
Submitted:
15 January 2026
Posted:
16 January 2026
You are already at the latest version
Abstract
Lipid sources are essential components in modern swine nutrition, not only due to their high energy density but also because of their positive effects on palatability, feed efficiency, and micronutrient absorption. However, rising raw material costs have encouraged the use of oxidized fats and oils (OxFO) as a cost-effective alternative in pig diets. These lipids, degraded by thermal and handling factors, undergo chemical alterations that negatively affect digestibility, energy metabolism, and animal health. This review critically examines the current scientific evidence regarding the impact of oxidized fat consumption in swine production systems. The physiological and biochemical mechanisms by which lipid oxidation products impair mitochondrial β-oxidation, cellular oxidative balance, energy efficiency, and meat quality are discussed. Moreover, the practical consequences on productive performance, muscle oxidative stability, and the expression of inflammatory and antioxidant markers are explored. Findings suggest that although the use of oxidized fats may offer economic savings, their metabolic and productive repercussions can compromise profitability and sustainability. The need to define safe inclusion thresholds (when replacement is not feasible), standardize analytical methods to assess oxidation status, and consider nutritional alternatives to mitigate adverse effects is emphasized.
Keywords:
oxidized fats
; swine nutrition
; energy metabolism
; oxidative stress
; meat quality
1. Introduction
Lipid sources represent one of the most energy-dense components in swine diets [1,2]. Beyond their caloric contribution, dietary fats improve palatability, reduce feed dustiness, and enhance the absorption of fat-soluble vitamins, collectively promoting feed efficiency and growth performance [3]. Consequently, the proper selection and handling of dietary lipid sources is critical to ensuring productive and economic performance in intensive pig production systems [4,5].).
However, the sustained increase in the cost of conventional feed ingredients has prompted the industry to consider more economical alternatives. Among these, recycled or low-quality fats and oils—often exhibiting varying degrees of oxidation—have gained attention [6,7]. Oxidized lipids are the result of chemical degradation processes induced by heat, light, oxygen, and pro-oxidant metals, leading to the formation of hydroperoxides, aldehydes, ketones, and polar compounds that compromise their nutritional value and safety [4,8]. The inclusion of oxidized fats and oils (OxFO) in swine diets has been associated with adverse effects on multiple physiological and productive parameters. Studies have reported reduced energy digestibility, decreased average daily gain, increased susceptibility to oxidative stress, and mitochondrial inefficiency [9,10,11,12]. These disruptions also manifest in meat quality, with increased volatile compound formation, reduced oxidative stability, and diminished sensory acceptance of the final product [13,14].
Despite the growing body of research documenting the negative impacts of OFO, the current literature lacks a systemic and integrative framework to assess their effects holistically. In this context, the application of theoretical models such as the General Systems Theory becomes relevant to analyze the interplay between animal, technological, and environmental components within production systems [15,16]. This theory suggests that biological systems—such as intensively reared pigs—should be evaluated considering the dynamic interactions between their internal components and the surrounding environment, particularly when the system is challenged by inputs of variable quality.
This review aims to provide a critical and schematic overview of the effects of OFO in swine feeding, with a focus on the physiological, biochemical, and technological changes associated with lipid peroxidation, lipid digestion and metabolism, animal energy efficiency, and meat quality. The goal is to offer an integrated perspective to guide future research and practical decision-making regarding the use of recycled lipids, within a framework of sustainability, metabolic efficiency, and animal health.
2. Methodological approach
For the characterization and schematic modeling of OxFO use in swine feeding and its relationship with energy metabolism, scientific literature was reviewed using major academic databases including PubMed, ScienceDirect, and Google Scholar. The collected information was analyzed through the lens of General Systems Theory (GST), a theoretical framework that supports the integration of multiple scientific disciplines to address complex problems in a holistic manner [15]. In real-world scenarios, processes and phenomena often defy classification within a single discipline due to their inherent complexity, resulting in what are known as complex systems [16,17]
A complex system is a conceptual abstraction of reality, organized as an integrated whole whose components are: (1) non-separable, (2) delineated by specific boundaries (with feedback mechanisms), and (3) not suitable for isolated study. To study a phenomenon in isolation is to remove it from the context in which its interactions unfold—a practice not applicable to biological systems, which are inextricably linked to their environment [16].
Swine production systems can be conceptualized as complex systems composed of the interaction of four key elements: context, human input, the animal itself, and technology [16]. In this review, emphasis was placed on the technological component, as it is a critical determinant of operational efficiency. This component manifests at two levels: a physical level, encompassing tools and processes used to reduce system variability (e.g., lipid source selection), and a biological level, involving applied knowledge to regulate physiological functions such as lipid digestion and metabolism [18].
Considering these dimensions, the impact of OxFO in swine feeding was examined through two analytical modeling approaches. The first was a general or “black box” approach, in which both endogenous and exogenous factors influencing energy metabolism are assessed without decomposing the system into its constituent parts. The second was a formal or “open box” model, allowing for a more precise exploration of internal relationships. This approach integrates concepts such as functional homogeneity, interdependence among components, cross-disciplinary knowledge, and evaluation under specific hypotheses [19]. Such a framework enables a deeper understanding of the physiological and productive consequences of oxidized fat intake in pigs.
To represent the metabolic changes induced by OxFO consumption, conceptual diagrams were used as analytical tools. These schematic representations help simplify and visualize key interactions within the biological system, omitting structural details that are not essential to understanding the phenomenon. Rather than reproducing the system in its entirety, the goal is to functionally represent observable or inferable relationships that are otherwise too complex to be analyzed collectively. This approach is consistent with the principles of GST, which emphasize that the purpose of modeling is not to mimic reality, but to partially reconstruct it through representations that facilitate its study [20].
3. Factors Attributable and Non-Attributable to the Biological Component (Pig) that Modify Energy Metabolism when Consuming Oxidized Fat Sources
3.1. Context
The continuous rise in the cost of feed ingredients in pig production has driven the industry to explore strategies to reduce feed-associated expenses, which can account for up to 70% of total production costs [7]. Among these strategies are: (i) improving feed efficiency through precision formulation, and (ii) incorporating alternative feed ingredients, such as oxidized fats and oils (OFO), increasingly sourced from the food processing industry due to their lower acquisition cost [6].
However, before incorporating new ingredients or by-products such as OxFO into swine diets, it is essential to understand their chemical composition and digestibility. This information is fundamental for accurate diet formulation and to determine whether these ingredients can be used safely and efficiently [21]. Although OxFO may initially appear as a cost-saving alternative, their use can be counterproductive [12,22]. Degraded fats compromise both palatability and digestibility, thereby reducing feed efficiency and potentially increasing total production costs. Moreover, the inclusion of OxFO in pig diets may negatively impact animal health by increasing susceptibility to disease, which further elevates production costs [10,23]. Meat quality may also be affected by OxFO inclusion, leading to reduced market value due to altered lipid profiles and increased oxidative damage [24,25]. Therefore, while oxidized fats may seem economically viable at first glance, the indirect costs associated with their use can ultimately outweigh the initial savings, compromising the overall profitability and sustainability of swine production systems.
3.2. Human Component
Despite the pig being one of the most extensively studied livestock species, specific physiological processes—particularly those related to metabolic responses to oxidized fat consumption and their effects on productivity and meat quality—remain insufficiently integrated [12,24,26]. The fragmented nature of the available evidence hinders the development of objective nutritional strategies and limitations for the safe and efficient use of OxFO in swine diets. While feed cost reduction is a clear incentive [27], a comprehensive understanding of the factors that modulate energy metabolism in pigs consuming oxidized lipids is still lacking. This limitation complicates the decision-making process and underscores the need for systems-based approaches to identify causal relationships and regulatory points within the biological network affected by OxFO intake (Figure 1).
3.3. Animal Component
Animal production systems are artificial constructs in which the animal depends entirely on human intervention to meet its physiological needs [28]. Thus, the greater the extent to which the biological component [the animal] is supported within the system, the greater the productive output. However, achieving this requires not merely the physical presence of the animal in the system, but a deep understanding of its biology [15]
Addressing the challenges and physiological effects associated with the inclusion of OxFO in swine feeding necessitates a shift from a relatively simple system—composed of human, animal, and technology—to a more complex configuration: the modulation of energy metabolism in pigs in response to oxidized lipid intake (Figure 2). Starting from this analytical premise, and by advancing our understanding of this phenomenon, it becomes feasible to regulate and strategically control the use of OxFO in swine nutrition.
3.4. Technological Component
From the perspective of General Systems Theory, the technological component in the study of OxFO in swine nutrition refers to the set of tools, processes, and scientific applications used to analyze, control, and mitigate the negative effects of these lipids on animal metabolism and product quality. This component integrates transversally across multiple critical subtopics within the pig production system.
In the analysis of lipid peroxidation, technology is reflected in the use of advanced analytical methods to quantify oxidative damage. In the domain of energy utilization and efficiency, technologies such as indirect calorimetry and mitochondrial biomarkers are employed to assess the metabolic impact of oxidized lipids. Likewise, in the study of lipid digestion and metabolism, digestive technologies—including exogenous enzymes and emulsifiers—are applied to enhance the absorption of degraded fats. Finally, in meat quality evaluation, the technological component includes instrumental tools for the analysis of color, volatile compound profiles, and texture, as well as nutritional strategies to preserve muscle oxidative stability. Collectively, these technological applications are essential for understanding and intervening in swine production systems that incorporate OxFO into the diet, supporting evidence-based decision-making and precision nutrition practices.
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
4. Lipid Peroxidation
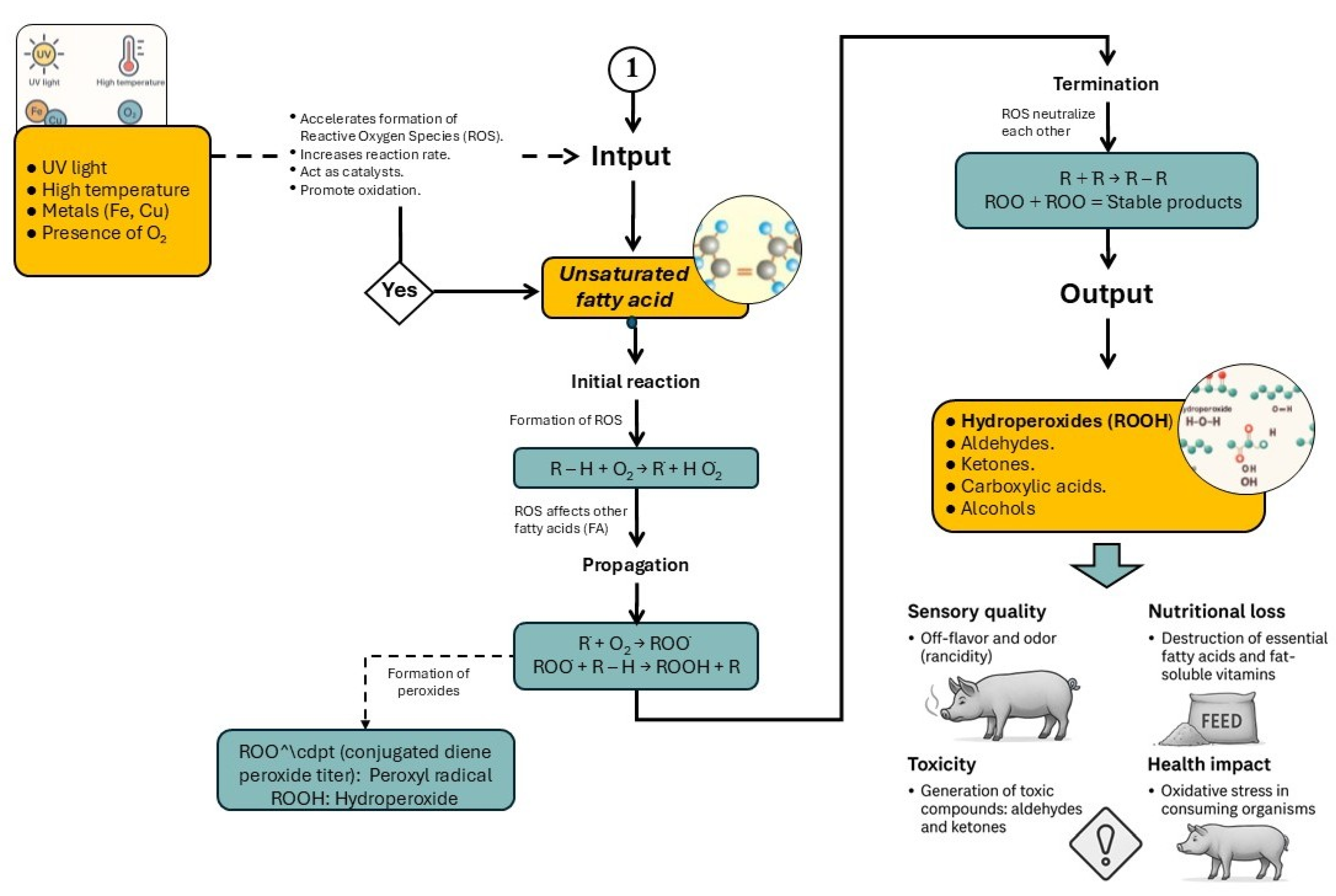
The peroxidation of fats and oils is a complex process influenced by several factors, including the degree of fatty acid saturation, temperature, and the presence of oxygen, transition metals (e.g., Cu and Fe), undissociated salts, water, and various non-lipid compounds [4]. Lipid peroxidation proceeds through three main stages: initiation, propagation, and termination (Figure 3), with each phase both consuming and generating a variety of chemical compounds [29]. The lipid hydroperoxides formed during the initial stages of peroxidation not only degrade lipid quality but also lead to the formation of secondary and tertiary oxidation products. These include aldehydes, ketones, alcohols, hydrocarbons, volatile organic acids, and epoxide compounds—all of which can exert detrimental effects on animal productivity and health [4,8]. The accumulation of these oxidative by-products in feed lipids reduces palatability, disrupts nutrient digestibility, and contributes to oxidative stress at the cellular level.
Due to the complexity of lipid peroxidation and the wide array of compounds formed and degraded during the process, accurately quantifying the degree of peroxidation in feed ingredients remains a significant challenge [30]. Consequently, no single method can adequately characterize or predict the oxidative status of lipids [4]. Various techniques are used to assess lipid oxidation, including spectrophotometric and chromatographic methods, each with distinct advantages and limitations. The choice of method depends on factors such as the sample type, target compounds, and analytical resources available.
Figure 2.
Structured diagram of the porcine biological system integrating physiological, technological, and contextual variables associated with the consumption of oxidized lipids. Numbers 1, 2, 3, and 4 serve as visual connectors referring to complementary sub-diagrams, where the involved functional mechanisms are described in greater detail.
Figure 2.
Structured diagram of the porcine biological system integrating physiological, technological, and contextual variables associated with the consumption of oxidized lipids. Numbers 1, 2, 3, and 4 serve as visual connectors referring to complementary sub-diagrams, where the involved functional mechanisms are described in greater detail.
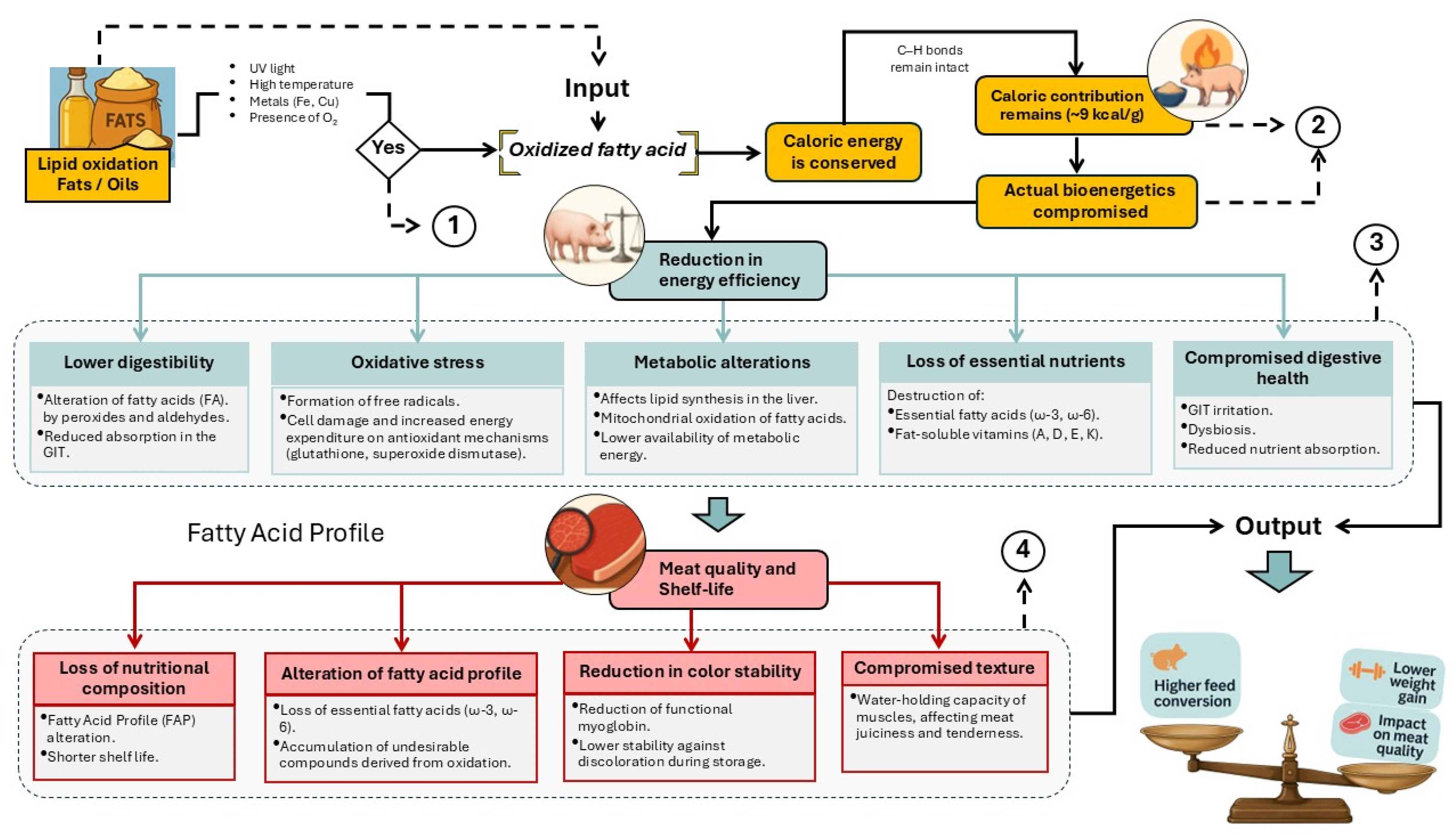
Figure 3.
Phases and key compounds generated during lipid peroxidation: initiation, propagation, and termination. Modified from Belitz et al. [29]. .
Figure 3.
Phases and key compounds generated during lipid peroxidation: initiation, propagation, and termination. Modified from Belitz et al. [29]. .
The inclusion of oxidized fats in animal diets can negatively impact feed palatability, nutrient digestibility, and overall performance [5,26]. In pigs, consumption of diets containing OxFO has been associated with reduced feed efficiency and lower weight gain [12]. Additionally, lipid peroxidation can compromise the nutritional quality of ingredients by reducing the bioavailability of fat-soluble vitamins and essential antioxidants [12]. Oxidative by-products may also interact with other biomolecules—such as proteins and nucleic acids—altering their structure and function, and contributing to the development of pathological conditions [8,29]. Notably, the intake of OxFO has been shown to induce oxidative stress in tissues, a condition linked to cellular damage and impaired reproductive function in animals [12,31].
5. Energy Utilization in Pigs Fed Oxidized Fats
Energy efficiency in pig production is a critical determinant of performance and profitability. Fats and oils serve as concentrated sources of energy in swine diets; however, they are rarely evaluated at the biological level due to the complexity, cost, and labor-intensive nature of the procedures involved. Such evaluations often require controlled experimental conditions and the use of live animals, making them impractical for routine quality control in feed manufacturing [4,32].
Lipid sources used in swine nutrition vary widely in their physicochemical characteristics—including color, fatty acid profile, free fatty acid content, degree of saturation/unsaturation, saponification value, and impurities [4,5]. Regarding the oxidative status of fats used in pig diets, a broad range of oxidation levels has been reported across different lipid sources (Table 1) [33].
Although the number of studies specifically addressing the use of oxidized fats in swine nutrition is still limited, available evidence indicates that diets containing OxFO have detrimental effects on pig growth performance [5]. These effects include reduced weight gain, slower growth rates, increased systemic oxidative stress, and lower energy digestibility [4,9,12,26]. The reduction in productive performance observed in pigs fed OxFO-rich diets has been closely linked to impaired energy utilization mechanisms (Figure 4).
The amount of usable energy a pig can derive from dietary lipids depends on species, age, and the chemical composition of the fat source [34]. For lipids, metabolizable energy (ME) typically accounts for 98–99% of digestible energy (DE), and net energy (NE) represents approximately 88–90% of ME [9,35]. However, accurate prediction of fat utilization is complicated by endogenous fat losses (EFL). In growing pigs, reported EFL values range from 1.33 to 14.02 g/kg of dry matter intake, reflecting variability driven by the type and amount of fat in the diet [34]. Diets rich in unsaturated fatty acids have been shown to increase fecal fat excretion due to microbial hydrogenation of unsaturated fatty acids in the hindgut (Jørgensen et al., 1993).
Consequently, both overestimation and underestimation of dietary lipid energy content are common. This variability results from differences not only among lipid sources but also within the same source, especially when the energy value has not been empirically determined [2]. For instance, NE values of choice white grease measured experimentally differed by up to 14% compared to estimates based on NRC (2012) equations [34]. To improve estimation accuracy, it has been proposed to incorporate detailed fatty acid profiles into predictive models [1], as natural variation in lipid composition can significantly impact energy availability—particularly in fats and oils derived from the food industry.
Industrial processes such as rendering, refining, and cracking can chemically alter fat structure, affecting both ME and oxidative susceptibility. Additionally, improper processing, transport, storage, and handling can dilute the effective energy content of lipids. Contributing factors include moisture, impurities, unsaponifiable compounds (e.g., waxes and complex lipids), and non-elutable material (NEM) [34]. These dilutive elements can reduce lipid energy content by up to 46% [36]. Such findings underscore the potential for misestimating both energy contribution and lipid quality when repurposing high-grade fats originally intended for human consumption in swine diets [33]. As lipids oxidize, their saturation degree increases—a factor that plays a central role in the negative impact of OxFO on swine energy metabolism [5,12].
6. Consumption of Oxidized Fats and Oils and Their Relationship with Energy Efficiency In Pigs
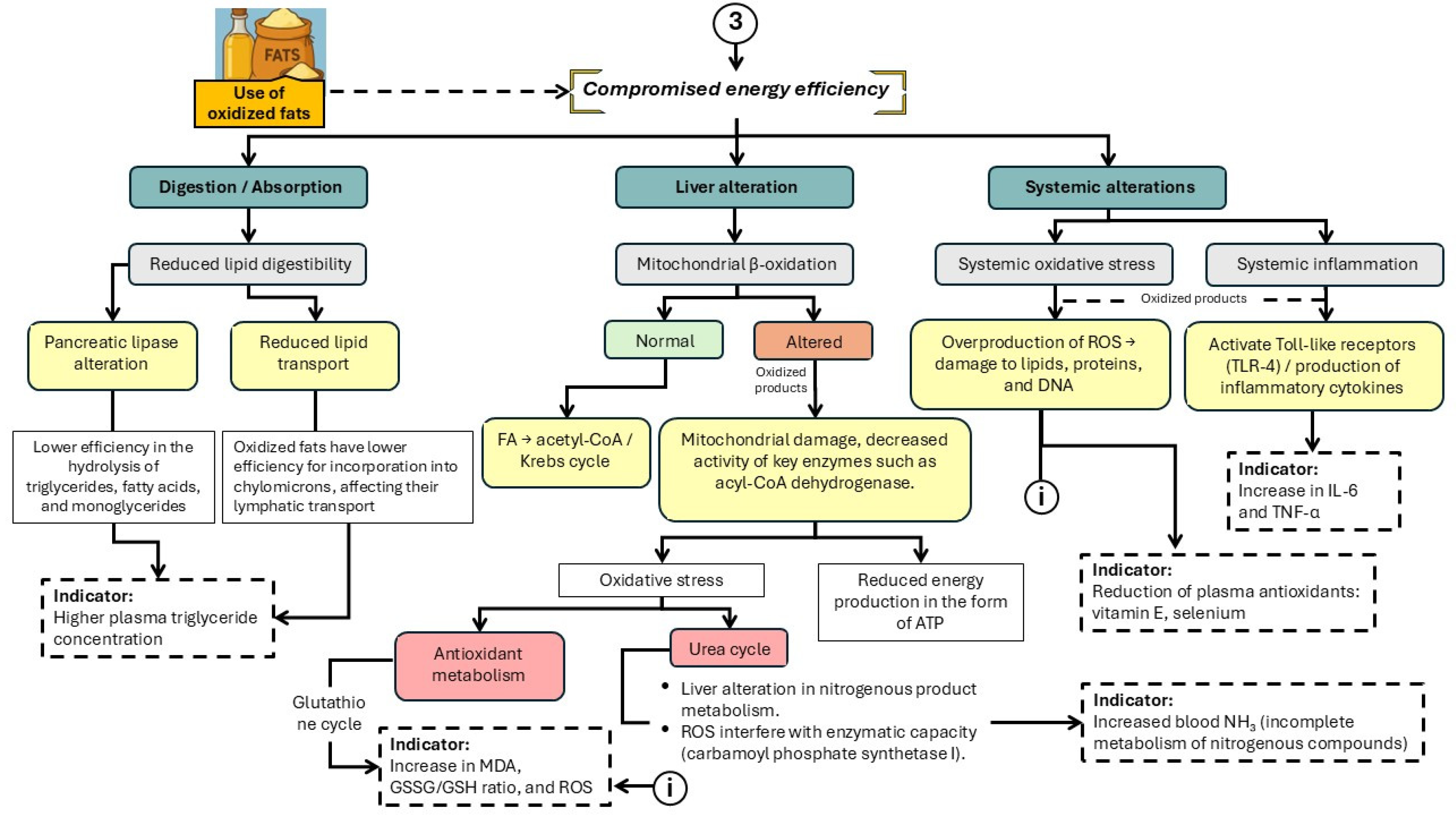
The use of OxFO in swine nutrition poses an emerging risk to both energy efficiency and metabolic health. While lipids serve as a concentrated energy source in diets for growing pigs, oxidation—caused by inadequate processing, storage, or reuse—leads to the formation of reactive compounds that may disrupt multiple metabolic pathways [37] (Figure 5).
The schematic representation summarizes the main physiological disruptions induced by OFO consumption. These include reduced lipid digestibility and mitochondrial damage, which impair β-oxidation and ATP production [38] In addition, systemic effects such as oxidative stress, inflammation mediated by Toll-like receptor 4 (TLR-4), and alterations in the urea and glutathione cycles have been observed. These disruptions compromise overall energy efficiency and are associated with elevated plasma markers of cellular damage, including malondialdehyde (MDA), the glutathione redox ratio (GSSG/GSH), interleukin-6 (IL-6), and tumor necrosis factor-alpha (TNF-α) [39,40,41].
6.1. Lipid Digestion and Metabolism
Under normal physiological conditions, lipid digestion in pigs begins in the oral cavity, where salivary enzymes such as lingual lipase and esterases contribute to the initial hydrolysis of triglycerides. This activity is particularly important in neonatal piglets [42], although its relevance decreases with age [43]. Mastication facilitates the release and subsequent emulsification of dietary lipids. In the stomach, triglycerides are partially converted into diglycerides and free fatty acids (FFA), which can trigger hormonal responses such as the release of cholecystokinin (CCK), a key regulator of intestinal motility and satiety [44,45]. However, the contribution of gastric lipases in pigs is limited, as they are inactivated under acidic pH conditions [43,46].
Figure 5.
Main physiological alterations associated with the consumption of oxidized fats and oils that interfere with energy efficiency in pigs. .
Figure 5.
Main physiological alterations associated with the consumption of oxidized fats and oils that interfere with energy efficiency in pigs. .
The principal site of lipid digestion is the small intestine, where more than 70% of dietary fats still reach the lumen in triglyceride form [47]. Non-emulsified fats are initially coated with bile salts and phospholipids, allowing the action of pancreatic enzymes including lipase, colipase, and cholesterol esterase [48,49]. Colipase stabilizes lipase at the lipid–water interface, facilitating triglyceride hydrolysis into monoglycerides and FFA (Delorme et al., 2011; Lo and Tso, 2009). These products are incorporated into mixed micelles for transport to the enterocytes, where they are absorbed either by passive diffusion (short- and medium-chain fatty acids) or via active transport mechanisms (long-chain fatty acids) [50,51]
However, the inclusion of OxFO in the diet can significantly disrupt this process (Wang et al., 2020; Liu et al., 2023). At the digestive level, OxFO may impair pancreatic lipase activity and reduce the efficiency of lipid emulsification and transport, particularly by altering the incorporation of triglycerides into chylomicrons [52]. These disruptions result in reduced lipid absorption, increased circulating triglycerides, and lower energy availability. In addition, OxFO has been shown to inhibit mixed micelle formation and potentially compete with de novo lipids at the absorption interface [53].
In post-absorptive metabolism, oxidized fatty acids may induce mitochondrial dysfunction in the liver, especially by impairing β-oxidation through reduced activity of key enzymes such as acyl-CoA dehydrogenase, thereby compromising ATP production. Moreover, oxidative lipid by-products generate reactive oxygen species (ROS), triggering systemic oxidative stress, intestinal inflammation, and activation of immune pathways such as Toll-like receptor 4 (TLR-4), leading to the release of proinflammatory cytokines including IL-6 and TNF-α [54,55]. These alterations not only reduce the energy efficiency of the diet but also impair intestinal health, cellular membrane integrity, and the bioavailability of essential nutrients such as vitamin E and selenium, both of which are reduced in animals fed oxidized lipids [55].
6.2. Mitochondrial β-Oxidation
Under normal physiological conditions, β-oxidation is the primary catabolic pathway for the energy utilization of fatty acids in pigs. This process takes place within the mitochondrial matrix and begins with the transport of long-chain fatty acids into the mitochondria via the carnitine–carnitine palmitoyltransferase I (CPT-I) system, located on the outer mitochondrial membrane [56]. Once inside, fatty acids undergo sequential degradation by a set of enzymes—acyl-CoA dehydrogenase, enoyl-CoA hydratase, 3-hydroxyacyl-CoA dehydrogenase, and 3-ketoacyl-CoA thiolase—producing acetyl-CoA, NADH, and FADH₂. These products feed into the Krebs cycle and the electron transport chain to support ATP synthesis [57].
However, when pigs consume OxFO, this process is impaired. Lipid metabolism inherently produces reactive oxygen species (ROS), which disrupt mitochondrial function and increase oxidative stress, thereby reducing energy efficiency [11,58]. Excess ROS damage mitochondrial membranes and oxidative machinery, leading to reduced ATP production and negatively impacting growth and feed conversion efficiency [59].
In pigs fed OFO, significant reductions in the NADH/NAD⁺ ratio and in cytochrome c oxidase (COX) activity have been reported, indicating compromised metabolic coupling and decreased respiratory chain efficiency [60,61]. At the systemic level, elevated plasma acylcarnitine concentrations have been observed, reflecting an accumulation of incompletely oxidized fatty acid intermediates and mitochondrial β-oxidation dysfunction [59].
From an enzymatic standpoint, chronic consumption of oxidized lipids impairs the activity of key enzymes such as long-chain acyl-CoA dehydrogenase (LCAD) and CPT-I, with activity reductions of up to 40% reported [62,63]. This inhibition limits fatty acid entry into mitochondria and delays their catabolism, resulting in the accumulation of free fatty acids and increased hydrogen peroxide (H₂O₂) production, a toxic byproduct that exacerbates oxidative damage in hepatic and muscular tissues [64].
The metabolic consequence of this imbalance is lipotoxicity, characterized by the accumulation of intermediates such as diacylglycerols and ceramides that interfere with cellular energy homeostasis [65]. In pigs, this condition is reflected in elevated plasma concentrations of non-esterified fatty acids (NEFA) [66]. This increase in NEFA is not only a marker of lipid mobilization but is also associated with hepatic dysfunction, insulin resistance, and accelerated protein catabolism. These findings demonstrate that the consumption of OxFO compromises mitochondrial β-oxidation, not only reducing the pig’s ability to extract energy from lipids but also promoting a pro-oxidative environment that impairs cellular health, growth, and overall production efficiency.
7. Consumption of Oxidized Fats and Its Relationship with Meat Quality
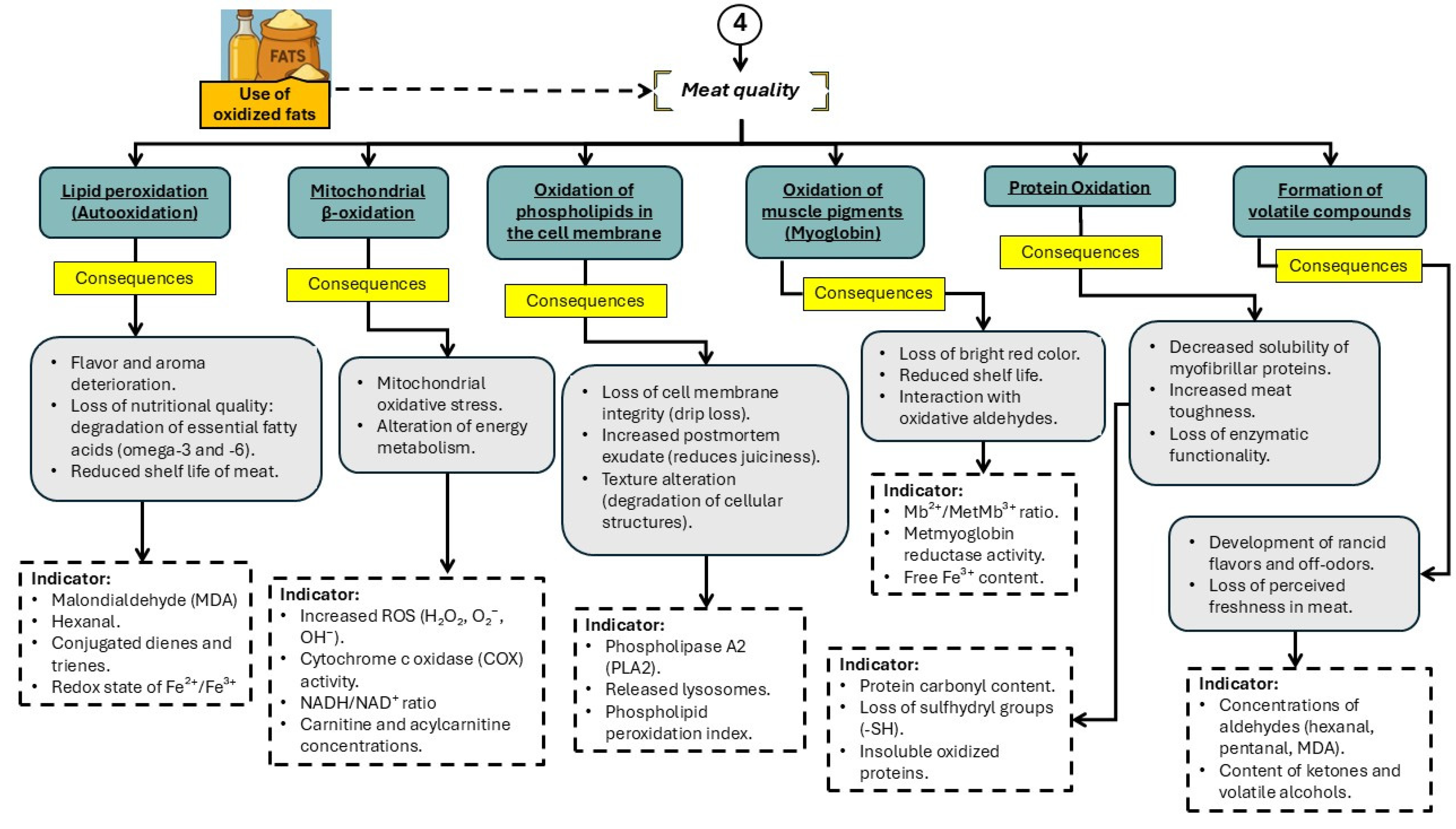
As previously discussed, the use of OxFO in swine diets is a common practice in the pork industry due to their low cost and availability [27]. However, beyond adversely affecting animal metabolism, growth, and health, the inclusion of OFO also has detrimental consequences on the final quality of pork [67,68,69]. This impact is primarily linked to the disruption of several key metabolic pathways, including lipid peroxidation, mitochondrial β-oxidation, phospholipid oxidation in cell membranes, myoglobin oxidation, protein oxidation, and the generation of volatile compounds (Figure 6). These alterations compromise important meat quality traits such as oxidative stability, color, tenderness, water-holding capacity, and sensory attributes. The accumulation of oxidative products in muscle tissue, including malondialdehyde (MDA) and protein carbonyls, has been associated with lipid rancidity, discoloration, and increased drip loss—factors that directly reduce consumer acceptance and shelf life of pork products.
7.1. Lipid Peroxidation and Oxidative Stress
The consumption of oxidized fats introduces lipid hydroperoxides and free radicals into the pig’s organism, promoting oxidative stress and disrupting cellular homeostasis [70]. Lipid peroxidation involves the reaction of polyunsaturated fatty acids with molecular oxygen, generating highly reactive secondary products such as malondialdehyde (MDA) and 4-hydroxynonenal (HNE), which can trigger cell damage and programmed cell death (apoptosis) in various tissues [71,72]. These peroxidation products compromise the integrity of cellular membranes, increasing membrane permeability and impairing cellular function—factors that directly impact meat stability.
Figure 6.
Main physiological alterations associated with the consumption of oxidized fats and oils that affect meat quality in pigs.
Figure 6.
Main physiological alterations associated with the consumption of oxidized fats and oils that affect meat quality in pigs.
Diets enriched with oxidized fats have been shown to increase MDA levels in plasma and muscle tissues (Table 2), with values ranging from 2.5 to 4.0 nmol/g of fresh meat. In contrast, pigs fed fresh lipids exhibit MDA levels below 2.0 nmol/g [14,71]. Likewise, hexanal—a volatile compound derived from n-6 fatty acid peroxidation—has been detected at concentrations exceeding 1.2 µg/g in deteriorated pork (Table 2), contributing to rancid flavors and reduced consumer acceptance [13].
Oxidative stress induced by lipid peroxidation also disrupts the activity of endogenous antioxidant defense systems. In pigs fed oxidized lipids, the activity of key enzymes such as superoxide dismutase (SOD) and catalase (CAT) is reduced (Table 2), compromising the neutralization of reactive oxygen species (ROS) [31]. Furthermore, the concentration of reduced glutathione (GSH)—a key cellular antioxidant—can decrease by up to 40%, enhancing the propagation of oxidative damage in muscle tissues and accelerating postmortem degradation of meat [73].
Another critical consequence of lipid peroxidation is its impact on inflammation and immune response. Ingested oxidized lipids activate pro-inflammatory signaling pathways such as nuclear factor kappa B (NF-κB), leading to elevated production of cytokines including interleukin-6 (IL-6) and tumor necrosis factor alpha (TNF-α) [74]. In pigs exposed to high-OxFO diets, plasma IL-6 levels can increase up to 2.5-fold compared to pigs fed fresh lipids, suggesting a chronic inflammatory state that may impair growth and overall welfare.
The deterioration of oxidative stability due to lipid peroxidation not only shortens meat shelf life but also compromises its sensory and nutritional attributes [75]. Lipid oxidation in skeletal muscle results in the degradation of essential fatty acids and the loss of fat-soluble vitamins, such as vitamins E and A, thereby reducing the nutritional value of the final product [76].
7.2. Oxidation of Phospholipids in Cellular Membranes
One of the less visible but critically harmful effects of feeding pigs oxidized lipid sources is the structural and functional damage to cellular membranes. These membranes, composed largely of phospholipids containing polyunsaturated fatty acids, are particularly susceptible to lipid peroxidation in the presence of free radicals and reactive oxygen species (ROS) [75,77]. Oxidation of these lipids compromises not only the physical integrity of the lipid bilayer but also essential functions such as ion transport, intracellular signaling, and transmembrane receptor function [78]. In monogastric animals such as pigs, chronic exposure to oxidized lipids has been associated with a significant increase in phospholipid hydroperoxide levels, destabilizing both plasma and mitochondrial membranes [5,33].
This structural damage triggers a cascade of functional alterations. Loss of membrane integrity facilitates the release of lysosomal enzymes and activates apoptotic pathways, undermining cell viability and ultimately affecting the quality of muscle tissue [79]. Mitochondrial membranes, in particular, are highly vulnerable to this process, leading to decreased electron transport efficiency and increased ROS production—factors that perpetuate oxidative damage in tissues [80].

Table 2.
Oxidative stress biomarkers in tissues potentially affecting meat quality in pigs fed fresh vs. oxidized lipids .
Table 2.
Oxidative stress biomarkers in tissues potentially affecting meat quality in pigs fed fresh vs. oxidized lipids .
| Biomarker | Tissue | Lipid Source | Effect on Meat | Reference | |
|---|---|---|---|---|---|
| Fresh | Oxidized | ||||
| MDA | Meat, nmol/g | <0.5 | >2.0 | Accelerates rancidity and reduces oxidative stability. | Boler et al. [31] Degroote et al. [93] |
| Plasma, nmol/mL | 9.15 | 12.51 | |||
| Hexanal | Meat, µg/g | <0.5 | >1.2 | Contributes to the development of rancid flavors. | Fernando et al. [91] |
| SOD | Liver, % reduction | - | 10-20 | Decreases the antioxidant capacity of the muscle, promoting lipid oxidation. | Lindblom et al. [94] |
| Erythrocytes, U/mg | 3.2 | 3.0 | Keller et al. [95] | ||
| Catalasa | Erythrocytes, U/g | 94 | 64 | Reduces protection against peroxides, increasing oxidative instability. | Keller et al. [95] |
| GPx | Erythrocytes, U/g | 97 | 74 | Lower antioxidant capacity in the muscle, accelerating meat deterioration. | Keller et al. [95] |
| Plasma, U/L | 307 | 334 | Degroote et al. [93] |
||
| FRAP | Plasma, µmolFe/L | 135 | 144 | Degroote et al. [93] | |
| IL-6 | Plasma, pg/mL | 43 | 85 | Associated with inflammatory processes that affect muscle quality. | Silva-Guillen et al. [74] |
| TNF-α | Plasma, pg/mL | 23 | 199 | May induce proteolysis and affect meat texture. | Silva-Guillen et al. [74] |
MDA: Malondialdehyde; SOD: Superoxide dismutase; GPx: glutathione peroxidase.
Furthermore, phospholipid oxidation alters membrane fluidity, negatively affecting the function of critical transmembrane proteins such as insulin receptors and glucose transporters, with significant metabolic implications [81]. In vivo studies have also reported reduced activity of Na⁺/K⁺-ATPase, a key enzyme for maintaining membrane potential. Impaired function of this enzyme compromises ionic homeostasis and contributes to cellular deterioration in animals exposed to oxidized lipids [82]. In this context, phospholipid oxidation represents a critical link in the chain of events leading to the deterioration of meat quality in pigs fed degraded fats.
7.3. Myoglobin Oxidation, Protein Oxidation, and Formation of Volatile Compounds
The oxidative stability of meat is a major determinant of its sensory, functional, and nutritional quality [83]. In this context, the oxidation of key components such as myoglobin, structural proteins, and lipids leads to a cascade of degradative changes that directly affect the color, flavor, texture, and shelf life of meat products [83]. In pigs fed oxidized fats, these processes are intensified by increased levels of reactive oxygen species (ROS) and secondary lipid peroxidation products, which exacerbate postmortem oxidative deterioration of muscle tissue [4,11,12].
Myoglobin oxidation: Myoglobin is a heme protein that serves as an oxygen reservoir and transporter in muscle and is the principal determinant of meat color. Its redox state defines the visual appearance of the product: oxymyoglobin (MbO₂) imparts a bright red hue, whereas metmyoglobin (MetMb) confers an undesirable brown coloration [84]. Under oxidative stress—especially in the presence of elevated ROS and reactive aldehydes from lipid peroxidation—the conversion of MbO₂ to MetMb is accelerated [85]. This conversion not only impairs the appearance of the product but also reflects loss of freshness and advanced oxidation. In pigs fed oxidized fats, MetMb levels have been reported to increase by up to 40% compared to pigs fed fresh lipids, significantly compromising visual quality [86]. Furthermore, this process is associated with reduced muscle reducing capacity, as evidenced by decreased activity of metmyoglobin reductase, an enzyme that normally contributes to the regeneration of the desirable red color [85].
Protein oxidation: Skeletal muscle proteins play critical structural and functional roles, including in muscle contraction, nutrient transport, and water retention [87]. Oxidation of these proteins—induced by dietary oxidized lipids—leads to irreversible modifications such as side-chain carbonylation, cross-link formation, and protein fragmentation [88]. These alterations negatively impact texture, reduce myofibrillar protein solubility, and impair water-holding capacity. In swine studies, elevated protein carbonyl content and significant losses of sulfhydryl (-SH) groups have been observed, both of which are indicators of severe structural damage [89]. Additionally, protein oxidation interferes with postmortem enzymatic activity of calpains and cathepsins, disrupting controlled proteolysis during aging and thereby reducing final meat tenderness [90].
Formation of volatile compounds: Oxidative degradation of lipids and proteins generates a wide array of volatile compounds that negatively affect the aroma and flavor of meat. These include aldehydes (e.g., hexanal, pentanal, heptanal), ketones, alcohols, and organic acids—byproducts of polyunsaturated fatty acid and amino acid oxidation [13,85]. These volatiles are responsible for the development of rancid, metallic, and off-flavor notes, which lower product acceptability. Hexanal concentrations in meat from pigs fed OFO have been reported to be up to three times higher than those from pigs fed fresh fats [91]. Moreover, these volatiles can interact with proteins and muscle pigments, further exacerbating oxidative deterioration. The typical profile of desirable volatile compounds, such as short-chain alcohols and lactones, is replaced by secondary aldehydes that contribute to sensory rejection [92].
Together, the oxidation of myoglobin, proteins, and lipids—and the resulting formation of volatile compounds—represent synergistic mechanisms of meat deterioration in pigs fed oxidized lipids. Incorporating natural or synthetic antioxidants into the diet is a key strategy for mitigating these effects and preserving the sensory and functional quality of the final product.
8. General Considerations
While the use of OxFO in swine diets may offer a cost-effective strategy for reducing formulation expenses, it carries significant implications for productive efficiency, energy metabolism, animal health, and meat quality. Scientific evidence consistently shows that lipid oxidation products profoundly disrupt key digestive and metabolic processes, including lipid digestion, intestinal absorption, mitochondrial β-oxidation, and cellular redox balance, ultimately compromising the animal’s physiological homeostasis.
From a production standpoint, the inclusion of oxidized lipids has been associated with reduced feed efficiency, slower growth rates, impaired immune response, and decreased oxidative stability of tissues. These alterations can translate into lower overall economic returns, despite the lower initial cost of the lipid source. Additionally, the final product quality is adversely affected by phenomena such as myoglobin oxidation, formation of undesirable volatile compounds, and oxidative damage to phospholipids and structural proteins—impairing the sensory, technological, and nutritional attributes of pork.
Despite these risks, the use of OxFO remains common in certain production systems, particularly those facing economic constraints or lacking quality control capabilities. Therefore, it is crucial to promote improved lipid ingredient monitoring through reliable analytical methods and to establish science-based inclusion limits. Furthermore, the adoption of antioxidant additives, protective dietary formulations, and targeted nutritional strategies is essential to mitigate the adverse effects of oxidative stress induced by degraded lipids. Collectively, these considerations highlight the need to re-evaluate the use of OxFO in swine nutrition—not merely from an immediate economic standpoint, but through a comprehensive lens that integrates metabolic efficiency, animal welfare, and long-term sustainability.
9. Future Perspectives
Advancing our understanding of the impact of OxFO in swine nutrition requires integrative approaches that go beyond the assessment of energy metabolism and product quality to encompass the sustainability of the production system. Safe inclusion thresholds must be experimentally established, and reliable mitochondrial and digestive biomarkers should be validated for the early detection of functional impairments. In parallel, it is necessary to optimize the use of rapid analytical technologies to assess lipid oxidation in real time, and to explore mitigation strategies involving antioxidants, emulsifiers, and functional additives. Finally, the integration of omics tools and economic–environmental modeling will allow for more accurate evaluations of the cost–benefit ratio of OxFO use, guiding decision-making towards more efficient, safe, and sustainable production systems.
Author Contributions
Conceptualization, L.H.L. and G.O.; methodology, G.O. and E.G.N.; validation, L.H.L., G.O. and M.A.P.; investigation, L.H.L., G.O., E.G.N. and M.A.P.; writing—original draft preparation, L.H.L., G.O., E.G.N. and M.A.P.; writing—review and editing, L.H.L., G.O., E.G.N. and M.A.P.; visualization, L.H.L. and G.O.; supervision, L.H.L. and M.A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Research Project SIGI, No. 12312036470.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data supporting the findings of this study are available from publicly accessible databases, including PubMed, ScienceDirect, and Google Scholar.
Acknowledgments
We thank the Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias (INIFAP-México) for financially supporting the Research Project SIGI, No. 12312036470.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kellner, T.A.; Patience, J.F. The Digestible Energy, Metabolizable Energy, and Net Energy Content of Dietary Fat Sources in Thirteen- and Fifty-Kilogram Pigs. J Anim Sci 2017, 95, 3984–3995. [Google Scholar] [CrossRef]
- NRC Nutrients Requirements of Swine; Eleventh Revised Edition, Ed.; 12va ed.; The National Academies Press: Washington, DC: , 2012; 2012.
- Kil, D.Y.; Ji, F.; Stewart, L.L.; Hinson, R.B.; Beaulieu, A.D.; Allee, G.L.; Patience, J.F.; Pettigrew, J.E.; Stein, H.H. Net Energy of Soybean Oil and Choice White Grease in Diets Fed to Growing and Finishing Pigs. J Anim Sci 2011, 89, 448–459. [Google Scholar] [CrossRef]
- Shurson, G.C.; Kerr, B.J.; Hanson, A.R. Evaluating the Quality of Feed Fats and Oils and Their Effects on Pig Growth Performance. J Anim Sci Biotechnol 2015, 6, 1–11. [Google Scholar] [CrossRef]
- Kerr, B.J.; Kellner, T.A.; Shurson, G.C. Characteristics of Lipids and Their Feeding Value in Swine Diets. J Anim Sci Biotechnol 2015, 6, 1–23. [Google Scholar] [CrossRef]
- Mateos, G.G.; Corrales, N.L.; Talegón, G.; Aguirre, L. Pig Meat Production in the European Union-27: Current Status, Challenges, and Future Trends. Anim Biosci 2024, 37, 755–774. [Google Scholar] [CrossRef]
- Roppa, L.; Duarte, M.E.; Kim, S.W. Pig Production in Latin America. Anim Biosci 2024, 37, 786–793. [Google Scholar] [CrossRef]
- Frankel, N. Lipid Oxidation. Prog Lipid Res 1980, 19, 1–22. [Google Scholar] [CrossRef]
- Li, E.; Liu, H.; Li, Y.; Liu, L.; Wang, F.; Li, D.; Zhang, S. Determination of Net Energy Content of Dietary Lipids Fed to Growing Pigs Using Indirect Calorimetry. J Anim Sci 2018, 96, 2184–2194. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Lee, Ya.W.; Chang, Ch. Effect of Oxidized Frying Oil and Vitamin C Levels on the Hepatic Xenobiotic-Metabolizing Enzyme System of Guinea Pigs. J Nutr Sci Vitaminol 2000, 46, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Lindblom, S.C.; Dozier, W.A.; Shurson, G.C.; Kerr, B.J. Digestibility of Energy and Lipids and Oxidative Stress in Nursery Pigs Fed Commercially Available Lipids. J Anim Sci 2017, 95, 239. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Chen, C.; Kerr, B.J.; Weber, T.E.; Johnston, L.J.; Shurson, G.C. Influence of Thermally Oxidized Vegetable Oils and Animal Fats on Growth Performance, Liver Gene Expression, and Liver and Serum Cholesterol and Triglycerides in Young Pigs. J Anim Sci 2014, 92, 2960–2970. [Google Scholar] [CrossRef]
- Domínguez, R.; Pateiro, M.; Gagaoua, M.; Barba, F.J.; Zhang, W.; Lorenzo, J.M. A Comprehensive Review on Lipid Oxidation in Meat and Meat Products. Antioxidants 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.B.; Solva, M.V. Da; Lannes, S.C.D.S. Lipid Oxidation in Meat: Mechanisms and Protective Factors - a Review. Food Science and Technology (Brazil) 2018, 38, 1–15. [CrossRef]
- Bertalanffy, L. Teoria general de los sistemas; Primera; Fondo de Cultura Económico: México, 1976. [Google Scholar]
- Ortiz, B.P.A.; Delgado, R.A.; Gómez, R.F. Sistemas Alejados Del Equilibrio : Un Lenguaje Para El Diálogo Transdisciplinario; Universidad Autónoma de Tlaxcala; Clave Editorial; 2016; ISBN 9786074373455. [Google Scholar]
- García, R. Sistemas Complejos; Primera; Gedisa, S.A.: Barcelona, 2006. [Google Scholar]
- Soni, R.P.; Katoch, M.; Ladohia, R. Integrated Farming Systems-A Review. Journal of Agriculture and Veterinary Science 2014, 7, 36–42. [Google Scholar] [CrossRef]
- Arnold, G.W.; de Wit, C.T. Critical Evaluation of Systems Analysis in Ecosystems Research and Management; Waneningen Centre for Agricultural Publishing and Documentation, 1976. [Google Scholar]
- Matsuoka, Y.; Ghosh, S.; Kitano, H. Consistent Design Schematics for Biological Systems: Standardization of Representation in Biological Engineering. J R Soc Interface 2009, 6. [Google Scholar] [CrossRef]
- Patience, J.F.; Rossoni-Serão, M.C.; Gutiérrez, N.A. A Review of Feed Efficiency in Swine: Biology and Application. J Anim Sci Biotechnol 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Derouchey, J.M.; Hancock, J.D.; Hines, R.H.; Maloney, C.A.; Lee, D.J.; Cao, H.; Dean, D.W.; Park, J.S. Effects of Rancidity and Free Fatty Acids in Choice White Grease on Growth Performance and Nutrient Digestibility in Weanling Pigs. J Anim Sci 2004, 82, 2937–2944. [Google Scholar] [CrossRef]
- Dibner, J.J.; Atwell, C.A.; Kitchell, M.L.; Shermer, W.D.; Ivey, F.J. Feeding of Oxidized Fats to Broilers and Swine: Effects on Enterocyte Turnover, Hepatocyte Proliferation and the Gut Associated Lymphoid Tissue. Animal Feed Science Technology 1996, 62, 1–13. [Google Scholar] [CrossRef]
- Alonso, V.; Najes, L.M.; Provincial, L.; Guillén, E.; Gil, M.; Roncalés, P.; Beltrán, J.A. Influence of Dietary Fat on Pork Eating Quality. Meat Sci 2012, 92, 366–373. [Google Scholar] [CrossRef]
- Teye, G.A.; Sheard, P.R.; Whittington, F.M.; Nute, G.R.; Stewart, A.; Wood, J.D. Influence of Dietary Oils and Protein Level on Pork Quality. Effects on Muscle Fatty Acid Composition, Carcass, Meat and Eating Quality. Meat Sci 2006, 73, 157–165. [Google Scholar] [CrossRef]
- Rosero, D.S.; Odle, J.; Moeser, A.J.; Boyd, R.D.; Van Heugten, E. Peroxidised Dietary Lipids Impair Intestinal Function and Morphology of the Small Intestine Villi of Nursery Pigs in a Dose-Dependent Manner. British Journal of Nutrition 2015, 114, 1985–1992. [Google Scholar] [CrossRef]
- Canakci, M. The Potential of Restaurant Waste Lipids as Biodiesel Feedstocks. Bioresour Technol 2007, 98, 183–190. [Google Scholar] [CrossRef]
- Juárez, C.A. Teoría de Sistemas En La Producción Animal, 2da ed.; Editorial Académica Española: España, 2019; ISBN 9786074245431. [Google Scholar]
- Belitz, H.D.; Grosch, W.; Schieberle, P. Lipids. In Food Chemistry; Springer Berlin Heidelberg: Berlin, 2009; pp. 158–245. ISBN 9783540699330. [Google Scholar]
- Das, U.N. Essential Fatty Acids: Biochemistry, Physiology and Pathology. Biotechnol J 2006, 1, 420–439. [Google Scholar] [CrossRef] [PubMed]
- Boler, D.D.; Fernández-Dueñas, D.M.; Kutzler, L.W.; Zhao, J.; Harrell, R.J.; Campion, D.R.; Mckeith, F.K.; Killefer, J.; Dilger, A.C. Effects of Oxidized Corn Oil and a Synthetic Antioxidant Blend on Performance, Oxi-Dative Status of Tissues, and Fresh Meat Quality in Finishing Barrows. J. Anim. Sci 2012, 90, 5159–5169. [Google Scholar] [CrossRef] [PubMed]
- Van Kempen, T.A.; Mccomas, S. Infrared Spectroscopy as a Tool for Assessing Fat Quality. Journal of Applied Poultry Research 2002, 11, 191–201. [Google Scholar] [CrossRef]
- Liu, P.; Kerr, B.J.; Chen, C.; Weber, T.E.; Johnston, L.J.; Shurson, G.C. Methods to Create Thermally Oxidized Lipids and Comparison of Analytical Procedures to Characterize Peroxidation. J Anim Sci 2014, 92, 2950–2959. [Google Scholar] [CrossRef]
- Wealleans, A.L.; Bierinckx, K.; di Benedetto, M. Fats and Oils in Pig Nutrition: Factors Affecting Digestion and Utilization. Anim Feed Sci Technol 2021, 277. [Google Scholar] [CrossRef]
- Van Milgen, J.; Noblet, J.; Dubois, S. Nutritional Models Energetic Efficiency of Starch, Protein and Lipid Utilization in Growing Pigs. J Nutr 2001, 131, 1309–1318. [Google Scholar] [CrossRef]
- Wealleans, A.L.; Bierinckx, K.; Witters, E.; di Benedetto, M.; Wiseman, J. Assessment of the Quality, Oxidative Status and Dietary Energy Value of Lipids Used in Non-Ruminant Animal Nutrition. J Sci Food Agric 2021, 101, 4266–4277. [Google Scholar] [CrossRef]
- Hong, C.; Huang, Y.; Cao, S.; Wang, L.; Yang, X.; Hu, S.; Gao, K.; Jiang, Z.; Xiao, H. Accurate Models and Nutritional Strategies for Specific Oxidative Stress Factors: Does the Dose Matter in Swine Production? J Anim Sci Biotechnol 2024, 15. [Google Scholar] [CrossRef]
- Dibner, J.J.; Atwell, C.A.; Kitchell, M.L.; Shermer, W.D.; Ivey, F.J. Animal Feed Feeding of Oxidized Fats to Broilers and Swine: Effects on Enterocyte Turnover, Hepatocyte Proliferation and the Gut Associated Lymphoid Tissue. Anim Feed Sci Technol 1996, 62, 1–13. [Google Scholar] [CrossRef]
- Bogoriani, N.W.; Sudiarta, I.W. Effect of Used Cooking Oil of the Stress Oxidative and Inflammation on Wistar Rats. Biomedical and Pharmacology Journal 2016, 9, 899–907. [Google Scholar] [CrossRef]
- Ammouche, A.; Rouaki, F.; Bitam, A.; Bellal, M.M. Effect of Ingestion of Thermally Oxidized Sunflower Oil on the Fatty Acid Composition and Antioxidant Enzymes of Rat Liver and Brain in Development. Ann Nutr Metab 2002, 46, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Koo, N.; Min, D.B. Reactive Oxygen Species, Aging, and Antioxidative Nutraceuticals. Compr Rev Food Sci Food Saf 2004, 3, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Dicklin, M.E.; Robinson, J.L.; Lin, X.; Odle, J. Ontogeny and Chain-Length Specificity of Gastrointestinal Lipases Affect Medium-Chain Triacylglycerol Utilization by Newborn Pigs. Jurnal of Animal Science 2006, 84, 818–825. [Google Scholar] [CrossRef]
- Moreau, H.; Gargouri, Y.; Lecat, D.; Junien, J.L.; Verger, R. Screening of Preduodenal Lipases in Several Mammals. Biochimica et Biophysics Acta 1988, 959, 241–252. [Google Scholar] [CrossRef]
- Mclaughlin, J.; Grazia Luca, M.; Jones, M.N.; Dockray, G.J.; Thompson, D.G. Fatty Acid Chain Length Determines Cholecystokinin Secretion and Effect on Human Gastric Motility. Gastroenterology 1999, 116, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Little, T.J.; Russo, A.; Meyer, J.H.; Horowitz, M.; Smyth, D.R.; Bellon, M.; Wishart, J.M.; Jones, K.L.; Feinle-Bisset, C. Free Fatty Acids Have More Potent Effects on Gastric Emptying, Gut Hormones, and Appetite than Triacylglycerides. Gastroenterology 2007, 133, 1124–1131. [Google Scholar] [CrossRef]
- Armand, M. Lipases and Lipolysis in the Human Digestive Tract: Where Do We Stand? Curr Opin Clin Nutr Metab Care 2007, 10, 156–164. [Google Scholar] [CrossRef]
- Jones, P.J.H.; Rideout, T. Jones, P.J.H., Rideout, T., 2012. Lipids, Sterols, and Their Metabolites. In: Ross, A.C., Caballero, B., Cousins, R.J., Tucker, K.L., Ziegler, T.R. (Eds.), Modern Nutrition in Health and Disease, Pp. 65–87. In Modern Nutrition in Health and Disease; Ross, A.C., Caballero, B., Cousins, R.J., Tucker, K.L., Ziegler, T.R., Eds.; 2012; pp. 65–87.
- Watanabe, S.; Lee, K.Y.; Chang, T.-M.; Berger-Ornstein, L.; Chey, W.Y.; Berger-Orn-Stein, L.; Role, W.Y.C. Role of Pancreatic Enzymes on Rel of Cholecystokinin-Pancreozymin in Esponse to Fat. Am J Physiol Gastrointest Liver Physiol 1988, 254, G837–G842. [Google Scholar] [CrossRef]
- Wilfart, A.; Montagne, L.; Simmins, H.; Noblet, J.; van Milgen, J. Effect of Fibre Content in the Diet on the Mean Retention Time in Different Segments of the Digestive Tract in Growing Pigs. Livest Sci 2007, 109, 27–29. [Google Scholar] [CrossRef]
- Kindel, T.; Lee, D.M.; Tso, P. The Mechanism of the Formation and Secretion of Chylomicrons. Atheroscler Suppl 2010, 11, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Lairon, D. Digestion and Absorption of Lipids. In Designing Functional Foods: Measuring and Controlling Food Structure Breakdown and Nutrient Absorption; Elsevier Ltd., 2009; pp. 68–93. ISBN 9781845694326. [Google Scholar]
- Zhang, J.Y.; Hu, J.; Kim, I.H. Supplemental Effects of Coated Omega-3 Polyunsaturated Fatty Acids to Basal Diet on Improving Growth Performance, Nutrients Digestibility, Fecal Lactobacillus Count and Fecal Score in Weaning Pigs. Anim Feed Sci Technol 2020, 269. [Google Scholar] [CrossRef]
- Zhang, L.L.; Zhang, H.; Li, Y.; Wang, T. Effects of Medium-Chain Triglycerides on Intestinal Morphology and Energy Metabolism of Intrauterine Growth Retarded Weanling Piglets. Arch Anim Nutr 2017, 71, 231–245. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, X.; Yang, R.; Ma, Q.; Qi, W.; Sanidad, K.Z.; Park, Y.; Kim, D.; Decker, E.A.; Zhang, G. Thermally Processed Oil Exaggerates Colonic Inflammation and Colitis-Associated Colon Tumorigenesis in Mice. Cancer Prevention Research 2019, 12, 741–750. [Google Scholar] [CrossRef]
- Mboma, J.; Leblanc, N.; Angers, P.; Rocher, A.; Vigor, C.; Oger, C.; Reversat, G.; Vercauteren, J.; Galano, J.M.; Durand, T.; et al. Effects of Cyclic Fatty Acid Monomers from Heated Vegetable Oil on Markers of Inflammation and Oxidative Stress in Male Wistar Rats. J Agric Food Chem 2018, 66, 7172–7180. [Google Scholar] [CrossRef]
- McGarry, J.D.; Brown, N.F. The Mitochondrial Carnitine Palmitoyltransferase System. From Concept to Molecular Analysis. Eur J Biochem 1997, 244, 1–14. [Google Scholar] [CrossRef]
- Mannaerts, G.P.; Veldhoven, P.P. Van; Casteels, M. Peroxisomal Lipid Degradation Via-and-Oxidation in Mammals. Cell Biochem Biophys 2000, 32, 73–87. [CrossRef] [PubMed]
- Gondret, F.; Louveau, I.; Mourot, J.; Duclos, M.J.; Lagarrigue, S.; Gilbert, H.; Van Milgen, J. Dietary Energy Sources Affect the Partition of Body Lipids and the Hierarchy of Energy Metabolic Pathways in Growing Pigs Differing in Feed Efficiency. J Anim Sci 2014, 92, 4865–4877. [Google Scholar] [CrossRef]
- Vianey-Saban, C.; Guffon, N.; Fouilhoux, A.; Acquaviva, C. Fifty Years of Research on Mitochondrial Fatty Acid Oxidation Disorders: The Remaining Challenges. J Inherit Metab Dis 2023, 46, 848–873. [Google Scholar] [CrossRef]
- Battino, M.; Quiles, J.L.; Huertas, J.R.; Ramirez-Tortosa, M.C.; Cassinello, M.; Mañas, M.; Lopez-Frias, M.; Mataix, J. Feeding Fried Oil Changes Antioxidant and Fatty Acid Pattern of Rat and Affects Rat Liver Mitochondrial Respiratory Chain Components. J Bioenerg Biomembr 2002, 34, 127–134. [Google Scholar] [CrossRef]
- Huertas, J.R.; Battino, M.; Mataix, F.J.; Lenaz, G. Cytochrome Oxidase Induction after Oxidative Stress Induced by Adriamycin in Liver of Rats Fed with Dietary Olive Oil. Biochem Biophys Res Commun 1991, 181, 375–382. [Google Scholar] [CrossRef]
- Koneru, P.; Ranganath, P. Fatty Acid Oxidation Disorders: An Update. Genetic Clinics 2025, 18, 10–22. [Google Scholar]
- Bebernitz, G.R.; Schuster, H.F. The Impact of Fatty Acid Oxidation on Energy Utilization: Targets and Therapy. Curr Pharm Des 2002, 8, 1199–1227. [Google Scholar] [CrossRef] [PubMed]
- Abbas, G.; Salman, A.; Rahman, S.U.; Ateeq, M.K.; Usman, M.; Sajid, S.; Zaheer, Z.; Younas, T. Aging Mechanisms: Linking Oxidative Stress, Obesity and Inflammation. Matrix Science Medica 2017, 1, 30–33. [Google Scholar] [CrossRef]
- Saponaro, C.; Gaggini, M.; Carli, F.; Gastaldelli, A. The Subtle Balance between Lipolysis and Lipogenesis: A Critical Point in Metabolic Homeostasis. Nutrients 2015, 7, 9453–9474. [Google Scholar] [CrossRef]
- Ostrowska, E.; Cross, R.F.; Muralitharan, M.; Bauman, D.E.; Dunshea, F.R. Effects of Dietary Fat and Conjugated Linoleic Acid on Plasma Metabolite Concentrations and Metabolic Responses to Homeostatic Signals in Pigs. British Journal of Nutrition 2002, 88, 625–634. [Google Scholar] [CrossRef]
- Powles, J.; Wiseman, J.; Cole, D.J.A.; Jagger, S. Prediction of the Apparent Digestible Energy Value of Fats given to Pigs. Animal Science 1995, 61, 149–154. [Google Scholar] [CrossRef]
- Mendoza, S.M.; van Heugten, E. Effects of Dietary Lipid Sources on Performance and Apparent Total Tract Digestibility of Lipids and Energy When Fed to Nursery Pigs. J Anim Sci 2014, 92, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, H.; Fernández, J.A. Chemical Composition and Energy Value of Different Fat Sources for Growing Pigs. Acta Agriculturae Scandinavica A: Animal Sciences 2000, 50, 129–136. [Google Scholar] [CrossRef]
- Hao, Y.; Xing, M.; Gu, X. Research Progress on Oxidative Stress and Its Nutritional Regulation Strategies in Pigs. Animals 2021, 11. [Google Scholar] [CrossRef]
- Mind, B.; Ahn, D.U. Mechanism of Lipid Peroxidation in Meat and Meat Products -A Review. Food Sci Biotechnol 2005, 14, 152–163. [Google Scholar]
- Pamplona, R.; Costantini, D. Molecular and Structural Antioxidant Defenses against Oxidative Stress in Animals. Am J Physiol Regul Integr Comp Physiol 2011, 301, 843–863. [Google Scholar] [CrossRef] [PubMed]
- Macho-González, A.; Garcimartín, A.; López-Oliva, M.E.; Bastida, S.; Benedí, J.; Ros, G.; Nieto, G.; Sánchez-Muniz, F.J. Can Meat and Meat-Products Induce Oxidative Stress? Antioxidants 2020, 9, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Silva-Guillen, Y.V.; Arellano, C.; Boyd, R.D.; Martinez, G.; Van Heugten, E. Growth Performance, Oxidative Stress and Immune Status of Newly Weaned Pigs Fed Peroxidized Lipids with or without Supplemental Vitamin e or Polyphenols. J Anim Sci Biotechnol 2020, 11. [Google Scholar] [CrossRef]
- Arowolo, F.K.; Yang, X.; Blaser, M.E.; Nicholson, A.M.; Hosokawa, E.; Booth, J.R.; Jobsis, C.T.; Russell, R.L.; Meudt, J.J.; Reichert, J.L.; et al. Presence of Lipid Oxidation Products in Swine Diet Lowers Pork Quality and Stability during Storage. Meat Sci 2020, 160. [Google Scholar] [CrossRef]
- Dragoev, S.G. Lipid Peroxidation in Muscle Foods: Impact on Quality, Safety and Human Health. Foods 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Pieszka, M. Polish Journal of Food and Nutrition Sciences Effect of Vegetable Oils Supplementation in Pig Diets on Lipid Oxidation and Formation of Oxidized Forms of Cholesterol in Meat. Pol. J. Food Nutr. Sci 2007, 57, 509–516. [Google Scholar]
- Zheng, Y.; Sun, J.; Luo, Z.; Li, Y.; Huang, Y. Emerging Mechanisms of Lipid Peroxidation in Regulated Cell Death and Its Physiological Implications. Cell Death Dis 2024, 15, 859. [Google Scholar] [CrossRef]
- Zhang, R.; Vooijs, M.A.; Keulers, T.G.H. Key Mechanisms in Lysosome Stability, Degradation and Repair. Mol Cell Biol 2025, 45, 212–224. [Google Scholar] [CrossRef]
- Suski, J.M.; Lebiedzinska, M.; Bonora, M.; Pinton, P.; Duszynski, J.; Wieckowski, M.R. Relation between Mitochondrial Membrane Potential and ROS Formation. Methods in Molecular Biology 2012, 810, 183–205. [Google Scholar] [CrossRef]
- Tracey, T.J.; Steyn, F.J.; Wolvetang, E.J.; Ngo, S.T. Neuronal Lipid Metabolism: Multiple Pathways Driving Functional Outcomes in Health and Disease. Front Mol Neurosci 2018, 11. [Google Scholar] [CrossRef]
- Srikanthan, K.; Shapiro, J.I.; Sodhi, K. The Role of Na/K-ATPase Signaling in Oxidative Stress Related to Obesity and Cardiovascular Disease. Molecules 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Gómez, I.; Janardhanan, R.; Ibañez, F.C.; Beriain, M.J. The Effects of Processing and Preservation Technologies on Meat Quality: Sensory and Nutritional Aspects. Foods 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Adepu, K.K.; Anishkin, A.; Adams, S.H.; Chintapalli, S.V. A Versatile Delivery Vehicle for Cellular Oxygen and Fuels or Metabolic Sensor? A Review and Perspective on the Functions of Myoglobin. Physiol Rev 2024, 104, 1611–1642. [Google Scholar] [CrossRef]
- Lynch, M.P.; Faustman, C. Effect of Aldehyde Lipid Oxidation Products on Myoglobin. J Agric Food Chem 2000, 48, 600–604. [Google Scholar] [CrossRef]
- Young, O.A.; West, J. Meat Color. In Meat Science and Applications; MARCEL DEKKER, INC.: New York, 2001; pp. 52–82. [Google Scholar]
- Xiong, Y.L. Muscle Proteins. In Proteins in Food Processing; Elsevier, 2017; pp. 127–148. ISBN 9780081007228. [Google Scholar]
- Domínguez, R.; Pateiro, M.; Munekata, P.E.S.; Zhang, W.; Garcia-Oliveira, P.; Carpena, M.; Prieto, M.A.; Bohrer, B.; Lorenzo, J.M. Protein Oxidation in Muscle Foods: A Comprehensive Review. Antioxidants 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Kong, B.; Liu, Q.; Liu, J. Physicochemical Change and Protein Oxidation in Porcine Longissimus Dorsi as Influenced by Different Freeze-Thaw Cycles. Meat Sci 2009, 83, 239–245. [Google Scholar] [CrossRef]
- Mohan, K.; Banerjee, R.; Maheswarappa, N.B. Recent Developments in Postmortem Aging and Evaluation Methods. In Meat Quality Analysis: Advanced Evaluation Methods, Techniques, and Technologies; Elsevier, 2019; pp. 81–99. ISBN 9780128192337. [Google Scholar]
- Fernando, L.N.; Berg, E.P.; Grün, I.U. Quantitation of Hexanal by Automated SPME for Studying Dietary Influences on the Oxidation of Pork. Journal of Food Composition and Analysis 2003, 16, 179–188. [Google Scholar] [CrossRef]
- Schilling, M.W.; Pham-Mondala, A.J.; Dhowlaghar, N.; Campbell, Y.L.; Dinh, T.T.; Tolentino, A.C.; Williams, J.B.; Xiong, Y.L. Changes in the Volatile Composition of Fresh Pork Sausage with Natural Antioxidants during Long-Term Frozen Storage. Meat and Muscle Biology 2019, 3, 194–209. [Google Scholar] [CrossRef]
- Degroote, J.; Wang, W.; Vergauwen, H.; De Smet, S.; Van Ginneken, C.; Michiels, J. Impact of a Dietary Challenge with Peroxidized Oil on the Glutathione Redox Status and Integrity of the Small Intestine in Weaned Piglets. Animal 2019, 13, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Lindblom, S.C.; Gabler, N.K.; Dilger, R.N.; Olson, Z.F.; Loving, C.L.; Kerr, B.J. Influence of Feeding Thermally Peroxidized Soybean Oil on Oxidative Status in Growing Pigs. J Anim Sci 2018, 96, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Keller, U.; Brandsch, C.; Eder, K. The Effect of Dietary Oxidized Fats on the Antioxidant Status of Erythrocytes and Their Susceptibility to Haemolysis in Rats and Guinea Pigs. J Anim Physiol Anim Nutr (Berl) 2004, 88. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic “black box” representation of the inputs and outputs of the biological system associated with the consumption of oxidized fats and oils.
Figure 1.
Schematic “black box” representation of the inputs and outputs of the biological system associated with the consumption of oxidized fats and oils.
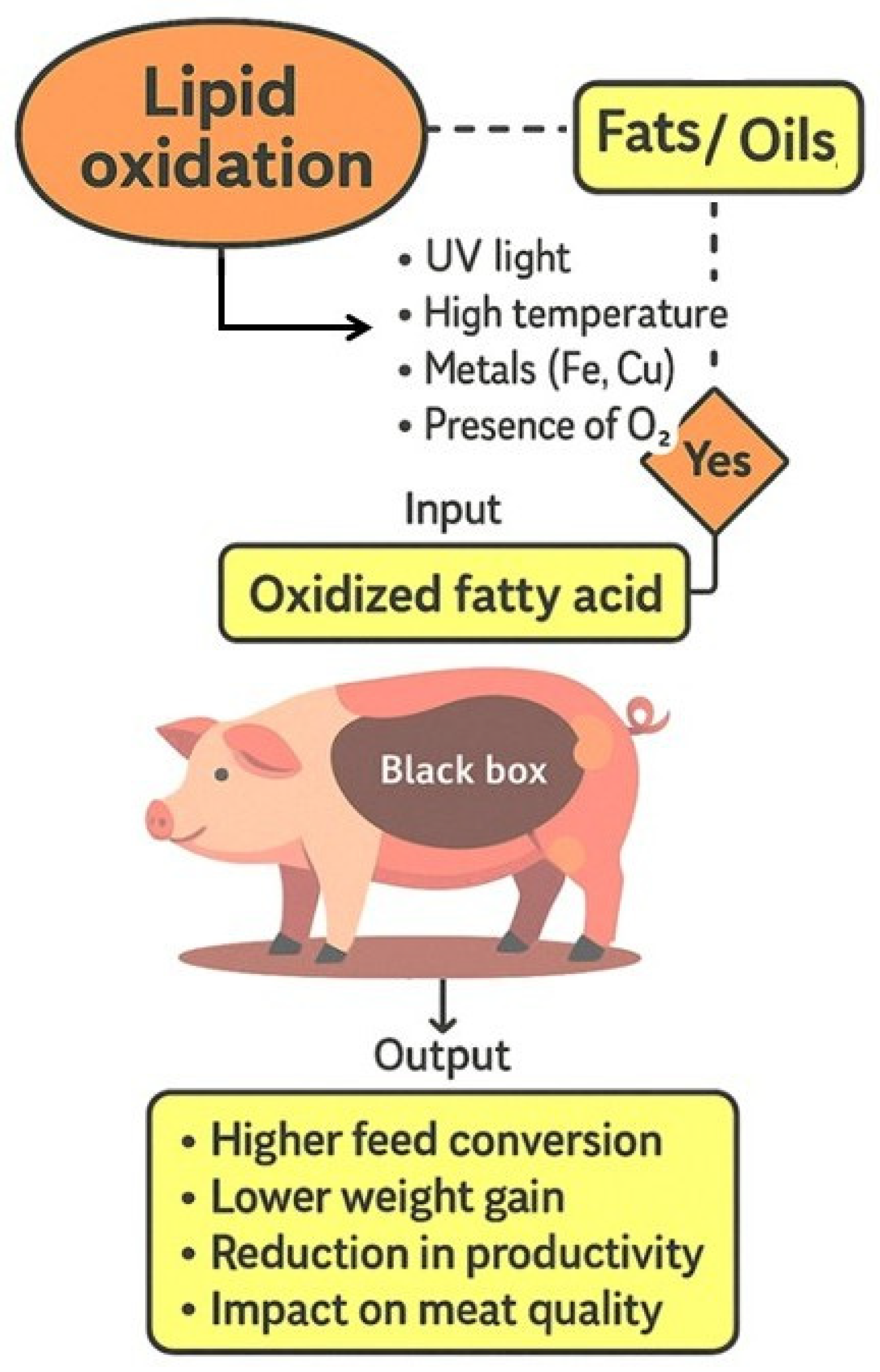
Figure 4.
Energy utilization in pigs consuming oxidized fats and oils. Developed based on the findings reported by Kil et al.; Li et al.; Lindblom et al. and Liu et al. [3,9,11,12]. .
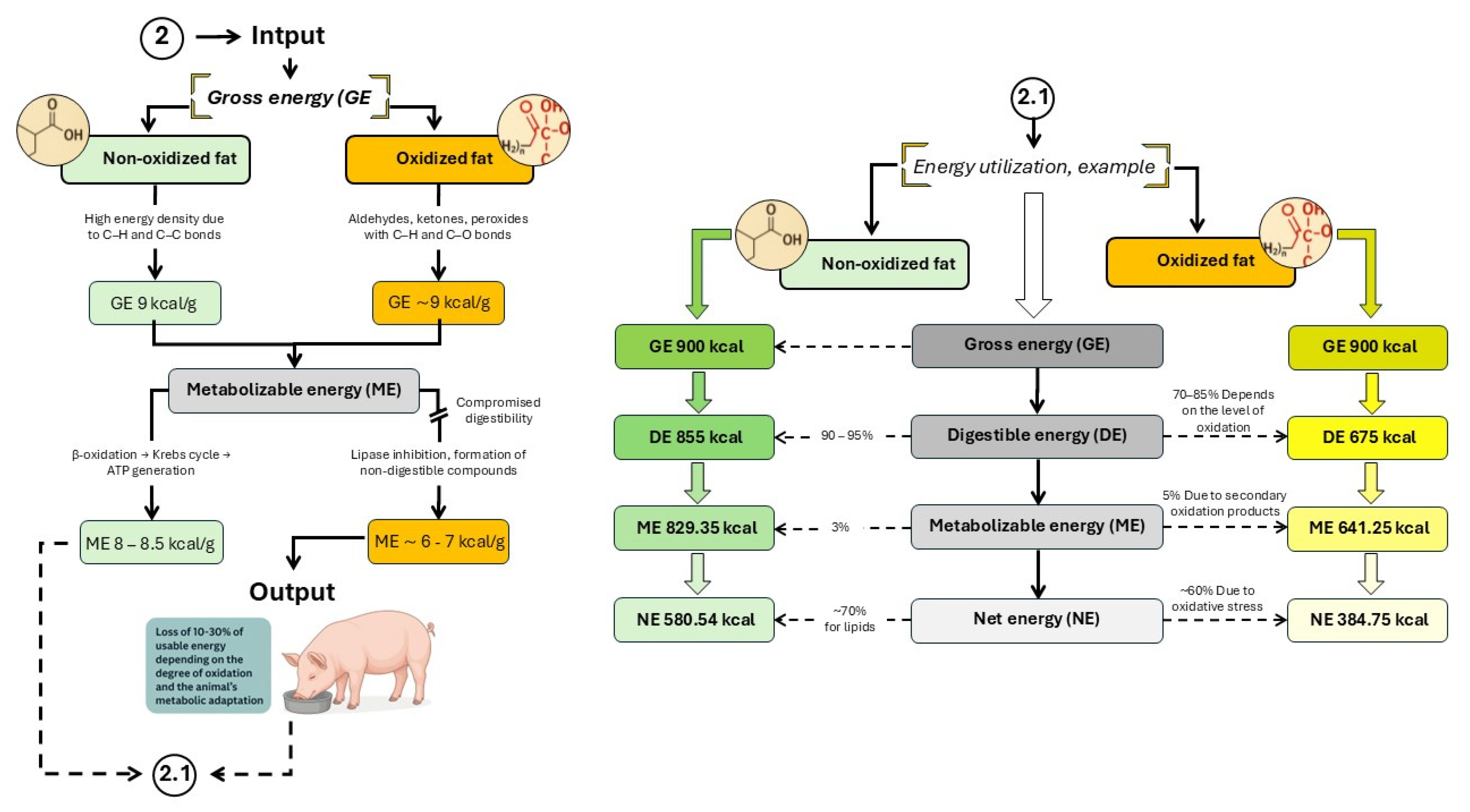
Table 1.
Chemical indicators of lipid oxidation in different fat sources subjected to thermal treatments. Modified from Liu et al. (2014b).
Table 1.
Chemical indicators of lipid oxidation in different fat sources subjected to thermal treatments. Modified from Liu et al. (2014b).
| Corn oil | Canola oil | Chicken fat | Tallow | |||||||||
| OL | SO | RO | OL | SO | RO | OL | SO | RO | OL | SO | RO | |
| PV1, mEq/kg | 1 | 151 | 2 | 1 | 239 | 12 | 1 | 57 | 2 | 1 | 29 | 3 |
| p-anisidina2 | <1 | 61.4 | 142.9 | 1 | 37.0 | 154.8 | 3 | 88 | 22 | 4 | 120 | 19 |
| TBARS3, μmol/kg | 16 | 225 | 119 | 45 | 968 | 622 | 79 | 151 | 58 | 58 | 61 | 41 |
| Hexanal, mg/kg | <1 | 390 | 83 | 1 | 180 | 59 | 3 | 88 | 22 | 4 | 120 | 19 |
| 2, 4-decadienal, mg/kg | 72 | 3,728 | 1,345 | 7 | 1,091 | 511 | 30 | 442 | 169 | 47 | 261 | 125 |
| HNE4, μmol/kg | 0 | 194 | 594 | 0 | 105 | 221 | 0 | 2 | 0 | 0 | 13 | 6 |
| AOM5, mEq/kg | 103 | 575 | 528 | 112 | 419 | 533 | 4 | 298 | 5 | <2 | 6 | 446 |
| OSI6, h | 8.4 | <1.0 | <1.0 | 9.2 | <1.0 | <1.0 | 24.6 | <1.0 | <1.0 | 12.1 | <1.0 | <1.0 |
OL original lipids (lipids were stored as received without antioxidants or heating); SO = slow oxidation (SO lipids were heated for 72 h at 95°C with constant compressed air flow rate at 12 L/min); RO = lipids (RO lipids were heated for 7 h at 185°C with constant compressed air flow rate at 12 L/min). 1PV = Peroxide value. 2There is no unit for the p-anisidine value. 3TBARS = Thiobarbituric acid reactive substances. 4HNE = 4-hydroxynonenal. 5AOM = Active oxygen method (measured as PV after 20 h of oxidation). 6OSI = Oxygen stability index (time to exponential increase in conductivity when analyzed at 110°C).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



