Submitted:
16 January 2026
Posted:
16 January 2026
You are already at the latest version
Abstract
The use of light (photons) delivered clinically from laser or light-emitting diodes (LED), is referred to as photobiomodulation therapy (PBMt). Increasingly PBMt is accepted particularly in dental practice for pain or pre-emptive anaesthesia. Understanding its mechanism of effectiveness is the key to its increasing acceptance. Of major importance to this is how PBMt affects not only the neurons but also the Schwann cells and fibroblasts of the peripheral nervous system which are unique in morphology and function. The specific roles of the neuronal cells of the dorsal root and trigeminal ganglia, now include consideration of the axon initial segment responsible for the initiation of the action potential and the T junction from which the distal and proximal axons arise which are complex but central to normal function. This cellular complexity, organization and function is discussed leading to a review of the mechanism of effectiveness of PBMt demonstrated by clinical trials in both medicine and dentistry. This review provides evidence of the involvement of the cytoskeleton, mitochondrial organization particularly related to fast and slow axonal flow and mitochondrial membrane potential in response to light in somatosensory neurons and nerves.
Keywords:
photobiomodulation (PBM)
; laser
; somatosensory neuron
; nociceptor
; cytoskeleton
; fast axonal flow (FAF)
; axon initial segment (AIS)
; analgesia
; pain relief
; Schwann cell
1. Introduction
Pain remains a problematic and universally important unsolved subject with treatments for acute and chronic pain varied as to their effectiveness. Increasingly among treatments is the use of light i.e., phototherapy or photobiomodulation therapy (PBMt). Although it is accepted that the retina responds to light at varying wavelengths and that Vitamin D is best obtained from sunlight, the idea of using light for pain remains outside mainstream medicine’s accepted practice though is becoming more widely used in standard dental therapy. While there are well-designed clinical trials showing PBMt effectiveness for painful musculoskeletal conditions [1,2,3] and, in dentistry, as a pre-emptive anaesthesia [4,5] the mechanism of its effectiveness is not well understood. Clinically, PBMt is delivered to sites of pathology at specific parameters including wavelength, dose, time protocol, area of delivery and more. However, reference to the unique and complex anatomy of peripheral nervous system (PNS) neurons demonstrates its key role in understanding how PBM affects such neurons which transduce noxious stimuli from the periphery into pain sensation in the central nervous system (CNS). Recent research demonstrating the unique anatomy of these first-order neurons highlights the important function of the axon initial segment (AIS), the complexity of the T-junction and the bifurcation forming the distal and proximal axons [6,7]. This complex organization has changed the focus in unravelling the response of the PNS to PBM. In addition, this review includes evidence of the essential role of Schwann cells in the dorsal root ganglia (DRG) [8] and more recently PNS-derived fibroblasts which promote neurite outgrowth in adult dorsal root ganglion (DRG) neurons [9]. Many studies of the effect of photons either as laser irradiation (LI) or from light-emitting diodes (LED) on fibroblasts demonstrate multiple effects. These include activation and transformation relevant to the above effects [10,11,12] with both Schwann cells and fibroblasts being now recognised as being essential for nerve function in development, maintenance, disease and pain. The role of these cells will now need to be included in understanding PBM effectiveness.
2. The importance of the Peripheral Nervous System Organization
The PNS is generally considered to be made up of somatosensory first order neurons however peripheral nerves include not only the afferent axons transmitting sensory information to the CNS but also motor axons arising from ventral horn cells of the spinal cord and sympathetic axons. The sensory nerve cell bodies lie within the DRG and PNS-related ganglia such as the trigeminal nerve ganglia in humans are of particular interest in understanding how noxious and other PNS stimuli are transmitted and perceived as pain.
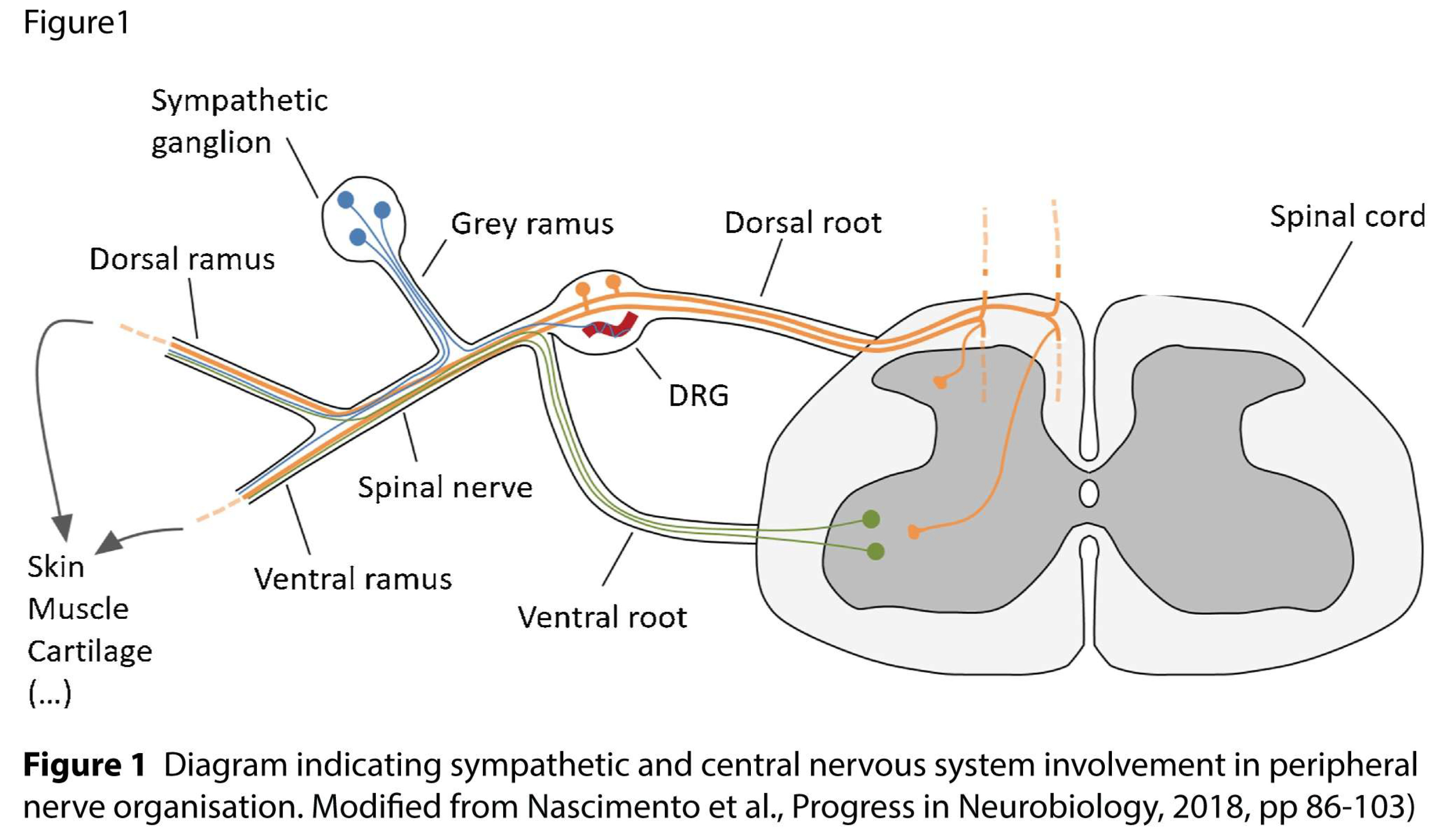
2.1. Dorsal Root Ganglia
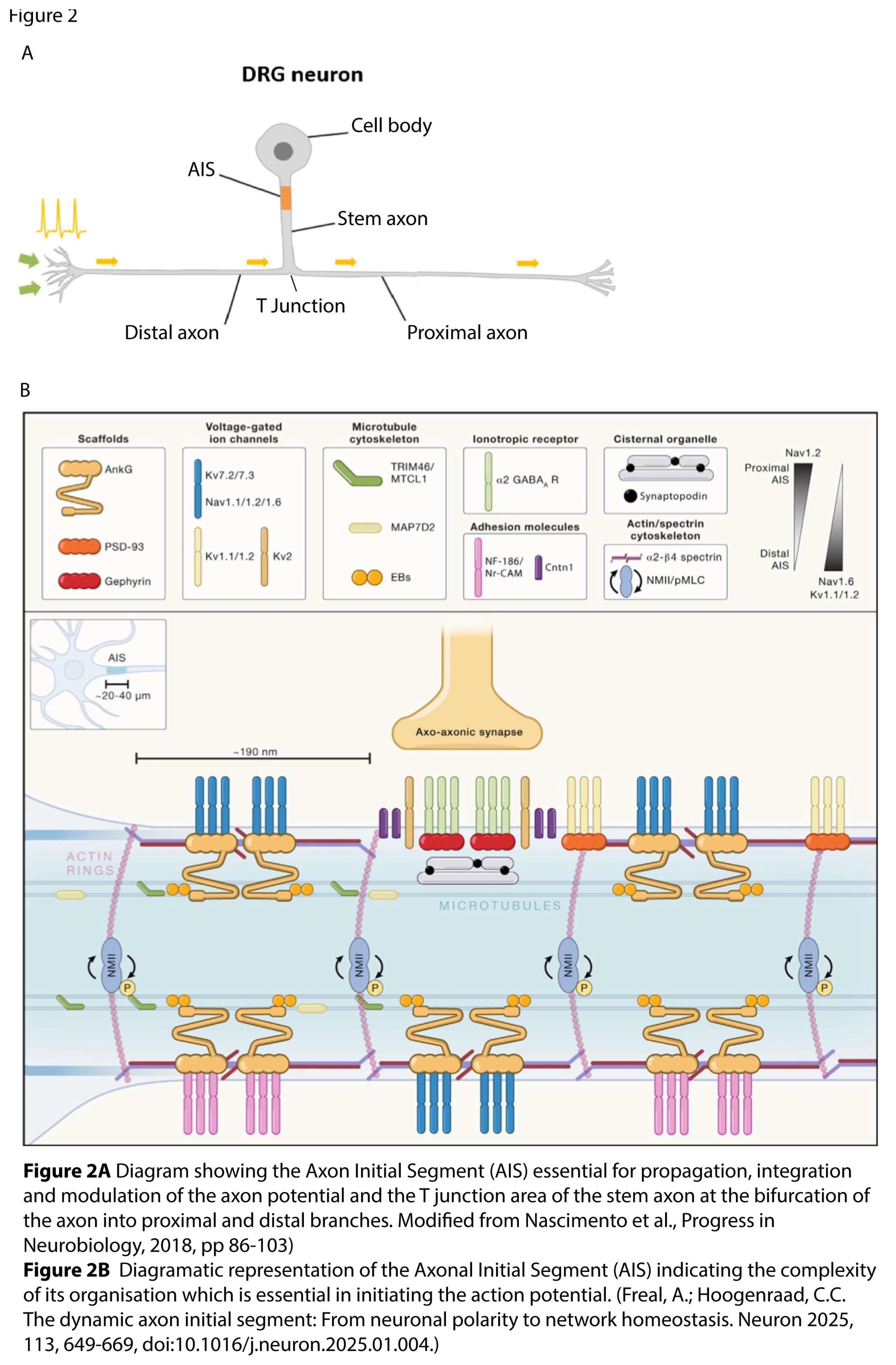
During human development the DRG neuronal cell body initially develops a pseudo-unipolar axon. Interestingly this was first described by Cajal [13] and occurs in unmyelinated C, Aδ and Aβ myelinated neurons. The initial area of the axon arising from the cell body, the ‘stem axon’ or ‘axon initial segment’ (AIS) with a specialized T junction where the axon bifurcates to form the proximal and distal arms. This T junction zone is now recognised as playing a major role in the propagation, integration and modulation of the action potential [6,7,14,15,16].
The distal axon in humans the such as those innervating the legs can be up to a meter long, or as in blue whales, 30 meters [17] further emphasizing the importance of understanding the role of the T junction. This development and specialization of the neuronal cell body and its axonal specialization is dependent on the associated Schwann cells and satellite cells [6] both of neural crest origin. While PNS and CNS axons are referred to as myelinated or unmyelinated this is based on examination at the light microscope level. Unmyelinated axons are ensheathed by uncompacted Schwann cells whereas myelinated axons are characterized by spiraling compacted ensheathment of individual Schwann cells along each axonal length.
2.2. The Schwann Cell
While PNS and CNS axons are referred to as myelinated or unmyelinated this is based on examination at the light microscope level. Unmyelinated axons are ensheathed by uncompacted Schwann cells whereas myelinated axons are characterized by spiraling compacted ensheathment of individual Schwann cells along each axonal length.
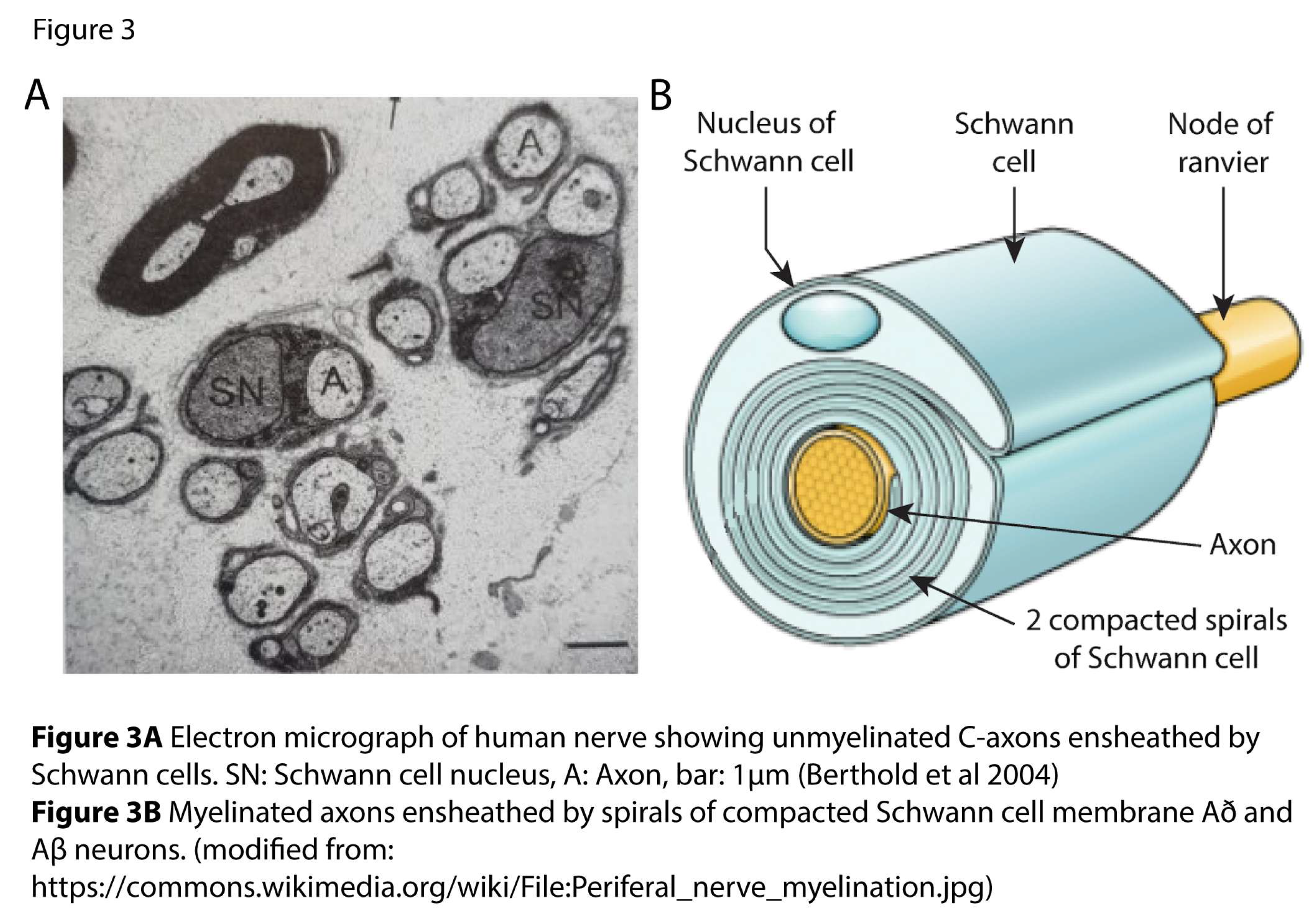
Considering the complex structure and function of DRG neuronal cells, it is more relevant to refer to such neurons as C-neurons or Aα and Aβ neurons rather than the common use of ‘fibre”. This nomenclature will therefore be used throughout, as this review emphasizes the importance of considering the role of the cell body as well as the axonal regions. Myelinated axons, the Að and Aβ neurons, are further characterized by nodes of Ranvier which develop where each Schwann cell abuts the other. The nodal and paranodal architecture is a complex arrangement of axon and Schwann cell molecules [18] and is essential for normal nerve conduction as its configuration affects nerve function, maintenance and response to injury or damage. In contrast the C neurons with their smaller diameter axons are unmyelinated but also ensheathed either individually or in groups within grooves of the Schwann cells. The number of Schwann cell spirals around each internode determines nerve cells as Aβ or Að neurons. The speed of conduction along myelinated axons depends on the number of compacted spirals of Schwann cell ensheathment with thinly myelinated Aβ neurons characterized by 100 or more spirals of compacted Schwann cells and conduction velocities of 30-120ms. Myelinated Aδ neurons have a conduction velocity of 4-30 Ms. These neurons respond to light touch and pressure while unmyelinated C neurons lying within Schwann cells grooves with conduction velocities of < 2.5 ms convey noxious sensation and thermoreception.
Within the relevant spinal cord segments, proximal axons of the first order neurons synapse with dendrites of the CNS second order afferent neurons within the dorsal horn of the spinal cord with their axons ascending to relevant neurons within the cortical centres of the brain [19]. Peripheral nerve axons unlike those of the CNS do not form dendrites despite this being frequently stated in peripheral nerve literature and diagrams. However their proximal axonal endings synapsing within the dorsal horn are MAP2 positive, characteristic of CNS axonal dendrites [20]. Again, of equal importance to neuronal function and generally overlooked is the recognition of the role of glial cells including not only SC but also the nerve cell body ensheathing satellite cells. Both are of neural crest origin and essential for normal nervous [8] system function but also in repair with a surprising proportion of SC precursors also able to transition to ‘neuronal’ fibroblasts. These remain within the nerve, providing extracellular matrix molecules, mainly collagen, that occupy the space among different axonal fibres [21]. Indeed, recent studies have discovered new functions for SC precursors as a class of multipotent progenitors, which can generate several different cell types [8,22,23]. This configuration too, is essential for normal function and in development, as well as maintenance and repair of the neuron. To complicate matters further, Abdo et al. have reported nociceptive SC which signal neuronal nociceptors in the upper dermal/epidermal region [24].
3. Discussion of the Cytoskeleton
3.1. Axon Initial Segment
The recent defining of the active role of the stem axon, the axon initial segment (AIS) and its bifurcation into distal (to skin and muscles) and proximal (to the spinal cord) axons of Aα, Aδ, and C neurons emphasizes the centrality of the cytoskeleton in neuronal structure and function [7]. Of particular note, the AIS is a highly specialized compartment in PNS and also CNS neurons, including somatosensory neurons, with their prominent nucleated cell bodies, axonal stem and long axonal processes [6]. The AIS is essential for two major functions: generating and modulating action potentials and maintaining neuron polarity [25]. Recent studies of the AIS junction report axonal microtubule organization varies between proximal and distal axons with the proximal axon having a higher density of microtubules than the peripheral axon. The AIS is also critical in regulating the transmission of bio-electrical signals from the neuron’s cell body to its axon in controlling fast and slow transport as well as the distribution of proteins, organelles [8]. It is also of particular interest in understanding the role of mitochondrial transport along the intra-axonal cytoskeleton of relevance to PBM/PT effectiveness. The stem axon, AIS and the bifurcation is also the central area for triphosphorylation of mitochondria which is reliant primarily on neuronal cell body metabolism. Axons in contrast to their neuronal cell bodies have relatively few ribosomes and other cytoplasmic components and are dependent on mitochondrial phosphorylation and the delivery of these ATP- rich mitochondria to the proximal and distal axons along the intra-axonal cytoskeleton to maintain both structure and function of the neuron.
Dephosphorylated mitochondria, other organelles including neuropeptides are transported retrogradely back to the cell body by dynein molecular motor proteins. Figure 4
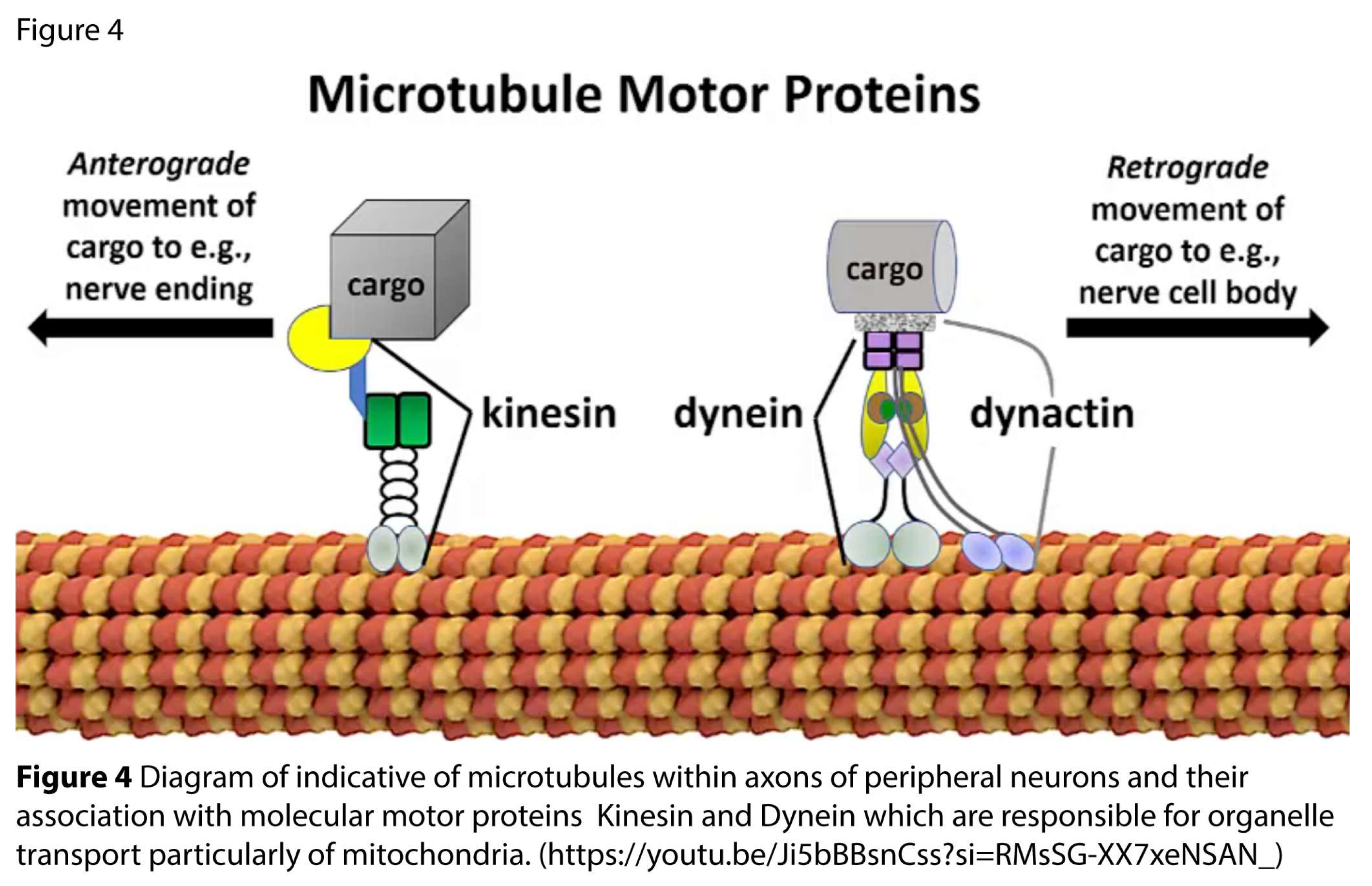
The cytoskeleton within the AIS undergoes rapid and extensive structural and functional changes in response to alterations in neuronal activity levels. Such plasticity has emerged as a key regulator of the unique neuronal morphology both within the ganglia and of the AIS and the T junction. The roles of the distal and proximal axons are further reflected in the unique organization of cytoskeletal intra-axonal microtubules. These highly dynamic microtubules composed of α and βtubulin dimers, undergo rapid de- and repolarization with a half-life of 10 minutes, an energy intense process [26]. They extend distally from the nucleus in the cell body to skin, receptors, muscles, and proximally to synapse with second-order neurons within the relevant spinal cord segment. Microtubules provide the infrastructure for fast axonal flow both anterograde and retrograde along the length of each axon from the cell body to the periphery or CNS synapse. Triphosphorylation of mitochondria occurs within the neuronal cell body area at the AIS, and these ATP-rich mitochondria are carried by kinesin molecular motor proteins ‘walking’ anterogradely along microtubules.
4.2. Fast Axonal Flow (FAF)
Microtubules provide the infrastructure for fast axonal flow both anterograde and retrograde along the length of each axon from the cell body to the periphery or CNS synapse. Triphosphorylation of mitochondria occurs within the neuronal cell body area at the AIS, and these ATP-rich mitochondria are carried by kinesin molecular motor proteins ‘walking’ anterogradely along microtubules.
(https://www.youtube.com/watch?v=y-uuk4Pr2i8 – Accessed 3-1-26). Dephosphorylated mitochondria, other organelles including neuropeptides are transported retrogradely back to the cell body by dynein molecular motor protein. This transport system is referred to as fast axonal flow (FAF) and is important not only for mitochondrial movement along the axon but is also equally important for the translocation of other signalling molecules from peripheral terminals to the cell body in the DRG. This includes receptors, transcription factors such as the neurotrophins, nerve growth factor (NGF) and brain-derived nerve growth factor (BDNF) associated with nerve injury and pro-inflammatory cytokines such as TNFα which are transported retrogradely to the cell body to initiate gene expression associated with pain modulation, nerve and tissue repair [27].
Understanding of the structural complexity of the AIS, its composition and function and its important role in preventing various neurological diseases is a recent finding. Although its molecular structure and organization is tightly regulated it remains vulnerable to neuronal damage and injury. Importantly, the AIS is able to respond to changes in synaptic input, neural activity, or injury resulting in the AnkyrinG-protein or ‘master organizer’ orchestrating rearrangement of the AIS membrane proteins and cytoskeleton [7]. This adaptive plasticity allows neurons to fine-tune their excitability and maintain proper excitation/inhibition (E/I) balance within neural networks. Due to this adaptive potential the AIS is proposed to be the central site of interaction with photons where PBM can modulate neuronal function.
4. What Is Photobiomodulation?
Photobiomodulation/therapy (PBM/PBMt) describes the interaction between photons and cellular activity to modulate cell physiology to achieve a therapeutic effect such as pain relief. PBMt has now replaced older synonyms such as low-level laser therapy (LLLT) or low-power laser therapy (LPLT) to encompass the nature of the device delivering light other than laser such as that from light-emitting diodes (LED) {Anders, 2015 #3553}. Although the term PBM is increasingly used there are significant differences between laser, producing coherent light and LED, the latter with a divergent beam, which potentially alter the therapeutic outcomes, even when delivering the same wavelengths. Figure 5.
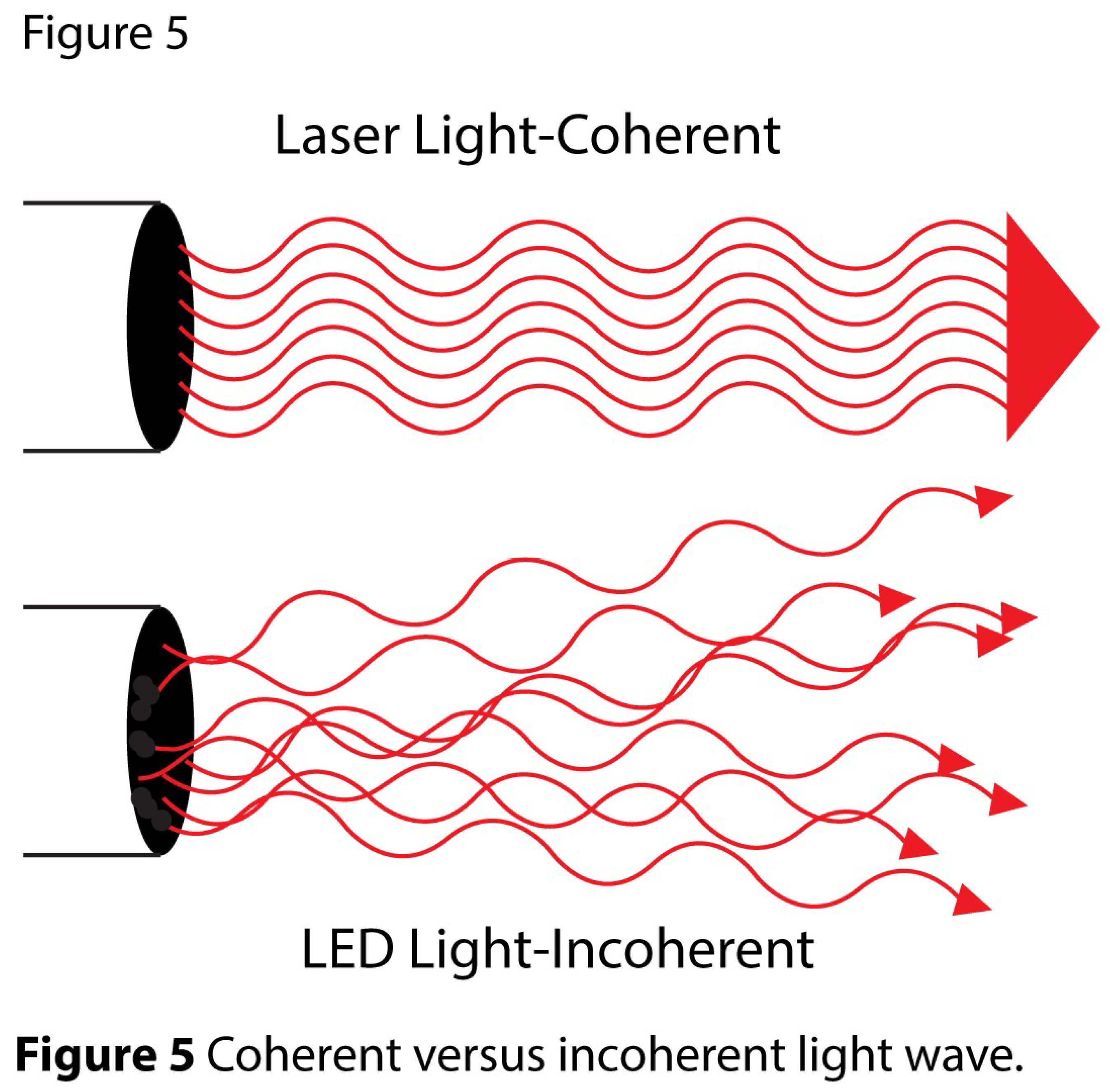
Laser photons are monochromatic, parallel in direction and coherent. In contrast, LED has a broader spectrum of wavelengths in its output, is non-coherent and is divergent. A full discussion of therapeutic differences is beyond the scope of this review, but it is generally accepted that laser penetrates more deeply than LED and will therefore have different therapeutic applications for deep anatomical structures such as facet joints while LED is suitable for superficial conditions such as eczema.
5. Experimental Studies of PBM
PBMt has been used pragmatically since the late 60’s initially in the clinical domains of wound healing, tissue repair and pain management, without a full understanding of the mechanisms. Effects on compact cells such as fibroblasts [29] and macrophages [30], lacking the axonal processes of neurons, and anti-inflammatory effects [31] were explored in some of the earliest studies of PBM. Understanding PBM effects on the peripheral nervous system and in particular nociceptors with reference to modulating pain has been a more recent development. With increasing knowledge of the morphology and function of somatosensory neurons, the involvement of light effects on nerves has now assumed more prominence in defining a neurophysiological mechanism for pain relief. Research has been directed to examining not only how an intact nerve responds to light of different wavelengths and how individual neurons also respond but how different anatomical segments of the neuron can be modified by photons.
5.1. DRG Studies
5.1.1. Human DRG Study—In Vivo
Illustrating this approach to defining PBM effects on different segments of somato-sensory neurons has led clinical researchers to target the cell body of neurons within DRG and to unravel the role of the cytoskeleton in modulating pain with PBM. The central role of the DRG as described earlier, as a therapeutic target for PBM-related pain modulation has been demonstrated in a human trial of low back pain [32] and a further two animal experiments which explore molecular changes in DRG and associated pain-behavior changes [33,34]. In the human trial Holanda et al. delivered laser directly to the DRG at the lumbar spine to patients with back pain, at the level of the second lumbar vertebra (L2) via a fibre-optic device. This was compared with lidocaine and radiofrequency ablation of L2 nerve roots, both standard interventional procedures in pain medicine. In the PBM treatment group, a continuous wave (cw), 808 nm wavelength diode laser (Photon Lase III1DCM, Brazil), with an output power of 100 mW was used in a single treatment. An 18-gauge needle was placed in the second lumbar intervertebral foramen with the fiberoptic laser device positioned a millimeter from the L2 DRG. Photons were delivered through a 600 mm optical fiber placed in the G18 needle. The tip of the fiber extended 5 mm beyond the tip of the needle in the second lumbar intervertebral foramen. The beam spot size was 0.003 cm2, irradiance 35W/cm2, exposure time 84 seconds, energy density 2800J/cm2, total energy was 8.4 J. The low back pain score was assessed by the visual analogue scale (VAS) and Pain Relief Scale (PRS) pre, post-procedure and in 1 month follow up. Remarkably, a single PBM session resulted in significant pain relief for all participants, with over 50% experiencing complete pain alleviation on the treatment day. Additionally, 1 month post-treatment, 70% of patients reported over 50% pain relief. These results support the clinical validity and feasibility of targeting the DRG though this technique requires highly specialized training and hospital facilities. Direct application of PBM to DRG in human subjects as demonstrated by Holanda et al. has led to Jenkins et al. to describe this technique as a new subcategory of PBMt, called transient small neural inhibition via PBM (tSNIP) [35] emphasizing the specific inhibitory effect of laser on C neurons.
5.1.2. Animal DRG Studies
Two animal experiments supported the findings of Holanda et al. demonstrating significant effects of laser on DRG showing reduction in both pain-related behaviors and molecular markers. In the first experiment, Chen et al. studied the effects of LI with wavelength of transcutaneously delivered 808± 5 nm, power 300 mW, cw on DRG neurons in the lumbar spine in a chronic compression of the dorsal root ganglia (CCD) model [33]. This is a validated model used to study neural responses and pain-related behaviors. CCD rats stainless-steel rods inserted into the fourth and fifth lumbar intervertebral foramen, and the rats were then subjected to 0 or 8 J/cm2 LI for 8 consecutive days following CCD surgery. Pain and heat stimuli were used to test for hyperalgesia following CCD. The levels of TNF-a, IL-1b and growth-associated protein- 43 (GAP-43) messenger RNA (mRNA) expression were measured via real-time PCR, and protein expression levels were analysed using immunohistochemical analyses. LI significantly decreased sensitivity to pain and heat stimuli in the CCD groups. The expression of the pro-inflammatory cytokines TNF-a and IL-1b were increased following CCD, but were reduced by LI. Furthermore, the expression of GAP- 43 was enhanced by LI. GAP-43 is a neuronal repair gene that localizes exclusively to nerve fibers and is present in regenerating peripheral nerves suggesting that not only is pain and destructive neuroinflammation controlled by LI, but neuronal repair and regeneration is also promoted. The second study by De Sousa et al. examined the pain threshold in the right hind-paw of mice, after PBM (810 nm, cw laser, for 120s, spot size 1 cm2 or 6 cm2, energy density of 1.2–36 J/cm2, power density: 50mW/cm2) was applied to various anatomical locations [34]. The pain threshold, measured with von Frey filaments, was increased more than 3-fold by PBM to the lower back overlying the DRG. PBM to other neural structures along the same spinal pathway such as the head, neck and ipsilateral (right) paw also produced an analgesic effect but the optimal effect was that to the DRG. No effect occurred when LI was applied to areas outside the nerve root distribution. The optimal effect occurred 2–3 hours post-PBM and disappeared by 24 hours. Seven daily irradiations showed no development of tolerance. Type 1 metabotropic glutamate receptors decreased, and prostatic acid phosphatase and tubulin-positive varicosities were increased as shown by immunofluorescence of DRG samples.
5.1.3. Studies of DRG Neuron Cultures
The above studies of LI delivered directly or transcutaneously to DRG illustrating the change in pain measures (in humans) and pain behaviors (in animals) along with significant changes in biochemical markers highlight the importance of the DRG as a target for PBM with the central role of the AIS and stem axon as a critical neuronal regulator of the whole neuron metabolism and function. Few studies, however, have investigated structural changes in the cytoskeleton of neurons which occur following LI which provide a basis for understanding the analgesic response and changes in biochemical markers to support these effects. One of the earliest studies to demonstrate the effects of LI on the cytoskeleton in neurons was by Chen et al. in 1993. Chen used 830nm, 20mW, cw laser at doses of either 6J over 5 minutes or 15J to isolated mouse DRG neurons for 15 minutes demonstrating “beading” or varicosity formation along the length of the neurons [36]. Varicosity formation reflects a depolymerization of microtubule dimers with disruption of cytoskeletal structure as well as function. Most relevant for the pain-relieving effects of laser was the decrease in the number of neurons containing substance P and calcitonin-gene related peptide (CRGP), which occur only in neuron with the characteristics of nociceptors.
In a similar vein, in a series of studies between 1990 and 1994, Wesselmann et al. used Nd: YAG laser (1064nm) to demonstrate multiple effects on cytoskeletal morphology and function as well as modulating action potentials amplitudes in in vitro rat nerve. [37,38,39,40,41,42,43]. Disruption of fast axonal flow (FAF) was seen with an experimental model in which horseradish peroxidase (HRP) was applied to the proximal end of severed rat tibial nerve which had been irradiated seven days earlier and changes in HRP density in the dorsal root ganglia observed [41]. HRP is a cell marker which can be used to retrogradely label sensory neurons and is a marker of axonal transport. In nerves exposed to laser, there was a selective decrease in the number of small dorsal root ganglion cells labelled with HRP compared with no change in larger sensory neurons or motor neurons [44]. These data demonstrate that 1064nm LI selectively disrupted retrograde flow in Aδ and C nociceptors. Additionally, Wesselmann also demonstrated a preferential impairment of action potentials in small, slowly conducting sensory nerves compared to larger afferent nerves [39]. Importantly these studies by Chen and Wesselmann demonstrate the selective nature of laser photons of two different near infra-red (NIR) wavelengths on small diameter nociceptors, with implications for analgesic effects.
5.1.4. Microtubule Depolymerization and Mitochondrial Membrane Potential (MMP)
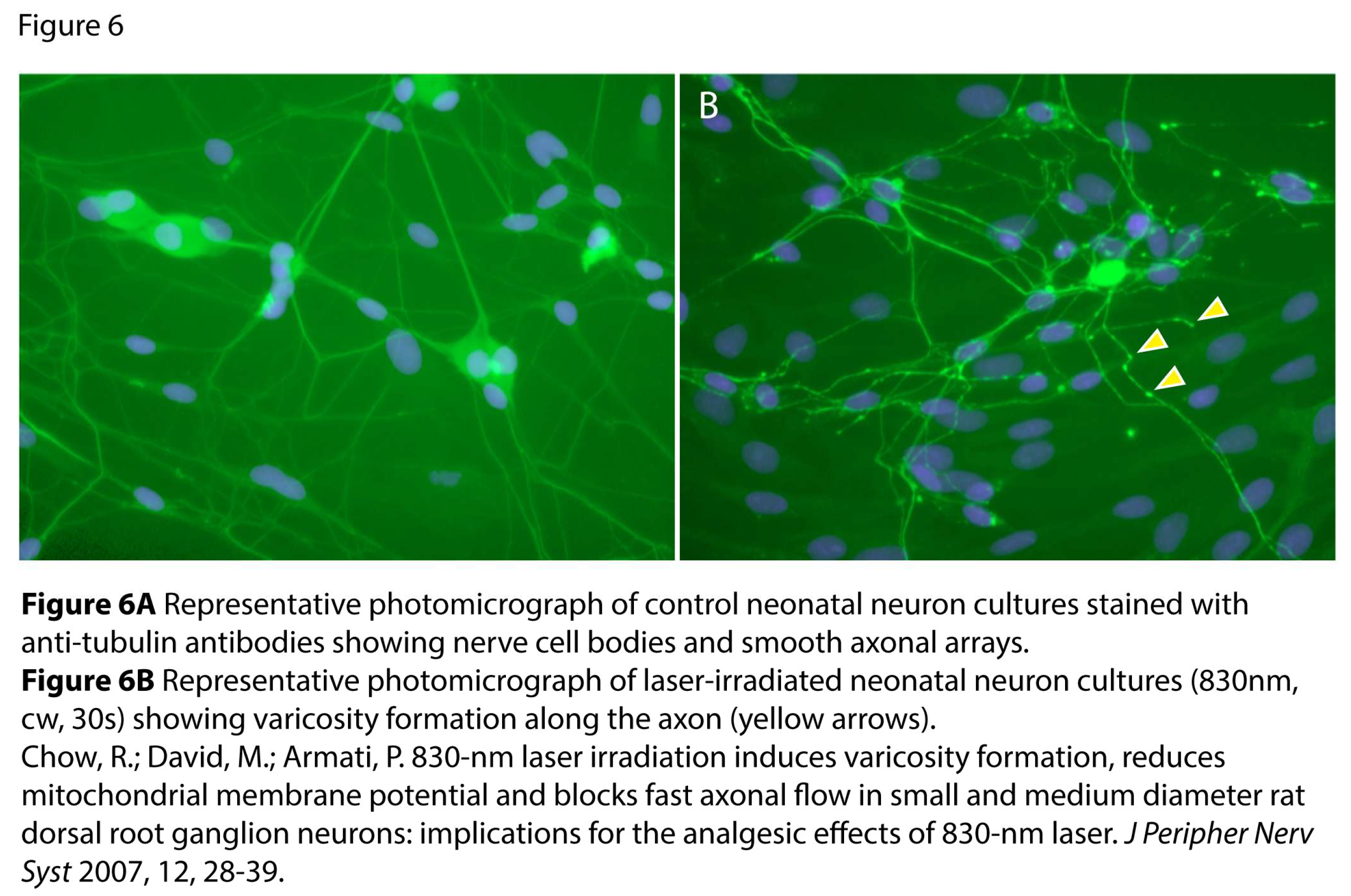
To further understand the mechanism of action of the therapeutic effects of PBM in modulating pain as in the previous studies we examined structural changes occurring in the microtubules of the cytoskeleton using rat DRG neuron cultures at wavelengths of 650, 808, 830 LI. In 2007 we used a cell culture model of neonatal rat DRG neurons to examine PBM effects of 830nm LI using live imaging and immunohistochemistry. We demonstrated reversible disruption to the cytoskeleton with varicosity formation representing depolymerization of α and β tubulin dimers, the constituents of microtubule subunits, using 830nm, 300mW, cw laser for 30s, 9J, PD: 300mW/cm2, ED 8.3J/cm2 stained with anti-β-tubulin antibodies specific for neurons [45]. Observation over 24 hours showed reversal of “varicosities”. Figure 6a and Figure 6b
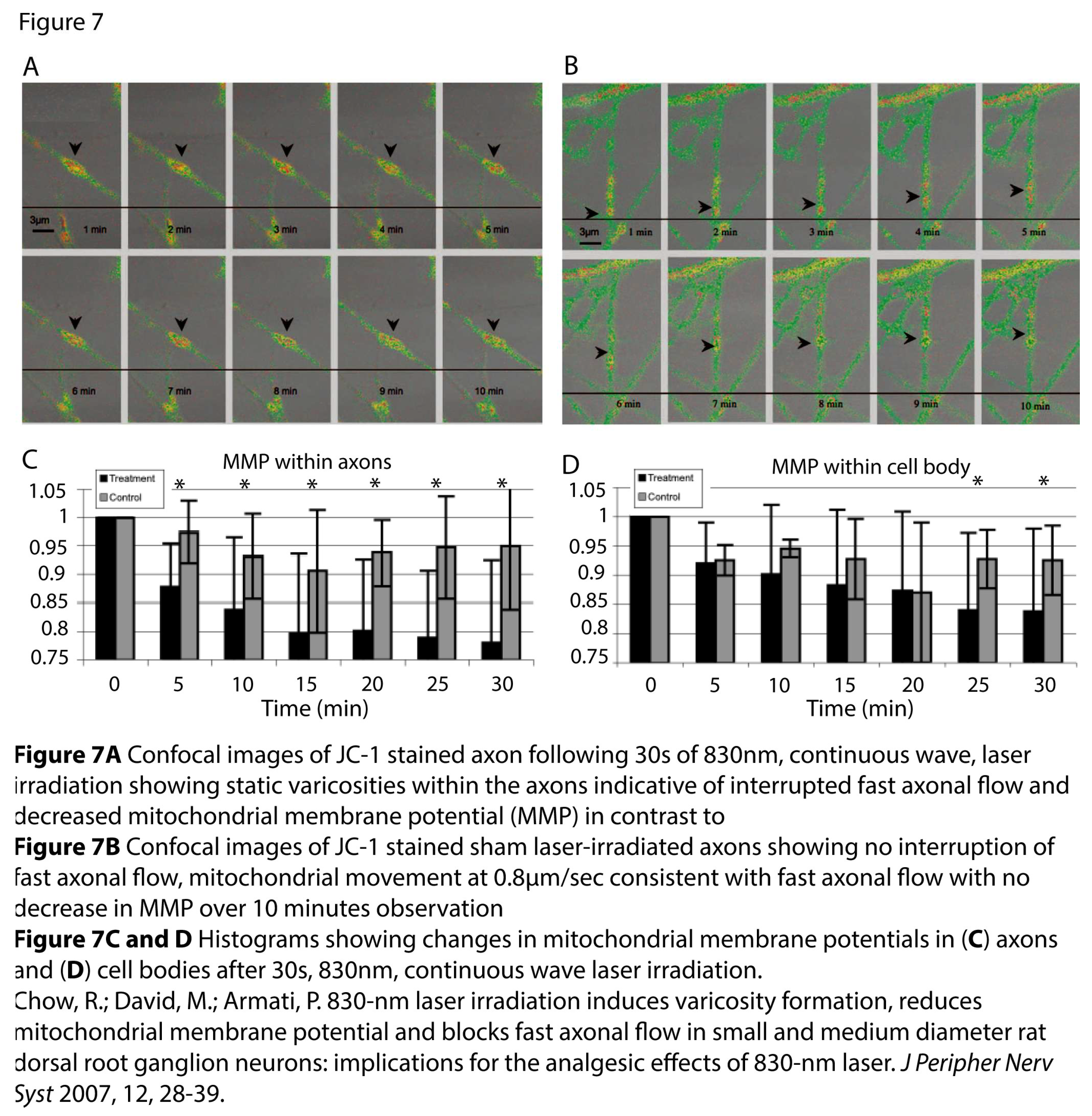
In addition, live-imaging using confocal microscopy demonstrated the cessation of movement of mitochondria along the microtubules with mitochondria “piling up” in varicosities. Simultaneously the mitochondrial membrane potential (MMP) of the mitochondria showed a decline from high to low energy reflecting a depletion of ATP within the mitochondria. (A) Real time confocal images of a JC-1 stained axon after 30s of 830nm, cw, laser irradiation showing static varicosity indicating blockade of FAF and decrease in MMP over 10 mins of observation. (B) Real-time confocal images of JC-1 stained, non-laser irradiated control axons showing movement of mitochondria at 0.8µm/s, consistent with FAF and no decrease in MMP over 10 min of observation. Figure 7.
In line with her pilot study of PBM to DRG to relieve back pain, Holanda et al. also used 808nm, 960mW, cw, PD: 300mW/cm2 at exposures times of 2, 5, 30, 60 and 120s in a culture model of rat DRG neuron and an in vivo spared nerve injury (SNI) model of neuropathic pain to examine effects on the cytoskeleton as well as functional outcomes [32]. Using similar methodology including immunohistochemical staining with anti-β tubulin antibody they replicated the same features as varicosity formation primarily in small diameter neurites (<30µm), identifying them as nociceptors and only at duration exposures of >120s in larger diameter neurites. Consistent with this finding they also demonstrated functional changes in a neuropathic pain model where they tested cold and heat allodynia using Von Freys hairs, showing an improvement with laser compared with control (no LI). Mechanical allodynia was unchanged.
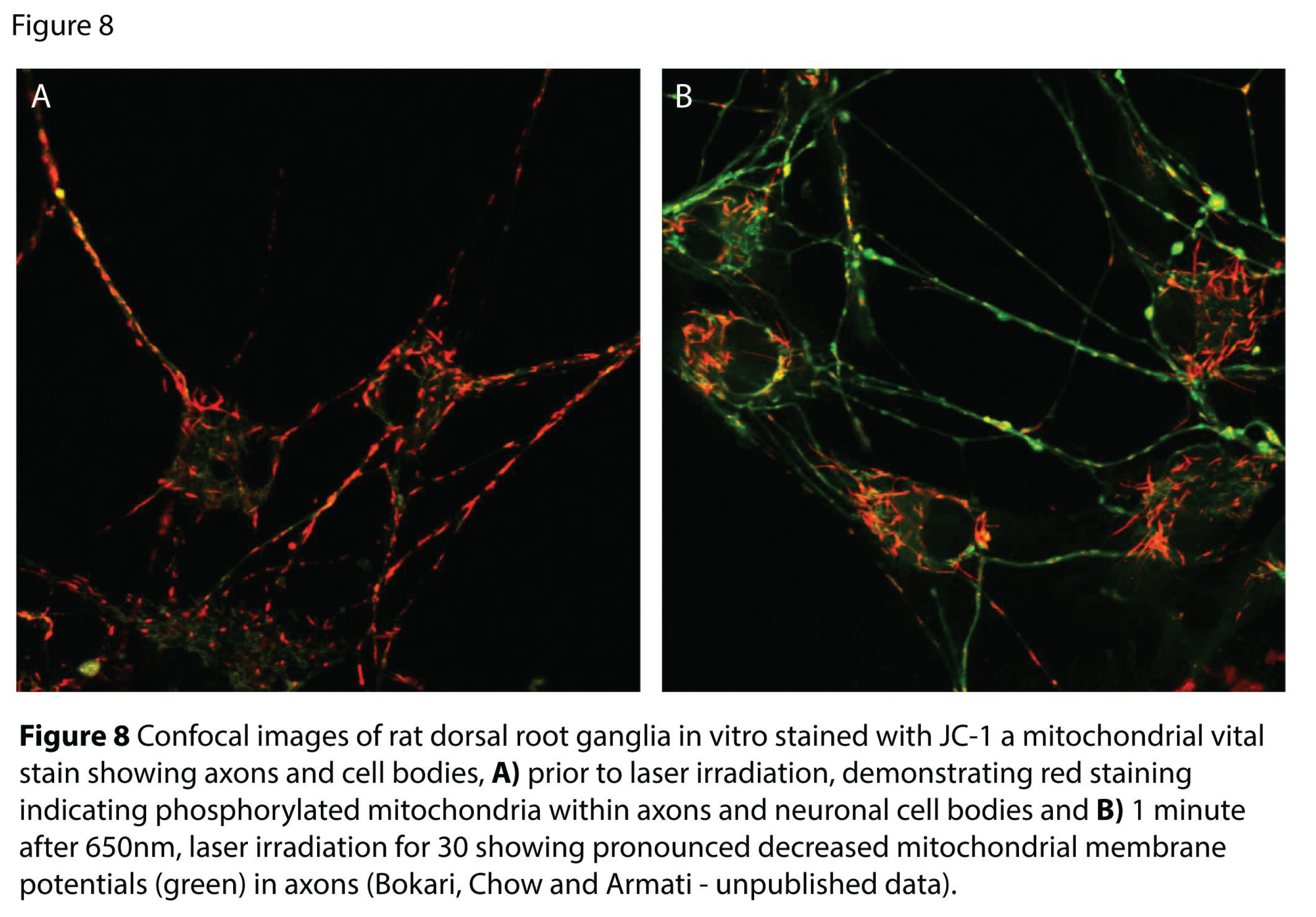
Similarly we also demonstrated similar morphological and functional changes in the microtubules with reversible varicosity formation following 650nm laser as seen with 830nm laser, using the same cultured neonatal DRG neuron model. Importantly at the beginning of this study and extending the understanding of the translational perspective of these findings, TRPV-1 fluorescence was also examined in the same series of experiments. This confirmed that the DRG neurons in culture were predominantly small and medium diameter neurons consistent with their nociceptive-specific function. No differences in fluorescence intensity between TRPV-1 antibody labelled irradiated neurons and control neurons were observed. The effects of 650nm laser for 15 secs (ED: 5.1 J/cm2), 30 secs (10.2 J/cm2) or 60 secs (20.4 J/cm2) to DRG neurons were examined commencing 3 minutes after laser irradiation, over a 10-minute period. No significant change was observed in the MMP (ψmm) along axons irradiated for 15secs compared to non-irradiated neurons, however, MMP of DRG neurons irradiated for 30 secs resulted in lower mean MMP values (low energy seen as green in Figure 8) in comparison to non-irradiated neurons (high energy seen as red) which significantly decreased by the 8th minute (p<0.05, P value = 0.04). MMP along axons of rat DRG neurons irradiated for 60 secs resulted in significantly lower mean MMP values throughout the ten-minute imaging period in comparison to nonirradiated neurons. Figure 8a and Figure 8b
In this series of experiments, not only was the MMP of axons evaluated, but also the MMP in axon hillocks were examined with no change observed in the MMP of the axon hillock following either 15 secs or 30 secs irradiation. Sham LI cultures showed no significant changes in MMP of cell bodies, axonal hillocks or axons.
As with our studies of 830nm laser we also investigated changes in FAF of mitochondria following 650nm laser. Both 30secs and 60secs of 650nm laser significantly decreased FAF in irradiated neurons compared to non-irradiated neurons (P value: <0.0001). FAF was significantly slowed below the normal range (0.5-2μm/sec) for both irradiated groups. The average FAF was 0.17μm/sec for mitochondria irradiated for 30 secs and imaged three minutes PI. This rate had increased to 0.27μm/sec 24 hrs. after irradiation. The average FAF was 0.07μm/sec for mitochondria irradiated for 60 secs and imaged three minutes post-irradiation (PI). This rate increased slightly to 0.11μm/sec 24 hours after irradiation. In non-irradiated neurons, the average FAF rate was 0.50μm/sec. Average FAF of mitochondria irradiated for 30 secs or 60 secs and imaged 3 minutes later plotted over the 10-minute imaging period did not increase significantly. The same was observed for values plotted 24hrs post irradiation. However, for neurons irradiated three minutes and 24hrs PI showed a decreasing trend in FAF in irradiated neurons compared to non-irradiated neurons.For the first time, this study shows that a single dose of 650nm LI at EDs ranging from 5.25 - 21 J/cm2 caused the formation of axonal varicosities. These varicosities and mitochondrial clusters were observed in neurons irradiated for all LI durations (ED: 5.25, 10.5 and 21J/cm2) and fixed 1hr PI. There was no difference in the number of varicosities although the varicosities and their mitochondrial clusters were still present 24hrs PI for all LI durations. The repeated dose of LI 24hrs apart did not increase or significantly affect the varicosities in irradiated neurons irrespective of end time points. Pooled average FAF of mitochondria in neurons irradiated for 30secs & 60secs and imaged three minutes and 24 hours PI. Neurons irradiated for 30secs & 60secs and imaged three minutes PI was significantly lower (p<0.05) than non-irradiated neurons (P values: <0.0001). FAF of mitochondria 24hrs after irradiation showed a lesser significance between non-irradiated and 30secs irradiated neurons (P value: 0.011) compared to 60secs irradiated neurons (P value: <0.0001). This suggests an initial recovery of FAF following 30secs LI.
6. Depolarisation Blockade
Studies using DRG neuron cultures illustrate reversible microtubule disruption which occurs with several wavelengths from red to infrared. Importantly these effects occur in small diameter nociceptors indicating that PBM is selective for unmyelinated C neurons. Critically, microtubule depolymerization initiates a cascade of further disruption to nerve function, particularly to ATPase requiring enzymes such as Na2+K+ATPase, a voltage-gated sodium channel setting the baseline for generation of action potentials, causing depolarisation blockade. The implication for this selective effect is that PBM acts directly on nociceptors to block action potential generation underpinning its use therapeutic application as a treatment for acute and chronic pain. As described previously, the neuronal cytoskeleton, in particular the microtubules and the associated molecular motors, provide the infrastructure for the maintenance of structure and function of anterograde fast axonal flow, which delivers ATP-rich mitochondria for all nerve functions, including bioelectric signalling. The Na2+K+ATPase pump maintains an action potential threshold for both somatosensory and motor neurons and is initiated by depolarisation of resting potentials.
Disruption to the Na2+K+ATPase pump has been demonstrated by Kudoh et al. using a rat saphenous nerve in which 830nm, cw, 60mW laser altered Na2+K+ATPase activity in a biphasic manner with suppression at high doses, leading to depolarisation blockade [47]. This may provide a mechanism for the reduction in action potential amplitude and slowed nerve conduction leading to analgesic/anaesthetic effects of PBM seen experimentally and clinically. Many other neuronal functions require ATP and the disruption to Na2+K+ATPase is only one of other possibilities where disruption could occur.
7. PBM and Modulation of Action Potentials
When a nociceptive stimulus occurs, either through illness or injury an action potential is initiated and transmitted via Aδ and C somatosensory neurons which synapse with various laminae in the spinal cord. Second and third order neurons then convey that signal to the pain matrix where it is perceived as pain [48]. The function of nociceptive nerves and the generation of a pain signal is dependent on the integrity of the microtubular infrastructure of the cytoskeleton. Given that we have demonstrated that PBM at wavelengths of 650nm, 830nm, 808nm and 1064nm can disrupt the cytoskeleton we propose that one of the mechanisms for the pain modulating effects of PBM seen clinically is the inhibition of action potential generation due to this disruption [49,50].
7.1. PBM and Effects on Action Potentials
Further exploring the laser-induced slowing of action potentials is a review which identified clearly the action of multiple wavelengths in both human and animal models [51].
Using the same wavelengths as with our DRG neuronal cultures, Yan et al. in a series of experiments using an in vivo sciatic nerve model demonstrated that laser irradiation at 650nm and 808 nm slowed conduction velocity (increased latency) and amplitude of somatosensory evoked potentials (SSEPs) and compound muscle action potentials (CMAPs) when applied transcutaneously at four sites along the course of the sciatic nerve. This occurred within 10 minutes of application and for the 650nm wavelength a small but significant effect persisted at 24 hours but had returned to normal by 48 hours [49]. This study was not able to identify a specific, selective effect on C neuron conduction described in other animal studies such as that by Wesselmann who demonstrated selective slowing of conduction velocity, following 1064nm in C nociceptors.
7.2. PBM-Induced Neural Blockade in Human Studies
In human subjects reduction in amplitude of action potentials and/or increase in latency has been described in studies of transcutaneous application over median nerve [52,53,54,55,56,57,58], superficial radial [59], sural nerve [60,61] including LED [62] and in one study of Trigeminal Nerve [63].
More recent studies have also demonstrated similar reductions in action potential and/or latency in ulnar nerve in human subjects [64,65]. Not all studies demonstrate inhibitory effects, particularly when LED is used [66], however, there is strong evidence that red to infra-red laser wavelengths can slow nerve conduction velocity and reduce action potential amplitude in a dose-dependent manner, when administered transcutaneously. Selective inhibition of C neurons was not identified in these studies.
7.2. PBM-Induced Neural Blockade in Animal Studies
A series of animal experiments in which LI was applied either transcutaneously or directly to nerve demonstrated not only reduction in action potentials and selective nociceptor blockade but also inhibition of pro-inflammatory evoked potentials. Noxiously evoked potentials elicited by mechanical [67], thermal [68,69,70] or chemical stimulation, such as formalin, turpentine, or bradykinin injection [71,72,73,74] were reduced or completely inhibited by LI to the exposed nerve at various points along the trunk. Studies of LI on experimental models of guinea pig, rat, mouse, cat, ferret, rabbit, or dog nerve were evaluated using varying methodologies including electrically evoked CAPs [72,75,76,77,78,79], SSEPs [80], with the inhibition of the amplitude of action potentials being the dominant finding.
A small number of studies in animal nerve-spinal cord models demonstrated inhibitory effects on evoked potentials within DRG or spinal cord following laser to peripheral nerve delivered either directly or transcutaneously. Kono identified reduction in spinal cord evoked potentials in cats following direct He-Ne laser, 1mW to sural nerve [75]. Wakabayashi et al. demonstrated that GaAlAs (830 nm, 350 mW) LI of the cervical surface of a tooth suppressed late discharges in responses evoked in wide-dynamic range neurons (WDR) of the trigeminal subnucleus caudal (TSC) neurons by tooth pulp stimulation but did not affect the early discharges of the TSC [71].
The early discharges in tooth pulp-evoked response of TSC neuron were electrophysiologically defined as spikes induced by inputs from Aδ afferent neurons according to the first spike latencies, and the late discharges were assumed to be evoked by excitatory inputs from C neurons [71].
Similarly, Shimoyama et al. demonstrated that prolonged (30 min) irradiation of the skin overlying the hind paw with He-Ne laser (8.5 mW, 632.8 nm) laser resulted in the attenuation of the formalin-induced discharge activities of the dorsal horn neurons at L1 – L3 in anesthetized rats [81]. More recently, the effects of laser irradiation (808nm, 750mW, 180s) of exposed sciatic nerve on the neuronal firing in lamina II of the spinal dorsal horn of a rat evoked by mechanical stimulation with von Frey filaments were examined [82]. The evoked potentials from nociceptors were inhibited from 5 minutes after laser irradiation and persisted for 3 hours. Sham irradiation did not alter the firing frequency. Histopathological evaluation revealed no damage to the sciatic nerve due to laser irradiation.
Buzza et al. 2024 have also demonstrated selective C neuron inhibition using 808nm 60mW, 4 minutes application in an exposed sciatic nerve model [83]. The rationale for defining tSNIP through its specific therapeutic effect was to distinguish it from general PBM applications, to reduce confusion regarding mechanisms and to promote standardization in this specialized area. Studies by Buzza using exposed nerve replicate effects seen in the series of animal experiments described earlier but the extrapolation to practical applications remains limited clinically. Importantly, Holanda et al. has implemented a clinical protocol using direct laser stimulation of DRG in patients suffering from back pain as described previously.
8. Clinical Application of PBM
8.1. PBM in Medical Conditions
Although the study by Holanda et al. provides a new model for clinical application delivering laser via fibreoptics directly to the DRG of patients with back pain it remains in the realm of highly specialised techniques. From the earliest days of clinical application of PBM LI and more latterly LED irradiation have been applied transcutaneously to achieve therapeutic effects in both nociceptive and neuropathic pain conditions.
Non-specific neck pain is one of the areas in which PBM has demonstrated effectiveness. Chow et al. treated 140 patients with non-specific neck pain using transcutaneously delivered 830nm, 300mW continuous wave (cw) laser, 9J per point at 20.1J/cm2 showing significant benefit in pain and quality of life [84]. A systematic review of PBM in the treatment of neck pain which included a total of 820 patients, established benefit for up to 22 weeks. Wavelengths used included 632.8nm, 780 nm, 820-830nm and 904 nm and doses used varied between 1-5.9/point with treatment times varying between 30s to 4 minutes. We propose that in the cervical spine the medial branch of the dorsal root lying over and supplying the facet joints is within the penetration depths of near-infrared lasers (~3cm) and that the medical procedure of facet joint blockade with local anaesthetics can be achieved at least partially by transcutaneous application of laser irradiation at the appropriate doses resulting in depolarisation blockade in these nerves. Other clinical areas where systematic reviews of clinical applications of PBM have also demonstrated favourable outcomes include knee osteoarthritis [85], diabetic peripheral neuropathy [86], shoulder tendinopathy[87,88], neuropathic pain [89], chemotherapy related neuropathy[90] and musculoskeletal pain [91]. Other clinical trials and reviews have reported favourable outcomes in PBM delivered transcutaneously in back pain [92,93], trigeminal neuralgia [94], post-herpetic neuralgia [95], Fibromyalgia [96] and crystalline arthropathies [97].
Other clinical areas where systematic reviews of clinical applications of PBM have also demonstrated favourable outcomes include knee osteoarthritis {Stausholm, 2019 #3967}, diabetic peripheral neuropathy {Korada, 2023 #4790}, shoulder tendinopathy{Haslerud, 2015 #4797;de Lara Quagliotto, 2025 #4783}, neuropathic pain {de Andrade, 2016 #4802}, chemotherapy related neuropathy{Argenta, 2017 #2440} and musculoskeletal pain {Clijsen, 2017 #4803}. Other clinical trials have reported favourable outcomes in PBM delivered transcutaneously in back pain, trigeminal neuralgia, post-herpetic neuralgia, fibromyalgia and crystalline arthropathies {Masoumipoor, 2014 #2682}.
8.2. PBM in Dentistry
Use of PBM routinely in dentistry is also increasing with both experimental and clinical trials showing PBM effectiveness for acute and chronic pain relief/analgesia and pre-emptive anaesthesia. Chan et al. conducted a double-blind, randomized, clinical trial and investigated the effectiveness and underlying mechanism of neural inhibition of pulsed Nd:YAG laser (1064nm) induction of pulpal analgesia compared with 5% EMLA anaesthetic cream. Forty-four paired premolars from 44 orthodontic patients requiring bilateral premolar extraction from either dental arch were randomly assigned to the ‘Laser plus Sham-EMLA’ or ‘EMLA plus Sham-Laser treatment group [50]. Analgesia was tested by an Electric Pulp Tester (EPT)and the cutting of a standardized cavity, which was terminated when participants reported sensitivity. Visual Analogue Scale (VAS) and numbness were recorded. This trial confirmed that a pulsed Nd:YAG laser at the specified parameters effectively induced pulpal analgesia, by suppression of the intradental nerve responses to both electrical and mechanical stimuli and provides an innovative, painless, non-invasive alternative to drug-induced analgesia for restorative dental procedures. Prior animal studies reflecting dental application found similar findings of reduced axon potentials in different experimental models. One study by Orchardson et al. aimed to simulate the effects of lasing dentine on pulpal nerve function [98].
The methodology involved threading rat spinal nerve roots through the prepared pulp canal of a 10 mm long tooth root segment which was mounted in a perspex bath. The protruding ends of the nerve were placed on platinum wire electrodes used to elicit and to record compound nerve action potentials (CAPS). Laser energy (average power = 0.3-3.0 W) was applied to the surface of the root segment using a pulsed Nd:YAG dental laser (dLase 300). When the laser probe tip was placed in static contact with the tooth surface, the nerve CAP was irreversibly abolished within 60 s of lasing at 1.0 - 3.0 W power. When the laser tip was moved to and from over the root surface in a scanning mode, similar levels of radiation produced less marked effects. In the latter mode, CAP attenuation increased with increasing power and duration of lasing. After 60 s lasing at 0.3 W, the CAP size was 95% ( 5, S.D.) of the pre-lasing controls value; with 2.0 W the CAP was reduced to 54% . The CAP recovered to 90% of control levels after lasing at powers up to 1.5 W but reached only 72% of control values after lasing at 2.0 W power. This study demonstrated that laser radiation applied to dentine caused a dose-dependent block of action potential conduction in nerve fibres in the underlying pulp chamber. It also emphasised the importance of an optimal dose of the Nd:YAG laser to achieve reversible action potential blockade without damage to the nerve. Wakabayshi et al. also demonstrated a similar inhibitory effect using gallium-aluminum-arsenide (GaAlAs) 830nm, 350mW, CW laser on responses evoked in trigeminal subnucleus caudal neurons by tooth pulp stimulation in anesthetized Wistar rats. The pulp of the lower incisor was electrically stimulated, and the evoked action potentials were extracellularly recorded in the ipsilateral caudal neurons. The laser beam was applied on the cervical surface of the stimulated incisor. The rate of firing discharges and the numbers of spikes evoked in the caudal neurons were compared before and after laser irradiation. Laser irradiation suppressed the late discharges in the response of the caudal neurons which were evoked by excitatory inputs from C neurons but did not suppress the early discharges evoked by inputs from Aδ afferents. This indicates that laser irradiation at these parameters for 120 s through the tooth structures inhibited the excitation of unmyelinated fibers of the pulp without affecting fine myelinated fibers. Authors concluded that low power LI suppressed action potentials by depolarization blockade of C neurons. Both the Orchardson and Wakabayahsi studies provided a validated basis for the later studies of Chan et al. There are also systematic reviews of PBM in dental applications including, temporomandibular joint disorders [99,100,101], endodontics [102], burning mouth syndrome [103] and orthodontic pain [104].
9. Discussion
The translational link between the critical role of the cytoskeleton and photonic disruption of intra-axonal microtubule dimers resulting in a cascade of events leading to nerve conduction disruption, specifically in nociceptors, is established.
The blockade of FAF, which in turn stops transport of ATP-rich mitochondria via the motor proteins both kinein and dynein which prevents ‘recharging’/phosphorylation of axonal voltage gated ion channels, inactivating sodium channels has also been demonstrated [105]. This series of disruptions in the structure of neurons initiated by photon absorption results in slowing of nerve conduction, selective for nociceptors, and light-induced depolarisation blockade leading clinically to pain modulation. From a translational perspective this supports the hypothesis of a neurophysiological mechanism of PBM in the treatment of pain.
Indeed, it is suggested that based on these effects PBM has the potential as a non-pharmacological therapy to assist patients in reducing opioid dependence and manage pain [106].
9.1. The Cytoskeleton and Pharmacological Agents
This putative mechanism of PBM aligns with the capability of certain drugs to disrupt the integrity the microtubules, including drugs such as Colchicine, in the treatment of gout or Vinca Alkaloids in cancer treatments. Such disruption has also been demonstrated with local anaesthetic agents whose mechanism of action results in part from their effect on cellular cytoskeletal systems [107,108,109]. The local anaesthetic lidocaine has been shown to specifically impair microtubule assembly in vitro by inhibiting polymerization of tubulin subunits [110] as well as kinesin-based axonal flow [111]. The specificity of this effect with local anesthetics and the rapidity of onset of their clinical effectiveness in relieving pain underpins a parallel process with photons. Understanding PBM in the context of these pharmacological agents provides further insight into the clinical application of PBM.
9.2. Neuroplasticity and PBM
PBM therapy has been used clinically for the treatment of pain for more than four decades, including neck pain [112], back pain [93,113], knee osteoarthritis [85] and tendinopathies [114]. Understanding the mechanism of the clinically significant pain modulating effects of PBMt has focused largely on anti-inflammatory studies or on wound healing and tissue repair. It is important, however, to differentiate the direct, local anti-inflammatory effects of PBM which are relevant to the management of acute pain [115] from neurophysiological effects mediated directly by changes in the cytoskeleton and disruption to FAF.
In the context of anti-inflammatory effects many studies document PBM effects on specific cell types such as mast cells, macrophages, fibroblasts and neutrophils. A small number of studies have addressed the potential for a neurophysiological basis but have not been rigorously pursued. As pain is ultimately a response initiated by the activation of nociceptors, Aδ and C neurons of the somatosensory nervous system by noxious stimuli, examining LI effects on these nerves is a logical step. A greater understanding of the complexity of somatosensory nerves, the multiple feedback loops from afferent stimulation of the PNS to the spinal cord and the “Pain Matrix” involving multiple centres in the CNS leading to pain perception [48,116], have led to a more nuanced understanding of how photons may affect nerve activity and function. Additionally, the phenomenon of neuroplasticity, the brain’s capacity to reorganize itself by forming new neural connections, is central to modern neuroscience [117] and to understanding PBM’s capacity to initiate long term pain relief.
Plasticity continues throughout the lifespan, supporting learning, memory, and recovery from injury or disease. Thus, the capacity of the nervous system to respond to targeted neuromodulation with PBM as a novel neuroplasticity-based intervention for pain, can now be more clearly understood as an adaptive mechanism.
9.3. PBM in Prevention of Acute Pain Progressing to Chronic Pain
The ability of PBM to relieve pain with acute analgesic and anaesthetic effects by blocking nerve conduction occurs within a time frame of minutes [51]. This is most effectively demonstrated clinically in PBM in induction of dental anaesthesia [50]. However, one of the goals of pain medicine is to prevent the transition from acute to chronic pain which occurs in approximately 10% of patients who have surgery. This typically begins as acute postoperative pain that is difficult to control but soon transitions into a persistent pain condition with neuropathic features that are unresponsive to opioids [118]. When injury occurs, especially nerve injury, there is an immediate release of nerve growth factor (NGF) which is transported to the cell body via retrograde fast axonal flow, stimulating gene transcription. This response mitigates further damage and initiates repair and regeneration. Similarly, tumor necrosis factor (TNF-α) is a key molecule in peripheral nerve injury being both protective and destructive [119]. This and other pro-inflammatory cytokines, such as brain-derived neurotrophin growth factor (BDNF), interleukin 6 (IL-6) are produced in Schwann cells and fibroblasts within peripheral nerve [120] and are associated with the development of central sensitisation, hyperalgesia and mechanical hypersensitivity [121,122]. TNF-α, for example, is conveyed by axonal retrograde transport to neuronal cell bodies of DRG. In the mouse model which can take up to 6 hours with movement at 200-300mm/day peaking between day 2 and day 5 post injury. Another important example of such transport is that of the trafficking of c-Jun N-terminal kinase (JNK) signalling molecules which stimulates nerve repair following axonal injury [123]. Lindwall et al. in a series of experiments demonstrated retrograde axonal transport of JNK and related molecules which was inhibited by the application of a cytoskeletal disruptor, vinblastine. When translocated to the cell body in the DRG via signalling mechanisms of retrograde trafficking these pronocioceptive proteins induce transcription of proinflammatory peptides including substance P and receptors TRPV1 and Nav1.8 gated sodium channels [124]. This mechanism links locally generated signals at the site of injury (or surgery) in the form of transcription factors to long-term nociceptive sensitization within the peripheral axons of sensory neurons to remote molecular effects within the cell bodies. There are several relevant examples where LI can disrupt the transport of these pro-nociceptive molecules. Expression of IL-6 which increases the synthesis of CREB, a transcription factor which is retrogradely trafficked to the cell body and induces “hyperalgesic priming”, is suppressed by 904nm laser at an ED of 1.97J/cm2 in gingival fibroblasts. This study identified a biphasic response with increases in IL-6 at 0.84-1.4J/cm2 [125]. In a model of spinal cord injury 660nm, cw laser 100mW; ED of 0.5J/cm2 and a PD of 819mW/cm2 suppressed IL-6 to control levels [126]. In an inflammatory model of knee arthritis in rats 810nm laser PD: 5000mW/cm2 delivering the light with total energy of 1-10J reduced IL-6. This study too showed a biphasic effect with no change in IL-6 levels at the higher dose level of 10J [127]. In another model of acute inflammation TNFα levels were reduced by irradiation 650nm laser with output powers varying from 1-5mW at power densities of 12.5, 31.25 and 62.5mW/cm2. Only the ED 31.25 mW/cm2 showed significant reduction in TNFα levels. De Oliveira Martins et al. showed improvement in sensory symptoms in a rat model of alveolar nerve injury in which laser decreased expression of brain derived nerve growth factor (BDNF) by 40% compared with controls [128]. In contrast 810nm, 50mW, at 1 or 4.4J/cm2 increased NGF expression on day 20 following irradiation in human Schwann cells, with the potential influence on nerve regeneration [129]
The ability of LI to block these signalling molecules has potentially important therapeutic benefits for preventing the transition from acute to chronic pain but, unlike drugs such as colchicine and vinblastine, both cytoskeletal disruptors, is not limited by serious side effects. Similarly, attempts at pharmacological intervention directed to NGF blockade as a potential treatment of neuropathic pain have also been hampered by serious side effects [130].
10. Conclusion
The importance of the neuronal cytoskeleton in maintaining the integrity of the nervous system as well as responding to acute and chronic painful pathological conditions is well understood. however, few studies have examined the specific effects of photons on the cytoskeleton in the PNS. The observation that laser can modulate the infrastructure of the cytoskeleton in somato-sensory nerves has important clinical implications. Both bio-electric signalling and molecular transport in nerves are disrupted. These observations can be hypothesized as mechanisms for the observed clinical analgesic and anesthetic effects of PBM especially for long-term pain relief, prevention of transition of acute to chronic pain, treatment of neuropathic pain and pre-emptive treatment efficacy in reducing pain. Though multiple complex mechanisms underlie the efficacy of PBM further research into how PBM modifies the cytoskeleton is likely to provide a new avenue for understanding its therapeutic benefits as a novel neuroplasticity-modulating modality in the context of modern neuroscience. This review reveals new insights into how PBM can directly modulate the somatosensory nervous system and harness neuroplasticity to modify the “Pain Matrix” for long-term pain management.
Author Contributions
Conceptualization, RC and PJA.; writing—original draft preparation, RC and PJA.; writing—RC and PJA.; project administration, PJA. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
Dr Adahir Labrador-Garrido, Postdoctoral Research Associate, University: of Sydney Central Clinical School | Brain and Mind Centre, Faculty of Medicine and Health, the University of Sydney, M02F, Neuroinflammation Group, prepared the images for the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| DRG | Dorsal Root Ganglion |
| PBM | Photobiomodulation |
| AIS | Axon initial segment |
| LI | Laser irradiation |
| MMP | Mitochondrial membrane potential |
| FAF | Fast axonal flow |
References
- Hennessy, S.N.; Corcoran, G.D. Low-level laser therapy in osteoarthritic pain: A narrative review with an approach to integrated clinical use. Osteoarthr Cartil Open 2025, 7, 100685. [Google Scholar] [CrossRef]
- Khaleghi, A.; Salari, B.; Mafi, S.; Fekrazad, R. Photobiomodulation therapy in the management of myofascial pain dysfunction syndrome: A randomized triple-blind clinical trial. Photodiagnosis Photodyn Ther 2025, 56, 105239. [Google Scholar] [CrossRef] [PubMed]
- Cidral-Filho, F.; Donatello, N.N.; Lugtu, C.; Hewitson, A. Photobiomodulation on shoulder and neck pain and disability: A comprehensive review. Lasers Med Sci 2024, 39, 263. [Google Scholar] [CrossRef]
- Kulkarni, S.; Walsh, L.J.; Bhurani, Y.; George, R. Assessment of the onset of analgesia and length of analgesia following the use of PBM with different wavelengths: a clinical study. Lasers Med Sci 2024, 39, 236. [Google Scholar] [CrossRef]
- Liang, R.; George, R.; Walsh, L.J. Pulpal response following photo-biomodulation with a 904-nm diode laser: a double-blind clinical study. Lasers Med Sci 2016, 31, 1811–1817. [Google Scholar] [CrossRef]
- Nascimento, A.I.; Mar, F.M.; Sousa, M.M. The intriguing nature of dorsal root ganglion neurons: Linking structure with polarity and function. Prog Neurobiol 2018, 168, 86–103. [Google Scholar] [CrossRef] [PubMed]
- Fréal, A.; Hoogenraad, C.C. The dynamic axon initial segment: From neuronal polarity to network homeostasis. Neuron 2025, 113, 649–669. [Google Scholar] [CrossRef]
- Koike, T. Distinctive glial cells in the dorsal root ganglion: their morphology and functions. Anat Sci Int 2025, 100, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Moog, M.; Ballesteros, A.; Ernfors, P.; Swartz, K. Molecular and electrophysiological characterization of cutaneous Schwann cells. Proceedings of the Biophysical Society Annual Meeting 2025, 2025, 604a. [Google Scholar] [CrossRef]
- Mokoena, D.R.; Houreld, N.N.; Dhilip Kumar, S.S.; Abrahamse, H. Photobiomodulation at 660 nm Stimulates Fibroblast Differentiation. Lasers Surg Med 2020, 52, 671–681. [Google Scholar] [CrossRef]
- Etemadi, A.; Sadatmansouri, S.; Sodeif, F.; Jalalishirazi, F.; Chiniforush, N. Photobiomodulation Effect of Different Diode Wavelengths on the Proliferation of Human Gingival Fibroblast Cells. Photochem Photobiol 2021, 97, 1123–1128. [Google Scholar] [CrossRef]
- Shaikh-Kader, A.; Houreld, N.N.; Rajendran, N.K.; Abrahamse, H. Levels of Cyclooxygenase 2, Interleukin-6, and Tumour Necrosis Factor-alpha in Fibroblast Cell Culture Models after Photobiomodulation at 660 nm. Oxid Med Cell Longev 2021, 2021, 6667812. [Google Scholar] [CrossRef]
- y Cajal, S. Degeneration & regeneration of the nervous system; Hafner Publishing Company, 1959; Volume 1. [Google Scholar]
- Colom, B.; Poitelon, Y.; Huang, W.; Woodfin, A.; Averill, S.; Del Carro, U.; Zambroni, D.; Brain, S.D.; Perretti, M.; Ahluwalia, A.; et al. Schwann cell-specific JAM-C-deficient mice reveal novel expression and functions for JAM-C in peripheral nerves. Faseb j 2012, 26, 1064–1076. [Google Scholar] [CrossRef] [PubMed]
- Sundt, D.; Gamper, N.; Jaffe, D.B. Spike propagation through the dorsal root ganglia in an unmyelinated sensory neuron: a modeling study. J Neurophysiol 2015, 114, 3140–3153. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.C.; Murillo, B.R.; Bessa, R.; Ribeiro, R.; Ferreira da Silva, T.; Porfírio-Rodrigues, P.; Martins, G.G.; Brites, P.; Kneussel, M.; Misgeld, T.; et al. Axon-specific microtubule regulation drives asymmetric regeneration of sensory neuron axons. Elife 2025, 13. [Google Scholar] [CrossRef]
- Vogl, A.W.; Lillie, M.A.; Piscitelli, M.A.; Goldbogen, J.A.; Pyenson, N.D.; Shadwick, R.E. Stretchy nerves are an essential component of the extreme feeding mechanism of rorqual whales. Curr Biol 2015, 25, R360–361. [Google Scholar] [CrossRef]
- Scherer, S.S.; Arroyo, E.J. Recent progress on the molecular organization of myelinated axons. J Peripher Nerv Syst 2002, 7, 1–12. [Google Scholar] [CrossRef]
- Purves, D.; Augustine, G.; Fitzpatrick, D.; Katz, L.; LaMantia, A.; McNamara, J.; Williams, S. Projections from the Cerebellum. In Neuroscience; Sinauer Associates, 2001. [Google Scholar]
- Hanani, M.; Spray, D.C. Emerging importance of satellite glia in nervous system function and dysfunction. Nature reviews 2020, 21, 485–498. [Google Scholar] [CrossRef]
- Jessen, K.R.; Mirsky, R. The Success and Failure of the Schwann Cell Response to Nerve Injury. Front Cell Neurosci 2019, 13, 33. [Google Scholar] [CrossRef]
- Kastriti, M.E.; Faure, L.; Von Ahsen, D.; Bouderlique, T.G.; Boström, J.; Solovieva, T.; Jackson, C.; Bronner, M.; Meijer, D.; Hadjab, S.; et al. Schwann cell precursors represent a neural crest-like state with biased multipotency. Embo j 2022, 41, e108780. [Google Scholar] [CrossRef]
- Furlan, A.; Adameyko, I. Schwann cell precursor: a neural crest cell in disguise? Dev Biol 2018, 444 Suppl 1, S25–s35. [Google Scholar] [CrossRef]
- Abdo, H.; Calvo-Enrique, L.; Lopez, J.M.; Song, J.; Zhang, M.D.; Usoskin, D.; El Manira, A.; Adameyko, I.; Hjerling-Leffler, J.; Ernfors, P. Specialized cutaneous Schwann cells initiate pain sensation. Science 2019, 365, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Freal, A.; Hoogenraad, C.C. Neuronal Cytoskeleton: Presynaptic Boutons as Hotspots for Activity-Dependent Microtubule Nucleation. Curr Biol 2019, 29, R1307–R1309. [Google Scholar] [CrossRef]
- Pellegrini, L.; Wetzel, A.; Granno, S.; Heaton, G.; Harvey, K. Back to the tubule: microtubule dynamics in Parkinson’s disease. Cell Mol Life Sci 2017, 74, 409–434. [Google Scholar] [CrossRef]
- Koltzenburg, M.; Bennett, D.L.; Shelton, D.L.; McMahon, S.B. Neutralization of endogenous NGF prevents the sensitization of nociceptors supplying inflamed skin. The European journal of neuroscience 1999, 11, 1698–1704. [Google Scholar] [CrossRef]
- Anders, J.J.; Lanzafame, R.J.; Arany, P.R. Low-level light/laser therapy versus photobiomodulation therapy. Photomed Laser Surg 2015, 33, 183–184. [Google Scholar] [CrossRef]
- Lam, T.S.; Abergel, R.P.; Meeker, C.A.; Castel, J.C.; Dwyer, R.M.; Uitto, J. Laser Stimulation of Collagen Synthesis in Human Skin Fibroblasts. Lasers in Life Sciences 1986, 1, 61–77. [Google Scholar]
- Young, S.; Dyson, M.; Bolton, P. Effect of light on calcium uptake by macrophages. Laser Therapy 1990, 2, 53–57. [Google Scholar] [CrossRef]
- Honmura, A.; Yanase, M.; Obata, J.; Haruki, E. Therapeutic effect on GaAlAs Diode Laser Irradiation on Experimentally Induced Inflammation in Rats. Lasers in Surgery & Medicine 1992, 12, 441–449. [Google Scholar]
- Holanda, V.M.; Chavantes, M.C.; Wu, X.; Anders, J.J. The mechanistic basis for photobiomodulation therapy of neuropathic pain by near infrared laser light. Lasers Surg Med 2017, 49, 516–524. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Wang, Y.-H.; Wang, C.-Z.; Ho, M.-L.; Kuo, P.-L.; Huang, M.-H.; Chen, C.-H. Effect of Low Level Laser Therapy on Chronic Compression of the Dorsal Root Ganglion. PloS one 2014, 9, e89894. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, M.V.P.; Kawakubo, M.; Ferraresi, C.; Kaippert, B.; Yoshimura, E.M.; Hamblin, M.R. Pain management using photobiomodulation: Mechanisms, location, and repeatability quantified by pain threshold and neural biomarkers in mice. J Biophotonics 2018, 11, e201700370. [Google Scholar] [CrossRef]
- Jenkins, M.W.; Buzza, A.; Skubal, A.C.; Moffitt, M.A.; Anders, J.J. Transient Selective Neural Inhibition via PBM. Photobiomodul Photomed Laser Surg 2024, 42, 574–576. [Google Scholar] [CrossRef]
- Chen, M.; Shimada, K.; Fujita, K.; Ishii, J.; Hirata, T.; Fujisawa, H. Neurite elongation from cultured dorsal root ganglia is inhibited by Ga-Al-As diode laser irradiation. Laser Life Sci 1993, 5, 237–242. [Google Scholar]
- Wesselmann, U.; Kerns, J.; Rymer, W. Laser effects in myelinated and nonmyelinated fibres in the rat peroneal nerve: a quantitative ultrastructural analysis. Exp Neurol 1994, 129, 257–265. [Google Scholar] [CrossRef]
- Wesselmann, U.; Kerns, J.; Rymer, W. Laser effects on myelinated and non-myelinated axons in rat peroneal nerve. Soc Neurosci Abstr 1992, 18, 134. [Google Scholar]
- Wesselmann, U.; Lin, S.; Rymer, W. Effects of Q-switched Nd:YAG laser irradiation on neural impulse propagation: II. Dorsal Roots and Peripheral Nerves. Physiol Chem Phys Med NMR 1991, 23, 81–100. [Google Scholar] [PubMed]
- Wesselmann, U.; Lin, S.; Rymer, W. Effects of Q-switched Nd:YAG laser irradiation on neural impulse propagation: I. Spinal Cord. Physiol Chem Phys Med NMR 1991, 23, 67–80. [Google Scholar] [PubMed]
- Wesselmann, U.; Lin, S.; Rymer, W. Selective decrease of small sensory neurons in lumbar dorsal root ganglia labeled with horseradish peroxidase after Nd:YAG laser irradiation of the tibial nerve in the rat. Exp Neurol 1991, 111, 251–262. [Google Scholar]
- Wesselmann, U.; Rymer, W.; Lin, S. Effect of pulsed infrared lasers on neural condition and axoplasmic transport in sensory nerves. In Proceedings of the Laser Surgery: Advanced Characterization, Therapeutics, and Systems II, 1990; pp. 319–333. [Google Scholar]
- Lin, S.-F.; Wesselmann, U.; Rymer, W.Z. Pulsed laser radiation effects on the distribution of conduction velocities in sciatic nerve of the rat. In Proceedings of the Laser Surgery:Advanced Characterization, Therapeutics and Systems II. SPIE Proceedings, Belingham, WA, 1990; pp. 334–342. [Google Scholar]
- Wesselmann, U.; Rymer, W.Z. Retrograde Horseradish Peroxidase Transport in Motor Axons after Nd:YAG Laser Irradiation of the Tibial Nerve in Rats. Experimental Neurology 1993, 119, 147–152. [Google Scholar] [CrossRef]
- Chow, R.; David, M.; Armati, P. 830-nm laser irradiation induces varicosity formation, reduces mitochondrial membrane potential and blocks fast axonal flow in small and medium diameter rat dorsal root ganglion neurons: implications for the analgesic effects of 830-nm laser. J Peripher Nerv Syst 2007, 12, 28–39. [Google Scholar] [CrossRef]
- Bokhari, L.; Chow, R.; Yan, W.; Armati, P. The effects of 650nm continuous wave laser irradiation on rat dorsal root ganglion neurons and sciatic nerve: relevance to neural mechanisms of low-level laser therapy for chronic pain. In Proceedings of the 18th International Society for Lasers in Medicine and Surgery (ISLMS), Tokyo, Japan, 1 December 2009, 2009. [Google Scholar]
- Kudoh, C.; Inomata, K.; Okajima, K.; Motegi, M.; Ohshiro, T. Effects of 830nm Gallium Aluminium Garsenide diode laser radiation on rat saphenous nerve sodium-potassium-adenosine triphosphatase activity: a possible pain attenuation mechanism examined. Laser Ther 1989, 1, 63–67. [Google Scholar]
- Motzkin, J.C.; Basbaum, A.I.; Crowther, A.J. Neuroanatomy of the nociceptive system: From nociceptors to brain networks. Int Rev Neurobiol 2024, 179, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Chow, R.; Armati, P.J. Inhibitory effects of visible 650-nm and infrared 808-nm laser irradiation on somatosensory and compound muscle action potentials in rat sciatic nerve: implications for laser-induced analgesia. J Peripher Nerv Syst 2011, 16, 130–135. [Google Scholar] [CrossRef]
- Chan, A.; Armati, P.; Moorthy, A.P. Pulsed Nd: YAG laser induces pulpal analgesia: a randomized clinical trial. J Dent Res 2012, 91, 79S–84S. [Google Scholar] [CrossRef] [PubMed]
- Chow, R.; Armati, P.; Laakso, E.L.; Bjordal, J.M.; Baxter, G.D. Inhibitory effects of laser irradiation on peripheral mammalian nerves and relevance to analgesic effects: a systematic review. Photomed Laser Surg 2011, 29, 365–381. [Google Scholar] [CrossRef]
- Walker, J. Temporary suppression of clonus in humans by brief photostimulation. Brain Res 1985, 340, 109–113. [Google Scholar] [CrossRef]
- Baxter, G.; Allen, J.; Bell, A. The effect of low-energy density laser irradiation upon human median nerve conduction latencies. J Physiol 1991, 435, 63. [Google Scholar]
- Baxter, G.; Bell, A.; Allen, J.; Ravey, J. Laser mediated increase in median nerve conduction latencies. Irish J Med Sci 1991, 160, 145. [Google Scholar]
- Baxter, G.D.; Allen, A.J.; Bell, J.; Ravey, J.; Diamantopoulos, C. Effect of laser (830nm) upon conduction in the median nerve. Amer Soc Laser Med Surg Abstr 1991, 3, 79. [Google Scholar]
- Baxter, G.C.; Walsh, D.M.; Allen, J.M.; Lowe, A.S.; Bell, A.J. Effects of low intensity infrared laser irradiation upon conduction in the human median nerve in vivo. Exp Physiol 1994, 79, 227–234. [Google Scholar] [CrossRef]
- Lowe, A.; Baxter, G.; Walsh, D.; Allen, J. Effect of low-intensity laser irradiation (830nm) upon skin temperature and antidromic conduction latencies in human median nerve: relevance of radiant exposure. Laser Surg Med 1994, 14, 40–46. [Google Scholar] [CrossRef]
- Walsh, D.; Baxter, G.; Allen, J. The effect of 820nm laser irradiation upon conduction in the frog (Rana Temporaria) sciatic nerve in vitro. Laser Therapy 1995, 7, 5–10. [Google Scholar] [CrossRef]
- Snyder-Mackler, L.; Bork, C.E. Effect of helium-neon laser irradiation on peripheral sensory nerve latency. Phys Ther 1988, 68, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Cambier, D.; Blom, K.; Witvrouw, E.; Ollevier, G.; De Muynck, M.; Vanderstraeten, G. The influence of low intensity infrared laser irradiation on conduction characteristics of peripheral nerve: A randomised, controlled, double blind study on the sural nerve. Laser Med Sci 2000, 15, 195–200. [Google Scholar] [CrossRef]
- Hadian, M.; Moghagdam, B. The effects of low power laser on electrophysiological parameters of sural nerve in normal subjects: a comparison between 670 and 780nm wavelengths. Acta Med Iran 2003, 41, 138–142. [Google Scholar]
- Vinck, E.; Coorevits, P.; Cagnie, B.; De Muynck, M.; Vanderstraeten, G.; Cambier, D. Evidence of changes in sural nerve conduction mediated by light emitting diode irradiation. Lasers Med Sci 2005, 20, 35–40. [Google Scholar] [CrossRef]
- Nelson, A.; Friedman, M. Somatosensory trigeminal evoked potential amplitudes following low-level laser and sham irradiation over time. Laser Ther 2001, 13, 60–64. [Google Scholar] [CrossRef]
- Safavi-Farokhi, Z.; Bakhtiary, A.H. The effect of infrared laser on sensory radial nerve electrophysiological parameters. Electromyogr Clin Neurophysiol 2005, 45, 353–356. [Google Scholar]
- Suganthirababu, P.; Prathap, L.; Alagesan, J.; Kumaresan, A.; Srinivasan, V.; D., G. Neurophysiological effect of low level laser therapy on Ulnar Nerve. Biomedicine 2021, 3, 543–546. [Google Scholar] [CrossRef]
- Telemeco, T.A.; Schrank, E.C. The Effect of Light Therapy on Superficial Radial Nerve Conduction Using a Clustered Array of Infrared Super luminous Diodes and Red Light Emitting Diodes. J Lasers Med Sci 2013, 4, 17–24. [Google Scholar]
- Orchardson, R.; Whitters, C. Effect of HeNe and pulsed Nd:YAG laser irradiation on intradental nerve responses to mechanical stimulation of dentine. Laser Surg Med 2000, 26, 241–249. [Google Scholar] [CrossRef]
- Tsuchiya, D.; Kawatani, M.; Takeshige, C. Laser irradiation abates neuronal responses to nociceptive stimulation of rat-paw skin. Brain Res Bull 1994, 34, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Mezawa, S.; Iwata, K.; Naito, K.; Kamogawa, H. The possible analgesic effect of soft-laser irradiation on heat nociceptors in the cat tongue. Archs Oral Biol 1988, 33, 693–694. [Google Scholar] [CrossRef]
- Kasai, S.; Kono, T.; Sakamoto, T.; Mito, M. Effects of low-power laser irradiation on multiple unit discharges induced by noxious stimuli in the anesthetized rabbit. J Clin Laser Med Surg 1994, 12, 221–224. [Google Scholar] [CrossRef]
- Wakabayashi, H.; Hamba, M.; Matsumoto, K.; Tachibana, H. Effect of irradiation by semiconductor laser on responses evoked in trigeminal caudal neurons by tooth pulp stimulation. Laser Surg Med 1993, 13, 605–610. [Google Scholar] [CrossRef]
- Shimoyama, M.; Fukuda, Y.; Shimoyama, N.; IIjima, K.; Mizuguchi, T. Effect of HeNe laser irradiation on synaptic transmission of the superior cervical ganglion in the rat. J Clin Laser Med Surg 1992, 10, 337–342. [Google Scholar] [CrossRef]
- Sato, T.; Kawatani, M.; Takeshige, C.; Matsumoto, I. Ga-Al-As laser irradiation inhibits neuronal activity associated with inflammation. Acupunct Electrother Res 1994, 19, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Jimbo, K.; Noda, K.; Suzuki, H.; Yoda, K. Suppressive effects of low-power laser irradiation on bradykinin evoked action potentials in cultured murine dorsal root ganglia cells. Neurosci Lett 1998, 240, 93–96. [Google Scholar] [CrossRef]
- Kono, T.; Kasai, S.; Sakamoto, T.; Mito, M. Cord dorsum potentials suppressed by low power laser irradiation on a peripheral nerve in the cat. J Clin Laser Med Surg 1993, 11, 115–118. [Google Scholar] [CrossRef]
- Miura, A.; Kawatani, M. Effects of diode laser irradiation on sensory ganglion cells from the rat. Pain Res 1996, 11, 175–183. [Google Scholar] [CrossRef]
- Arber, S.; Boskov, D.; Rymer, W. Effects of CO2 and HeNe laser irradiation on rat sciatic nerve in vitro. In Proceedings of the Laser Tissue Interaction - P Soc Photo-Opt Ins SPIE, Los Angeles, California, USA, 1990; pp. 196–204. [Google Scholar]
- Tsuchiya, D.; Kawatani, M.; Takeshige, C.; Sato, T.; Matsumoto, I. Diode laser irradiation selectively diminishes slow component of axonal volleys to dorsal roots from the saphenous nerve in the rat. Neurosci Lett 1993, 161, 65–68. [Google Scholar] [CrossRef]
- Kasai, S.; Kono, T.; Yasuhiro, Y.; Kotani, H.; Sakamoto, T.; Mito, M. Effect of low-power laser irradiation on impulse conduction in anaesthetized rabbits. J Clin Laser Med Surg 1996, 14, 107–109. [Google Scholar] [CrossRef]
- Kao, M.-C.; Lin, F.-Y.; Chiu, H.C. Laser effect on somatosensory potential of the peripheral nerve. Amercian Society for Laser Medicine & Surgery Abstracts 1989, 30–31. [Google Scholar]
- Shimoyama, N.; Iijima, K.; Shimoyama, M.; Mizuguchi, T. The effects of helium-neon laser on formalin-induced activity of dorsal horn neurons in the rat. J Clin Laser Med Surg 1992, 10, 91–94. [Google Scholar] [CrossRef]
- Uta, D.; Ishibashi, N.; Konno, T.; Okada, Y.; Kawase, Y.; Tao, S.; Kume, T. Near-Infrared Photobiomodulation of the Peripheral Nerve Inhibits the Neuronal Firing in a Rat Spinal Dorsal Horn Evoked by Mechanical Stimulation. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Buzza, A.; Tapas, K.; Zhuo, J.; Anders, J.J.; Lewis, S.J.; Jenkins, M.W.; Moffitt, M. Selective neural inhibition via photobiomodulation alleviates behavioral hypersensitivity associated with small sensory fiber activation. Lasers Surg Med 2024, 56, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Chow, R.T.; Barnsley, L.B.; Heller, G.Z. The effect of 300mW, 830nm laser on chronic neck pain: a double-blind, randomized, placebo-controlled study. Pain 2006, 124, 201–210. [Google Scholar] [CrossRef]
- Stausholm, M.B.; Naterstad, I.F.; Joensen, J.; Lopes-Martins, R.A.B.; Saebo, H.; Lund, H.; Fersum, K.V.; Bjordal, J.M. Efficacy of low-level laser therapy on pain and disability in knee osteoarthritis: systematic review and meta-analysis of randomised placebo-controlled trials. BMJ Open 2019, 9, e031142. [Google Scholar] [CrossRef]
- Korada, H.Y.; Arora, E.; Maiya, G.A.; Rao, S.; Hande, M.; Shetty, S.; Gundmi, S.; Anche, P.; Amravadi, S. Effectiveness of Photobiomodulation Therapy on Neuropathic Pain, Nerve Conduction and Plantar Pressure Distribution in Diabetic Peripheral Neuropathy - A Systematic Review. Curr Diabetes Rev 2023, 19, e290422204244. [Google Scholar] [CrossRef]
- Haslerud, S.; Magnussen, L.H.; Joensen, J.; Lopes-Martins, R.A.; Bjordal, J.M. The efficacy of low-level laser therapy for shoulder tendinopathy: a systematic review and meta-analysis of randomized controlled trials. Physiother Res Int 2015, 20, 108–125. [Google Scholar] [CrossRef]
- de Lara Quagliotto, G.; Manchope, M.P.; Hilario, R.; Zubeldia, V.; Stacheslki, R.A.; de Carvalho, A.R.; Buzanello, M.R.; Bertolini, G.R.F. Photobiomodulation associated with physical exercise in shoulder impingement syndrome. Systematic review with meta-analysis. Photochem Photobiol 2025. [Google Scholar] [CrossRef] [PubMed]
- de Andrade, A.L.; Bossini, P.S.; Parizotto, N.A. Use of low level laser therapy to control neuropathic pain: A systematic review. J Photochem Photobiol B 2016, 164, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Argenta, P.A.; Ballman, K.V.; Geller, M.A.; Carson, L.F.; Ghebre, R.; Mullany, S.A.; Teoh, D.G.; Winterhoff, B.J.; Rivard, C.L.; Erickson, B.K. The effect of photobiomodulation on chemotherapy-induced peripheral neuropathy: A randomized, sham-controlled clinical trial. Gynecol Oncol 2017, 144, 159–166. [Google Scholar] [CrossRef]
- Clijsen, R.; Brunner, A.; Barbero, M.; Clarys, P.; Taeymans, J. Effects of low-level laser therapy on pain in patients with musculoskeletal disorders: a systematic review and meta-analysis. Eur J Phys Rehabil Med 2017, 53, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Liao, C.D.; Hong, J.P.; Hsu, W.C.; Wu, C.W.; Chen, H.C. Effects of laser therapy on chronic low back pain: A systematic review and meta-analysis of randomized controlled trials. Clin Rehabil 2022, 36, 289–302. [Google Scholar] [CrossRef]
- Huang, Z.; Ma, J.; Chen, J.; Shen, B.; Pei, F.; Kraus, V.B. The effectiveness of low-level laser therapy for nonspecific chronic low back pain: a systematic review and meta-analysis. Arthritis Res Ther 2015, 17, 360. [Google Scholar] [CrossRef]
- Ibarra, A.M.C.; Biasotto-Gonzalez, D.A.; Kohatsu, E.Y.I.; de Oliveira, S.S.I.; Bussadori, S.K.; Tanganeli, J.P.C. Photobiomodulation on trigeminal neuralgia: systematic review. Lasers Med Sci 2021, 36, 715–722. [Google Scholar] [CrossRef]
- Mukhtar, R.; Fazal, M.U.; Saleem, M.; Saleem, S. Role of low-level laser therapy in post-herpetic neuralgia: a pilot study. Lasers Med Sci 2020, 35, 1759–1764. [Google Scholar] [CrossRef]
- Navarro-Ledesma, S.; Carroll, J.; Burton, P.; Ana, G.M. Short-Term Effects of Whole-Body Photobiomodulation on Pain, Quality of Life and Psychological Factors in a Population Suffering from Fibromyalgia: A Triple-Blinded Randomised Clinical Trial. Pain Ther 2023, 12, 225–239. [Google Scholar] [CrossRef]
- Soriano, F.; Campana, V.; Moya, M.; Gavotto, A.; Simes, J.; Soriano, M.; Soriano, R.; Spitale, L.; Palma, J. Photobiomodulation of pain and inflammation in microcrystalline arthropathies: experimental and clinical results. Photomed Laser Surg 2006, 24, 140–150. [Google Scholar] [CrossRef]
- Orchardson, R.; Peacock, J.M.; Whitters, C.J. Effects of pulsed Nd:YAG laser radiation on action potential conduction in.
- nerve fibres inside teeth in vitro. Journal of Dentistry 1998, 26, 421–426. [CrossRef] [PubMed]
- Díaz, L.; Restelli, L.; Valencia, E.; Atalay, D.I.; Abarca, J.M.; Gil, A.C.; Fernández, E. Effectiveness of low-level laser therapy on temporomandibular disorders. A systematic review of randomized clinical trials. Photodiagnosis Photodyn Ther 2025, 53, 104558. [Google Scholar] [CrossRef] [PubMed]
- Candido-do-Prado, L.G.; Ribeiro-Silva, V.H.A.; Simoes-Barbosa, A.F.; Mazzi-Chaves, J.F.; Magri, L.V. Differential effectiveness of photobiomodulation in muscular and articular temporomandibular disorders: a systematic review and critical appraisal. Lasers Med Sci 2025, 40, 487. [Google Scholar] [CrossRef]
- Shukla, D.; Muthusekhar, M.R. Efficacy of low-level laser therapy in temporomandibular disorders: A systematic review. Natl J Maxillofac Surg 2016, 7, 62–66. [Google Scholar] [CrossRef]
- Sarangi, P.; Suman, S.; Satapathy, S.K.; Meher, R.; Das, A.; Pradeep, D.M. The Pros and Cons of Dental Laser Therapy in Conservative Dentistry and Endodontics-A Systematic Review. J Pharm Bioallied Sci 2024, 16, S3083–s3085. [Google Scholar] [CrossRef]
- Khemiss, M.; Dammak, N.; Lajili, O.; Yacoub, S.; Ben Khelifa, M. Efficacy of laser therapy on primary burning mouth syndrome: a systematic review. J Oral Facial Pain Headache 2024, 38, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Sonesson, M.; De Geer, E.; Subraian, J.; Petrén, S. Efficacy of low-level laser therapy in accelerating tooth movement, preventing relapse and managing acute pain during orthodontic treatment in humans: a systematic review. BMC Oral Health 2016, 17, 11. [Google Scholar] [CrossRef]
- Scheuer, T. Regulation of sodium channel activity by phosphorylation. Semin Cell Dev Biol 2011, 22, 160–165. [Google Scholar] [CrossRef]
- Ross, G. Photobiomodulation Therapy: A Possible Answer to the Opioid Crisis. Photobiomodul Photomed Laser Surg 2019, 37, 667–668. [Google Scholar] [CrossRef]
- Poste, G.; Papahadjopoulos, D.; Nicholson, G. Local anaesthetics affect transmembrane cytoskeletal control of mobility and distribution of cell surface receptors. P Nat Acad Sci USA 1975, 72, 4430–4434. [Google Scholar] [CrossRef]
- Nicolson, G.; JR., S.; Poste, G. Effects of local anaesthetics on cell morphology and membrane-associated cytoskeletal organization in BALB3/3T3. J Cell Biol 1976, 68, 395–402. [Google Scholar] [CrossRef]
- Tanelian, D.L.; Markin, V.S. Biophysical and functional consequences of receptor-mediated nerve fiber transformation. Biophys J 1997, 72, 1092–1108. [Google Scholar] [CrossRef]
- Haschke, R.H.; Byers, M.R.; Fink, B.R. Effects of lidocaine on rabbit brain microtubular protein. J Neurochem 1974, 22, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.; Muto, E.; Mashimo, T.; Iwane, A.H.; Yoshiya, I.; Yanagida, T. Direct inhibition of microtubule-based kinesin motility by local anesthetics. Biophys J 2000, 78, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Chow, R.T.; Johnson, M.I.; Lopes-Martins, R.A.; Bjordal, J.M. Efficacy of low-level laser therapy in the management of neck pain: a systematic review and meta-analysis of randomised placebo or active-treatment controlled trials. Lancet 2009, 374, 1897–1908. [Google Scholar] [CrossRef]
- Glazov, G.; Yelland, M.; Emery, J. Low-level laser therapy for chronic non-specific low back pain: a meta-analysis of randomised controlled trials. Acupunct Med 2016, 34, 328–341. [Google Scholar] [CrossRef]
- Naterstad, I.F.; Joensen, J.; Bjordal, J.M.; Couppe, C.; Lopes-Martins, R.A.B.; Stausholm, M.B. Efficacy of low-level laser therapy in patients with lower extremity tendinopathy or plantar fasciitis: systematic review and meta-analysis of randomised controlled trials. BMJ Open 2022, 12, e059479. [Google Scholar] [CrossRef] [PubMed]
- Bjordal, J.; Johnson, M.; Iverson, V.; Aimbire, F.; Lopes-Martins, R. Photoradiation in acute pain: A systematic review of possible mechanisms of action and clinical effects in randomized placebo-controlled trials. Photomed Laser Surgery 2006, 24, 158–168. [Google Scholar] [CrossRef]
- Hu, Y.; Ma, J.; Chen, B.; Pang, J.; Liang, W.; Wu, W. The Duration of Chronic Pain Can Affect Brain Functional Changes of the Pain Matrix in Patients with Chronic Back Pain: A Resting-State fMRI Study. J Pain Res 2024, 17, 1941–1951. [Google Scholar] [CrossRef]
- Gazerani, P. The neuroplastic brain: current breakthroughs and emerging frontiers. Brain Res 2025, 1858, 149643. [Google Scholar] [CrossRef]
- Glare, P.; Aubrey, K.R.; Myles, P.S. Transition from acute to chronic pain after surgery. Lancet 2019, 393, 1537–1546. [Google Scholar] [CrossRef]
- Shubayev, V.I.; Myers, R.R. Axonal transport of TNF-alpha in painful neuropathy: distribution of ligand tracer and TNF receptors. J Neuroimmunol 2001, 114, 48–56. [Google Scholar] [CrossRef]
- Murwani, R.; Armati, P. Peripheral nerve fibroblasts as a source of IL-6, TNFalpha and IL-1 and their modulation by IFNgamma. Journal of Neurological Sciences 1998, 161, 99–109. [Google Scholar] [CrossRef]
- Fregnan, F.; Muratori, L.; Simoes, A.R.; Giacobini-Robecchi, M.G.; Raimondo, S. Role of inflammatory cytokines in peripheral nerve injury. Neural Regen Res 2012, 7, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Guptarak, J.; Wanchoo, S.; Durham-Lee, J.; Wu, Y.; Zivadinovic, D.; Paulucci-Holthauzen, A.; Nesic, O. Inhibition of IL-6 signaling: A novel therapeutic approach to treating spinal cord injury pain. Pain 2013, 154, 1115–1128. [Google Scholar] [CrossRef] [PubMed]
- Lindwall, C.; Kanje, M. Retrograde axonal transport of JNK signaling molecules influence injury induced nuclear changes in p-c-Jun and ATF3 in adult rat sensory neurons. Mol Cell Neurosci 2005, 29, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Sorkin, A.; Von Zastrow, M. Signal transduction and endocytosis: close encounters of many kinds. Nat Rev Mol Cell Biol 2002, 3, 600–614. [Google Scholar] [CrossRef]
- Harorli, O.T.; Hatipoglu, M.; Erin, N. Effect of Photobiomodulation on Secretion of IL-6 and IL-8 by Human Gingival Fibroblasts In Vitro. Photobiomodul Photomed Laser Surg 2019, 37, 457–464. [Google Scholar] [CrossRef]
- Mojarad, N.; Janzadeh, A.; Yousefifard, M.; Nasirinezhad, F. The role of low level laser therapy on neuropathic pain relief and interleukin-6 expression following spinal cord injury: An experimental study. J Chem Neuroanat 2018, 87, 60–70. [Google Scholar] [CrossRef]
- Pallotta, R.C.; Bjordal, J.M.; Frigo, L.; Leal Junior, E.C.; Teixeira, S.; Marcos, R.L.; Ramos, L.; Messias Fde, M.; Lopes-Martins, R.A. Infrared (810-nm) low-level laser therapy on rat experimental knee inflammation. Lasers Med Sci 2012, 27, 71–78. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Martins, D.; Martinez dos Santos, F.; Evany de Oliveira, M.; de Britto, L.R.; Benedito Dias Lemos, J.; Chacur, M. Laser therapy and pain-related behavior after injury of the inferior alveolar nerve: possible involvement of neurotrophins. J Neurotrauma 2013, 30, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, S.O.; Golestaneh, A.F.; Shafiee, A.; Hafizi, M.; Omrani, H.A.; Soleimani, M. Effects of low level laser therapy on proliferation and neurotrophic factor gene expression of human schwann cells in vitro. J Photochem Photobiol B 2012, 107, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Belanger, P.; West, C.R.; Brown, M.T. Development of pain therapies targeting nerve growth factor signal transduction and the strategies used to resolve safety issues. J Toxicol Sci 2018, 43, 1–10. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



