
Submitted:
14 January 2026
Posted:
15 January 2026
You are already at the latest version
Abstract
Cowpea (Vigna unguiculata) is a crop of major socioeconomic importance in Northern and Northeastern Brazil, but its productivity is strongly constrained by infestations of the black aphid Aphis craccivora, which causes direct damage through sap feeding and indirect losses by transmitting plant viruses. Although insecticides are widely used, their intensive application raises concerns related to resistance development and environmental impacts. Silicon (Si) has been investigated as a sustainable alternative for enhancing plant resistance to biotic and abiotic stresses by strengthening structural barriers and activating biochemical defense pathways. This study evaluated the influence of silicon fertilization on the plant–insect interaction between cowpea genotypes and A. craccivora. Three bioassays were conducted under greenhouse and laboratory conditions to assess aphid preference among genotypes, the effect of silicon on insect attractiveness, and plant susceptibility with and without silicon supplementation. Rice husk biochar and a commercial silicon source were used as Si fertilizers. Although the landrace genotype accumulated higher silicon levels, this did not result in reduced aphid populations or mitigation of plant damage. A negative correlation was observed between silicon accumulation and chlorophyll content, suggesting silicon-associated physiological adjustments under herbivory stress. Overall, under the evaluated conditions, silicon application did not confer effective resistance against A. craccivora, indicating that its role in integrated pest management strategies for cowpea requires further investigation.
Keywords:
induced resistance
; aphid herbivory
; sustainable fertilization
; host plant response
; integrated pest management
1. Introduction
Cowpea (Vigna unguiculata (L.) Walp.) plays a central role in food security and rural livelihoods in Northern and Northeastern Brazil, where it is widely cultivated by smallholder farmers. In these regions, traditional landraces are still commonly used, contributing to the conservation of genetic diversity and locally adapted traits that are essential for crop resilience under low-input production systems [1,2].
Despite its importance, cowpea productivity is strongly constrained by the black aphid, Aphis craccivora Koch (Hemiptera: Aphididae), a highly adaptable and cosmopolitan pest. This species rapidly colonizes young plants, causing direct damage through continuous sap extraction and indirect losses associated with the transmission of plant viruses [3]. Its short life cycle and high reproductive rate favor explosive population growth, often leading to severe infestations. Although chemical control remains the most common management strategy, its recurrent use raises concerns regarding environmental contamination, disruption of natural enemy communities, and the selection of resistant aphid populations [4]. These limitations highlight the need for alternative and complementary approaches within integrated pest management programs.
Among the strategies explored to reduce reliance on synthetic insecticides, silicon (Si) fertilization has received increasing attention. Although not classified as an essential element, silicon is known to enhance plant tolerance to both biotic and abiotic stresses by strengthening cell walls and modulating biochemical defense responses. Silicon accumulation in plant tissues may act as a physical barrier to herbivores and, in some cases, stimulate the production of secondary metabolites involved in plant defense. In this context, agricultural by-products such as rice husk-derived biochar represent a promising and low-cost source of silicon, particularly for small-scale farming systems.
Several studies have reported positive effects of silicon on plant growth, stress tolerance, and resistance to insect pests [5]. However, responses are highly variable and depend on plant genotype, insect feeding guild, and the form and timing of silicon application. In cowpea, evidence regarding the effectiveness of silicon against sap feeding insects such as aphids remains limited and sometimes contradictory, especially when traditional landraces are considered.
Despite the increasing use of silicon based fertilization as a sustainable tool for pest management, it remains unclear whether silicon supplementation can effectively modify the interaction between cowpea genotypes and the black aphid A. craccivora, particularly in terms of aphid preference and plant susceptibility.
This study was based on the following hypotheses: (i) silicon fertilization alters the plant–insect interaction by reducing the attractiveness of cowpea plants to A. craccivora; (ii) increased silicon accumulation in plant tissues decreases aphid performance and population growth; and (iii) silicon-induced physiological changes contribute to enhanced plant tolerance under aphid herbivory. Therefore, the objective of this study was to evaluate the role of silicon fertilization using rice husk biochar and a commercial silicon source on the interaction between cowpea genotypes and the black aphid A. craccivora. Specifically, we investigated aphid preference among genotypes, the effect of silicon on insect attractiveness, and plant susceptibility under silicon supplemented and non-supplemented conditions through a series of greenhouse and laboratory bioassays.
2. Materials and Methods
2.1. Experimental Site and Plant Material
The bioassays were conducted under controlled conditions in a greenhouse and in the Plant Protection Laboratory at the Federal University of Piauí, Professora Cinobelina Elvas Campus (UFPI–CPCE), located in Bom Jesus, Piauí, Brazil (9°05′04.4″ S; 44°19′37.5″ W). According to the Köppen climate classification, the region presents a tropical savanna climate with a dry winter season (Aw) [6].
Seeds of the cowpea landrace used in this study were obtained from a smallholder farmer in the municipality of Redenção do Gurguéia, Piauí, Brazil (9°29′10″ S; 44°35′15″ W).
For the genotype preference assay, eight cowpea genotypes were evaluated: six improved cultivars supplied by Embrapa Meio-Norte (BRS Aracê, TVu, BRS Tumucumaque, BRS Guariba, BRS Gurguéia, and BRS Novaera), one landrace collected from a local farmer (Crioulo), and one commercial variety purchased from a local market (Canetinha). Plants of the Canetinha variety were also used for the maintenance of the black aphid colony throughout the experimental period.
2.2. Soil Analysis
The soil used in the experiments was collected from the experimental area of the Federal University of Piauí (UFPI) and classified as a dystrophic Yellow Latosol according to the Brazilian Soil Classification System [7]. Chemical and particle-size characterization was performed following the procedures described by Teixeira et al. [8].
Soil silicon concentration was determined using the colorimetric molybdate method, as described by Vieira and Silva [9]. The results of the soil chemical and granulometric characterization are presented in Appendix A.1.
2.3. Rice Husk Carbonization (Biochar Production)
The rice husk used in this study was obtained from an industrial rice-processing facility. The carbonization process was carried out in a rectangular kiln made of galvanized steel sheets. Combustion was manually controlled and monitored until the material reached the characteristic black coloration indicative of the desired carbonization stage.
To interrupt the thermal process, the biochar was moistened and subsequently dried under natural solar radiation. After drying, the material was stored in sealed containers to prevent moisture absorption. A representative sample was collected and subjected to chemical analysis following the methodology described by Miyazawa et al. [10] to determine mineral nutrient contents. The biochar presented a silicon concentration of 13.54 g kg−1. The results of the chemical characterization of the biochar are provided in Appendix A.2.
2.4. Bioassay 1: Preference Among Vigna unguiculata Cultivars
The assay was conducted using eight cowpea (Vigna unguiculata) genotypes: BRS Aracê, TVu, BRS Tumucumaque, BRS Guariba, BRS Gurguéia, BRS Novaera, a landrace variety (Crioulo), and a commercial variety (Canetinha). For comparative purposes, four genotypes with previously reported resistance levels were included: BRS Tumucumaque, BRS Gurguéia, and BRS Novaera (susceptible), and BRS Guariba (resistant), as described by Silva et al. [11].
Before sowing, seeds were treated with the fungicide Vitavax®–Thiram 200 SC. The genotypes were sown in 500 mL disposable polystyrene cups, perforated at the bottom to ensure adequate drainage and aeration. Each cup was labeled and filled with 700 g of soil. Two seeds were planted per cup, and the plants were irrigated daily to prevent water stress.
Seven days after sowing, thinning was performed to maintain one plant per cup. Each plant represented an experimental unit, and the experiment followed a completely randomized design in an 8 × 7 arrangement (eight treatments and seven replicates). Irrigation was monitored twice daily throughout the experimental period to avoid any water deficit.
At sixteen days after sowing, when plants reached the V3 growth stage, each plant was infested with five apterous adult female aphids of Aphis craccivora, characterized by their glossy black coloration. All insects were five days old and obtained from a previously standardized colony. Aphids were allowed to move freely among the evaluated genotypes, characterizing the assay as a free-choice test.
Two days after infestation (DAI), the number of live adult aphids present on each plant was recorded, and the insects were subsequently removed. Four days after infestation, the number of live nymphs on each genotype was quantified. Based on these data, the ratio between the number of nymphs and the initial number of adults (nymph/adult ratio) was calculated, following the methodology proposed by Silva et al. [12].
The evaluated variables in this bioassay were: number of adults, number of nymphs, and the nymph/adult ratio.
2.5. Bioassay 2: Preference of Aphis craccivora for Vigna unguiculata Under Silicon Fertilization
This assay was conducted in plastic pots containing 2.5 kg of soil. Liming and base fertilization were performed according to regional recommendations [13]. The experimental design was completely randomized in a 2 × 3 factorial scheme, consisting of two cowpea genotypes (the landrace Crioulo and the cultivar BRS Novaera), three treatments (two silicon sources and a control), and ten replicates. The cultivar BRS Novaera was selected because of its previously reported higher capacity for silicon uptake [14].
The silicon sources used were rice husk biochar (18.5 g per pot) and a commercial silicon oxide product (0.537 g per pot). Both sources were adjusted to supply 0.1 g of silicon per kg of soil, following the protocol adapted from Leite et al. [14]. Prior to sowing, seeds were treated as described in Bioassay 1.
Silicon sources were incorporated into the soil at the time of pot filling, according to the previously determined rates. Five seeds were sown per pot, and thinning was performed seven days after emergence, leaving only the most vigorous plant in each pot, which was considered the experimental unit. Throughout the experiment, plants were manually irrigated twice daily, in the morning and afternoon.
At sixteen days after sowing, when plants reached the V3 growth stage, each plant was infested with five apterous adult female aphids of Aphis craccivora, characterized by their glossy black coloration. All insects were five days old and obtained from a previously standardized colony. Aphids were allowed to move freely among the evaluated plants, characterizing the assay as a free-choice test. Two days after infestation (DAI), the number of live adult aphids present on each plant was recorded and the insects were subsequently removed. Four days after infestation, the number of live nymphs on each experimental unit was quantified.
In addition, leaf chlorophyll content was measured before and after infestation using a portable chlorophyll meter (ClorofiLOG CFL 1030, Falker). After nymph removal, the first fully expanded trifoliate leaf from each plant was collected for silicon content analysis following the methodology described by Miyazawa et al. [10]. Aboveground plant tissues were also harvested to determine fresh mass and dry mass after oven-drying at forced air circulation for 72 h.
The variables evaluated in this bioassay were: number of adults, number of nymphs, nymph/adult ratio, chlorophyll content, and silicon accumulation [12].
2.6. Bioassay 3: Susceptibility of Silicon-Fertilized Vigna unguiculata (Landrace) to Aphis craccivora over Time
This bioassay was conducted in plastic pots filled with 10 kg of soil. Liming and base fertilization were performed according to regional recommendations [13]. A total of 36 experimental units were established in a completely randomized design, consisting of three treatments (two silicon sources and a control) with twelve replicates each.
The silicon sources used were rice husk biochar (74 g per pot) and a commercial silicon oxide product (2.151 g per pot). Both sources were adjusted to supply 0.1 g of silicon per kg of soil, following the protocol adapted from Leite et al. [14].
Seeds were treated with fungicide as described for Bioassay 1. Silicon sources were incorporated into the soil at the time of pot preparation. Five seeds were sown per pot (Figure 1a), and thinning was performed seven days after sowing, leaving one plant per pot, which was considered the experimental unit. Throughout the experiment, plants were manually irrigated twice daily.
Ten days after sowing, plants were infested with ten apterous adult female aphids of Aphis craccivora, all five days old and obtained from a previously standardized colony. Aphids were allowed to move freely among the evaluated plants, characterizing the assay as a free-choice test (Figure 1b).
Evaluations were carried out at 7, 14, 21, and 28 days after infestation (DAI). The following parameters were assessed: leaf chlorophyll content (Figure 1c), aphid population level (POP), and seedling damage (DM). Population levels and plant damage were evaluated according to the criteria proposed by Jackai and Singh [15], using a visual rating scale from 1 to 9 for aphid population density and from 1 to 5 for damage severity.
2.7. Statistical Analysis
Data were analyzed separately for each bioassay using RStudio software (version 12.1, 2023). For Bioassay 2, the effects of silicon treatments and cowpea genotypes on aphid biological parameters and chlorophyll content were evaluated using a two-way analysis of variance (ANOVA). The significance of main effects and interactions was assessed using the F-test at a 5% probability level (p ≤ 0.05).
When significant differences were detected, treatment means were compared using Tukey’s honestly significant difference (HSD) test at p ≤ 0.05. Graphs were generated using SigmaPlot software (version 15.0, Systat Software, 2022), and Pearson’s correlation analyses were performed in RStudio.
3. Results
3.1. Bioassay 1 – Preference Among Cowpea Genotypes
Differences among cowpea genotypes were detected in relation to the number of Aphis craccivora nymphs (Figure 2). The lowest nymph densities were recorded on the Canetinha and BRS Tumucumaque genotypes, whereas the landrace (Crioulo) showed the highest infestation levels. In contrast, no significant differences were observed among genotypes for the number of adult aphids, indicating a similar initial colonization pattern across treatments.
These results suggest that, although adult aphid settlement did not vary among genotypes, differences in nymph production were sufficient to discriminate levels of susceptibility among the evaluated materials.
3.2. Bioassay 2 – Preference Under Silicon Fertilization
Silicon fertilization did not significantly affect the number of adult or nymph aphids in either of the cowpea genotypes evaluated (Figure 3). Regardless of the silicon source applied, aphid abundance remained statistically similar to the control treatment. However, silicon accumulation differed between genotypes, with the landrace showing higher silicon content in plant tissues than the cultivar BRS Novaera.
Chlorophyll concentrations measured before and after infestation also did not differ significantly between treatments or between genotypes (Figure 4). These results indicate that, under the conditions tested, silicon fertilization did not alter either aphid preference or the main physiological parameters associated with photosynthetic capacity.
Similarly, when data were analyzed considering silicon sources independently of genotype, no significant differences were detected for aphid abundance, silicon accumulation, or chlorophyll-related parameters (Figure 5 and Figure 6). These findings reinforce that, in this experimental context, silicon fertilization did not modify the interaction between cowpea plants and A. craccivora.
A significant interaction between genotype and silicon treatment was observed for initial chlorophyll b content (p ≤ 0.01; Figure 7). Under control conditions, BRS Novaera exhibited higher chlorophyll b values than the landrace. However, when silicon sources were applied, chlorophyll b levels in the landrace increased and became similar to those observed in BRS Novaera.
Pearson correlation analysis revealed a strong positive relationship between the number of adult aphids and the number of nymphs, indicating that higher adult abundance was associated with increased offspring production (Figure 8). A negative correlation was observed between the number of adults and the nymph/adult ratio, suggesting an inverse relationship between adult density and relative reproductive output.
Strong correlations were also detected among chlorophyll variables, particularly between chlorophyll a, chlorophyll b, and total chlorophyll at both the beginning and the end of the experiment. In contrast, silicon accumulation showed a moderate negative correlation with final chlorophyll a, b, and total chlorophyll levels, indicating that higher silicon content tended to be associated with lower chlorophyll values. No significant correlations were found between silicon accumulation and aphid abundance, suggesting that silicon content did not directly influence aphid population density.
3.3. Bioassay 3 – Susceptibility over Time
In the time-course evaluation of the landrace genotype subjected to different silicon sources, significant effects were associated with the evaluation period rather than with silicon treatments.
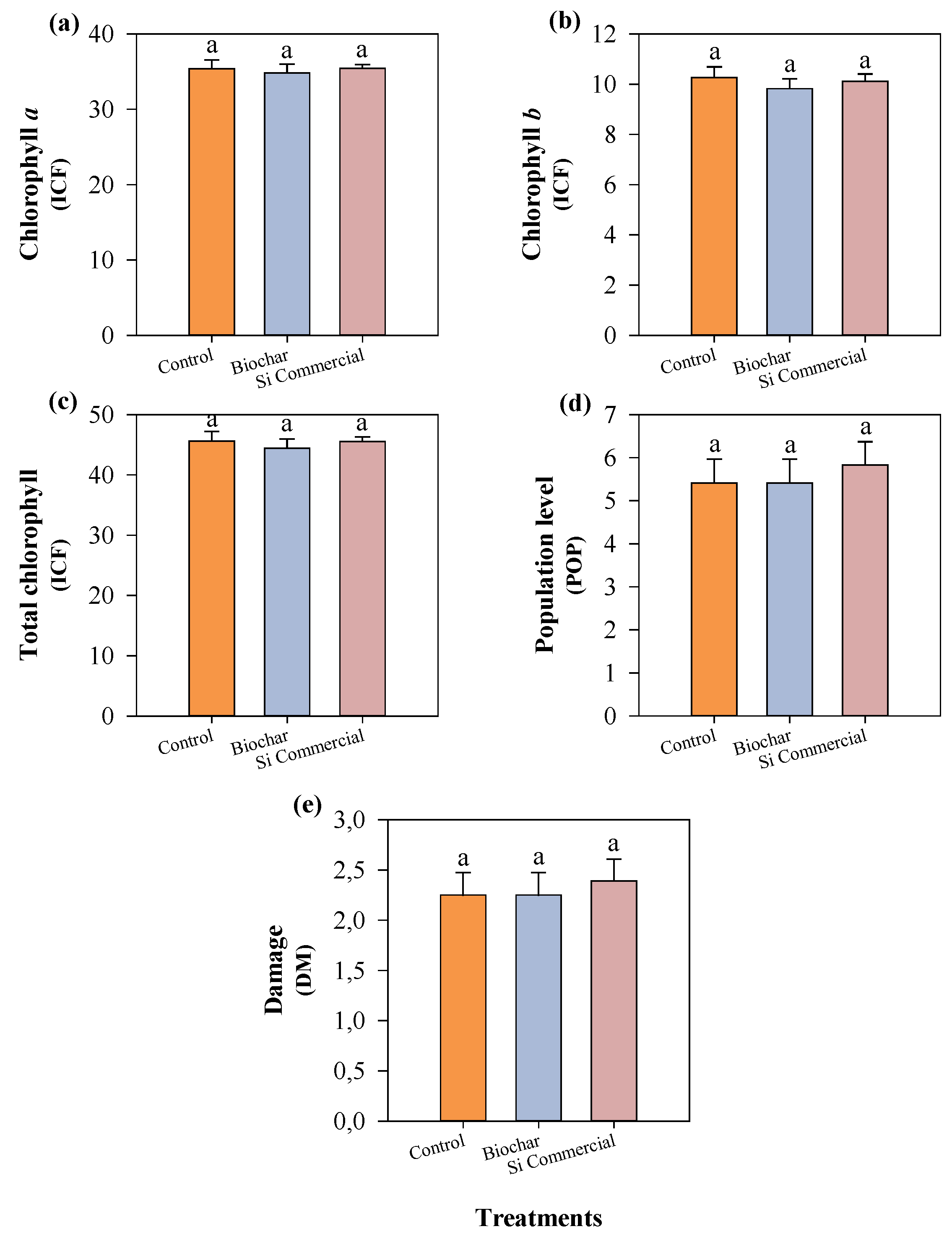
Chlorophyll a and total chlorophyll decreased significantly in the fourth evaluation (Figure 9). Chlorophyll b reached its highest values during the third evaluation and subsequently declined. Aphid population levels increased progressively over time, peaking at the third evaluation, while plant damage scores also increased consistently, with significant differences among evaluation dates (p ≤ 0.05).
The increase in chlorophyll levels during the third week coincided with the peak in aphid population and the intensification of plant damage. Nevertheless, neither rice husk biochar nor commercial silicon altered any of the evaluated parameters throughout the experimental period (Figure 10).
4. Discussion
This study evaluated the effects of silicon on the management of Aphis craccivora in Vigna unguiculata, focusing on resistance mechanisms (antixenosis and antibiosis) and plant physiological responses. Although silicon treatments (rice husk biochar and commercial silicon) did not promote significant reductions in black aphid populations or improve chlorophyll maintenance, these outcomes can be explained by a combination of biological, environmental, and genetic factors.
Plant resistance to insects involves direct mechanisms, such as antibiosis and antixenosis, as well as indirect defenses mediated by the attraction of natural enemies. In Bioassay 1, the statistical uniformity observed for adult aphids suggests that the evaluated genotypes did not differ in terms of attractiveness or suitability for adult development of A. craccivora. In contrast, variation in the number of nymphs represents an indicator of antibiosis [16]. A plant-mediated effect on insect biology was detected in the BRS Tumucumaque and Canetinha genotypes, which showed lower offspring production and reduced development. The absence or low expression of antixenosis or antibiosis, even in the cultivar BRS Guariba previously described as resistant [17,18] may be associated with the occurrence of distinct A. craccivora biotypes.
Aphid biotypes can overcome plant resistance mechanisms through genetic adaptations [19]. In addition, reports of A. craccivora biotypes in the African continent [20] reinforce the hypothesis that the aphid population used in this study may differ from those against which Brazilian cowpea varieties have shown resistance. Plant insect interactions are also strongly influenced by environmental factors. Genotypes previously described as resistant may exhibit susceptibility under different environmental conditions [21]. Furthermore, the expression of resistance genes can be inhibited by epistatic interactions, compromising plant defense [22].
Silicon is widely recognized for its ability to enhance plant resistance to biotic and abiotic stresses. It accumulates in cell walls as amorphous silica, forming physical barriers against herbivorous insects and microorganisms [23]. In addition, silicon may induce systemic resistance by stimulating the production of phenolic compounds, lignin, and antioxidant enzymes [24]. However, in the present study, the higher silicon accumulation observed in the landrace genotype in Bioassay 2 did not result in detectable benefits in aphid population control or chlorophyll maintenance.
Although some studies have demonstrated that silicon can reduce insect infestation [25], responses vary according to plant species, insect species, application method, and experimental conditions. For instance, lower densities of black aphid nymphs have been reported in lima bean when silicon was applied simultaneously to foliage and soil, with the effect attributed to antibiosis-based resistance [26]. Nevertheless, the effectiveness of silicon depends on its interaction with hormonal signaling pathways that regulate the expression of defense-related genes [24].
Herbivory activates complex hormonal pathways, particularly those mediated by jasmonic acid (JA) and salicylic acid (SA). JA is mainly associated with defenses against chewing insects, whereas SA plays a central role in responses to sap-feeding insects [27]. These pathways often display antagonistic interactions, in which the activation of one may suppress the other, thereby limiting the effectiveness of defensive responses [28,29]. This antagonism may help explain the absence of significant silicon effects in the present study, as SA activation induced by aphids may have inhibited JA-dependent responses, such as the accumulation of defensive secondary metabolites.
In addition, studies have shown that silicon can influence these hormonal pathways. Stimuli that activate the JA pathway promotes greater silicon accumulation in Brachypodium distachyon [30]. However, the predominance of SA signaling in response to sap-feeding insects such as aphids may reduce silicon accumulation and limit its effectiveness in inducing resistance.
In Bioassay 3, which evaluated susceptibility over time, the reduction in total chlorophyll levels during the fourth week can be attributed to the increase in damage caused by black aphid infestation, which compromises the photosynthetic capacity of plants. Biotic stress may initially stimulate compensatory mechanisms, such as increased chlorophyll content, but these responses tend to fail under severe damage conditions [31]. The negative correlations observed between silicon accumulation and final chlorophyll levels suggest that silicon may be associated with physiological stress responses, including the reallocation of energetic resources toward defense at the expense of photosynthesis [32]. However, the lack of significant correlations between silicon accumulation and insect abundance indicates that, under the conditions of this experiment, silicon did not directly influence aphid population dynamics.
The absence of significant effects of rice husk biochar and commercial silicon across all evaluated variables suggests that, under the conditions tested, silicon did not provide additional advantages for the management of A. craccivora. Nevertheless, it is important to consider that exposure time, application rates, and application methods may have influenced these results.
Future studies should incorporate biochemical and molecular analyses to better understand the interaction between silicon and hormonal signaling pathways, as well as its influence on the production of secondary metabolites. Field experiments are also necessary to assess silicon effectiveness under diverse environmental conditions and against different A. craccivora biotypes. In addition, research on the effects of silicon on the attractiveness of natural enemies, such as predators and parasitoids, may provide valuable insights into its potential role in integrated pest management programs.
5. Conclusions
The cowpea landrace evaluated in this study proved to be susceptible to infestation by Aphis craccivora, with no evidence of resistance mediated by antixenosis or antibiosis mechanisms. The application of silicon sources, including rice husk biochar and commercial silicon, did not induce significant defensive responses against A. craccivora in any of the evaluated genotypes. Although plants exhibited physiological adjustments to stress caused by aphid infestation, such as an initial increase in chlorophyll levels, the landrace genotype was unable to maintain homeostasis as herbivory damage progressed.
Author Contributions
João Vitor Soares Morais: Conceptualization; Investigation; Methodology; Writing—review & editing; Thayline Rodrigues de Oliveira: Data curation, Investigation; Methodology, Visualization; Ana Clara Marcelino de Moura: Data curation; Methodology; Visualization; Sara dos Santos Nunes: Data curation; Methodology; Visualization; Danile Marques Pacheco: Data curation; Formal analysis; Software; Writing—review & editing Bruno Ettore Pavan: Formal analysis; Writing—review & editing Jose Bruno Malaquias: Formal analysis; Writing—review & editing; Funding acquisition; Supervision; Jenilton Gomes da Cunha: Data curation; Formal analysis; Software; Writing—review & editing. Luciana Barboza Silva: Conceptualization; Investigation; Funding acquisition; Writing—original draft; Writing—review & editing; Supervision; Resources; Project administration.
Funding
This research was funded by Coordination for the Improvement of Higher Education Personnel (CAPES), Brazil, for the scholarship and resource provided, and the Federal University of Piauí, Brazil, for providing logistical support.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
The authors gratefully acknowledge the institutional support provided by the Fundação de Amparo à Pesquisa do Estado do Piauí (FAPEPI), and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), which were fundamental to the development of this research. Institutional support from the Universidade Federal do Piauí (UFPI), Campus Professora Cinobelina Elvas (CPCE), is duly acknowledged, particularly for providing laboratory infrastructure, administrative assistance, and technical support throughout the execution of the study.
Conflicts of Interest
The authors declare that there is no conflict of interest regarding the publication of this article.
Abbreviations
The following abbreviations are used in this manuscript:
| DIC | Delineamento Inteiramente Casualizado |
| DAI | Dias Após Infestação |
Appendix A
Appendix A.1

Table A1.
Caracterização química e granulométrica do solo utilizado no experimento.
| pH | H+Al | Al | Ca | Mg | K | SB | T | Si | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| (H2O) | ----------------------------- cmolc dm-3 ---------------------------- | -----mg kg-1---- | |||||||||
| 5,0 | 3,35 | 0,40 | 1,08 | 0,17 | 0,05 | 1,30 | 4,65 | 6,42 | |||
| P | Cu | Mn | Fe | Zn | m | V | M.O | Argila | Silte | Areia | |
| ------------------ mg dm-3 -------------------- | -------%------- | --------------------g kg-1----------------- | |||||||||
| 6,2 | 0,04 | 18,49 | 137 | 0,04 | 23,5 | 28 | 10,19 | 219 | 24 | 757 | |
Note: H+AL: acidez potencial; Al: alumínio; Ca: cálcio; Mg: magnésio; K: potássio; SB: soma de bases; T: CTC pH 7; Si: Silício; P: fósforo; Cu: cobre; Mn: manganês; Fe: ferro; Zn: zinco; m: saturação por alumínio; V: saturação de bases; e MO: matéria orgânica.
Appendix A.2
Table A2.
Caracterização química da casca de arroz como fonte de silício utilizado no experimento.
| Ca | Mg | P | K | Si | Cu | Mn | Fe | Zn | |
|---|---|---|---|---|---|---|---|---|---|
| ---------m-g kg-1------------ | ----------g kg-1--------- | --------------------Mg kg-1-------------------- | |||||||
| 0,1 | 0,1 | 0,54 | 0,5 | 13,54 | 1,85 | 241 | 125 | 16,35 | |
Note: Ca: cálcio; Mg: magnésio; P: fósforo; K: potássio; Si: Silício; Cu: cobre; Mn: manganês; Fe: ferro e Zn: zinco.
References
- LIMA, A. F.; SILVA, E. G. A.; IWATA, B. F. Agriculturas e agricultura familiar no Brasil: uma revisão de literatura. Retratos de Assentamentos 2019, v. 22(n. 1), 50–68. [Google Scholar] [CrossRef]
- PUNEETH, G. M.; GOWTHAMI, R.; KATRAL, A.; LAXMISHA, K. M.; VASUDEVA, R.; SINGH, G. P.; ARCHAK, S. On-farm crop diversity, conservation, importance and value: a case study of landraces from Western Ghats of Karnataka, India. Scientific Reports 2024, v. 14(n. 1), 10712. [Google Scholar] [CrossRef] [PubMed]
- SILVA, L. C.; NERE, D. R.; FERREIRA, A. D. C. L.; BLEICHER, E. Resistance in advanced cowpea lines to Aphis craccivora Koch, 1854 (Hemiptera: Aphididae). Revista Agro@ mbiente On-line 2019, v. 13, 185–195. [Google Scholar] [CrossRef]
- SAYED, S. M.; ALOTAIBI, S. A.; GABER, N.; ELARRNAOUTY, S. S. Evaluation of five medicinal plant extracts on Aphis craccivora (Hemiptera: Aphididae) and its predator, Chrysoperla carnea (Neuroptera: Chrysopidae) under laboratory conditions. Insects 2020, v. 11(n. 6), 398. [Google Scholar] [CrossRef]
- KOVÁCS, S.; KUTASY, E.; CSAJBÓK, J. The multiple role of silicon nutrition in alleviating environmental stresses in sustainable crop production. Plants 2022, v. 11(n. 9), 1223. [Google Scholar] [CrossRef]
- HOLANDA, R. M.; MEDEIROS, R. M. Classificação climática pelo método de Köppen e Thornthwaite em Bom Jesus do Piauí, Brasil. Revista Pantaneira 2019, v. 16., 57–68. Available online: https://periodicos.ufms.br/index.php/revpan/article/view/9075.
- SANTOS, H. G.; JACOMINE, P. K. T.; ANJOS, L. H. C.; OLIVEIRA, V. A.; LUMBRERAS, J. F.; COELHO, M. R.; ALMEIDA, J. A.; ARAÚJO FILHO, J. C.; OLIVAIRA, J. B.; CUNHA, T. J. F. Sistema Brasileiro de Classificação de Solos, 2 ed.; 2018. [Google Scholar]
- TEIXEIRA, P. C.; DONAGEMMA, G. K.; FONTANA, A.; TEIXEIRA, W. G. Manual de métodos de análise de solo. In Revista e ampliada, 3 ed.; 2017; p. pp. 573. [Google Scholar]
- VIEIRA, W.; SILVA, F. C.; Silva, F. C. Análises de fertilizantes minerais, organominerais e corretivos. In Manual de análises químicas de solos, plantas e fertilizantes, 2 ed.; Embrapa/CNPTIA, 2009; pp. 191–233. [Google Scholar]
- MIYAZAWA, M.; PAVAN, M. A.; MURAOKA, T.; CARMO, C. A. F. S.; MELO, W. J.; Silva, F. C. Análise de tecido vegetal. In Manual de análises químicas de solos, plantas e fertilizantes, 2 ed.; 2009; pp. 191–233. [Google Scholar]
- SILVA, L.C.; NERE, D. R.; BLEICHER, E.; BARBOSA, A. V. C.; TAVARES, J. M. Demographic parameters of black aphid in cowpea cultivars. Ciência Agrícola 2018, v. 16(n. 1), 69–76. Available online: http://www.seer.ufal.br/index.php/revistacienciaagricola/article/view/3782/3659.
- SILVA, J. F. Resistência genética de Vigna unguilata (L.) Walp. À Aphis craccivora e amostragem. Dissertação (mestrado); Universidade Federal do Ceará, Centro de Ciências Agrárias, Departamento de Fitotecnia, Programa de Pós-Graduação em Agronomia/Fitotecnia: Fortaleza, 2011. Available online: https://repositorio.ufc.br/handle/riufc/8495.
- SOUSA, D. M. G.; LOBATO, E. Cerrado: correção do solo e adubação. In Embrapa Informação Tecnológica; 2004; p. 416. [Google Scholar]
- LEITE, W. S.; MIRANDA, R. S.; ROCHA, M. M.; DUTRA, A. F.; SANTOS, A. S.; SILVA, A. C.; BRITO, F. M.; SOUSA, R. S.; ARAÚJO, A. S. F.; NASCIMENTO, C. W. A.; ALCÂNTARA NETO, F. Silicon alleviates drought damage by increasing antioxidant and photosynthetic performance in cowpea. Journal of Agronomy and Crop Science 2023, v. 209(n. 6), 772–787. [Google Scholar] [CrossRef]
- JACKAI, L. E. N.; SINGH, S. R. Screening techniques for host plant resistance to cowpea insect pests. 1988. Available online: https://biblio.iita.org/documents/U88ArtJackaiScreeningNothomNodev.pdf-c3c0b879438fec8d62d56908894651f5.pdf.
- BISCHOFF, A. M.; ARAUJO, E. S.; BENATTO, A.; ZIMMERMANN, R. C.; OLIVEIRA, M. C. H.; ROSA, J. M.; BERNARDI, D.; ZAWADNEAK, M. A. C. Evidence of antibiosis resistance of four strawberry cultivars against Duponchelia fovealis (Lepidoptera: Crambidae). Crop Protection 2023, v. 168, 106213. [Google Scholar] [CrossRef]
- MORAES, J. G. L.; BLEICHER, E. Preferência do pulgão-preto, Aphis craccivora Koch, a diferentes genótipos de feijão-de-corda, Vigna unguiculata (L.) Walp. Ciência Rural 2007, v. 37(n. 6), 1554–1557. [Google Scholar] [CrossRef]
- SILVA, L.C.; NERE, D. R.; BLEICHER, E.; BARBOSA, A. V. C.; TAVARES, J. M. Demographic parameters of black aphid in cowpea cultivars. Ciência Agrícola 2018, v. 16(n. 1), 69–76. Available online: http://www.seer.ufal.br/index.php/revistacienciaagricola/article/view/3782/3659. [CrossRef]
- VAN EMDEN, H. F.; HARRINGTON, R. Aphids as crop pests. In CAB International, 2 ed.; 2017. [Google Scholar]
- OFUYA, T. I. Control of the cowpea aphid, Aphis craccivora Koch (Homoptera: Aphididae), in cowpea, Vigna unguiculata (L.) Walp. Integrated Pest Management Reviews 1997, n. 2, 199–207. [Google Scholar] [CrossRef]
- SILVA, J. F.; BLEICHER, E. Resistência de genótipos de feijão-de-corda ao pulgão-preto. Pesquisa Agropecuária Brasileira 2010, v.45, 1089–1094. [Google Scholar] [CrossRef]
- JANDER, G. Revisiting plant-insect interactions. Annual Review of Plant Biology 2018, 69, 147–171. [Google Scholar]
- EPSTEIN, E. Silicon: its manifold roles in plants. Annals of applied Biology 2009, v. 155(n. 2), 155–160. [Google Scholar] [CrossRef]
- RANJAN, A.; SINHA, R.; BALA, M.; PAREEK, A.; SINGLA-PAREEK, S. L.; SINGH, A. K. Silicon-mediated abiotic and biotic stress mitigation in plants: Underlying mechanisms and potential for stress resilient agriculture. Plant Physiology and Biochemistry 2021, v. 163, 15–25. [Google Scholar] [CrossRef]
- Yan, G.; Huang, Q.; Zhao, S.; Xu, Y.; He, Y.; Nikolic, M.; NIKOLIC, N.; LIANG; Zhu, Y.Z. Silicon nanoparticles in sustainable agriculture: synthesis, absorption, and plant stress alleviation. Frontiers in Plant Science 2024, v. 15, 1393458. [Google Scholar] [CrossRef]
- PORTELA, G. L. F.; SILVA, P. R. R..; GIRÃO FILHO, J. E.; PÁDUA, L. E. M.; MELO JÚNIOR, L. C. Silício como indutor de resistência no controle do pulgão preto Aphis craccivora Koch, 1854 em fava Phaseolus lunatus. Arquivos do Instituto Biológico 2019, v. 86, e0512018. [Google Scholar] [CrossRef]
- ERB, M.; REYMOND, P. Molecular interactions between plants and insect herbivores. Annual review of plant biology 2019, v. 70(n. 1), 527–557. [Google Scholar] [CrossRef]
- THALER, J. S.; HUMPHREY, P. T.; WHITEMAN, N. K. Evolution of jasmonate and salicylate signal crosstalk. Trends in plant science 2012, v. 17(n. 5), 260–270. [Google Scholar] [CrossRef]
- JOHNSON, S. N.; ROWE, R. C.; HALL, C. R. Aphid feeding induces phytohormonal cross-talk without affecting silicon defense against subsequent chewing herbivores. Plants 2020, v. 9(n. 8), 1009. [Google Scholar] [CrossRef]
- JOHNSON, S. N.; HARTLEY, S. E.; RYALLS, J. M. W.; FREW, A.; HALL, C. R. Targeted plant defense: silicon conserves hormonal defense signaling impacting chewing but not fluid-feeding herbivores. Ecology 2021, v. 102(n. 3). [Google Scholar] [CrossRef]
- CHAVES, V. V. Trocas gasosas e fluorescência da clorofila “a” em genótipos de cana-de-açúcar infestados por cigarrinha-das-raízes Mahanarva fimbriolata. Dissertação (Mestrado em Fitotecnia), Universidade Federal de Viçosa, Viçosa, 2015. [Google Scholar]
- COSKUN, D.; DESHMUKH, R.; SONAH, H.; MENZIES, J. G.; REYNOLDS, O.; MA, J. F.; KRONZUCKER, H. J.; BÉLANGER, R. R. The controversies of silicon’s role in plant biology. New Phytologist 2019, 221(1), 67–85. [Google Scholar] [CrossRef]
Figure 1.
Germination of cowpea seeds four days after sowing (a); transfer of black aphids to landrace cowpea plants (b); and the ClorofiLOG device used for chlorophyll measurements (c).
Figure 1.
Germination of cowpea seeds four days after sowing (a); transfer of black aphids to landrace cowpea plants (b); and the ClorofiLOG device used for chlorophyll measurements (c).

Figure 2.
Feeding preference of the black aphid (Aphis craccivora) on cowpea (Vigna unguiculata) genotypes. Means followed by the same lowercase letters do not differ significantly according to Tukey’s test (p ≤ 0.05). Bars represent the mean ± standard error.
Figure 2.
Feeding preference of the black aphid (Aphis craccivora) on cowpea (Vigna unguiculata) genotypes. Means followed by the same lowercase letters do not differ significantly according to Tukey’s test (p ≤ 0.05). Bars represent the mean ± standard error.
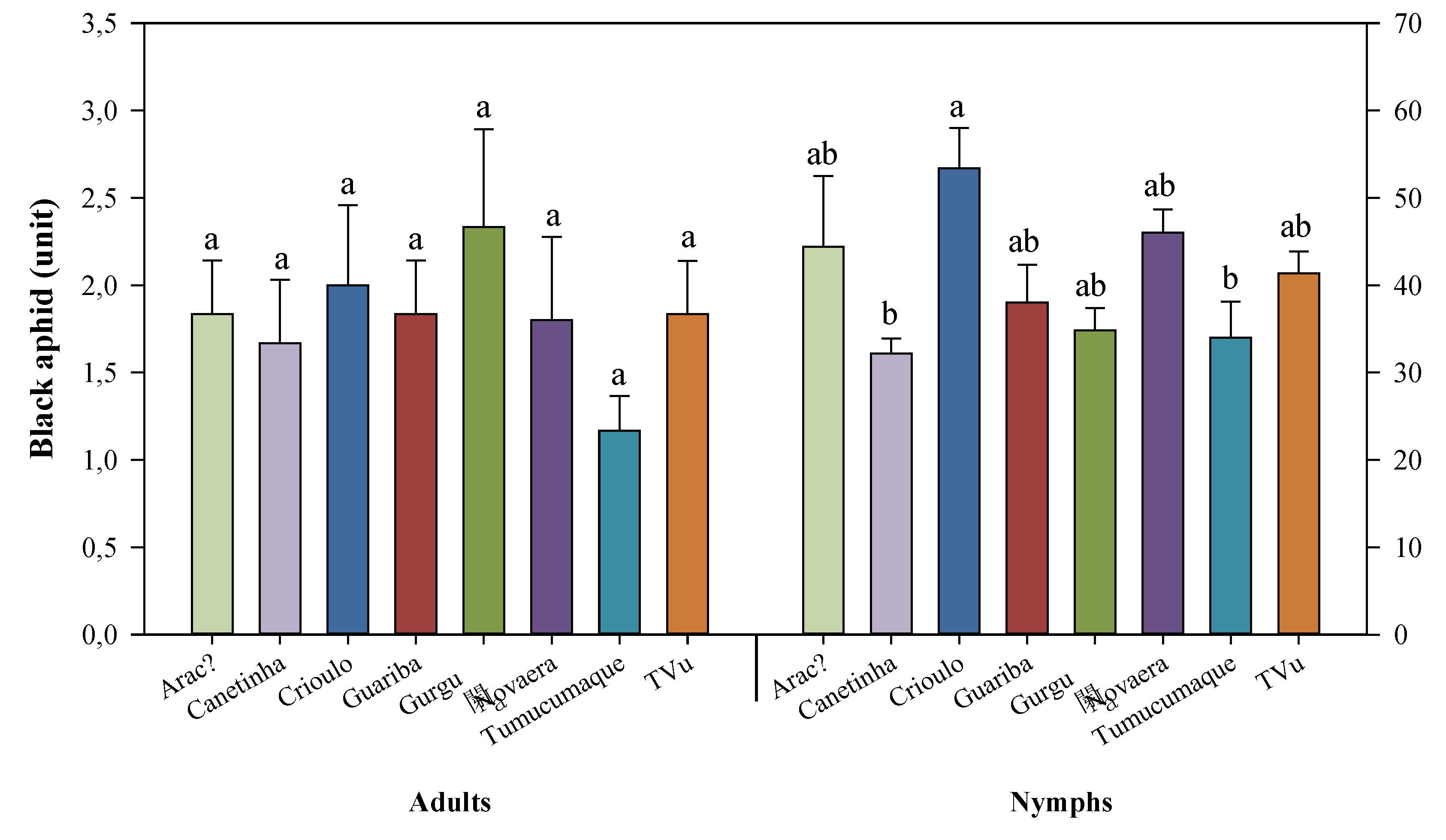
Figure 3.
Number of adult aphids after two days (a), number of nymphs after four days (b), nymph/adult ratio (c), and silicon accumulation (d) in two cowpea genotypes cultivated with different silicon sources and exposed to black aphid herbivory. Means followed by the same lowercase letters do not differ significantly according to the F-test (p ≤ 0.05). Bars represent the mean ± standard error.
Figure 3.
Number of adult aphids after two days (a), number of nymphs after four days (b), nymph/adult ratio (c), and silicon accumulation (d) in two cowpea genotypes cultivated with different silicon sources and exposed to black aphid herbivory. Means followed by the same lowercase letters do not differ significantly according to the F-test (p ≤ 0.05). Bars represent the mean ± standard error.
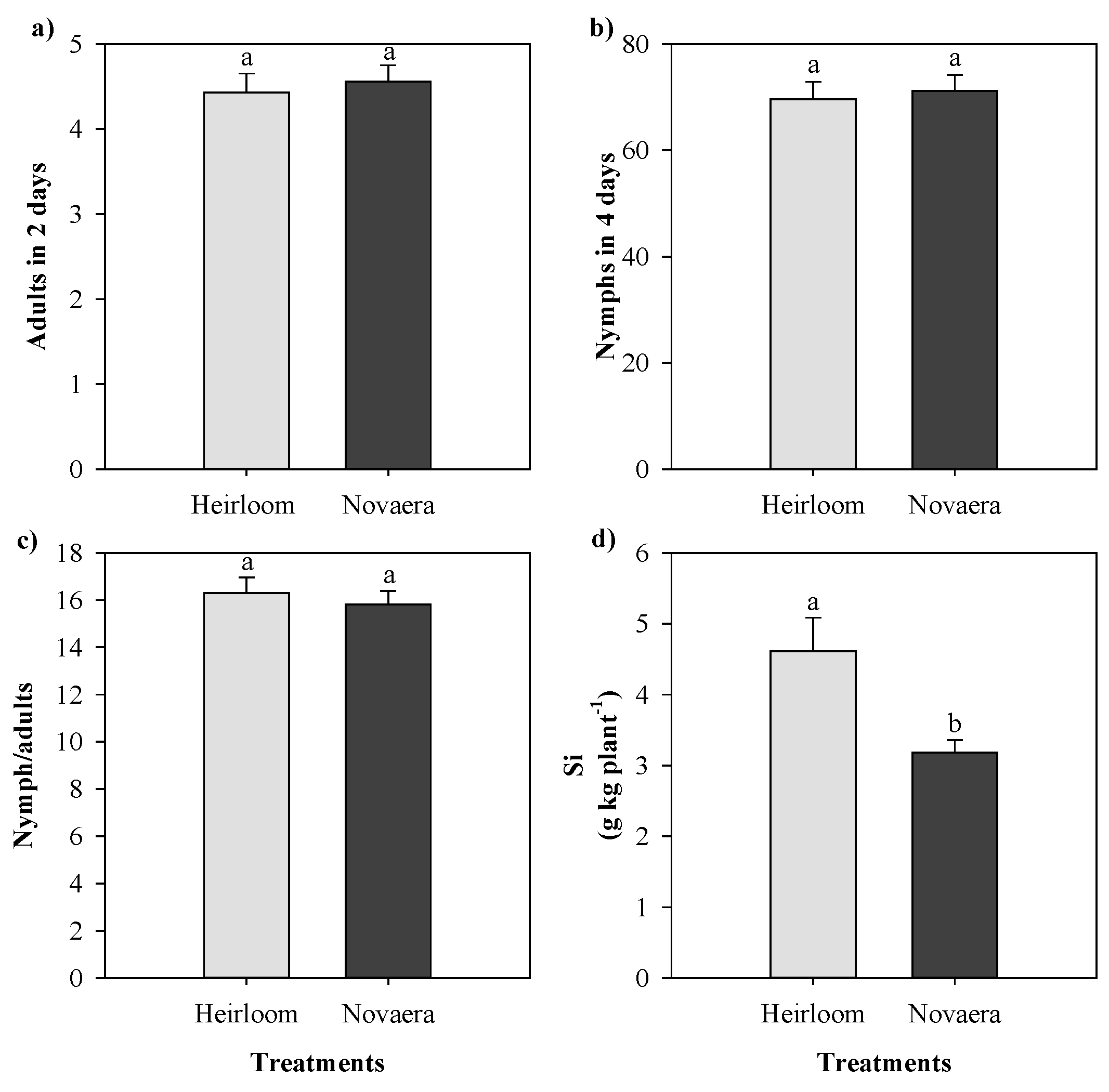
Figure 4.
Initial chlorophyll a (a), final chlorophyll a (b), initial total chlorophyll (c), final total chlorophyll (d), and final chlorophyll b (e) in two cowpea genotypes cultivated with different silicon sources before and after black aphid infestation. Means followed by the same lowercase letters do not differ significantly according to the F-test (p ≤ 0.05). Bars represent the mean ± standard error.
Figure 4.
Initial chlorophyll a (a), final chlorophyll a (b), initial total chlorophyll (c), final total chlorophyll (d), and final chlorophyll b (e) in two cowpea genotypes cultivated with different silicon sources before and after black aphid infestation. Means followed by the same lowercase letters do not differ significantly according to the F-test (p ≤ 0.05). Bars represent the mean ± standard error.
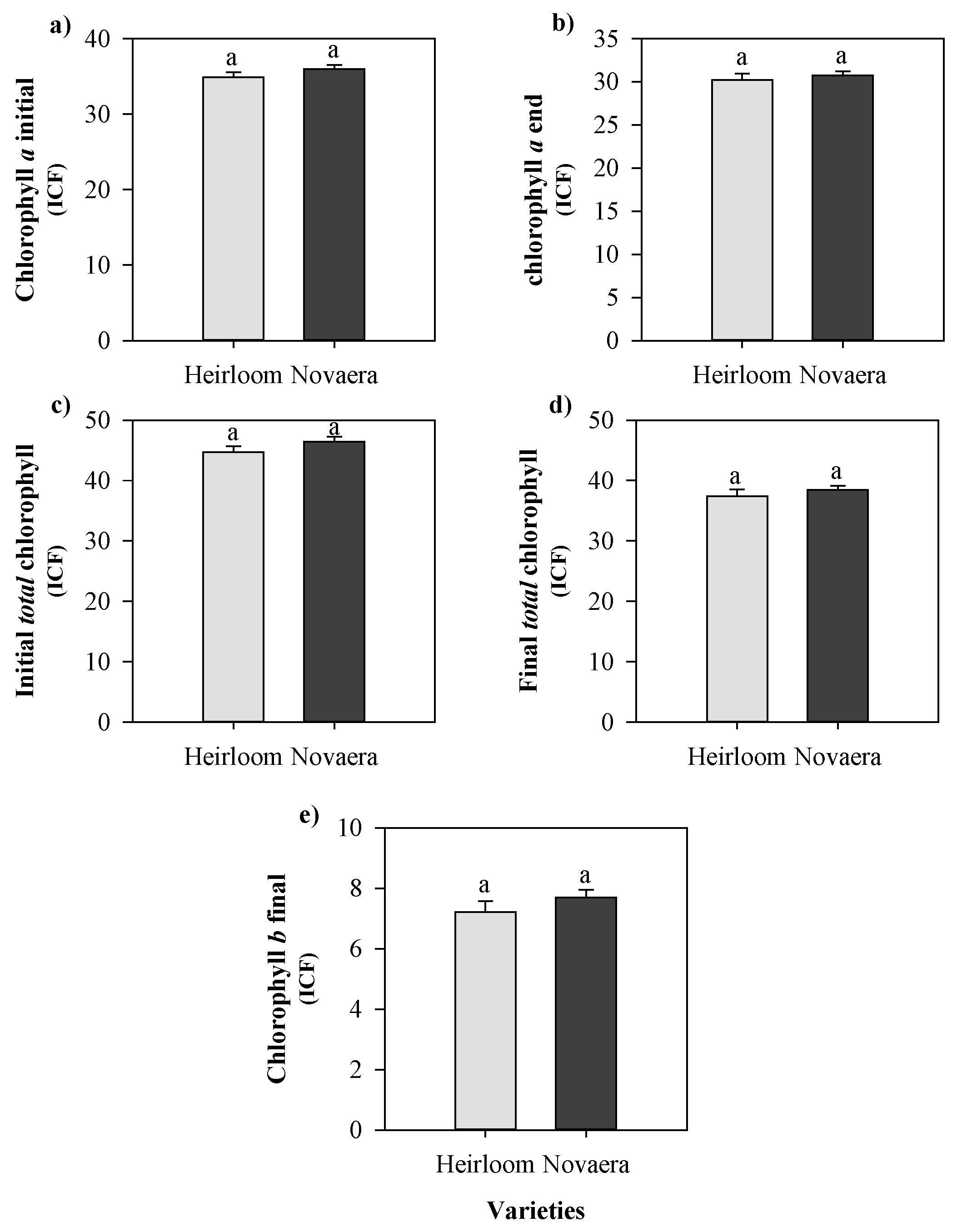
Figure 5.
Number of adult aphids after two days (a), number of nymphs after four days (b), nymph/adult ratio (c), and silicon accumulation (d) in cowpea plants cultivated with different silicon sources and exposed to black aphid herbivory. Means followed by the same lowercase letters do not differ significantly according to the F-test (p ≤ 0.05). Bars represent the mean ± standard error.
Figure 5.
Number of adult aphids after two days (a), number of nymphs after four days (b), nymph/adult ratio (c), and silicon accumulation (d) in cowpea plants cultivated with different silicon sources and exposed to black aphid herbivory. Means followed by the same lowercase letters do not differ significantly according to the F-test (p ≤ 0.05). Bars represent the mean ± standard error.
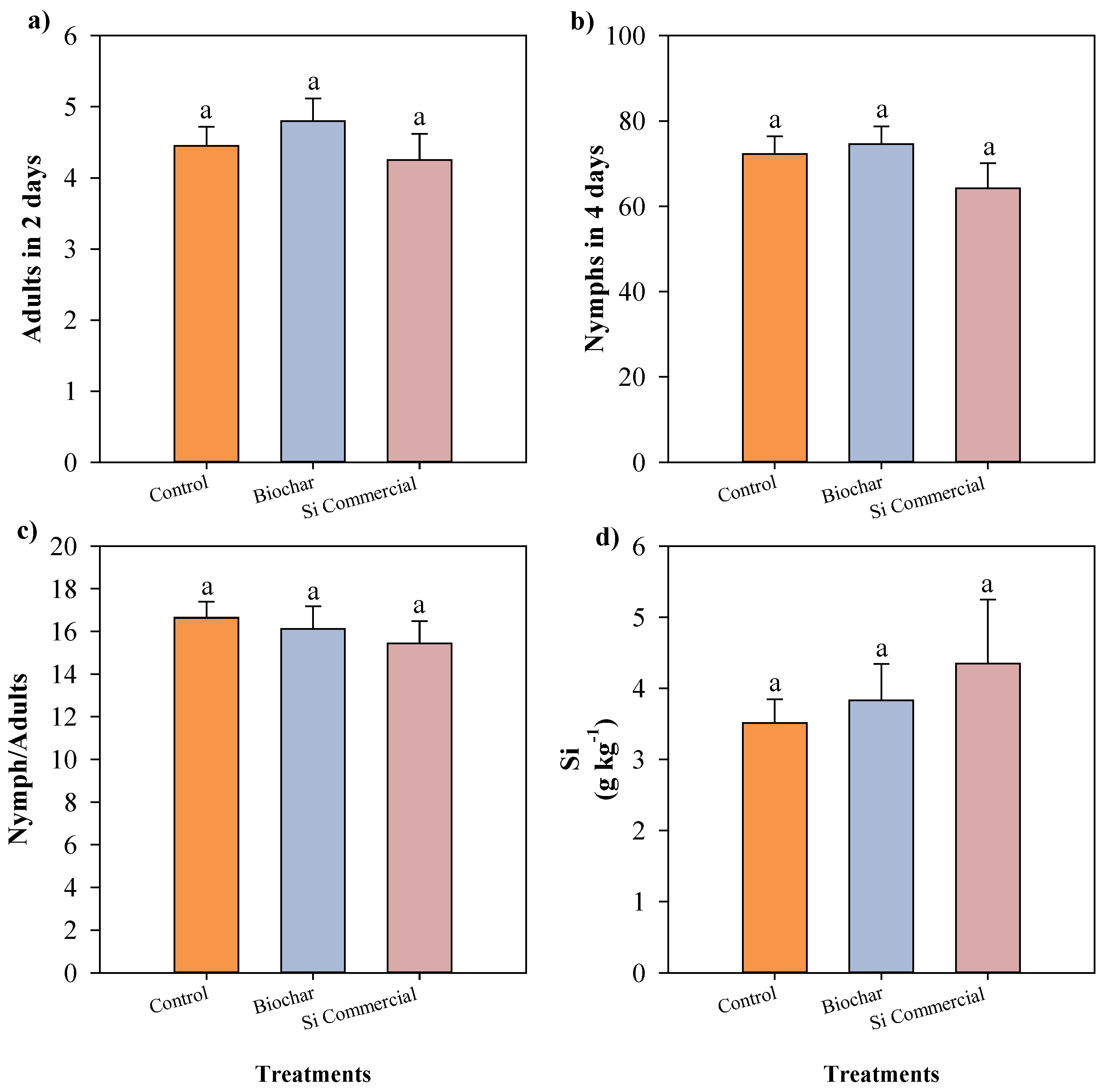
Figure 6.
Initial chlorophyll a (a), final chlorophyll a (b), initial total chlorophyll (c), final total chlorophyll (d), and final chlorophyll b (e) in cowpea plants cultivated with different silicon sources before and after black aphid infestation. Means followed by the same lowercase letters do not differ significantly according to the F-test (p ≤ 0.05). Bars represent the mean ± standard error.
Figure 6.
Initial chlorophyll a (a), final chlorophyll a (b), initial total chlorophyll (c), final total chlorophyll (d), and final chlorophyll b (e) in cowpea plants cultivated with different silicon sources before and after black aphid infestation. Means followed by the same lowercase letters do not differ significantly according to the F-test (p ≤ 0.05). Bars represent the mean ± standard error.
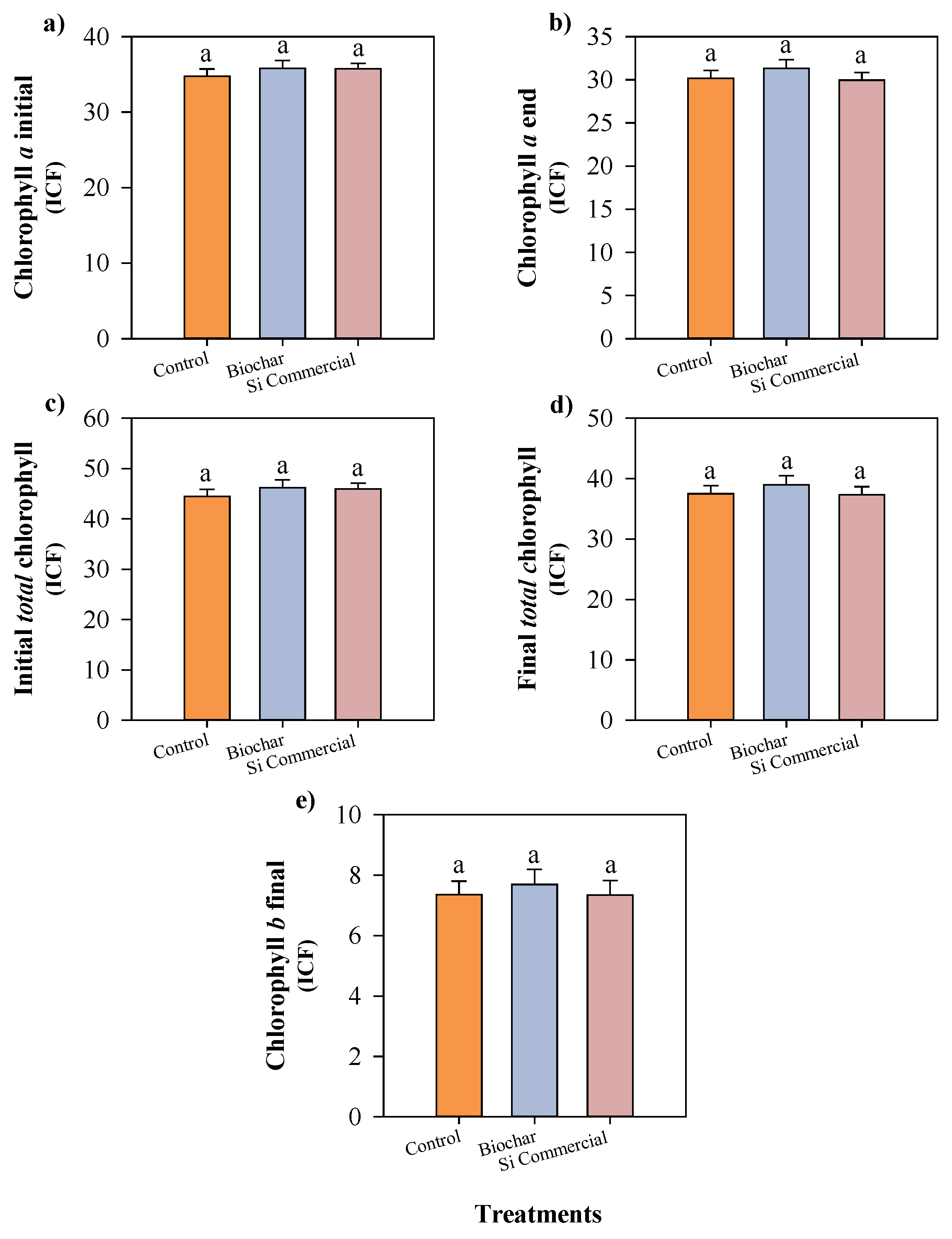
Figure 7.
Initial chlorophyll b levels in cowpea plants of the landrace (Crioulo) and BRS Novaera subjected to different silicon treatments. Means followed by the same uppercase letters among treatments and lowercase letters among genotypes do not differ significantly according to Tukey’s test (p ≤ 0.05). Bars represent the mean ± standard error.
Figure 7.
Initial chlorophyll b levels in cowpea plants of the landrace (Crioulo) and BRS Novaera subjected to different silicon treatments. Means followed by the same uppercase letters among treatments and lowercase letters among genotypes do not differ significantly according to Tukey’s test (p ≤ 0.05). Bars represent the mean ± standard error.
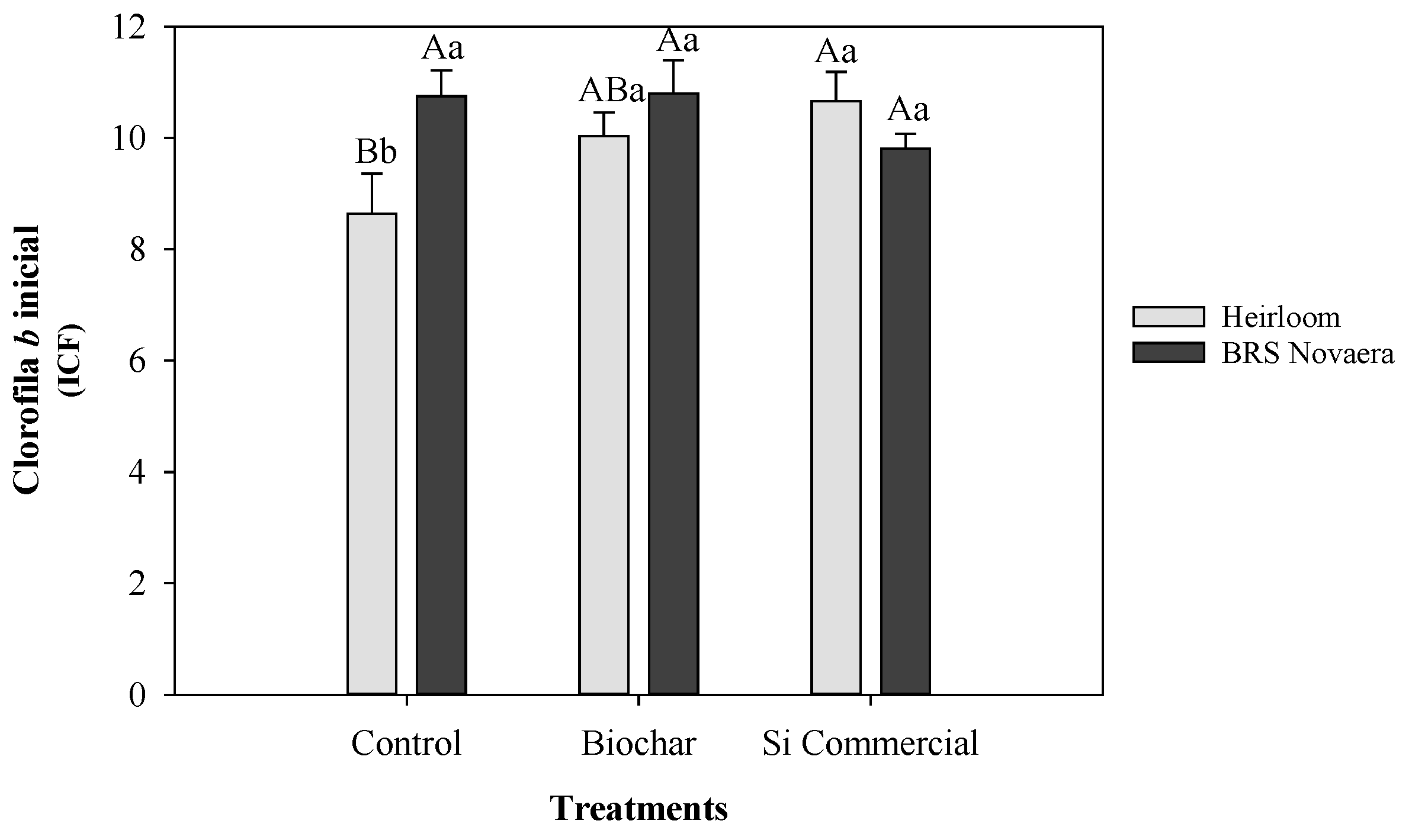
Figure 8.
Pearson correlation matrix among the following variables: number of adults at 48 h, number of nymphs at four days, nymph/adult ratio (NA), initial chlorophyll a, initial chlorophyll b, initial total chlorophyll, final chlorophyll a, final chlorophyll b, final total chlorophyll, and silicon accumulation. Values closer to 1 indicate strong positive correlations, whereas values closer to −1 indicate strong negative correlations.
Figure 8.
Pearson correlation matrix among the following variables: number of adults at 48 h, number of nymphs at four days, nymph/adult ratio (NA), initial chlorophyll a, initial chlorophyll b, initial total chlorophyll, final chlorophyll a, final chlorophyll b, final total chlorophyll, and silicon accumulation. Values closer to 1 indicate strong positive correlations, whereas values closer to −1 indicate strong negative correlations.
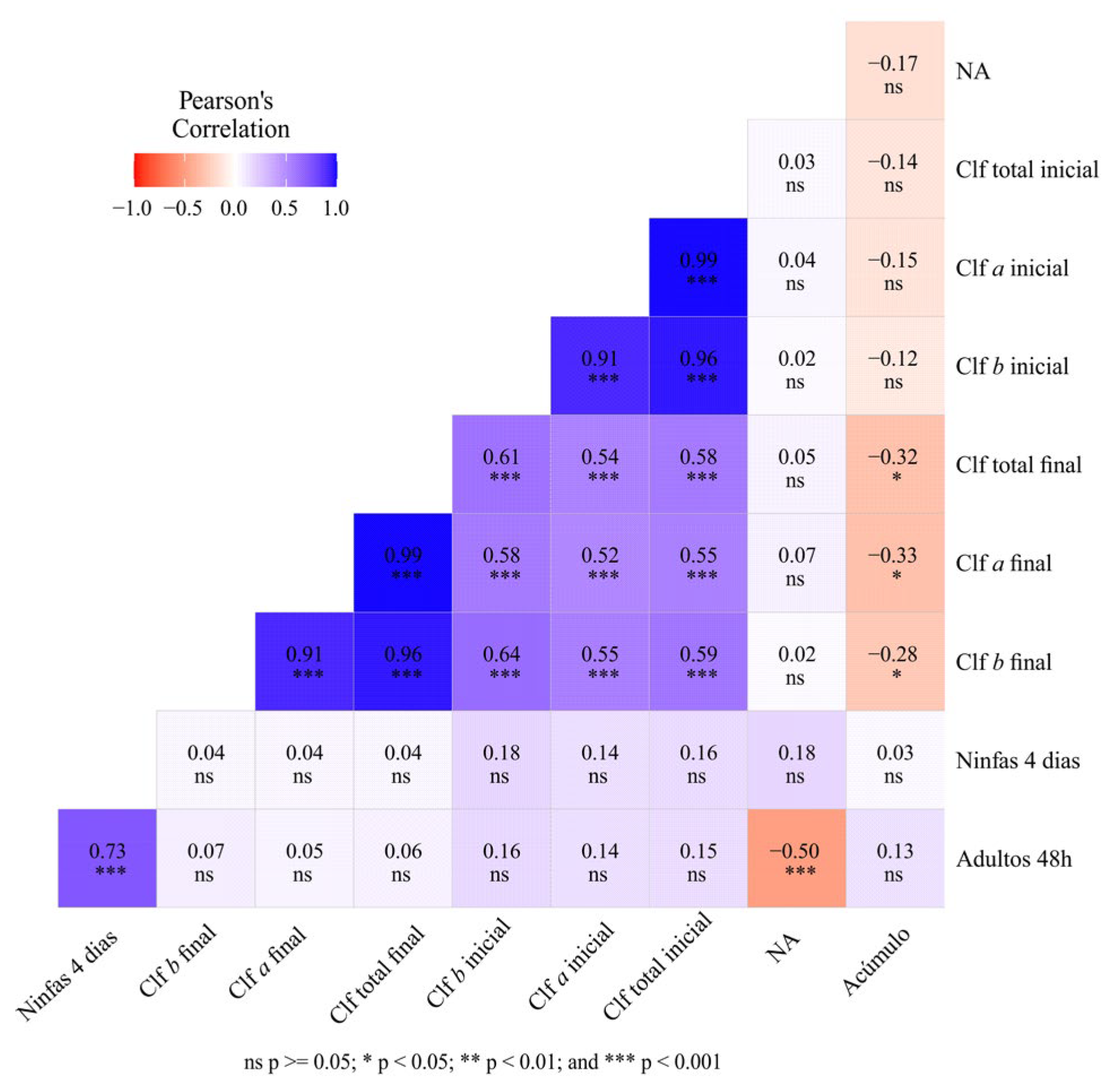
Figure 9.
Chlorophyll a (a), chlorophyll b (b), total chlorophyll (c), aphid population level (d), and damage to cowpea seedlings cultivated with different silicon sources and subjected to black aphid herbivory, evaluated at 7, 14, 21, and 28 days after infestation (e). Means followed by the same lowercase letters do not differ significantly according to the F-test (p ≤ 0.05). Bars represent the mean ± standard error.
Figure 9.
Chlorophyll a (a), chlorophyll b (b), total chlorophyll (c), aphid population level (d), and damage to cowpea seedlings cultivated with different silicon sources and subjected to black aphid herbivory, evaluated at 7, 14, 21, and 28 days after infestation (e). Means followed by the same lowercase letters do not differ significantly according to the F-test (p ≤ 0.05). Bars represent the mean ± standard error.
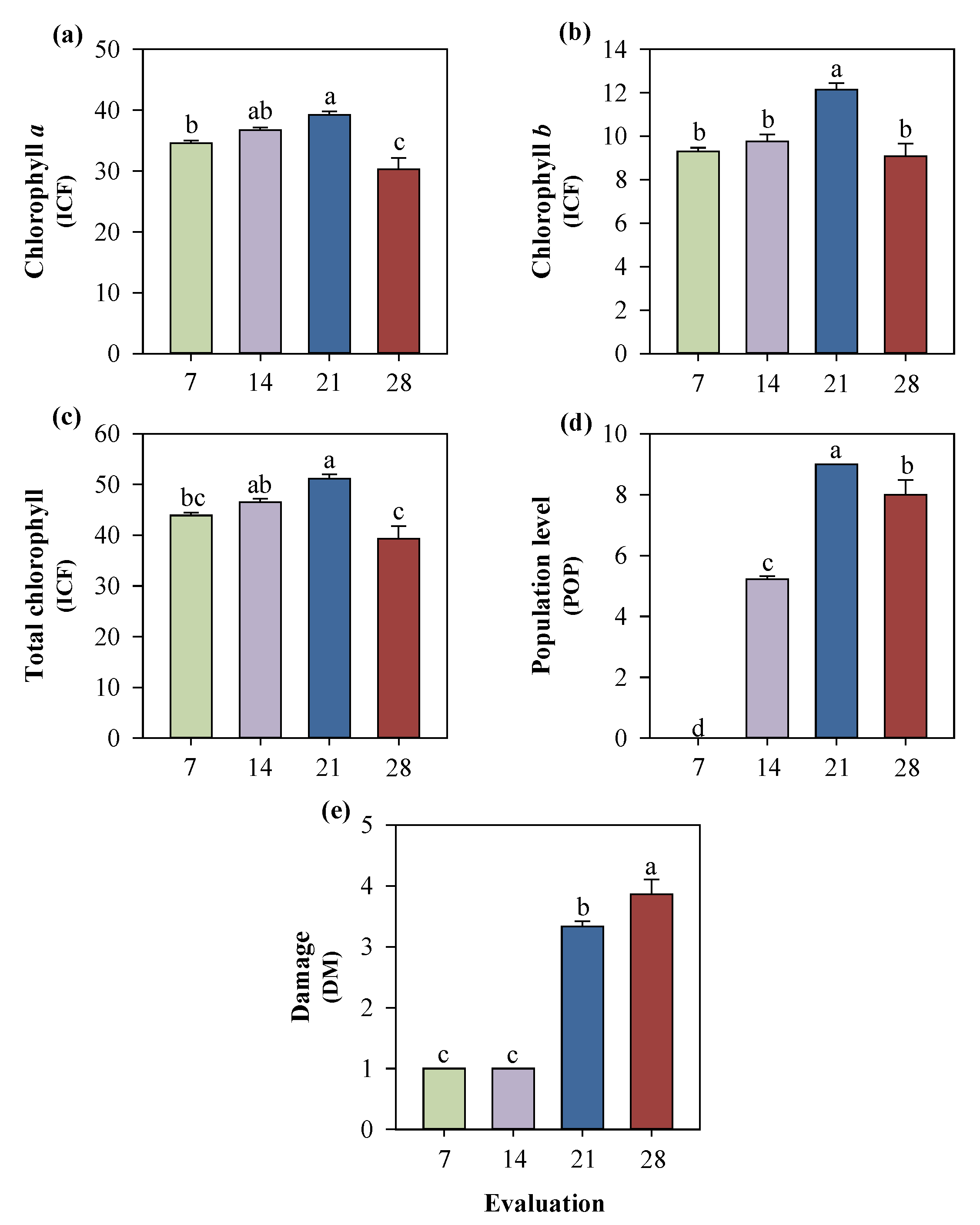
Figure 10.
Chlorophyll a (a), chlorophyll b (b), total chlorophyll (c), aphid population level (d), and plant damage (e) in cowpea plants cultivated with different silicon sources and subjected to black aphid herbivory. Means followed by the same lowercase letters do not differ significantly according to the F-test (p ≤ 0.05). Bars represent the mean ± standard error.
Figure 10.
Chlorophyll a (a), chlorophyll b (b), total chlorophyll (c), aphid population level (d), and plant damage (e) in cowpea plants cultivated with different silicon sources and subjected to black aphid herbivory. Means followed by the same lowercase letters do not differ significantly according to the F-test (p ≤ 0.05). Bars represent the mean ± standard error.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



