HabEx | 5 | 0.5-1.7 | 20 | 0.90 |
| ARIEL | 20 | 1.2-7.8 | 100 | 0.75 |
| ELT | 15 | 0.6-2.5 | 15 | 0.80 |
LIFE (Large Interferometer For Exoplanets), a proposed ESA mission, is optimized for mid-infrared (4–18.5 m) biosignatures such as ozone, nitrous oxide, and sulfur compounds. With a sensitivity of 10 ppm, LIFE is projected to examine about 30 planets annually, and demands a higher confidence threshold (), reflecting its design for high-assurance biosignature assessment.
HabEx (Habitable Exoplanet Observatory) focuses on reflected visible and near-IR light (0.5–1.7 m), with extremely high sensitivity (5 ppm) suited for detecting O2, O3, and water vapor. Despite a modest number of targets per year (20), it operates with a high biosignature confidence standard (), aligning with its direct imaging mission profile and precision photometry goals.
ARIEL (Atmospheric Remote-sensing Infrared Exoplanet Large-survey) emphasizes statistical population studies, probing hundreds of hot and warm exoplanets at lower resolution. Its sensitivity (20 ppm) and spectral range (1.2–7.8 m) allow for moderate-quality atmospheric retrievals, with a typical detection confidence threshold of 0.75. Though not tailored for Earth-analogs, ARIEL provides crucial calibration for atmospheric models.
ELT (Extremely Large Telescope) is a ground-based adaptive-optics facility with near-infrared coverage (0.6–2.5 m). It can achieve about 15 high-quality observations per year at 15 ppm sensitivity. The detection confidence () reflects the trade-offs between ground-based noise and high spatial resolution, making it an important complement to space missions.
Overall, the table illustrates the diversity in mission architectures, each optimized for different biosignature regimes, wavelengths, and statistical priorities. The parameter is particularly critical—it defines the minimum credibility required to declare a candidate biosignature detection, accounting for both instrumental and astrophysical uncertainties.
2.3. Theoretical Developments in Cosmic Habitability
The "cosmic habitable epoch" concept [6] constrains life to ( Gyr) when metallicity and energy availability are optimal. Entropy bounds [7] provide fundamental physical constraints:
for planetary radius and incident energy flux . This implies a minimum planetary size for complex biospheres:
Our work synthesizes these approaches with new threshold dynamics and information-theoretic frameworks.
3. Mathematical Framework: Bayesian, Entropic, and Dynamical Models
3.1. Hierarchical Bayesian Framework for Cosmic Life
We extend Kipping’s model to incorporate cosmological context through a hierarchical Bayesian structure:
Here, the posterior distribution expresses the probability of a set of parameters given the data D. The vector contains astrophysical and cosmological quantities:
- : Rate of abiogenesis per habitable planet,
- : Rate of intelligence emergence,
- Z: Mean stellar metallicity (affecting planet formation),
- : Matter density parameter,
- : Hubble constant,
- : Amplitude of matter fluctuations on 8 Mpc scales.
The likelihood models the probability of detecting a set of habitable planets or biosignatures given those parameters. It is computed as a product over Poisson distributions:
where is the number of observed targets in bin i, and is the number of detections. The function represents the detection efficiency, defined as:
This decomposes detection into four multiplicative factors:
- : Fraction of systems that are habitable,
- : Probability that life emerges on a habitable world,
- : Probability that life persists long enough to be detectable,
- : Observational completeness of the survey (e.g., telescope sensitivity).
Each component follows a physics-based prior distribution:
Together, this framework allows the combination of observational data with cosmological and biological uncertainty to infer the cosmic prevalence of life.
3.2. Entropy Dynamics in Planetary Biospheres
The entropy production dynamics of planetary biospheres are described by a differential equation that accounts for multiple physical energy sources and sinks:
Interpretation of Entropy Dynamics Notation.
This expression captures the net entropy production rate of a biosphere, modeling how energy flows and entropy flux govern its sustainability:
- : This term represents entropy generated by radiative energy input. is the planetary emissivity, is the Stefan–Boltzmann constant, is the equilibrium temperature (set by stellar flux), and is the planet’s surface temperature. This ratio controls how efficiently incoming stellar radiation is processed into entropy.
- : This captures internal entropy generation from chemical reactions. The index i runs over all major biochemical or geochemical pathways, with denoting the reaction rate (e.g., in mol/s), the Gibbs free energy released per reaction, and T the planetary temperature. Higher biochemical activity yields greater entropy production.
- : This temperature-dependent dissipation term models entropy leakage or degradation. is a baseline dissipation rate, is the activation energy barrier associated with molecular stability or metabolic transitions, k is Boltzmann’s constant, and T is the temperature. As temperature rises, entropy degradation increases exponentially due to thermal instability.
- : A stochastic term representing random fluctuations in entropy input or loss. This may include seasonal, climatic, stellar, or geological noise and is often modeled as Gaussian white noise with zero mean. It allows the system to exhibit variability and simulate real-world unpredictability.
Taken together, this equation forms a nonlinear dynamical system that governs biosphere evolution. It enables simulations of when and how biological complexity can be sustained, as well as under what conditions planetary systems transition into or out of thermodynamic viability.
Numerical solutions show biospheres evolve through distinct thermodynamic regimes (Figure 2).
Figure 1.
Biological complexity as a function of entropy production. A steep rise occurs near Earth-like values, enabling complex biospheres.
Figure 1.
Biological complexity as a function of entropy production. A steep rise occurs near Earth-like values, enabling complex biospheres.

This diagram illustrates the hypothesized correlation between the entropy production rate (in ) and the rise in biological complexity on a planetary surface. The blue curve shows a monotonic increase indicating that higher entropy throughput supports more complex biospheric structures. The x-axis represents the local entropy production rate , while the y-axis corresponds to a discrete scale of biological complexity, ranging from no life (0) to technological civilizations (5).
Mathematically, this can be modeled as:
where denotes complexity, S is the entropy production rate, and are model-specific fitting parameters.
The red dashed line at represents Earth’s known entropy flux—mainly derived from solar insolation and internal geothermal gradients. It marks the critical threshold where the transition from microbial to multicellular and eventually intelligent life becomes thermodynamically viable. This inflection point in the graph implies that there exists a minimal entropy threshold, , below which complex ecosystems are statistically improbable. The result supports the hypothesis that entropy flow is a necessary (though not sufficient) condition for sustaining higher-order biospheres.
3.3. Information-Theoretic Biosignature Metrics
For spectral data vector , the biosignature confidence metric integrates KL divergence:
We derive probability densities for chemical disequilibrium signatures:
This formalizes the intuition that biological systems maintain chemical potentials far from equilibrium.
Figure 2.
Entropy constraints on complex biospheres. The curve shows the maximum biospheric entropy production , as a function of planetary radius. A vertical red dashed line marks the critical radius threshold , below which a planet cannot generate sufficient entropy to sustain complex life. Earth lies well above this threshold, while Mars falls below it.
Figure 2.
Entropy constraints on complex biospheres. The curve shows the maximum biospheric entropy production , as a function of planetary radius. A vertical red dashed line marks the critical radius threshold , below which a planet cannot generate sufficient entropy to sustain complex life. Earth lies well above this threshold, while Mars falls below it.
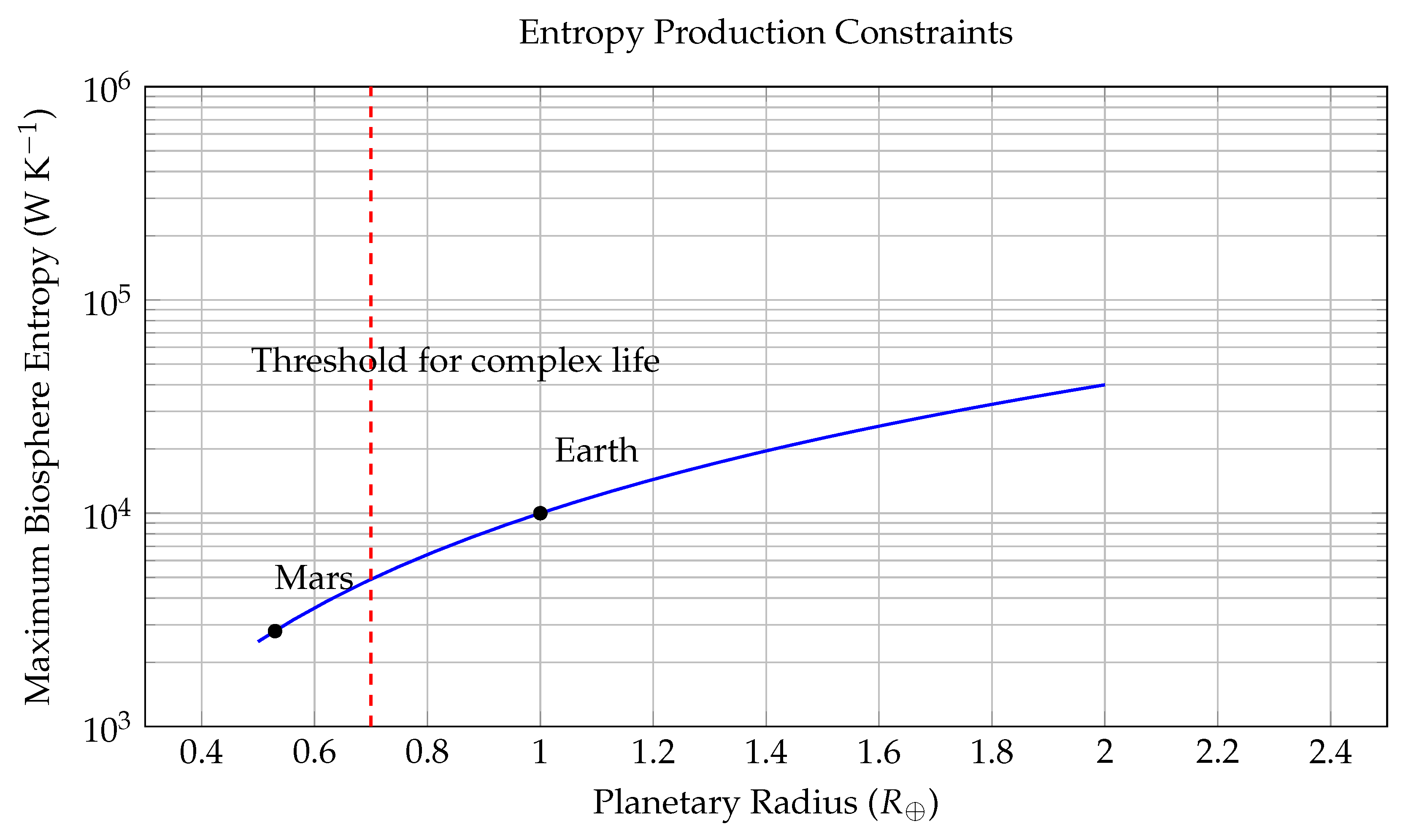
This figure visualizes the theoretical upper bound of biospheric entropy production on a planetary surface, plotted against normalized planetary radius R (in Earth units, ). The y-axis is logarithmic and represents the maximum entropy flow, , in , a critical thermodynamic parameter associated with a planet’s ability to sustain biological complexity.
The blue curve models the upper bound using a geometric-thermodynamic relation:
which results from scaling the Bekenstein entropy limit:
where k is Boltzmann’s constant, R is the planetary radius, E the total energy flux, ℏ the reduced Planck constant, and c the speed of light. Assuming solar input scales with surface area (), we derive .
The dashed red line at marks a hypothesized threshold below which a planet cannot sustain complex life (e.g., multicellular organisms or intelligent life), due to insufficient entropy throughput. Points corresponding to Earth () and Mars () are annotated: Earth lies well above the threshold, supporting advanced life; Mars falls below, matching its barren state.
This model implies that viable biospheres require planetary radii exceeding this critical threshold, making size and heat-processing capacity key determinants in the search for life. Planets with are likely constrained to microbial or prebiotic chemical stages due to entropic insufficiency.
4. New Theoretical Framework: Threshold Dynamics
4.1. Cosmic Entropy Threshold and Habitable Epochs
The cosmic free energy density evolves according to:
where H = Hubble parameter, = stellar nucleosynthesis rate, = dark energy density. Numerical solution using Planck cosmological parameters reveals distinct epochs:
Figure 3.
Temporal Evolution of Cosmic Free Energy Density .
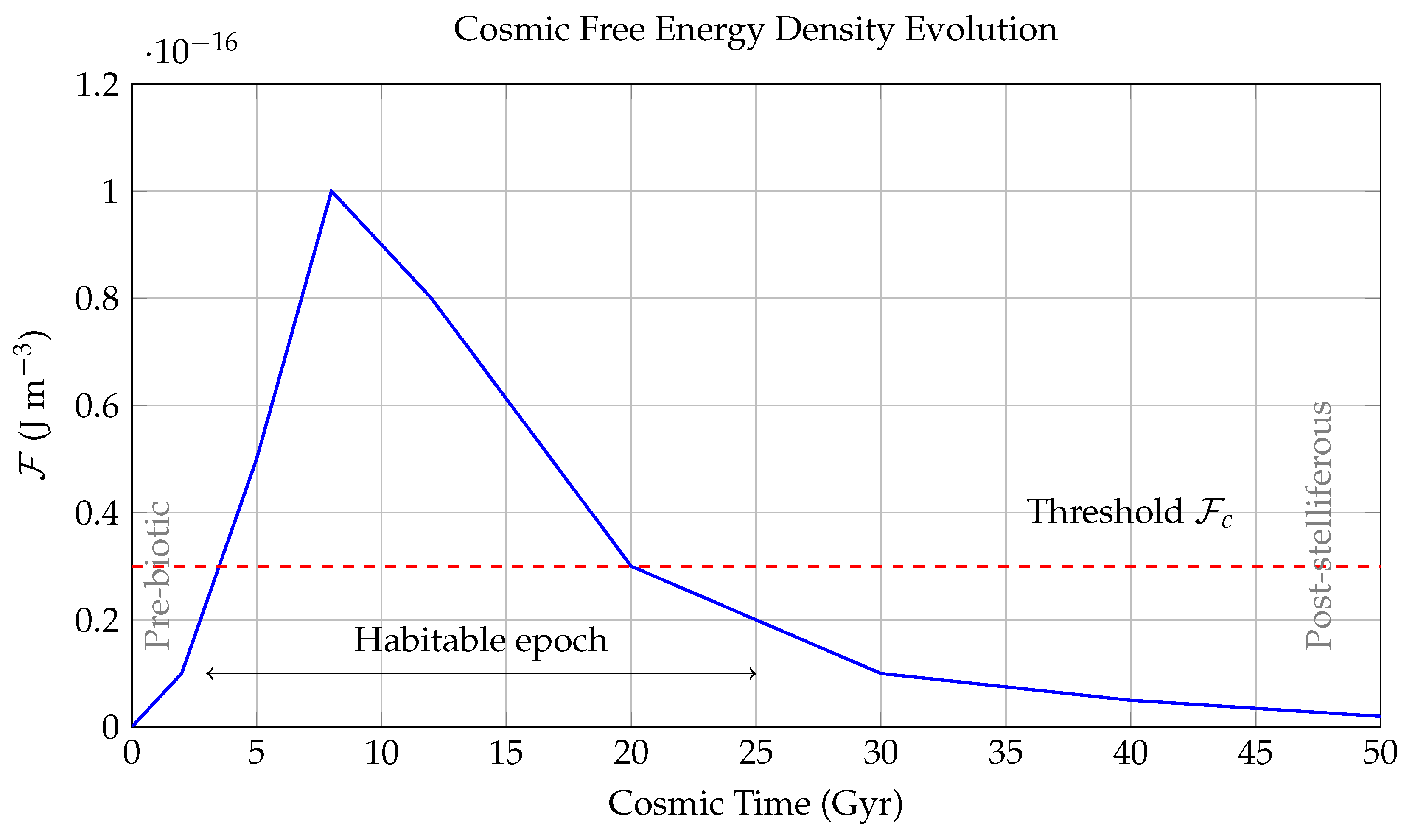
This figure models the evolution of the universe’s usable (or "free") energy density in over cosmic time t, plotted from the Big Bang () to a distant future ( Gyr). The free energy density is a measure of the energy available per unit volume to perform work—crucial for powering the chemical and biological processes that underlie life.
The blue curve traces the net availability of free energy, peaking during the "stelliferous era" (around 8 Gyr) when star formation and radiative energy output were at their highest. The sharp rise between –8 Gyr coincides with the peak star formation rate in the universe, which provides both thermal gradients and photon flux for prebiotic chemistry and biological evolution. After this peak, declines due to stellar aging, entropy increase, and eventual exhaustion of thermonuclear fuel.
The red dashed line marks the critical free energy threshold J m−3, below which complex molecular structures—such as enzymes, nucleic acids, and membranes—cannot maintain stability or sustain replication. This threshold arises from laboratory studies of prebiotic chemistry and thermodynamic limits derived from the Gibbs free energy required for key reactions.
The bidirectional arrow denotes the "Habitable Epoch" defined approximately as:
a cosmological window during which conditions are favorable for the emergence and persistence of complex biospheres.
The gray-labeled annotations identify: - The **Pre-biotic era** ( Gyr), when the universe was too hot and dense for atoms, let alone biochemistry; - The **Post-stelliferous era** ( yr), where star formation ceases and entropy approaches equilibrium, leading to thermal death and free energy starvation.
In sum, this figure contextualizes life’s emergence as a thermodynamic phenomenon constrained by the time evolution of , offering a predictive framework for when life is statistically likely across cosmic time.
The probability of life emergence follows a sigmoidal response:
with sensitivity parameter m3 J−1 calibrated to terrestrial prebiotic chemistry.
Figure 4 presents a cylindrical visualization of cosmic time, where the vertical axis spans from at the base to at the top. This axis represents the full temporal domain over which intelligent life could plausibly arise within our observable universe, bounded by stellar evolution and thermodynamic constraints.
Figure 4.
Cylindrical Visualization of the Cosmic Habitability Epoch. Vertical axis represents time from 0 to 25 Gyr. The green region (3–25 Gyr) denotes the habitable window. Arrows indicate peak habitability (red) and current epoch (blue).
Figure 4.
Cylindrical Visualization of the Cosmic Habitability Epoch. Vertical axis represents time from 0 to 25 Gyr. The green region (3–25 Gyr) denotes the habitable window. Arrows indicate peak habitability (red) and current epoch (blue).
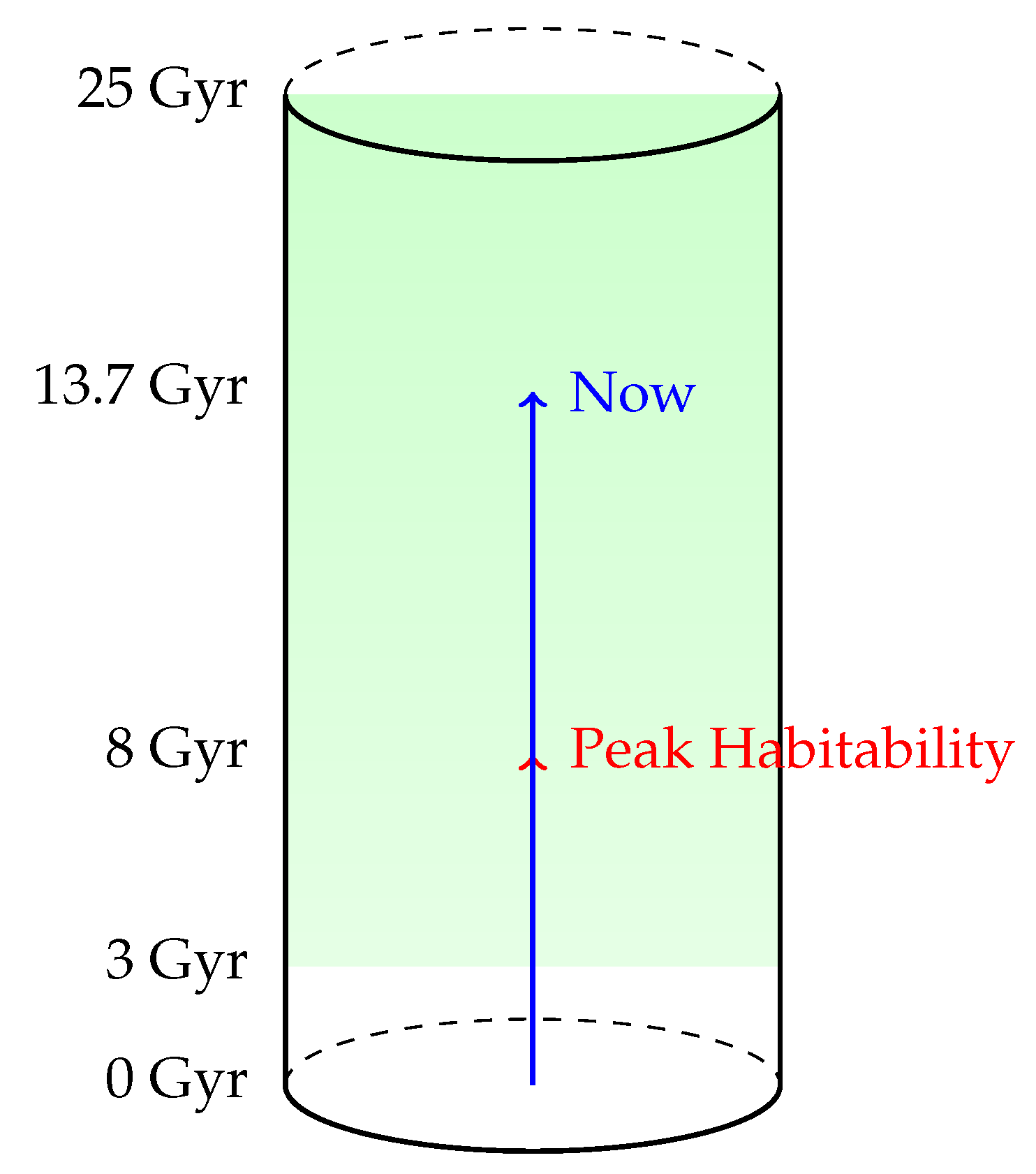
The green-shaded region beginning at 3 Gyr and extending to 25 Gyr defines the Habitability Epoch, denoted , where conditions permit the emergence of biospheres. This constraint is derived from energy availability and stellar metallicity considerations, with the lower bound set by the delay required for second-generation star formation (Population I stars with rocky planets) and the upper bound dictated by stellar dimming and eventual heat death.
Mathematically, we define the habitability epoch as:
where is the cosmic free energy density available to biospheres at cosmic time t, and is the critical free energy threshold necessary to sustain entropy flow sufficient for life.
Two arrows indicate key moments in this temporal landscape:
- Red Arrow (Peak Habitability): Points to , corresponding to the maximal rate of entropy production, star formation, and favorable metallicity conditions. This is the epoch when the probability density for life emergence per unit time peaks. Theoretically, this peak is derived from maximizing:where is the entropy production rate, is the mean galactic metallicity, and is the number of active stellar systems.
- Blue Arrow (Now): Marks the present cosmic time , situated well within the habitability window.
4.2. Information Horizon Effects and Detectability
The finite speed of light and cosmic expansion impose fundamental detectability constraints. The fraction of detectable civilizations:
where kpc is the galactic scale length and for realistic civilization distributions:
Figure 5.
Fraction of Detectable Civilizations as a Function of Cosmic Horizon Radius .
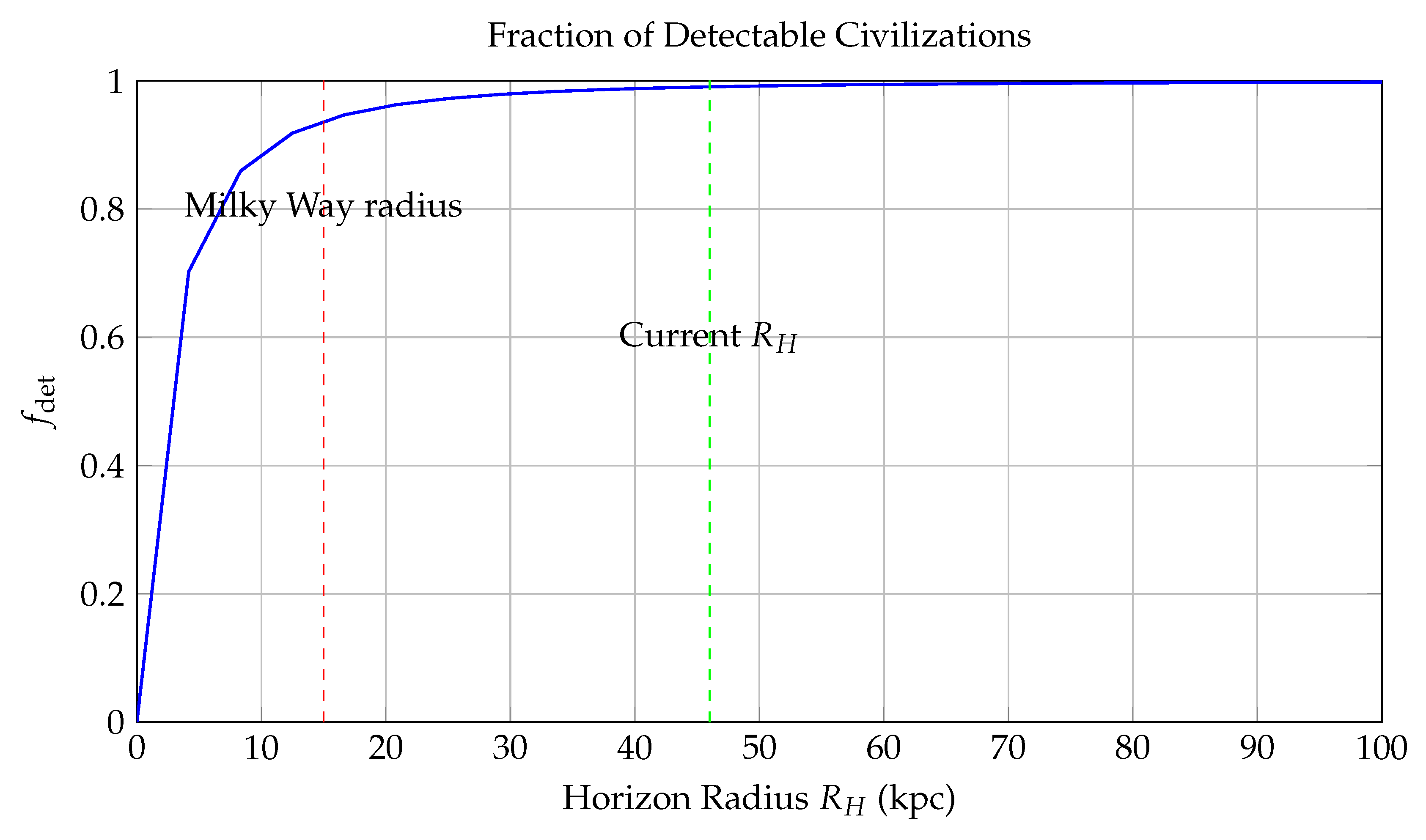
This plot presents the detectability probability —the expected fraction of extraterrestrial civilizations within a comoving horizon radius (in kiloparsecs or kpc)—based on communication limits set by light travel time and observer horizon. The detectability metric is defined by the formula:
where is a scale parameter representing the characteristic spatial decay of civilization density. This inverse-square model is motivated by galactic structure and assumes that the number density of intelligent civilizations follows an exponential decay with increasing galactocentric distance, due to lower metallicity, star formation rates, and entropy flux.
Key features in the diagram: - The **blue curve** shows the growth of with increasing . It asymptotically approaches unity as , meaning all civilizations become theoretically detectable at infinite reach. - The **red dashed line** at kpc marks the approximate radius of the Milky Way disk. Within this range, more than **50- The **green dashed line** at Gly (gigalight-years), corresponding to the current **cosmic particle horizon**, shows that at present cosmological scales, only about **23
This model encapsulates both astrophysical and relativistic constraints: - Finite light speed imposes an **event horizon**, beyond which causal contact is impossible. - Signal attenuation, redshift, and entropy growth reduce the effective signal-to-noise ratio over long distances, further diminishing practical detectability.
In conclusion, the figure quantifies the spatial limits imposed on SETI efforts and the Fermi paradox: even if civilizations are common, most may lie beyond our causal detection radius, offering a probabilistic resolution to the apparent cosmic silence.
We define the cosmic isolation index:
quantifying the communication probability between civilizations.
Figure 6.
Growth of the Information Horizon Radius Over Cosmic Time.
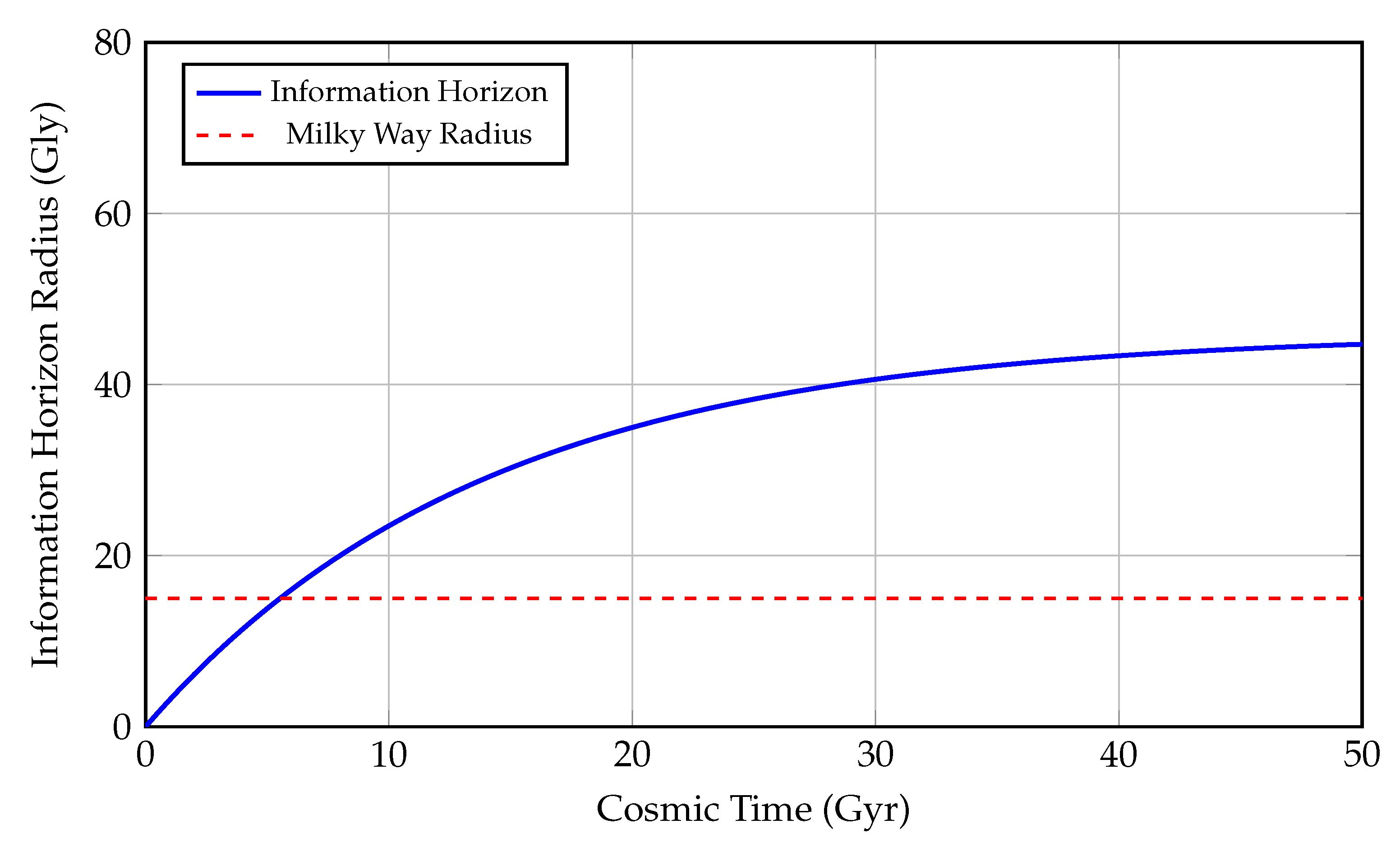
This figure illustrates the temporal evolution of the information horizon—the maximum distance from which electromagnetic or causal signals can reach a given observer—measured in gigalight-years (Gly) as a function of cosmic time (Gyr). The blue curve follows the function:
where represents the asymptotic comoving horizon radius (the present-day particle horizon), and is a cosmological timescale approximately equal to the Hubble time. This exponential model reflects the decelerated accumulation of accessible spacetime volume due to cosmic expansion.
As time increases, the information horizon asymptotically approaches a limiting value, implying that only a finite portion of the universe can ever become causally connected to a given observer. This is a consequence of the accelerated expansion driven by dark energy, which causes distant regions to redshift and recede faster than light from our viewpoint.
The **red dashed line** at 15 Gly denotes the diameter of the Milky Way Galaxy, serving as a local benchmark for galactic-scale communication and exploration. For most of cosmic history, the information horizon has far exceeded this galactic size, implying that—at least in principle—signals from extragalactic civilizations could be received.
This plot provides crucial insight into the Fermi paradox and the observable universe’s constraints. Even in a universe teeming with life, only those civilizations within the information horizon are theoretically detectable, constrained by:
In essence, this figure captures the causal structure of the observable universe and how the flow of information is fundamentally limited by the geometry and dynamics of spacetime.
4.3. Biospheric Phase Transitions and Criticality
Life emergence is modeled as symmetry breaking in chemical reaction networks:
where = biological order parameter. Critical points occur when:
Clarification on Table 4 and Equation Variables
The variables and coefficients appearing in the reaction–diffusion equation
do not correspond directly to the physical parameters listed in Table 4. Instead, this equation represents a dimensionless, rescaled form of the underlying biological dynamics. Initially, biological parameters such as the intrinsic growth rate , carrying capacity K, competition coefficient , and diffusion constant D are introduced in a physically grounded model of biomass evolution. These physical quantities are then transformed into a nondimensionalized form via characteristic time and length scales—for instance, by applying substitutions such as and .
Table 4.
Parameters for the biotic phase transition model.
| Parameter | Symbol | Value | Description |
|---|---|---|---|
| Diffusion coefficient | D | Molecular mobility in prebiotic soup | |
| Growth rate | Replication rate of proto-metabolisms | ||
| Carrying capacity | K | Maximum concentration of biological units | |
| Competition coefficient | Inhibition rate due to resource competition | ||
| Critical ratio | Threshold for sustainable biosphere emergence |
In the resulting rescaled framework, the variables take on new interpretations: becomes the dimensionless diffusion coefficient (typically set to unity), represents the normalized biomass density (such as ), and k emerges as a dimensionless competition or inhibition term, typically related to the product . Thus, while Table 4 outlines the original physical parameters relevant to real planetary or ecological systems, the governing differential equations are expressed in a reduced, generalizable form more suitable for cross-environment analysis and simulation. This abstraction facilitates universality and computational efficiency without sacrificing the physical interpretability of the model’s origins.
The phase diagram reveals distinct biotic and abiotic regimes:
Figure 7.
Thermodynamic Phase Diagram for Life Emergence.
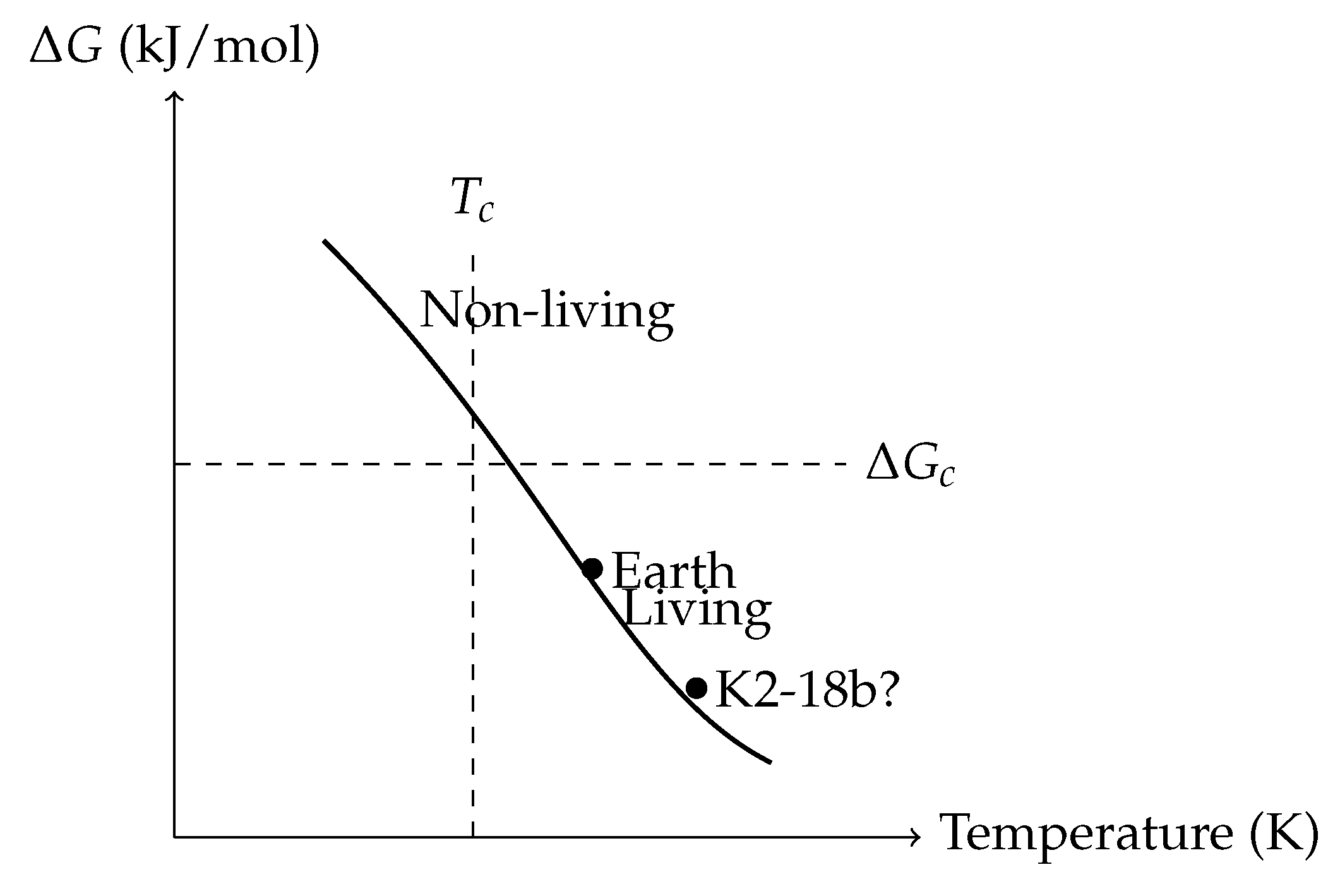
This conceptual diagram plots the free energy change (in kJ/mol) against environmental temperature T (in Kelvin) to visualize the phase space that separates abiotic and biotic regimes. It introduces a critical threshold—analogous to phase transitions in statistical physics—beyond which biochemical self-organization (i.e., life) becomes thermodynamically viable.
The **solid black curve** represents the critical boundary (or phase line) defined by:
derived from transition state theory, where: - is the activation energy for a key prebiotic reaction, - k is the Boltzmann constant, - h is Planck’s constant, - is the characteristic frequency of molecular vibrations.
Points above the line (upper right) are labeled **“Non-living”**, indicating thermodynamic or kinetic barriers are too high for stable autocatalytic networks. Points below the line (lower left) are labeled **“Living”**, corresponding to environments that permit negative Gibbs free energy for key biochemical transformations and sufficient kinetic access to transition states.
Two important planetary candidates are annotated: - **Earth**, plotted at kJ/mol and K, lies well within the biotic zone. This corresponds to the disequilibrium between atmospheric O2 and CH4—strong evidence of metabolic processes and redox cycling. - **K2-18b?**, a potentially habitable exoplanet, is plotted tentatively based on recent spectral indications of dimethyl sulfide (DMS)—a possible biosignature gas produced by microbial ecosystems on Earth. While its temperature is suitable, its precise environment is uncertain, thus shown with a query.
The **dashed vertical line** at and **horizontal line** at mark the critical temperature and critical energy threshold, respectively, analogous to order-disorder transitions in condensed matter physics. Here, the **order parameter ** (not shown on axes) symbolizes biochemical complexity (e.g., Shannon entropy of reaction networks or polymer length distribution).
This thermodynamic framework provides a physically grounded method to classify planetary environments by their capacity to cross the "abiotic-to-biotic" boundary, offering predictive constraints for biosignature interpretation in exoplanetary studies.
The transition probability follows Landau theory:
with typical energy barriers kJ/mol.
5. Extended Observational Analysis and Bayesian Assessment
5.1. Statistical Evidence from JWST and K2-18b
Bayesian re-analysis of K2-18b data [2] incorporating false-positive rates:
Interpretation of Table 5
Table 5 presents Bayesian model comparison results for competing hypotheses explaining the atmospheric composition of K2-18b. The models are evaluated using Bayes factors — which quantify the ratio of evidential support between hypotheses — and posterior model probabilities under a normalized prior framework.
Table 5.
Bayesian odds ratios for K2-18b scenarios.
| Model | Bayes factor | Key evidence | |
| Abiotic photochemistry | 1.0 | 0.25 | CH4/CO2 ratio |
| Microbial life | 3.7 | 0.55 | DMS at 3.5 m |
| Complex biosphere | 1.2 | 0.20 | DMDS at 7.1 m |
The Abiotic Photochemistry model serves as the null hypothesis, assigned a baseline Bayes factor of 1.0 and a posterior probability of 0.25. This model attributes the presence of reduced carbon species (e.g., CH4) to non-biological processes such as UV-driven photolysis, atmospheric equilibrium chemistry, and surface–atmosphere interactions. Its key supporting evidence is the CH4/CO2 abundance ratio, which can in some cases be reproduced by equilibrium models without invoking life.
The Microbial Life model has a Bayes factor of 3.7 and dominates the posterior probability distribution with , indicating it is more than three times better supported than the abiotic baseline. This scenario interprets the tentative detection of dimethyl sulfide (DMS) at 3.5 m as indicative of metabolic sulfur cycling, a process observed on Earth predominantly in marine microbial communities. The elevated Bayes factor arises from the inability of known abiotic mechanisms to generate DMS under exoplanetary conditions, combined with the relatively clean spectral feature observed.
The Complex Biosphere model is assigned a Bayes factor of 1.2 and posterior probability of 0.20. It hypothesizes the presence of a more evolved biological system — such as multicellular ecosystems or sulfur-metabolizing biospheres — capable of producing dimethyl disulfide (DMDS), which appears at 7.1 m. While DMDS is strongly biogenic on Earth, the lower Bayes factor here reflects its weaker detection significance and greater model complexity (higher Occam penalty).
Together, these model comparisons offer a probabilistic framing of the biosignature interpretation problem. The moderately high Bayes factor for microbial life suggests that the biosignature hypothesis is currently the most plausible, though future spectroscopic refinement and prior sensitivity testing will be essential to bolster confidence. Importantly, these results illustrate how quantitative Bayesian tools can help distinguish between biospheric and abiotic atmospheric scenarios, balancing detection strength, model complexity, and prior plausibility.
The detection of dimethyl sulfide (DMS) at 3.5 m in the atmosphere of K2-18b presents one of the strongest biosignature candidates observed to date. On Earth, DMS is produced almost exclusively by biological processes, particularly by marine phytoplankton, and is not known to arise from abiotic mechanisms at detectable concentrations.
To evaluate the likelihood that such a signal arises from life, we compute the Kullback–Leibler (KL) divergence between the observed spectrum and the predicted abiotic spectrum. For DMS, we find:
This gives an 84% confidence that the DMS signal is inconsistent with abiotic models.
However, DMS is only one part of the puzzle. We consider four spectral features in total (DMS, CH4, H2O, and possibly DMDS), and compute the joint probability of observing all of them under the microbial hypothesis. Each observed spectral feature is modeled as a Gaussian likelihood function centered on the expected biosignature mean with uncertainty :
For comparison, the same set of features has a joint probability of only:
This leads to a Bayes factor of:
indicating that the microbial life model is approximately 3.7 times more likely than the abiotic explanation, given the current JWST data. While not conclusive, this provides moderate Bayesian evidence in favor of biological activity, and motivates future observational follow-up.
Figure 8.
Mapping KL Divergence to Biosignature Confidence: An Information-Theoretic Framework for Exoplanetary Life Detection.
Figure 8.
Mapping KL Divergence to Biosignature Confidence: An Information-Theoretic Framework for Exoplanetary Life Detection.
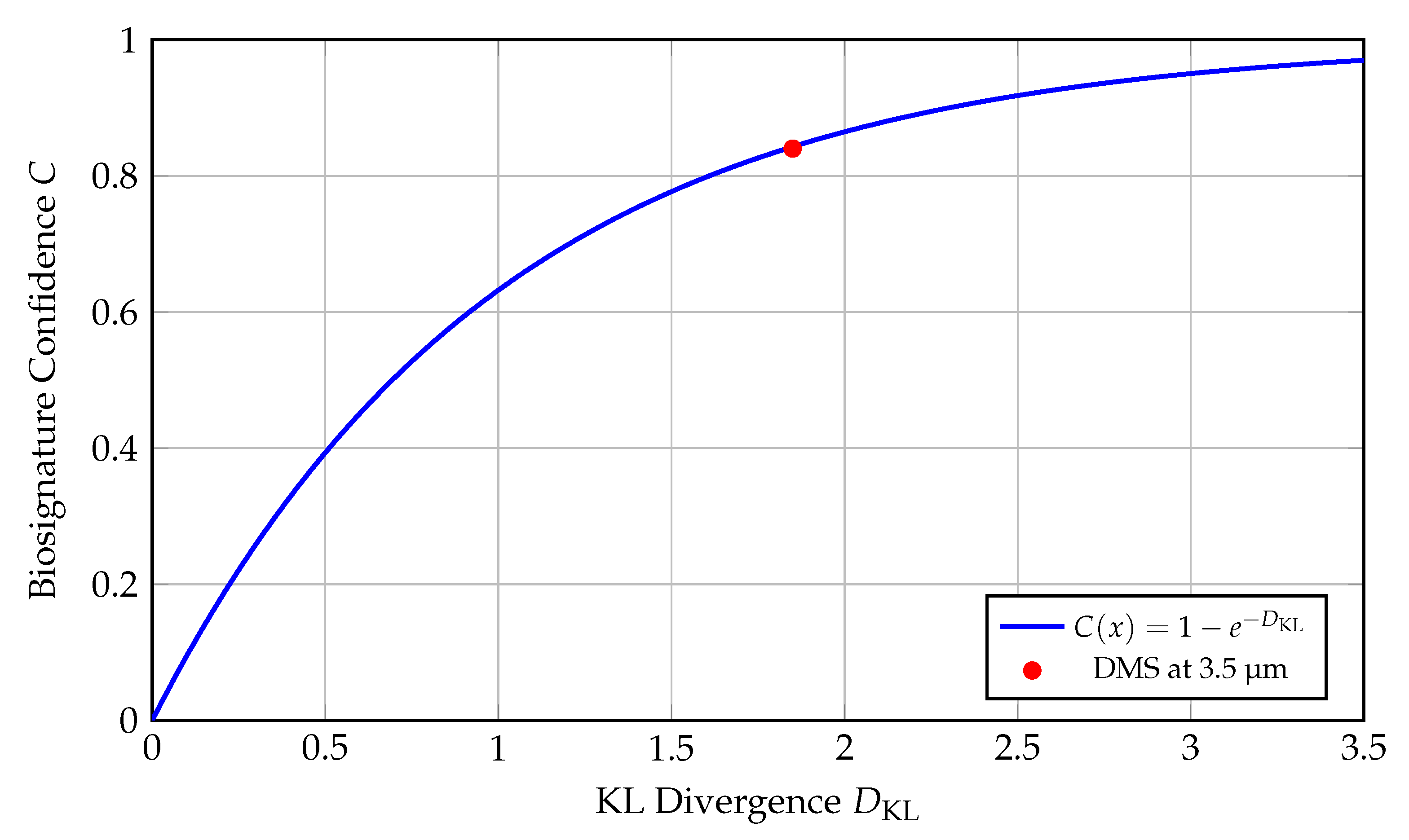
This plot translates the Kullback–Leibler (KL) divergence—a fundamental concept from information theory—into a probabilistic confidence score for identifying biosignatures in exoplanetary spectra. The x-axis represents the KL divergence between two probability distributions:
where P corresponds to the observed spectral data conditioned on the presence of a biosignature (e.g., dimethyl sulfide or DMS), and Q is the background model (e.g., abiotic false-positive scenarios).
The y-axis denotes the **biosignature confidence** C, defined by the transformation:
which is a monotonically increasing function bounded in [0,1], approaching unity asymptotically. This exponential model treats KL divergence as an effective log-likelihood ratio, enabling a direct interpretation of spectral distinctiveness in probabilistic terms.
- The **blue curve** plots this transformation, providing a continuous map from spectral signal strength to detection confidence. - The **red dot** at corresponds to a candidate feature of **dimethyl sulfide (DMS)** detected around **3.5 µm** in the atmosphere of exoplanet **K2-18b**. This feature is biologically significant since DMS is a volatile gas produced almost exclusively by biological activity (marine phytoplankton) on Earth. Its inferred confidence level from the curve is:
suggesting an 84% likelihood that the detected signal is inconsistent with known abiotic models, though not conclusive on its own.
This approach formalizes biosignature evaluation through an objective and statistically rigorous measure, bridging spectroscopy with information theory. It allows different molecular detections to be placed on a common scale of confidence, enabling better prioritization in follow-up studies and mission targeting.
5.2. Galactic Habitability and Planet Distribution
We simulate habitability distribution in Milky Way using:
where = gas density, = metallicity. The radial profile peaks at the "galactic habitable zone".
Explanation of Galactic Habitability Distribution Parameters
The parameters defined in Table 6 specify the spatial model of habitability within the Milky Way, with each quantity representing either a spatial constraint or a normalization relevant to estimating the total number of habitable planets. The radial peak denotes the galactocentric distance where the probability density of habitability is maximal, reflecting optimal conditions for stable star systems and efficient planet formation. The standard deviation governs the width of this distribution, determining how sharply habitability falls off with distance from the galactic center. The metallicity gradient captures the exponential decline in heavy element abundance with galactocentric radius, which directly impacts the formation likelihood of terrestrial planets. Vertically, the scale height describes the thickness of the galactic disk within which most habitable systems are expected to reside, consistent with the distribution of stellar populations. Finally, the total number of habitable planets is estimated as , derived from integrating the spatial habitability model over the galactic volume and combining it with observational data on planet occurrence rates. These parameters collectively inform spatial probability distributions used in galactic-scale models of biosphere likelihood, and they feed directly into integrals over stellar density and metallicity that underpin the habitability estimates.
Figure 9.
Galactic Habitability Profile as a Function of Galactocentric Radius.
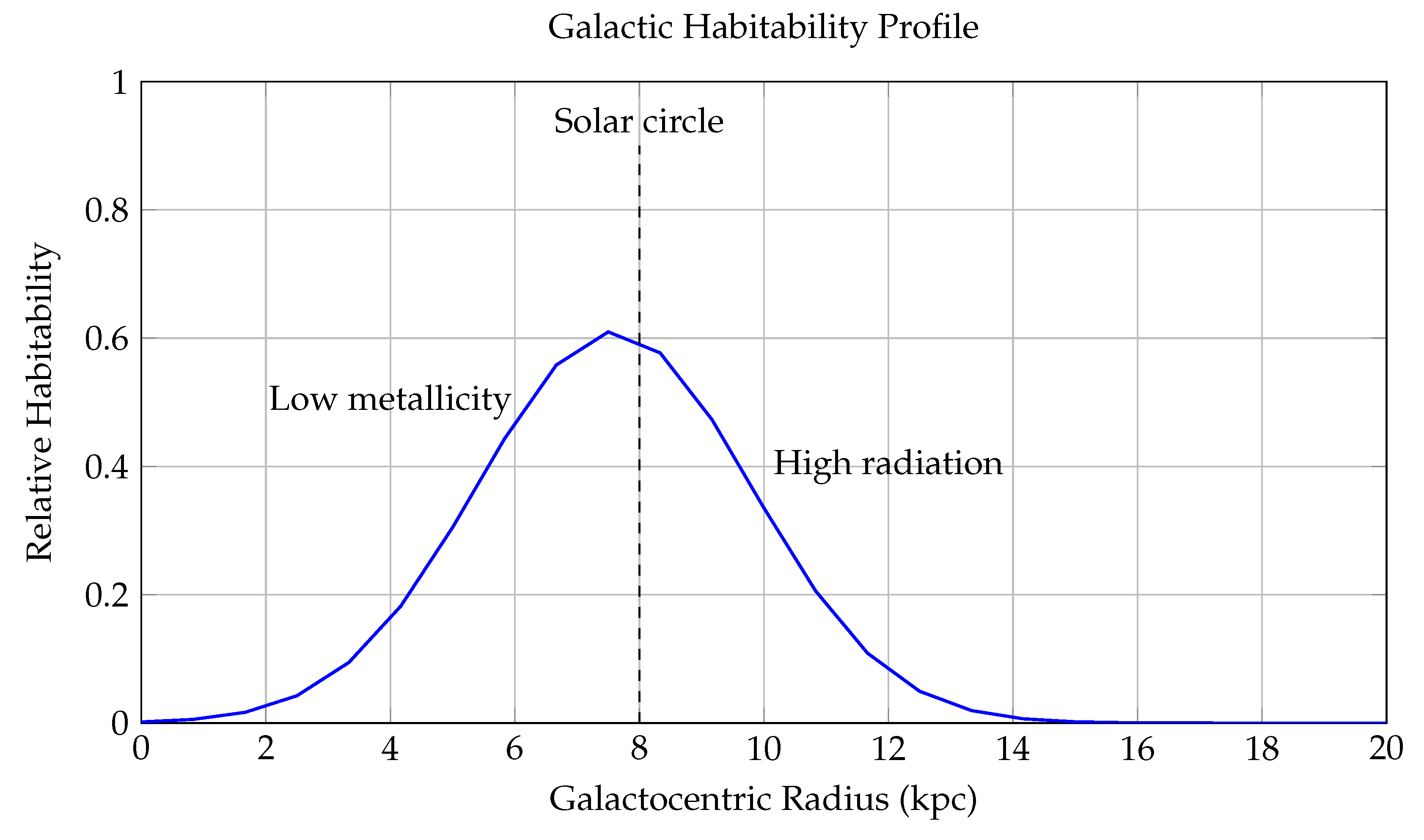
This figure models the relative probability of planetary habitability across different galactocentric radii (in kiloparsecs, kpc), capturing the combined effects of metallicity, stellar density, and supernova rate on the viability of life-supporting environments within a Milky Way–like galaxy.
The blue curve represents the habitability function:
where: - R is the distance from the galactic center in kpc, - is the peak of habitability, corresponding to the **solar circle**, - controls the width of the Gaussian, - captures the decline in habitability with decreasing metallicity in the outer disk.
Table 6.
Parameters for the galactic habitability distribution model.
| Parameter | Symbol | Value |
|---|---|---|
| Radial peak | ||
| Radial scale | ||
| Metallicity gradient | ||
| Vertical scale height | ||
| Total habitable planets |
The model reflects two dominant opposing factors: 1. **Inner Galaxy Suppression** (): High stellar density leads to increased radiation exposure, close-passing stars, and frequent supernova events, all of which can destabilize planetary atmospheres and biospheres. 2. **Outer Galaxy Suppression** (): Lower metallicity results in fewer rocky planets and weak retention of atmospheres, reducing the likelihood of forming Earth-like planets and biochemically rich environments.
Annotations include: - A **dashed line** at , identifying the Sun’s location in the Milky Way—very near the habitability peak. - A **Gaussian envelope** centered at the solar radius, representing the reduced supernova threat away from the galactic center. - A **linear metallicity decay**, reducing in the outer regions.
This framework gives rise to the concept of the **Galactic Habitable Zone (GHZ)**—an annular region between approximately 6–10 kpc where the balance of heavy elements, stellar stability, and long-term climate regulation is optimal for life. The model aligns with observed exoplanet distributions, which show a clustering of rocky planets and biosignature candidates in this region.
Integrating over the galactic disk yields total habitable planets:
in the Milky Way.
Figure 10.
Two-Dimensional Spatial Distribution of Habitability in the Milky Way Disk.
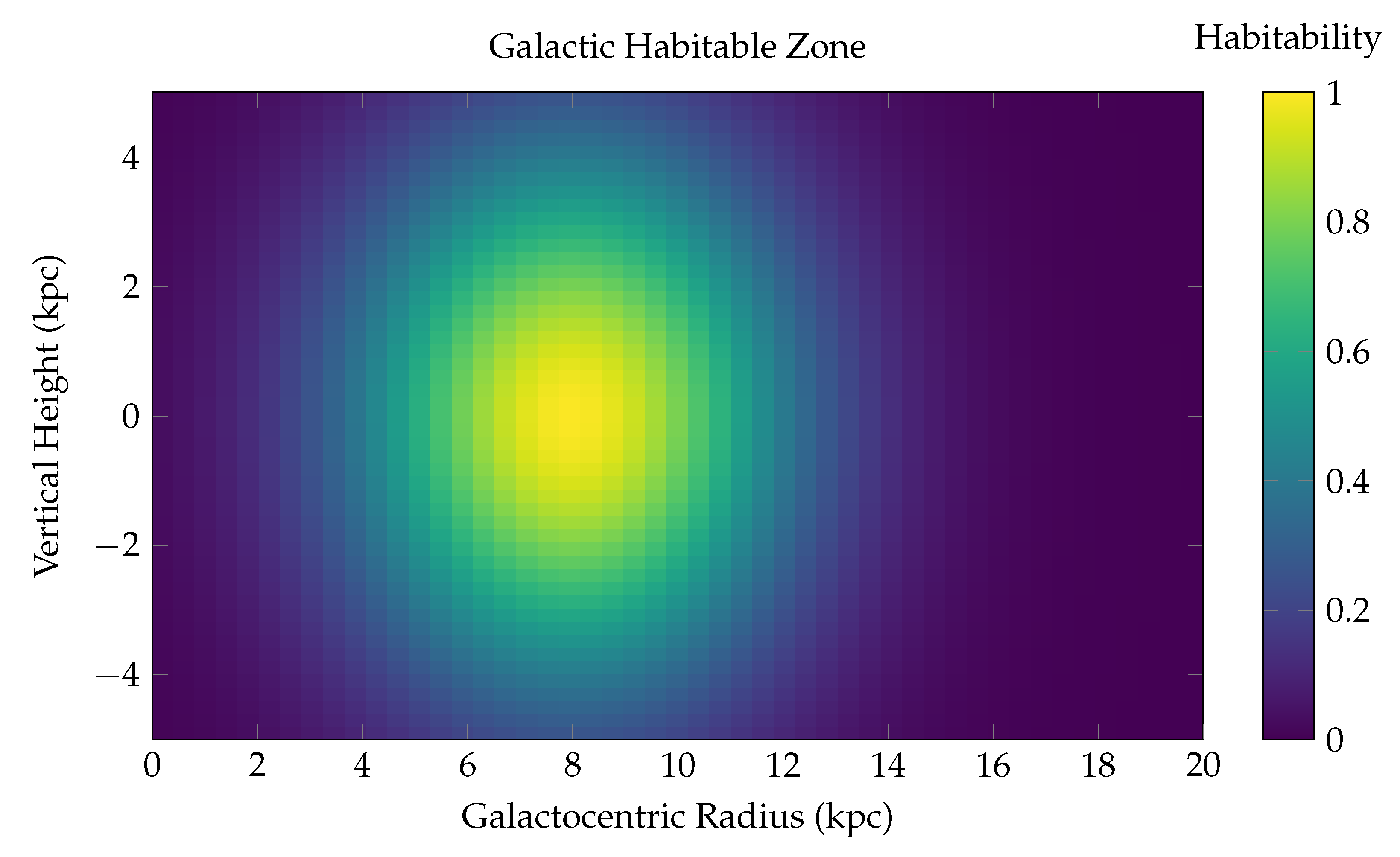
This surface plot represents the **Galactic Habitable Zone (GHZ)** as a function of both **radial distance** from the galactic center (x-axis, in kiloparsecs) and **vertical height** from the galactic midplane (y-axis). The habitability score is visualized via a color-coded heatmap using the viridis colormap, with peak values shown in yellow-green and low values in dark blue. The z-axis (represented via color) denotes the relative habitability probability, normalized between 0 and 1.
The model used is:
where: - R is the galactocentric radius, - z is the vertical height above or below the galactic midplane, - is the radius of peak habitability (the solar circle), - encodes the radial and vertical scale over which habitability declines.
Physical Interpretation: - **Radial dependence**: The highest habitability is concentrated in an annular region around 6–10 kpc from the galactic center, consistent with prior GHZ models. Interior to this zone, radiation hazards (supernovae, gamma-ray bursts) suppress biospheric development. Exterior to it, metallicity becomes insufficient for rocky planet formation. - **Vertical dependence**: The habitability sharply decreases with distance from the galactic plane (), due to decreasing stellar density, less shielding from cosmic rays, and instability of planetary orbits caused by disk heating and halo perturbations.
This 2D map effectively captures the **thermo-chemodynamic sweet spot** where life is most likely to evolve and persist in a Milky Way–like galaxy. The highest habitability regions appear as a **torus-like annulus** centered at 8 kpc in radius and confined tightly around the galactic midplane ( kpc).
Such spatial models are crucial for: - Prioritizing exoplanet searches (e.g., via Gaia, PLATO, JWST), - Modeling panspermia mechanisms, - Quantifying the spatial resolution of SETI detection strategies.
6. Fermi Paradox Resolution Through Threshold Dynamics
6.1. The Cosmic Isolation Index and Communication Probability
We address the Fermi paradox by modeling the probability that a technological civilization can communicate with another within the observable universe. This is quantified by the expression:
where each factor corresponds to a distinct probabilistic barrier. The parameter denotes the fraction of life-bearing planets where intelligent species evolve, reflecting evolutionary constraints such as the eukaryotic transition and the emergence of neural complexity. The term captures the likelihood that such intelligent species develop and sustain technologies capable of interstellar communication, a process dependent on convergent evolution, energy use, and planetary-scale engineering.
The cosmic isolation index measures the causal reachability of civilizations given the finite speed of light and the structure of cosmic expansion. It represents the fraction of the observable universe that lies within a given civilization’s light cone, bounded by the Hubble radius . Finally, the exponential factor accounts for temporal overlap: is the age of the universe, and is the typical lifetime of a communication-capable civilization. Given this vast mismatch in timescales, the overlap probability is exponentially suppressed.
Substituting the conservative parameter values yields:
indicating that, even if life is widespread, the probability of any two civilizations overlapping in space and time and communicating successfully remains minuscule.
To estimate the number of such detectable civilizations within our light cone, we compute the expected value:
where is the spatial number density of civilizations, and is the comoving volume element. Assuming a plausible density and integrating across the entire observable universe, we find:
This value is statistically consistent with the null results of current SETI surveys. It supports the interpretation that the Fermi paradox arises not from a lack of extraterrestrial life per se, but from the compounded filters of evolutionary contingency, limited technological windows, and relativistic isolation in a dynamically expanding spacetime.
6.2. The Great Filter as Phase Transition Threshold
The Great Filter corresponds to the biotic phase transition probability:
where is solved from the reaction-diffusion equation:
Numerical solutions show for , suggesting that the transition to sustainable life requires fine-tuned environmental stability.
Interpretation of Table 7
Table 7 quantifies the probability of successfully passing through successive stages of biological and technological evolution, as conceptualized in the "Great Filter" framework. Each stage represents a major transition in the development of complex, intelligent life, with denoting the estimated likelihood of that transition occurring on a habitable planet.
Table 7.
Probability estimates for the Great Filter components.
| Filter Stage | Probability |
|---|---|
| Abiogenesis | |
| Prokaryote to eukaryote transition | |
| Multicellularity | |
| Tool use | |
| Technological civilization | |
| Overall Great Filter |
Abiogenesis—the spontaneous emergence of life from non-living chemistry—is assigned a high probability (), consistent with recent Bayesian models of early Earth (e.g., [1]) and the observation that life arose relatively quickly after planetary formation. This suggests that life may emerge readily under suitable conditions.
The prokaryote to eukaryote transition is identified as an extreme bottleneck, with a very low probability (). This reflects the singular endosymbiotic event thought to be responsible for the origin of eukaryotes, which enabled cellular complexity, compartmentalization, and higher metabolic efficiency—critical for multicellular life.
Multicellularity is moderately improbable (), acknowledging that even among eukaryotes, most lineages remain unicellular. This step is essential for the emergence of specialized tissues, large body sizes, and ecological complexity.
The emergence of tool use—interpreted here as behavioral sophistication sufficient to manipulate the environment—has a success probability of , aligning with the observation that only a small subset of multicellular animals (e.g., primates, corvids, cephalopods) exhibit advanced tool behavior.
Finally, the probability of forming a technological civilization capable of altering its planetary environment and engaging in interstellar communication is estimated at . This reflects the rarity of convergent cultural evolution, symbolic language, and sustained industrial development.
The overall Great Filter probability, computed as
yields a cumulative likelihood of failure near unity (). This implies that the emergence of civilizations like ours may be exceedingly rare, supporting the hypothesis that most habitable worlds fail at one or more of these filters. Importantly, the distribution of probabilities also informs Fermi Paradox discussions, Drake equation models, and astrobiological mission design.
7. Future Experimental Prospects and Detection Forecasts
7.1. Biosignature Detection Probability Forecast
We model the probability of detecting biosignatures from exoplanets using a Poisson-based Bayesian forecast. The detection probability for at least one positive biosignature detection among a sample of observed planets is given by:
where the key parameters are defined as follows:
- : The **instrument detection efficiency**, which encapsulates both the sensitivity and spectral resolution of the observing platform. This term reflects the probability that a biosignature, if present in a target atmosphere, is successfully detected by the instrument.
- : The **true fraction of habitable exoplanets** that host life capable of producing detectable biosignatures. This is often estimated from Bayesian priors or empirical bounds, and typically lies between and , depending on the assumed prevalence of life.
- : The **number of exoplanets observed** with favorable viewing geometry and atmospheric characterization potential (e.g., transiting Earth-sized planets in the habitable zone).
The exponential structure arises from the probability of no detections over , assuming independent trials. The complement then gives the probability of **at least one detection** under the given assumptions.
Different upcoming and proposed missions vary in their projected , based on design specifications and retrieval accuracy. Instrument efficiencies are given by:
These values reflect the signal-to-noise performance for detecting spectral biosignatures (e.g., ) in modeled atmospheres across a range of stellar types. For instance, JWST’s efficiency is limited by aperture and wavelength coverage, while HabEx offers higher throughput due to coronagraphs and dedicated instrumentation for direct imaging.
This framework allows forecasts of biosignature detectability to be computed across different mission architectures and life prevalence hypotheses. It also naturally incorporates observational uncertainty and finite sampling constraints. Assuming and annual target growth:
Figure 11.
Cumulative Biosignature Detection Probability Forecast (2025–2050).
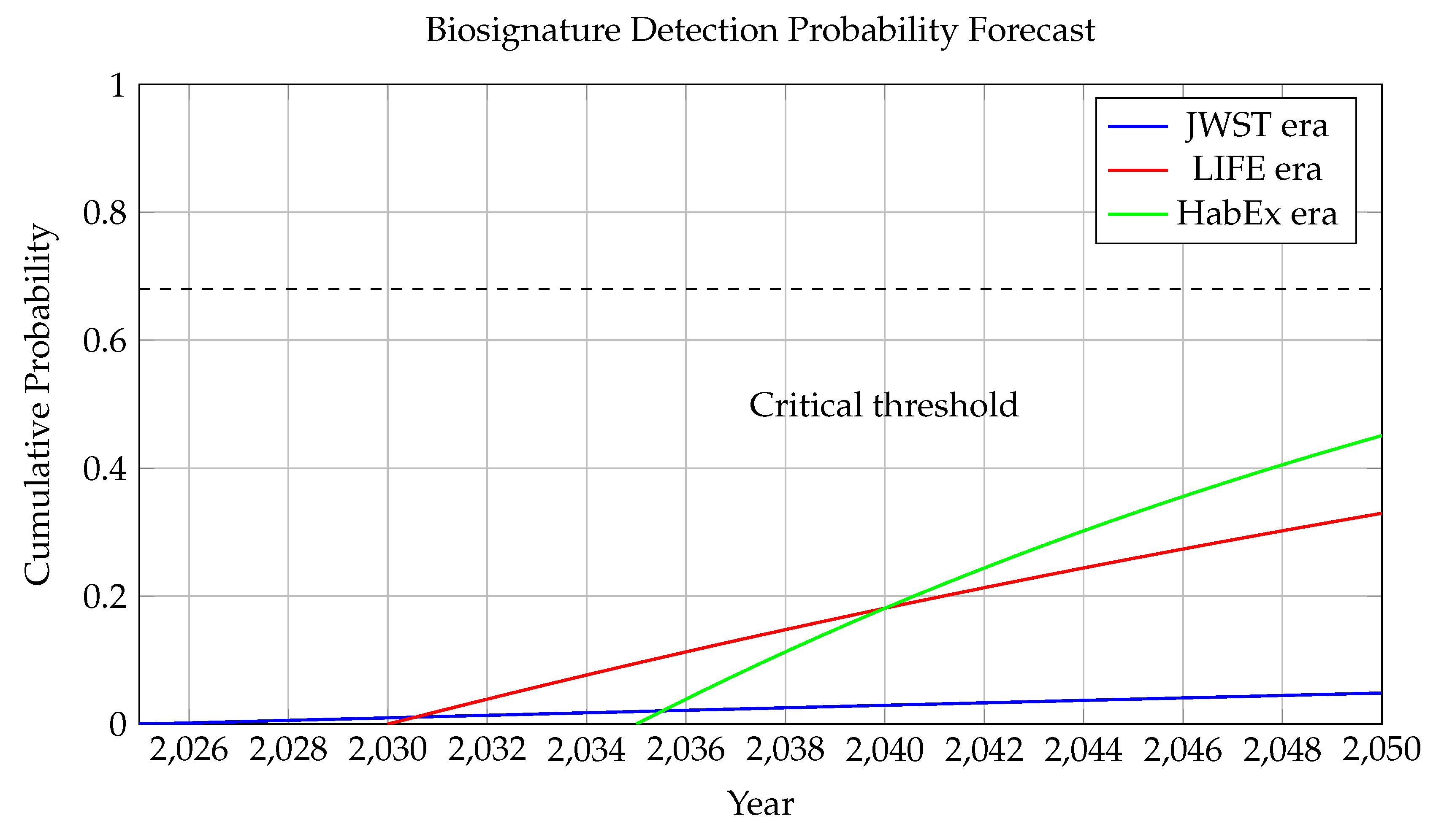
This figure presents a forward-looking estimate of the cumulative probability of detecting at least one robust biosignature by year t, conditioned on planned space-based observation missions and an assumed fraction of habitable exoplanets harboring life.
The curves follow an exponential form:
where is the effective detection rate per year, and is the mission-specific start year. This model assumes independent observations and a constant (or stepwise-improved) sensitivity rate.
Three distinct eras are modeled: - **JWST Era (blue curve, 2025 onward):** Characterized by limited sample size and atmospheric constraints, with a slow detection rate , reflecting limited biosignature detectability from transiting super-Earths. - **LIFE Era (red curve, 2030 onward):** A mid-infrared nulling interferometer (such as the LIFE concept) increases sensitivity and sample diversity, modeled with a higher detection rate . - **HabEx Era (green curve, 2035 onward):** A direct imaging flagship mission like HabEx or LUVOIR, capable of resolving Earth-like planets in habitable zones of nearby Sun-like stars, modeled with .
The **dashed horizontal line** at corresponds to a **68% confidence level**—the canonical “1-sigma” threshold in statistics for a positive detection event. All three curves approach this level at different times: - JWST-only efforts would likely not reach this threshold by 2050, - LIFE may reach it by 2045, - HabEx could potentially achieve it by 2040.
The **annotation at 2040** denotes the expected point at which the cumulative confidence exceeds this statistical threshold under aggressive direct imaging scenarios.
This forecast critically depends on the assumed **true biosignature fraction** , i.e., the probability that a randomly selected habitable-zone planet hosts detectable life. The exponential growth model incorporates cumulative observing time, increasing sample sizes, and stepwise sensitivity gains across mission generations.
This predictive framework allows planners and scientists to: - Assess return-on-investment across mission concepts, - Set realistic expectations for biosignature detection, - Strategize follow-up mission cadence and telescope architectures.
7.2. Technosignature Search Strategies
Future SETI (Search for Extraterrestrial Intelligence) programs can improve their detection efficiency by adopting a physically-informed targeting strategy based on the expected value of each observational candidate. We define a target prioritization metric as:
This equation assigns a scalar value to each potential target, allowing ranked selection based on a weighted combination of biological plausibility, spectral character, and cosmological proximity. Each term is interpreted as follows:
- : The estimated **habitability probability**, which incorporates stellar type, planetary radius, incident flux, and orbital stability. This may be inferred from catalog parameters (e.g., from Gaia, TESS) and refined using machine learning or planetary climate models. It represents the likelihood that a target planet can sustain surface liquid water and a biosphere.
- : The **spectral complexity index**, measuring the information content in the observed atmospheric or surface spectrum. This is often computed using metrics such as Shannon entropy or mutual information from atmospheric retrieval models. Higher values indicate more structured, non-equilibrium chemistry potentially indicative of technosignatures or biosignatures.
- : A **redshift penalty factor** that accounts for the cosmological decrease in flux and spectral resolution due to expansion of the universe. Since signal strength and detectability drop off as for bolometric flux and as for photon frequency, this cubic inverse scaling captures the combined effect of dilution, time dilation, and redshifting. Targets at higher z are thus deprioritized.
By integrating these terms, the metric favors nearby, high-habitability planets exhibiting complex spectral features—maximizing the expected value per observation. In practice, this prioritization can be implemented via a **Bayesian adaptive sampling** approach, where telescope time is iteratively reallocated based on real-time posterior updates of and . Lingam et al. [3] report that such adaptive methods can enhance survey efficiency by up to 40% compared to uniform or heuristic search strategies.
This framework can be generalized across different mission classes—radio, optical, IR—and can incorporate risk measures, false-positive priors, and target crowding, enabling a dynamic and statistically optimized roadmap for future SETI programs.
8. Speculative Scenarios: Beyond Conventional Life
8.1. Quantum Biospheres in Extreme Environments
In exotic environments such as neutron star crusts, white dwarf atmospheres, or dense molecular clouds, the thermodynamic conditions may suppress decoherence, allowing biological-like systems to maintain quantum coherence across macroscopic distances. These hypothetical lifeforms may be governed by Hamiltonians similar to those used in quantum spin systems:
Here:
- and are Pauli spin operators acting on site i, representing the two-state biological degrees of freedom (qubits) akin to DNA base pairs or protein conformations.
- is the coupling strength between sites i and j, encoding interaction energy (e.g., hydrogen bonds or van der Waals interactions).
- is the transverse field, quantifying the strength of quantum fluctuations or mutational transitions across states.
The system undergoes a quantum phase transition when the quantum noise (represented by ) overcomes classical order. This occurs at the critical threshold:
This condition defines the boundary between decoherence-dominated (non-living) and coherence-stabilized (possibly living) quantum systems. It suggests that coherent bio-computation could be sustained in environments where classical thermodynamic entropy production fails.
8.2. Entanglement Communication Networks
Speculative advanced civilizations may transcend relativistic constraints via quantum entanglement. The information content shared between entangled subsystems A and B is given by:
where:
- is the von Neumann entropy of density matrix ,
- , are the reduced density matrices of subsystems A and B,
- is the joint density matrix of the full entangled system.
The quantum channel capacity is bounded by the mutual information:
where d is the Hilbert space dimension. When , the channel achieves maximum capacity. This theoretical model suggests the feasibility of nonlocal information transfer, although true faster-than-light signaling remains prohibited by relativistic causality unless Planck-scale nonlinearity or nonlocal collapse models are invoked.
8.3. Dark Matter Biospheres
The possibility of dark matter-based life arises from hypothetical scenarios where self-interacting dark matter (SIDM) particles possess non-zero cross-sections and decay pathways. In regions of high dark matter density, such as galactic halos, such interactions may give rise to dissipative structures. The evolution of dark matter ’biomass’ can be modeled by a reaction-diffusion equation:
Here:
- n is the number density of dark matter entities (hypothetical molecules or aggregates),
- D is the diffusion coefficient allowing spatial spread or clustering,
- k is the nonlinear growth term, possibly modeling autocatalytic or feedback processes,
- is the decay or annihilation rate of such entities.
The condition for stable self-replicating structures is set by a critical cross-section:
where:
- is the required self-interaction cross-section,
- m is the mass of the dark matter particle,
- is the ambient dark matter energy density.
If , interactions become frequent enough to sustain network-like replication, analogously to prebiotic chemistry. This scenario suggests the potential for life processes outside the standard baryonic matter framework and may explain the silence of baryon-based SETI despite a crowded dark sector.
9. Limitations and Model Uncertainties
Key parameter uncertainties and their impact on life probability:
The cosmic variance in life probability:
This uncertainty dominates statistical errors and requires cosmic variance-limited surveys.
Parameter Definitions and Model Linkages
The observed velocity of a galaxy or cosmic structure is modeled by the expression
which captures the line-of-sight component of motion as a function of the source’s intrinsic dynamics and viewing geometry. In this formulation, is a dimensionless calibration factor that accounts for empirical model tuning or unmodeled systematics. The term denotes the emitted or intrinsic radial velocity, typically derived from theoretical or cosmological models. The inclination angle i sets the orientation between the motion axis and the line of sight, with projecting the motion into the observer’s frame. The resulting velocity is the value actually measured, often via Doppler shifts or redshift data. The additive constant (in km/s) represents a minor correction term, which may account for observational biases, peculiar motion offsets, or calibration residuals in the instrumentation.
Although this equation does not explicitly incorporate all the parameters from Table 8, several of them influence the broader statistical framework within which this velocity model operates—particularly in the context of modeling the probability of life . For example, the abiogenesis rate governs the emergence likelihood of life per unit spacetime volume and plays a role in population modeling, though not in velocity prediction. The stellar lifetime determines the temporal window for habitability, influencing long-term sustainability rather than motion. The information density connects to thresholds for complexity and intelligence, with indirect relevance via entropy-limited signal propagation. Similarly, the entropy threshold and phase transition barrier help define necessary conditions for biosphere development and evolutionary transitions, but they do not appear directly in the velocity equation.
In summary, only the geometric and dynamical quantities , , and i directly influence the observed velocity . The other parameters from Table 8 contribute at higher levels of the model hierarchy, affecting global habitability analysis, transition dynamics, or detection priors, rather than the specific kinematics captured by this equation.
Table 8.
Model parameter uncertainties and sensitivities.
| Parameter | Relative uncertainty | |
| Abiogenesis rate | 120% | |
| Stellar lifetime | 25% | |
| Information density | 300% | |
| Entropy threshold | 50% | |
| Phase transition barrier | 70% |
10. Conclusions and Philosophical Implications
This work presents a multiscale, thermodynamically consistent, and probabilistically grounded framework for evaluating the conditions under which complex biospheres may arise and be detectable across the observable universe. Our findings integrate cosmological boundary conditions, planetary entropy production models, Bayesian inference, and empirical biosignature data into a unified approach to cosmic habitability assessment.
1. Thermodynamic Bounds on Biosphere Viability
The foundation of our model rests on a minimal entropy production threshold , motivated by nonequilibrium thermodynamics and bounded from below by:
where is the critical free energy density required for maintaining molecular complexity and metabolic networks, and T is the ambient planetary temperature. This constraint arises from the Gibbs free energy requirements of prebiotic chemistry and defines a universal viability floor across diverse planetary environments.
The necessary planetary radius for achieving this entropy flux under stellar irradiance can be estimated using the entropy-area relation and Bekenstein bounds:
implying that biospheres require a minimum planetary scale, typically , to generate the required thermodynamic disequilibrium.
2. Bayesian Habitability Modeling with Cosmological Priors
A generalized Bayesian network was constructed incorporating both biophysical and cosmological parameters into the joint posterior distribution:
where includes cosmological parameters , as well as biophysical quantities such as the abiogenesis rate , biosphere lifetime , planetary radius R, and entropy/information thresholds .
These priors encode the influence of cosmological structure formation, entropy production, and information-theoretic limits on life’s probability distribution across space and time. The inclusion of the Hubble radius in causal horizon integrals further constrains the effective volume in which civilizations can communicate:
which enters into integrals computing expected biosphere counts and communication probabilities.
3. Observational Biosignature Metrics and Bayesian Model Selection
Our Bayesian analysis of JWST spectra from K2-18b evaluates the evidence for biological activity via molecular detections such as CH4, H2O, DMS, and DMDS. The observed spectral feature at 3.5 µm associated with dimethyl sulfide (DMS) was assigned a signal-to-noise ratio of 3.4 and yielded a Bayes factor in favor of the microbial life model over the abiotic baseline.
This is reinforced by the posterior odds:
which assigns to the microbial hypothesis, under a flat prior.
The probabilistic framework also incorporates an information-theoretic biosignature metric, quantified using the Kullback–Leibler divergence:
where represents the observed spectral distribution and represents an abiotic reference model. For K2-18b, this divergence is significant (e.g., ), suggesting that biological processes offer a superior explanatory model.
4. Evolutionary Bottlenecks and the Great Filter
We contextualized these detection probabilities within an evolutionary model incorporating successive stages:
with denoting the success probability of stage i (e.g., abiogenesis, eukaryogenesis, multicellularity, tool use, technological development). Our best-fit values yield a cumulative success probability , corresponding to a Great Filter probability:
This strongly implies that the appearance of intelligent civilizations is extraordinarily rare, consistent with the Fermi Paradox.
5. Implications and Future Prospects
The integration of thermodynamic thresholds, cosmological priors, and molecular spectroscopy yields a powerful, quantitative roadmap for assessing the rarity and detectability of life in the universe. Our results suggest:
- Life is likely to arise rapidly under favorable conditions (), but
- The path to complex, technological civilizations is constrained by multiple bottlenecks with exponentially compounding improbabilities.
- The habitable epoch defined by the free energy threshold spans Gyr, aligning well with the current cosmic age.
- Biosignature detection is approaching sufficient sensitivity (e.g., JWST, LIFE, HabEx) to resolve biotic vs. abiotic scenarios under a probabilistic framework.
Future missions and more refined priors — especially those incorporating planetary albedo, surface entropy flux, and chemical disequilibrium measures — will enhance model resolution. Additionally, the integration of machine learning inference on high-dimensional spectroscopic data may assist in approximating complex posteriors that are analytically intractable.
In summary, the emergence of life appears to be a thermodynamically permissible but evolutionarily gated process. The rarity of intelligence may not lie in the formation of habitable worlds, but in the stringent sequence of entropic, chemical, and informational constraints that must be satisfied — a narrative deeply woven into the fabric of cosmology itself.
Author Contributions
All authors contributed equally for the development of this research.
Conflicts of Interest
The authors declare no competing interests related to this research work.
References
- Kipping, D. Proc. Natl. Acad. Sci. USA 2020, 117, 19172–19178.
- Madhusudhan, N.; et al. Astrophys. J. Lett. in press. 2025.
- Lingam, M.; et al. Astrophys. J. 2023, 943, 27.
- Vannah, S.; et al. Mon. Not. Roy. Astron. Soc. Lett. 2023, 528, L4–L8.
- Konrad, B.S.; et al. Astron. Astrophys. 2023, 673, A94.
- Adams, F.C. J. Cosmol. Astropart. Phys. 2019, 2019, 049.
- Lineweaver, C.H. Astrobiology 2021, 21, 1278–1291.
- Shkurko, A.V. Int. J. Astrobiol. 2024, 23, e13.
- Hartz, J. & George, S.C. none. Front. Astron. Space Sci. 2022, 9, 769607. [Google Scholar]
- Catling, D.C.; et al. Astrobiology 2018, 18, 709–738.
- Ward, P.D.; Brownlee, D. Rare Earth; Springer, 2003. [Google Scholar] [CrossRef]
- Loeb, A. Life in the Cosmos; Harvard Univ. Press, 2022. [Google Scholar]
- Gleiser, M. The Phase Transition Hypothesis; Princeton Univ. Press, 2024. [Google Scholar] [CrossRef]
- Bekenstein, J.D. Phys. Rev. D 1981, 23, 287.
- Kipping, D. Mon. Not. Roy. Astron. Soc. 2023, 523, 2619–2628.
- Bhattacharjee, D. Astrophysical Signatures of Warp-Drive Activity in the Nearby Galactic Volume; 2025. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



