
Submitted:
14 January 2026
Posted:
15 January 2026
You are already at the latest version
Abstract
This study provides an integrated morphological, histopathological, ecological, and experimental evaluation of the parasitic copepod Ergasilus celestis infecting the European eel Anguilla anguilla from Lake El-Manzala, Damietta, Egypt. The objectives were to redescribe adult female morphology, assess gill damage caused by infestation, analyze ecological factors affecting parasite prevalence and intensity, and evaluate in vitro antiparasitic activity of selected plant extracts. Ecological assessments involved examining 170 host fishes collected seasonally from October 2023 to September 2024. In vitro bioassays assessed the effects of Curcumin and Myrrh extracts on parasite survival. The results revealed pronounced morphological adaptations in the female parasite related to attachment and reproduction. Infestation induced severe histopathological damage, including epithelial hyperplasia, tissue erosion, and distortion of gill lamellae. Ecological analyses demonstrated a very highly significant seasonal pattern, with peak prevalence and intensity during spring, higher susceptibility in female hosts, and strong correlations between infestation intensity and environmental parameters (P < 0.001). In vitro assays showed a concentration-dependent reduction in parasite survival, with Curcumin exhibiting stronger antiparasitic activity than Myrrh. These findings demonstrate the influence of host biology, environmental factors, and plant extracts on E. celestis infestation, offering insights for effective parasite management and sustainable control in marine ecosystems.
Keywords:
parasitic copepods
; Ergasilus celestis
; ultrastructure
; histopathology
; in vitro
1. Introduction
Parasitic copepods of the family Ergasilidae are among the most widespread and ecologically significant ectoparasites affecting freshwater and brackish water fishes worldwide. Members of the genus Ergasilus have been reported on the gills of a diverse range of fish hosts, where females attach firmly using modified second antennae, often resulting in significant gill tissue damage and compromised respiratory function in infested hosts. Early foundational work outlined the general morphology and pathogenic impacts of ergasilid copepods, noting extensive tissue reactions including hyperplasia and lamellar fusion in infested gills, which reduce the effective respiratory surface area of fish hosts [1,2].
The family Ergasilidae includes approximately 30 genera and 163 species of the fish-parasitic copepod Ergasilus, in which parasitism is restricted to females. The majority of these species inhabit freshwater ecosystems and predominantly infest the gill tissues of their fish hosts [3,4]. Similar infestation dynamics have been recorded from African inland waters, where Ergasilus and other copepod taxa were identified as dominant gill parasites in cichlids and catfish, with infection levels influenced by environmental parameters and host characteristics [5]. Parasitic copepod infections constitute a key factor in the identification and characterization of these parasites, particularly in relation to the histopathological effects that negatively impact fish health. Exposure of fish to alkaline conditions, in combination with additional stressors such as parasitic infestations, triggers substantial physiological strain, leading to structural damage of the gills and disruption of antioxidant capacity, metabolic activity, and ionic homeostasis [2,6].
Beyond prevalence studies, recent advances in taxonomy and systematics have expanded the diversity of Ergasilus recognized from freshwater fishes in the Middle East and African lakes, including the description of new species and integrative taxonomic assessments combining morphological and molecular data [7,8,9,10,11]. In addition, research integrating histopathological, ultrastructural and molecular analyses has provided insights into host-parasite interactions and the physiological consequences of infestations, such as oxidative stress and epithelial damage in gill tissues [12,13,14,15].
Parasitic crustaceans belonging to the family Ergasilidae are recognized as significant pathogens in aquaculture systems, as their attachment to fish gills induces severe epithelial damage, respiratory impairment, physiological stress, and increased vulnerability to secondary infections, ultimately resulting in reduced growth performance and economic losses in cultured fish populations [16,17].
Despite previous contributions, available information on the infestation dynamics of Ergasilus celestis in Anguilla anguilla under Mediterranean–Nile Delta conditions remains limited. Existing studies have provided valuable insights; however, aspects related to seasonal variation and the influence of water quality parameters are still insufficiently explored. Furthermore, few investigations have integrated detailed morphological characterization with histopathological effects, ecological analyses, and in vitro assessments of natural antiparasitic agents within a single host–parasite framework. Addressing these underexplored aspects would substantially advance our understanding of parasite ecology and support the development of sustainable management strategies in marine water and freshwater ecosystems.
2. Martials and Methods
2.1. Study Area and Sampling Period
The present study was conducted on the European eel Anguilla anguilla collected from Lake El-Manzala, Damietta Governorate, Egypt. Fish samples were obtained seasonally over a one-year period from October 2023 to September 2024 to assess temporal variation in parasite prevalence and intensity. Lake El-Manzala (Figure 1) is located in the northeastern part of the Nile Delta, Egypt, approximately (between latitudes 31°15′ - 31°30′ N, and longitudes 32°00′ - 32°30′ E). Lake El-Manzala is a shallow brackish water lake characterized by fluctuating physicochemical conditions, making it a suitable ecosystem for investigating host–parasite–environment interactions. A total of 170 specimens of A. anguilla were randomly collected using local fishing gears. Fish were immediately transported to the laboratory in insulated containers for parasitological examination. The systematic position and nomenclature of the examined fish in the present study are as follows:
Family : Anguillidae.
e.g., : Anguilla anguilla [18]. English name: Common eel
2.2. Parasitological Examination
Each fish was examined externally, with particular attention given to the gills, which were carefully excised and inspected under dissecting microscope for the presence of parasitic copepods. Recovered parasites were gently detached using fine forceps, counted, and preserved in 10% formaldehyde for subsequent morphological analysis. Parasite prevalence and mean intensity were calculated following standard parasitological definitions. The parasitic copepods were identified using [19,20]. Identification, systematic position and nomenclature of the parasites under investigation are as follows:
Phylum: Arthropoda
Subphylum: Crustacea
Class: Maxillopoda
Subclass: Copepoda
Order: Poecilostomatoidea
Family: Ergasilidae [19]
Genus: Ergasilus [19]
Species: Ergasilus celestis [20]
2.3. Morphological Examination
Adult female specimens of Ergasilus celestis were cleared in lactic acid and examined under a compound light microscope. Detailed morphometric measurements were obtained using an ocular micrometer, and drawings were prepared with the aid of a camera lucida. Morphological terminology and identification criteria followed standard taxonomic keys for ergasilid copepods. Measurements are expressed in micrometers (μm) (Figure 2).
2.4. Examination of Parasitic Copepod by SEM
Collected parasitic copepods were fixed in 2.5% glutaraldehyde (pH 7.4), post-fixed in 4% osmium tetroxide, dehydrated in ascending ethanol (30–70%), coated with gold, and examined using a JEOL JSM 6510 LVSEM at Mansoura University, Egypt.
2.5. Histological Investigations
For histopathological assessment, gill arches from both infested and non-infested fish were excised and fixed in 10% neutral buffered formalin for 24–48 h. Fixed tissues were dehydrated through a graded ethanol series, cleared in xylene, and embedded in paraffin wax. Sections of 4–6 μm thickness were prepared using a rotary microtome and stained with hematoxylin and eosin (H&E). Histological sections were examined microscopically to evaluate tissue alterations associated with parasite attachment.
2.6. Ecological and Environmental Assessment
Water quality parameters were measured in situ and in the laboratory following standard procedures. Water temperature (°C), pH, and salinity were recorded using calibrated portable meters. Dissolved oxygen concentrations were determined using the Winkler titrimetric method according to [21].
Dissolved oxygen (DO) concentration was calculated using the following equation:
 where mL titrant represents the volume of sodium thiosulfate solution consumed during titration, B is the reagent blank volume, N denotes the normality of the sodium thiosulfate solution, and the constant 8 corresponds to the equivalent weight of oxygen (mg). The factor 1000 converts the calculated values to mg L−1. mL sample titrated refers to the volume of the water sample used for titration, while A represents the total volume (mL) of manganous sulfate and alkaline iodide reagents added. The correction factor (300−A) / 300 accounts for sample displacement by the added reagents in a standard 300 mL BOD bottle.
where mL titrant represents the volume of sodium thiosulfate solution consumed during titration, B is the reagent blank volume, N denotes the normality of the sodium thiosulfate solution, and the constant 8 corresponds to the equivalent weight of oxygen (mg). The factor 1000 converts the calculated values to mg L−1. mL sample titrated refers to the volume of the water sample used for titration, while A represents the total volume (mL) of manganous sulfate and alkaline iodide reagents added. The correction factor (300−A) / 300 accounts for sample displacement by the added reagents in a standard 300 mL BOD bottle.
These parameters were used to evaluate their relationship with parasite prevalence and mean intensity. Seasonal variation in infection parameters was assessed by grouping samples into four seasons (autumn, winter, spring, and summer). Host sex was determined to evaluate sex-related differences in parasite intensity.
2.7. In Vitro Evaluation of Plant Extracts
In vitro bioassays were conducted to assess the antiparasitic effects of Curcumin (Curcuma longa) and Myrrh (Commiphora myrrha) extracts against adult female E. celestis. Live parasites were exposed to a series of increasing concentrations of each extract in Petri dishes containing filtered water. Parasite survival time was recorded and compared with an untreated control group. Mean life span was calculated for each concentration.
2.8. Statistical Analysis
All statistical analyses were performed using SPSS 26.0 (IBM Corp., USA). Differences in parasite prevalence and mean intensity among seasons, host sexes, and environmental variables were evaluated using appropriate statistical tests, including t-tests and correlation analyses. Results were considered statistically significant at P < 0.05.
Statistical figures and graphical representations were generated using Python 3.0 to ensure accurate and reproducible visualization of data. Results are presented as mean ± standard error (SE)
3. Results
3.1. Female Morphology
The female Ergasilus celestis is characterized by a slender, elongated body that is markedly expanded anteriorly and gradually tapers toward the posterior end. The body length, excluding caudal setae and ovisacs, ranges from 900 to 1020 μm, with a width of 380 to 400 μm (Table 1). The body is divided into the prosome and urosome. The prosome comprises the cephalosome and mesosome, with the first mesosomal somite completely incorporated into the cephalosome, forming an elongated cephalothorax. This cephalothorax represents nearly half the total body length and lacks a distinct boundary with the incorporated somite except for a shallow lateral constriction. Anteriorly, it forms a short rostrum, while posteriorly it is transversely truncated. It measures 500–555 μm in length and 330–340 μm in width, with width decreasing posteriorly (Figure 3, Figure 4, Figure 5 and Figure 6). The mesosome includes three free thoracic somites (second to fourth), which are broader than long and progressively decrease in size toward the posterior. Females bear two prominent ovisacs attached posteriorly; each ovisac is elongated, slightly tapered distally, and measures 1030–1055 μm in length and 135–150 μm in width. Each sac contains approximately 75–85 large, spherical eggs that are clearly visible through the thin membrane. The urosome consists of three abdominal somites, each wider than long and slightly diminishing in width posteriorly. The first abdominal somite is the largest. The third abdominal somite is deeply notched posteriorly, forming two lobes, each bearing a caudal ramus. Each ramus is longer than wide, approximately equal to the combined length of the last two abdominal somites, and armed distally with three terminal setae, with the innermost being the longest (Table 1).
The cephalothorax bears a pair of antennules (1st antenna) and a pair of prehensile antennae (2nd antenna). The antennules are cylindrical, six-segmented, and 95–100 μm long and 30 μm in a maximum width (Table 1). The segments decrease in size distally except for the second, which is the largest; they carry numerous simple setae, with an armature formula of 2–9–3–4–3–5 (Figure 3B). The antennae are long, slender, and subchelate, measuring 345–360 μm in length and 110 μm in a maximum width and consist of four segments. They consist of a short basal coxobasis, a long first endopodal segment bearing a small medial inner spine, a narrow curved second endopodal segment forming a subchela, and a short terminal claw-like segment that is smooth and unarmed.
Five pairs of thoracic swimming legs arise from the first five thoracic somites. Legs 1–4 are biramous, with three-segmented exopods and endopods, except for the fourth leg which has a two-segmented exopod. The exopod consists of two segments while the endopod consists of three segments. Each segment of exopod and endopod is provided with serrate spines and unarmed setae arranged on the distal segmental margin. Armature of rami is shown in (Table 2). These legs display characteristic ornamentation including rasp-like spines, naked setae, and rows of spinules. The fifth leg is highly reduced and distinctly separated from the preceding pair. The urosome also includes the reduced fifth thoracic somite and the genital (sixth thoracic) somite, both clearly separated from the segments anterior to them (Figure 4, Figure 5 and Figure 6).
3.2. Histological Investigations
In normal gills of Anguilla anguilla, each gill arch gives rise to two rows of primary filaments, each supported by a cartilaginous core and bearing a single row of thin-walled secondary lamellae. These secondary lamellae are the primary site of gaseous exchange (Figure 7A). In contrast, infestation by the parasitic copepod Ergasilus celestis produced pronounced histopathological alterations at sites of attachment on the gill lamellae (Figure 7B and Figure 8A & B). Parasite attachment via the modified second antennae caused deep penetration into gill tissue, resulting in chronic epithelial hyperplasia, formation of epitheliocystis-like inclusions, and massive loss of filament tissues (Figure 7B and Figure 8A & B). These changes distorted the normal lamellar architecture and reduced the effective respiratory surface area, indicating impaired gas exchange.
3.3. Ecological Assessments
170 examined Anguilla anguilla fishes were collected during the present investigation from October 2023 until September 2024. 95 of them were infected with parasitic copepods (Ergasilids), Ergasilus celestis. Seasonal variation in the prevalence (%) of the parasitic copepod Ergasilus celestis infecting its host throughout the year. The prevalence showed marked seasonal differences, with the highest infection rate recorded during spring (57%), while markedly lower prevalence was observed in autumn (3%), winter (2%), and summer (1%). Statistical analysis revealed a highly significant effect of season on parasite prevalence (DF = 4, t = 7.53, P < 0.001), indicating strong seasonal influence on the distribution of E. celestis (Table 3 & Figure 9).
The Figure 9 illustrates a pronounced seasonal pattern, with spring showing the highest prevalence (90.5%), whereas markedly lower prevalence rates were recorded in autumn (4.8%), winter (3.2%), and summer (1.6%). Statistical analysis confirmed a very highly significant seasonal variation in parasite prevalence (DF = 4, t = 7.53, P < 0.001), indicating that spring represents the peak season for E. celestis infestation. The mean intensity of infestation exhibited clear seasonal variation (Table 4 & Figure 10), with a pronounced peak during spring (36.90 ± 10.90), whereas markedly lower intensities were recorded in autumn (0.50 ± 0.70), winter (0.80 ± 1.30), and summer (0.60 ± 0.89). Statistical analysis revealed a very highly significant increase in mean intensity during spring (t = −4.90, P < 0.001) and a highly significant difference during summer (t = −3.22, P < 0.001), while no significant seasonal effect was observed in autumn or winter (P > 0.05).
In the (Table 5 and Figure 11), the mean intensity of infestation was higher in females (17.30 ± 1.115) than in males (11.60 ± 1.011). Statistical analysis revealed a very highly significant difference between host sexes (t = 15.57, P < 0.001), indicating that female hosts exhibited a greater susceptibility to E. celestis infestation compared to males.
Statistical analysis in Table 6 revealed very highly significant associations between parasite mean intensity and all examined environmental variables (P < 0.001), indicating a strong influence of water quality parameters on parasite infestation levels.
3.4. In Vitro Evaluation of Curcumin and Myrrh Against Ergasilus celestis
In vitro exposure of the copepod parasite Ergasilus celestis to different concentrations of Curcumin and Myrrh extracts (Table 7 and Figure 12) resulted in a marked reduction in parasite life span compared with the control group (140 min). Curcumin exhibited a stronger concentration-dependent effect, with the shortest mean life span recorded at 15000 ppm (10 min). In contrast, Myrrh extract showed a gradual reduction in survival time, reaching 20 min at the highest concentration (7000 ppm).
Figure 12 showing both plant extracts induced a concentration-dependent reduction in parasite survival compared to the untreated control. Curcumin exhibited a stronger antiparasitic activity, with the highest concentration (15,000 ppm) causing the greatest reduction in mean life span. Myrrh also significantly reduced survival, reaching its maximum effect at 7,000 ppm. Asterisks (**) indicate statistically significant differences compared with the control (P < 0.01).
4. Discussion
The present study provides an integrated analysis of the parasitic copepod Ergasilus celestis infecting Anguilla anguilla, combining morphological, histopathological, ecological, and experimental approaches. Such comprehensive investigations remain limited for ergasilids copepods, particularly under the environmental conditions of the Nile Delta region and Mediterranean Sea. The morphological characteristics of the adult female E. celestis observed in the present study are largely consistent with the original descriptions of ergasilid copepods, particularly regarding body elongation, anterior expansion, and the development of prehensile second antennae adapted for firm attachment to gill tissues [20,22]. Minor variations in body size and appendage measurements compared with earlier reports may reflect host-specific adaptations or environmental influences, as previously suggested for other Ergasilus species [23,24]. The presence of large ovisacs containing numerous eggs highlights the high reproductive potential of this parasite, which may contribute to its persistence and seasonal outbreaks in natural fish populations. The fish gill is a multifunctional organ involved in respiration and key homeostatic processes, including osmoregulation, hormone metabolism, nitrogen excretion, and acid–base balance [25]. Histological examination revealed severe pathological alterations in the gills of infested fish, including epithelial hyperplasia, tissue erosion, and distortion of secondary lamellae. Similar pathological responses have been documented in fish infected with Ergasilus sieboldi and related species, where mechanical damage caused by the modified antennae leads to chronic inflammation and impaired gas exchange [2,26,27]. The marked reduction in effective respiratory surface area observed in the present study suggests that heavy infestations of E. celestis may compromise host fitness and increase susceptibility to secondary infections, particularly under environmental stress. The pronounced seasonal variation recorded in parasite prevalence and mean intensity, with peak infestation during spring, is in agreement with previous studies reporting higher ergasilid activity during warmer periods associated with increased host metabolism and parasite reproduction [28,29]. The significantly higher infestation levels observed in female hosts further support earlier findings that host sex may influence susceptibility to parasitic copepods, possibly due to physiological or hormonal differences [30,31]. Comparisons with recently described Ergasilus species also highlight interesting patterns. For instance, in a study from Lake Tanganyika, [24] described five species (including three new), using both morphological and molecular approaches. They reported that morphological characters such as segmentation of appendages, ornamentation of swimming legs, and shape of caudal rami are crucial for species delineation. The elongated cephalothorax and detailed segmentation in E. celestis, therefore, may represent taxonomically informative traits that help distinguish it from congeners. Ecologically, the well-developed swimming legs and biramous structure in the first four thoracic legs of E. celestis suggest an ability to navigate within the host’s gill environment, or potentially during free-swimming stages, similar to other Ergasilus species. This flexibility could facilitate host-finding or repositioning on the gill filaments, especially under varying flow or host movement. The strong associations between parasite mean intensity and water quality parameters, including temperature, salinity, pH, and dissolved oxygen, emphasize the critical role of environmental conditions in shaping host–parasite interactions. Similar correlations have been reported in African and Asian freshwater systems, where fluctuations in physicochemical parameters directly affected parasite development and transmission dynamics [5,32]. These findings suggest that environmental degradation or climate-driven changes could exacerbate ergasilid infestations in vulnerable fish populations. In agreement with our findings, infection prevalence was higher in females, whereas mean intensity and density were greater in males. Seasonally, both prevalence and intensity of Ergasilus infection peaked in spring, followed by autumn, winter, and summer [33]. These findings [34] are consistent with our study, as we also observed that abiotic factors, including water pH and flow rate, influence parasite abundance and prevalence. Similar to previous reports, higher pH was associated with increased parasite occurrence, while faster water flow reduced prevalence. Moreover, temperature appeared to enhance the occurrence of P. anguillae, aligning with patterns observed in our host population. This suggests that manipulating environmental conditions could be an effective approach to managing parasite infections in aquaculture systems.
The in vitro bioassays demonstrated a concentration-dependent reduction in parasite survival following exposure to Curcumin and Myrrh extracts, with Curcumin exhibiting superior antiparasitic activity. These results are consistent with previous reports highlighting the broad-spectrum antiparasitic and antimicrobial properties of plant-derived compounds [35,36]. The observed efficacy of natural extracts against E. celestis suggests their potential as environmentally friendly alternatives to synthetic chemotherapeutics, particularly in integrated parasite management strategies. Furthermore, future research should prioritize integrating molecular techniques to resolve its phylogenetic placement. Molecular studies on Ergasilus (e.g., 28S rDNA in E. sieboldi) have revealed paraphyly within the genus and emphasized the need for combining morphological and genetic data for accurate taxonomy. Given the detailed morphological traits of E. celestis, a phylogenetic analysis could clarify its relationship to other Ergasilus species and possibly reveal cryptic diversity.
5. Conclusions
The morphological profile of Ergasilus celestis underscores characteristic features of ergasilid copepods, as well as species-specific traits that may reflect its biology and ecology. The high egg production, specialized appendages, and segmented body make it a promising candidate for further taxonomic, ecological, and parasitological studies. Such research would not only clarify its taxonomy but also illuminate its role in host–parasite dynamics and potential impacts on fish health. Overall, the present study underscores the multifactorial nature of E. celestis infestations, driven by parasite biology, host characteristics, and environmental conditions. The integration of morphological, histopathological, ecological, and experimental data provides a robust framework for understanding the dynamics of ergasilid copepods in freshwater ecosystems.
Funding
This research received no external funding.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We are grateful to the scanning and Transmission electron microscope unit at Mansoura University, Egypt.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Kabata, Z. Ecology of fish parasites. Family Ergasilidae. In: The World of Copepods. Zoological Studies 2004, 43, 229–234. [Google Scholar]
- Shereen, A.F.; Shymaa, H. Pathobiological and ultrastructural studies of infested spotted seabass, Dicentrarchus punctatus in Damietta region, Egypt. JESP 2024, 54, 167–174. [Google Scholar] [CrossRef]
- Hadfield, K.A. History of discovery of parasitic Crustacea. In book: Parasitic Crustacea. In Smit, N.J., Bruce, N.L. and Hadfield, K.A. (Eds.). Pp, 7–72. In book: Parasitic Crustacea: State of knowledge and future trends. Zoological Monographs. 2019, 3, Springer: 481pp.
- Walter, T.C.; Boxshall, G. World of Copepods Database. Ergasilus Nordmann, 1832. Accessed through: World Register of Marine Species. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=128641 (accessed on 2024-10-2).
- Bouaye, C.S.A.; Kassi, G.B.; Yedehi, E.A.; Essetchi, P.K. Prevalence of the Parasitic Copepods Ergasilus latus and Lernea cyprinaceain African Catfish (Chrysichthys nigrodigitatus) in Relation to the Water Quality of Adiaké-Aby Lagoon,Côte d’Ivoire. Journal of Advances in Biology & Biotechnology 2025, 28, 1142–1153. [Google Scholar] [CrossRef]
- Endalew, A.; Wei, Y.; Jixiang, H.; Lynda, D.; Siqi, L.; Mohamed, F.; Yalun, D.; Rahma, H.; Mustafa, H.; Jun, Q. Ultrastructural, Osmoregulation, Physiological Metabolism, and Antioxidative Response of Red Tilapia (Oreochromis spp.) Under Acute Alkaline Stress. Fishes 2026, 11, 26. [Google Scholar] [CrossRef]
- Precious, P.; Nico, J.; Liesl Van, As.; Marllese, T.; Kerry, A.H. Integrative Morphological and Genetic Characterisation of the Fish Parasitic Copepod Ergasilus mirabilis Oldewage & van As, 1987: Insights into Host Specificity and Distribution in Southern Africa. Diversity 2023, 15, 965. [Google Scholar] [CrossRef]
- Robert, M.; Iveta, H.; Andrea, S.; Michal, B.; Radek, S.; Jasna, V.; Maria, S. Diversity and phylogeny of parasitic copepods of freshwater fishes from the Mediterranean and the Middle East. Parasitology 2025, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Du Xin, X.; Xu, X.; Chen, Z.; Zhang, Z.; Liu, Z.; Muhammad, S. Molecular Identification of Parasitic Copepods Using Cytochrome c Oxidase Subunit 1 from Chinese Marine Waters. Pakistan Journal of Zoology 2024, 1–7. [Google Scholar]
- Angela, M.; German, A.; Jose Celso, DO. M. Ergasilus tipurus n. sp. (Copepoda: Ergasilidae), A Parasite of Brazilian Amazon Fish Species. Acta Parasitologica 2019. [Google Scholar] [CrossRef]
- Marketa, O.; Maria, Y.; Lukas, V.; Jiri, H. Distribution and host range of a highly invasive parasitic copepod. Hydrobiologia 2025, 852, 2221–2239. [Google Scholar] [CrossRef]
- Basem, A.A.; Thamir, K.A.; Atheer, H.A. A New Ergasilus Nordmann, 1832 Species (Copepoda: Cyclopoida, Ergasilidae) from Gills of Two Freshwater Fishes at Al-Gharraf River, Southern Iraq. EJABF 2024, 28, 633–643. [Google Scholar]
- Robert, M.; Eva, R.; Maria, S. Species of Ergasilus von Nordmann, 1832 (Copepoda: Ergasilidae) from cichlid fishes in Lake Tanganyika. Parasitology 2023, 150, 579–598. [Google Scholar] [CrossRef]
- Shimaa, A.R.; Maha, M.R.; Ghada, E.A.; Eissa, A.E.; Mohamed, A.; Mahmoud, AO. Molecular characterization and phylogenetic analysis of parasitic copepoda; Ergasilus sieboldi isolated from cultured gilthead sea bream (Sparus aurata) in Egypt, associated with analysis of oxidative stress biomarkers. J Parasit Dis. 2022, 46, 1080–1089. [Google Scholar]
- Shereen A. F. (2014) Studies on parasitic copepods of economically important fishes using modern techniques. PhD Thesis, Damietta University, https://www.du.edu.eg/ 25/8/2014.
- Abou-Okada, M.; Rashad, M.; Ali, G.; Abdel-Radi, S.; Hassan, A. Oxidative stress, gene expression and histopathology of cultured gilthead sea bream (Sparus aurata) naturally co-infected with Ergasilus sieboldi and Vibrio alginolyticus. BMC Veterinary Research. 2023, 19, 277. [Google Scholar] [CrossRef]
- Ola, H.; Mohamed, M.; Ahmed, G.; Moaheda, E.; Noha, E.; Walaa, S.; Maha, R.; Ghada, A. The mitigative effect of selenium nanoparticles against aluminium nanoparticle toxicity in Nile Tilapia (Oreochromis niloticus): growth performance, antioxidant status, gene expression, and histopathological changes. Aquaculture International 2025, 33, 443. [Google Scholar] [CrossRef]
- Linnaeus, C. Systema Naturae: The System of Nature through the Three Kingdoms of Nature, according to Classes, Orders, Genera, and Species, with Characters, Differences, Synonyms, and Places, 1758, 10th ed., Vol. 1; Laurentii Salvii: Holmiae, Sweden.
- Von Nordmann, A. Micrographic Contributions to the Natural History of Invertebrate Animals, 1832, Part II. G. Reimer, Berlin.
- Roberts, L.S. Ergasilid copepods parasitic on freshwater fishes. Journal of Parasitology 1970, 56, 1028–1035. [Google Scholar]
- Strickland, J.D.H.; Parsons, T.R. A Practical Hand Book of Sea Water Analysis. Fisheries Res. Board Canada Bull., 1972, 167, 2nd ed., p.310.
- Kabata, Z. Parasitic Copepoda of British Fishes. London. The Ray Society. 1979, No. 152, 468pp.
- Kabata, Z. Parasites and diseases of fish cultured in the tropics. International Development Research Center. 1985, Taylor and Francis Inc. 318 pp.
- Míč, R.; Řehulková, E.; Seifertová, M. Species of Ergasilus von Nordmann, 1832 from African freshwater fishes. Parasitology 2023, 150(7), 579–598. [Google Scholar] [CrossRef] [PubMed]
- Erik, S. Parasites of wild sea bass Dicentrarchus labrax from Norway. Diseases of Aquatic Organisms 2002, 48, 209–212. [Google Scholar]
- Mohammed, E.El-Z.; Ola, A. AS.; Shereen, A.F.; Abdel Aziz, A.K. Evaluation of the effect of larval metamorphosis of some digenetic parasites within the digestive gland tissues of the freshwater ampullariid snails, Lanistes carinatus (Olivier, 1804). Parasitology International 2025, 108(10). [Google Scholar] [CrossRef]
- Piasecki, W.; Boxshall, G.; Panicz, R.; Eljasik, P. New identification traits of Tracheliastes maculatus (Copepoda: Lernaeopodidae), a parasite of bream, Abramis brama (Cyprinidae). International Journal for Parasitology: Parasites and Wildlife 2025, 27, 101108. [Google Scholar]
- Dhanya, P.; Amina, S. Parasitic copepod infestations in commercially exploited fishes from Kayamkulam backwater, Kerala, India. Journal of Parasitic Diseases 2019, 43, 245–252. [Google Scholar] [CrossRef]
- Ali, A.; Adday, T. Description of a new species of Dermoergasilus Ho & Do, 1982 (Copepoda: Ergasilidae) from the redbelly tilapia Coptodon zillii (Gervais) (Perciformes: Cichlidae) in Basrah, southern Iraq. Systematic Parasitology 2019, 96, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Ayman, J.; Andreas, R.; Bouchta, EL-M.; Yanez, MV.; Garcia, MC.; Munoz, M.; Moya, F.; El Arraf, S. Environmental Factors Changes and Its Association with the Catches Fluctuations of Atlantic Chub Mackerel (Scomber colias, Gmelin, 1789) in the South Alboran Sea (Western Mediterranean Sea). EJABF 2024, 28, 1727–1750. [Google Scholar]
- Shereen, A.F.; Salwa, A.; Zeyad, H. Ultrastructure of Lamproglena pulchella (Copepoda: Lernaeidae), a gill parasite on the freshwater fish, Leuciscus vorax, from Tigris River, Iraq. 2019, 23(4), 385–389. [Google Scholar] [CrossRef]
- Johnson, EM.; Andrade, FM; Sanchez, PS.; Hernadez, AG.; Li, C.; David Wells, RJ. Spatiotemporal and Environmental Effects on Demersal Fishes Along the Nearshore Texas Continental Shelf. Fishes 2025, 10, 632. [Google Scholar] [CrossRef]
- Sadguru, P.; Dharmendra, S. Seasonal variations in prevalence and intensity of gill lice, Ergasilus, a crustacean parasite on freshwater fishes of Balrampur, U.P. Int.J.Entomol.Res. 2024, 9, 58–61. [Google Scholar]
- Duane, E.B.; David, K.C. Occurrence of Ergasilus celestis (Copepoda) and Pseudodactylogryrus anguillae (Monogenea) among wild eels (Anguilla rostrata) in relation to stream flow, pH and temperature and recommendations for controlling their transmission among captive eels. Aquaculture 2000, 187, 261–274. [Google Scholar] [CrossRef]
- Ammar, A.; Ahmeda, A.; Abdelkarim, M.; El-Bab, A.; Amer, A.; Abdelnour, S.; El-Nawsany, M.; Mahmoud, A.; Nalel, M. Dietary Curcumin Nanoparticles Improve Growth Performance, Oxidative Status and Immune Response of European Seabass ( Dicentrarchus Labrax ). Annals of Animal Science 2024, 24. [Google Scholar] [CrossRef]
- Eissa, El-S.; Alaidaroos, B.; Jastanlah, S.; Munir, M.; Shafi, M.; Abd El-Aziz, Y.; Bazina, W.; Saadlah, I.; Eissa, M.; Paolucci, M.; Alaryah, F.; Abd El-Hamed, N.; Elgammal, M.; Saddony, S. Dietary Effects of Nano Curcumin on Growth Performances, Body Composition, Blood Parameters and Histopathological Alternation in Red Tilapia (Oreochromis sp.) Challenged with Aspergillus flavus. Fishes. 2023, 8, 208. [Google Scholar] [CrossRef]
Figure 1.
Location map of Lake Manzala showing the sampling site and fish collection at Shatta City, Damietta Governorate, Egypt.
Figure 1.
Location map of Lake Manzala showing the sampling site and fish collection at Shatta City, Damietta Governorate, Egypt.
Figure 2.
Diagram of the thoracic leg of E. celestis showing the dimension of the measurements. L1,first length; L2, second length; W, Total width.
Figure 2.
Diagram of the thoracic leg of E. celestis showing the dimension of the measurements. L1,first length; L2, second length; W, Total width.
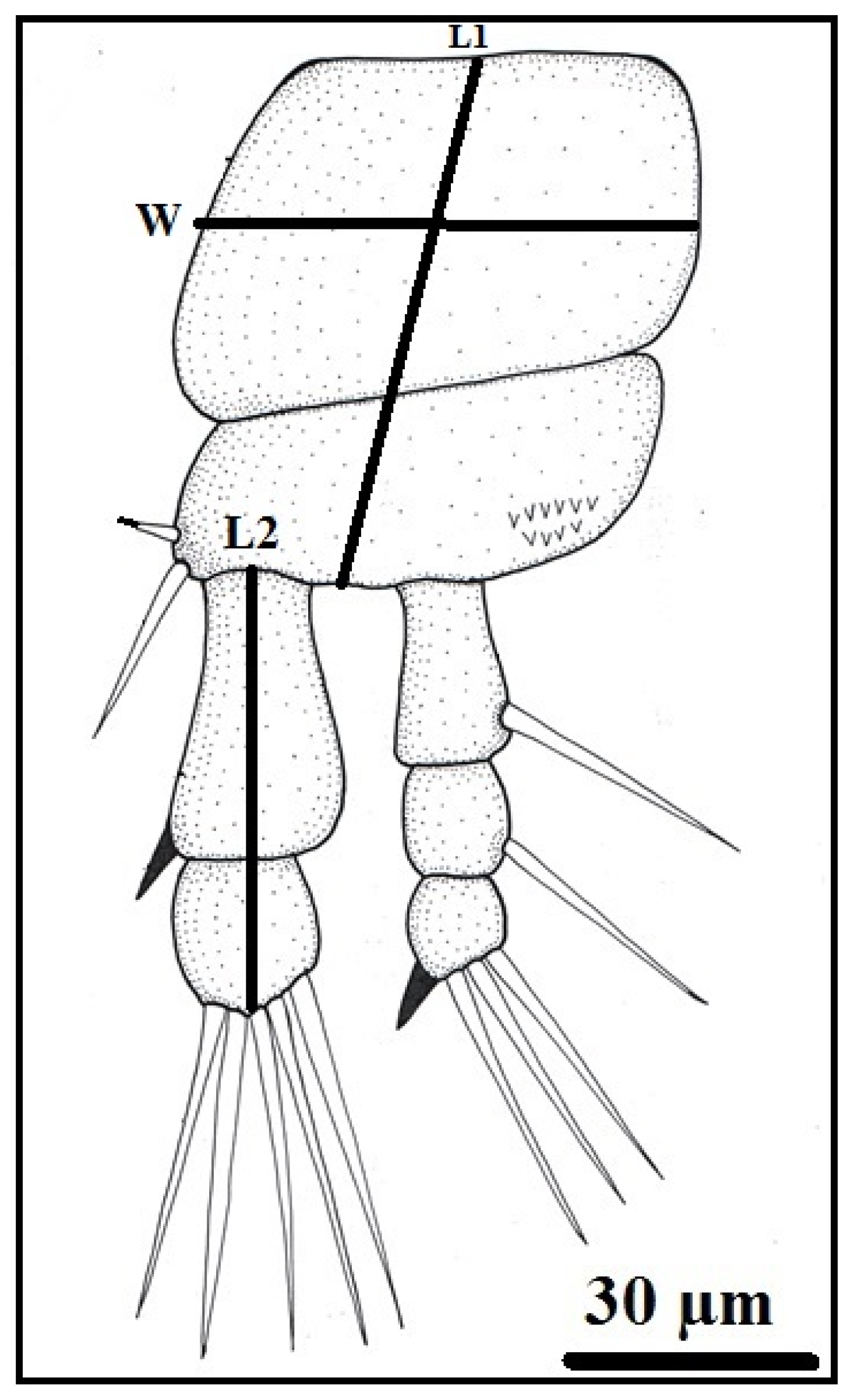
Figure 3.
Ergasilus celestis Roberts, 1970, adult female (A) dorsal body surface, antennule (An), antenna (Ant), cephalothorax (Ce), first thoracic swimming leg (1st Tsl), urosome (Ur), egg sac (Es); (B) the antennule; (C) the antenna. Scale bars in micrometers (µm).
Figure 3.
Ergasilus celestis Roberts, 1970, adult female (A) dorsal body surface, antennule (An), antenna (Ant), cephalothorax (Ce), first thoracic swimming leg (1st Tsl), urosome (Ur), egg sac (Es); (B) the antennule; (C) the antenna. Scale bars in micrometers (µm).
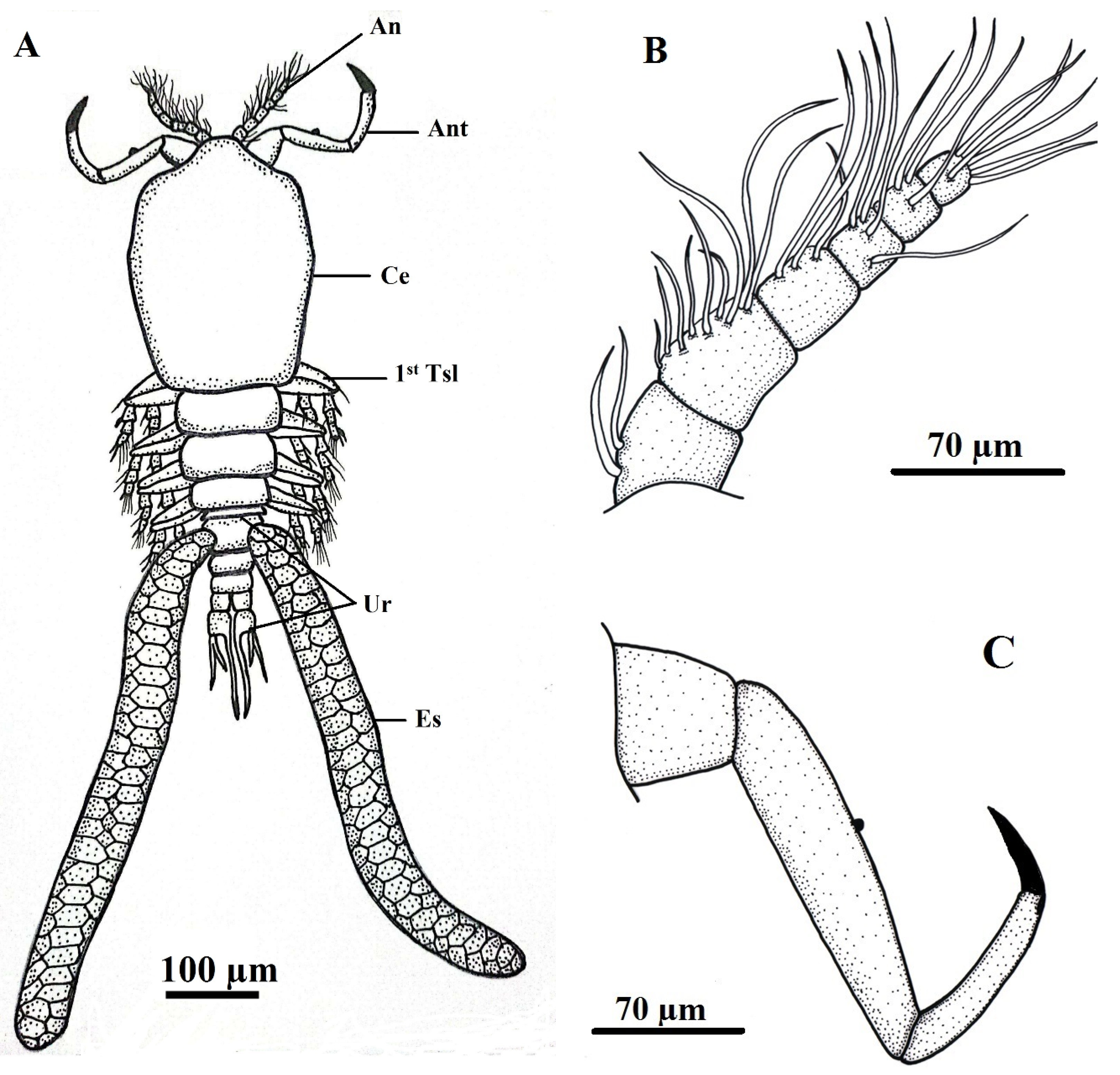
Figure 4.
Ergasilus celestis Roberts, 1970, Thoracic swimming legs of adult female (A) 1st thoracic leg; (B) 2nd & 3rd thoracic leg; (C) 4th thoracic leg; (D) Urosome. Scale bars in micrometers (µm).
Figure 4.
Ergasilus celestis Roberts, 1970, Thoracic swimming legs of adult female (A) 1st thoracic leg; (B) 2nd & 3rd thoracic leg; (C) 4th thoracic leg; (D) Urosome. Scale bars in micrometers (µm).
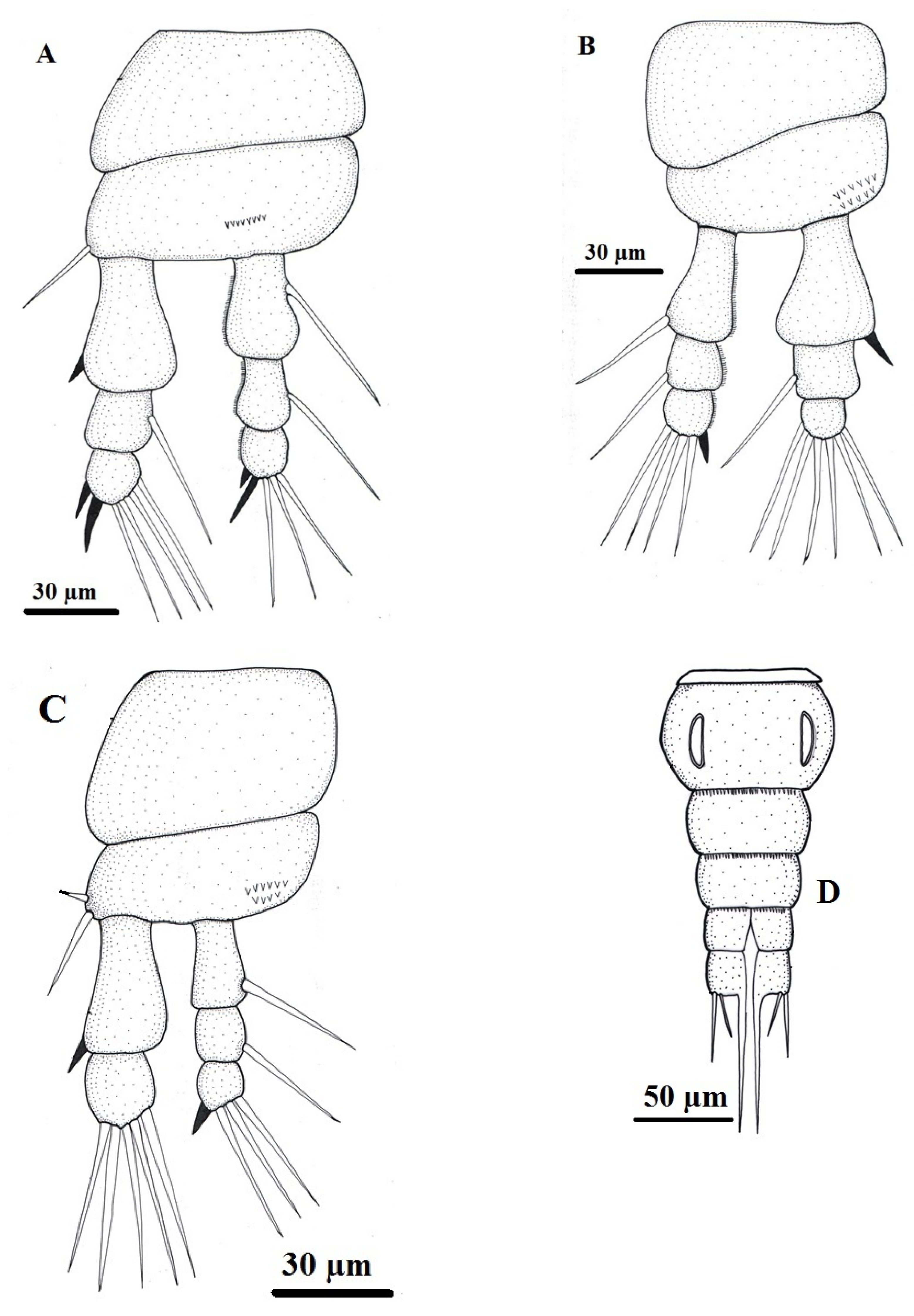
Figure 5.
Light micrographs of the female copepodian parasite, Ergasilus celestis Roberts, 1970 with lactic acid, (A) cephalothorax; (B) posterior body showing 3rd thoracic swimming leg (3rd Tsl), 4th thoracic swimming leg (4th Tsl), egg sac (Es); (C) latero-vental view with the four thoracic swimming legs (Tsls); (D) Inset enlarged of urosome, abdomen (Ab), genital segment (Gs), caudal ramus (Cr). Scale bars in micrometers (µm).
Figure 5.
Light micrographs of the female copepodian parasite, Ergasilus celestis Roberts, 1970 with lactic acid, (A) cephalothorax; (B) posterior body showing 3rd thoracic swimming leg (3rd Tsl), 4th thoracic swimming leg (4th Tsl), egg sac (Es); (C) latero-vental view with the four thoracic swimming legs (Tsls); (D) Inset enlarged of urosome, abdomen (Ab), genital segment (Gs), caudal ramus (Cr). Scale bars in micrometers (µm).
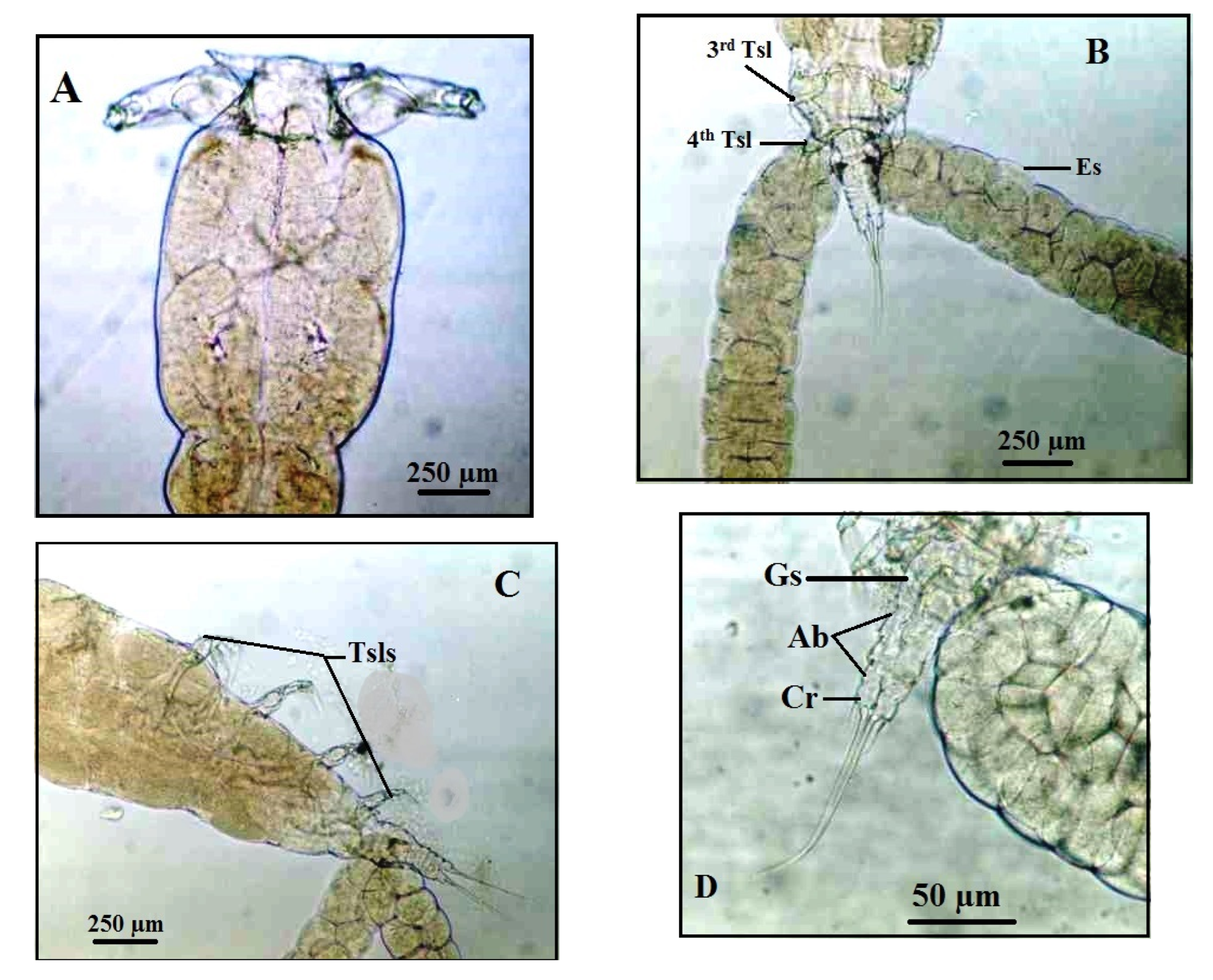
Figure 6.
Scanning electron micrographs of the female copepodian parasite, Ergasilus celestis Roberts, 1970, (A) Female body (lateral view), antenna (Ant), cephalothorax (Ce); the four thoracic swimming legs (Tsls), egg sac (Es); (B) showing urosome (Ur), egg sac (Es), 4th thoracic swimming leg (4th Tsl); (C) Female body (ventral view), antennule (An); (D) Cephalothorax ( ventral view).
Figure 6.
Scanning electron micrographs of the female copepodian parasite, Ergasilus celestis Roberts, 1970, (A) Female body (lateral view), antenna (Ant), cephalothorax (Ce); the four thoracic swimming legs (Tsls), egg sac (Es); (B) showing urosome (Ur), egg sac (Es), 4th thoracic swimming leg (4th Tsl); (C) Female body (ventral view), antennule (An); (D) Cephalothorax ( ventral view).
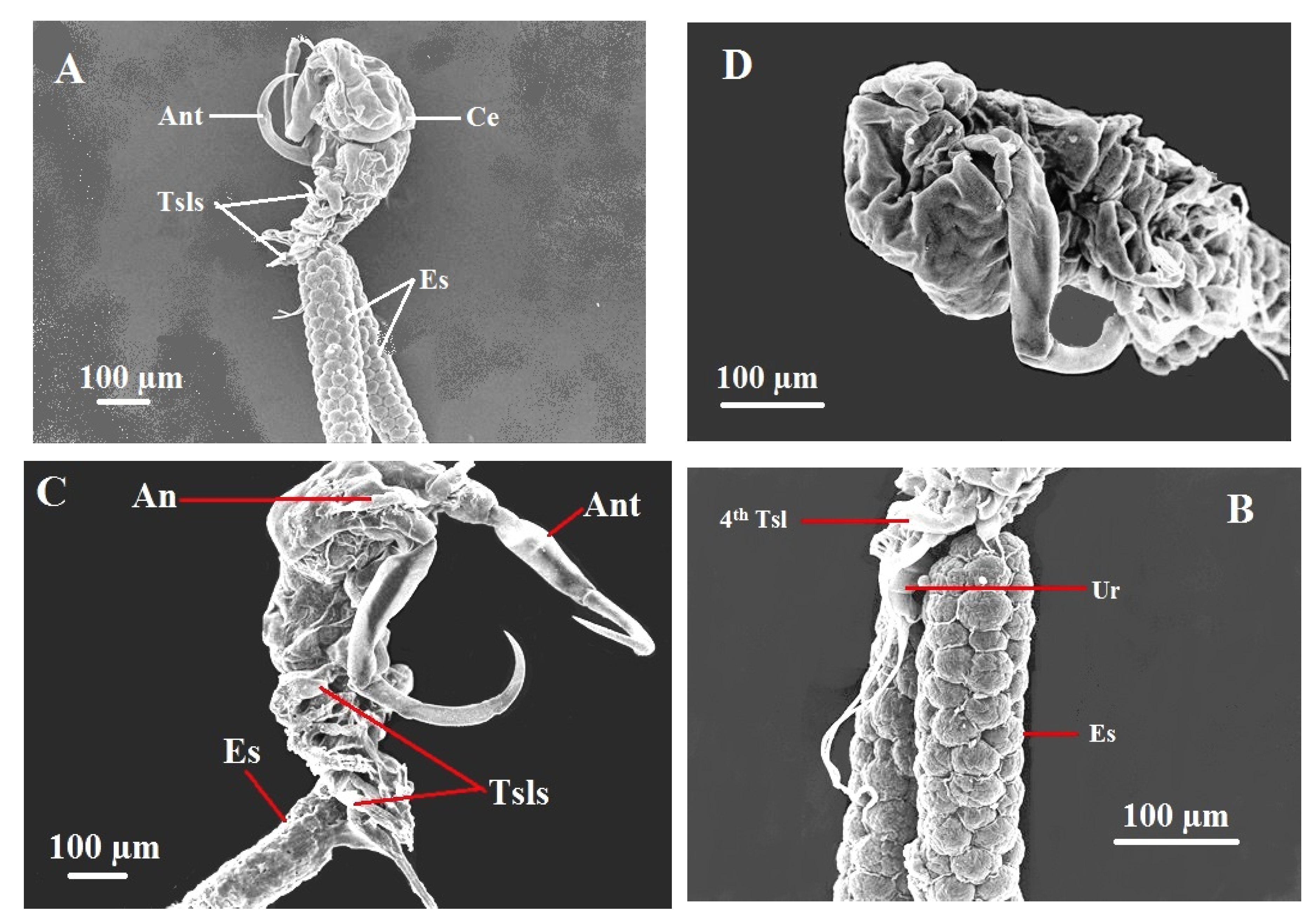
Figure 7.
Light micrographs through the gill filaments of common eel, Anguilla anguilla , (A) The normal gill histology, pgf, primary gill filament and sgl, secondary gill lamellae; (B) The infested gills showing histopathological findings associated with copepod parasite E. celestis (C), necrosis and gill destructions ( arrows), proliferation of gill epithelium and epitheliocystis-likeinclusions (*), Egg sac (Es).
Figure 7.
Light micrographs through the gill filaments of common eel, Anguilla anguilla , (A) The normal gill histology, pgf, primary gill filament and sgl, secondary gill lamellae; (B) The infested gills showing histopathological findings associated with copepod parasite E. celestis (C), necrosis and gill destructions ( arrows), proliferation of gill epithelium and epitheliocystis-likeinclusions (*), Egg sac (Es).
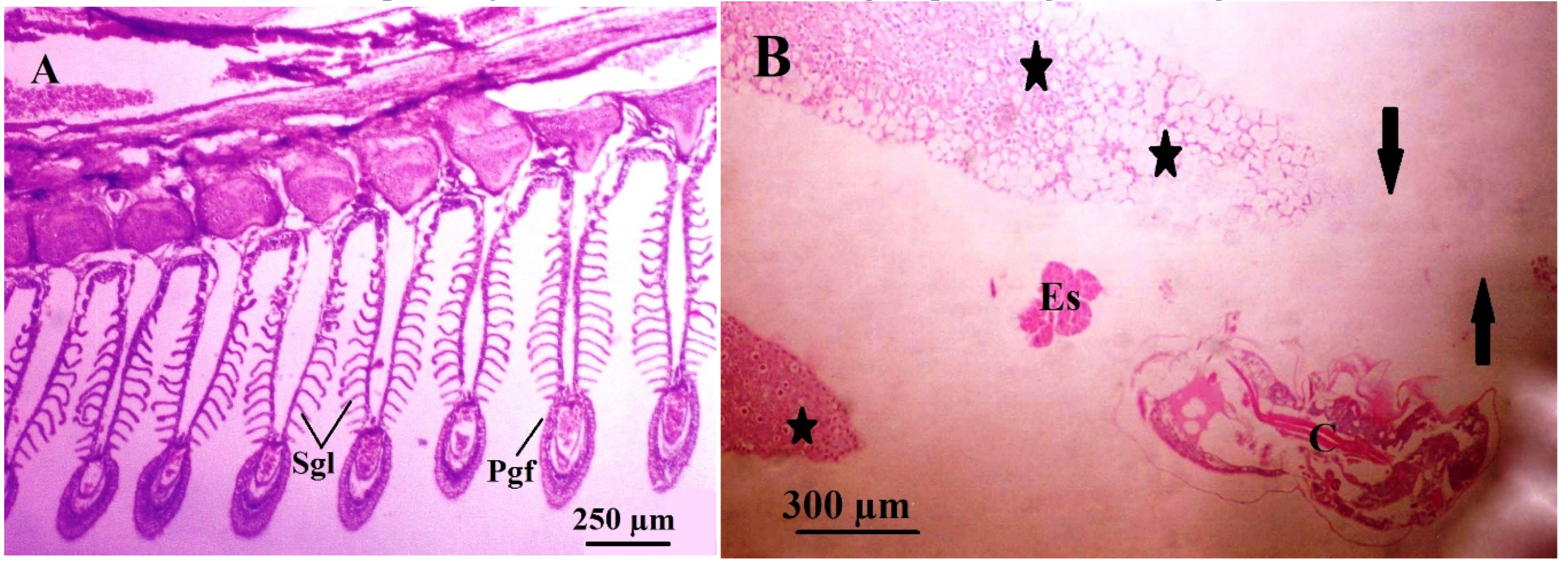
Figure 8.
Scanning electron micrographs (A & B) illustrating pathological impacts of the female copepodian parasite, Ergasilus celestis (C) showing Attachment of antenna (Ant) to grip gill filaments causing erosion and destruction of the gill filaments and lamellae (white and red stars).
Figure 8.
Scanning electron micrographs (A & B) illustrating pathological impacts of the female copepodian parasite, Ergasilus celestis (C) showing Attachment of antenna (Ant) to grip gill filaments causing erosion and destruction of the gill filaments and lamellae (white and red stars).
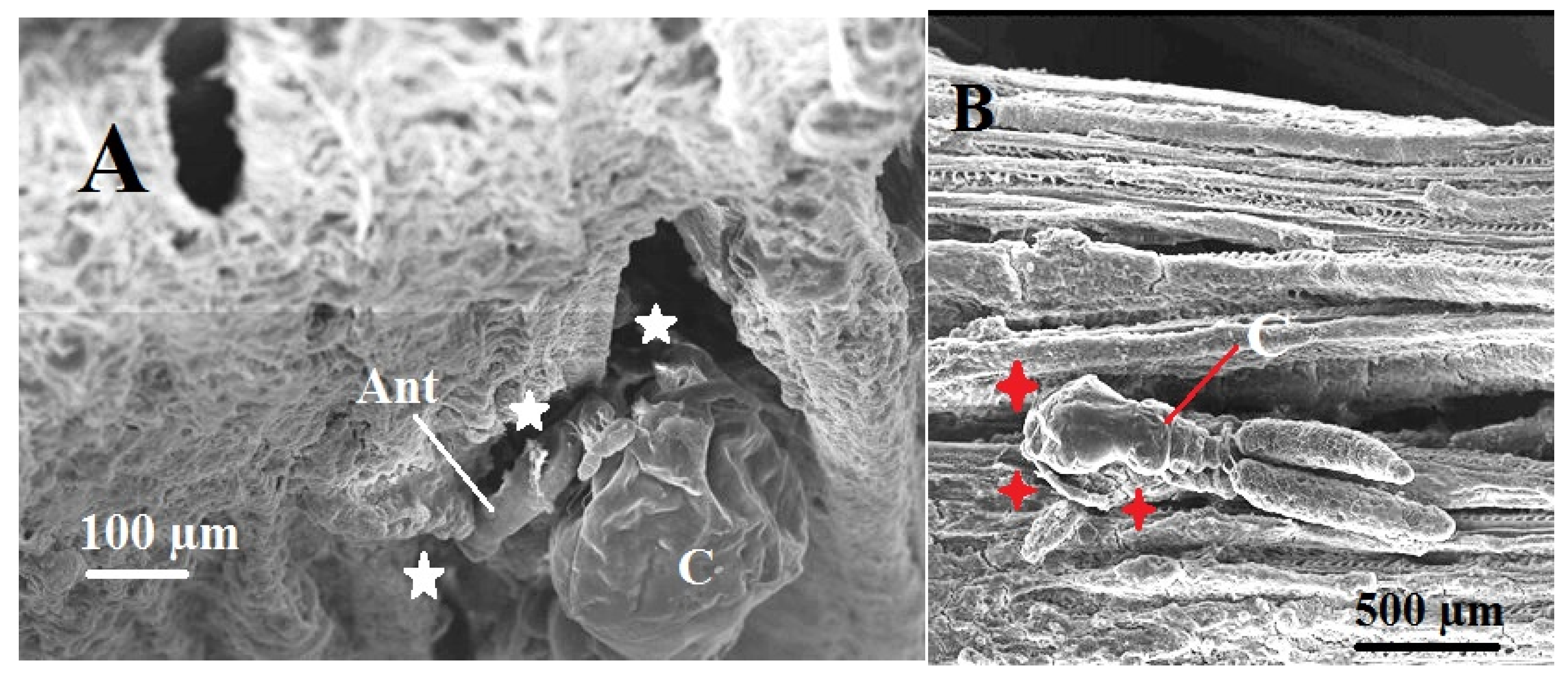
Figure 9.
Seasonal distribution of the prevalence (%) of the parasitic copepod Ergasilus celestis during different seasons of the year.
Figure 9.
Seasonal distribution of the prevalence (%) of the parasitic copepod Ergasilus celestis during different seasons of the year.
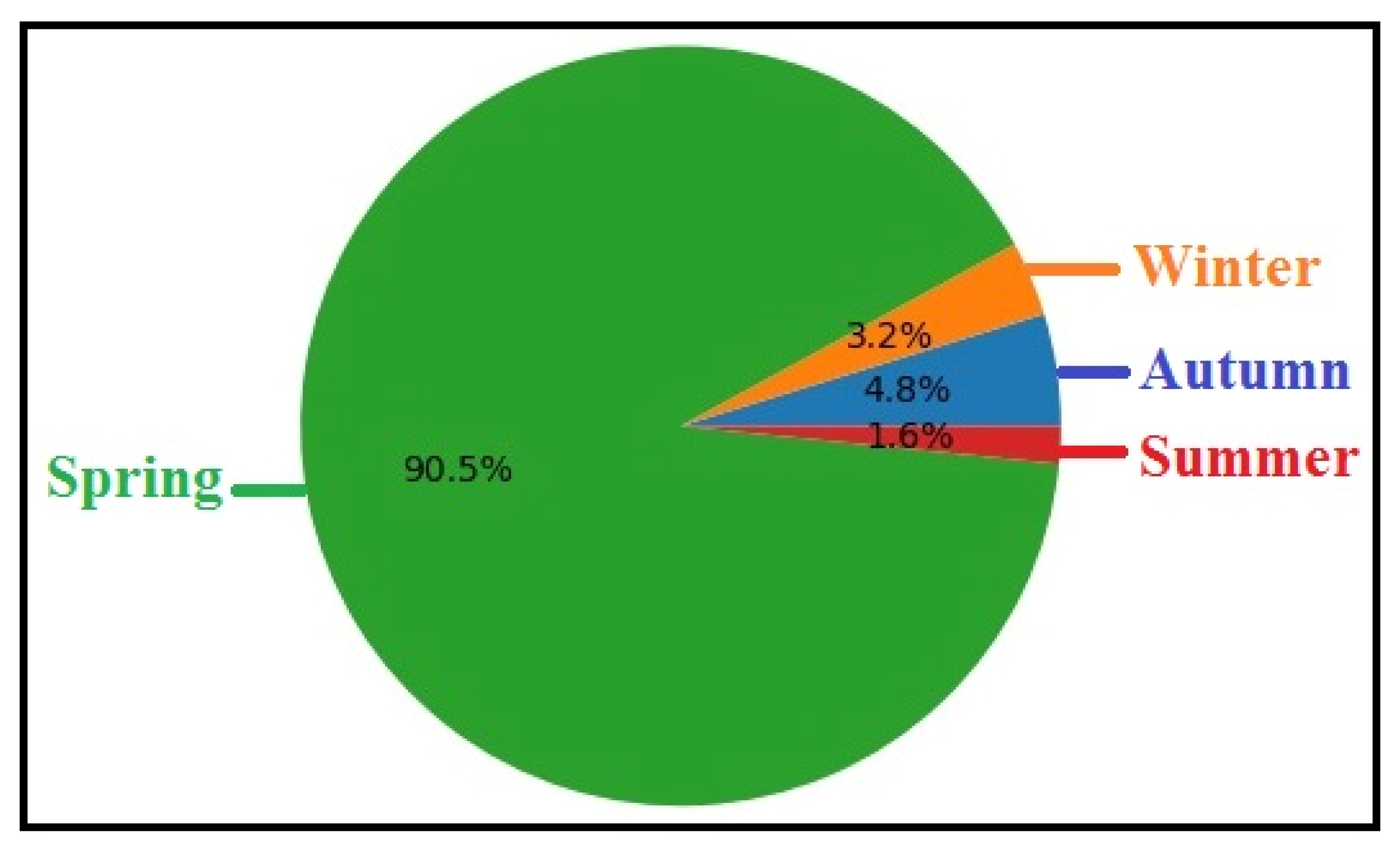
Figure 10.
Seasonal variation in the mean intensity (± SD) of the parasitic copepod Ergasilus celestis recorded from Lake El-Manzala, Damietta. The highest mean intensity was observed during spring, showing a very highly significant difference (***, P ≤ 0.001), whereas autumn and winter exhibited low and non-significant variations.
Figure 10.
Seasonal variation in the mean intensity (± SD) of the parasitic copepod Ergasilus celestis recorded from Lake El-Manzala, Damietta. The highest mean intensity was observed during spring, showing a very highly significant difference (***, P ≤ 0.001), whereas autumn and winter exhibited low and non-significant variations.
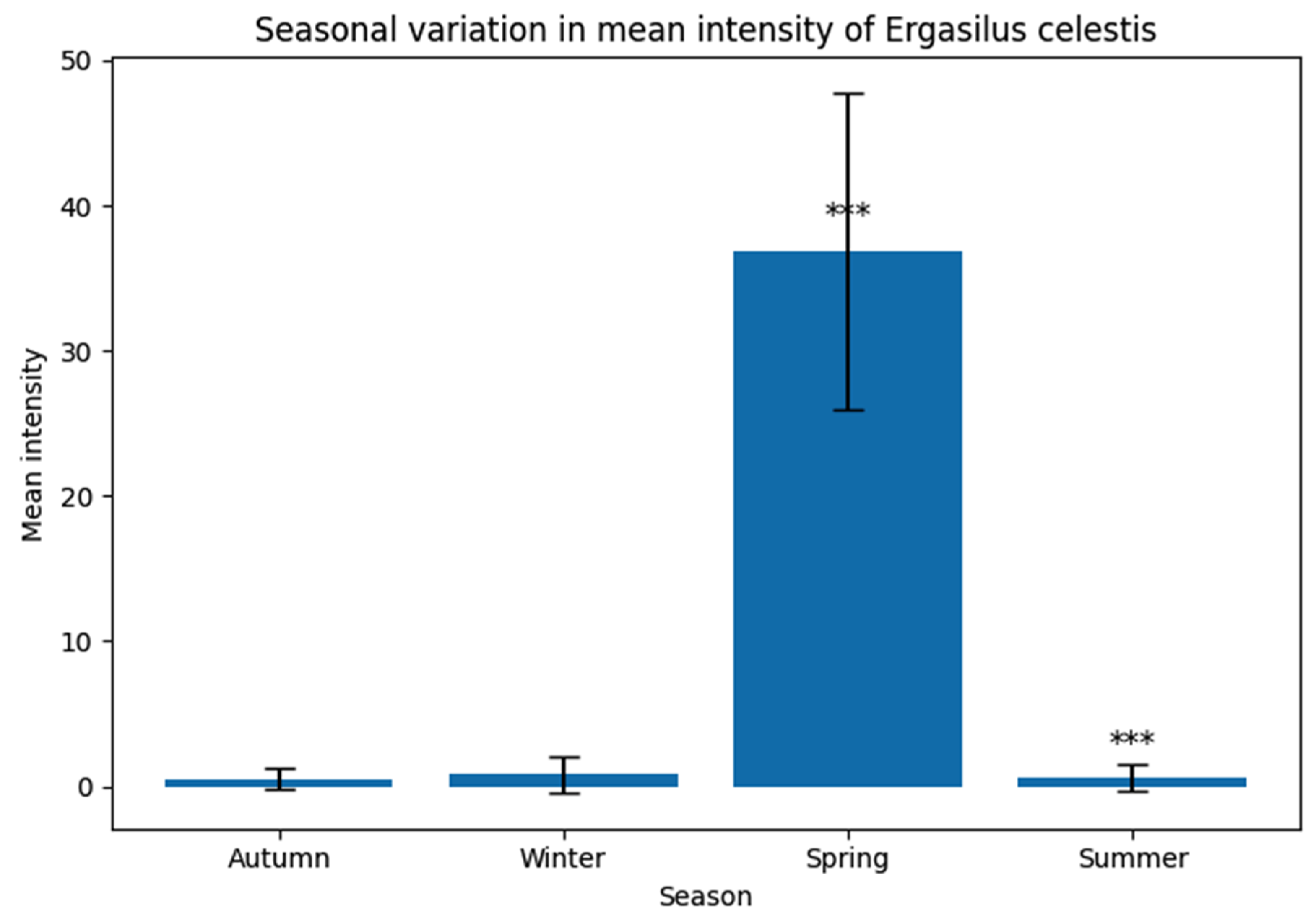
Figure 11.
Comparison of the mean intensity (± SE) of the parasitic copepod Ergasilus celestis infesting female and male hosts (Anguilla anguilla). Females showed a significantly higher mean intensity than males, with a very highly significant difference (***, P ≤ 0.001).
Figure 11.
Comparison of the mean intensity (± SE) of the parasitic copepod Ergasilus celestis infesting female and male hosts (Anguilla anguilla). Females showed a significantly higher mean intensity than males, with a very highly significant difference (***, P ≤ 0.001).
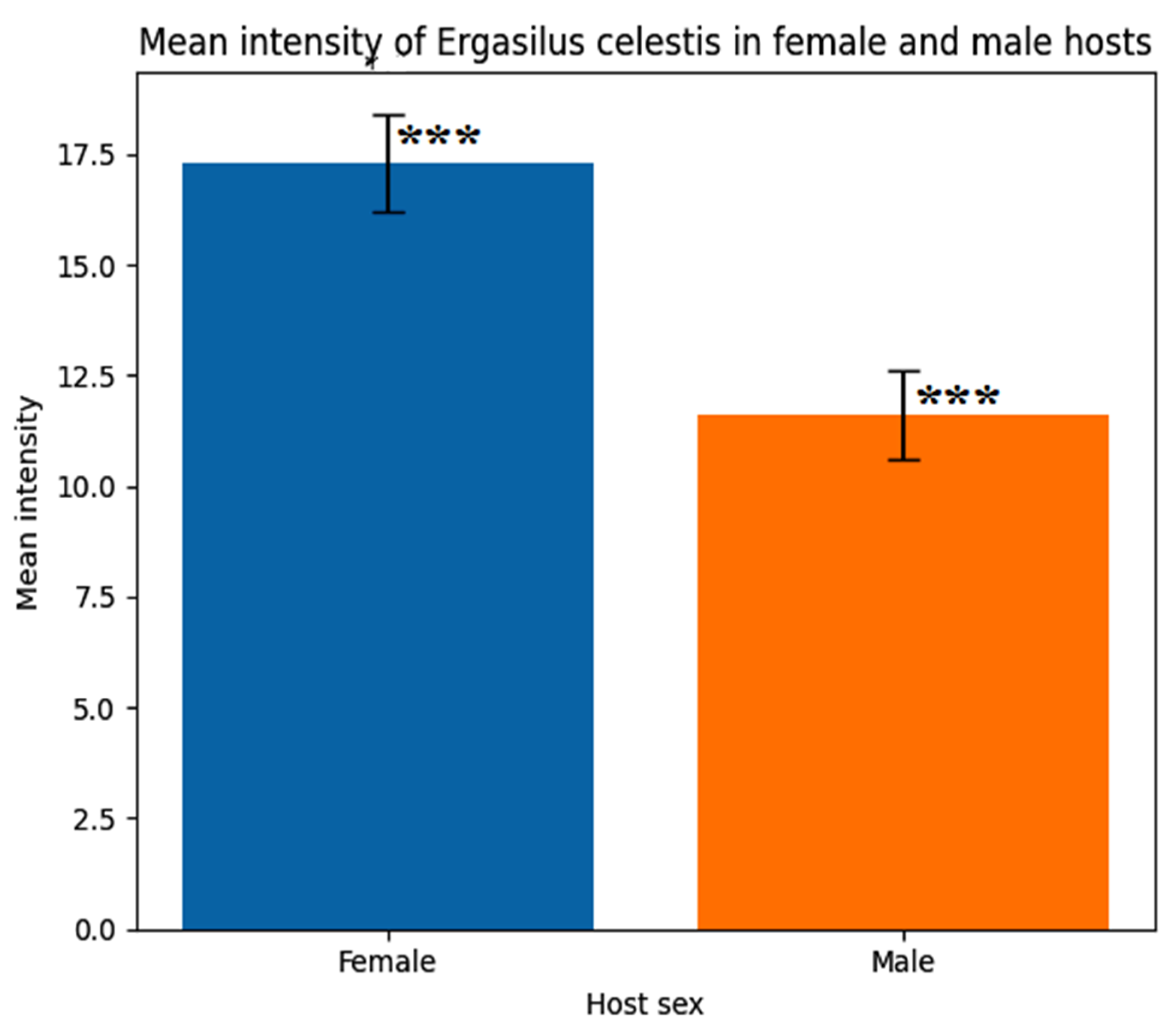
Figure 12.
In vitro effects of different concentrations of curcumin (Curcuma longa) and myrrh (Commiphora molmol) extracts on the mean life span (min) of the parasitic copepod Ergasilus celestis.
Figure 12.
In vitro effects of different concentrations of curcumin (Curcuma longa) and myrrh (Commiphora molmol) extracts on the mean life span (min) of the parasitic copepod Ergasilus celestis.
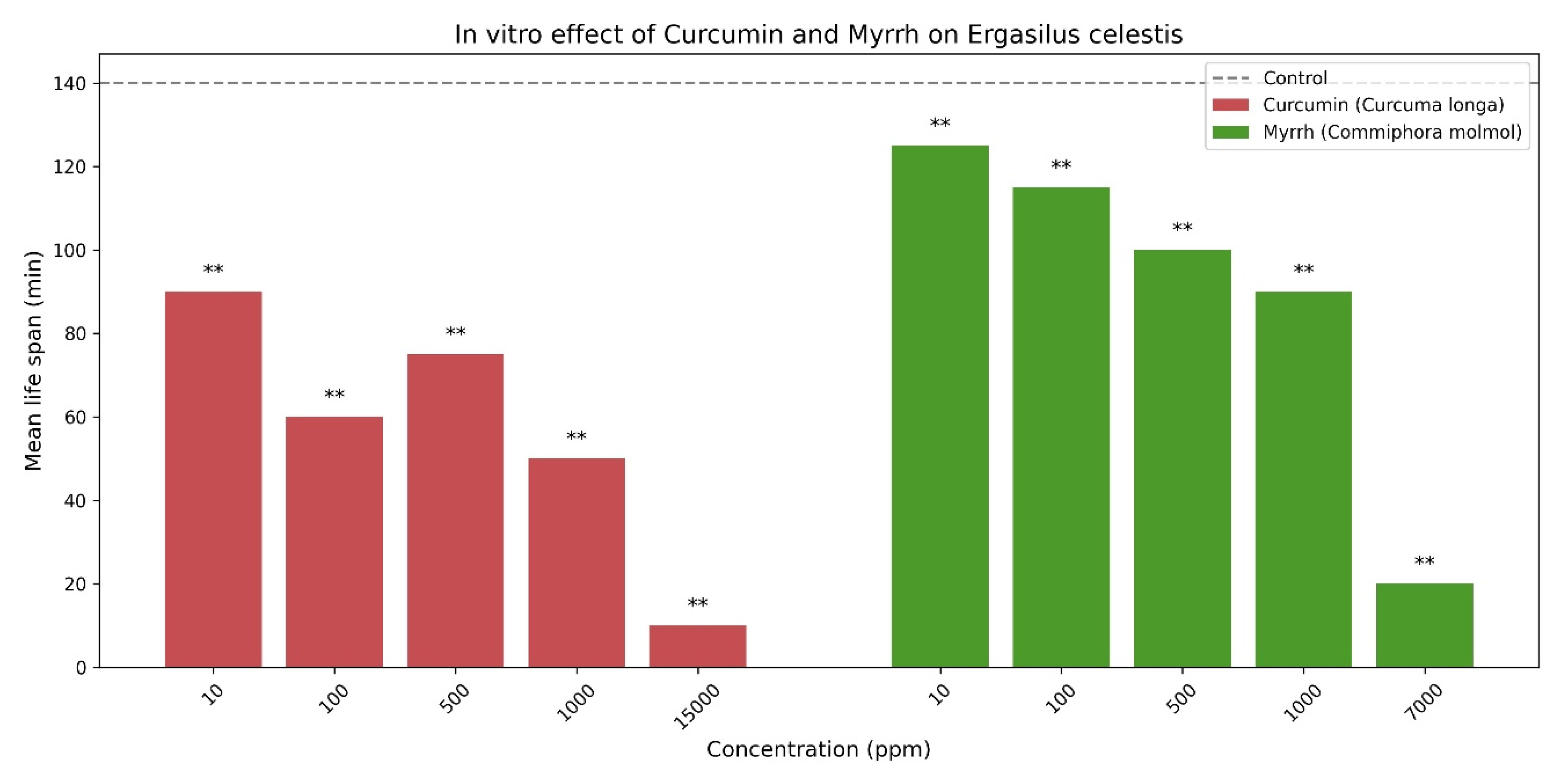

Table 1.
Morphometric measurements of the parasitic copepod, Ergasilus celestis Roberts, 1970.
| Characters | Maximum length | Maximum width |
| Female Body | 1020 | 400 |
| Cephalothorax | 555 | 340 |
| 1st antenna | 100 | 30 |
| 2nd antenna | 360 | 110 |
| 1st thoracic leg (L1) | 55 | - |
| 1st thoracic leg (L2) | 60 | - |
| 1st thoracic leg (W) | - | 75 |
| 2nd & 3rd thoracic leg (L1) | 55 | - |
| 2nd & 3rd thoracic leg (L2) | 60 | - |
| 2nd & 3rd thoracic leg (W) | - | 75 |
| 4th thoracic leg (L1) | 55 | - |
| 4th thoracic leg (L2) | 45 | - |
| 4th thoracic leg (W) | - | 50 |
| 5th thoracic leg | - | - |
| Urosome | 150 | 80 |
| Egg sac | 1055 | 150 |
| Egg abundance | Numerous approx. 75:85 eggs | |
(L1: first length, L2: second length, W: total width, measurements in microns).
Table 2.
The armature formula of the thoracic swimming legs of adult female Ergasilus celestis Roberts, 1970.
Table 2.
The armature formula of the thoracic swimming legs of adult female Ergasilus celestis Roberts, 1970.
| Thoracic legs | Coxa | Basis | Exopod | Endopod | ||||
| 1 | 2 | 3 | 1 | 2 | 3 | |||
| 1st leg | 0-0 | Ι-0 | Ι-0 | 0-1 | ΙΙ-4 | 0-1 | 0-1 | ΙΙ-3 |
| 2nd leg | 0-0 | 0-0 | Ι-0 | 0-1 | 0-6 | 0-1 | 0-1 | Ι-4 |
| 3rd leg | 0-0 | 0-0 | Ι-0 | 0-1 | 0-6 | 0-1 | 0-1 | Ι-4 |
| 4th leg | 0-0 | ΙΙ-0 | Ι-0 | 0-5 | - | 0-1 | 0-1 | Ι-3 |
Roman numerals = Spines Arabic numerals = Setae.
Table 3.
Seasonal prevalence (%) of parasitic copepod Ergasilus celestis at Lake El-Manzala, Damietta.
Table 3.
Seasonal prevalence (%) of parasitic copepod Ergasilus celestis at Lake El-Manzala, Damietta.
| Parasitic copepod | Season | Prevalence (%) | DF | t-value | P-value |
| Ergasilus celestis | Autumn | 3% | 4 | 7.53 | < 0.001*** |
| Winter | 2% | ||||
| Spring | 57% | ||||
| Summer | 1% |
P, probability; DF,Degree of freedom;*, significant; **, highly significant; ***, very highly significant.
Table 4.
Seasonal mean intensity of parasitic copepod Ergasilus celestis at Lake El-Manzala, Damietta.
Table 4.
Seasonal mean intensity of parasitic copepod Ergasilus celestis at Lake El-Manzala, Damietta.
| Parasitic copepod | Season | Mean | ±SD | t-value | P-value |
| Ergasilus celestis | Autumn | 0.500 | 0.70 | -0.23 | 0.804 |
| Winter | 0.800 | 1.30 | -0.9 | 0.279 | |
| Spring | 36.900 | 10.90 | -4.90 | < 0.001*** | |
| Summer | 0.600 | 0.89 | -3.22 | < 0.001*** |
P, probability; ±SD, standard deviation; ⃰ ⃰ ⃰ very highly significant (P < 0.001).
Table 5.
Statistical analysis of the mean intensity of parasitic copepod Ergasilus celestis infesting females and males of fishes.
Table 5.
Statistical analysis of the mean intensity of parasitic copepod Ergasilus celestis infesting females and males of fishes.
| Parasitic copepod | Host name | Host sex | Mean intensity | t-value | ±SE | P-value |
| Ergasilus celestis | Anguilla anguilla | Female | 17.30 | 15.570 | 1.115 | < 0.001*** |
| Male | 11.60 | 11.477 | 1.011 |
P, probability; ±SE, standard error of mean; ⃰ ⃰ ⃰ very highly significant (P < 0.001).
Table 6.
Relationship between mean intensity of Ergasilus celestis and water parameters at Lake El-Manzala, Damietta.
Table 6.
Relationship between mean intensity of Ergasilus celestis and water parameters at Lake El-Manzala, Damietta.
| Water parameters | Mean intensity | ±SE | t-value | P-value | ||
| Copepodian parasite site | Temperature | 25.23 | 3.61 | 0.420 | -55.502 | < 0.001*** |
| pH | 7.69 | 0.359 | -16.40 | < 0.001*** | ||
| Salinity | 24.23 | 5.587 | -36.615 | < 0.001*** | ||
| Oxygen content | 6.20 | 0.407 | -10.965 | < 0.001*** | ||
P, probability; ±SE, standard error of mean; *** very highly significant (P < 0.001).
Table 7.
In vitro effect of Curcumin and Myrrh extracts on the life span of Ergasilus celestis.
| Plant extractions | Concentration (ppm) | Mean life span (min) | Reduction vs control (%) |
| Control | 0 | 140 | - |
| Curcumin, Curcuma longa | 10 | 90 | 35.7 |
| 100 | 60 | 57.1 | |
| 500 | 75 | 46.4 | |
| 1000 | 50 | 64.3 | |
| 15000 | 10 | 92.9 | |
| Myrrh, Commiphora molmol | 10 | 125 | 10.7 |
| 100 | 115 | 17.9 | |
| 500 | 100 | 28.6 | |
| 1000 | 90 | 35.7 | |
| 7000 | 20 | 85.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



