
Submitted:
14 January 2026
Posted:
15 January 2026
You are already at the latest version
Abstract
The spontaneous emergence of macroscopic dissipative structures in systems driven by generalized chemical potentials is well-established in non-equilibrium thermodynamics. Some examples are, hurricanes, Bénard cells, reaction-diffusion patterns, and atmospheric/oceanic currents. Less recognized, however, are microscopic dissipative structures that form when the driving potential excites internal molecular degrees of freedom (electronic states and nuclear coordinates), typically via high-energy photons. The thermodynamic dissipation theory for the origin of life posits that the core biomolecules of all three domains of life originated as self-organized molecular dissipative structures—chromophores or pigments—that proliferated across the Archean ocean surface to absorb and dissipate the intense “soft” UV-C (205–280 nm) and UV-B (280–315 nm) solar flux into heat. Thermodynamic coupling to ancillary antenna and surface-anchoring molecules subsequently increased photon dissipation and enabled more complex dissipative processes, including modern photosynthesis, to dissipate lower-energy but higher-flux UV-A and visible light. Further thermodynamic coupling to abiotic geophysical cycles (e.g., diurnal, water cycles, winds, and ocean currents) ultimately produced today’s biosphere, efficiently dissipating the full incident solar spectrum well into the infrared. This paper details three examples of molecular dissipative structuring (nucleotides, fatty acids, pigments) and argues that dissipative structuring, rather than natural selection, is the fundamental creative force in biology at all levels of hierarchy.
Keywords:
origin of life
; abiogenesis
; non-equilibrium thermodynamics
; molecular dissipative structuring
; thermodynamic dissipation theory
; prebiotic chemistry
; natural selection
; thermodynamic selection
; biosphere
MSC: 92C05; 92C15; 92C40; 92C45; 80Axx; 82Cxx; 82B35; 82C26
1. Introduction
There are two kinds of structures in nature; equilibrium structures resulting from the maximization of the entropy (e.g., minimization of the Gibb’s potential at constant temperature and pressure), and non-equilibrium structures (or “processes”, since these are often dynamical) resulting from the optimization of the entropy production. Examples of equilibrium structures include crystalline structures and the solar system. Examples of non-equilibrium structures include hurricanes, Bénard cells, reaction-diffusion patterns, the water cycle and atmospheric/oceanic currents. These latter structures were given the name “dissipative structures” by Prigogine [1,2,3] who, building on the work of Onsager [4,5,6], developed the mathematical formalism for treating these structures in the latter half of the 20th century.
Although macroscopic dissipative structures, such as the examples given above, are well known, microscopic dissipative structures are less well known. These are formed through the excitation of internal molecular degrees of freedom (electronic states and nuclear coordinates). Examples include, ATP-driven molecular motors (kinesin, myosin walking on tracks) [7,8] and flagellar motor (run-and-tumble dissipation of proton gradient) [9,10], and protein production in the rybozyme driven by chemical potential [11].
We have identified a previously unrecognized class of microscopic dissipative structures fundamental to the origin of life; organic UV-C chromophores or pigments, dissipatively structured under the UV-C light flux at the Earth’s Archean surface from common precursors such as hydrogen cyanide (HCN), cyanogen (NCCN) and carbon dioxide (CO2) in water [12,13,14]. These structures in water solvent effectively absorb and convert high energy photons into many lower energy photons, thereby performing the non-equilibrium thermodynamic imperative of entropy production. Today, these UV-C chromophores are known as the fundamental molecules of life because they are in the three domains of life (archea, bacteria and eukaryote) and therefore at the very foundations of life. They include the nucleotides, amino acids, fatty acids, sugars, vitamins, coenzymes, cofactors, and pigments (Figure 1).
The objective of this paper is to describe the primordial (Archean) structuring of this newly recognized class of dissipative structures, the fundamental molecules, under the Archean UV-C photon potential. First, a brief introduction to Classical Irreversible Thermodynamic (CIT) theory of Prigogine and collaborators in the non-linear regime is provided. Next, the thermodynamics and dynamics of molecular dissipative structuring of chromophores occurring at the origin of life under the Archean surface UV-C light is described. Three examples (nucleic acids, fatty acids, and pigments), chosen from among the fundamental molecules, are then given. We then discuss how molecular dissipative structuring evolves and is still very much active today, but now under UV-A and visible light using ATP as an intermediate chemical potential. We compare this evolution through dissipative structuring and thermodynamic selection to evolution through natural selection of Darwinian and Gaia theories. Finally, we conclude that thermodynamic selection, including both stochastic and deterministic elements, rather than Darwinian natural selection, is the true fundamental creative force in biology.
2. Non-Linear Classical Irreversible Thermodynamic Theory
Processes, involving material and energy dynamics, are driven by what are known as generalized chemical potentials. Examples include the electric potential which promotes the movement of charged material (a current), temperature potentials which promote a flow of energy (heat flow), concentration potentials which promote the flow of matter (diffusion), chemical and photochemical potentials which promote molecular transformations (chemical and photochemical reactions), etc. The potentials divided by the ambient temperature, are known as generalized thermodynamic forces because the size of these quantities determines the strength of the corresponding generalized thermodynamic flows (e.g. heat flow, diffusion, or reaction rate).
When thermodynamic forces are sufficiently large, the relation between force and flow is no longer necessarily linear. Internal flows can lead to new internal forces, and so on, giving rise to a set of coupled and even catalytically coupled processes and, therefore, non-linear behavior. It is precisely this non-linearity between forces and flows that leads to numerous possible stationary solutions for the system and thus the large diversity of dynamics seen in the interaction of material with its environment, especially for carbon based material when subjected to a strong photochemical potential, a scenario which we call “life”.
The elements of non-linear CIT theory describing the dynamics and evolution of a material system held far out-of-equilibrium through interaction of the system with its environment, are the following;
- 1.
- The existence of at least one relatively constant applied external generalized thermodynamic potential defining the environment - the applied thermodynamic forces.
- 2.
- The spontaneous generation of internal generalized thermodynamic flows resulting from these applied external generalized forces and the possibility of new internal forces that these flows themselves generate.
- 3.
- The existence (in the asymptotic time limit) of various sets of these internal (to the system) forces and flows for non-linear systems for the same initial and boundary conditions, (i.e., multiple, locally stable, dissipative structures, or processes, at stationary states), each set of which can have a different rate of dissipation of the applied external potential (entropy production).
- 4.
- External or internal stochastic perturbations which, near a critical point, could cause the non-linear system to leave the local attraction basin in parameter space of one stationary state and evolve to that of another.
- 5.
- The stochastic (non-deterministic) tendency for evolution through perturbation to stationary states (dissipative structures) of generally greater dissipation (entropy production), particularly towards those states with positive feed-back (non-linear auto- or cross-catalytic), since these have a larger “attraction basin” in this generalized parameter space.
The following sections describe how the origin of life was a particular scenario of the carbon-based molecular dissipative structuring under the UV-C light potential of the early Archean, and how the continuation of this to the dissipative structuring of complexes of the fundamental molecules, led towards pigments of visible wavelengths and their distribution over the entire surface of Earth, thereby increasing global photon dissipation.
3. The Thermodynamics and Dynamics of Molecular Dissipative Structuring
The thermodynamics and dynamics of molecular dissipative structuring is similar to that of macroscopic dissipative structuring in that it involves the restructuring of material to foment the dissipation of an externally imposed generalized chemical potential. The difference is that it involves exciting inter-molecular degrees of freedom (rather than intra-molecular) producing structuring that foments the distribution of the conserved quantity (e.g., photon energy) over a greater number of microscopic degrees of freedom (e.g., a greater number of red-shifted photons). The macroscopic structures that arise are the concentration profiles of the different dissipatively structured molecules.
The “thermodynamic dissipation theory for the origin and evolution of life” (TDTOL) [12,13,14,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34] asserts that life arose as spontaneous molecular dissipative structuring of organic chromophores (pigments) to dissipate the incident UV-C solar light flux available at Earth’s surface throughout the Archean.
Such molecular dissipative structuring led to a “UV-C Pigment World” derived from common carbon-based precursors such as HCN, NCCN, and CO2 at the ocean surface under soft UV-C (205 to 285 nm) and soft UV-B (310-320 nm) photon wavelengths [34]. The soft UV-C photons arrived at the Earth’s surface throughout the Archean with an important integrated energy flux of W m−1 [16,24]. Photons in this region of the spectrum have sufficient energy to re-configure covalent bonds of carbon based molecules, but not enough energy to severely ionize and thereby dissociate or degrade these. The fossil record suggests that UV-C pigments (now known as the fundamental molecules of life – Figure 1) are indeed the first pigments to arise.
The hallmarks of direct photon dissipative structuring [25] are;
- 1.
- sufficient energy per photon to overcome activation barriers as well as sufficiently large photoreaction quantum efficiencies,
- 2.
- a general increase in photon extinction coefficients as the molecules evolve from simple precursors towards final pigments [25],
- 3.
- the formation of conical intersections connecting excited electronic states with the electronic ground state, allowing ultrafast (subpicosecond) radiationless dissipation (internal conversion),
- 4.
- a general trend towards increasing absorption of the greater intensity longer wavelengths of the prevailing surface solar spectrum,
- 5.
- molecular ionization energies greater than photon energies in the prevailing surface spectrum, inhibiting photon-induced degradation.
Product molecules can (but not necessarily) have a lower Gibb’s free energy than that of the precursor molecules from which they evolved. However, in thermal systems (chemical reactions) evolution to such a lower free energy state is not spontaneous if there are large energy barriers between configurations. With UV-C photons, coupling of the reactions to the impressed UV-C photon potential (photochemical reactions), allows the transformation to proceed over the barriers, and even to higher Gibb’s free energy configurations, at a rate dependent on, i) photon intensities at the different wavelengths, , ii) the absorption cross section of the molecule as a function of wavelength , and iii) the widths of the phase-space paths leading to the particular conical intersection on the electronic excited state potential energy surface (i.e., the quantum efficiencies ) for particular molecular transformations or internal conversion. Reverse transformations , or transformations to other possible products (e.g., ), under the UV light are less probable if the quantum efficiencies are smaller (smaller phase-space path on the excited potential energy surface) as compared to the quantum efficiency for internal conversion to the ground state .
Figure 2 explains this dissipative structuring (evolution) of the initial poorly absorbing precursor molecular concentration profile towards concentration profiles of greater photon dissipative efficacy under the impressed soft UV-C photon spectrum of the Archean.
4. Examples of Molecular Dissipative Structuring
Hydrogen cyanide (HCN), cyanogen (NCCN) and carbon dioxide (CO2) are likely precursors of life’s fundamental molecules [35,36]. The CO2 atmospheric concentration was likely 10 to 2500 times the modern concentration, with the lower limit being necessary to counteract the fainter young Sun for habitable surface temperatures [37]. HCN and NCCN are particularly important for the nucleobases [14,33,38]. The formation of HCN and NCCN in the N2-rich atmosphere of the Archean required first breaking the triple covalent bond between nitrogens, N≡N. The photodissociation energy ∼9.8 eV corresponds to a wavelength of 126.5 nm (close to the solar Lyman- line of 121.6 nm). Atomic nitrogen then attacks a carbon atom from CH or CH2 to form HCN [39]. Given the large Archean atmospheric abundance of N2 and CO2 and a carbon to oxygen ratio of C/O≥1, this is accomplished readily via photochemistry [40]. It has been estimated that HCN concentrations as high as M may have been common in the enriched microlayer of the Archean ocean surface [14].
In previous articles [14,25,28,33] we have provided detailed simulations of the photochemical molecular dissipative structuring of some of the fundamental molecules of life from the common precursors in water. The reader is referred to those articles for a complete analysis. Here we provide only an outline of the steps involved in relation to non-linear CIT theory while emphasizing the similarities of their thermodynamic origins for three categories of fundamental molecules, in spite of the diversity of their contemporary functions in life.
4.1. Nucleotides
The photochemical production of the nucleotide adenine from HCN in water has been studied experimentally by Ferrris and Orgel [38] and through time-dependent density functional theory by Boulanger [41]. In reference [14] we suggested that this was a molecular dissipative structuring process and identified the relevant photochemical reactions and demonstrated how photon dissipation increases with each step on route to adenine. We simulated the relevant photochemical and chemical reactions involved in the Archean dissipative structuring of adenine (Figure 3) occurring within a fatty acid vesicle floating on the ocean surface [14]. The process involves 5 molecules of hydrogen cyanide (HCN) in water which are converted in 7 steps into adenine under the soft UV-C spectrum (205-285 nm) of Figure 1.
After each step on route to adenine from HCN, the global photon dissipation of the concentration profile of the different molecules involved increases (Figure 4) – a hallmark of dissipative structuring (Section 3). Adenine, the final product, has the largest photon absorption cross section peaking at 260 nm, exactly where the incident Archean spectrum (Figure 1) peaks, and a conical intersection for sub-picosecond dissipation of the electronic excitation energy into heat of the molecule and local water environment.
4.2. Fatty Acid Vesicles
In the previous subsection, the dissipative structuring of adenine was assumed to occur within a fatty acid vesicle floating at the Archean ocean surface. The existence of such vesicles, which spontaneously form from hydrocarbon chains through Gibb’s free energy minimization, is a common assumption in origin of life scenarios. A mechanism postulated for the formation of fatty acids during the early Archean is that of heat activated Fischer-Tropsch polymerization of smaller hydrocarbon chains such as ethylene at the very high temperatures of deep ocean hydrothermal vents.
A more likely scenario is that of the dissipative structuring of hydrocarbon chains under UV-C photons from CO2 or CO in water at moderate temperatures on the ocean surface [28]. Indications that ultraviolet light may have played an important role in the formation of hydrocarbons have come from different experiments. For example, it was shown in the early 1960’s that irradiation with UV-C light of CO2 saturated water containing ferrous salts results in the production of formic acid and formaldehyde [42].
Fatty acid hydrocarbon tails can be extended through the sequential photon-induced polymerization of an initiator molecule such as ethylene. Photo-polymerization occurs through direct photon-induced cleavage of the initiator molecule producing a free-radical which subsequently attacks the carbon-carbon double bonds of an existing polymer.
Saturated fatty acids do not absorb in the UV except for disassociation at nm and the carboxyl head group which absorbs with a small peak at 207 nm [43]. Under the Archean UV-C flux, photon-induced deprotonation could lead to a double carbon bond forming at any point on the hydrocarbon tail. A single double carbon bond in the tail will absorb at 210 nm. Migration of the double bonds along the tail is known to occur [44] leading to conjugated bonds. Two double bonds in a conjugated configuration (diene) gives strong absorption at 215–230 nm. Three conjugated double bonds (triene) will lead to absorption at 258–265 nm, while those with 4 (tetraene) will lead to absorption at 290–304 nm [45]. The diene and triene absorptions lie within the important UV-C spectrum arriving at Earth’s surface during the Archean (Figure 1).
The steps involved in the dissipative structuring of fatty acids are presented in Figure 5 and are summarized as follows,
- 1.
- UV-C-induced reduction of CO2 and CO in water saturated with these to form ethylene,
- 2.
- UV-C-induced polymerization of ethylene to form long hydrocarbon tails with an even number of carbon atoms,
- 3.
- oxidation and hydrolysis events to stop the growing of the chain and form the carboxyl group,
- 4.
- UV-C-induced deprotonation of the tails to form a double bond,
- 5.
- double bond migration to give a conjugated diene or triene with a conical intersection and strong absorption within the Archean UV-C spectrum.
Polymerization of ethylene occurs over a large UV wavelength region but rates are more than two orders of magnitude larger at UV-C wavelengths (254 nm) than at UV-A (365 nm) [46]. Oxygen acts as a strong inhibitor to polymerization by rapidly reacting with the radical to form a peroxy-based radical which does not promote polymerization [46]. Such an oxidation reaction following hydrolysis is the origin of the carboxyl head group of the fatty acids. The presence of oxygen and the lack of surface UV-C light today means that hydrocarbon chain polymerization today at the ocean surface, although existing [47,48], is only a mere remnant of what it probably was at the origin of life.
Hydrocarbons having conjugated dienes, trienes, or tetraenes almost always have conical intersections [49] allowing rapid dissipation of the electronic excited state energy. Reaching the conical intersection when in the electronic excited state involves a twisting about two C=C bonds and decreasing one of the C-C-C angles producing a kink in the carbon backbone [49]. Therefore, as for the dissipative structuring of adenine, the same photons that dissipatively structured the fatty acid are the ones that will be dissipated efficiently by the final photochemical product (Figure 2).
These dissipatively structured fatty acids are, of course, robust to further photochemical reactions because of the sub-picosecond decay times of their electronic excited states (due to their conical intersections) which is too fast to allow appreciable further chemical transformation. These conjugated fatty acid structures are thus the final molecular dissipative structures.
Within a wide range of pH values, fatty acids will form spherical vesicles through Gibb’s free energy minimization. In order to maintain vesicle integrity at the hot surface temperatures of C prevalent (occurring near the equator) during the early Archean, these fatty acids would necessarily have been long ( C atoms) and cross linked through UV-C light which helps improve stability at high temperatures and over a wider range of pH values [28,50]. There is, in fact, a predominance of 16 and 18 carbon atom fatty acids in the whole available Precambrian fossil record [51,52].
4.3. Pigments
From within the thermodynamic dissipation theory of the origin of life, all fundamental molecules (those common to the three domains) are considered to have been produced as molecular dissipative structures in the form of UV-C chromophores (pigments) [13]. These chromophores evolved further to increase photon dissipative efficacy and to cover the entire Earth’s surface solar spectrum. For example, the nucleobases became information carrying molecules, with the information first related to improving UV-C dissipative efficacy [53] and the fatty acids became the vesicle structures [28] which, besides dissipating soft UV-C light themselves, provided shielding from hard UV-C light [54] as well as performing many other useful functions.
The thermodynamic imperative of increasing photon dissipation permitted, over time, more complex biosynthetic pathways to emerge for the dissipative structuring of pigments which absorb and dissipate photons of longer, visible, wavelengths of much higher photon intensity. Although these photons did not have sufficient energy individually for direct rearrangement of carbon covalent bonds, through biosynthetic pathways, they provided sufficient energy for the phosphorylation of adenine to produce ATP, which could then, in numbers and in combination with cofactors and coenzymes, provide chemical potential for molecular transformation of covalent bonds.
In this section, we consider a dissipatively structured UV-C chromophore evolving through dissipative structuring into one of the most important pigments in the visible for life, chlorophyll; the foundational molecule of visible photosynthesis [55]. The details of the dissipative structuring of chlorophyll under UV-C light have been presented elsewhere [55], here only an overview is presented.
The route to producing chlorophyll a and b from the common amino acid glutamic acid in most organisms today [56] is the route proposed for their production in the early Archean, although through direct dissipative structuring under UV-C photons instead of utilizing the chemical potential of ATP and the complex enzymes of today. This route is; Glutamic Acid → Glutamate-1-Semialdehyde (GSA) → 5-Aminolevulinic Acid (ALA) → Porphobilinogen (PBG) → Hydroxymethylbilane (HMB) → Uroporphyrinogen III → Coproporphyrinogen III → Protoporphyrinogen IX → Protoporphyrin IX → Mg-protoporphyrin → Mg-protoporphyrin monomethyl ester → Divinyl protochlorophyllide a → Monovinyl protochlorophyllide a → Chlorophyllide a → Chlorophyll a → Chlorophyll b.
Figure 6 plots the molar extinction coefficients of all resolved peak absorption wavelengths for all molecules listed above on route to chlorophyll a and b. As predicted for molecular dissipative structuring, the extinction coefficients increase in size and the peaks increase in number and towards higher photon intensities as the molecule evolves from the precursor L-glutamate to the final chlorophyll molecule. Protoporphyrin IX, and all later molecules, have a conical intersection for extremely rapid internal conversion (photon dissipation into heat) to the ground state [57].
Visible absorption also begins at the intermediate protoporphyrin IX. It may be that at this point in the evolution a primordial visible light photosynthesis became active, still based on photon dissipation but now through a more complex biosynthetic pathway using visible light and with complex enzymes likely vulnerable to UV-C light. It was, therefore, probably around this time ( Ga) that oxygenic photosynthesis began to fill the oceans and atmosphere with oxygen, leading to a protective ozone layer.
5. Dissipative Structuring with Thermodynamic Selection: The Fundamental Creative Force in Biology
Darwinian natural selection cannot be considered as the fundamental creative force in biology since it is not based on physical or chemical law. Rather, natural selection is only a symbolic metaphor, useful only at the level of organism. Attempts at application to non-organismal levels in the biological hierarchy leads to paradoxes and ambiguities [18,32].
Perhaps one of the most conspicuous indications of the ineptness of Darwinian theory is that it sheds no light on the origin of life. Although attempts have been made to stretch the fabric of traditional evolutionary theory to cover selection of individual molecules based on chemical stability or on the ability of molecules to sequester precursors (e.g., through chemical affinity). Neither of these two lines of research have proven fruitful.
Non-equilibrium thermodynamic theory in the non-linear regime, on the other hand, as developed by Onsager, Machlup, Prigogine, Nicolis, Glansdorff, and many others ([2,6,58]) provides a physical and chemical description of the complex dynamics of material in general interacting with its environment. Under this framework, processes (i.e., dissipative structures [3,59]) arise “spontaneously” under an external thermodynamic potential to dissipate this potential. Multiple, locally stable, stationary states exist for non-linear systems, and under perturbation the system may evolve from one state to another, governed by both fluctuations (statistical) and thermodynamic law (deterministic).
The great advantage of this thermodynamic framework over traditional evolutionary theory is that it is based on established fundamental law; the conservation laws, the second law of thermodynamics, and the continuity equations. Furthermore, the framework applies simultaneously to all biotic system levels, from fundamental organic molecules at the origin of life, up to the present biosphere. Stubborn problems and paradoxes inherent in Darwinian theory find resolution under this non-equilibrium thermodynamic framework and a physical thermodynamic explanation for the origin, persistence, and evolution of life can be provided [18].
This section describes the natural evolution carbon-based systems under thermodynamic selection of characteristics of structures (or processes), based stochastically on increases in the global rate of dissipation of the prevailing solar spectrum. Different mechanisms of thermodynamic selection are operative at the different biotic-abiotic levels; from the molecular level at the origin of life discussed above, up to the level of the present biosphere with non-linear coupling of both biotic and abiotic processes.
At the molecular level during the Archean (e.g., “The Pigment World” [34]) natural thermodynamic selection is that of molecular dissipative structuring as described above and presented in Figure 2. At this level (in fact, at all levels) photon dissipative efficacy is being selected and there is no correspondence with traditional Darwinian theory, which, in fact, fails here to provide a description of selection. For this reason, Darwinian theory provides no insights into the origin of life.
At the level of the organism, natural thermodynamic selection resembles Darwinian natural selection with the proviso that what is being selected is not the organisms tautologically defined “differential reproductive success” given it’s external environment, but rather the organisms “differential contribution to global entropy production” given it’s external environment.
This can be elucidated with the following specific example: A wolf endowed with characteristics giving it success at the kill is therefore also successful at dissipating the chemical potential in the body of their prey, but more importantly, in spreading, the non-utilized portion as excrement, serving as a catalyst (fertilizer) for plant growth and thus greater global ecosystem photon dissipation. This was very apparent in the greening (and therefore greater photon dissipation) of Yellowstone National Park following the reintroduction of wolves in 1995 after their extinction by over hunting in 1926 [60]. By increasing the conversion rate of prey to plant fertilizer, and by keeping the prey on the move (thus preventing overgrazing and spreading the nutrients of the prey) successful wolves foment the global entropy production of their ecosystem. Wolves contributing less to entropy production will be those not as efficient at the kill. The mechanism by which these wolves are selected against is indeed their weakening, and therefore lower probability of reaching reproductive age.
At this organismal level, the thermodynamic object of selection - differential entropy production, closely parallels the Darwinian object of selection - differential reproductive success. However, from the thermodynamic perspective, the description is not tautological. Selection is based on a measurable global physical variable - global ecosystem entropy production - not on a local, tautological, non-measurable quantity such as “survivability”. Long term experiments with bacteria indeed demonstrate that global dissipation, rather than local “fitness” is optimized in nature [61].
At the level of today’s biosphere, parasitic, symbiotic, and mutualistic interaction among species from all three domains of life occur, as well as the coupling of biotic with abiotic dissipative processes. This increases the efficacy of solar photon dissipation from the ultraviolet and well into the infrared through increasing the efficacy of plant and cyanobacterial growth, including the spread of organic pigments over the whole of Earth’s surface. This also foments abiotic dissipative processes such as the water cycle and ocean and wind currents [18,19,20,23]. Darwinian theory has no explanation for the driving force for the dynamics of ecosystems, suggesting instead that interactions among entities within the ecosystem are somehow the emergent result of an underlying individualistic organismal battle for survival.
Others argue that Darwinian theory, in fact, similarly applies directly to higher order entities, such as competition between species or clades. At these higher levels, however, competition loses significance since the number of entities in competition dwindles until reaching the limit of only one entity, the biosphere, which itself needs explanation for its observed evolution [62].
Reference is also made to Gaia theory where the selection of interactions between entities within the biosphere is supposed to occur such that it results in characteristics making life conditions more “favorable” to the majority of living organisms [63]. However, here again, tautology arises through the introduction of the word “favorable”. More correctly, under the non-equilibrium thermodynamic perspective, coupling of irreversible processes occurs through dissipative structuring such that the global entropy production generally increases. As for thermodynamic selection at both the molecular and organismal levels, selection at the biosphere level has both deterministic and stochastic elements, as will be explained next.
Thermodynamic selection at the biosphere level (existence for a finite time in a particular stationary state out many possible states in a non-linear system) is contingent upon stationary state stability, and this, in turn, as also observed for abiotic thermodynamic stationary states, is contingent upon entropy production (photon dissipation). Those internal or external fluctuations, macroscopic or microscopic, of any of the components of the biosphere which lead the biosphere to stationary states of greater global entropy production, are those fluctuations most likely be amplified (e.g., auto-catalytic or positive feedback processes).
In terms of thermodynamic forces and flows, a fluctuation in the biosphere may cause new thermodynamic forces to arise at any hierarchal level, giving rise to new generalized flows and the elimination of others. In this way, particular molecular concentration profiles, complexes of different molecules, individual organisms, communities, species, clades, ecosystems, and biospheres arise, wax and wane, or go extinct accordingly.
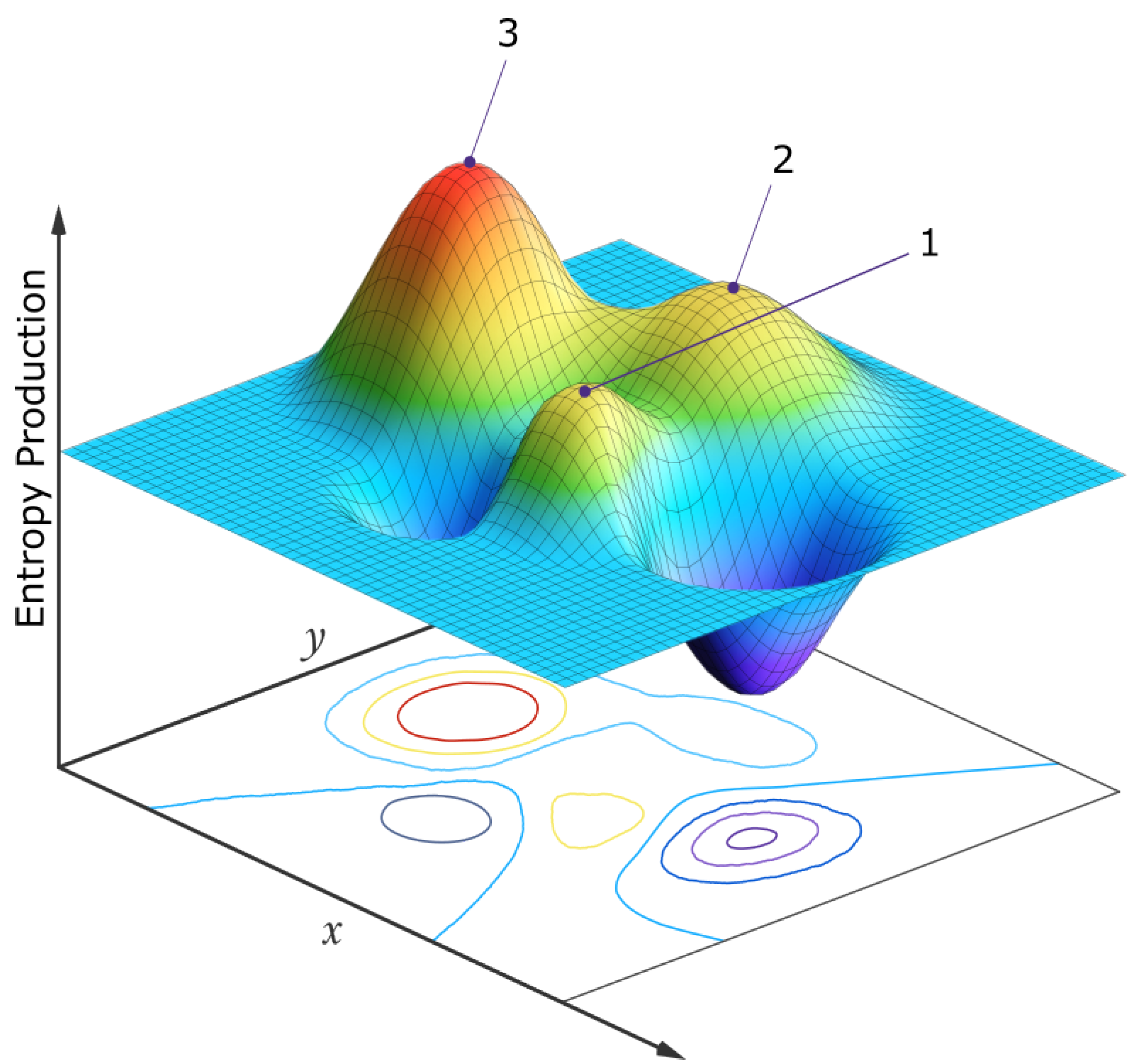
Since selection is contingent upon the photon dissipation of the entire biosphere, at any particular hierarchal level, the biotic units do not compete with each other, or struggle against their external environment, as imagined in the traditional Darwinian perspective, but rather form part of a quasi-stable global biotic-abiotic stationary state which “competes”, on stochastic fluctuation, with other similarly available stationary states of different photon dissipative efficacy in the neighborhood of a generalized phase space (e.g., molecular concentration space at the origin of life, or species populations space at the level of today’s ecosystem). Those stationary states, under the specific environmental conditions (principally the solar photon potential arriving at Earth’s upper atmosphere), which result in greater photon dissipation are generally more probable (stable) since they have a larger attraction basin and greater photon dissipation in this space, and thus more likely to be occupied at any given period in Earth’s history (Figure 7).
For the biosphere, the coupling of biological irreversible processes occurs at each hierarchal level, incorporating, as well, abiotic dissipative processes. For example, the water cycle is coupled through the heat of dissipation of photons in organic pigments in the leaves of plants or within cyanobacteria on the surfaces of the oceans, lakes, and wet soils. The coupling of the water cycle to photon dissipation in organic pigments is autocatalytic since more water in the water cycle implies a greater greening of Earth which implies more water in the water cycle [20,64]. The water cycle dissipates the infrared light resulting from the heat of photon dissipation in the leaves and cyanobacteria even further towards the infrared, finally emitting the energy into space at the cloud tops having an approximate black-body temperature of -14 µC (with an emission peak at m).
Finally, since today’s biosphere has both biotic and abiotic components coupled on many different levels and over different time scales, it is relevant to make a few remarks concerning the plasticity in time of biotic and abiotic irreversible process. Both biotic organisms and abiotic processes have the ability to adapt to a changing impressed thermodynamic potential. Biotic organisms today adapt through their organismal plasticity (e.g., the ability to migrate, the ability to survive off different thermodynamic potentials - heterotrophy, or through mutation of their genes and reproduction - plasticity at the species level). In contrast, abiotic processes have an inherent plasticity, for example, a change in size or direction of a hurricane in response to a change in the size or direction of the ocean surface temperature. The hurricane is, in fact, steered by cyanobacterial ocean surface concentration dissipating sunlight into surface heat [65], yet another biotic-abiotic coupling.
6. Conclusions
Dissipative structuring in biology has been ongoing since its first appearance as molecular dissipative structuring of the fundamental molecules under the UV-C surface light of the Archean, to today’s coupling of biology with abiotic dissipative structures, such as the water cycle and ocean and atmospheric currents. Today, the sum of these structures or processes make up the global dissipative structure known as the biosphere which dissipates sunlight well into the infrared, leading to Earth’s emission of the incident solar energy as far-infrared light with a peak at roughly 15 m.
Neo-Darwinian evolutionary theory suggests that the creative force in biology is natural selection through life’s struggle to survive an imposing external environment while competing for survival among other organisms. Gaia theory speaks of “life (collectively) shaping the environment for its collective benefit”. Both descriptions are tautological and ambiguous. Our non-equilibrium thermodynamic perspective, on the other hand, suggests that irreversible processes such as life are instead dissipative structures or processes arising to dissipate impressed generalized thermodynamic potentials. These processes are thermodynamic flows which rise and fall in response to changes in internal thermodynamic forces. Fluctuations near critical points take the global system to different stationary states (sets of quasi-stable flows and forces), in general toward greater stability, corresponding to increasing solar photon dissipation, as depicted in Figure 2 for the fundamental molecules at life’s origin, and as presented in Figure 7 for today’s biosphere.
The fundamental creative force in biology is therefore thermodynamic, giving rise to two categories of structures; equilibrium and non-equilibrium structures. The spontaneous formation of both these categories of structures is a thermodynamic process involving the variables entropy and entropy production, respectively. The general trend for equilibrium systems is towards maximum entropy while for non-equilibrium systems it is towards greater global entropy production. Structuring in both cases, and the evolution over time of structures or processes for the non-equilibrium case, can be described by physical/chemical principles derived from the second law of thermodynamics and the continuity relations.
Funding
This research received no external funding.
Data Availability Statement
No new data were created.
Acknowledgments
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ALA | 5-Aminolevulinic Acid |
| ATP | Adenosine triphosphate |
| CIT | Classical Irreversible Thermodynamic theory |
| CO2 | Carbon dioxide |
| DNA | Deoxyribonucleic acid |
| GSA | Glutamate-1-Semialdehyde |
| H2S | Hydrogen sulfide |
| HCN | Hydrogen cyanide |
| HMB | Hydroxymethylbilane |
| PBG | Porphobilinogen |
| RNA | Ribonucleic acid |
| SO2 | Sulfur dioxide |
| TDTOL | Thermodynamic Dissipation Theory for the Origin of Life |
| UV-A | Light within the region 315-400 nm |
| UV-B | Light within the region 280-315 nm |
| UV-C | Light within the region 100-280 nm |
| UV-C (hard) | Light in the region 100-205 nm |
| UV-C (soft) | Light within the region 205-285 nm |
References
- Prigogine, I. Introduction to Thermodynamics Of Irreversible Processes, third ed.; John Wiley & Sons, 1955. [Google Scholar]
- Prigogine, I. Introduction to Thermodynamics Of Irreversible Processes, third ed.; John Wiley & Sons: Hoboken, NJ, USA, 1967. [Google Scholar]
- Prigogine, I.; Nicolis, G. Biological order, structure and instabilities. Quarterly Reviews of Biophysics 1971, 4, 107–144. [Google Scholar] [CrossRef]
- Onsager, L. Reciprocal Relations in Irreversible Processes, I. Phys. Rev. 1931, 37, 405–426. [Google Scholar] [CrossRef]
- Onsager, L. Reciprocal Relations in Irreversible Processes, II. Phys. Rev. 1931, 38, 2265. [Google Scholar] [CrossRef]
- Onsager, L.; Machlup, S. Fluctuations and Irreversible Processes. Phys. Rev. 1953, 91, 1505–1512. [Google Scholar] [CrossRef]
- Vale, R.D.; Milligan, R.A. The way things move: Looking under the hood of molecular motor proteins. Science 2000, 288, 88–95. [Google Scholar] [CrossRef]
- Howard, J. Mechanics of Motor Proteins and the Cytoskeleton; Sinauer Associates: Sunderland, MA, 2001. [Google Scholar]
- Berg, H.C.; Brown, D.A. Chemotaxis in Escherichia coli analysed by three-dimensional tracking. Nature 1972, 239, 500–504. [Google Scholar] [CrossRef]
- Berg, H.C. E. coli in Motion; Springer: New York, 2003. [Google Scholar] [CrossRef]
- Zhang, B.; Cech, T.R. Peptide bond formation by in vitro selected ribozymes. Nature 1997, 390, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Michaelian, K. Thermodynamic origin of life. ArXiv 2009, arXiv:physics.gen. [Google Scholar]
- Michaelian, K. Thermodynamic dissipation theory for the origin of life. Earth Syst. Dynam. 2011, 224, 37–51. Available online: https://esd.copernicus.org/articles/2/37/2011/esd-2-37-2011.html. [CrossRef]
- Michaelian, K. The Dissipative Photochemical Origin of Life: UVC Abiogenesis of Adenine. Entropy 2021, 23. Available online: https://www.mdpi.com/1099-4300/23/2/217. [CrossRef] [PubMed]
- Meixnerová, J.; Blum, J.D.; Johnson, M.W.; Stüeken, E.E.; Kipp, M.A.; Anbar, A.D.; Buick, R. Mercury abundance and isotopic composition indicate subaerial volcanism prior to the end-Archean “whiff” of oxygen. [https://www.pnas.org/doi/pdf/10.1073/pnas.2107511118]. Proceedings of the National Academy of Sciences 2021, 118, e2107511118. [Google Scholar] [CrossRef] [PubMed]
- Sagan, C. Ultraviolet Selection Pressure on the Earliest Organisms. J. Theor. Biol. 1973, 39, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Meller, R.; Moortgat, G.K. Temperature dependence of the absorption cross sections of formaldehyde between 223 and 323 K in the wavelength range 225–375 nm. [https://agupubs.onlinelibrary.wiley.com/doi/pdf/10.1029/1999JD901074]. Journal of Geophysical Research: Atmospheres 2000, 105, 7089–7101. [Google Scholar] [CrossRef]
- Michaelian, K. The Non-Equilibrium Thermodynamics of Natural Selection: From Molecules to the Biosphere. Entropy 2023, 25. [Google Scholar] [CrossRef]
- Michaelian, K. The Biosphere. In chapter The biosphere: A thermodynamic imperative; Ishwaran, N., Ed.; INTECH: London, 2012; pp. 51–60. [Google Scholar]
- Michaelian, K. Biological catalysis of the hydrological cycle: life’s thermodynamic function. Hydrol. Earth Syst. Sci. 2012, 16, 2629–2645. Available online: www.hydrol-earth-syst-sci.net/16/2629/2012/. [CrossRef]
- Michaelian, K. A non-linear irreversible thermodynamic perspective on organic pigment proliferation and biological evolution. Conference Series Journal of Physics 475 2013, 012010. [Google Scholar] [CrossRef]
- Michaelian, K.; Santillán Padilla, N. DNA Denaturing through Photon Dissipation: A Possible Route to Archean Non-enzymatic Replication. bioRxiv. 2014. Available online: https://www.biorxiv.org/content/early/2014/11/24/009126.full.pdf. [CrossRef]
- Michaelian, K.; Simeonov, A. Fundamental molecules of life are pigments which arose and co-evolved as a response to the thermodynamic imperative of dissipating the prevailing solar spectrum. Biogeosciences 2015, 12, 4913–4937. Available online: https://bg.copernicus.org/articles/12/4913/2015/. [CrossRef]
- Michaelian, K. Thermodynamic Dissipation Theory of the Origin and Evolution of Life: Salient characteristics of RNA and DNA and other fundamental molecules suggest an origin of life driven by UV-C light; Self-published; Printed by CreateSpace: Mexico City, 2016; ISBN 9781541317482. [Google Scholar]
- Michaelian, K. Microscopic Dissipative Structuring and Proliferation at the Origin of Life. In Heliyon; 2017; 3, p. e00424. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5647473/. [CrossRef]
- Michaelian, K.; Simeonov, A. Thermodynamic explanation of the cosmic ubiquity of organic pigments. Astrobiol. Outreach 2017, 5, 156. Available online: https://www.longdom.org/open-access/thermodynamic-explanation-for-the-cosmic-ubiquity-of-organicpigments-2332-2519-1000156.pdf. [CrossRef]
- Michaelian, K. Homochirality through Photon-Induced Denaturing of RNA/DNA at the Origin of Life. Life 2018, 8. Available online: http://www.mdpi.com/2075-1729/8/2/21. [CrossRef]
- Michaelian, K.; Rodriguez, O. Prebiotic fatty acid vesicles through photochemical dissipative structuring. Revista Cubana de Química 2019, 31, 354–370. [Google Scholar]
- Michaelian, K.; Santillan, N. UVC photon-induced denaturing of DNA: A possible dissipative route to Archean enzyme-less replication. Heliyon 2019, 5, e01902. Available online: https://www.heliyon.com/article/e01902. [CrossRef]
- Michaelian, K. Photochemical Dissipative Structuring of the Fundamental Molecules of Life. Proceedings, 5th International Electronic Conference on Entropy and Its Applications; Session: Biological Systems 2019.
- Michaelian, K.; Cano, R.E. A Photon Force and Flow for Dissipative Structuring: Application to Pigments, Plants and Ecosystems. Entropy 2022, 24, 76. [Google Scholar] [CrossRef]
- Michaelian, K. Non-Equilibrium Thermodynamic Foundations of the Origin of Life. Foundations 2022, 2, 308–337. [Google Scholar] [CrossRef]
- Hernández, C.; Michaelian, K. Dissipative Photochemical Abiogenesis of the Purines. Entropy 2022, 24, 1027. [Google Scholar] [CrossRef]
- Michaelian, K. The Pigment World: Life’s Origins as Photon-Dissipating Pigments. Life 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Minard, R.D.; Matthews, C.N. HCN World: Establishing Proteininucleic Acid Life via Hydrogen Cyanide Polymers. Abstr. Pap. Am. Chem. Soc. 2004, 228, U963–U963. [Google Scholar]
- Matthews, C.N. Origins: Genesis, Evolution and Diversity of Life. In Series: Cellular Origin and Life in Extreme Habitats and Astrobiology; Seckbach, J., Ed.; Kluwer: Dordrecht, 2004; Vol. 6, chapter The HCN World, pp. 121–135. [Google Scholar]
- Catling, D.C.; Zahnle, K.J. The Archean atmosphere. Science Advances 2020, 6, eaax1420. [Google Scholar] [CrossRef] [PubMed]
- Ferris, J.P.; Orgel, L.E. An Unusual Photochemical Rearrangement in the Synthesis of Adenine from Hydrogen Cyanide. J. Am. Chem. Soc. 1966, 88, 1074–1074. [Google Scholar] [CrossRef]
- Trainer, M.G.; Jimenez, J.L.; Yung, Y.L.; Toon, O.B.; Tolbert, M.A. Nitrogen Incorporation in CH4-N2 Photochemical Aerosol Produced by Far UV Irradiation. In NASA archives; 2012. Available online: https://ntrs.nasa.gov/archive/nasa/casi.ntrs.nasa.gov/20120009529.pdf.
- Rimmer, P.B.; Rugheimer, S. Hydrogen cyanide in nitrogen-rich atmospheres of rocky exoplanets. Icarus 2019, 329, 124–137. [Google Scholar] [CrossRef]
- Boulanger, E.; Anoop, A.; Nachtigallova, D.; Thiel, W.; Barbatti, M. Photochemical Steps in the Prebiotic Synthesis of Purine Precursors from HCN. Angew. Chem. Int. 2013, 52, 8000–8003. [Google Scholar] [CrossRef]
- Getoff, N. Reduktion der Kohlensäure in wässeriger Lösung unter Einwirkung von UV-licht. Zeitschrift für Naturforschung B 1962, 17, 87–90. [Google Scholar] [CrossRef]
- Vicente, A.; Antunes, R.; Almeida, D.; Franco, I.J.A.; Hoffmann, S.V.; Mason, N.J.; Eden, S.; Duflot, D.; Canneaux, S.; Delwiche, J.; et al. Photoabsorption measurements and theoretical calculations of the electronic state spectroscopy of propionic, butyric, and valeric acids. Phys. Chem. Chem. Phys. 2009, 11, 5729–5741. [Google Scholar] [CrossRef]
- Mandal, T.K.; Chatterjee, S.N. Ultraviolet- and Sunlight-Induced Lipid Peroxidation in Liposomal Membrane. Radiation Research 1980, 83, 290–302. [Google Scholar] [CrossRef]
- Lewis, G.N.; Calvin, M. The Color of Organic Substances Reports UV absorption maxima for conjugated polyenes: diene 217 nm, triene 258 nm, tetraene 300 nm. Chemical Reviews 1949, 25, 273–328. [Google Scholar] [CrossRef]
- Bowman, C.N.; Kloxin, C.J. Toward an enhanced understanding and implementation of photopolymerization reactions. AIChE J. 2008, 54, 2775–2795. [Google Scholar] [CrossRef]
- Mochida, M.; Kitamori, Y.; Kawamura, K.; Nojiri, Y.; Suzuki, K. Fatty acids in the marine atmosphere: Factors governing their concentrations and evaluation of organic films on sea-salt particles. Journal of Geophysical Research: Atmospheres 2002, 107, AAC 1–1–AAC 1–10. Available online: https://agupubs.onlinelibrary.wiley.com/doi/pdf/10.1029/2001JD001278. [CrossRef]
- Wellen, B.A.; Lach, E.A.; Allen, H.C. Surface pKa of octanoic, nonanoic, and decanoic fatty acids at the air-water interface: applications to atmospheric aerosol chemistry. Phys. Chem. Chem. Phys. 2017, 19, 26551–26558. [Google Scholar] [CrossRef] [PubMed]
- Celani, P.; Garavelli, M.; Ottani, S.; Bemardi, F.; Robb, M.A.; Olivucci, M. Molecular “Trigger” for Radiationless Deactivation of Photoexcited Conjugated Hydrocarbons. J. Am. Chem. Soc. 1995, 117, 11584–11585. [Google Scholar] [CrossRef]
- Fan, Y.; Fang, Y.; Ma, L. The self-crosslinked ufasome of conjugated linoleic acid: Investigation of morphology, bilayer membrane and stability. Colloids and Surfaces B: Biointerfaces 2014, 123, 8–14. [Google Scholar] [CrossRef]
- Han, J.; Calvin, M. Occurrence of fatty acids and aliphatic hydrocarbons in a 3.4 billion-year-old sediment. Nature 1969, 224, 576–577. [Google Scholar] [CrossRef]
- Van Hoeven, W.; Maxwell, J.; Calvin, M. Fatty acids and hydrocarbons as evidence of life processes in ancient sediments and crude oils. Geochimica et Cosmochimica Acta 1969, 33, 877–881. [Google Scholar] [CrossRef]
- Mejía Morales, J.; Michaelian, K. Photon Dissipation as the Origin of Information Encoding in RNA and DNA. Entropy 2020, 22. Available online: https://www.mdpi.com/1099-4300/22/9/940. [CrossRef]
- Lechuga, I.; Michaelian, K. Fatty Acid Vesicles as Hard UV-C Shields for Early Life. Foundations 2023, 3, 99–114. [Google Scholar] [CrossRef]
- Michaelian, K.; Simeonov, A. The Dissipative Photochemical Origin of Photosynthesis. 2026. [Google Scholar]
- Niyogi, K.K.; Wolosiuk, R.A.; Malkin, R. Photosynthesis. In Biochemistry and Molecular Biology of Plants, 2 ed.; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; Wiley Blackwell: Chichester, West Sussex, 2015; pp. 515–519. [Google Scholar]
- Falahati, K.; Hamerla, C.; Huix-Rotllant, M.; Burghardt, I. Ultrafast photochemistry of free-base porphyrin: a theoretical investigation of B → Q internal conversion mediated by dark states This work shows that B → N conversion is barrierless ( 20 fs), followed by N → Q conversion via a conical intersection ( 100 fs), in agreement with experiments on porphine and related free-base porphyrins. Physical Chemistry Chemical Physics 2018, 20, 12483–12492. [Google Scholar] [CrossRef] [PubMed]
- Glansdorff, P.; Prigogine, I. Thermodynamic Theory of Structure, Stability and Fluctuations; Wiley - Interscience: Hoboken, NJ, USA, 1971. [Google Scholar]
- Prigogine, I.; Nicolis, G. On Symmetry-Breaking Instabilities in Dissipative Systems. J. Chem. Phys. 1967, 46, 3542. [Google Scholar] [CrossRef]
- Ripple, W.J.; Beschta, R.L. Trophic cascades in Yellowstone: The first 15years after wolf reintroduction. Biological Conservation 2012, 145, 205–213. [Google Scholar] [CrossRef]
- Barrick, J.E.; Yu, D.S.; Yoon, S.H.; Jeong, H.; Oh, T.K.; Schneider, D.; Lenski, R.E.; Kim, J.F. Genome evolution and adaptation in a long-term experiment with Escherichia coli. Nature 2009, 461, 1243–1247. [Google Scholar] [CrossRef] [PubMed]
- Swenson, R. The Cybernetics of Complex Systems: Self-organization, Evolution, and Social Change. In chapter End-directed physics and evolutionary ordering: Obviating the problem of the population of one; Intersystems Publications: Cambridge, MA, USA, 1991; pp. 41–60. [Google Scholar]
- Lovelock, J.E. The Ages of Gaia; A Biography of Our Living Earth; W. W. Norton&Company: New York, NY, USA, 1988. [Google Scholar]
- Kleidon, A.; Fraedrich, K.; Heimann, M.A. Green Planet Versus a Desert World: Estimating the Maximum Effect of Vegetation on the Land Surface Climate. Climatic Change 2000, 44, 471–493. [Google Scholar] [CrossRef]
- Gnanadesikan, A.; Emanuel, K.; Vecchi, G.A.; Anderson, W.G.; Hallberg, R. How ocean color can steer Pacific tropical cyclones. Geophysical Research Letters 2010, 37, L18802. Available online: http://arxiv.org/abs/https://agupubs.onlinelibrary.wiley.com/doi/pdf/10.1029/2010GL044514 [https://agupubs.onlinelibrary.wiley.com/doi/pdf/10.1029/2010GL044514. [CrossRef]
- Michaelian, K. Thermodynamic stability of ecosystems. Journal of Theoretical Biology 2005, 237, 323–335. Available online: https://www.sciencedirect.com/science/article/pii/S0022519305001839?via%3Dihub. [CrossRef] [PubMed]
Figure 1.
The spectrum of UV light available at Earth’s surface before the origin of life at approximately 3.9 Ga and until at least 2.9 Ga (curves black and red, respectively). The spectrum in the UV-C may even have persisted throughout the entire Archean until 2.5 Ga [15]. Atmospheric CO2, H2O, SO2 and probably some H2S, were responsible for the absorption of wavelengths shorter than ∼205 nm, and atmospheric aldehydes (e.g., formaldehyde and acetaldehyde, common photochemical products of CO2 and water) absorbed between about 285 and 305 nm [16,17]), approximately corresponding to the UV-B region (280 and 315 nm). By around 2.2 Ga (green curve), UV-C light at Earth’s surface was completely extinguished by oxygen and ozone (which we consider as life produced UV-C pigments) resulting from organisms performing oxygenic photosynthesis. The yellow curve corresponds to the present surface spectrum. Energy fluxes are for the Sun at the zenith. Over 50 of the fundamental molecules of life are plotted at their wavelengths of maximum absorption: nucleic acids (black), amino acids (green), fatty acids (violet), sugars (brown), vitamins, co-enzymes, and cofactors (blue), and pigments (red). We have asserted that these fundamental molecules were dissipatively structured as UV-C pigments under this light. The font size is roughly proportional to the relative size of the respective molar extinction coefficient of the pigment. Reproduced with permission from Michaelian [18].
Figure 1.
The spectrum of UV light available at Earth’s surface before the origin of life at approximately 3.9 Ga and until at least 2.9 Ga (curves black and red, respectively). The spectrum in the UV-C may even have persisted throughout the entire Archean until 2.5 Ga [15]. Atmospheric CO2, H2O, SO2 and probably some H2S, were responsible for the absorption of wavelengths shorter than ∼205 nm, and atmospheric aldehydes (e.g., formaldehyde and acetaldehyde, common photochemical products of CO2 and water) absorbed between about 285 and 305 nm [16,17]), approximately corresponding to the UV-B region (280 and 315 nm). By around 2.2 Ga (green curve), UV-C light at Earth’s surface was completely extinguished by oxygen and ozone (which we consider as life produced UV-C pigments) resulting from organisms performing oxygenic photosynthesis. The yellow curve corresponds to the present surface spectrum. Energy fluxes are for the Sun at the zenith. Over 50 of the fundamental molecules of life are plotted at their wavelengths of maximum absorption: nucleic acids (black), amino acids (green), fatty acids (violet), sugars (brown), vitamins, co-enzymes, and cofactors (blue), and pigments (red). We have asserted that these fundamental molecules were dissipatively structured as UV-C pigments under this light. The font size is roughly proportional to the relative size of the respective molar extinction coefficient of the pigment. Reproduced with permission from Michaelian [18].
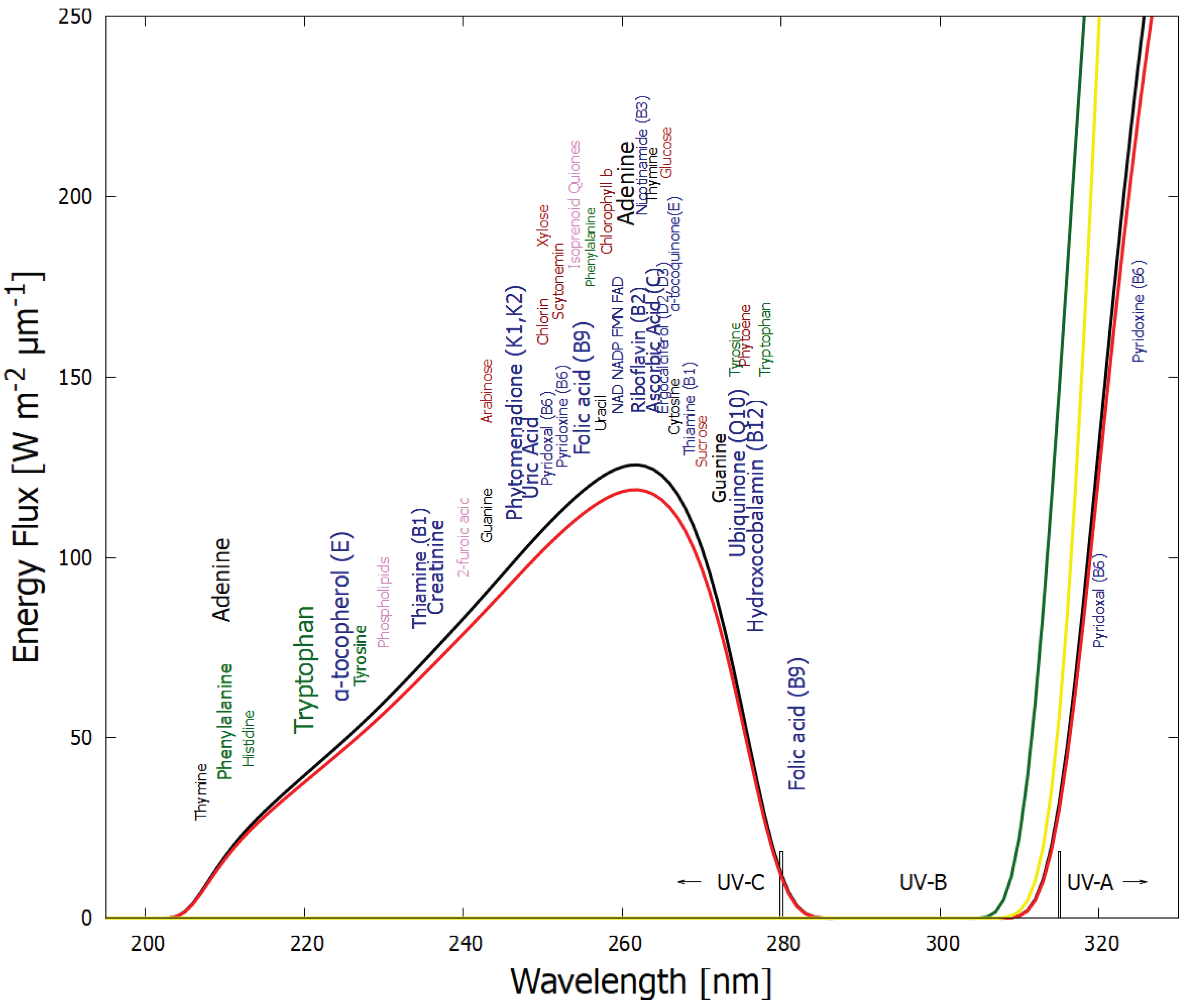
Figure 2.
Mechanism for the evolution of molecular structures towards ever greater photon dissipative efficacy (“molecular dissipative structuring”) on route to the fundamental molecules (in this example adenine). The high activation barriers between configurations mean that reactions will not proceed spontaneously but only through coupling to photon absorption events. Forward and backward rates depend on photon intensities I at the different wavelengths of maximum absorption for the two structures, and on the phase-space widths of paths on their excited potential energy surface qi leading to the conical intersection giving rise to the particular transformation. This implies, in general, different quantum efficiencies for the forward () and backward () reactions. Given different intensities of the incident spectrum at the absorption maxima, and since and (where the “…” represents quantum efficiencies for other possible molecular transformations), those stationary states (concentration profiles) with greater photon dissipative efficacy (higher photon intensity at absorption maxima and higher quantum efficiency for internal conversion ) will therefore gradually become dominant under a continuously impressed UV-C photon flux, independently of the sign or size of the difference in the Gibb’s free energies G of the molecules. This process, of selection of molecular concentration profiles of ever greater photon dissipative efficacy, driving general evolution towards the right in the diagram, we call natural thermodynamic selection. Taken with permission from Michaelian [14].
Figure 2.
Mechanism for the evolution of molecular structures towards ever greater photon dissipative efficacy (“molecular dissipative structuring”) on route to the fundamental molecules (in this example adenine). The high activation barriers between configurations mean that reactions will not proceed spontaneously but only through coupling to photon absorption events. Forward and backward rates depend on photon intensities I at the different wavelengths of maximum absorption for the two structures, and on the phase-space widths of paths on their excited potential energy surface qi leading to the conical intersection giving rise to the particular transformation. This implies, in general, different quantum efficiencies for the forward () and backward () reactions. Given different intensities of the incident spectrum at the absorption maxima, and since and (where the “…” represents quantum efficiencies for other possible molecular transformations), those stationary states (concentration profiles) with greater photon dissipative efficacy (higher photon intensity at absorption maxima and higher quantum efficiency for internal conversion ) will therefore gradually become dominant under a continuously impressed UV-C photon flux, independently of the sign or size of the difference in the Gibb’s free energies G of the molecules. This process, of selection of molecular concentration profiles of ever greater photon dissipative efficacy, driving general evolution towards the right in the diagram, we call natural thermodynamic selection. Taken with permission from Michaelian [14].
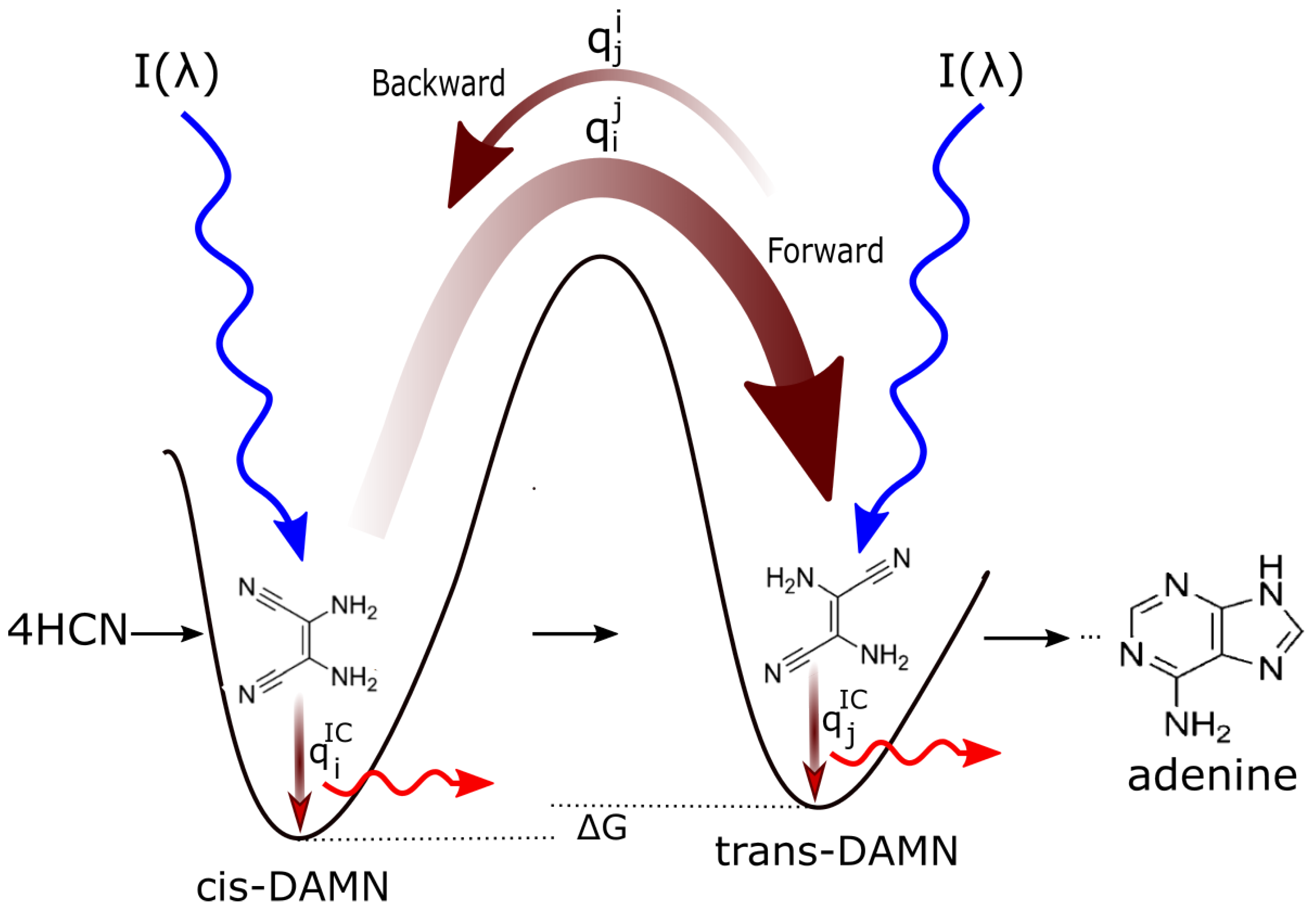
Figure 3.
The photochemical synthesis of adenine from 5 molecules of hydrogen cyanide (HCN) in water, as discovered by Ferris and Orgel (1966) [38,41]. Four molecules of HCN are transformed into the smallest stable oligomer (tetramer) of HCN, known as cis-2,3-diaminomaleonitrile (cis-DAMN) (2), which, under a constant UV-C photon flux isomerizes into trans-DAMN (3) (diaminofumaronitrile, DAFN). Further conversion on absorbing two more UV-C photons turns it into an imidazole intermediate, 4-amino-1H-imidazole-5-carbonitrile (AICN) (7). Hot ground state thermal reactions with another HCN molecule or its hydrolysis product formamide (or ammonium formate) leads to the purine adenine (8). This is a dissipative structuring process which ends in adenine, a pigment with a large molar extinction coefficient at 260 nm and a peaked conical intersection which promotes the dissipation of photons at the wavelength of maximum intensity of the Archean solar UVC spectrum (Figure 1). Adapted from Ferris and Orgel (1966) [38].
Figure 3.
The photochemical synthesis of adenine from 5 molecules of hydrogen cyanide (HCN) in water, as discovered by Ferris and Orgel (1966) [38,41]. Four molecules of HCN are transformed into the smallest stable oligomer (tetramer) of HCN, known as cis-2,3-diaminomaleonitrile (cis-DAMN) (2), which, under a constant UV-C photon flux isomerizes into trans-DAMN (3) (diaminofumaronitrile, DAFN). Further conversion on absorbing two more UV-C photons turns it into an imidazole intermediate, 4-amino-1H-imidazole-5-carbonitrile (AICN) (7). Hot ground state thermal reactions with another HCN molecule or its hydrolysis product formamide (or ammonium formate) leads to the purine adenine (8). This is a dissipative structuring process which ends in adenine, a pigment with a large molar extinction coefficient at 260 nm and a peaked conical intersection which promotes the dissipation of photons at the wavelength of maximum intensity of the Archean solar UVC spectrum (Figure 1). Adapted from Ferris and Orgel (1966) [38].
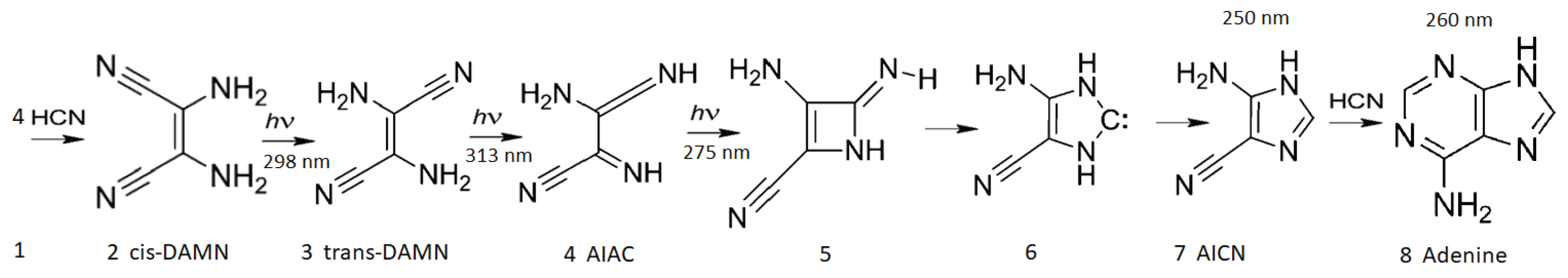
Figure 4.
The entropy production determined through simulation of the photochemical and chemical reactions on route to adenine [14] as a function of time in Archean days during the UV-C photochemical dissipative structuring process leading to adenine within a vesicle floating at the ocean surface at a temperature of µC. The entropy production generally increases monotonically, but not always. During the day (yellow parts of dashed line), entropy production is due to the dissipation of the UV-C spectrum into heat by the corresponding product concentration profile. At night, entropy production goes to zero (although thermal chemical reactions still occur during the night, this entropy production is small and not included in the figure). Taken with permission from Michaelian [14].
Figure 4.
The entropy production determined through simulation of the photochemical and chemical reactions on route to adenine [14] as a function of time in Archean days during the UV-C photochemical dissipative structuring process leading to adenine within a vesicle floating at the ocean surface at a temperature of µC. The entropy production generally increases monotonically, but not always. During the day (yellow parts of dashed line), entropy production is due to the dissipation of the UV-C spectrum into heat by the corresponding product concentration profile. At night, entropy production goes to zero (although thermal chemical reactions still occur during the night, this entropy production is small and not included in the figure). Taken with permission from Michaelian [14].
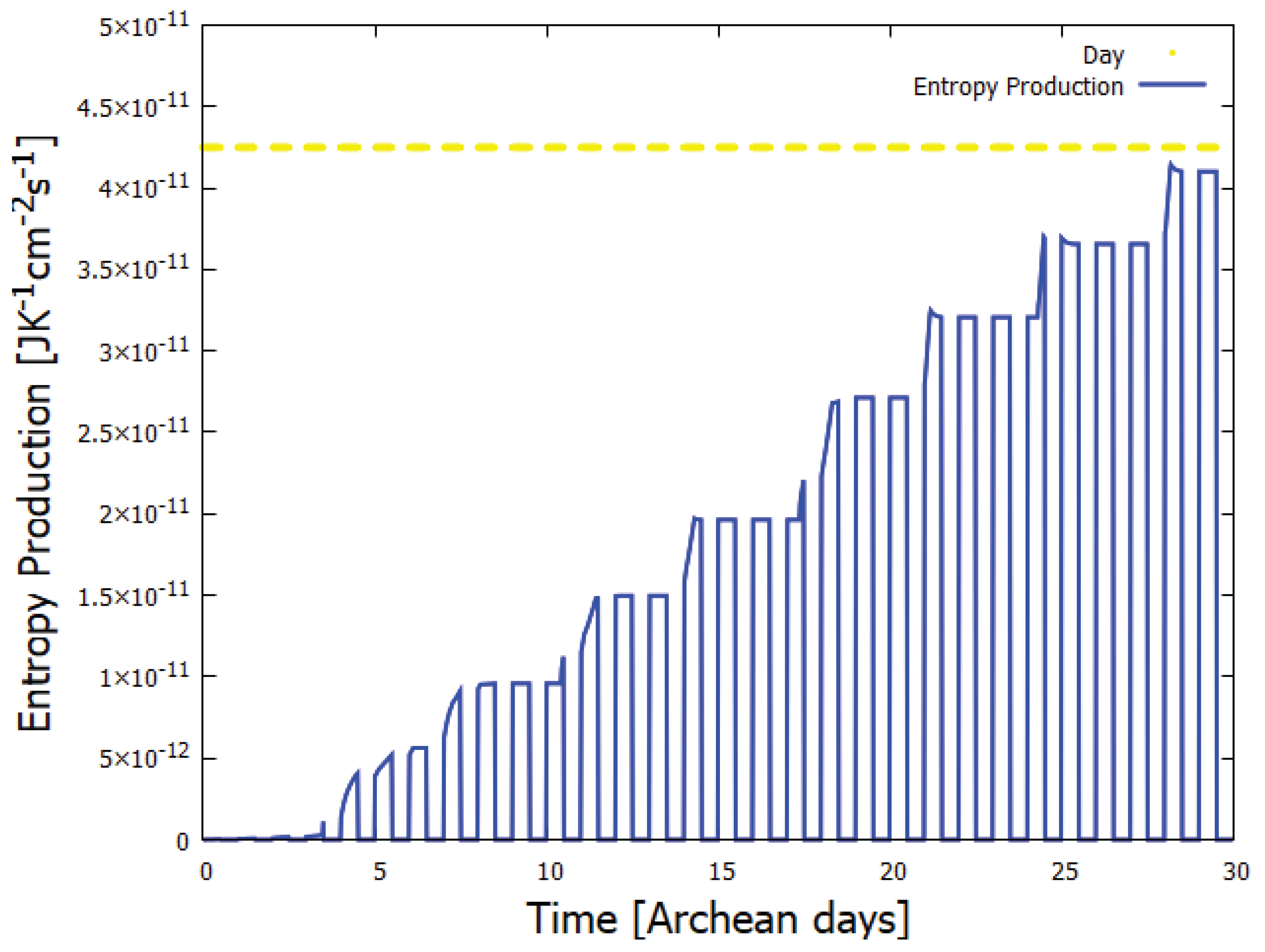
Figure 5.
The photochemical dissipative structuring of an 18 carbon atom fatty acid under UV-C+UV-B light. i) UV-C-induced reduction of CO2 and CO in water to form ethylene, ii) UV-C-induced polymerization of ethylene forming ever longer hydrocarbon chains, iii) oxidation and hydrolysis stop the growing of the chain and form the carboxyl group, iv) UV-C-induced deprotonation of the tail to form covalent double bonds, v) double bond migration to a conjugated triene, octadeca-12,14,16-trienoic acid (C18:3n-2), with a conical intersection. The final product absorbs strongly at nm [45] near to the peak of the incident Archean UV-C spectrum at nm (Figure 1).
Figure 5.
The photochemical dissipative structuring of an 18 carbon atom fatty acid under UV-C+UV-B light. i) UV-C-induced reduction of CO2 and CO in water to form ethylene, ii) UV-C-induced polymerization of ethylene forming ever longer hydrocarbon chains, iii) oxidation and hydrolysis stop the growing of the chain and form the carboxyl group, iv) UV-C-induced deprotonation of the tail to form covalent double bonds, v) double bond migration to a conjugated triene, octadeca-12,14,16-trienoic acid (C18:3n-2), with a conical intersection. The final product absorbs strongly at nm [45] near to the peak of the incident Archean UV-C spectrum at nm (Figure 1).
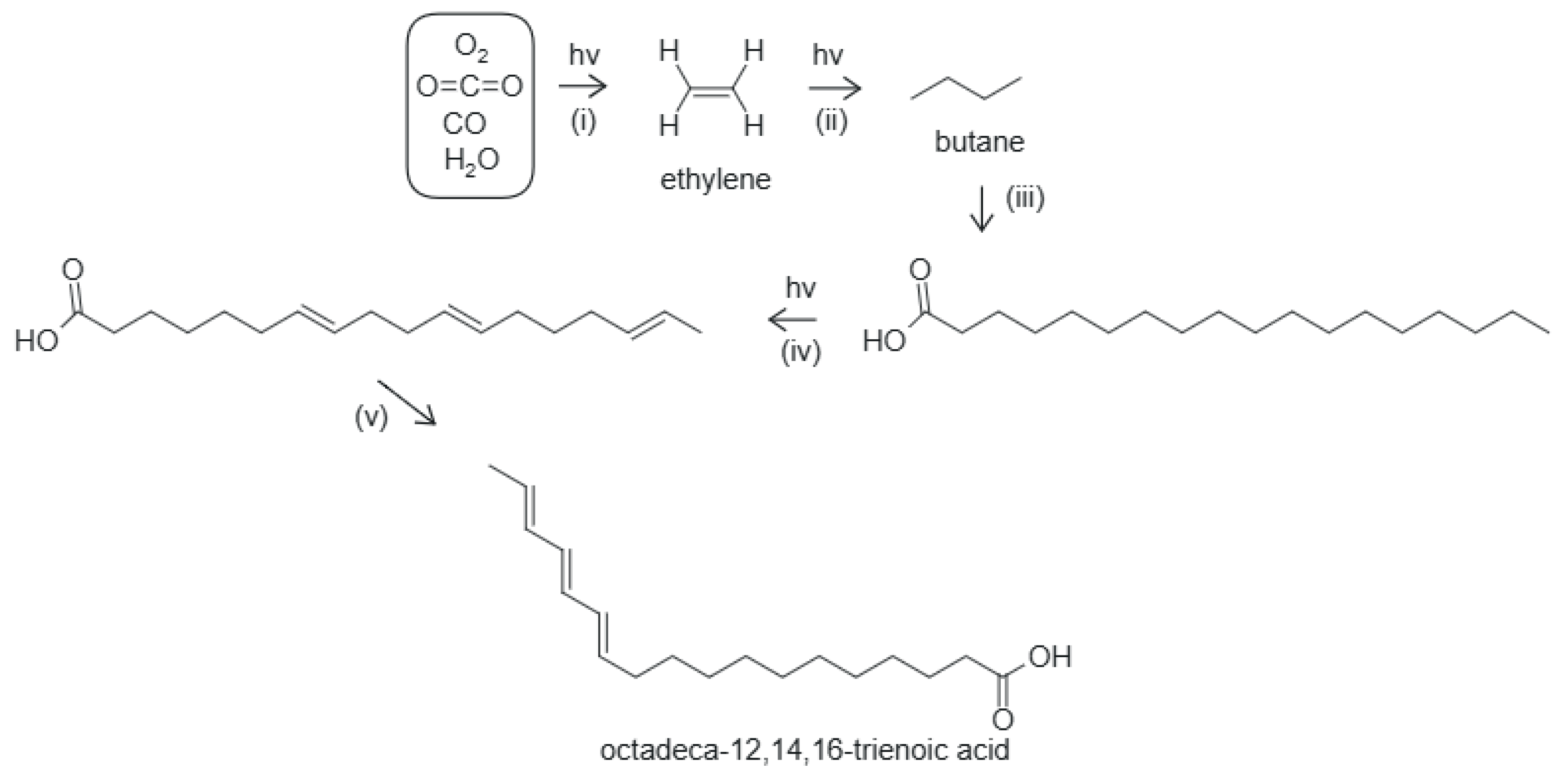
Figure 6.
The molar extinction coefficients at peak wavelengths for all molecules on route to chlorophyll a and b. The color code on the right gives the size of the extinction coefficient in units of M−1cm−1 and the location of the dots on the x-axis identifies the wavelengths of the resolved peaks. Protoporphyrin IX, and all later molecules, have a conical intersection for extremely rapid internal conversion to the ground state. The black curve represents Earth’s surface solar spectrum at the origin of life (3.85 Ga), the red curve is the same for 2.9 Ga, the green curve at 2.2 Ga and the yellow curve is today’s surface spectrum. Taken with permission from Michaelian and Simeonov [55].
Figure 6.
The molar extinction coefficients at peak wavelengths for all molecules on route to chlorophyll a and b. The color code on the right gives the size of the extinction coefficient in units of M−1cm−1 and the location of the dots on the x-axis identifies the wavelengths of the resolved peaks. Protoporphyrin IX, and all later molecules, have a conical intersection for extremely rapid internal conversion to the ground state. The black curve represents Earth’s surface solar spectrum at the origin of life (3.85 Ga), the red curve is the same for 2.9 Ga, the green curve at 2.2 Ga and the yellow curve is today’s surface spectrum. Taken with permission from Michaelian and Simeonov [55].
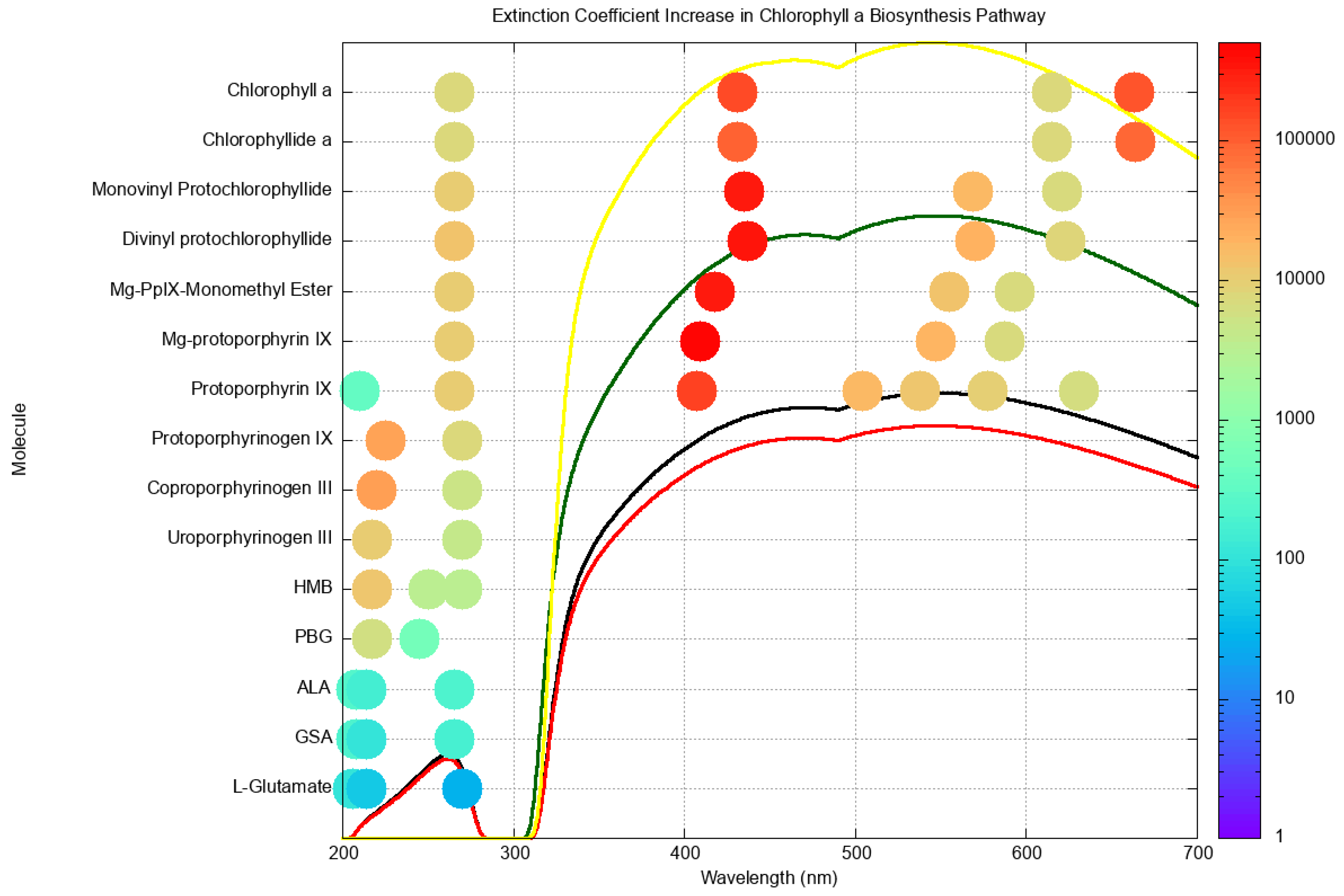
Figure 7.
A simplified 2-dimensional schematic representation of the entropy production surface (EPS) of a generalized phase space for a biosystem under a constant solar photon potential. The variables x and y at the origin of life may be, for example, the concentrations of different pigment molecules, while for an ecosystem of today, the variables may be the populations of different species. Three locally stable stationary states at local peaks in the entropy production surface are presented. On large enough external or internal perturbation, the system evolves from one stationary state to another. Although fluctuations are generally stochastic, the system will most often be found in those stationary states with a larger attraction basin and generally with a higher peak in photon dissipation (the stationary state labeled “3”). For molecules, this corresponds to concentration profiles with greater quantum efficiency for dissipation to the ground state through a conical intersection. For an ecosystem, this corresponds to animal and plant population profiles giving greater total photon dissipation (climax ecosystems). If the system began in stationary state 1, its most probable future evolution would be 1→2→3, but any combination is possible. For the biosphere, the x and y variables might be the number of species in two different clades and sub-peaks (not shown) corresponding to different species populations would exist on the main peaks and evolution would usually be local, among the sub-peaks. However, every once in a while a perturbation may be large enough (for example, an asteroid impact) to move the system from one main peak to another (e.g., 1 →3, mammals y becoming more prominent than dinosaurs x). Point, cyclic, or even chaotic dynamics are allowed superimposed on these peaks [66]. Autocatalytic stationary states have higher peaks and larger attraction basins in this generalized phase space and are thus more probable. The dimensionality of the generalized phase space is not fixed but evolves over time providing new “shorter” routes to larger peaks of entropy production (e.g., the re-introduction of a population of wolves into the ecosystem of Yellow Stone National Park, see text). Taken with permission from Michaelian [18].
Figure 7.
A simplified 2-dimensional schematic representation of the entropy production surface (EPS) of a generalized phase space for a biosystem under a constant solar photon potential. The variables x and y at the origin of life may be, for example, the concentrations of different pigment molecules, while for an ecosystem of today, the variables may be the populations of different species. Three locally stable stationary states at local peaks in the entropy production surface are presented. On large enough external or internal perturbation, the system evolves from one stationary state to another. Although fluctuations are generally stochastic, the system will most often be found in those stationary states with a larger attraction basin and generally with a higher peak in photon dissipation (the stationary state labeled “3”). For molecules, this corresponds to concentration profiles with greater quantum efficiency for dissipation to the ground state through a conical intersection. For an ecosystem, this corresponds to animal and plant population profiles giving greater total photon dissipation (climax ecosystems). If the system began in stationary state 1, its most probable future evolution would be 1→2→3, but any combination is possible. For the biosphere, the x and y variables might be the number of species in two different clades and sub-peaks (not shown) corresponding to different species populations would exist on the main peaks and evolution would usually be local, among the sub-peaks. However, every once in a while a perturbation may be large enough (for example, an asteroid impact) to move the system from one main peak to another (e.g., 1 →3, mammals y becoming more prominent than dinosaurs x). Point, cyclic, or even chaotic dynamics are allowed superimposed on these peaks [66]. Autocatalytic stationary states have higher peaks and larger attraction basins in this generalized phase space and are thus more probable. The dimensionality of the generalized phase space is not fixed but evolves over time providing new “shorter” routes to larger peaks of entropy production (e.g., the re-introduction of a population of wolves into the ecosystem of Yellow Stone National Park, see text). Taken with permission from Michaelian [18].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



