Submitted:
14 January 2026
Posted:
15 January 2026
You are already at the latest version
Abstract
The global health crisis driven by Antimicrobial Resistance (AMR) necessitates an urgent pivot toward novel therapeutic agents, with traditional medicinal plants serving as a critical resource. The Asteraceae genus Verbesina, particularly utilized in Mexican ethnobotany, has garnered scientific attention due to its potent bioactive profile against infection and inflammation. This review provides a comprehensive and critical synthesis of the pharmacological landscape of Verbesina species, focusing specifically on the dual role of its major secondary metabolites, the Sesquiterpene Lactones (SLs), as both cytotoxic and antimicrobial agents. We systematically compile and analyze reported in vitro data, including IC50 values from cancer and non-cancerous cell lines, and MIC values against clinically relevant drug-resistant strains like S. aureus and E. coli. A core focus is placed on establishing the therapeutic index (SI = IC50/MIC) for lead compounds, providing a crucial indicator of drug feasibility. Furthermore, we review the proposed molecular mechanisms of SL action, such as the crucial role of the α-methylene-γ-lactone moiety in alkylating cellular targets, which underpins both their antiproliferative and bactericidal effects. By critically bridging ethnopharmacology with modern mechanistic data, this review validates the translational potential of Verbesina metabolites and highlights clear directions for bioassay-guided isolation and optimization as next-generation anti-resistance scaffolds.
Keywords:
Verbesina
; sesquiterpene lactones
; cytotoxicity
; antimicrobial resistance
; MTT assay
; therapeutic index
; eudesmanes
; ethnobotany
; Asteraceae
1. Introduction
The rapid and relentless emergence of multidrug-resistant (MDR) microbial pathogens represents one of the most pressing and complex global health threats of the 21st century [1]. The World Health Organization (WHO) has categorized Antimicrobial Resistance (AMR) as a critical priority, forecasting a dire scenario where common infections and minor injuries could once again become life-threatening [2,3]. This crisis is exacerbated by the overuse of existing antibiotics and a severe slowdown in the discovery pipeline for new therapeutic agents with novel mechanisms of action [4]. Consequently, the scientific community faces an imperative to explore nature's vast chemical library for new scaffolds capable of overcoming established bacterial defense mechanisms. Historically, natural products have been the undisputed bedrock of modern pharmacology, with approximately 60% of current antibacterial and anticancer agents derived directly from natural sources or semi-synthetic modifications thereof [5]. These biologically evolved molecules possess unparalleled structural complexity, allowing them to interact with complex biological targets in unique ways.
Among the plant families recognized for their phytochemical richness and pharmacological significance, the Asteraceae (formerly Compositae) stands out [6]. This family is the largest in Mexico, a nation designated as "Megadiverse," which houses a high concentration of unique chemical diversity and is considered an "evolutionarily active zone" dueating to an accelerated diversification rate within the Asteroideae subfamily [7,8]. The chemical diversity within Asteraceae is staggering, yielding a wide array of specialized metabolites, including polyacetylenes, flavonoids, alkaloids, and notably, various classes of terpenoids [6,9,10].
Within this prolific family, the Sesquiterpene Lactones (SLs) form a pivotal class of secondary metabolites, characterized by an isoprenoid structure containing 15 carbon atoms and a lactone function [11]. These SLs are structurally classified into several major skeletal groups, including germacranolides, guaianolides, and eudesmanolides [10]. The SLs are of paramount interest to medicinal chemists due to the consistent presence of a highly reactive functional group: the α-methylene-γ-lactone moiety [11]. This structural feature is widely regarded as the primary pharmacophore responsible for their profound biological activities. The unsaturated lactone ring acts as a non-specific alkylating agent, undergoing a Michael-type addition reaction with biological nucleophiles, particularly the sulfhydryl (-SH) groups found in cysteine residues of proteins and peptides like glutathione [11]. This alkylation mechanism provides a clear, unifying hypothesis for their therapeutic potential across both microbial and proliferative targets.
The genus Verbesina itself encompasses over 300 species, with the greatest diversity concentrated in Mexico and the southwestern USA [12,13]. Phylogenetic analysis confirms Verbesina as a distinct lineage within the Heliantheae alliance [7,14], reinforcing its status as a reliable repository of potent specialized metabolites. This chemical potency is not accidental; the genus’s bioactivity is fundamentally linked to its ecological role as an aggressive chemical warrior [15,16]. Species like V. encelioides are notorious invasive weeds globally [17], capable of quickly dominating ecosystems due to potent allelopathic properties, which involve the competitive release of phytotoxic chemicals [15,18,19]. This high ecological competence and survival, even in contaminated environments, underscore the necessity and effectiveness of its underlying chemical arsenal [7,13].
Phytochemical investigation has consistently identified characteristic metabolites: the dominant SLs are primarily of the eudesmanolide type, a scaffold that accounts for almost half of all natural eudesmanolides found in the Asteraceae family [12,15]. These compounds are often isolated as cinnamoyloxy eudesmane conjugates, a defining chemical signature of the genus. Historically, the initial isolation and structural elucidation of these cinnamoyloxy esters from species like V. virginica established this complex scaffold as the functional heart of Verbesina's chemistry [20].
The genus's therapeutic relevance is deeply rooted in Mexican ethnobotany, where various species are integrated into local practices for treating ailments related to inflammation, gastrointestinal distress, and topical infections [21]. For instance, V. crocata is traditionally used for wound healing and antidiabetic activity [22,23]. Beyond anti-infective use, the therapeutic scope is rapidly expanding, with recent studies validating the plant’s dual function as a valuable resource for sustainable agriculture (acting as potent plant biostimulants that promote crop yield [5,13,18]) and neuroprotection (showing efficacy against Alzheimer's disease models [19]).
However, the powerful, non-selective chemical reactivity inherent in the α-methylene-γ-lactone group, the very mechanism responsible for efficacy presents a critical translational challenge: the inevitable duality of efficacy and toxicity [11]. The same alkylating mechanism that deactivates bacterial enzymes can interfere with vital homeostatic processes in non-target, healthy mammalian cells [24,25,26]. This potential non-selective toxicity is vividly underscored by the genus’s history, as many species contain highly toxic compounds like the guanidine alkaloid, galegine, which is fatal to livestock [8,27].
Therefore, a purely potent finding in an antimicrobial assay (low Minimum Inhibitory Concentration, MIC) or a cytotoxicity assay (low IC50) is insufficient. Success depends entirely on the compound’s ability to preferentially target pathogenic or diseased cells over healthy ones. This necessitates a critical quantitative comparison of these two parameters by calculating the Selectivity Index (SI = IC50/MIC) [28]. This metric is essential for advancing natural product research toward clinical application, providing the necessary assurance that potent compounds are also safe.
Despite compelling ethnobotanical and preliminary pharmacological data, a critical gap exists in the synthesized literature: the absence of a focused, comparative analysis that quantifies the SI for its constituent Sesquiterpene Lactones. This review aims to address this gap by providing a comprehensive, mechanistic, and quantitative analysis of the Verbesina genus's potential as a source of anti-resistance therapeutics. We will critically evaluate the literature regarding the cytotoxicity (IC50) and antimicrobial efficacy (MIC) of its specialized metabolites, synthesizing all available in vitro data to calculate and interpret the Therapeutic Index for key scaffolds. By focusing on the structural relationships, dual biological activities, and selective toxicity profiles, this review will delineate the most promising scaffolds for further chemical development, ultimately validating the translational potential of this rich Mexican medicinal resource in the fight against AMR (Figure 1).
2. Ethnopharmacology and Phytochemical Diversity of the Verbesina Genus
The rational investigation of the Verbesina genus, which remains the largest genus within the Heliantheae tribe, is firmly anchored in its definitive chemotaxonomic and phylogenetic placement. Molecular studies confirm that Verbesina belongs to the subtribe Verbesininae, a distinct and ancient lineage nestled within the core Heliantheae clade [2]. This confirmed taxonomic position refines the focus for all chemical investigation, supporting the established consistency of the genus's potent eudesmane SLs as strong chemotaxonomic markers [16]. This structural link is so vital that contemporary research has focused on assembling the complete chloroplast genome (plastome) for V. alternifolia, providing a robust genomic anchor necessary for targeted bioassay-guided isolation and standardization across genetically diverse populations [16].
2.1. Ethnopharmacology and Ecological Context
2.1.1. Chemotaxonomy and Global Distribution
The foundational relevance of the Verbesina genus is established through its chemotaxonomic and phylogenetic placement [2]. Molecular data firmly place Verbesina within a distinct clade, often recognized as the revised subtribe Verbesininae [2,29], nestled in the larger Heliantheae alliance clade, which accounts for approximately 26% of all species in the Asteraceae family [9]. This refined circumscription, separating Verbesina from other common allies, acts as a crucial DNA-based chemotaxonomic anchor that directs and refines future bioassay-guided isolation work [2]. Geographically, the genus is highly prolific, encompassing over 300 species with the greatest diversity concentrated in Mexico and the southwestern USA [9]. The widespread and robust distribution of Verbesina across central Mexican flora, including regions of high diversity like the Bajío, underscores its pharmacological importance [29]. Furthermore, Mexico is positioned as an "evolutionarily active zone" for this lineage, characterized by high speciation rates within the Asteroideae subfamily, which accelerates the diversification of the genus’s potent specialized metabolites [9]. The specific niche preference, such as V. virginica strictly inhabiting areas under the canopy, suggests that the plant's metabolic investment in synthesizing a robust chemical arsenal, including SLs, necessitates a sheltered microclimate to maximize chemical production [30]. The extensive geographical distribution and success are also supported by studies that investigate its cellular mechanics, including cytological analysis of V. encelioides, which has shown genetic variability often linked to the plant's aggressive ecological adaptation and the synthesis of secondary metabolites [31].
2.1.2. Mexican Ethnobotany: The Rationale for Anti-AMR
The pharmacological investigation of Verbesina is directly guided by its established history in Mexican traditional medicine [22], which involves a broader cultural reliance on potent, locally sourced medicinal flora throughout the nation [32]. For instance, V. persicifolia, or 'Huichim,' is a popular folk remedy confirmed for anti-diabetic activity [33] and is used for ailments including inflammatory, gastrointestinal, and anti-infective complaints like wounds that resist healing [34,35]. Similarly, the endemic V. crocata is traditionally applied topically for wounds and burns [22], and V. sphaerocephala is locally applied for treating gangrene and venous ulcers [36]. Scientific reports have successfully confirmed the efficacy for three traditional uses: wound healing, antidiabetic, and diuretic activity [23]. Most recently, V. persicifolia has been validated for applications related to the central nervous system, such as anticonvulsant activity [37], broadening its therapeutic scope beyond anti-infective use. The traditional use of these compounds for infections and inflammation provides the essential ethnopharmacological rationale for investigating its modern anti-AMR properties [35,38]. Furthermore, the anti-infective potential extends beyond the Mexican context, with species like the South American V. macrophylla used traditionally for bacterial and fungal infections of the urinary and respiratory tracts [39].
2.1.3. Ecological Resilience and Chemical Defense
The plant’s inherent bioactivity is fundamentally linked to its ecological success as an aggressive chemical warrior [40,41]. The high ecological competence of species like V. encelioides is evidenced by its recognition as a weed [42], and its aggressive colonization ability is enhanced by strong allelopathic properties [40]. The aggressive success is intrinsically linked to its ability to adapt to diverse and often disturbed habitats, with species like V. alternifolia surviving in fire-affected systems, highlighting the environmental pressures that drive the constant synthesis and selection of potent specialized metabolites [43]. This chemical defense system includes not only defense against herbivory but also compounds vital for pollinator attraction [43]. The plant's overall resilience is high, yet its physical defenses are limited against certain atmospheric stressors like ambient ozone, underscoring the vital role of specialized secondary metabolites for survival [4]. Crucially, the potent chemical arsenal of the genus is physically manufactured and housed in specialized defensive structures common across its tribe [44]. Anatomical studies confirm the ubiquitous presence of Linear Glandular Trichomes (LGTs) on the plant surfaces that are metabolically active and primarily accumulate terpenoids and other specialized metabolites [44]. This rich chemical arsenal extends beyond SLs to include a robust mixture of triterpenoids and phytosterols like β-Amyrin and Lupeol [45], which may possess anti-diabetes and antidyslipidemic properties, suggesting the non-SL fraction provides synergistic or mitigating activities important for the overall therapeutic profile [45]. Finally, the selection of Verbesina for drug discovery is justified by its high degree of genetic and ecological resilience, confirmed by molecular studies showing a significant genetic distance from cultivated Asteraceae and the capacity for producing diverse and potent specialized metabolites to survive in competitive environments [46].
2.2. The Core Chemical Landscape: Sesquiterpene Lactones
2.2.1. Sesquiterpene Lactones: Structure, Chemotaxonomy, and Stereochemistry
The chemical identity of the Verbesina genus is structurally defined by the Sesquiterpenoid class, which are overwhelmingly the most abundant specialized metabolites, constituting approximately 79% of all Terpenoids reported in the genus [21]. This rich biosynthetic output is concentrated around the bicyclic C15 eudesmane skeleton [15], a scaffold recognized for its wide range of biological activities, including anti-tumor, antifungal, and antibacterial properties (Figure 2) [10,47]. The historical foundation began with seminal research on the roots of V. virginica in 1961, which reported the isolation of α- and β-Verbesinol derivatives [48]. These compounds possessed the C15 sesquiterpene framework with a cis-decalin (eudesmane) ring system. Crucially, this study determined that the compounds existed naturally as isomeric sesquiterpene esters which, upon degradation, yielded p-hydroxycinnamic acid [48]. This established the existence of the defining cinnamoyloxy eudesmane scaffold, a structure that links a lipophilic terpenoid body to a phenolic acid (the cinnamoyl moiety) confirming the characteristic chemical structure that governs the genus’s potent bioactivity [48,49,50]. This overwhelming structural prevalence means that eudesmane cinnamates alone account for over a quarter (26.5%) of all reported Verbesina compounds, firmly validating the structural focus of this review [21]. The consistency of this chemical signature is reinforced by systematic investigations of South American Verbesina species (including V. glabrata, V. macrophylla, and V. luetzelburgii), which confirmed consistent chemical dominance with all species affording eudesmane or germacrane derivatives, nearly all bearing a cinnamate or related ester residue [51]. Further supporting this claim, a phytochemical investigation into the aerial parts of V. turbacensis (a species of ethnobotanical importance) successfully isolated four eudesmane derivatives (three of which were novel), further reinforcing the ubiquitous nature of this C15 bicyclic scaffold [52].
The early chemical investigation of the genus was often challenged by the complexity and high oxygenation of these terpenoids. For instance, the structural elucidation of Rupestrol from V. rupestris (Jamaica) detailed a pentahydroxy sesquiterpene of the eudesmane type, featuring hydroxyl groups at C-4, C-6, and C-14, alongside secondary hydroxyls at C-1 and C-9. Chemical reactivity tests, such as the inertness of Rupestrol cinnamate to periodic acid, were required to definitively eliminate C-3 as a site for a secondary alcohol, supporting the locations at C-1 and C-9 [53]. Further complexity was revealed in V. rupestris through the isolation of enantio-eudesmane sesquiterpenes, mirror-image isomers of the common eudesmane structure including Rupestrinol and γ-Chaenocephalol, both isolated as their cinnamate esters. The isolation of these eudesmane sesquiterpenes provided the necessary chemotaxonomic evidence to support the botanical reclassification of Chaenocephalus rupestris to Verbesina rupestris [54].

Table 1.
Structural Diversity of Key Sesquiterpenoids (SLs and Core Analogues) from the Verbesina Genus.
Table 1.
Structural Diversity of Key Sesquiterpenoids (SLs and Core Analogues) from the Verbesina Genus.
| Compound Name (Skeleton) | Structural Descriptor | Source Species | Biological Role |
|---|---|---|---|
| Verbesindiol (Eudesmane - Diol) |
Parent core; 4R,5S,6R,7S,10R configuration; Structural standard. |
V. virginica, V. eggersii |
Potent against prostate cancer cells. |
| CDE (Eudesmane Cinnamate) | 4β-cinnamoyloxy, 1β,3α-dihydroxyeudesm-7,8-ene; Alkylating pharmacophore. | V. persicifolia | Mild mitochondrial uncoupler. Patented for obesity/diabetes/cancer treatment. |
| Arbusculin derivative (Eudesmane Lactone) |
(1R,4S,4aS,7R)-1-methyl-7-(prop-1-en-2-yl)octahydro-2H-1,4a-(epoxymethano)naphthalen-4-yl acetate. | V. lanata | Against the protozoan L. major |
| Rupestrol (Pentahydroxy Eudesmane) |
Highly oxygenated enantio-eudesmane with C-9 oxygenation; isolated as cinnamate esters. | V. rupestris | Confirms complex stereochemistry/chemotaxonomic links. |
| Rupestrinol (Enantio-Eudesmane) |
Epimeric sesquiterpene isolated as cinnamate esters. | V. rupestris | Part of the complex enantio-eudesmane structural series. |
| Eudesmane Esters (Eudesmane Cinnamate) |
Phenolic acid (cinnamate/coumarate) conjugated eudesmanes. | V. virginica | Upholds the chemical signature of SLs conjugated with phenolic acids. |
| SL Acetylated Derivatives (Eudesmane/ Germacrane) | Compounds with acetate substituents. |
V. persicifolia | Acetylation shifts cytotoxicity mechanism to Complex II inhibition (Mitochondrial). |
| Germacrene-Cinnamate | 6β-cinnamoyloxy-1β-hydroxy-10α-methoxy-3-oxo-germacra-4,5Z-ene. | V. negrensis | Antibacterial against S. aureus. |
| Zempoaline C (Elemanolide) |
Elemanolide skeleton with fused α,β-unsaturated-γ-lactone moiety. | V. seattonii | Deploys alkylating pharmacophore via diverse skeletal type. |
| Verocephol (Amorphane Lactol) |
Unique γ-lactol; first reported amorphane skeleton. | V. sphaerocephala | Marks high biosynthetic diversity (C15 precursors). |
| Cadinenes (Sesquiterpenoid) |
New Cadinene-type SLs confirmed by X-ray. | V. sphaerocephala | Structural diversity increases selective lead probability. |
| Parthenin Analog (SL) | SLs induce cytotoxicity via Michael addition to -SH groups. | (General SL mechanism) | α-methylene-γ-lactone is the alkylating pharmacophore. |
| V. macrophylla EO (Sesquiterpenes) | Essential Oil dominated by C15 Sesquiterpenes (Germacrene D). | V. macrophylla | Antimicrobial/anti-inflammatory action with very low host toxicity |
| Anthelmintic Action (Sesquiterpenes / Phenolics) | Essential oil shows 100% adulticidal activity against Habronema muscae. | V. alternifolia | Causes degenerative changes to parasite morphology (SEM confirmed). |
Establishing a reliable Structure-Activity Relationship (SAR) requires an unequivocal definition of the molecule’s C15 scaffold, particularly its stereochemistry [55]. This was achieved through a series of meticulous structural confirmations. The p-coumaryl ester of the new sesquiterpenediol, Verbesindiol, was isolated from the herbaceous parts of V. virginica, and alkaline hydrolysis confirmed it was an Eudesmane derivative with hydroxyl groups located at the tertiary C-4 position and a second at C-6. The relative and absolute configuration was definitively established (4R, 5S, 6R, 7S, 10R) using 1H-NMR, 13C-NMR, and Circular Dichroism (CD) spectroscopy [20]. This structural work culminated in the X-ray crystallographic analysis of the parent diol, which provided the absolute configuration and definitively established the Verbesindiol core as a 4α,6β-dihydroxyeudesmane with a cis-fused decalin ring system. This structure, characterized by the specific orientation of the hydroxyls at C-4 and C-6, serves as the absolute standard for correlating cinnamoyloxy eudesmane scaffolds with their observed biological activities [56]. A detailed reinvestigation of V. eggersii consistently confirmed the ubiquity of Verbesindiol derivatives across the genus, noting that this species is rich in cinnamoyloxy eudesmane SLs and yielded further derivatives, including 15-hydroxy derivatives, a rearranged eudesmane acid, and a benzofuran derivative, highlighting the propensity of the Asteraceae to produce structurally diverse scaffolds that originate from the same basic terpenoid precursors [57].
The necessity for definitive characterization is further demonstrated by studies on Mexican endemic species. The aerial part of V. virgata, for example, afforded two new C-4 cinnamates, including 4β-cinnamoyloxy-1β,2α-dihydroxyeudesm-7-en, whose structure and stereochemistry were unambiguously determined by X-ray diffraction analysis. This analysis confirmed the cis-ring junction and the equatorial orientation of the C-1 hydroxyl group. The study also confirmed the presence of vicinal hydroxyl groups through periodate cleavage, demonstrating that these natural molecules possess chemical functionality capable of undergoing specific reactions with biological targets [58]. The ubiquitous presence of these Eudesmane cinnamates is further confirmed by their isolation from V. sordescens, which afforded two new eudesmane cinnamates alongside a typical Verbesina blend of terpenoids and polyacetylenes [24]. The ubiquitous nature of the eudesmane skeleton in the genus continues to be proven, as an investigation into V. virginica yielded two new cinnamoyl-esterified eudesmane compounds, both featuring the core eudesmane ring system modified by substitution with hydroxyl groups at C-1 and C-4. Crucially, one compound was esterified with p-coumaric acid and the other with cinnamic acid, reinforcing the defining chemical characteristic of the genus: SLs conjugated with phenolic acids [59]. The ongoing investigation continues to define the absolute configuration of key eudesmane derivatives from species like V. turbacensis using single-crystal X-ray crystallographic analysis, reinforcing the reliance on advanced structural data to reliably map a compound's efficacy to its three-dimensional shape [60].
The importance of stereochemical precision for reliable SAR is highlighted by findings regarding the eudesmanetriol mono-cinnamate isolated from V. persicifolia. This compound, identified as 4,10-dimethyl-7-isopropyl-trans-decalin-1,3,4-triol-7-ene-4-trans cinnamate, revealed a correction to the configuration of its C-4 asymmetric center compared to previous assignments in other Verbesina species, confirming the trans-decalin eudesmane backbone in certain highly active lead molecules [61]. A core example of the characteristic scaffold is 4β-cinnamoyloxy, 1β,3α-dihydroxyeudesm-7,8-ene, isolated from the hexane extract of V. persicifolia [34]. This eudesmane derivative is pivotal because its core scaffold, as shown through studies on the closely related CDE, can exert profound biological effects not solely through direct toxicity but through the modulation of fundamental cellular processes. For instance, the eudesmane skeleton has been shown to act as a natural mild uncoupler in liver mitochondria [62]. This critical evidence underscores the necessity of rigorously quantifying the IC50 (cytotoxicity) and MIC (efficacy) of these highly substituted eudesmane derivatives to assess their feasibility as drugs with a safe Therapeutic Index.
2.2.2. Sesquiterpenoid and Monoterpenoid Diversity
While the cinnamoyloxy eudesmane scaffold is the recognized chemotaxonomic marker, the Verbesina genus is, at its core, a dedicated biosynthetic factory for C15 molecules, exhibiting remarkable structural diversity that extends across multiple sesquiterpenoid skeletal types [63]. This complexity ensures that the core bioactive mechanism, the α-methylene-γ-lactone pharmacophore is deployed via numerous molecular architectures [10,64,65]. The biosynthesis of these diverse SLs often begins with the germacrane skeleton, which is widely considered the common precursor to the complex array of eudesmane derivatives [50,51]. Evidence for this pathway includes the co-occurrence of germacrane derivatives alongside eudesmanes across many species [24,51]. A novel, active SL derivative, 6β-cinnamoyloxy-1β-hydroxy-10α-metoxy-3-oxo-germacra-4,5Z-ene, was structurally characterized following bioassay-guided isolation from the chloroformic extract of V. turbacensis [38].
This structural variability is not limited to bicyclic and macrocyclic scaffolds. Investigations into V. occidentalis revealed the presence of six new sesquiterpenes structurally elucidated as derivatives of muurolene, alongside Verboccidentafuran, a novel trisubstituted furan derivative (C15H20O) [66]. Furthermore, the genus’s reliance on multiple SL scaffolds includes the Elemanolides, such as Zempoalines C and D, isolated from V. seattonii, whose highly characteristic elemane skeleton with a fused α,β-unsaturated-γ-lactone moiety was structurally established via X-ray crystallography [64]. Most recently, a comprehensive study on V. sphaerocephala successfully isolated six cadinene-type SLs, five of which were previously undescribed. The accurate structures of these compounds were elucidated by X-ray analyses, highlighting that the genus biosynthesizes multiple SL scaffolds from common C15 precursors, increasing the probability of finding a lead molecule with a distinct and selective mechanism of action [63]. The genus also biosynthesizes the unique amorphane sesquiterpene γ-lactol, Verocephol, from V. sphaerocephala, marking the first reported occurrence of a γ-lactol with an amorphane skeleton [67].
Beyond the non-volatile SLs, the Essential Oil (EO) represents a volatile, lipophilic line of chemical defense [23] and is overwhelmingly dominated by sesquiterpenes (99.2% total) [68,69]. The EO of V. macrophylla, for example, is heavily dominated by the characteristic C15 sesquiterpenes Germacrene D (37.3%) and its derivative, Germacrene D-4-ol (17.0%) [23,69]. Similarly, the EO of V. negrensis is dominated by α-pinene (43.1%), a known antibacterial monoterpene. This structural partitioning of anti-infective activity into volatile like α-pinene and non-volatile SLs fractions is strategic: the mechanism of action for such monoterpenes typically involves the destruction of the cellular integrity of the microorganism, inhibiting respiration and ion transport, which offers an alternative pharmacological pathway to the Michael addition mechanism characteristic of SLs [70]. The consistent presence of these sesquiterpenes reinforces the centrality of the terpene pathway to the plant's chemical strategy [69].
This diverse arsenal is synthesized and housed within specialized plant structures. Anatomical analysis of V. macrophylla revealed that lipids, terpenes, and alkaloids are physically manufactured and stored in specialized leaf secretory ducts via an elaborate, energy-intensive mechanism known as granulocrine secretion. This elaborate and conserved production mechanism underscores the fundamental, evolved importance of these secondary metabolites for the plant's defense system [69]. The complexity extends to C10 monoterpenoids, evidenced by the isolation of the C10 alcohol isoepicampherenol and bornyl esters (e.g., (-)-bornyl ferulate and (-)-bornyl p-coumarate) from V. rupestris [71]. These bornyl hydroxycinnamic esters were also confirmed in V. turbacensis [72,73], demonstrating that the plant utilizes multiple chemical classes to exert its powerful pharmacological effects. However, this inherent diversity leads to high chemotypic variability across different populations, with the relative percentages of key sesquiterpenes fluctuating significantly depending on the plant’s maturity. This variability complicates standardization efforts, emphasizing the need for robust protocols to ensure the consistency and reproducibility of IC50 and MIC data [68].
2.2.3. Non-SL Chemical Arsenal and Biosynthetic Complexity
The total pharmacological profile of Verbesina extracts is a complex outcome of a dense and multi-layered chemical defense system that extends far beyond the dominant SLs [11,74]. Qualitative phytochemical screening confirms the simultaneous presence of nearly all major secondary metabolite classes including Alkaloids, Glycosides, Saponins, Phenols, Tannins, Flavonoids, and Terpenoids, a comprehensive chemical diversity that necessitates a thorough investigation of synergistic effects for accurate biological interpretation [74].
Guanidine Alkaloids: The Dual Challenge of Toxicity and Therapy:
A critical challenge for advancing Verbesina metabolites to clinical application lies in managing the presence of guanidine alkaloids, a class chemically distinct from the dominant SLs [75]. This class presents a unique therapeutic duality. On one hand, toxicological investigations into V. encelioides confirmed the presence of the highly potent, non-selective toxin galegine (3-methyl-2-butenyl-guanidine) [27,76], which is responsible for severe sheep and cattle losses and is a documented hazard for grazing livestock [17,27]. This toxin's acute lethality underscores the non-selective nature of the raw plant's defense system [17]. On the other hand, the genus yields therapeutically valuable analogs, notably from V. caracasana, including the monomeric hypotensive agent caracasanamide and the dimeric, cyclobutane-containing caracasandiamide [77,78,79,80,81,82,83]. These specialized guanidino-amide alkaloids represent a significant chemical class for cardiovascular therapy [75]. The natural co-occurrence of the lethal toxin galegine with chemically related, but non-toxic supportive molecules like N-2,3-dihydroxy-3-methylbutyl-acetamide in V. encelioides provides strong proof-of-concept that selectively isolating a non-cytotoxic therapeutic agent is chemically feasible, emphasizing the imperative for rigorous bioassay-guided fractionation [76,84].
Phenolic Compounds, Flavonoids, and Supportive Metabolites:
The extract's capacity for anti-inflammatory and antioxidant activity is substantially defined by a high content of phenolic compounds and flavonoids [18,85]. The overall bioactivity is reinforced by the presence of a diverse class of flavonol diglycosides, including quercetin 3-galactoside-7-glucoside and quercetin 3-xyloside-7-glucoside from V. encelioides [86,87], and the novel Rhamnocitrin-3-O-glucuronide from V. myriocephala [88]. More recent analyses have identified high concentrations of the anti-inflammatory and antioxidant agent Rutin (quercetin-3-O-rhamnoglucoside) in V. sphaerocephala [39] and catechin derivatives (catechin-3-glycoside isomers) and phylloflavan in V. crocata [36,43]. The biological role of these non-SL components is critical, as they include sitosterol derivatives which exhibit anti-inflammatory, antineoplastic, and immune-modulatory activities. This dense profile of phenolic acids such as, p-coumaric acid, gallic acid, caffeic acid, etc. [17,85] is directly linked to the allelopathic activity of the genus; HPLC analysis of V. encelioides root leachate confirmed that phenolic compounds are functionally involved in the chemical interference with competing flora [3]. Furthermore, early phytochemical work confirmed the presence of highly reactive phenolic conjugates, including caffeic acid ethyl ester (a compound previously unknown as a natural product) and caffeic acid borneol ester in the aerial parts of species like V. angustifolia and V. greenmannii [89].
The extensive chemical profile is confirmed by comparative spectroscopic analyses, such as Raman spectroscopy on wild species like V. virginica, which exhibits a distinctly measurable spectrum based on marker bands assigned to scaffold molecules such as phenylpropanoids and carotenoids, supporting the fundamental premise that these specialized secondary metabolites produce unique chemical fingerprints that correlate with the diverse biological activities reported [90]. However, while the genus produces an array of defensive compounds, physiological studies reveal intrinsic limitations in their generalized antioxidant capacity (AA) against environmental stressors. In V. occidentalis (crown-beard), leaf AA content is relatively low (2-4 μmol/g fresh weight) compared to other resilient species. Critically, as the season progresses, V. occidentalis exhibits a diminished capacity to convert oxidized DHA back to active AA (reduced redox state), suggesting a vulnerability in its natural defense cycle against sustained oxidative stress. These intrinsic physiological limitations highlight the necessity for the plant to employ potent, externally acting, high-efficacy compounds, such as SLs, for sustained protection against biotic and abiotic challenges [91].
Triterpenoids, Alkaloids, and Specialized Defense Structures:
The genus exhibits further remarkable biosynthetic capacity through the production of Triterpenoid Saponins [92]. A comprehensive investigation of V. virginica yielded six new compounds (Verbesinosides derivatives) possessing a novel 15,27-cyclooleanane-type triterpenoid skeleton [93], distinct from the oleanolic acid derivative Copteroside E isolated from V. suncho [94]. The presence of these saponins, along with other non-SL triterpenoids such as friedelin, epifriedelin, α- and β-amyrin, and lupeol in V. encelioides [11,45,95], is vital as they contribute to the extract's complexity, with saponins potentially influencing membrane permeability and thereby enhancing the cellular penetration of other lipophilic compounds [94].
This complex defensive system also includes a diverse range of Acetylenic compounds and their cyclized sulfur derivatives, the thiophenes, which are recognized as chemotaxonomic markers of the Heliantheae tribe [96]. The investigation of Mexican Verbesina species confirms the genus's rich, yet structurally diverse, chemical composition, reinforcing the structural complexity of these specialized compounds. The roots of V. angustifolia contained the linear C13 polyacetylene pentainene, along with the diterpene acids. Similarly, the roots of V. oncophora yielded polyacetylene pentainene and a diterpene alcohol [89]. A reinvestigation of the roots of V. virginica (collected in Louisiana) further yielded the ubiquitous polyacetylene pentainene and caryophyllene along with eudesmane derivatives [97]. Thiophenes, such as C13-Monothiophenes and C13-Dithio compounds, are known for their strong biological activity due to their ROS-generating photoactivation potential [96]. The complete profile even extends to specialized heterocycles, evidenced by the isolation of the only known 2-isopropyliden-2H-benzofuran-3-one from V. luetzelburgii [98].
Non-Traditional Bioactive Classes and Technological Application
Recent analytical studies have significantly broadened the understanding of the genus's therapeutic potential by identifying non-traditional bioactive classes. The neuroprotective activity of the V. encelioides flower extract, for example, is chemically linked to carotenoids, such as Spirilloxanthin (12.38-13.24% of the total composition), and the diterpene lactone Ginkgolide A. These compounds demonstrate efficacy by binding to the enzyme Acetylcholinesterase (AChE) [19]. Furthermore, the genus's defensive strategy employs structural macromolecules, with biochemical analysis of V. encelioides identifying a low molecular weight 14 kDa protein characterized as a host defense compound exhibiting moderate antimicrobial activity [99]. The roots of V. encelioides also contain fundamental biomolecules that contribute to the extract's complexity, including the amino acids nor-leucine and ornithine and the carbohydrates D-ribose and sucrose [11,100].
The plant's rich organic matter and protein profile is fundamentally linked to its bioactivity as a growth-promoting component [12]. More fundamentally, the high performance is linked to the genus’ known content of organic matter, carbohydrates, and proteins, suggesting they provide essential growth-promoting components which complement the high-potency, often toxic, defensive nature of the Sesquiterpene Lactones. The bioactivity of the extracts is attributed to their complex profile of organic compounds and proteins, enabling enzymatic activation for nutrient assimilation. Although key minerals like Nitrate and Potassium were not detected in the aqueous extracts, the presence of other nutrients like Ammonia, Ca, Mg, and Phosphate was confirmed [13]. This comprehensive chemical complexity is now leveraged for nanotechnology; the hot water extract of V. encelioides and the aqueous extract of V. crocata are successfully used for the "green synthesis" of Silver Nanoparticles (AgNPs), where the compounds facilitate the reduction and chelation/stabilization of the resulting nanoparticles through redox processes [101,102].
3. Cytotoxic Activity of Verbesina Metabolites: Focus on IC50 Profiles
The inherent chemical reactivity of SLs, driven by the α-methyleneγ-lactone moiety, ensures their broad-spectrum biological activity, encompassing both antimicrobial and antiproliferative effects. While this potency is desirable, the historical lack of quantitative cytotoxic investigation has often relegated the genus to a preliminary research stage [21]. Early studies noted that out of over thirty-eight phytochemically characterized Verbesina species, only four had been biologically investigated, with limited IC50 data available [21]. Consequently, the primary objective of this section is to compile and analyze the recently available quantitative cytotoxicity data, providing a contemporary and rigorous assessment of the genus’s true antiproliferative potential against cancerous and non-cancerous models, thereby establishing the crucial host-toxicity component for the final Selectivity Index (SI) calculation.
3.1. Anticancer Potential Across Cell Lines
The cytotoxic potential of Verbesina compounds is confirmed by consistent activity across both crude extracts and purified isolates, validating the eudesmane scaffold as an inherently bioactive template. Methanolic extracts of V. encelioides demonstrated significant and quantifiable cytotoxic activity against diverse human cancer models, displaying highly consistent potency across three tumor cell lines after 48 hours of exposure. The extract achieved half-maximal growth inhibition (GI50) values of 26.32 ± 2.42 μg/L against MCF-7 (Breast Adenocarcinoma), 28.65 ± 2.89 μg/L against NCI-H460 (Non-Small Lung Cancer), and 26.82 ± 8.54 μg/L against SF-268 (CNS Cancer). This consistency across disparate cancer origins suggests a broad-acting, fundamental cytotoxic mechanism, which is a hallmark of the reactive SLs that define the genus’s chemical profile [25].
However, the efficacy is strongly dependent on the extraction method used. Comparative studies using both ethanolic (VE) and aqueous (VA) crude extracts of V. encelioides against the human colon cancer cell line HCT-116 revealed a stark difference in potency. The ethanolic extract (VE) exhibited significantly higher potency with an IC50 of 310 μg/mL (0.31 mg/mL), which was twice as potent as the aqueous extract (VA) at 650 μg/mL (0.65 mg/mL). This difference confirms the strategic necessity of concentrating the lipophilic compounds, primarily Sesquiterpene Lactones and triterpenoids, through non-polar extraction methods to maximize the yield of active lead molecules [26].
The intrinsic activity of the core eudesmane scaffold itself, even before cinnamoyl esterification, is confirmed by low micromolar cytotoxicity. The key sesquiterpenediol, Verbesindiol (4α, 6β-dihydroxyeudesmane), isolated from V. virginica, demonstrated potent activity against advanced, castration-resistant prostate cancer cell lines. Specifically, Verbesindiol achieved an IC50 of 4.0 ± 0.6 μM against PC-3 (Androgen-independent) and 5.2 ± 0.6 μM against DU 145 (Androgen-independent) cells. This demonstrates that the parent eudesmane skeleton is inherently cytotoxic, positioning Verbesinol derivatives as excellent starting points for SAR investigation aimed at optimizing selectivity [103].
Crucially, isolated SLs, the primary focus of this review, also demonstrate high potency. The core eudesmane scaffold, 4β-cinnamoyloxy,1β,3α-dihydroxyeudesm-7,8-ene, exhibited potent antiproliferative activity (GI50) against a broad panel of human tumor cell lines, with values ranging from 14.7 μM to 32.3 μM [34]. This finding is complemented by related research on V. persicifolia, which isolated an eudesmane triol mono-cinnamate reported to display promising anti-inflammatory, anti-hypoglycemic, and powerful peroxynitrite scavenging activities, along with measurable cytotoxicity against different tumor cell lines. This array of activity across multiple compounds in V. persicifolia directly establishes it as a major source of selectively toxic scaffolds [61]. This translational potential is further substantiated by a Mexican patent detailing the isolation of the eudesmane 4CDE (a core scaffold), which is claimed for the treatment of diabetes mellitus type 2, cancer, obesity, and as an anti-inflammatory, directly validating the high-value, multifaceted application of this genus [104].
However, not all crude extracts exhibit high cytotoxicity. The hydroethanolic extract from the stem of V. turbacensis (a species traditionally used for inflammation) demonstrated a low cytotoxic effect against the human hepatoma cell line HepG2. After 24 hours of exposure, the extract showed minimal toxicity (IC50 > 500 μg/mL), and even after 48 hours, the IC50 remained relatively low at 334.6 μg/mL. This low-to-moderate cytotoxicity reinforces the argument that the high-potency effects of SLs are often diluted in crude hydroethanolic extracts, necessitating chemical purification to isolate the potent molecules [73].
Finally, the potential for cellular specificity in the crude extracts is an important consideration for SI optimization. The alcoholic extract of V. encelioides exhibited a differential cytotoxic response across human cancer cell lines. While the extract was strongly cytotoxic to HepG2 cells, reducing viability to 53% at 1000 μg/mL, it proved non-cytotoxic to A-549 (human lung cancer) cells, even at the highest concentrations tested. This inherent degree of cellular specificity suggests that isolating key compounds may yield a selective drug candidate with a favorable SI [105].
3.2. Molecular Mechanisms of Cytotoxicity
The potent IC50 profiles observed in Verbesina metabolites are fundamentally linked to the inherent, non-selective chemical reactivity of the SLs, which is primarily executed through mitochondria-mediated apoptosis induced by the generation of Reactive Oxygen Species (ROS) [26,105]. This mechanism is consistent with the predicted alkylating action of the α-methylene-γ-lactone pharmacophore against nucleophilic sites on cellular proteins and peptides (Figure 3) [26].
3.2.1. Oxidative Stress and the Apoptotic Cascade
Mechanistic studies using the ethanolic extract (VE) of V. encelioides against the HCT-116 colon cancer cell line definitively established the full cascade of cell death. The initial event involves oxidative stress induction; exposure to the VE significantly increased the level of Lipid Peroxidation (LPO) (up to 74%) while simultaneously causing a substantial Glutathione (GSH) depletion (up to 46%), strongly indicating a pro-oxidant mechanism. This chemical stress directly leads to a dose-dependent surge in intracellular ROS production, which subsequently disrupts the integrity of the mitochondrial membrane. This disruption results in a significant loss of Mitochondrial Membrane Potential (MMP) (up to 54% decline), leading to mitochondrial damage [26].
The cytotoxic mechanism is further confirmed at the molecular level. The disruption of the MMP triggers a classic apoptotic cascade, characterized by the significant upregulation of pro-apoptotic genes. Specifically, the extract significantly upregulated the expression of Bax (by 2.1-fold), p53 (by 2.9-fold), caspase-3 (by 2.2-fold), and caspase-9 (by 2.5-fold), while simultaneously downregulating the anti-apoptotic gene Bcl-2 (by 0.55-fold). The resulting activation of caspases confirms that programmed cell death is the primary mode of action [26]. This mechanism is consistent with separate studies on the alcoholic extract of V. encelioides against HepG2 cells, which also found that the primary cause of cell death was oxidative stress, resulting in mitochondrial damage and genotoxic potential evidenced by extensive DNA damage quantified by the Comet Assay. This DNA damage likely triggers the observed G2/M cell cycle arrest (up to ~50% at 1000 μg/mL), leading to cell death upon repair failure ( [105]. Importantly, this mechanism appears to be non-folate related, as the V. encelioides methanol extract displayed no inhibitory effect on DHFR enzyme activity, supporting the hypothesis that the cytotoxic action proceeds via non-enzymatic Michael addition rather than competitive enzyme antagonism.
3.2.2. Scaffold Dependency and Mitochondrial Targeting
Mechanistic studies focusing on isolated pure eudesmane scaffolds confirm that the cytotoxic mechanism is definitively mapped to the mitochondria, but the mode of action is structure-specific, a crucial finding for SAR and SI optimization. One identified mechanism involves mild uncoupling of the respiratory chain. Studies on the pure eudesmane CDE (4β-cinnamoyloxy,1β, 3α-dihydroxyeudesm-7,8-ene), a characteristic scaffold of the genus, revealed that it acts as a natural mild uncoupler in rat liver mitochondria (RLM). This effect involves CDE inducing a drop in the mitochondrial membrane potential (ΔΨ) and causing a bioenergetic collapse. Mechanistically, CDE is proposed to interact with the respiratory chain, particularly at the level of cytochrome C oxidase (COX), leading to the generation of H2O2 and subsequent ROS production. This critical finding confirms that the cytotoxic or anti-proliferative action of Verbesina SLs is not confined to simple alkylation but includes complex regulation of cellular energy metabolism, a crucial pathway in both cancer and microbial persistence [62].
A second, more nuanced mechanism was revealed by the comprehensive mechanistic investigation of 4β-cinnamoyloxy,1β,3α-dihydroxyeudesm-7,8-ene and its derivatives (Compound 1), which differentiated the mode of action based on subtle functionalization of the eudesmane skeleton. This work identified two distinct pathways for apoptosis. The first pathway involves Mitochondrial Permeability Transition (MPT) induction: the native hydroxyl groups on the eudesmane moiety (found in the lead Compound 1 and the hydrogenated Compound 3) were found to be crucial for inducing Ca2+-dependent MPT, which leads to mitochondrial swelling and the release of pro-apoptotic factors (Cyt C and AIF). The second pathway involves Respiratory Chain Inhibition: conversely, when the hydroxyl groups were masked by acetate substituents (in the diacetate Compound 2 and hydrogenated diacetate Compound 4), the mechanism shifted dramatically. These acetylated compounds caused injury by directly inhibiting Succinate dehydrogenase of the mitochondrial respiratory chain and inducing a general uncoupling effect on oxidative phosphorylation (Figure 4). This complex mechanistic picture demonstrates that subtle structural modifications on the eudesmane skeleton drastically alter the cellular target, which is a key consideration for chemically optimizing the SI [34].
3.2.3. Long-Term Cytotoxic Profile
Understanding the long-term cytotoxic profile of these scaffolds is essential for translating acute IC50 data into a true therapeutic advantage. Further mechanistic studies on Verbesindiol, the non-esterified core scaffold, revealed a complex profile. While the compound demonstrated high acute cytotoxicity (84-97% cell death) in the SRB assay, suggesting excellent initial anti-cancer potency, clonogenic assays (which measure the ability of cancer cells to form colonies after drug washout) showed that compound 6 had no detectable effects on colony-forming ability. This surprising result suggests that the initial cytotoxic mechanism of Verbesindiol does not prevent the long-term regenerative capacity of the cells, differentiating its mode of action from other Verbesina metabolites that do cause G2/M arrest and inhibit colony formation. This complexity emphasizes that achieving a true therapeutic advantage relies not just on acute IC50 values, but on understanding the compound's selective long-term mechanism of action [103].
3.3. Toxicity to Non-Cancerous Cells
The determination of a favorable SI is the central translational challenge for Verbesina metabolites, a necessity rooted in the genus’s long-standing documentation as a poisonous plant [8,27]. This history provides the historical imperative for rigorous toxicity screening (IC50/CC50) in modern pharmacology [27]. The non-selective and potentially fatal nature of the crude plant material is demonstrated by the potent guanidine alkaloid, galegine (3-methyl-2-butenyl-guanidine), which is classified as a documented hazard for grazing livestock in the United States. Exposure to galegine causes severe pathological effects and death in sheep, with observed symptoms consistent with congestion of the lungs and fluid in the chest cavity [11,27]. The high cytotoxic activity of this known poison is confirmed by laboratory testing, where isolated Galegine was found to be highly potent against the HepG2 (Hepatocellular Carcinoma) cell line, exhibiting an IC50 value of 10.9 μg/mL [87]. This high degree of cytotoxicity for a known poison further underscores the absolute necessity of rigorous bioassay-guided isolation and chemical optimization to establish a sufficient SI for any therapeutic lead compound [14]. The historical toxicological findings also revealed a crucial co-occurrence of these non-selective toxins with chemically related, but experimentally verified non-toxic supportive molecules, such as the N-2,3-dihydroxy-3-methylbutyl-acetamide isolated alongside galegine in V. encelioides. This functional differentiation provides crucial proof-of-concept that selectively isolating a non-cytotoxic therapeutic agent from the complex crude mixture is chemically feasible [76].
The core challenge remains the lack of specificity inherent to the reactive SLs. Quantitative data on the core eudesmane scaffold, Compound 1 (4β-cinnamoyloxy,1β,3α-dihydroxyeudesm-7,8-ene) (Figure 4), demonstrated a pronounced lack of specificity toward tumor cell lines and the non-tumorigenic human mesothelial cells (MeT-5A). The GI50 value for Compound 1 on the non-tumorigenic MeT-5A cells (32.3 μM) was found to be directly within the range of its toxicity against tumor cells (14.7 μM against A431). This direct demonstration of non-selective cytotoxicity underscores the central challenge of the entire review: the necessity of maximizing the efficacy (MIC) while minimizing the host toxicity (IC50 to healthy cells) to achieve a favorable SI 34]. The necessity for further toxicity evaluation remains a critical concern for the translational use of all Verbesina extracts; while some aqueous extracts of V. crocata demonstrated diuretic activity without obvious signs of toxicity in rats, the probable presence of the potent toxin galegine makes the overall toxicity of the plant a primary barrier to advancing novel scaffolds [23]. Furthermore, the strong allelopathic history of the genus provides a clear historical and contemporary imperative for rigorous toxicity screening [17].
Despite these concerns, quantitative data strongly supports the hypothesis that selective separation of the therapeutic scaffold can yield low-toxicity agents. An investigation into the antiprotozoal activity of V. encelioides isolated six major constituents, including the triterpenoids pseudotaraxasterol and its derivatives, and β-sitosterol glycosides. When tested against human embryonic lung fibroblasts (MRC-5), a standard non-cancerous cell line, all isolated compounds were determined to be non-cytotoxic with high values (CC50 > 64 μg/mL). This result contrasts sharply with the activity of many conventional chemotherapy agents and provides crucial evidence of the genus’s potential for a favorable SI [106]. This finding is supported by the study of the crude alcoholic extract of V. encelioides, which showed a strong cytotoxic response against HepG2 and MCF-7 cancer cells but was non-cytotoxic to A-549 (human lung cancer) cells. This differential response to specific cell lines suggests an inherent degree of cellular specificity, indicating that isolating key compounds may yield a selective drug candidate [105].
Furthermore, the toxicity of the EO fraction, which is dominated by sesquiterpene hydrocarbons, also demonstrates a favorable safety profile. The acute toxicity of the V. macrophylla EO was evaluated in mice, with the median lethal dose (LD50) estimated to be greater than 5000 mg/kg (p.o.). The oil exhibited a low rate of hemolysis in human red blood cells (hRBCs), with the highest rate being only 2.14 ± 0.10% at 5000 μg/mL. A hemolysis rate of less than 5% is considered a favorable safety indicator for drug application, suggesting that the EO compounds, despite being lipophilic, interact minimally with the human cellular membrane [39]. The necessity of balancing efficacy with host safety is further reinforced by reports that anthelmintic activity has been linked to high levels of the toxic guanidine alkaloid, galegine, and nitrates in Verbesina alcoholic extracts, reinforcing the core challenge for developing a safe therapeutic agent: maximizing efficacy while rigorously ensuring the selective isolation of compounds away from known toxins like galegine [107].
4. Antimicrobial Efficacy and Anti-AMR Mechanisms
The established chemical arsenal of the Verbesina genus, dominated by SLs, C13 polyacetylenes, and volatile terpenes, translates directly into a potent, broad-spectrum defense system that provides activity against bacterial, fungal, protozoal, and parasitic pathogens. This inherent chemical capability supports the ethnopharmacological use of Verbesina species in treating infections and wounds. The rigorous quantification of this efficacy, often expressed as the MIC, is the critical second metric required to calculate the SI and validate the translational potential of its specialized metabolites [21].
4.1. Activity Against Clinically Relevant Pathogens
The anti-infective capacity of Verbesina metabolites is confirmed across numerous pathogenic kingdoms, ranging from human pathogens to agricultural pests, underscoring the broad-spectrum nature of the plant's chemical defense. Even prior to chemical enhancement, crude extracts of V. encelioides demonstrated reliable anti-infective activity against a range of pathogens, validating their initial use in drug discovery screens. For instance, crude methanolic extracts of V. encelioides exhibited effectiveness against the bacteria Escherichia coli and Vibrio cholerae, as well as the fungi Aspergillus flavus and Aspergillus niger. This robust baseline activity suggests that the chemical potency of the genus's natural metabolites alone is sufficient to warrant further investigation and optimization [108].
Furthermore, the potent anti-fungal properties are highlighted in a process patent that claims the use of V. encelioides extract for treating dermatophyte infections. The extracts demonstrated anti-mycotic activity against clinically relevant fungi, including Microsporum canis, Trichophyton Rubrum, and Malassezia furfur, providing quantitative in vitro validation of the genus’s broad-spectrum anti-infective capacity [109]. The total hydroalcoholic extract and its resulting fractions possess significant and varied antimicrobial properties, demonstrating high activity against Gram-positive bacteria (Staphylococcus aureus and Bacillus subtilis) and fungi (Aspergillus fumigatus and Candida albicans), while showing moderate activity against the Gram-negative bacterium Escherichia coli. Notably, the total hydroalcoholic extract exhibited antifungal activity against C. albicans superior to the reference drug Amphotericin B in the diffusion agar technique. The ethyl acetate fraction generally maintained higher activity than the petroleum ether fraction in the antimicrobial assays, suggesting the concentration of moderately polar, active principles [87].
The efficacy of isolated SLs, the focus of this review, is measured in the low micromolar range. The pure isolated germacrene SL exhibited measurable antibacterial activity against two clinically relevant Gram-positive pathogens, with the MIC determined to be 64 μg/mL against both Staphylococcus aureus (ATCC 29213) and Enterococcus faecalis (ATCC 29212) [38]. SLs isolated from Verbesina species also demonstrate significant activity against fungi and oomycetes, showcasing a broad-spectrum anti-infective capacity. In the study of V. lanata, the crude ethyl acetate extract showed a MIC100 of 35 μg/mL against Plasmopara viticola (grapevine downy mildew). Importantly, when tested as pure compounds, several isolated eudesmane SLs showed MIC100 values below that of the crude extract, confirming they are the primary active principles. The high potency of these pure SLs is critical, as exemplified by compounds such as 6β-cinnamoyloxy-3β,4α-dihydroxyeudesmane, which achieved an MIC100 of 3.9 μg/mL against P. viticola, and 6β-cinnamoyloxy-1β-hydroxyeudesm-4-en-3-one, with an MIC100 of 5.0 μg/mL. This potency, up to 7-fold greater than the crude extract, highlights the necessity of purification for maximizing efficacy and accurately calculating the Therapeutic Index. The eudesmane sesquiterpenes were further confirmed as the primary active constituents responsible for significant antifungal activity against P. viticola, with five compounds achieving MIC100 values less than 10 μg/mL [55].
The EO fraction, which contains volatile terpenes, also demonstrates significant, quantifiable inhibitory effects. The EO from V. negrensis confirmed efficacy against common human pathogens, achieving an MIC of 500 μL/mL against Staphylococcus aureus (ATCC 25923) and 350 μL/mL against Enterococcus faecalis (ATCC 29212). While this EO showed no activity against Gram-negative bacteria such as E. coli or Klebsiella pneumoniae at the tested concentrations, the confirmed efficacy against Gram-positive pathogens is consistent with broader Asteraceae literature [70]. The essential oil of V. macrophylla showed both antibacterial and antifungal activity against strains causing respiratory and urinary tract infections, with MIC values varying from 8 to 512 μg/mL for bacteria and 4 to 32 μg/mL for C. albicans. Notably, the EO's activity against K. pneumoniae (MIC 16 μg/mL) was observed to be superior to the standard antibiotic Amikacin (MIC 32 μg/mL). Furthermore, it achieved potent antifungal activity against Candida albicans (MIC 4 μg/mL) [39].
Beyond bacteria, Verbesina compounds demonstrate broad-spectrum activity against protozoal and parasitic agents [106]. Studies on the Argentinean species V. subcordata showed significant trypanocidal activity against Trypanosoma cruzi (the causative agent of Chagas' disease), with the non-polar dichloromethane extract achieving 97.7 ± 0.2% growth inhibition at 100 μg/mL, suggesting that the lipophilic SLs and terpenoids are responsible for the parasiticidal effect [65]. Expanding on this, aqueous extracts of the native Brazilian species V. macrophylla exhibit potent activity against the same protozoan, eliminating 100% of the epimastigote form of T. cruzi after treatment with a moderate concentration of 200 μg/mL. The inflorescence extract also demonstrated significant fungicidal efficacy, inhibiting the growth of C. tropicalis by 41% [110]. Activity against other Neglected Tropical Diseases (NTDs) is also moderate; for instance, benzyl 2,6-dimethoxy benzoate from V. encelioides showed an IC50 of 38.1 μg/mL against Trypanosoma brucei [106]. The genus also provides strong efficacy in agricultural biocontrol, demonstrating strong nematicidal activity against the economically destructive root-knot nematode, Meloidogyne javanica. The most potent activity was found in the aqueous extract of fresh flowers of V. encelioides, and incorporating the plant material into nematode-infested soil significantly reduced the number of viable juvenile nematodes [11,111].
Finally, non-small molecule defense components confirm that multiple fractions of Verbesina exhibit broad-spectrum activity. An isolated 14 kDa protein from V. encelioides validated the susceptibility of multiple pathogens through zone of inhibition assays, confirming activity across Gram-negative bacteria (E. coli, P. aeruginosa), Gram-positive bacteria (S. aureus, E. faecalis), and Fungal Pathogens (A. flavus, F. oxysporum) [99]. The extracts also demonstrate potent antifungal activity against the problematic necrotrophic fungus Botrytis cinerea, with the aqueous stem extract exhibiting the highest antifungal activity (IC50 of 0.10 mg/mL) [85].
Furthermore, several Verbesina species demonstrate significant in vitro activity against both Gram-positive and Gram-negative bacteria, directly supporting their potential as anti-infective agents. Methanolic extracts of V. sphaerocephala exhibited remarkable antibacterial efficacy, demonstrating growth inhibition that was statistically non-significant from the ampicillin positive control (10 mg/mL) in several tests. Against E. coli (Gram-negative), the leaf extract achieved 95.00 ± 5.00% growth inhibition, and against S. aureus (Gram-positive), the leaf extract showed 90.00 ± 5.00% growth inhibition [36]. Interestingly, the study suggested that the antimicrobial effect was negatively correlated with the total phenolic and flavonoid content, implying that less abundant, possibly more potent, lipophilic compounds, such as the characteristic Sesquiterpene Lactones may be the primary drivers of the antibacterial action, or that a synergistic effect among the diverse metabolites is present.
The efficacy of this wide-ranging arsenal is reinforced by the conclusion that the plant's overall anti-infective potency relies heavily on a synergistic effect between its many constituents, including SLs, triterpenoids, and flavonoids [106]. Furthermore, the widespread crisis of anthelmintic resistance necessitates alternative control strategies, for which Verbesina extracts provide a promising phytomedicine source. The anthelmintic activity in the Egyptian context has previously been linked to high levels of the toxic guanidine alkaloid, galegine, and nitrates in Verbesina alcoholic extracts. This reinforces the core challenge for developing a safe therapeutic agent: maximizing anti-parasitic efficacy while rigorously ensuring the selective isolation of compounds away from known toxins like galegine [107].
4.2. Molecular Mechanisms of Antimicrobial Action
The anti-infective efficacy observed in Verbesina extracts is the result of a multi-layered defense strategy, employing distinct mechanisms to disrupt fundamental processes in diverse microbial kingdoms1. While the cytotoxic mechanism of SLs is primarily through Michael addition to nucleophilic proteins, the genus deploys non-SL metabolites that operate through separate cytotoxic pathways, often resulting in complex synergistic interactions.
4.2.1. The Core Mechanistic Hypothesis: Alkylation and Membrane Disruption
The primary hypothesis for SL action in microbial systems mirrors that in mammalian cytotoxicity: the highly reactive α-methylene-γ-lactone moiety acts as a non-specific alkylating agent. This mechanism, which involves non-enzymatic Michael addition to nucleophilic sites on biological proteins, distinguishes the SLs from many conventional antibiotics that target specific enzymes like DHFR (dihydrofolate reductase). Studies have demonstrated that the V. encelioides methanol extract displayed no inhibitory effect on DHFR enzyme activity despite robust cytotoxic efficacy, strongly supporting this non-enzymatic mechanism [25].
This alkylation mechanism leads to significant structural damage to the target cells. For instance, ultrastructural analysis of T. cruzi epimastigotes exposed to V. macrophylla extract revealed extensive damage, including cytoplasmic and nuclear disorganization, vacuolization, and swelling of reservosomes. This disruption, confirmed via Sytox Green fluorescence, indicates a compromised plasma membrane. The mode of action, disrupting the crucial cell membrane barrier reinforces the importance of identifying compounds that bypass conventional resistance pathways. Researchers strongly suggest that the sesquiterpene lactones present in V. macrophylla contribute to this membrane-disrupting and T. cruzi activity, further establishing the SL class as the driver of anti-infective effects in the genus [110].
4.2.2. Specialized Mechanisms of Non-SL Metabolites
The broad spectrum of activity observed in crude extracts (cytotoxicity, anthelmintic, and anti-fungal) relies heavily on the synergistic interaction of diverse metabolites, each acting via a different, specialized mechanism [66]. This co-occurrence of various chemical classes suggests a multi-layered defense and multiple modes of action that contribute to the overall biological effects observed in crude extracts. For instance, the bornyl caffeic acid ester and related compounds combine a terpene scaffold with a phenolic acid, structures known for antioxidant and anti-inflammatory roles, hinting at complex synergistic interactions between the various metabolites [89]. Other specialized metabolites rely on light-activated mechanisms for cytotoxicity. Thiophenes (sulfur-containing acetylenes) are highly susceptible to photoactivation, generating singlet oxygen and other ROS when exposed to UV-A light. This mechanism leads to broad-spectrum toxicity, often by causing irreparable damage to the microbial cell membrane and DNA [96].
The anti-protozoal efficacy of Verbesina compounds is supported by targeted molecular mechanisms beyond non-selective cell damage. The bornyl esters isolated from V. turbacensis were found to possess potent inhibitory activity against the cysteine protease rhodesain. Rhodesain is the major cysteine protease utilized by the parasite Trypanosoma brucei rhodesiense (the causative agent of African trypanosomiasis/sleeping sickness). This targeted enzyme inhibition confirms that Verbesina metabolites can achieve high efficacy by directly interfering with a specific, critical virulence factor of a neglected tropical pathogen [72].
Other specialized metabolites rely on light-activated mechanisms for cytotoxicity. Thiophenes (sulfur-containing acetylenes) are highly susceptible to photoactivation, generating singlet oxygen and other ROS when exposed to UV-A light. This mechanism leads to broad-spectrum toxicity, including anti-fungal and anti-bacterial activity, often by causing irreparable damage to the microbial cell membrane and DNA. The inclusion of these photoactive metabolites in Verbesina extracts further supports the hypothesis that the gross biological effect is a complex synergistic interaction between multiple compound classes, each operating via a distinct mechanism [96].
The oil extract of V. alternifolia, which demonstrated potent adulticidal anthelmintic properties against the equine parasite Habronema muscae, revealed a mechanism linked to physical and nutritional interference. Scanning Electron Microscopy (SEM) analysis revealed irreversible degenerative changes to the parasite's external morphology. Changes included shrinking, detachment, and distortion of the cuticle, the basic entry route for anthelmintic drugs and deformation of the lips and papillae. This mechanism suggests that the active components, attributed to the presence of terpenoids, flavonoids, and aromatic compounds, facilitate their effect through passive diffusion and subsequent membrane and structural disruption. Furthermore, the formation of aggregates in and around the buccal capsule (anterior digestive tract) suggests a disruptive action that may interfere with the nematode's nutrition, potentially leading to undernourishment and mortality, similar to the proposed action of SLs targeting cellular processes [107].
The antifungal mechanism is also multi-faceted. The fungicidal effect against Botrytis cinerea is hypothesized to be due to a synergistic effect of the identified phenolic compounds. Individual phenolics present disrupt key fungal processes in multiple ways: Protocatechuic acid affects membrane permeability; Vanillic acid inhibits ergosterol synthesis; and Quercetin alters cell membrane composition through oxidative stress [85]. Similarly, the potent anti-cyanobacterial effect of the aqueous extract of V. encelioides against Microcystis aeruginosa involves significant physiological disruption, causing a marked decrease in the content of Chlorophyll-a and carotenoids, demonstrating that the compounds interfere with photosynthesis and ultimately induce cell death [112].
4.2.3. Ecological and Biosynthetic Context
The sheer chemical arsenal confirmed throughout the genus is a functional manifestation of potent allelopathic properties. The economic and ecological pressure exerted by V. encelioides is directly linked to the use of defensive secondary compounds that enable it to resist both pests and competing flora [42]. This allelopathic effect suggests that the active metabolites, whether SLs or acetylenic compounds have mechanisms of action capable of disrupting fundamental cell survival processes (membrane integrity and energy metabolism) that are common across diverse microbial kingdoms [40,74].
The defense mechanism is further supported by direct anatomical evidence: Histochemical staining confirms that the synthesis and storage of Verbesina's key metabolites occur directly within specialized glandular trichomes. These trichomes actively accumulate terpenoids (including sesquiterpenes) and phenolic/fluorescent metabolites. This direct evidence of a structural defense system confirms that the compounds responsible for antimicrobial activity and cytotoxicity are natural defensive agents designed for broad-spectrum biotic challenge. [44]. The biosynthetic process supporting this defense is highly elaborate. In V. macrophylla, specialized leaf secretory ducts actively produce and store defensive materials, including terpenes and alkaloids, to combat herbivores and microbes via a complex process known as granulocrine secretion. This elaborate, energy-intensive mechanism, where secretion begins in the plastids of specialized transfer cells, confirms that anti-infective and anti-herbivory activities are a primary ecological function of the Verbesina genus, supporting the hypothesis that these compounds possess powerful, broad-spectrum bioactivity [69]. This non-selectivity further emphasizes the necessity of rigorously quantifying the SI to ensure any potent anti-AMR lead compound spares healthy mammalian cells [40].
Finally, the layered defense strategy is also mediated by macromolecules; proteins from young leaves of V. encelioides exhibit moderate antimicrobial activity against various bacterial and fungal species, demonstrating a chemical and biological defense strategy [11]. The plant's allocation strategy is predicted by the Limiting Resource Model (LRM), which suggests that when the host plant experiences stress (parasitism), it increases the allocation to shoot mass, directing more resources to the area where many of its chemical defenses (SLs) are sequestered, potentially suggesting a prioritized defense response to biotic stress [6].
4.3. Combating Resistance: New Approaches
The potential of the Verbesina genus to address resistance extends beyond direct antimicrobial activity, encompassing therapeutic strategies that support the host immune system, target chronic diseases, and leverage modern nanotechnology for enhanced efficacy. This multifaceted utility highlights the strategic value of the plant as a bioprospecting source for complex, next-generation scaffolds.
4.3.1. Host-Mediated and Anti-Inflammatory Strategies
The established traditional use of Verbesina for wound healing and inflammation provides compelling in vivo evidence supporting its systemic anti-infective and anti-inflammatory potential. Topical application of the methanolic extract (VCME) of V. crocata to incisional wounds in a murine model demonstrated significant acceleration of the healing process, reducing the final closure time (FCT) to 12 days, which was faster than the positive control (Recoveron®) and the negative control (Vaseline). Crucially, the VCME significantly increased the tensile strength of the healed tissue (700 ± 148.3 g) compared to the positive control (640 ± 259.9 g), indicating superior collagen organization [22,23]. Histological analysis confirmed that the extract advanced the healing process to the remodeling phase by promoting the synthesis of mature collagen and significantly reducing the number of fibroblasts. This efficacy is attributed to the extract’s anti-inflammatory action (linked to sitosterol glycoside and known sesquiterpenes) and its potential for immune modulation (phylloflavan activates macrophages), demonstrating the extract's ability to control the initial infective and inflammatory phases of wound repair [22].
The precise structural requirement for this anti-inflammatory activity is defined by the core eudesmane scaffold's functionalization. Pure 4β-cinnamoyloxy, 1β,3α-dihydroxyeudesm-7,8-ene demonstrated a notable anti-inflammatory activity, achieving 83.18% edema inhibition in vivo (ED50 = 0.43 μM). This effect is strongly dependent on the integrity of the cinnamate moiety: chemical modifications such as acetylation and hydrogenation resulted in a significant reduction in anti-inflammatory activity (up to 60% reduction for the hydrogenated derivative), underscoring a prevailing role of the unsaturated double bonds of the cinnamate scaffold in this biological effect [34]. This mechanism of reducing the inflammatory phase is further supported by the EO fraction, which demonstrated strong anti-inflammatory and antipyretic activities statistically similar to or exceeding commercial drugs like Dexamethasone and Dipyrone, respectively. The anti-inflammatory effect (carrageenan-induced peritonitis model) is associated with the reduction of the pro-inflammatory cytokines TNF-α (up to 73.52%) and IL-1β (up to 87.71%), consistent with the activity of its major components, β-caryophyllene and Germacrene D, which are potent inhibitors of pro-inflammatory cytokine production [39].
This mechanism of host-defense enhancement is further validated by studies on V. oerstediana, which demonstrated crucial immunomodulatory activity. Extracts of V. oerstediana (VEOEBA) significantly enhance the antimicrobial efficacy of human monocytes and neutrophils against Staphylococcus aureus infections. Mechanistically, this enhancement is attributed to the extract's ability to significantly stimulate the production of Neutrophil Extracellular Traps (NETs). NETs are extracellular fibers composed of DNA-histone complexes that entrap and kill bacteria, representing a critical, non-phagocytic defense mechanism, suggesting that Verbesina compounds can serve as adjunctive AMR therapies by boosting the host's innate immune response [113].
4.3.2. Non-Infectious Therapeutic Applications (Chronic Disease and SAR)
The chemical versatility of the genus ensures its therapeutic utility extends to high-value areas outside of acute infection, particularly in managing chronic human diseases. The guanidino-amide alkaloids are a significant class of potent hypotensive agents [75]. The novel monomer caracasanamide from V. caracasana demonstrated a dose-dependent hypotensive effect, reducing mean blood pressure while simultaneously increasing cardiac inotropism (dP/dt) and respiratory function [79,80,114]. Critically, this compound does not decrease cardiac inotropism or chronotropism at nontoxic doses, in contrast to most other hypotensive drugs tested, making it a clearly advantageous therapeutical aid for moderate arterial hypertension [77,80]. The pursuit of structural analogs, such as the dimeric caracasandiamide [78] and prenylagmatine analogs [82,83], demonstrated that subtle variations within the alkaloid structure significantly enhance pharmacological action, providing a clear path for chemical modification and optimization of the genus’s cardiovascular agents [82,84,114].
Beyond cardiovascular agents, the genus demonstrates efficacy in managing metabolic syndrome. The extract of V. persicifolia was found to significantly alter glucose tolerance, enhance glucose uptake in skeletal muscle, and notably inhibit glycogenolysis in the liver, suggesting it is effective in controlling blood glucose following a heavy carbohydrate diet [33]. Furthermore, the ethanolic extract of V. encelioides roots exhibited significant hypolipidemic activity in rats, resulting in a measured decrease in total cholesterol, triglycerides (TGL), and very low-density lipoproteins (VLDL), while maintaining or increasing high-density lipoproteins (HDL) [115]. The genus's therapeutic scope is rapidly expanding to include neuroprotection and anticonvulsant activity; the methanolic extract of V. persicifolia demonstrated highly significant anticonvulsant activity in the zebrafish seizure model, resulting in a 100 survival rate after a PTZ-induced seizure event at a non-toxic concentration [37]. Aqueous extracts of V. crocata also exhibit measurable diuretic activity, comparable to the positive control furosemide in increasing sodium and potassium clearance and excretion [23].
The most compelling evidence for a favorable SI drived from a Verbesina species is provided by research on V. lanata. Compounds closely related to the genus's characteristic scaffolds were evaluated for anti-protozoal activity against Leishmania major promastigotes, showing dose- and time-dependent activity with IC50 values ranging from 4.9 to 25.3 μM. Crucially, the toxicity against normal murine macrophages (CC50) ranged from 432.5 to 620.7 μM, yielding a high SI of up to 88.2 for Arbusculin Derivative ((1R,4S,4aS,7R)-1-methyl-7-(prop-1-en-2-yl)octahydro-2H-1,4a-(epoxymethano)naphthalen-4-yl acetate). This SI value is substantially better than that of the standard drug, meglumine antimoniate (glucantime), which had an SI of 40.0. This quantitative comparison provides definitive validation that eudesmane scaffolds can yield lead compounds with a high therapeutic window [15].
4.3.3. Technological Valorization and Biocontrol Applications
The chemical complexity of Verbesina allows its extracts to be leveraged in modern nanotechnology, offering a rapid, safe, and environmentally friendly avenue for combating AMR. The methanolic and ethanolic extracts of V. encelioides contain natural reducing agents ( polyphenols, flavonoids, and terpenoids) capable of performing the green synthesis of silver nanoparticles (AgNPs). These fabricated AgNPs then exhibit superior antimicrobial action compared to the extracts alone against both bacteria (E. coli and V. cholerae) and fungi (Aspergillus spp.) [101,108]. This successful synthesis is further supported by the use of purified saponin extracts from V. encelioides as natural reducing and stabilizing agents for AgNPs used in dermal applications, such as the management of acne [92]. The successful synthesis using V. crocata further supports the viability of this strategy, with the synthesized Ag/AgCl-NP's exhibiting sizes less than 100 nm, indicating strong potential for enhanced antimicrobial and anti-biofilm activity [102]. The broad utility of Verbesina metabolites suggests non-traditional applications in combating resistance, as phenolic and other compounds within the aqueous extract serve as natural reducing agents for the green synthesis of AgNPs, which exhibit enhanced antimicrobial activity against pathogens like E. coli and Vibrio cholerae. This suggests Verbesina extracts can be directly leveraged in novel drug delivery and anti-AMR formulations [11].
Furthermore, the plant's aggressive ecological resilience is being valorized for biocontrol applications. The nematicidal capability of Verbesina opens a new avenue for AMR research by targeting biocontrol agents that reduce reliance on synthetic pesticides [111]. Aqueous extracts of the plant show strong nematicidal activity against the root-knot nematode Meloidogyne javanica, indicating its potential as a green manure or natural nematicide. Furthermore, the plant’s aggressive ecological resilience is being valorized for biocontrol applications. A patent claims an anti-termite herbal composition derived in part from the flowers and leaves of V. encelioides, demonstrating its commercial potential as an eco-friendly pesticide. This supports the strategy of leveraging the plant's natural chemical defense system, driven by compounds like alkaloids and saponins, as a low-cost, sustainable biopesticide solution [116].
The demonstrated allelopathic potential of V. encelioides offers an innovative route for biocide valorization as an environmentally friendly control agent for cyanobacterial blooms in eutrophic water bodies. The aqueous extract of the aerial parts of V. encelioides exhibits a strong anti-cyanobacterial effect on the growth of the harmful species, Microcystis aeruginosa, with a maximum Inhibitory Rate of 93% achieved at 1 mg/mL concentration. This efficacy is noted to be due to the compounds interfering with photosynthesis and ultimately inducing cell death [112].
The severe agricultural impact of V. encelioides as a weed and as a reservoir host for thrips (vectors for Tomato Spotted Wilt Virus, TSWV) strongly justifies the development of Verbesina-derived compounds as high-value, natural biopesticides [1,5,8]. Beyond biopesticides, the genus exhibits potential for industrial valorization. Early screenings for renewable energy sources identified V. alternifolia as a rubber-producing species yielding a quantifiable percentage of hydrocarbon (0.4%), confirming its utility as a source of low-molecular-weight polyisoprenes and hydrocarbon feedstocks [117]. Conversely, the extracts demonstrate significant agricultural potential by acting as growth biostimulants, particularly in high-value crops like strawberry (Fragaria ananassa). Application of aqueous extracts of V. sphaerocephala and V. fastigiata resulted in notable increases in productive parameters, including up to a 207.14% Agronomic Efficiency Index (IEA) for fruit and flower production [13]. The intrinsic chemical resilience of V. encelioides also opens a high-value application in phytoremediation, suggesting its potential role in the stabilization or phytoextraction of heavy metals from contaminated mine waste [7].
5. Critical Analysis: The Therapeutic Index and Drug Feasibility
The successful translation of a natural product into a pharmaceutical lead depends entirely on its ability to demonstrate preferential toxicity, which is quantified by the Selectivity Index (SI = IC50/MIC)[28]. The inherent chemical characteristic of the Verbesina genus, defined by highly reactive SLs and potent alkaloids, presents a formidable challenge: the strong potential for efficacy (MIC) is inextricably linked to the risk of host toxicity (IC50). This section critically analyzes the compiled data, providing quantitative and structural arguments to assess the feasibility of isolating and optimizing SL scaffolds with an intrinsically superior therapeutic window.
5.1. Calculation and Interpretation of the Selectivity Index (SI)
The determination of the SI is complicated by the presence of a diverse chemical arsenal, yet quantitative evidence confirms that a favorable SI is achievable through targeted isolation (Figure 5 and Table 2).
5.1.1. Quantitative Evidence for a Favorable Selectivity Index
The most compelling quantitative evidence for a favorable SI derived from a Verbesina species is provided by research on V. lanata. This study evaluated compounds closely related to the genus's characteristic eudesmane scaffolds for anti-protozoal activity against Leishmania major promastigotes, yielding IC50 values ranging from 4.9 to 25.3 μM. Crucially, the toxicity against normal murine macrophages (CC50) ranged from 432.5 to 620.7 μM. This differential resulted in a high SI of up to 88.2 for Arbusculin Derivative [47]. This SI value is substantially better than that of the standard drug, meglumine antimoniate (glucantime), which had an SI of 40.0. This quantitative comparison provides definitive validation that eudesmane scaffolds can yield lead compounds with a high therapeutic window [15].
This potential for selectivity is also demonstrated qualitatively in other fractions. The EO of V. macrophylla provides a strong qualitative argument for a favorable SI. The EO is toxicologically safe (low hemolysis in human cells and LD50 > 5000 mg/kg) while simultaneously exhibiting potent antimicrobial activity (MIC 16 μg/mL against K. pneumoniae and MIC 4 μg/mL against C. albicans). The low toxicity is correlated with the known safety profile of its major components. The finding that the essential oil is a potent antimicrobial and anti-inflammatory agent with a wide toxicological safety window validates the ethnopharmacological use and suggests that the essential oil of V. macrophylla may be a basis for developing drugs with natural anti-AMR effects [39]. Similarly, strong trypanocidal activity (up to 100% elimination at 200 μg/mL) against T. cruzi combined with the confirmed toxicological safety of the essential oil of the same species strongly indicates the potential for a favorable SI [110]. Furthermore, the measured MIC of 64 μg/mL for a pure isolated SL ((1S,6R,7S,10S,Z)-6-hydroxy-10-isopropyl-7-methoxy-3,7-dimethyl-4-oxocyclodec-2-en-1-yl cinnamate) against key Gram-positive pathogens provides a critical benchmark for the efficacy component of the SI calculation [38]. This further justifies the mandatory strategy of purification and structural investigation to identify and ultimately modify these powerful lead molecules [38,55]. This mechanism of host-defense enhancement is further validated by studies on V. oerstediana, which demonstrated crucial immunomodulatory activity by stimulating NET release without significantly affecting the viability of the neutrophils (viable NETosis), suggesting an ideal pharmacological profile for developing adjunctive AMR therapies [113].
5.1.2. The Duality Challenge and Evidence for Mitigation
The core translational challenge for Verbesina SLs is their non-selective reactivity [62]. Experimental data confirms the characteristic duality of efficacy and potential toxicity inherent to the genus's metabolites [12].
Studies provide key evidence of dose-dependent biological activity. Full-strength root leachate significantly suppressed the root and shoot growth of radish seedlings (a toxicity model), demonstrating a potent, non-selective effect typical of high concentrations. Conversely, a lower, optimized concentration (1:4 dilution of root leachate with high N fertilization) actually promoted radish root growth by up to +36.5%, indicating that lower concentrations can result in beneficial (stimulatory) effects [3]. This duality confirms the complex task of optimizing the therapeutic window, as the same extract contains compounds that can both severely inhibit and promote growth, demanding precise chemical isolation [3,12]. This high performance is fundamentally linked to the extract’s dual nature: acting as allelopathic agents that inhibit seed germination, but also as potent biostimulants that significantly enhance crop growth and yield [18]. Furthermore, the prominent antioxidant activity of the extracts, with the ethyl acetate fraction showing high capacity in assays like Ferric Reducing Power, reinforces the conclusion that the total efficacy profile of Verbesina relies on the complex synergistic interaction between potent (toxic) primary metabolites (like SLs and galegine) and high-concentration support molecules (like phenolic acids and flavonoids), necessitating the targeted isolation and study of all classes for optimal therapeutic outcomes [85].
Quantitative data models this separation challenge. Potency is concentrated in the crude extract and the ethyl acetate fraction, often considered the primary source of SLs [87]. This finding is supported by the comparison demonstrating that the ethanolic extract (VE) is twice as potent as the aqueous extract (VA). The greater efficacy of the VE strongly suggests that the core bioactive agents responsible for the observed potent ROS-mediated apoptosis are concentrated within the non-polar fraction [26]. Therefore, optimizing the therapeutic window requires separating the potent efficacy-driving compounds (likely the highly reactive SLs) from the toxicity-mitigating or synergistic non-SL components (like the less-toxic triterpenoids and β-sitosterol glycosides) [106]. This process minimizes the co-extraction of inactive, highly polar compounds and potentially concentrates the therapeutic molecules away from water-soluble toxins (like the alkaloid galegine), thereby optimizing the input material for subsequent isolation and structural modification aimed at generating a more favorable SI [26].
The severity of the SI challenge is mitigated by the isolation of less-toxic fractions [106]. The low cytotoxicity found for the hydroethanolic V. turbacensis extract (IC50 of 334.6 μg/mL on HepG2) is a critical component for the future calculation of its SI. A high IC50 against a generalized cancer cell line, when compared to a low MIC, would yield a favorable SI, suggesting a low host toxicity [73]. Furthermore, the study on CDE from V. persicifolia introduces a compelling argument for the genus's potential selectivity, classifying CDE as a "mild" uncoupler. The authors stress that since V. persicifolia is widely used in Mexican traditional medicine for various complaints, administering CDE should not have dangerous toxic side-effects [62]. This high level of tolerability, coupled with robust efficacy at significantly lower doses (5 mg/kg and 10 mg/kg), strongly supports the potential for a favorable SI for the extract's neuroprotective properties [19].
5.1.3. The Need for Rigorous Validation
The overall analysis reinforces the contemporary and direct validation for the central objective of the review: to rigorously compile the necessary IC50 data and calculate the SI to bridge the gap between proven high efficacy and unassessed safety [55]. The inherent allelopathic/phytotoxic property of the raw Verbesina extract is confirmed, which directly relates to the central challenge of the SI [102]. The strong in vivo wound healing performance of V. crocata suggests that the complex chemical mixture provides a favorable overall therapeutic profile, successfully controlling the inflammatory phase while promoting organized tissue remodeling [22]. The successful valorization of Verbesina requires isolating the beneficial compounds from the potential toxins (like the guanidine alkaloid, galegine) to ensure its role as a safe, sustainable biostimulant or as a high-SI therapeutic agent [18].
5.2. Structure-Activity Relationship (SAR)
The establishment of reliable SAR for Verbesina metabolites is the indispensable prerequisite for designing less toxic, highly selective drug candidates with a favorable SI. This process mandates meticulously linking the complex C15 scaffold and its functional groups to the observed dual cytotoxic (IC50) and antimicrobial (MIC) activities.
5.2.1. Foundational Structural Requirements and Stereochemical Precision
SAR metrics are dependent on unequivocal structural gold standards, which must resolve the complex stereochemistry and conformation of the bicyclic core. Errors in stereochemistry directly translate to flawed SAR conclusions regarding which compounds possess the most favorable therapeutic index [50,55].
The cis-Decalin Core: Early structural studies confirmed the complex stereochemistry of the Verbesina sesquiterpenes. The Verbesinols were found to be stereomeric with junenol, establishing the complex, rigid cis-ring juncture of the verbesinol core [48]. The X-ray confirmation of Verbesindiol's structure was vital because it provided the definitive assignment of the cis-ring fusion and the stereochemistry of C-4 and C-6, correcting earlier ambiguous interpretations [56]. Without this foundational structural evidence, pharmacological data would be difficult to reliably map to the true molecular scaffold. This rigorous structural confirmation is a necessary precursor to conducting meaningful SAR analysis and correlating the presence of the phenolic ester moiety with the observed acute cytotoxicity (IC50) or anti-AMR efficacy (MIC) [59].
Stereochemical Corrections and Analysis: Establishing a rigorous SAR requires painstaking chemical elucidation. The definitive structure of Rupestrol was established through a series of chemical conversions, including acetylation, hydrolysis, and periodate oxidation, demonstrating the complexity required for rigorous SAR determination [53]. The use of Nuclear Overhauser Effect (NOE) experiments validated the configuration of the C-4 hydroxyl group in Verbesindiol derivatives, confirming the original assignment and contradicting proposed changes by other research groups [57]. 1H-NMR spectroscopy corrected earlier assignments for α- and β-Verbesinol esters by measuring very small coupling constants (J5,6 and J6,7), establishing that the compounds likely adopt absolute configurations where the C-7β-configuration of the isopropyl group is consistently observed [97]. The absolute configuration of Rupestrinol was determined via the Circular Dichroism (CD) curve of its derived ketone, suggesting it has the same absolute configuration as the parent compound Rupestrol [54]. The precise stereochemical determination, which confirmed the C-6 hydroxyl and C-10 methyl groups were cis and axial, is foundational for establishing accurate SAR metrics, as this axial arrangement dictates the spatial presentation of the C-6 cinnamate ester pharmacophore [20].
Conformational Flexibility: The X-ray analysis of C-4 cinnamates confirmed the existence of two crystallographically independent molecules in the crystal, differing only in the conformations of the side chains, highlighting the inherent conformational flexibility of the molecule that can impact its biological interaction [58]. The complexity of structural diversity in eudesmane derivatives can be explained by the germacrene precursor existing in different conformations, which ultimately results in eudesmane derivatives with different configurations [50]. Similarly, precise SAR analysis in the Elemanolide series (Zempoaline C) required definitive X-ray analysis due to their conformational flexibility [64]. The successful characterization of V. turbacensis eudesmanes employed two-dimensional NMR experiments, including COSY, NOESY, and HMBC, to achieve detailed structural and stereochemical assignments [52]. Furthermore, the X-ray determination of Verocephol's structure provided the necessary resolution for accurately interpreting the molecule's inherent reactivity to its observed activities [67].
5.2.2. Functional Group Dependence and Mechanistic Shift
The structural diversity of the SLs, including highly oxygenated enantio-eudesmanes like Rupestrol [53] and novel cadinene-type SLs [63], demonstrates that the core cytotoxic and antimicrobial action is highly dependent on functional group placement [47]. This precise structural work is the prerequisite for comparing the efficacy (MIC) and cytotoxicity (IC50) of different scaffolds to achieve a favorable SI [66].
The Alkylating Pharmacophore: The potential for any SL to succeed as a therapeutic agent is intimately linked to the highly reactive α-methylene-γ-lactone moiety, which acts as a template for non-reversible alkylation. The persistent co-occurrence of the cinnamate ester (the α,β-unsaturated moiety that acts as the electrophile) with other terpenoids reinforces the hypothesis that the genus's potent effects are driven by these functionalized compounds [24].
Hydroxyls, Acetyls, and Mechanism: A pivotal structure-activity study on derivatives of Compound 1 revealed that subtle structural modifications drastically alter the cellular target, even when GI50 values remain comparable [34]. Native Hydroxyls (Compound 1 and 3, Figure 4) are crucial for inducing Ca2+-dependent Mitochondrial Permeability Transition (MPT). This effect is preventable by MPT inhibitors, supporting the role of the hydroxyl groups in MPT induction. Conversely, masking the hydroxyl groups with Acetate Substituents (Compound 2 and 4, Figure 4) shifts the mechanism dramatically. These acetylated derivatives cause injury primarily by directly Inhibiting Complex II (Succinate dehydrogenase) of the mitochondrial respiratory chain and inducing an uncoupling effect on oxidative phosphorylation.
Structural Features for Activity: The biological activity of eudesmane sesquiterpenoids is strongly determined by the types, positions, and quantity of substituent groups. Anti-inflammatory activity, for instance, is significantly influenced by substitutions at pivotal positions like C-8, C-11, and C-13. An increase in the number of hydroxy substituents often correlates with an enhancement in anti-inflammatory efficacy. Furthermore, the terminal double bond between C-4 and C-15 (Δ4(15) double bond) could significantly enhance this activity [15].
5.2.3. SAR Challenges and Translational Optimization
The successful application of SAR must account for the genus's chemical complexity and the translational necessity of reducing toxicity [10].
Targeting Alkaloid SAR for Analogue Design: Design principles established for the guanidino-amide alkaloids offer a parallel path for SL modification efforts. For hypotensive activity in this class, the presence of the double bond was found to be of major importance, while the absence of methoxy groups on the aromatic ring resulted in a measurable increase in hypotensive activity [114].
Translational Strategy: A major strategy for optimizing the SI is the development of structural analogues aimed at reducing toxicity while enhancing effectiveness. This involves generating monofunctional alkylants or using chemical modifications to introduce substituents that alter the activity and safety profile [10].
The Absolute Requirement: The initial step requires establishing a reliable SI for potent SLs, which is only meaningful after meticulous chemical elucidation provides the unequivocal structural gold standard [55]. Critically, the determination of the exact stereochemistry for the eudesmane derivatives from V. turbacensis provides necessary benchmarks, as correlations drawn between potency (MIC/IC50) and subtle structural differences would be scientifically unfounded without this foundational crystallographic work [60]. The successful assembly of the 152,050 bp plastome of V. alternifolia provides a crucial genetic reference point that can help minimize the impact of geographical and chemotypic variability, supporting the goal of standardization necessary for reproducible SAR studies [16].
6. Discussion and Translational Direction
The data compiled within this review strongly validate the ethnopharmacological use of the Verbesina genus, confirming that its specialized metabolites, particularly the Sesquiterpene Lactones (SLs), provide a high-potency chemical arsenal against Antimicrobial Resistance (AMR). However, the inherent duality of efficacy and host toxicity, driven by the reactive α-methylene-γ-lactone pharmacophore defines the core translational challenge. Moving beyond promising MIC and IC50 data requires a rigorous roadmap focused on maximizing the Selectivity Index (SI) through chemical optimization, standardization, and targeted bioassay-guided fractionation.
6.1. The Imperative for Chemical Optimization and Analogue Development
The SAR analysis (Section 5.2) demonstrated that subtle changes in the SL scaffold, such as masking hydroxyl groups with acetate substituents can drastically shift the cellular mechanism, from inducing Mitochondrial Permeability Transition (MPT) to inhibiting respiratory Complex II [34]. This complexity confirms that simple isolation of the natural product is insufficient; future success hinges on advanced medicinal chemistry to modify and detoxify the highly active natural scaffolds.
Mitigating Toxicity via Semi-Synthesis: The primary goal is to mask the non-selective reactivity of the α-methylene-γ-lactone core while preserving antimicrobial efficacy. The most promising strategy is the semi-synthesis of structural analogues aimed at reducing general cytotoxicity [10]. Only compounds with a rigorously confirmed favorable SI can proceed toward this chemical optimization.
Synthetic Access and Scale-Up: For many valuable but scarce metabolites, such as the benzofuran-3-one derivative from V. luetzelburgii natural abundance is too low for comprehensive in vivo and SI testing, underscoring the necessity of a robust synthetic organic chemistry program [98]. This capability is also critical for the scale-up and optimization of the guanidino-amide alkaloid scaffolds, as evidenced by the successful total synthesis of hypotensive analogs [75,83].
6.2. Standardization and Bioassay-Guided Isolation
Translational research on Verbesina metabolites is complicated by high natural variability, requiring strict control over raw material quality and isolation protocols.
- Addressing Chemical Variability: The measured IC50 and MIC values are susceptible to ontogenetic and geographical chemical variation, as the relative percentage of key sesquiterpenes can fluctuate significantly depending on the plant’s maturity [68]. Future work must implement high-resolution analytical standards like HPLC profiling to define standardized chemotypes and minimize this variability, thereby ensuring the reproducibility of the SI calculation.
- Genetic Anchoring: The successful assembly of the complete plastome for V. alternifolia provides a crucial genetic reference point, which should be leveraged to map and minimize the impact of genetic and chemotypic variability when working with new accessions [16].
- Targeted Toxicity Assessment: Rigorous SI calculation requires toxicity profiles (IC50 on non-cancerous lines) for every potent isolate to bridge the gap between high efficacy and unassessed safety [55]. Furthermore, the scarcity of information on many ethnobotanically important species, such as V. crocata, demands comprehensive characterization of active principles and toxicity assessments to elaborate safe pharmaceutical products [23].
- Biotechnology for Consistency: To circumvent the challenges of field-collected material, the successful biosynthesis of key triterpenoids like lupeol in V. encelioides cell cultures offers a high-value pathway for the scale-up and standardization of non-toxic support molecules [94]. This difficulty has prompted translational research into novel process engineering. A specific process patent demonstrates the development of an optimized hexane extraction method for V. persicifolia that dramatically increases the yield and purity of the lead eudesmane 4CDE (up to 3 g/kg of fresh material), successfully solving the scalability challenge for this high-value scaffold [104].
7. Conclusion
The Verbesina genus is a verified repository of complex, evolved chemical defenses, offering structurally unique sesquiterpenoid scaffolds with proven dual cytotoxic and antimicrobial activities. By identifying lead compounds with a favorable Selectivity Index (SI), and leveraging modern chemical synthesis and rigorous standardization protocols, the potential of this rich Mexican medicinal resource can be fully unlocked. The future of anti-AMR drug discovery is complex, but the multifaceted chemical mechanisms and translational feasibility demonstrated by Verbesina metabolites position them as promising next-generation scaffolds in the fight against antibiotic resistance.
Author Contributions
Conceptualization, C.A.L.-R. and T.J.P.; methodology, T.J.P. and K.I.V.-H.; software, F.H.-R.; validation, C.A.L.-R., F.H.-R., and T.J.P.; formal analysis, K.I.V.-H. and T.J.P.; investigation, C.A.L.-R. and K.I.V.-H.; resources, F.R.R.-M. and F.H.-R.; data curation, C.A.L.-R. and K.I.V.-H.; writing—original draft preparation, T.J.P. and K.I.V.-H.; writing—review and editing, C.A.L.-R. and F.H.-R.; visualization, F.H.-R.; supervision, F.R.R.-M., F.H.-R., and C.A.L.-R.; project administration, F.R.R.-M.; funding acquisition, F.H.-R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Grichar, W.J.; Sestak, D.C. Control of Golden Crownbeard (Verbesina encelioides) in Peanut (Arachis hypogaea) with Postemergence Herbicides. Peanut Sci. 1998, 25, 39–43. [Google Scholar] [CrossRef]
- Panero, J.L.; Jansen, R.K.; Clevinger, J.A. Phylogenetic relationships of subtribe Ecliptinae (Asteraceae: Heliantheae) based on chloroplast DNA restriction site data. Am. J. Bot. 1999, 86, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Inderjit; Asakawa, C.; Dakshini, K.M.M. Allelopathic potential of Verbesina encelioides root leachate in soil. Can. J. Bot. 2000, 77, 1419–1424. [Google Scholar] [CrossRef]
- Chappelka, A.H.; Neufeld, H.S.; Davison, A.W.; Somers, G.L.; Renfro, J.R. Ozone Injury on Cutleaf Coneflower (Rudbeckia laciniata) and Crown-Beard (Verbesina occidentalis) in Great Smoky Mountains National Park. Environ. Pollut. 2003, 125, 53–59. [Google Scholar] [CrossRef]
- Grichar, W.J.; Jordan, D.L.; Prostko, E.P. Weed control and peanut (Arachis hypogaea L.) response to formulations of imazapic. Crop Prot. 2012, 36, 31–36. [Google Scholar] [CrossRef]
- Evans, B.; Borowicz, V. Verbesina alternifolia Tolerance to the Holoparasite Cuscuta gronovii and the Impact of Drought. Plants 2013, 2, 635–649. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Chit, E.; Carrillo-González, R.; González-Chávez, M.d.C.A.; Vibrans, H.; Yáñez-Espinosa, L.; Delgado-Alvarado, A. Riparian plants on mine runoff in Zimapan, Hidalgo, Mexico: Useful for phytoremediation? Int. J. Phytoremediation 2016, 18, 861–868. [Google Scholar] [CrossRef]
- Sharma, R.; Chandel, V.; Rishi, N. A new variant of Croton yellow vein mosaic virus naturally infecting wild sunflower in India. VirusDis. 2018, 29, 513–519. [Google Scholar] [CrossRef]
- Rivera, P.; Villaseñor, J.L.; Terrazas, T.; Panero, J.L. The importance of the Mexican taxa of Asteraceae in the family phylogeny. J. Syst. Evol. 2021, 59, 935–952. [Google Scholar] [CrossRef]
- Kaur, A.; Kaur, S.; Jandrotia, R.; Singh, H.P.; Batish, D.R.; Kohli, R.K.; Rana, V.S.; Shakil, N.A. Parthenin—A Sesquiterpene Lactone with Multifaceted Biological Activities: Insights and Prospects. Molecules 2021, 26, 5347. [Google Scholar] [CrossRef]
- Kaur, K.; Sidhu, M.C.; Ahluwalia, A.S. A Systematic Review of an Invasive Plant Species: Verbesina encelioides (Cav.) Benth. & Hook. f. Ex A. Gray. Int. J. Pharm. Sci. Res. 2022, 13, 3824–3832. [Google Scholar] [CrossRef]
- Velasco-Ramírez, A.P.; Velasco-Ramírez, A.; Hernández-Herrera, R.M.; Ceja-Esquivez, J.; Velasco-Ramírez, S.F.; Ramirez-Anguiano, A.C.; Torres-Morán, M.I. The Impact of Aqueous Extracts of Verbesina sphaerocephala and Verbesina fastigiata on Germination and Growth in Solanum lycopersicum and Cucumis sativus Seedlings. Horticulturae 2022, 8, 652. [Google Scholar] [CrossRef]
- Velasco-Ramírez, A.P.; Velasco-Ramírez, A.; Hernández-Herrera, R.M.; Murguia-Lizalde, D.M.; Velasco-Ramírez, S.F. Extractos de Verbesina sphaerocephala y Verbesina fastigiata promueven el desarrollo en maceta de Fragaria ananassa. Agroind. Sci. 2022, 12, 29–37. [Google Scholar] [CrossRef]
- Mehal, K.K.; Kaur, A.; Singh, H.P.; Batish, D.R. Investigating the phytotoxic potential of Verbesina encelioides: effect on growth and performance of co-occurring weed species. Protoplasma 2023, 260, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.-X.; Zhao, H.-Y.; Peng, C.; Liu, F.; Xiong, L. Eudesmane-type sesquiterpenoids: Structural diversity and biological activity. Heliyon 2024, 10, e35270. [Google Scholar] [CrossRef]
- Tomasello, S.; Manzo, E.; Karbstein, K. Comparative plastome assembly of the yellow ironweed (Verbesina alternifolia) using Nanopore and Illumina reads. Front. Plant Sci. 2024, 15, 1429494. [Google Scholar] [CrossRef]
- Mehal, K.K.; Sharma, A.; Kaur, A.; Kalia, N.; Kohli, R.K.; Singh, H.P.; Batish, D.R. Modelling the ecological impact of invasive weed Verbesina encelioides on vegetation composition across dryland ecosystems of Punjab, northwestern India. Environ. Monit. Assess. 2023, 195, 725. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Pérez, A.R.; Hernández-Herrera, R.M.; Velasco-Ramírez, S.F.; Velasco-Ramírez, A.; Ramírez-Anguiano, A.C.; Velasco-Ramírez, A.P. Effect of Botanical Extracts from Verbesina sphaerocephala on the Growth and Yield of Tomato Under Greenhouse Conditions. Acta Agrobot. 2025, 78, 201390. [Google Scholar] [CrossRef]
- Verma, V.; Rao, L.; Chaudhary, M.; Jain, S.; Pinapati, K.K.; Dey, A.; Sharma, S.; Srivastava, N. Spirilloxanthin from Verbesina encelioides Flower Extract has Neuroprotective Benefits in Scopolamine-Induced Memory Impaired Rats that Resemble Alzheimer’s Disease. Mol. Neurobiol. 2025, 62, 12602–12615. [Google Scholar] [CrossRef] [PubMed]
- Herz, W.; Kumar, N. Aromatic and other constituents of four Verbesina species: Structure and stereochemistry of verbesindiol. Phytochemistry 1981, 20, 247–250. [Google Scholar] [CrossRef]
- Mora, F.D.; Alpan, L.; McCracken, V.J.; Nieto, M. Chemical and Biological Aspects of the Genus Verbesina. Nat. Prod. J. 2013, 3, 140–150. [Google Scholar] [CrossRef]
- García-Bores, A.M.; Álvarez-Santos, N.; López-Villafranco, M.E.; Jácquez-Ríos, M.P.; Aguilar-Rodríguez, S.; Grego-Valencia, D.; Espinosa-González, A.M.; Estrella-Parra, E.A.; Hernández-Delgado, C.T.; Serrano-Parrales, R.; González-Valle, M. R.; Benítez-Flores, J. C. Verbesina crocata: A Pharmacognostic Study for the Treatment of Wound Healing. Saudi J. Biol. Sci. 2020, 27, 3113–3124. [Google Scholar] [CrossRef]
- Heyerdahl-Viau, I.; López-Naranjo, F.; Córdova-Moreno, R.; Urbina-Pastrana, A.; Martínez-Núñez, J.M. A Brief Review of Verbesina crocata (Cav.) Less., a Scantly Studied Medicinal Plant. Res. J. Pharmacogn. 2023, 10, 51–56. [Google Scholar] [CrossRef]
- Bohlmann, F.; Zdero, C.; Robinson, H.; King, R.M. Two New Eudesmane Cinnamates from Verbesina sordescens. Phytochemistry 1982, 21, 1663–1664. [Google Scholar] [CrossRef]
- Albalawi, M.A.D.; Bashir, N.A.O.; Tawfik, A. Anticancer and Antifolate Activities of Extracts of Six Saudi Arabian Wild Plants Used in Folk Medicine. J. Life Sci. 2015, 9, 334–340. [Google Scholar] [CrossRef]
- Farshori, N.N. Verbesina encelioides-Induced Cytotoxicity and Mitochondria-Mediated Apoptosis in Human Colon Cancer Cells through ROS Generation. Biologia 2021, 76, 2711–2720. [Google Scholar] [CrossRef]
- Oelrichs, P.B.; Vallely, P.J.; MacLeod, J.K.; Lewis, I.A.S. Isolation of Galegine from Verbesina enceloiodes. J. Nat. Prod. 1981, 44, 754–755. [Google Scholar] [CrossRef]
- Indrayanto, G.; Putra, G.S.; Suhud, F. Chapter Six - Validation of in-vitro bioassay methods: Application in herbal drug research. In Profiles of Drug Substances, Excipients and Related Methodology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 46, pp. 273–307. [Google Scholar] [CrossRef]
- Villaseñor, J.L.; Ortiz, E. La Familia Asteraceae en la Flora del Bajío y de Regiones Adyacentes. Acta Bot. Mex. 2012, 100, 259–291. [Google Scholar] [CrossRef]
- Van Auken, O.W.; Leonard, W.J. Why is Verbesina virginica (Frostweed, Asteraceae) Not Found in Grasslands? Phytologia 2016, 98, 76–88. https://www.phytologia.org/uploads/2/3/4/2/23422706/98_1_76-88vanauken_verbesina_in_grasslands_rev_12-29-15.pdf.
- Kaur, K.; Ramanpreet; Gupta, R.C.; Kumari, S. Cyto-Morphological Studies of Some Dicot Plants from Rajasthan (India). Cytologia 2015, 80, 353–362. [Google Scholar] [CrossRef]
- Domínguez-Barradas, C.; Cruz-Morales, G.E.; González-Gándara, C. Plantas de uso medicinal de la Reserva Ecológica "Sierra de Otontepec", municipio de Chontla, Veracruz, México. CienciaUAT 2015, 9, 41–52. [Google Scholar] [CrossRef]
- Perez G., R.M.; Perez G., S.; Zavala, M.A.; Perez G., S.C. Effect of Agarista mexicana and Verbesina persicifolia on Blood Glucose Level of Normoglycaemic and Alloxan-diabetic Mice and Rats. Phytother. Res. 1996, 10, 351–353. [Google Scholar] [CrossRef]
- Dalla Via, L.; Mejia, M.; García-Argáez, A.N.; Braga, A.; Toninello, A.; Martínez-Vázquez, M. Anti-Inflammatory and Antiproliferative Evaluation of 4β-Cinnamoyloxy,1β,3α -Dihydroxyeudesm-7,8-ene from Verbesina persicifolia and Derivatives. Bioorg. Med. Chem. 2015, 23, 5816–5828. [Google Scholar] [CrossRef]
- Martínez Alfaro, M.Á.; Evangelista Oliva, V.; Mendoza Cruz, M.; Morales Garcia, G.; Toledo Olazcoaga, G.; Wong León, A. Catálogo de plantas útiles de la Sierra Norte de Puebla, México. Cuadernos IBUNAM 27. Universidad Nacional Autónoma de México, Instituto de Biología, 2001. https://cepe-taxco.bibliotecas.unam.mx:84/cgi-bin/koha/opac-detail.pl?biblionumber=2513.
- Rodríguez-Valdovinos, K.Y.; Salgado-Garciglia, R.; Vázquez-Sánchez, M.; Álvarez-Bernal, D.; Oregel-Zamudio, E.; Ceja-Torres, L.F.; Medina-Medrano, J.R. Quantitative Analysis of Rutin by HPTLC and In Vitro Antioxidant and Antibacterial Activities of Phenolic-Rich Extracts from Verbesina sphaerocephala. Plants 2021, 10, 475. [Google Scholar] [CrossRef]
- López-Rosas, C.A.; González-Periañez, S.; Pawar, T.J.; Zurutuza-Lorméndez, J.I.; Ramos-Morales, F.R.; Olivares-Romero, J.L.; Saavedra Vélez, M.V.; Hernández-Rosas, F. Anticonvulsant Potential and Toxicological Profile of Verbesina persicifolia Leaf Extracts: Evaluation in Zebrafish Seizure and Artemia salina Toxicity Models. Plants 2025, 14, 1078. [Google Scholar] [CrossRef]
- Mora, F.D.; Alpan, L.; De Tommasi, N.; McCracken, V.J.; Nieto, M. New Antibacterial Germacrene from Verbesina negrensis. Planta Med. 2013, 79, 707–710. [Google Scholar] [CrossRef]
- Veras, B.O.d.; Oliveira, J.R.S.d.; Lima, V.L.M.d.M.; Navarro, D.M.d.A.F.; Aguiar, J.C.R.d.O.F.d.; Mourad, G.M.d.M.; Silva, J.W.d.; Assis, C.R.D.d.; Gorlach-Lira, K.; Assis, P.A.C.d.; Barbosa, J.I.d.S.; Melo, M.R.C.S.d.; Oliveira, M.B.M.d.; Silva, M.V.d.; Lopes, A.C.S. The essential oil of the leaves of Verbesina macrophylla (Cass.) S.F.Blake has antimicrobial, anti-inflammatory and antipyretic activities and is toxicologically safe. J. Ethnopharmacol. 2021, 265, 113248. [Google Scholar] [CrossRef]
- Al-Sodany, Y.M.; Issa, A.A.; Kahil, A.A.; Ali, E.F. Diversity of Soil Cyanobacteria in Relation to Dominant Wild Plants and Edaphic Factors at Western Saudi Arabia. Annu. Res. Rev. Biol. 2018, 26, 1–14. [Google Scholar] [CrossRef]
- Essinger, J.F., Jr.; Gan, L.; Behrens, N.J. Stable Liquid Formulations and Methods of Using the Same. U.S. Patent Application 20200260722 A1, 20 August 2020. https://portal.unifiedpatents.com/patents/patent/US-11109588-B2.
- Grichar, W.J.; Sestak, D.C. Herbicide Systems for Golden Crownbeard (Verbesina encelioides) Control in Peanut. Peanut Sci. 2000, 27, 23–26. [Google Scholar] [CrossRef]
- Van Nuland, M.E.; Haag, E.N.; Bryant, J.A.M.; Read, Q.D.; Klein, R.N.; Douglas, M.J.; Gorman, C.E.; Greenwell, T.D.; Busby, M.W.; Collins, J.; LeRoy, J.T.; Schuchmann, G.; Schweitzer, J.A.; Bailey, J.K. Fire Promotes Pollinator Visitation: Implications for Ameliorating Declines of Pollination Services. PLoS ONE 2013, 11, e79853. [Google Scholar] [CrossRef] [PubMed]
- Aschenbrenner, A.-K.; Horakh, S.; Spring, O. Linear Glandular Trichomes of Helianthus (Asteraceae): Morphology, Localization, Metabolite Activity and Occurrence. AoB PLANTS 2013, 5, plt028. [Google Scholar] [CrossRef]
- Sultana, S.; Ali, M.; Mir, S.R. Chemical Constituents From the Aerial Roots of Ficus benghalensis L., Leaves of Nyctanthes arbor-tristis L. and Roots of Verbesina encelioides (Cav.) Benth. et Hook. f. J. Pharm. Biosci. 2018, 6, 16–26. [Google Scholar] [CrossRef]
- Bonchev, G.N.; Vassilevska-Ivanova, R. Fingerprinting the genetic variation and intergeneric hybrid dynamics in the family Asteraceae (genera Helianthus, Echinaceae, Tagetes and Verbesina) using iPBS markers. Biologia 2019, 75, 457–464. [Google Scholar] [CrossRef]
- Wu, Q.-X.; Shi, Y.-P.; Jia, Z.-J. Eudesmane Sesquiterpenoids from the Asteraceae Family. Nat. Prod. Rep. 2006, 23, 699–734. [Google Scholar] [CrossRef]
- Gardner, P.D.; Park, G.J.; Albers, C.C. α- and β-Verbesinol. Sesquiterpene Alcohols of the cis-Decalin Series. J. Am. Chem. Soc. 1961, 83, 1511–1512. [Google Scholar] [CrossRef]
- Bohlmann, F.; Lonitz, M. Neue Eudesman-Derivate und andere Sesquiterpene aus Verbesina-Arten [New Eudesmane Derivatives and Other Sesquiterpenes from Verbesina Species]. Chem. Ber. 1978, 111, 254–263. [Google Scholar] [CrossRef]
- Jakupovic, J.; Ellmauerer, E.; Jia, Y.; Bohlmann, F.; Dominguez, X.A.; Schmeda-Hirschmann, G. Further Eudesmane Derivatives from Verbesina Species. Planta Med. 1987, 53, 39–42. [Google Scholar] [CrossRef]
- Bohlmann, F.; Grenz, M.; Gupta, R.K.; Dhar, A.K.; Ahmed, M.; King, R.M.; Robinson, H. Eudesmane Derivatives from Verbesina Species. Phytochemistry 1980, 19, 2391–2397. [Google Scholar] [CrossRef]
- Amaro-Luis, J.M.; Ramírez, I.; Delgado-Méndez, P.; Jorge, Z.D. Eudesmane Derivatives from Verbesina turbacensis. J. Braz. Chem. Soc. 2002, 13, 352–357. [Google Scholar] [CrossRef]
- Box, V.G.S.; Chan, W.R.; Taylor, D.R. The Structure of Rupestrol a New Sesquiterpenoid from Verbesina rupestris (Urb.) Blake. Tetrahedron Lett. 1971, 12, 4371–4374. [Google Scholar] [CrossRef]
- Box, V.G.S.; Bardouille, V.; Chan, W.R. Enantio-Eudesmane Sesquiterpenes from Verbesina rupestris. Phytochemistry 1977, 16, 987–990. [Google Scholar] [CrossRef]
- Ramseyer, J.; Thuerig, B.; De Mieri, M.; Schärer, H.-J.; Oberhänsli, T.; Gupta, M.P.; Tamm, L.; Hamburger, M.; Potterat, O. Eudesmane Sesquiterpenes from Verbesina lanata with Inhibitory Activity against Grapevine Downy Mildew. J. Nat. Prod. 2017, 80, 3296–3304. [Google Scholar] [CrossRef]
- Herz, W.; Kumar, N.; Blount, J.F. Crystal Structure and Stereochemistry of Verbesindiol. J. Org. Chem. 1982, 47, 1785–1786. [Google Scholar] [CrossRef]
- Banerjee, S.; Jakupovic, J.; Bohlmann, F.; King, R.M.; Robinson, H. A Rearranged Eudesmane and Further Verbesindiol Derivatives from Verbesina eggersii. Phytochemistry 1985, 24, 1106–1108. [Google Scholar] [CrossRef]
- Martínez, M.; de Vivar, A.R.; Ortega, A.; Quintero, M. de L.; García, C.; Fronczek, F.R. Eudesmane Triols from Verbesina virgata. Phytochemistry 1983, 22, 979–982. [Google Scholar] [CrossRef]
- Xu, W.H.; Liang, Q.; Liang, Z.S.; Li, X.C. Two New Eudesmane Derivatives from Verbesina virginica. Chin. Chem. Lett. 2010, 21, 693–695. [Google Scholar] [CrossRef]
- Bruno-Colmenárez, J.; Amaro-Luis, J.M.; Ramírez, I.; Delgado-Méndez, P.; Díaz de Delgado, G. Absolute Configuration of Two Eudesmane Derivatives from Verbesina turbacensis H.B.K. J. Chem. Crystallogr. 2010, 40, 110–115. [Google Scholar] [CrossRef]
- Vazquez Cadanedo, A.; Díaz-T, E.; Jankowski, C.K.; Dako, E.; Doiron, J.; Jiménez-E, M. On the Structure of Triolcinnamate from Verbesina persicifolia. Indian J. Chem. B 2015, 54, 1342–1345. https://www.researchgate.net/publication/283489292_On_the_structure_of_triolcinnamate_from_Verbesina_persicifolia.
- Dalla Via, L.; García-Argáez, A.N.; Braga, A.; Martínez-Vázquez, M.; Grancara, S.; Martinis, P.; Agostinelli, E.; Toninello, A. An Eudesman Derivative from Verbesina persicifolia D.C. as a Natural Mild Uncoupler in Liver Mitochondria. A New Potential Anti-obesity Agent? Curr. Pharm. Des. 2014, 20, 253–261. [Google Scholar] [CrossRef]
- Arciniegas, A.; Pérez-Castorena, A.L.; Villaseñor, J.L.; Romo de Vivar, A. Cadinenes and other metabolites from Verbesina sphaerocephala A. Gray. Biochem. Syst. Ecol. 2020, 93, 104183. [Google Scholar] [CrossRef]
- Ortega, A.; Maldonado, E.; Fronczek, F.R.; Delord, T.J.; Chiari, G. Elemanolides from Verbesina seattonii. Phytochemistry 1985, 24, 1755–1760. [Google Scholar] [CrossRef]
- Selener, M.G.; Elso, O.; Grosso, C.; Borgo, J.; Clavin, M.; Malchiodi, E.L.; Cazorla, S.I.; Flavia, F.; Sülsen, V.P. Anti-Trypanosoma cruzi Activity of Extracts from Argentinean Asteraceae Species. Iran. J. Pharm. Res. 2019, 18, 1854–1861. [Google Scholar] [CrossRef]
- Bohlmann, F.; Lonitz, M. Neue Sesquiterpene aus Verbesina occidentalis. Phytochemistry 1978, 17, 453–455. [Google Scholar] [CrossRef]
- Salmón, M.; Soriano-Garcia, M.; Toscano, R.A.; Cárdenas, J.; Miranda, R.; Vargas, F.; Angeles, E. Verocephol, a Unique Amorphane Sesquiterpene γ-Lactol. J. Org. Chem. 1985, 50, 4171–4172. [Google Scholar] [CrossRef]
- Albuquerque, M.R.J.R.; Canuto, K.M.; Pessoa, O.D.L.; Nunes, E.P.; Nascimento, R.F.; Silveira, E.R. Essential Oil Composition of Verbesina diversifolia DC. Flavour Fragr. J. 2006, 21, 634–636. [Google Scholar] [CrossRef]
- Bezerra, L.D.A.; Mangabeira, P.A.O.; de Oliveira, R.A.; Costa, L.C.D.B.; Da Cunha, M. Leaf Blade Structure of Verbesina macrophylla (Cass.) F. S. Blake (Asteraceae): Ontogeny, Duct Secretion Mechanism and Essential Oil Composition. Plant Biol. 2018, 20, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Mora, F.D.; Rojas, Y.L.; González, V.; Velasco, J.; Díaz, T.; Ríos, N.; Rojas-Fermin, L.B.; Carmona, J.; Silva, B.; Nieto, M. Chemical Composition and in vitro Antibacterial Activity of the Essential Oil of Verbesina negrensis from the Venezuelan Andes. Nat. Prod. Commun. 2015, 10, 1309–1310. [Google Scholar] [CrossRef]
- Box, V.G.S.; Chan, W.R. Terpenoids from Verbesina rupestris. Phytochemistry 1975, 14, 583. [Google Scholar] [CrossRef]
- Ogungbe, I.V.; Crouch, R.A.; Haber, W.A.; Setzer, W.N. Phytochemical Investigation of Verbesina turbacensis Kunth: Trypanosome Cysteine Protease Inhibition by (-)-Bornyl Esters. Nat. Prod. Commun. 2010, 5, 1161–1166. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Gallardo, K.; Alvarez-Ortega, N.; Olivero-Verbel, J. Cytotoxicity of Nine Medicinal Plants from San Basilio de Palenque (Colombia) on HepG2 Cells. Plants 2023, 12, 2686. [Google Scholar] [CrossRef]
- Fawzy, M.A.; Hifney, A.F.; Issa, A.A.; Gareib, G. Phytochemical Constituents and Allelopathic Effects of Some Medicinal Plants Extract on the Soil Algal Diversity. J. Agric. Sci. Technol. A 2013, 3, 1000–1009. https://qikan.cqvip.com/Qikan/Article/Detail?id=48303743&from=Qikan_Search_Index.
- Botta, B.; Carmignani, M.; Volpe, A.R.; Botta, M.; Corelli, F.; Delle Monache, G. Novel Hypotensive Agents from Verbesina Caracasana: Structure, Synthesis and Pharmacology. Curr. Med. Chem. 2003, 10, 1845–1862. [Google Scholar] [CrossRef]
- Eichholzer, J.V.; Lewis, I.A.S.; MacLeod, J.K.; Oelrichs, P.B.; Vallely, P.J. Galegine and a new dihydroxyalkylacetamide from Verbesina enceloiodes. Phytochemistry 1982, 21, 97–99. [Google Scholar] [CrossRef]
- Delle Monache, G.; Delle Monache, F.; Carmignani, M.; Bonnevaux, S.C.; Espinal, R.; De Luca, C.; Botta, B. Guanidine Derivatives Having Hypotensive Activity, Composition Containing Them and Process for Obtaining Them. U.S. Patent 5,059,624, 22 October 1991. https://worldwide.espacenet.com/patent/search/family/011261768/publication/EP0330629A2?q=EP330629A2.
- Delle Monache, G.; Botta, B.; Delle Monache, F.; Espinal, R.; De Bonnevaux, S.C.; De Luca, C.; Botta, M.; Corelli, F.; Dei, D.; Gacs-Baitz, E.; Carmignani, M. Caracasandiamide, a Truxinic Hypotensive Agent from Verbesina caracasana. Bioorg. Med. Chem. Lett. 1996, 6, 233–238. [Google Scholar] [CrossRef]
- Delle Monache, G.; Botta, B.; Delle Monache, F.; Espinal, R.; De Bonnevaux, S.C.; De Luca, C.; Botta, M.; Corelli, F.; Carmignani, M. Caracasanamide, a Novel Hypotensive Agent from Verbesina caracasana. Bioorg. Med. Chem. Lett. 1992, 2, 415–418. [Google Scholar] [CrossRef]
- Delle Monache, G.; Botta, B.; Delle Monache, F.; Espinal, R.; De Bonnevaux, S.C.; De Luca, C.; Botta, M.; Corelli, F.; Carmignani, M. Novel Hypotensive Agents from Verbesina caracasana. 2. Synthesis and Pharmacology of Caracasanamide. J. Med. Chem. 1993, 36, 2956–2963. [Google Scholar] [CrossRef]
- Carmignani, M.; Volpe, A.R.; Delle Monache, F.; Botta, B.; Espinal, R.; De Bonnevaux, S.C.; De Luca, C.; Botta, M.; Corelli, F.; Tafi, A.; Ripanti, G.; Delle Monache, G. Novel Hypotensive Agents from Verbesina caracasana. 6. Synthesis and Pharmacology of Caracasandiamide. J. Med. Chem. 1999, 42, 3116–3125. [Google Scholar] [CrossRef] [PubMed]
- Delle Monache, G.; Volpe, A.R.; Delle Monache, F.; Vitali, A.; Botta, B.; Espinal, R.; De Bonnevaux, S.C.; De Luca, C.; Botta, M.; Corelli, F.; Camignani, M. Further Hypotensive Metabolites from Verbesina caracasana. Bioorg. Med. Chem. Lett. 1999, 9, 3249–3254. [Google Scholar] [CrossRef]
- Carmignani, M.; Volpe, A.R.; Botta, B.; Espinal, R.; De Bonnevaux, S.C.; De Luca, C.; Botta, M.; Corelli, F.; Tafi, A.; Sacco, R.; Delle Monache, G. Novel Hypotensive Agents from Verbesina caracasana. 8. Synthesis and Pharmacology of (3,4-Dimethoxycinnamoyl)-N1-agmatine and Synthetic Analogs. J. Med. Chem. 2001, 44, 2950–2958. [Google Scholar] [CrossRef]
- Compagnone, R.S.; Bermudez, J.; Ibáñez, G.; Díaz, B.; Garrido, M.R.; Israel, A.; Suárez, A.I. New Guanidine Alkaloids from the Leaves of Verbesina peraffinis. Nat. Prod. Commun. 2008, 3, 511–514. [Google Scholar] [CrossRef]
- Rodríguez-Valdovinos, K.Y.; Salgado-Garciglia, R.; Hernández-García, A.; Saavedra-Molina, A.; del Río-Torres, R.E.N.; López-Meza, J.E.; Monribot-Villanueva, J.L.; Guerrero-Analco, J.A.; Medina-Medrano, J.R. Antioxidant and Antifungal Activities and Characterization of Phenolic Compounds Using Ultra-High Performance Liquid Chromatography and Mass Spectrometry (UPLC-MS) of Aqueous Extracts and Fractions from Verbesina sphaerocephala Stems. Plants 2024, 13, 2791. [Google Scholar] [CrossRef]
- Glennie, C.W.; Jain, S.C. Flavonol 3,7-Diglycosides of Verbesina enceloiodes. Phytochemistry 1980, 19, 157–158. [Google Scholar] [CrossRef]
- Abbas, F.A.; El Sayed, Z.I.; Dora, G.A.; Ateya, A.-M.; Samy, S. Phytochemical and Biological Studies of Verbesina encelioides (Cav.) Benth. and Hook. Asian J. Phytomed. Clin. Res. 2016, 4, 108–120. http://www.ajpcrjournal.com/article/PHYTOCHEMICAL%20AND%20BIOLOGICAL%20STUDIES%20OF%20VERBESINA_ENCELIOIDES%20(CAV.)%20BENTH%20AND%20HOOK.pdf.
- Wagner, H.; Iyengar, M.A.; Seligmann, O.; Herz, W. Rhamnocitrin-3-Glucuronid in Verbesina myricephala. Phytochemistry 1974, 13, 493–494. [Google Scholar] [CrossRef]
- Bohlmann, F.; Zdero, C. Neue Terpen-Inhaltsstoffe aus Verbesina-Arten. Phytochemistry 1976, 15, 1310–1311. [Google Scholar] [CrossRef]
- Farber, C.; Sanchez, L.; Kurouski, D. Confirmatory Non-Invasive and Non-Destructive Identification of Poison Ivy Using a Hand-Held Raman Spectrometer. RSC Adv. 2020, 10, 21530–21534. [Google Scholar] [CrossRef]
- Burkey, K.O.; Neufeld, H.S.; Souza, L.; Chappelka, A.H.; Davison, A.W. Seasonal Profiles of Leaf Ascorbic Acid Content and Redox State in Ozone-Sensitive Wildflowers. Environ. Pollut. 2006, 143, 427–434. [Google Scholar] [CrossRef]
- Verma, V.; Chaudhary, M.; Srivastava, N. Antioxidative Properties of Isolated Saponins of Verbesina encelioides (Cav.) Benth. & Hook. f. ex Gray and SEM Studies of Synthesized Green Nanoparticles for Acne Management. Plant Sci. Today 2019, 6, 575–582. [Google Scholar] [CrossRef]
- Xu, W.-H.; Jacob, M.R.; Agarwal, A.K.; Clark, A.M.; Liang, Z.-S.; Li, X.-C. Verbesinosides A−F, 15,27-Cyclooleanane Saponins from the American Native Plant Verbesina virginica. J. Nat. Prod. 2009, 72, 1022–1027. [Google Scholar] [CrossRef] [PubMed]
- Cerda-Garcia-Rojas, C.M.; Zamorano, G.; Chávez, M.I.; Catalán, C.A.N.; Joseph-Nathan, P. 1H and 13C NMR Study of Copteroside E Derivatives. Magn. Reson. Chem. 2000, 38, 494–499. [Google Scholar] [CrossRef]
- Singh, R.; Jain, R. Biopotentialities of Verbesina encelioides Cell Cultures. Indian J. Plant Physiol. 2009, 13, 224–230. https://www.semanticscholar.org/paper/BIOPOTENTIALITIES-OF-VERBESINA-ENCELIOIDES-CELL-Singh-Jain/cfc158b31d2d1dd7db47d5690a8c583b0f788dc6.
- Christensen, L.P.; Lam, J. Acetylenes and Related Compounds in Heliantheae. Phytochemistry 1991, 30, 11–49. [Google Scholar] [CrossRef]
- Bohlmann, F.; Zdero, C. Zur konfiguration von α- und β-Verbesinolcumarat. Phytochemistry 1979, 18, 1751–1752. [Google Scholar] [CrossRef]
- Pergomet, J.L.; Larghi, E.L.; Kaufman, T.S.; Bracca, A.B.J. First Total Synthesis of the Only Known 2-Isopropyliden-2H-Benzofuran-3-one Isolated from V. luetzelburgii. RSC Adv. 2017, 7, 5242–5250. [Google Scholar] [CrossRef]
- Divya Ramakrishnan, C.K.; Doss, D.; Vijayabharathi, A. Biochemical and Antimicrobial Characterization of an Underexploited Medicinal Plant Verbesina encelioides. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3407–3416. [Google Scholar] [CrossRef]
- Sindhu, R.K.; Vasudeva, N.; Sharma, S.K. Pharmacognostical and Preliminary Phytochemical Investigations on Verbesina encelioides Benth. Roots. J. Herb. Med. Toxicol. 2010, 4, 113–118. https://www.researchgate.net/publication/239524726_Pharmacognostical_and_preliminary_phytochemical_investigations_on_Verbesina_encelioides_Benth_Roots.
- Kushwaha, H.B.; Malik, C.P. Nanofabrication of Silver Nanoparticles From the Stem and Leaf Extract of Verbesina encelioides. Natl. Acad. Sci. Lett. 2012, 35, 555–563. [Google Scholar] [CrossRef]
- Ramírez-Herrera, A.D.; Ceja-Buenrostro, J. de J.; Miranda-Torres, A.; Reynoso-Marín, F.J. Verbesina crocata como agente biosintético de nanopartículas de plata/cloruro de plata y su evaluación en plántulas de rábano. Tópicos Investig. Cienc. Tierra Mater. 2025, 12, 38–45. [Google Scholar] [CrossRef]
- Shaffer, C.V.; Cai, S.; Peng, J.; Robles, A.J.; Hartley, R.M.; Powell, D.R.; Du, L.; Cichewicz, R.H.; Mooberry, S.L. Texas Native Plants Yield Compounds with Cytotoxic Activities against Prostate Cancer Cells. J. Nat. Prod. 2016, 79, 531–540. [Google Scholar] [CrossRef]
- Universidad Nacional Autónoma de México. Process for obtaining eudesmane sesquiterpene 4a-cinnamoiloxy-1b,3a-dihydroxyeudesm-7,8-eno(4CDE). MX 2013014284 A. 5 June 2015. https://worldwide.espacenet.com/patent/search/family/054261219/publication/MX2013014284A?q=MX2013014284A.
- Al-Oqail, M.M.; Siddiqui, M.A.; Al-Sheddi, E.S.; Saquib, Q.; Musarrat, J.; Al-Khedhairy, A.A.; Farshori, N.N. Verbesina encelioides: Cytotoxicity, Cell Cycle Arrest, and Oxidative DNA Damage in Human Liver Cancer (HepG2) Cell Line. BMC Complement. Altern. Med. 2016, 16, 126. [Google Scholar] [CrossRef]
- Ezzat, S.M.; Salama, M.M.; Mahrous, E.A.; Maes, L.; Pan, C.-H.; Abdel-Sattar, E. Antiprotozoal Activity of Major Constituents from the Bioactive Fraction of Verbesina encelioides. Nat. Prod. Res. 2016, 31, 676–680. [Google Scholar] [CrossRef]
- Abdel-Wahab, A.; Abdel-Maogood, S.Z.; Mahdy, O.A. Scanning Electron Microscopic Changes in Habronema muscae after In Vitro Exposure to Plant Extract (Verbesina alternifolia). J. Egypt. Soc. Parasitol. 2017, 47, 47–54. https://jesp.journals.ekb.eg/article_77965_1c54bbae5f06ac169e35f7a1a9176f73.pdf. [CrossRef]
- Bhati-Kushwaha, H.; Malik, C.P. Biopotential of Verbesina encelioides (Stem and Leaf Powders) in Silver Nanoparticle Fabrication. Turk. J. Biol. 2013, 37, 645–654. [Google Scholar] [CrossRef]
- Wankhade, S.; Rai, M.K. A Product and Process of Extraction of Asteraceae Plant Material for Treatment of Dermatophyte Infections. IN 3395/MUM/2010. 14 December 2010. https://patentscope.wipo.int/search/en/detail.jsf?docId=IN211588671.
- Bezerra, L.A.; Carvalho, L.P.; Mello, É. de O.; Taveira, G.B.; de Melo, E.J.T.; Gomes, V.M.; da Cunha, M. Fungicidal and trypanocidal activity of aqueous extracts of leaves and inflorescences of Verbesina macrophylla (Asteraceae). Acta Sci. Biol. Sci. 2024, 46, e71523. [Google Scholar] [CrossRef]
- Oka, Y. Nematicidal Activity of Verbesina encelioides against the Root-Knot Nematode Meloidogyne javanica and Effects on Plant Growth. Plant Soil 2012, 355, 311–322. [Google Scholar] [CrossRef]
- Lahlali, M.; Manaut, N.; Loudiki, M.; Douma, M. First Assessment of the Anti-Cyanobacterial Potentialities of the Invasive Weed Verbesina encelioides Against Microcystis aeruginosa Growth. Desalination Water Treat. 2023, 310, 150–156. [Google Scholar] [CrossRef]
- Yaseen, R.; Branitzki-Heinemann, K.; Moubasher, H.; Setzer, W.N.; Naim, H.Y.; von Köckritz-Blickwede, M. In Vitro Testing of Crude Natural Plant Extracts from Costa Rica for Their Ability to Boost Innate Immune Cells against Staphylococcus aureus. Biomedicines 2017, 5, 40. [Google Scholar] [CrossRef] [PubMed]
- Corelli, F.; Dei, D.; Delle Monache, G.; Botta, B.; De Luca, C.; Carmignani, M.; Volpe, A.R.; Botta, M. Synthesis and Preliminary Pharmacological Evaluation of Analogues of Caracasanamide, a Hypotensive Natural Product. Bioorg. Med. Chem. Lett. 1996, 6, 653–658. [Google Scholar] [CrossRef]
- Sindhu, R.K.; Vasudeva, N.; Sharma, S.K. Hypolipidemic Potential of Ethanolic Extract of Verbesina encelioides Benth. Roots. Acta Pol. Pharm. 2011, 68, 301–304. https://www.ptfarm.pl/wydawnictwa/czasopisma/acta-poloniae-pharmaceutica/110/-/13479.
- Banasthali University. Anti-Termite Herbal Composition And Process For Preparation Thereof. IN 2010/MUM/3395. 14 December 2017. https://patentimages.storage.googleapis.com/33/1b/a1/98dbb5da2bfd4d/WO2011027362A2.pdf.
- Carr, M.E.; Bagby, M.O. Tennessee Plant Species Screened for Renewable Energy Sources. Econ. Bot. 1987, 41, 78–85. [Google Scholar] [CrossRef]
Figure 1.
The Duality Challenge and Translational Road Map.
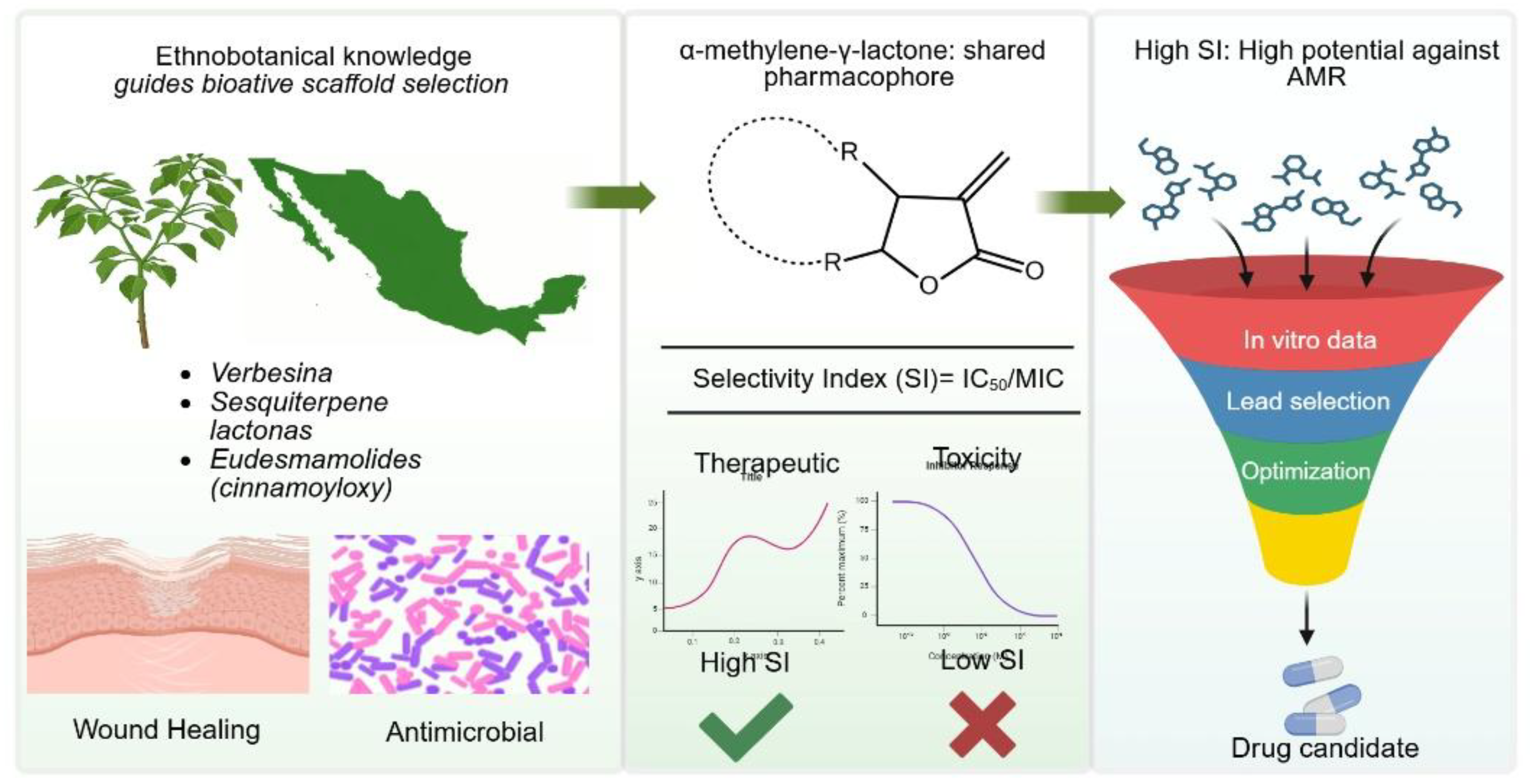
Figure 2.
Core SL Skeletons and Pharmacophore.
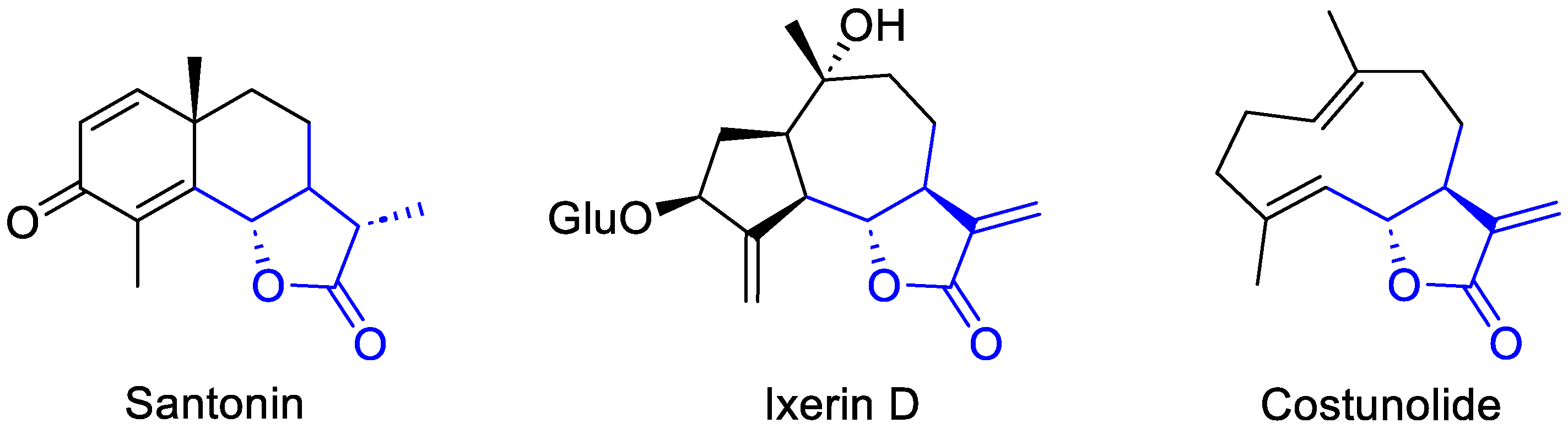
Figure 3.
Dual molecular mechanisms of action of Verbesina sesquiterpene lactones. The α-methylene-γ-lactone pharmacophore mediates distinct biological effects depending on the cellular target. (A) Bacterial cell: sesquiterpene lactones penetrate and disrupt the bacterial membrane, alkylate thiol-containing proteins via Michael-type addition, cause enzyme inactivation and ion leakage, and ultimately lead to loss of metabolic homeostasis and bacterial cell death. (B) Mammalian cell: intracellular accumulation of sesquiterpene lactones promotes reactive oxygen species (ROS) overproduction, mitochondrial dysfunction, cytochrome c release, caspase activation, and apoptosis, resulting in host cell cytotoxicity. This mechanistic asymmetry underlies the therapeutic duality of these compounds and highlights the importance of the selectivity index (SI = IC₅₀/MIC) for translational feasibility.Principio del formularioFinal del formulario.
Figure 3.
Dual molecular mechanisms of action of Verbesina sesquiterpene lactones. The α-methylene-γ-lactone pharmacophore mediates distinct biological effects depending on the cellular target. (A) Bacterial cell: sesquiterpene lactones penetrate and disrupt the bacterial membrane, alkylate thiol-containing proteins via Michael-type addition, cause enzyme inactivation and ion leakage, and ultimately lead to loss of metabolic homeostasis and bacterial cell death. (B) Mammalian cell: intracellular accumulation of sesquiterpene lactones promotes reactive oxygen species (ROS) overproduction, mitochondrial dysfunction, cytochrome c release, caspase activation, and apoptosis, resulting in host cell cytotoxicity. This mechanistic asymmetry underlies the therapeutic duality of these compounds and highlights the importance of the selectivity index (SI = IC₅₀/MIC) for translational feasibility.Principio del formularioFinal del formulario.
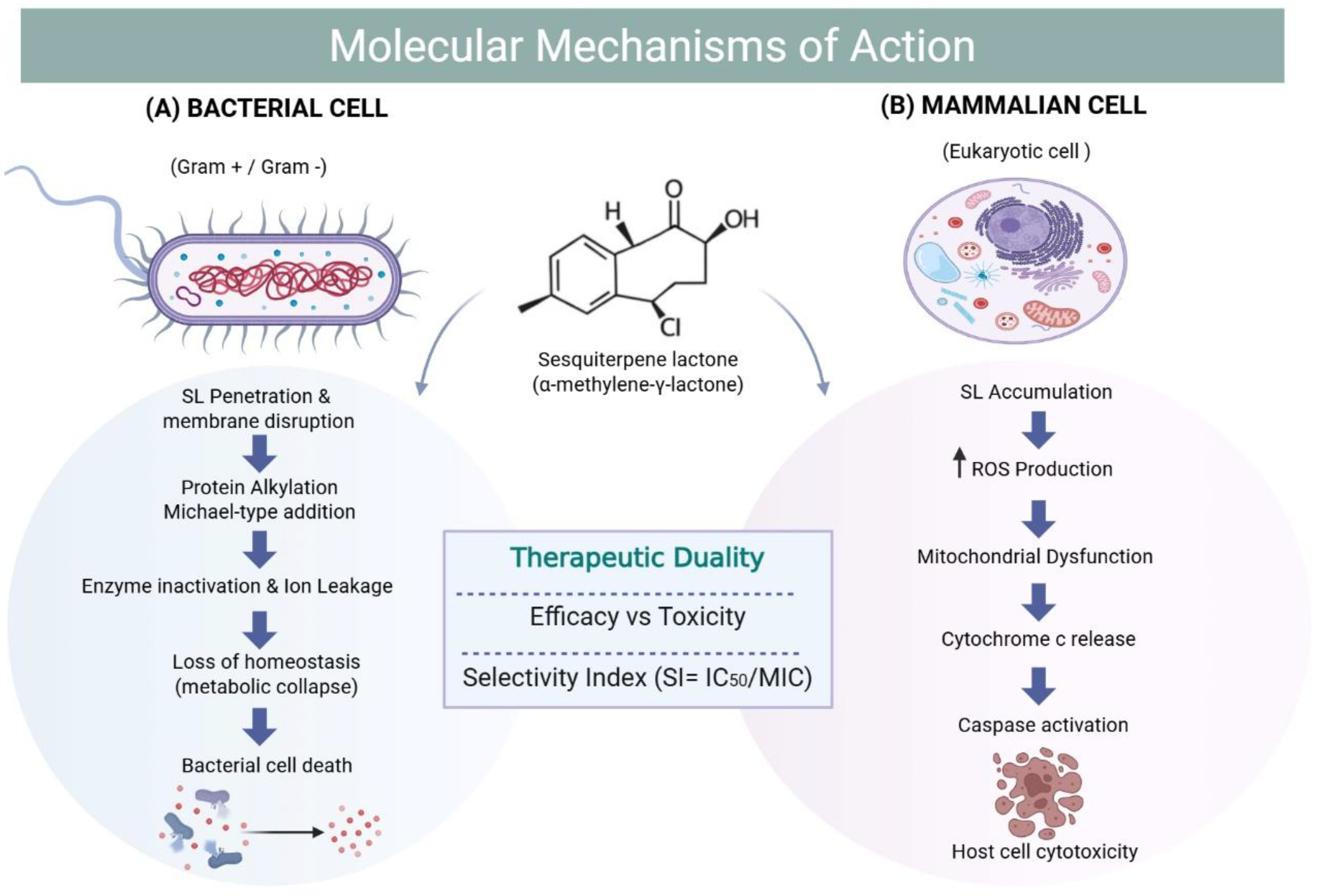
Figure 4.
Synthesis of 4β-cinnamoyloxy,1β,3α-dihydroxyeudesm-7,8-ene derivatives 2–4.
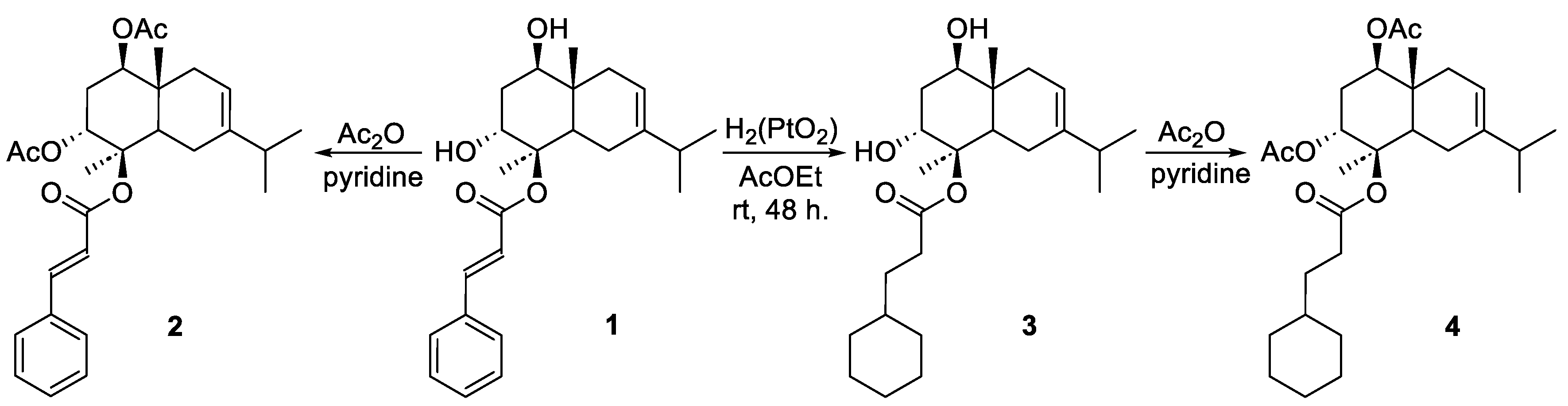
Figure 5.
Translational Feasibility: The SI Comparison.
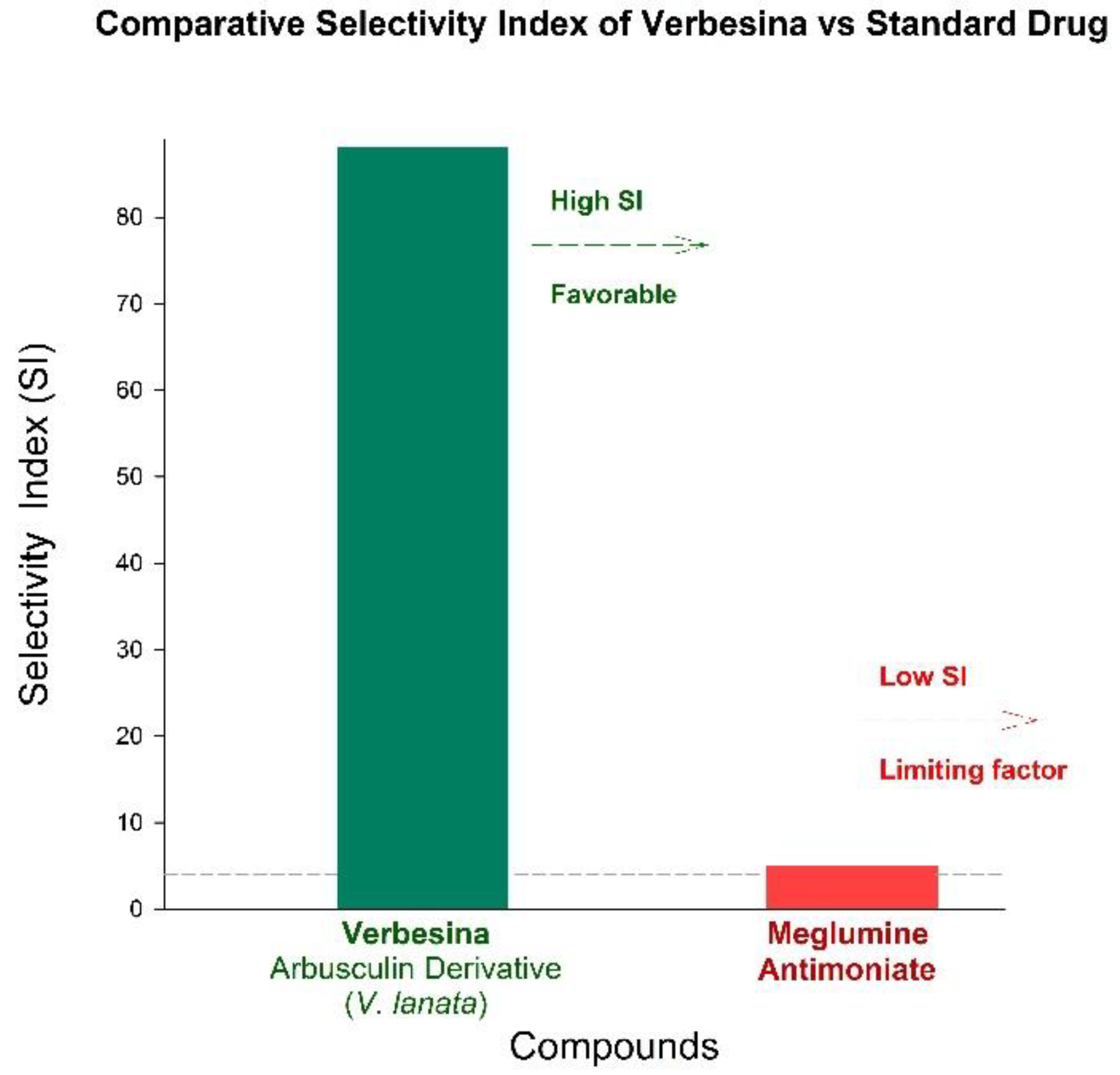
Table 2.
Comparative Analysis of Selectivity Index (SI) and Efficacy-Toxicity Duality for Verbesina Metabolites and Extracts.
Table 2.
Comparative Analysis of Selectivity Index (SI) and Efficacy-Toxicity Duality for Verbesina Metabolites and Extracts.
| Category | Compound / Extract |
Efficacy (Target & Value) |
Toxicity (Cell Line & Value) |
Selectivity Index (SI) |
|---|---|---|---|---|
| I. Confirmed High Selectivity (SI >> 1) | ||||
| Anti-Protozoal Lead [47] | Arbusculin Derivative (V. lanata) |
L. major amastigotes, 4.9 - 25.3 μM |
NMM, 432.5 - 620.7 μM |
Up to 88.2 |
| Non-Cytotoxic Triterpenes [106] | Pseudotaraxasterol Derivatives (from V. encelioides) |
Protozoa (Antiprotozoal activity), 32.2 - 48.2 μg/mL |
HELF, (MRC-5) (CC50), > 64 μg/mL |
>1.3−2.0 |
| Host-Immune Booster [113] | V. oerstediana Acetone Bark Extract |
S. aureus, Significantly boosts killing at 10 μg/mL |
Neutrophils (Viability), Non-cytotoxic at 10 μg/mL |
High (Qualitative) |
| II. Efficacy & Duality Challenges | ||||
| Efficacy Benchmark (Germacrene) [38] |
Germacrene SL (V. negrensis) |
S. aureus / E. faecalis (MIC), 64 μg/mL |
CC50 on non-cancerous line: > 64 μg/mL | ≈1 (Implied) |
| Duality Case (Mild Uncoupler) [62] | CDE (from V. persicifolia) | Undocumented | RLM (Ceff), 20-100 μM (Mild Uncoupling) | ≈1 (Implied) |
| Cytotoxicity Benchmark (Extract) [73] |
V. turbacensis Hydroethanolic Extract |
(Low Activity assumed) | HepG2 cells (IC50), 334.6 μg/mL | High (Qualitative) |
| Toxicity Benchmark (Isolated Toxin) [87] |
Galegine (Alkaloid) (from V. encelioides) | (Toxicity focus) | Hep G2 cells (IC50), 10.9 μg/mL | Very Low SI (Implied) |
| Extract Duality (Allelopathy) [3] |
V. encelioides Root Leachate |
Inhibition (Full Strength), -25.7% (RRG) | Stimulation (Low Dilution), +36.5% (RRG) |
Concentration- Dependent |
SI: Selectivity Index (CC50/IC50 or CC50/MIC); NMM: Normal murine macrophages; HELF: Human embryonic lung fibroblasts; RLM: Rat Liver Mitochondria; RRG: Radish Root Growth; IC50: standard inhibitory, MIC: Minimum Inhibitory Constant; CC50: Cytotoxicity; GI50: growth inhibition; The SI for the Germacrene Efficacy Benchmark (MIC = 64 μg/mL) is inferred (≥ 1) as a specific non-cytotoxicity CC50 is currently undocumented, although correlated non-toxic triterpenoid CC50 values for V. encelioides exceed this concentration; Arbusculin Derivative: (1R,4S,4aS,7R)-1-methyl-7-(prop-1-en-2-yl)octahydro-2H-1,4a-(epoxymethano)naphthalen-4-yl acetate; Germacrene SL: (1S,6R,7S,10S,Z)-6-hydroxy-10-isopropyl-7-methoxy-3,7-dimethyl-4-oxocyclodec-2-en-1-yl cinnamate; CDE: (4β-cinnamoyloxy- 1β,3α-dihydroxyeudesm-7,8-ene).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



