
Submitted:
15 January 2026
Posted:
15 January 2026
You are already at the latest version
Abstract
The Apennine wolf (Canis lupus italicus) increasingly inhabits human-modified land-scapes, where exposure to anthropogenic environments may influence pathogen cir-culation and antimicrobial resistance (AMR). Despite this relevance, no shotgun met-agenomic data are available for this subspecies during rehabilitation. A juvenile male wolf admitted to a Wildlife Rehabilitation Center (WRC) after traumatic injury and treated with multiple antibiotics was sampled at admission (T0) and after 11 months of rehabilitation (T1). Shotgun metagenomic sequencing (Illumina NovaSeq) was used to characterize fecal microbial communities, potential pathogens, and antimicrobial resistance genes (ARGs) using Kraken2 and CARD-RGI. Bacterial diversity increased from T0 to T1. Microbial composition shifted from Enterobacterales-dominated pro-files to more diverse communities. Reads associated with animal and human patho-gens were detected at both time points, together with human-associated taxa and viral reads at T1. ARGs were abundant (444 at T0; 417 at T1), mainly involving efflux pumps and β-lactamases. Genes related to Highest Priority Critically Important Anti-microbials—including mcr variants, van clusters, and oxazolidinone resistance deter-minants—were identified. Shotgun metagenomics revealed marked microbiome changes and high ARG diversity in an Apennine wolf during rehabilitation. These findings highlight wolves as potential sentinels of environmental AMR and emphasize the importance of biosecurity measures in WRCs.
Keywords:
wild animal
; apennine wolf
; wildlife rehabilitation
; shotgun metagenomics
; fecal microbiome
; resistome
; antimicrobial resistance genes
; environmental antimicrobial resistance
; one health
1. Introduction
Throughout the 19th and mid-20th centuries, large carnivores in Europe declined significantly due to human persecution, habitat loss, and fragmentation [1,2]. However, in recent decades, species such as Eurasian lynx (Lynx lynx), brown bear (Ursus arctos), and grey wolf (Canis lupus) have rebounded, reoccupying former territories [2]. This recovery is largely attributed to conservation policies, protective legislation, tailored management practices (e.g., damage prevention and compensation programs), increasing public acceptance and changes in land use [1,2,3,4]. While urban areas and human population density have generally increased, many rural areas have seen depopulation and a return of natural vegetation, enhancing habitat suitability and connectivity for large carnivores in Eastern and Central Europe, allowing for long-distance dispersal [5,6]. Recently, wolves have been recorded in nearly all EU countries (except islands), with established populations in most mainland states [1,7]. Wolves have proven particularly adaptable and successful at colonizing human-dense landscapes [1], with reproducing packs near residential and industrial zones in Germany, France, western Poland, western Finland, southern Sweden, and Italy [8,9].
This expansion creates novel interfaces between wildlife, livestock, and humans, influencing contact rates, species diversity, and population density, all of which affect cross-species pathogen transmission risk [10]. Worldwide, Wildlife Rehabilitation Centres (WRCs) treat diseased, injured, or weakened animals, providing valuable insights into wildlife–human interactions [11,12]. Additionally, health data collected at WRCs can also shed light on pathogens circulating in wild populations [13]. Pathogens are natural components of ecosystems and influence the population dynamics of wild vertebrates. Their emergence is shaped by host–environment interactions including factors such as climate change, land use, and animal management [1].
As apex predators, wolves serve as indicators of ecosystem health, reflecting pathogen occurrence and distribution due to their adaptability to different habitats and contacts with various prey species and scavengers [14,15]. Furthermore, the social behavior of wolves, which often live in groups, increases the likelihood of detecting in a single animal pathogens related to within-pack transmission [16,17,18].
Wolves have also been recognized as useful indicators of environmental contamination by antibiotic-resistant bacteria [19,20,21,22,23], which remains a major public health concern and the main focus of numerous studies aimed at characterizing environmental contamination through investigations on wildlife [24].
Next-generation sequencing approaches have been widely applied to characterize pathogens in wolves and to trace the microbial community evolution from wolves to domestic dogs [25,26,27,28,29]. Deep sequencing has provided unprecedented insights into the structure and function of microbial communities over time and across environments, insights that traditional culture-dependent methods could not achieve [30,31]. Studies of wildlife-associated bacterial communities have shown temporal fluctuations, significant social influences, and shifts related to diet changes resulting from habitat degradation [32,33,34,35]. Metagenomic approaches have also been employed to explore the resistome in wild animals, providing data on the influence of human activities and environmental conditions [36,37,38,39].
Building on these observations, the application of shotgun metagenomics to investigate the microbiome and resistome in wild animals at WRCs may provide valuable insights into populations of origin. Additionally, it may contribute to the development of evidence-based conservation and biosecurity strategies. In this view, the aim of this study was to investigate changes in fecal microbiome and resistome of a long-term rehabilitated Apennine wolf (Canis lupus italicus) through shotgun metagenomic sequencing.
2. Materials and Methods
A one-year-old male Apennine wolf (Canis lupus italicus) was rescued alive and admitted to the veterinary clinic of the Wildlife Research Center (Maiella National Park, Italy) for medical care and rehabilitation. The wolf had been found trapped in an illegal snare set for wild boars. It was subsequently recovered by a veterinary officer of the local health authority, who administered a chemical restraint using Zoletil® (a combination of tiletamine and zolazepam) at an approximate dosage of 5 mg/kg. The sedated animal was then transported to the WRC veterinary facility.
Fecal samples were collected at two distinct time points: T0, upon the animal's initial recovery at the Wildlife Research Center (WRC), and T1, following the completion of its rehabilitation in the veterinary facility. A period of 11 months elapsed between the T0 and T1 sample collections. Both procedures were conducted without causing disturbance to the animal. Subsequent to undergoing a pancarpal arthrodesis, the wolf regained near-total motor function and was ultimately released into an authorized wildlife enclosure for wolves.
Total DNA was extracted from each fecal sample by MagPurix 12A Nucleic Acid Extraction System (Zinexts Life Science Corp., Taipei, Taiwan). The DNA extracts were measured with NanoDrop One (Thermo Scientific) and Qubit Fluorimeter (Invitrogen) for yield and purity check. DNA libraries, prepared with a Nextera Library Preparation Kit (Illumina), were subsequently sequenced with the NovaSeq 6000 (Illumina), using 2 × 150 paired-end sequencing per flow cell. The reads were deposited under BioProject PRJNA1101237.
FASTQ quality check was performed using FastQC v0.12.1 and taxonomic classification was conducted through the Kraken2 Metagenomics pipeline [40,41,42]. Krona plots of the classification were generated for each sample [43].
Antimicrobial resistance analysis was achieved by CARD-RGI (Comprehensive Antibiotic Resistance Database – Resistance Gene Identifier) software [44,45], and Krona plots of the resistance mechanisms were designed for each sample [43].
Statistical analyses were performed using Stata/SE 17.0 (StataCorp LLC, College Station, TX, USA). Microbial and resistance gene diversity was assessed using Shannon and Simpson diversity indices. Differences in read counts of potential pathogens and antimicrobial resistance gene families between T0 and T1 were evaluated using either the Mann–Whitney U test or two-sample t-test, depending on data distribution assessed through Shapiro–Wilk normality testing. Statistical significance was set at p < 0.05.
3. Results
3.1. Animal Clinical Data
The animal had sustained a lacerated wound from an illegal metal snare trap in the right carpal region, resulting in exposure of the radiocarpal joint. The animal’s Body Condition Score was 5/9 [46] with a weight of 26.3 kg. The animal was treated with a surgical procedure and with enrofloxacin (50 mg/mL) at 5 mg/kg q12 h and clindamycin (600 mg/4 mL) at 10 mg/kg q12 h for 10 days. Based on the antibiotic susceptibility testing, the antibiotic therapy was replaced with cefovecin (80 mg/mL) at 8 mg/kg (single administration). The antibiotic susceptibility tests were performed on colonies of Streptococcus canis, Escherichia coli, and Pseudomonas aeruginosa recovered from the carpal wound and intra-articular swab [22]. These isolates showed phenotypic resistance to amoxicillin/clavulanic acid, ampicillin, benzylpenicillin, chloramphenicol, clindamycin, cefotaxime, doxycycline, erythromycin, enrofloxacin, cephalexin, cephalotin, cefovecin, gentamicin, tetracycline, marbofloxacin, neomycin, trimethoprim/sulfamethoxazole, nitrofurantoin, and imipenem [22].
The diet consisted primarily of sheep carcasses, provided on average twice weekly, and roe deer carcasses, provided approximately once a week. No dietary supplements or commercial pet foods were administered.
The key environmental shifts introduced by the wolf's 11-month rehabilitation period—compared to its preceding 11–12 months in the wild—included: (i) the dietary change; (ii) the antibiotic treatments; (iii) confinement in a washable-walled enclosure with a straw bedding substrate, cleaned on average every 48–72 hours; and (iv) proximity to a human caretaker (at a distance of 50–150 cm) during feeding and medication, which occurred approximately every 48 hours. While the caretaker used personal protective equipment, a face mask was not consistently worn.
3.2. Basic Data Statistics
Quality analysis allowed to obtain a total of 229,557,801 reads per sample T0 and 229,646,438 reads for sample T1.
3.3. Taxonomic Identification
Six taxonomic levels of classification and abundances were produced for each sample, from phylum to species. The overall taxonomic distribution of T0 and T1 samples revealed that the highest proportion of contigs mapped to bacteria (98.6% T0 and 81.9% T1), followed by viruses and archaea (~1%) (Figure 1).
Details of taxonomic classification for both time points are reported in Table S1. A total of 74 phyla, 173 families, 476 genera, 1,274 species were annotated. In detail, the phyla Chloroflexota, Planctomycetota, Verrucomicrobiota, Chlamydiota, Thermodesulfobacteriota, Nitrososphaerota, Nucleocytoviricota, Artverviricota and Cossaviricota were described only in T0, while the phyla Gemmatimonadota, Acidobacteriota, Verrucomicrobiota, Thermodesulfobacteriota, Phixviricota and Pisuviricota were identified only in T1. The orders with the highest abundance were Enterobacterales (94.2%) in T0 and Enterobacterales (26.3%), Bacteroidales (22.4%) and Eubacteriales (20.4%) in T1. The highest abundance of families was described for Enterobacteriaceae (92.8%) in T0 and for Enterobacteriaceae (26.2%), Hominidae (19%) and Bacteroidaceae (17.7%) in T1. The relative abundance of genera is reported in Figure 2 and the highest abundance was described for Escherichia spp. in T0 (92%) and T1 (26.2%).
The highest abundant bacterial species were Escherichia coli (79.9%) in T0 and Escherichia coli (26.2%), Homo sapiens (19%), Phocaeicola vulgatus (15.7%), Flavonifractor plautii (13.2%) in T1 (Figure 3).
The majority of viral species detected in wolf feces at T0 and T1 belonged to the genus Phapecoctavirus (85%) at T0 and to genus Krischvirus at T1 (71%) (Figure 4). In addition, sequences related to Severe Acute Respiratory Syndrome Coronavirus 2 (0.03%) were reported in wolf feces at T1.
The number of reads related to potential pathogens identified in wolf feces are reported in Table 1. No statistically significant difference was observed between the number of reads of potential pathogens detected at T0 and T1 (p=0.2865). The diversity indices increased from T0 to T1 (Shannon index: 0.97 at T0 and 2.3 at T1; Simpson index:0.35 at T0 and 0.84 at T1).
3.4. Antibiotic Resistance Genes
The most described resistance mechanism was related to the antibiotic efflux in both samples (T0: 71.2% T1: 77.6%), followed by antibiotic inactivation (T0: 17.2% T1:14.2%) (Figure 5).
The antibiotic efflux resistance mechanism is related to resistance nodulation cell division (RND) (T0: 50% T1: 54%), major facilitator superfamily (MFS) (T0: 28% T1: 26%), ATP-binding cassette (ABC) (T0: 10% T1:10%), small multidrug resistance (SMR) (T0:1% T1:1%) and multidrug and toxic compound extrusion (MATE) transporter (T0:0.5% T1:0.5%). In addition, these efflux pump confer resistance to aminocoumarin, aminoglycosides, fluoroquinolones, cephalosporins, glycylcycline, penam, tetracycline, rifamycin, phenicol, macrolide, disinfecting agents and antiseptics in bacterial species detected in both samples.
The antibiotic inactivation mechanism is correlated to resistance against monobactam, carbapenem, cephalosporins, penam, penem, aminoglycosides, macrolide, tetracycline and cephamycin in both samples.
The antibiotic target alteration mechanism was associated to peptide, macrolide, lincosamide, streptogramin, aminoglycoside resistance in sample T0 and T1. In addition, the Erm 23S ribosomal RNA methyltransferase was identified only in sample T1.
The antibiotic target protection mechanism was associated to tetracycline, macrolide and streptogramin resistance in sample T0 and to tetracycline, macrolide, streptogramin and fluoroquinolones resistance in sample T1.
The antibiotic target replacement was associated to sulfonamide and diaminopyrimidine resistance in both samples T0 and T1 and the reduced permeability to antibiotics was related to peptide, monobactam, carbapenem, cephalosporin, cephamycin, penam and penem resistance in both samples.
A total of 444 resistance genes were detected in sample T0 and 417 resistance genes were described in sample T1 (Table S2). The relative abundance of drug classes associated with resistance genes are reported in Figure 6. The most detected drug class was penam in T0 and aminoglycosides in T1.
The most detected resistance genes in fecal samples are reported in Figure 7 and are related to antibiotic efflux pumps conferring resistance to fluoroquinolone (acrB, acrF, evgS, ermA, ermB, mdtF), cephalosporin (acrB, acrF), glycylcycline (acrB), penam (acrB, acrF mdtF, evgS), tetracycline (acrB, evgS), rifamycin (acrB), phenicol (acrB), peptide (acrB, YojI), aminoglycoside (APH(6)-Id, kdpE, baeS), macrolide (mdtF, evgS, mphA), aminocoumarin (mdtB, mdtC, baeS), sulfonamide (sul1), nitroimidazole (msbA), phosphonic acid (mdtG), disinfecting agents and antiseptics (acrB, mdtN, mdtO).
The Shannon index for resistance genes was 4.32 at T0 and decreased to 3.79 at T1. Similarly, the Simpson index for resistance genes showed values of 0.97 at T0 and 0.95 at T1. Finally, the resistance genes associated with medically important antimicrobials were detected in fecal samples. Specifically, these included resistance genes linked to exclusively authorized for human use molecules, as well as those authorized for use in both animals and humans and categorized as Highest Priority Critically Important Antimicrobials (HPCIAs) and Critically Important Antimicrobials (CIAs) (Table 2). The Shannon index for medically important antibiotics was 4.34 at T0 and decreased to 3.76 at T1. Similarly, the Simpson index showed values of 0.97 at T0 and 0.96 at T1. No statistically significant difference was observed between the number of reads of medically important antimicrobial genes detected at T0 and T1, except for the class of aminoglycosides (Table 2).
4. Discussion
In this study, the shotgun metagenomic tool was applied for the first time to describe the microbial community, as well as the changes in the resistome and taxonomic composition in fecal samples of an Apennine wolf exposed to antibiotics and to modifications of habitat and feeding behavior related to recovery in a WRC. Despite the limited number of samples, the results obtained provide valuable contributions to updating the data available in the literature on Apennine wolf and to designing future studies.
Many research experiences have demonstrated that wolves can persist in human-dominated landscapes [47,48,49,50]. This adaptability is likely facilitated by high degrees of ecological and behavioral plasticity. Consequently, wolves not only represent a valuable source of health data within wildlife rehabilitation centers but also emerge as critical ecological sentinels. As apex predators, they can provide crucial insights into the state of the environmental microbiome and resistome. Their exposure to human activities and infrastructures could position them as key indicators of how anthropogenic pressures are altering microbial communities and driving the selection and dissemination of microorganisms and antibiotic resistance genes.
The results of taxonomic identification revealed a substantial discrepancy of groups distribution with similar previous investigations that reported the predominance of Firmicutes, Bacteroides and Fusobacteria phyla [30,51]. Similar findings were obtained in Italy, comparing the fecal microbiome of four pack living in two different geographic areas, the Maremma Regional Park in Mediterranean coastal range and the Central Italian Alps, respectively. This composition is considered consistent with carnivorous diet, rich in proteins derived from mammalian prey [52]. Conversely, our study showed in T0 sample a marked dominance of Enterobacterales order, belonging to Pseudomonadota phylum, that remains the largest group in T1 sample, but at the lower prevalence, followed by Bacteroidales and Eubacteriales microorganisms.
The recovery area for the wolf in this study is covered by existing dietary research conducted by the Maiella National Park [53]. Wild boar was the predominant prey item throughout the year for all wolf packs investigated, accounting for 67% of the diet. Other wild ungulates appeared less significant: roe deer and red deer comprised 13.3% and 7.7%, respectively, while chamois accounted for 4%. Other than wild prey, livestock constituted only 5.87% and fruits 3.13%. The prevalence of wild ruminants and minimal reliance on domestic animals within the diet was additionally corroborated for the same geographic area by research involving GPS-equipped radio-collars on animals belonging to five distinct packs [54]. Therefore, the composition of microbiota in wolf under study appears to be unrelated to the diet considered typical for the species. Other factors, such as the individual variability and the health status of the animal at the time of the capture, may have influenced these results.
The differences in the Shannon and Simpson indices between T0 and T1 indicate an increase in bacterial diversity over time, likely driven by the introduction of new bacterial species or a more balanced distribution among the species already present. These variations may be influenced by changes in diet, habitat, or antibiotic therapy, as previously suggested for wolves and other mammal species [26,29,31].
Although no statistically significant differences were observed in the reads of potentially pathogenic species identified at T0 and T1, it is noteworthy that several potential pathogens, both viral and bacterial species, relevant for animal and human health were identified. The available studies on the Apennine wolf have reported positive samples for selected viral (canine distemper virus, protoparvoviruses, canine adenoviruses, canine coronavirus, pseudorabies virus, circovirus, cyclovirus, and hepatitis E virus) and bacterial pathogens (Listeria monocytogenes, Yersinia spp.) [55,56,57,58,59,60,61,62,63]. Furthermore, phenotypic resistance to penicillins, cephalosporins, phenicols, macrolides, quinolones, carbapenems, tetracyclines, and trimethoprim/sulfamethoxazole has been documented in Staphylococcus pseudintermedius, Enterococcus faecalis, Escherichia coli, Klebsiella oxytoca, Streptococcus dysgalactiae, Leclercia adecarboxilata, Streptococcus canis, Pseudomonas aeruginosa, and other Enterobacteriaceae isolated from this species [64]. In addition to these species, reads related to Brucella, Campylobacter, Coxiella, Legionella genera were reported in the current study. The detection of viruses such as human mastadenovirus C and severe acute respiratory syndrome-related coronavirus underscore the influence of anthropogenic environments on the fecal microbial profile of wolves, as confirmed by the high number of reads related to Homo sapiens.
Genetic determinants of virulence (Shiga toxin-producing Escherichia coli) and resistance (extended-spectrum β-lactamase-producing Enterobacteriaceae) have also already been detected in samples from Apennine wolves by means of end-point PCR [64]. In this study, the application of advanced sequencing methodologies allowed us to obtain new insights on antibiotic resistance determinants in this species. The Shannon and Simpson indices for resistance genes detected at T0 and T1 suggest a relatively high diversity at both time points, with a slight decrease in resistance gene diversity over time. This could indicate a minor shift in the composition or distribution of certain resistance genes while maintaining overall high variability. These findings suggest that the use of broad-spectrum antibiotics, followed by therapy guided by antibiogram results, may be an effective strategy for preserving the animal resistome during the recovery time. Furthermore, regarding critically important antibiotics, no high variations were observed in the Shannon and Simpson indices, mirroring the overall trend for resistance genes. A statistically significant difference was found only in the mean reads associated with the aminoglycoside class between T0 and T1. Indeed, antibiotic selection pressure is not the only evolutionary force involved in shaping and modifying the resistome [65,66]. Ecological factors, human interactions, and the interconnectedness of ecosystems also contribute to shaping the distribution patterns of resistance genes [67].
The genes optrA, cfrE, cfrD, and cfrB, which are considered emerging resistance determinants to oxazolidinones [68], were identified at both T0 and T1. Additionally, the mcr-5.2 and mcr-1.11 genes, associated with emerging colistin resistance [69], and van cluster genes, correlated with glycopeptide resistance [70], were detected. The presence of emerging resistance genes related to antibiotic classes critical in nosocomial infections highlights the need for further studies involving wolves as potential sentinels of environmental contamination with resistance determinants. These findings, along with the identification of reads that may be associated with pathogens prioritized by the WHO [69 (71)], such as carbapenem-resistant Acinetobacter baumannii, carbapenem-resistant Enterobacterales, carbapenem-resistant Pseudomonas aeruginosa, vancomycin-resistant Enterococcus faecium, and methicillin-resistant Staphylococcus aureus, emphasize how wildlife recovery efforts are not only crucial for conservation purposes but also provide valuable information about human, animal, and environmental health. Thus, rescued animals serve as sentinels of circulating pathogens and resistance determinants in the environment, offering essential additional data to guide strategies and action plans [72].
Finally, it is important to consider that these data underline the need for caution when handling rescued animals to mitigate health risks for operators, due to the zoonotic potential of some detected pathogens and the relevance of medically important antibiotic resistance genes. Additionally, the reverse transfer of pathogens or resistance profiles that could compromise species conservation should also be avoided. The presence of reads does not guarantee the occurrence of microorganisms capable of infecting a new host nor does it demonstrate the presence of an infection in the host where it was identified [73]. However, it provides valuable evidence for future studies and highlights the importance of preventive biosecurity measures to protect operators and ensure the conservation of wildlife species in WRCs.
5. Conclusions
This study provides the first metagenomic description of microbiome and resistome changes in an Apennine wolf during long-term rehabilitation. Clear shifts in microbial composition and the detection of a wide range of antimicrobial resistance genes, including determinants of critical relevance for human medicine, underline the sensitivity of wolves to environmental and anthropogenic influences.
Despite being limited to a single case, these findings highlight the value of rehabilitated wildlife as sentinels of pathogen and resistance circulation in human-impacted ecosystems and emphasize the need for strict biosecurity protocols in rehabilitation facilities. Further research involving larger cohorts is required to clarify how rehabilitation practices and human–wildlife interfaces contribute to shaping microbial communities and antimicrobial resistance patterns in large carnivores.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1. Table S1: Taxonomic classification of microbial reads across six levels (phylum, class, order, family, genus, species) for both time points. Table S2: Comprehensive list of antimicrobial resistance genes (ARGs) detected in fecal samples at T0 and T1. Information includes gene name, gene family, resistance mechanism, associated antimicrobial class, and estimated read counts.
Author Contributions
Conceptualization, C.S., S.A. and C.E.D.F.; methodology, C.S. and C.E.D.F.; validation, C.S. and C.E.D.F.; formal analysis, C.S. and C.E.D.F.; investigation, S.A., F.D.T. and A.A.; resources, S.A. and C.E.D.F. ; data curation, C.S., S.A., F.D.T. and C.E.D.F.; writing—original draft preparation, C.S., S.A., F.D.T. and A.A.; writing—review and editing F.M. and C.E.D.F.; supervision, S.A., F.M. and C.E.D.F.; funding acquisition, S.A. and C.E.D.F. All: authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Ministry of Environment, Italy, WOLFNEXT Project (Direttiva 6372/UDCM 31 Marzo 2021).
Institutional Review Board Statement
This study is derived from the application of conservation actions under the WOLFNEXT Project (Direttiva 6372/UDCM 31 Marzo 2021, Ministry of Environment, Italy).
Informed Consent Statement
Not applicable.
Data Availability Statement
The original data presented in the study are openly available in BioProject PRJNA1101237 in National Center for Biotechnology Information (NCBI) and are included in the article or supplementary material. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ABC: ATP-binding cassette AMR: Antimicrobial resistance ARG: Antimicrobial resistance gene BCS: Body condition score CARD: Comprehensive Antibiotic Resistance Database CIA: Critically Important Antimicrobial DNA: Deoxyribonucleic acid FASTQ: Standard text-based format for sequencing reads and quality scores GC: Guanine–cytosine GPS: Global Positioning System HPCIA: Highest Priority Critically Important Antimicrobial MATE: Multidrug and toxic compound extrusion MFS: Major facilitator superfamily NCBI: National Center for Biotechnology Information NGS: Next-generation sequencing RND: Resistance–nodulation–cell division RNA: Ribonucleic acid SMR: Small multidrug resistance WHO: World Health Organization WRC: Wildlife Rehabilitation Centre |
References
- Chapron, G.; Kaczensky, P.; Linnell, J.D.C.; et al. Recovery of Large Carnivores in Europe’s Human-Dominated Landscapes. Science 2014, 346, 1517–1519.
- Cimatti, M.; Ranc, N.; Benítez-López, A.; et al. Large Carnivore Expansion in Europe Is Associated with Human Population Density and Land Cover Changes. Divers. Distrib. 2021, 27, 602–617.
- Dressel, S.; Sandström, C.; Ericsson, G. A Meta-Analysis of European Attitudes Toward Bears and Wolves. Conserv. Biol. 2015, 29, 565–574.
- Milanesi, P.; Puopolo, F.; Zellweger, F. Landscape Features, Human Disturbance or Prey Availability? What Shapes the Distribution of Large Carnivores in Europe? Land 2022, 11, 1807.
- Bartoń, K.A.; Zwijacz-Kozica, T.; Zięba, F.; Sergiel, A.; Selva, N. Bears without Borders: Long-Distance Movement in Human-Dominated Landscapes. Glob. Ecol. Conserv. 2019, 17, e00541.
- Martínez-Abraín, A.; Jiménez, J.; Jiménez, I.; et al. Ecological Consequences of Human Depopulation of Rural Areas on Wildlife: A Unifying Perspective. Biol. Conserv. 2020, 252, 108860.
- Fernández-Gil, A.; da Rocha, C.; Ferreira Pinto, S.M.; Di Silvestre, I. Large Carnivore Management Plans in EU Member States; 2018.
- Blanco, J.C.; Sundseth, K. The Situation of the Wolf (Canis lupus) in the European Union – An In-Depth Analysis; European Commission: Brussels, Belgium, 2023.
- Zanni, M.; Brogi, R.; Merli, E.; Apollonio, M. The Wolf and the City: Insights on Conservation in the Anthropocene. Anim. Conserv. 2023, doi:10.1111/acv.12858.
- Hassell, J.M.; Begon, M.; Ward, M.J.; Fèvre, E.M. Urbanization and Disease Emergence: Dynamics at the Wildlife–Livestock–Human Interface. Trends Ecol. Evol. 2017, 32, 55–67.
- Long, R.B.; Krumlauf, K.; Young, A.M. Characterizing Trends in Human–Wildlife Conflicts in the American Midwest Using Wildlife Rehabilitation Records. PLoS ONE 2020, 15, e0238805.
- Molony, S.; Baker, P.; Garland, L.; Cuthill, I.; Harris, S. Factors That Can Be Used to Predict Release Rates for Wildlife Casualties. Anim. Welf. 2007, 16, 361–367.
- Vezyrakis, A.; Bontzorlos, V.; Rallis, G.; Ganoti, M. Two Decades of Wildlife Rehabilitation in Greece: Major Threats, Admission Trends and Treatment Outcomes from a Prominent Rehabilitation Centre. J. Nat. Conserv. 2023, 73, 126372.
- Malmberg, J.L.; White, L.A.; VandeWoude, S. Bioaccumulation of Pathogen Exposure in Top Predators. Trends Ecol. Evol. 2021, 36, 411–420.
- Ståhlberg, S.; Apollonio, M. Scavenger Activity and Anti-Predator Behaviour in an Apennine Wolf Area. Sustainability 2023, 15, 11056.
- Johnson, M.R.; Boyd, D.K.; Pletscher, D.H. Serologic Investigations of Canine Parvovirus and Canine Distemper in Relation to Wolf (Canis lupus) Pup Mortalities. J. Wildl. Dis. 1994, 30, 270–273.
- Molnar, B.; Duchamp, C.; Mostl, K.; Diehl, P.A.; Betschart, B. Comparative Survey of Canine Parvovirus, Canine Distemper Virus and Canine Enteric Coronavirus Infection in Free-Ranging Wolves of Central Italy and South-Eastern France. Eur. J. Wildl. Res. 2014, 60, 613–624.
- Di Francesco, C.E.; Smoglica, C.; Paoletti, B.; et al. Detection of Selected Pathogens in Apennine Wolf (Canis lupus italicus) by a Non-Invasive GPS-Based Telemetry Sampling of Two Packs from Majella National Park, Italy. Eur. J. Wildl. Res. 2019, 65, 84.
- Simões, R.; Ferreira, C.; Gonçalves, J.; et al. Occurrence of Virulence Genes in Multidrug-Resistant Escherichia coli Isolates from Iberian Wolves (Canis lupus signatus) in Portugal. Eur. J. Wildl. Res. 2012, 58, 677–684.
- Gonçalves, A.; Igrejas, G.; Radhouani, H.; et al. Antimicrobial Resistance in Faecal Enterococci and Escherichia coli Isolates Recovered from Iberian Wolf. Lett. Appl. Microbiol. 2013, 56, 268–274.
- Garcês, A.; Pires, I. European Wild Carnivores and Antibiotic-Resistant Bacteria: A Review. Antibiotics 2023, 12, 1725.
- Smoglica, C.; Angelucci, S.; Di Tana, F.; et al. Antibiotic Resistance in the Apennine Wolf (Canis lupus italicus): Implications for Wildlife and Human Health. Antibiotics 2023, 12, 950.
- Di Francesco, A.; Salvatore, D.; Ranucci, A.; Gobbi, M.; Morandi, B. Antimicrobial Resistance in Wildlife: Detection of Antimicrobial Resistance Genes in Apennine Wolves from Central Italy. Vet. Res. Commun. 2024, 48, 1941–1947.
- Torres, R.T.; Carvalho, J.; Cunha, M.V.; et al. Temporal and Geographical Research Trends of Antimicrobial Resistance in Wildlife—A Bibliometric Analysis. One Health 2020, 11, 100198.
- Marcacci, M.; Ancora, M.; Mangone, I.; et al. Whole Genome Sequence Analysis of the Arctic-Lineage Strain Responsible for Distemper in Italian Wolves and Dogs through a Fast and Robust Next Generation Sequencing Protocol. J. Virol. Methods 2014, 202, 64–68.
- Liu, Y.; Liu, B.; Liu, C.; et al. Differences in the Gut Microbiomes of Dogs and Wolves: Roles of Antibiotics and Starch. BMC Vet. Res. 2021, 17, 112.
- Conceição-Neto, N.; Godinho, R.; Álvares, F.; et al. Viral Gut Metagenomics of Sympatric Wild and Domestic Canids, and Monitoring of Viruses: Insights from an Endangered Wolf Population. Ecol. Evol. 2017, 7, 4135–4146.
- Sarchese, V.; Fruci, P.; Palombieri, A.; et al. Molecular Identification and Characterization of a Genotype 3 Hepatitis E Virus Strain Detected in a Wolf Faecal Sample, Italy. Animals 2021, 11, 3465.
- Chen, L.; Sun, M.; Xu, D.; et al. Gut Microbiome of Captive Wolves Is More Similar to Domestic Dogs than to Wild Wolves Indicated by Metagenomics Study. Front. Microbiol. 2022, 13, 1027188.
- Wu, X.; Zhang, H.; Chen, J.; et al. Analysis and Comparison of the Wolf Microbiome under Different Environmental Factors Using Three Different Data of Next Generation Sequencing. Sci. Rep. 2017, 7, 11332.
- Bergner, L.M.; Orton, R.J.; da Silva Filipe, A.; et al. Using Noninvasive Metagenomics to Characterize Viral Communities from Wildlife. Mol. Ecol. Resour. 2019, 19, 128–143.
- Amato, K.R.; Yeoman, C.J.; Kent, A.; et al. Habitat Degradation Impacts Black Howler Monkey (Alouatta pigra) Gastrointestinal Microbiomes. ISME J. 2013, 7, 1344–1353.
- Tung, J.; Barreiro, L.B.; Burns, M.B.; et al. Social Networks Predict Gut Microbiome Composition in Wild Baboons. eLife 2015, 4, e05224.
- Bobbie, C.B.; Mykytczuk, N.C.S.; Schulte-Hostedde, A.I. Temporal Variation of the Microbiome Is Dependent on Body Region in a Wild Mammal (Tamiasciurus hudsonicus). FEMS Microbiol. Ecol. 2017, 93, fix081.
- Grieneisen, L.E.; Livermore, J.; Alberts, S.; Tung, J.; Archie, E.A. Group Living and Male Dispersal Predict the Core Gut Microbiome in Wild Baboons. Integr. Comp. Biol. 2017, 57, 770–785.
- Miller, E.A.; Ponder, J.B.; Willette, M.; Johnson, T.J.; VanderWaal, K.L. Merging Metagenomics and Spatial Epidemiology to Understand the Distribution of Antimicrobial Resistance Genes from Enterobacteriaceae in Wild Owls. Appl. Environ. Microbiol. 2020, 86, e00571-20.
- Skarżyńska, M.; Leekitcharoenphon, P.; Hendriksen, R.S.; et al. A Metagenomic Glimpse into the Gut of Wild and Domestic Animals: Quantification of Antimicrobial Resistance and More. PLoS ONE 2020, 15, e0242987.
- Jia, T.; Chang, W.S.; Marcelino, V.R.; et al. Characterization of the Gut Microbiome and Resistomes of Wild and Zoo-Captive Macaques. Front. Vet. Sci. 2022, 8, 778556.
- Zou, S.; Hu, R.; Liang, S.; et al. Assessment of Health Risk of Antibiotics Resistance Genes from Human Disturbed Habitat to Wild Animals: Metagenomic Insights into Availability and Functional Changes of Gut Microbiome. Ecotoxicol. Environ. Saf. 2024, 285, 117117.
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast Metagenomic Sequence Classification Using Exact Alignments. Genome Biol. 2014, 15, R46.
- Lu, J.; Salzberg, S.L. Ultrafast and Accurate 16S rRNA Microbial Community Analysis Using Kraken 2. Microbiome 2020, 8, 124.
- FASTQ Toolkit. Illumina BaseSpace Sequence Hub Apps.
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive Metagenomic Visualization in a Web Browser. BMC Bioinform. 2011, 12, 385.
- McArthur, A.G.; Waglechner, N.; Nizam, F.; et al. The Comprehensive Antibiotic Resistance Database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357.
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2020, 48, D517–D525.
- Laflamme, D.P. Development and Validation of a Body Condition Score System for Dogs. J. Am. Anim. Hosp. Assoc. 1997, 33, 126–135.
- Eggermann, J.; da Costa, G.F.; Guerra, A.M.; Kirchner, W.H.; Petrucci-Fonseca, F. Presence of Iberian Wolf (Canis lupus signatus) in Relation to Land Cover, Livestock and Human Influence in Portugal. Mamm. Biol. 2011, 76, 217–221.
- Llaneza, L.; Lopez-Bao, J.V.; Sazatornil, V. Insights into Wolf Presence in Human-Dominated Landscapes: The Relative Role of Food Availability, Humans and Landscape Attributes. Divers. Distrib. 2012, 18, 459–469.
- Ahmadi, M.; Lopez-Bao, J.V.; Kaboli, M. Spatial Heterogeneity in Human Activities Favors the Persistence of Wolves in Agroecosystems. PLoS ONE 2014, 9, e108080.
- Kuijper, D.P.J.; Sahlen, E.; Elmhagen, B.; et al. Paws without Claws? Ecological Effects of Large Carnivores in Anthropogenic Landscapes. Proc. R. Soc. B 2016, 283, 20161625.
- DeCandia, A.L.; Cassidy, K.A.; Stahler, D.R.; et al. Social environment and genetics underlie body site specific microbiomes of Yellowstone National Park gray wolves (Canis lupus). Ecol. Evol. 2021, 11, 9472–9488.
- Nardi, F.; Lazzeri, L.; Iannotti, N.; et al. Analysis of Scat for Gut Microbiome Identification in Wolves from a Mediterranean and an Alpine Area. Diversity 2023, 15, 37. [CrossRef]
- Valerio, A.; Antonucci, A.; Giuliani, A.; et al. Wolf Prey Selection and Food Availability in the Multi-Prey Ecosystem of Majella National Park, Abruzzo. In Proceedings of the XIX Congress of Italian Ecology Society, Bolzano, Italy, 15–18 September 2009.
- Antonucci, A.; Valerio, A.; Petrizzelli, L.; et al. Maiella Wolves Do Not Like Livestock? Twenty Years of Research and Experience on Feeding Ecology of Apennine Wolf (Canis lupus italicus). In Proceedings of Wolves Across Borders Conference, Stockholm, Sweden, 8–11 May 2023.
- Di Sabatino, D.; Lorusso, A.; Di Francesco, C.E.; et al. Arctic Lineage-Canine Distemper Virus as a Cause of Death in Apennine Wolves (Canis lupus) in Italy. PLoS ONE 2014, 9, e82356.
- Zaccaria, G.; Malatesta, D.; Scipioni, G.; et al. Circovirus in Domestic and Wild Carnivores: An Important Opportunistic Agent? Virology 2016, 490, 69–74.
- Alfano, F.; Dowgier, G.; Valentino, M.P.; et al. Identification of Pantropic Canine Coronavirus in a Wolf (Canis lupus italicus) in Italy. J. Wildl. Dis. 2019, 55, 504–508.
- Balboni, A.; Musto, C.; Kaehler, E.; et al. Genetic Characterization of Canine Adenovirus Type 1 Detected by Real-Time PCR in an Oral Sample of an Italian Wolf (Canis lupus). J. Wildl. Dis. 2019, 55, 737–741.
- Amoroso, M.G.; Di Concilio, D.; D’Alessio, N.; et al. Canine Parvovirus and Pseudorabies Virus Coinfection as a Cause of Death in a Wolf (Canis lupus) from Southern Italy. Vet. Med. Sci. 2020, 6, 600.
- Ndiana, L.A.; Lanave, G.; Desario, C.; et al. Circulation of Diverse Protoparvoviruses in Wild Carnivores, Italy. Transbound. Emerg. Dis. 2021, 68, 2489–2502.
- Ndiana, L.A.; Lanave, G.; Vasinioti, V.; et al. Detection and Genetic Characterization of Canine Adenoviruses, Circoviruses, and Novel Cycloviruses from Wild Carnivores in Italy. Front. Vet. Sci. 2022, 9, 331.
- Bertelloni, F.; Cagnoli, G.; Ebani, V.V. Survey on the Occurrence of Zoonotic Bacterial Pathogens in the Feces of Wolves (Canis lupus italicus) Collected in a Protected Area in Central Italy. Microorganisms 2024, 12, 2367.
- Moreno, A.; Musto, C.; Gobbi, M.; et al. Detection and Molecular Analysis of Pseudorabies Virus from Free-Ranging Italian Wolves (Canis lupus italicus) in Italy—A Case Report. BMC Vet. Res. 2024, 20, 9.
- Foti, M.; Siclari, A.; Mascetti, A.; Fisichella, V. Study of the Spread of Antimicrobial-Resistant Enterobacteriaceae from Wild Mammals in the National Park of Aspromonte (Calabria, Italy). Environ. Toxicol. Pharmacol. 2018, 63, 69–73.
- McEwen, S.A.; Collignon, P.J. Antimicrobial Resistance: A One Health Perspective. Microbiol. Spectr. 2018, 6, AR-BA-0009-2017.
- Kim, D.W.; Cha, C.J. Antibiotic Resistome from the One-Health Perspective: Understanding and Controlling Antimicrobial Resistance Transmission. Exp. Mol. Med. 2021, 53, 301–309.
- Pradier, L.; Bedhomme, S. Ecology, More Than Antibiotics Consumption, Is the Major Predictor for the Global Distribution of Aminoglycoside-Modifying Enzymes. eLife 2023, 12, e77015.
- Brenciani, A.; Morroni, G.; Schwarz, S.; Giovanetti, E. Oxazolidinones: Mechanisms of Resistance and Mobile Genetic Elements Involved. J. Antimicrob. Chemother. 2022, 77, 2596–2621.
- Kardos, G.; Laczkó, L.; Kaszab, E.; et al. Phylogenetic Analysis of the Genes in D-Ala-D-Lactate Synthesizing Glycopeptide Resistance Operons: The Different Origins of Functional and Regulatory Genes. Antibiotics 2024, 13, 573.
- Shahzad, S.; Willcox, M.D.P.; Rayamajhee, B. A Review of Resistance to Polymyxins and Evolving Mobile Colistin Resistance Gene (mcr) among Pathogens of Clinical Significance. Antibiotics 2023, 12, 1597.
- World Health Organization (WHO). Bacterial Priority Pathogens List 2024: Bacterial Pathogens of Public Health Importance to Guide Research, Development and Strategies to Prevent and Control Antimicrobial Resistance; WHO: Geneva, Switzerland, 2024.
- Costa, M.M.; Pinto da Cunha, N.; Hagnauer, I.; Venegas, M. A Retrospective Analysis of Admission Trends and Outcomes in a Wildlife Rescue and Rehabilitation Center in Costa Rica. Animals 2023, 14, 51.
- Quince, C.; Walker, A.W.; Simpson, J.T.; et al. Shotgun Metagenomics, from Sampling to Analysis. Nat. Biotechnol. 2017, 35, 833–844.
Figure 1.
Krona chart representing taxonomic composition of the whole microbial community associated with wolf feces at sampling time T0 (A) and T1 (B), revealed by metagenome sequencing.
Figure 1.
Krona chart representing taxonomic composition of the whole microbial community associated with wolf feces at sampling time T0 (A) and T1 (B), revealed by metagenome sequencing.
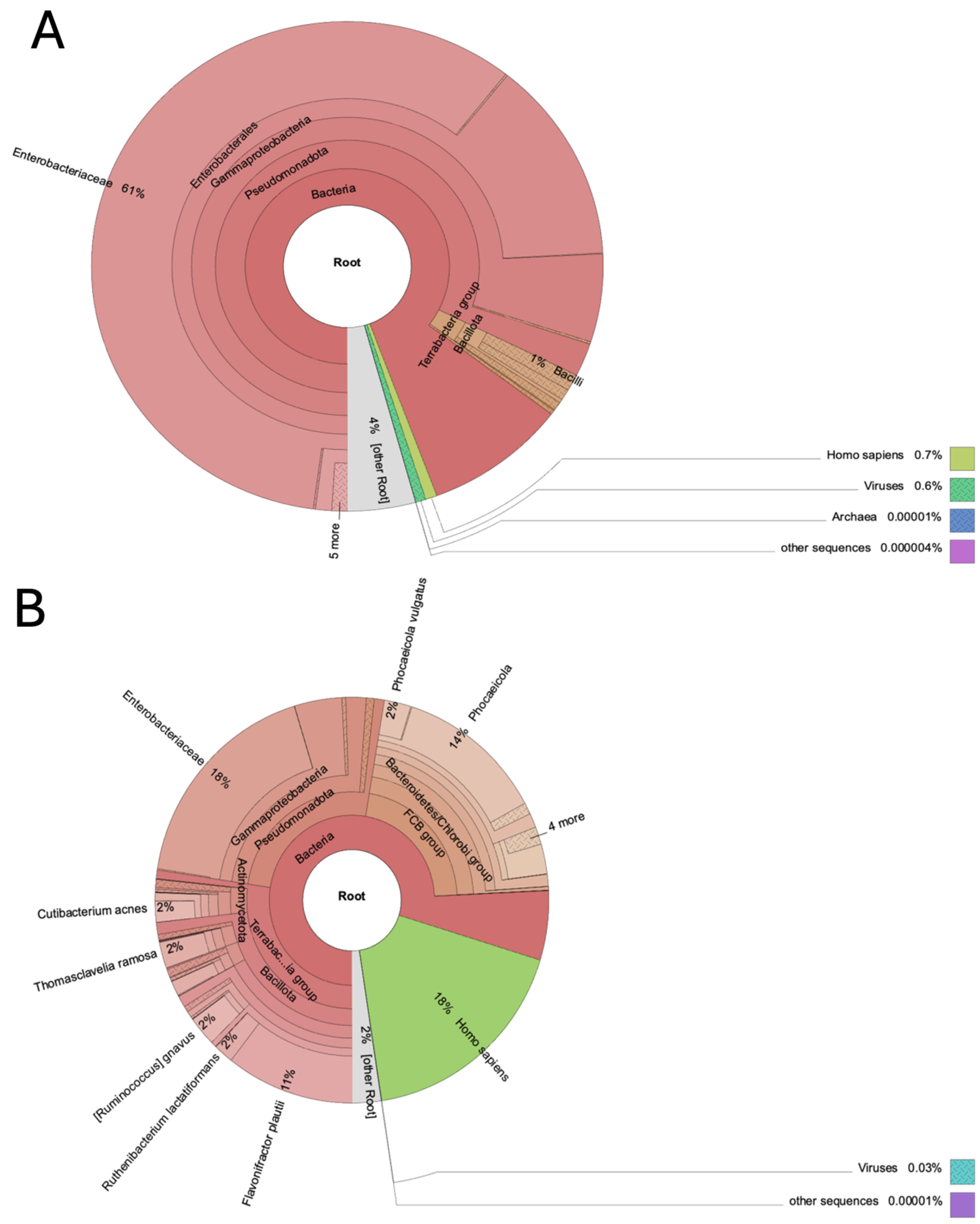
Figure 2.
Two radial charts showing relative abundance of taxonomic genera in wolf feces at time T0 and T1. Genera taxa with relative abundances greater than 0.05% were reported.
Figure 2.
Two radial charts showing relative abundance of taxonomic genera in wolf feces at time T0 and T1. Genera taxa with relative abundances greater than 0.05% were reported.
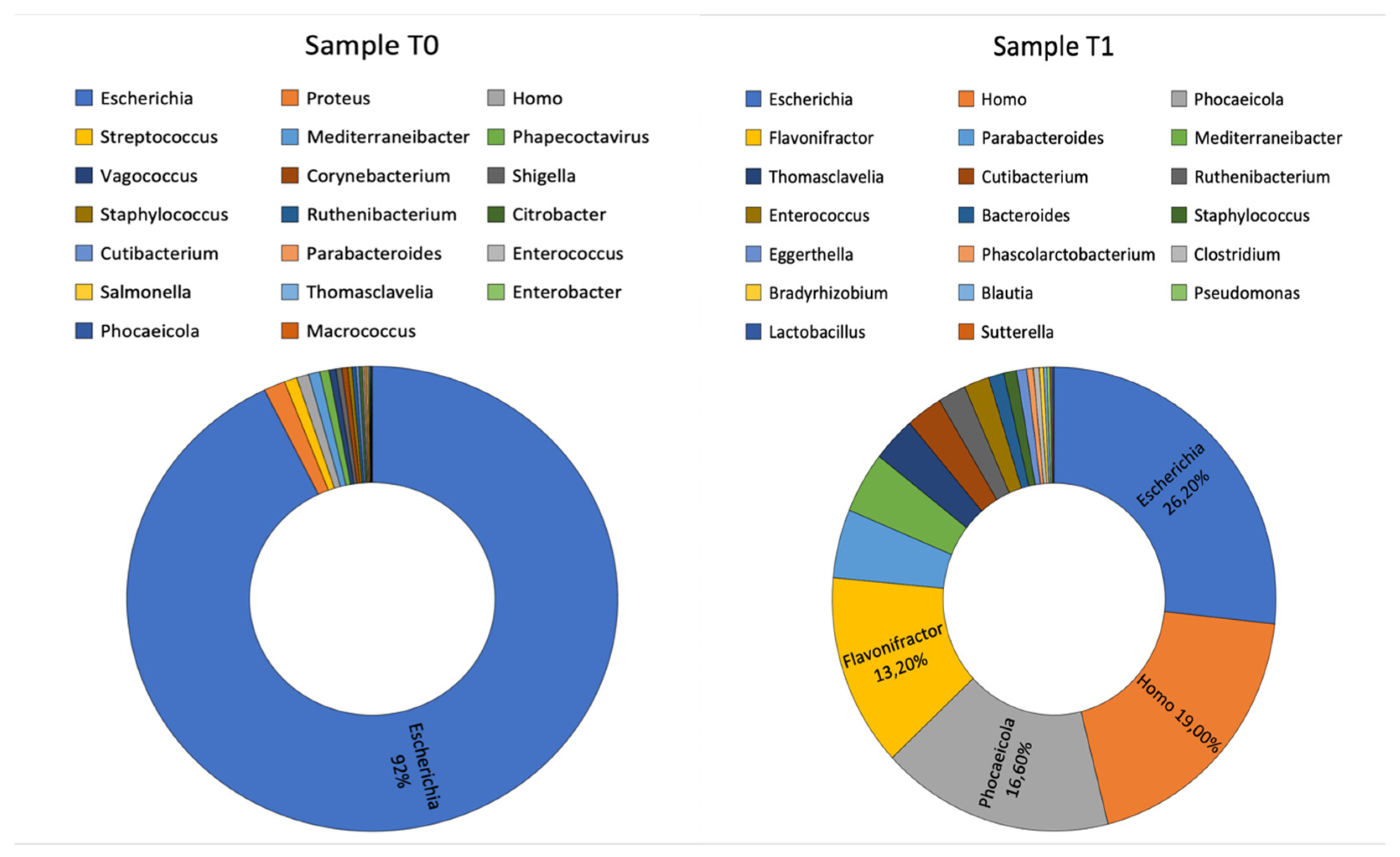
Figure 3.
Bar chart showing the relative abundances of the most recurrent bacterial species in wolf feces at T0 (blue) and T1 (orange).
Figure 3.
Bar chart showing the relative abundances of the most recurrent bacterial species in wolf feces at T0 (blue) and T1 (orange).
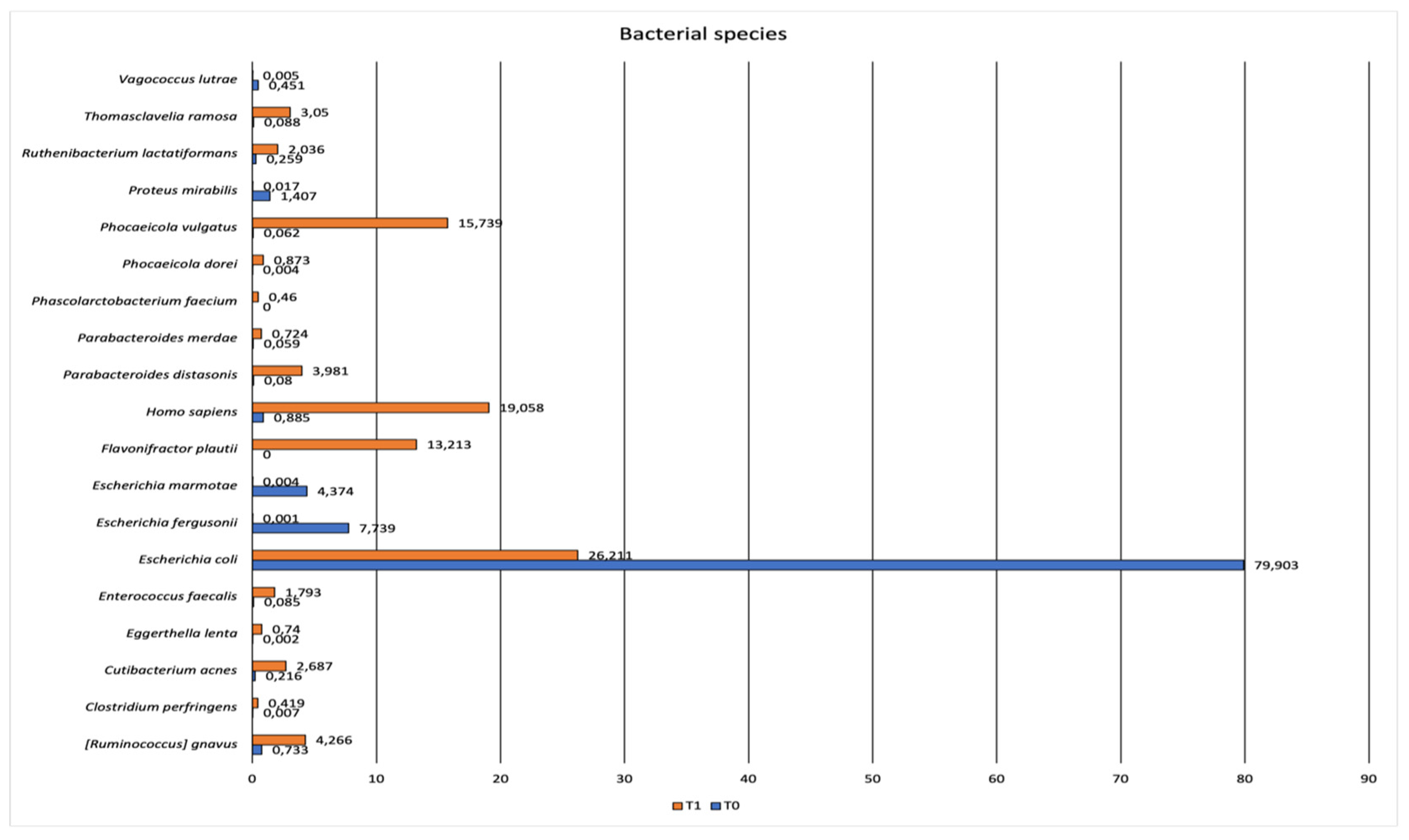
Figure 4.
Krona chart representing taxonomic composition of the whole virus community associated with wolf feces at T0 (A) and T1(B).
Figure 4.
Krona chart representing taxonomic composition of the whole virus community associated with wolf feces at T0 (A) and T1(B).
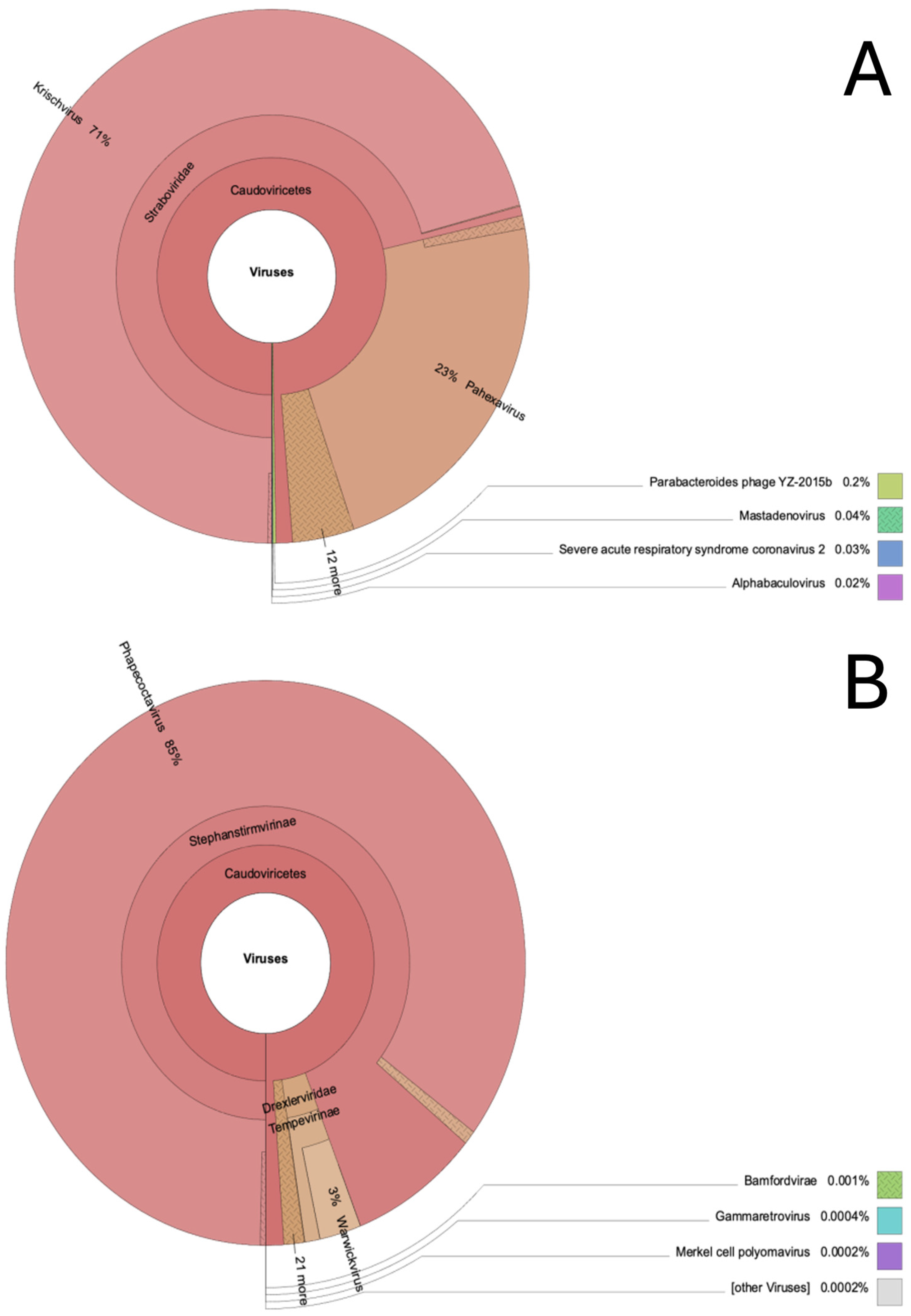
Figure 5.
Bar chart showing the relative abundances of the resistance mechanism in wolf feces at T0 (blue) and T1 (orange).
Figure 5.
Bar chart showing the relative abundances of the resistance mechanism in wolf feces at T0 (blue) and T1 (orange).
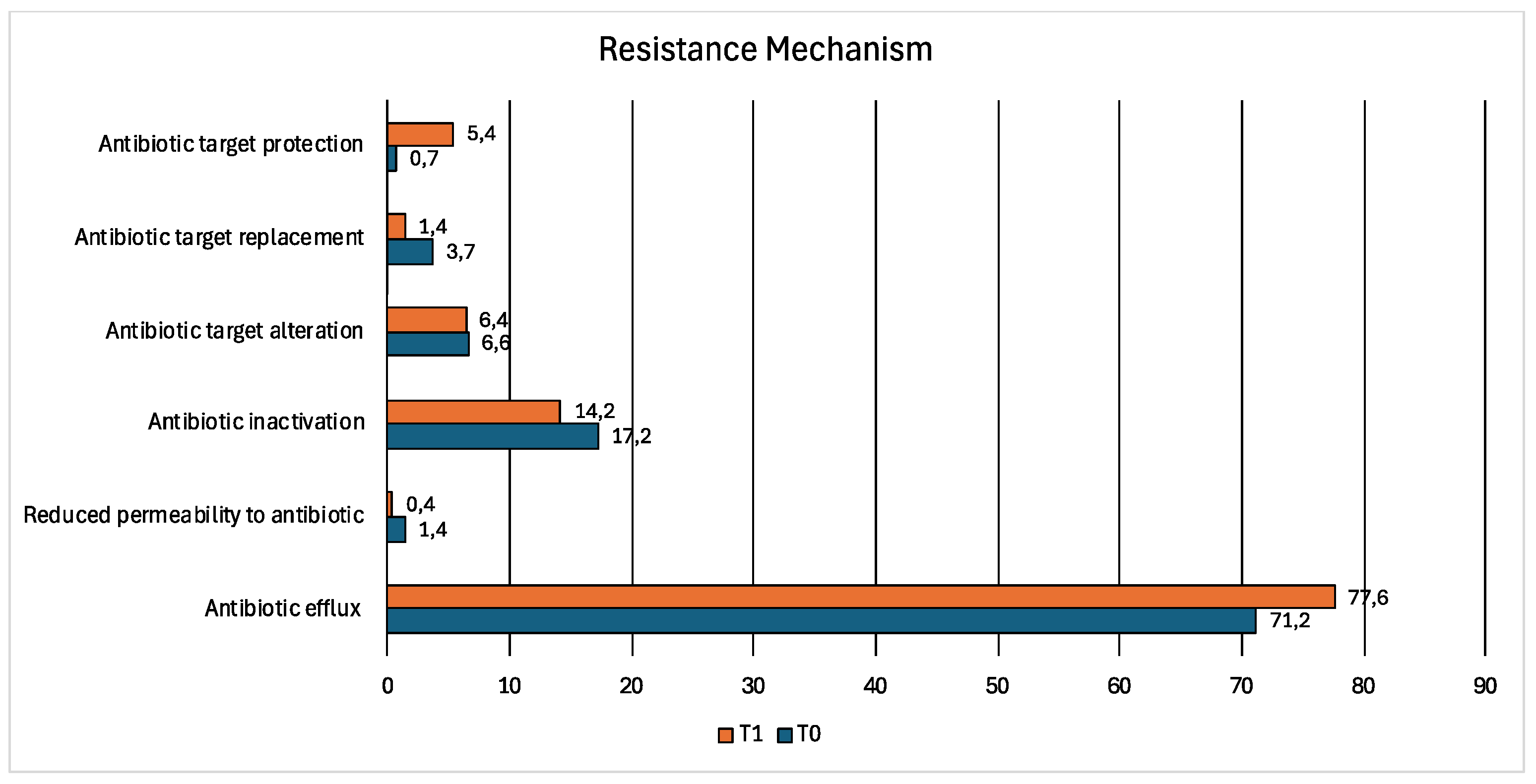
Figure 6.
Bar chart showing the relative abundance of drug classes associated with resistance gene reads in wolf fecal samples at T0 (blue) and T1 (orange).
Figure 6.
Bar chart showing the relative abundance of drug classes associated with resistance gene reads in wolf fecal samples at T0 (blue) and T1 (orange).
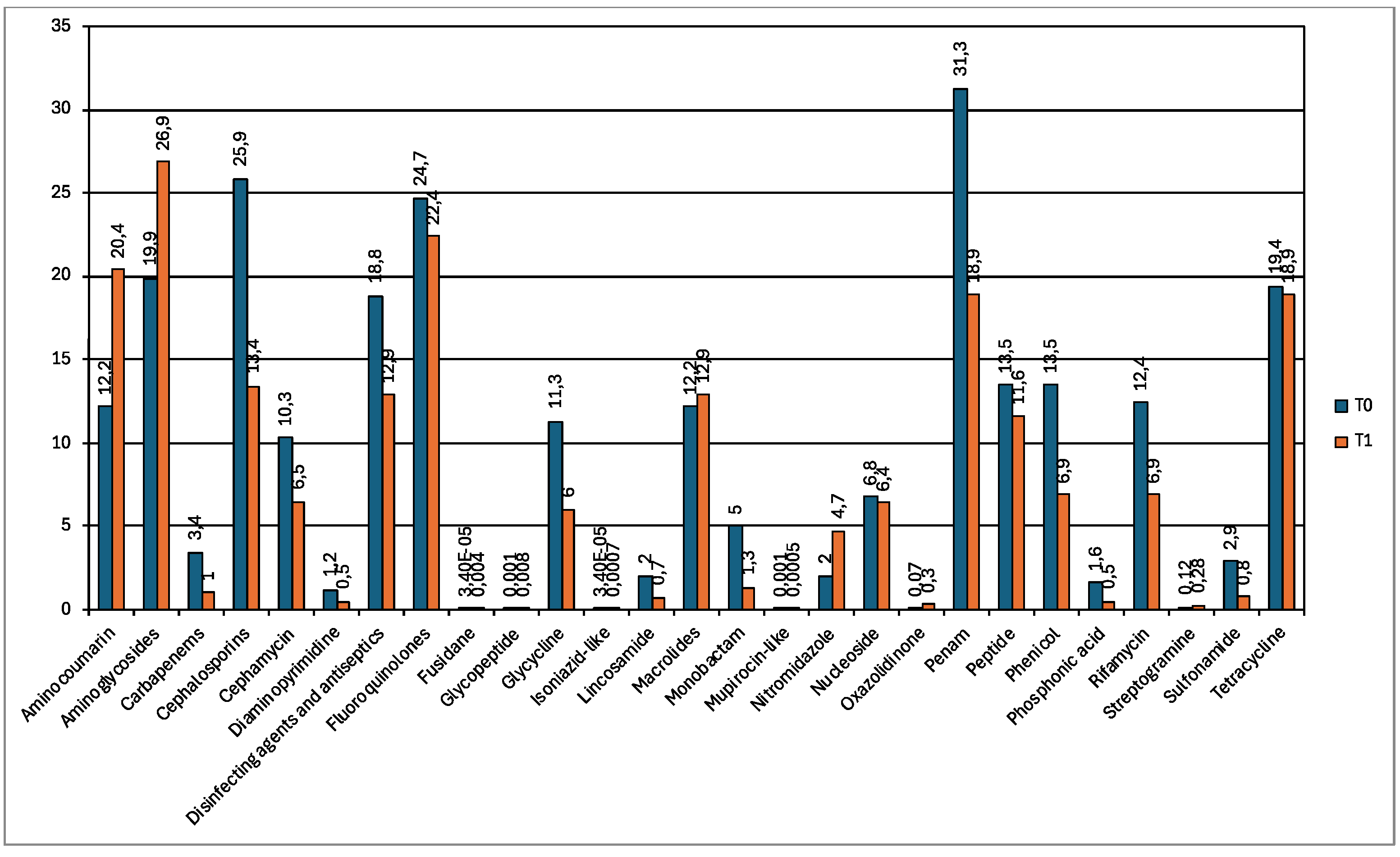
Figure 7.
Bar chart showing the relative abundances of the most recurrent resistance genes in wolf feces at T0 (blue) and T1 (orange).
Figure 7.
Bar chart showing the relative abundances of the most recurrent resistance genes in wolf feces at T0 (blue) and T1 (orange).
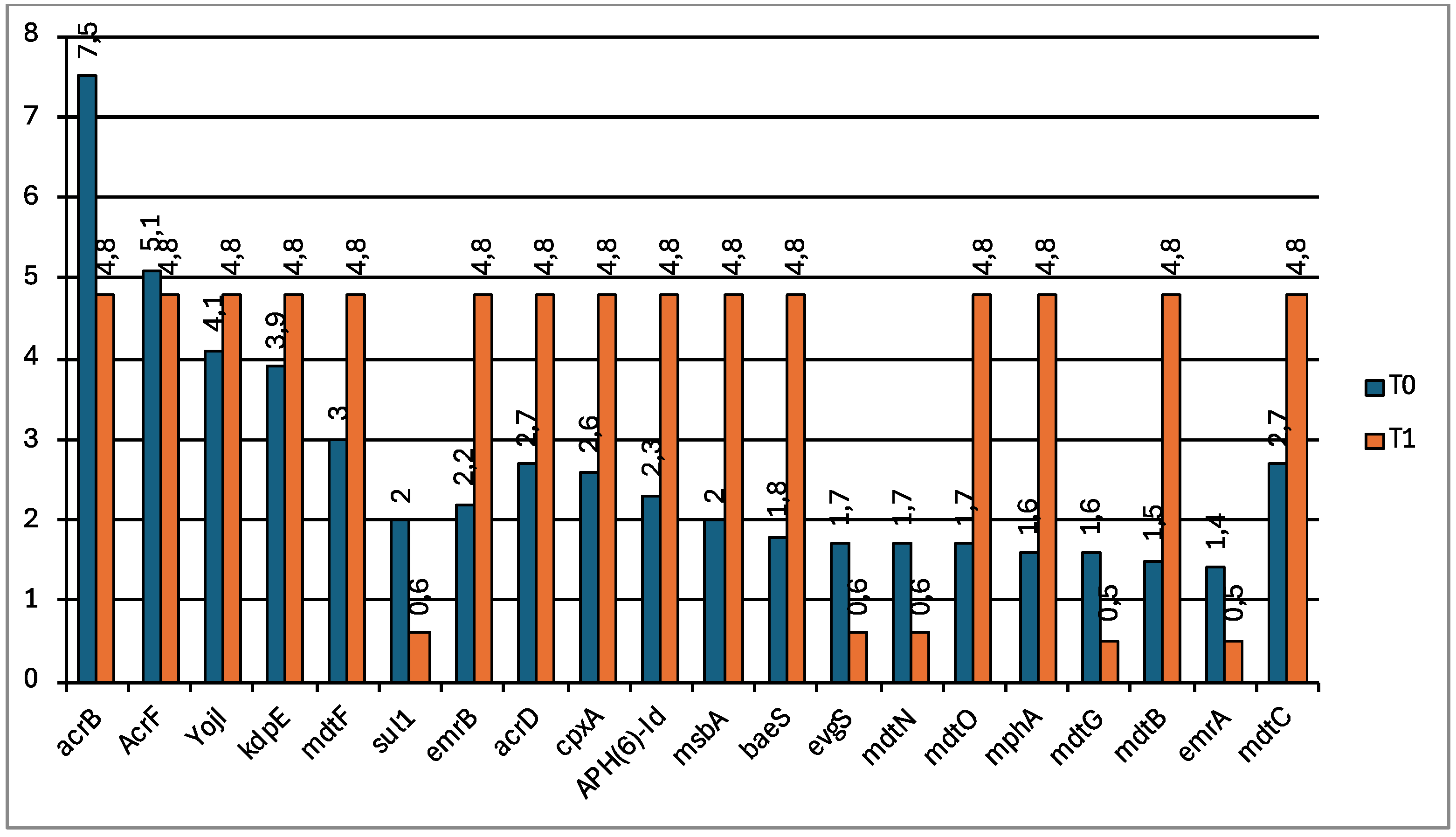

Table 1.
Sequence of potential pathogens identified in wolf feces samples at T0 and T1.
| Potential pathogens | Estimate_reads T0 | Estimate_reads T1 |
| Acinetobacter baumannii | - | 284 |
| Brucella anthropi | - | 193 |
| Brucella pseudogrignonensis | 3440 | - |
| Burkholderia pseudomallei | 23 | - |
| Campylobacter coli | - | 54 |
| Campylobacter concisus | - | 70 |
| Campylobacter jejuni | 732 | - |
| Campylobacter upsaliensis | 1012 | - |
| Capnocytophaga canimorsus | 402 | 672 |
| Clostridium botulinum | 6 | 1450 |
| Clostridium novyi | 72 | - |
| Clostridium perfringens | 5241 | 172512 |
| Clostridioides difficile | 13106 | 7160 |
| Coxiella burnetii | 18 | - |
| Cutibacterium acnes | 171303 | 1107244 |
| Enterococcus faecalis | 67458 | 738853 |
| Escherichia coli | 63237325 | 10802453 |
| Haemophilus parainfluenzae | 763 | 17919 |
| Helicobacter canis | 24 | - |
| Human mastadenovirus C | 2 | 4 |
| Klebsiella pneumoniae | 2127 | 1993 |
| Legionella pneumophila | - | 11 |
| Listeria monocytogenes | - | 26 |
| Mycobacterium intracellulare | - | 39 |
| Neisseria gonorrhoeae | - | 626 |
| Neisseria meningitidis | 86 | 160 |
| Morganella morganii | 15350 | 9563 |
| Pseudomonas aeruginosa | 1344 | 6049 |
| Pseudomonas alcaligenes | - | 3106 |
| Pseudomonas putida | 924 | 2485 |
| Proteus mirabilis | 1113530 | 6972 |
| Severe acute respiratory syndrome-related coronavirus | - | 4 |
| Staphylococcus aureus | 33503 | 650 |
| Staphylococcus caprae | 5 | 12 |
| Staphylococcus epidermidis | 23997 | 151866 |
| Staphylococcus hominis | 18967 | 23442 |
| Staphylococcus hyicus | 14351 | - |
| Staphylococcus pseudintermedius | 124572 | 21 |
| Staphylococcus saprophyticus | 18 | 194 |
| Streptococcus agalactiae | 226154 | 6948 |
| Streptococcus canis | 26216 | 13 |
| Streptococcus dysgalactiae | 189164 | 54 |
| Streptococcus equinus | 9740 | 133 |
| Streptococcus pyogenes | 28121 | 64 |
| Streptococcus suis | 6873 | 1262 |
| Streptococcus uberis | 34711 | 44 |
| Vibrio cholerae | - | 51 |
| Yersinia enterocolitica | 26 | 20 |
Table 2.
Antimicrobial resistance gene families of medically important antimicrobials detected in fecal samples at T0 and T1 and p value.
Table 2.
Antimicrobial resistance gene families of medically important antimicrobials detected in fecal samples at T0 and T1 and p value.
| Antibiotic classes | AMR Gene Family | ||
| Medically important antibiotics | T0 | T1 | p value |
| Cephalosporins | RND antibiotic efflux pump, TEM beta-lactamase, EC beta-lactamase, SHV beta-lactamase, ampC-type beta-lactamase, OXA beta-lactamase, CTX-M beta-lactamase, ACT beta-lactamase, SRT beta-lactamase, class A LRA beta-lactamase, OKP beta-lactamase, SCO beta-lactamase, CepA beta-lactamase, CblA beta-lactamase, LAP beta-lactamase | RND antibiotic efflux pump,TEM beta-lactamase, EC beta-lactamase, SHV beta-lactamase, CMY beta-lactamase, MFS antibiotic efflux pump, ampC-type beta-lactamase, SMR antibiotic efflux pump OXA beta-lactamase CTX-M beta-lactamase CfxA beta-lactamase CblA beta-lactamase ADC beta-lactamases CIA beta-lactamase SRT beta-lactamase | 0.39532 |
| Authorized for use in humans only | |||
| Carbapenems | OXA beta-lactamase, SHV beta-lactamase, ACT beta-lactamase, KPC beta-lactamase, subclass B3 PEDO beta-lactamase, MFS antibiotic efflux pump, RND antibiotic efflux pump | OXA beta-lactamase, SHV beta-lactamase, IND beta-lactamase, CIA beta-lactamase, CfiA beta-lactamase, RND antibiotic efflux pump, MFS antibiotic efflux pump | 0.5063 |
| Monobactam | TEM beta-lactamase, RND antibiotic efflux pump | TEM beta-lactamase, RND antibiotic efflux pump | 0.7482 |
| Glycopeptides | vanS gene in vanA cluster, vanXY gene in vanC cluster, vanY gene in vanA cluster, vanR gene in vanC cluster, vanX gene in vanB cluster, Cfr 23S ribosomal RNA methyltransferase, Bleomycin resistant protein | vanS gene in vanA cluster, vanS gene in vanE cluster, vanY gene in vanA cluster, vanR gene in vanM cluster, Bleomycin resistant protein, RND antibiotic efflux pump | 0.8341 |
| Glycyclines | RND antibiotic efflux pump, ABC antibiotic efflux pump; MFS antibiotic efflux pump; multidrug and toxic compound extrusion (MATE) transporter | RND antibiotic efflux pump, ABC antibiotic efflux pump, MFS antibiotic efflux pump multidrug and toxic compound extrusion (MATE) transporter |
0.8179 |
| Oxazolidinone | MFS antibiotic efflux pump Miscellaneous ABC-F subfamily ATP-binding cassette ribosomal protection proteins Cfr 23S ribosomal RNA methyltransferase |
MFS antibiotic efflux pump Cfr 23S ribosomal RNA methyltransferase RND antibiotic efflux pump |
0.4693 |
| Isoniazid | MFS antibiotic efflux pump | MFS antibiotic efflux pump | |
| HPCIA | |||
| Quinolones | MFS antibiotic efflux pump, RND antibiotic efflux pump, ABC antibiotic efflux pump, MATE transporter, quinolone resistance protein (qnr) |
MFS antibiotic efflux pump, RND antibiotic efflux pump, ABC antibiotic efflux pump, MATE transporter, quinolone resistance protein (qnr) |
0.6372 |
| Polymyxins | MCR phosphoethanolamine transferase | MCR phosphoethanolamine transferase | |
| CIA | |||
| Aminoglycosides | KdpDE, RND antibiotic efflux pump, APH(6), ANT(3''), ANT(4'), ANT(6), ANT(9), AAC(6'), AAC(3), APH(2''), APH(3'), APH(3''), APH(4) | RND antibiotic efflux pump, MFS antibiotic efflux pump, SMR antibiotic efflux pump, 16S rRNA methyltransferase (A1408), 16S rRNA methyltransferase (G1405), APH(6), ANT(3'') ANT(4'), ANT(6), ANT(9) AAC(6'), AAC(3), APH(2'') APH(3'), APH(3''), APH(4) |
0.0004 |
| Ansamycins | RND antibiotic efflux pump, ABC antibiotic efflux pump, rifamycin-resistant beta-subunit of RNA polymerase (rpoB), SMR antibiotic efflux pump, rifampin ADP-ribosyltransferase (Arr), MFS antibiotic efflux pump | RND antibiotic efflux pump, ABC antibiotic efflux pump, rifamycin-resistant beta-subunit of RNA polymerase (rpoB), SMR antibiotic efflux pump, MFS antibiotic efflux pump, RbpA bacterial RNA polymerase-binding protein |
0.6115 |
| Macrolides | MFS antibiotic efflux pump; RND antibiotic efflux pump, MPH, SMR antibiotic efflux pump, msr-type ABC-F protein, Erm 23S ribosomal RNA methyltransferase, ABC antibiotic efflux pump | MFS antibiotic efflux pump, RND antibiotic efflux pump, MPH, SMR antibiotic efflux pump, msr-type ABC-F protein Erm 23S ribosomal RNA methyltransferase, non-erm 23S ribosomal RNA methyltransferase (G748), ABC antibiotic efflux pump |
0.3549 |
ABC: ATP-binding cassette; CIA: Critically Important Antimicrobial; HPCIA: Highest Priority Critically Important Antimicrobial; MATE: multidrug and toxic compound extrusion; MFS: major facilitator superfamily; MPH: macrolide phosphotransferase; RND: resistance-nodulation-cell division; SMR: small multidrug resistance.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



