
Submitted:
09 January 2026
Posted:
09 January 2026
You are already at the latest version
Abstract
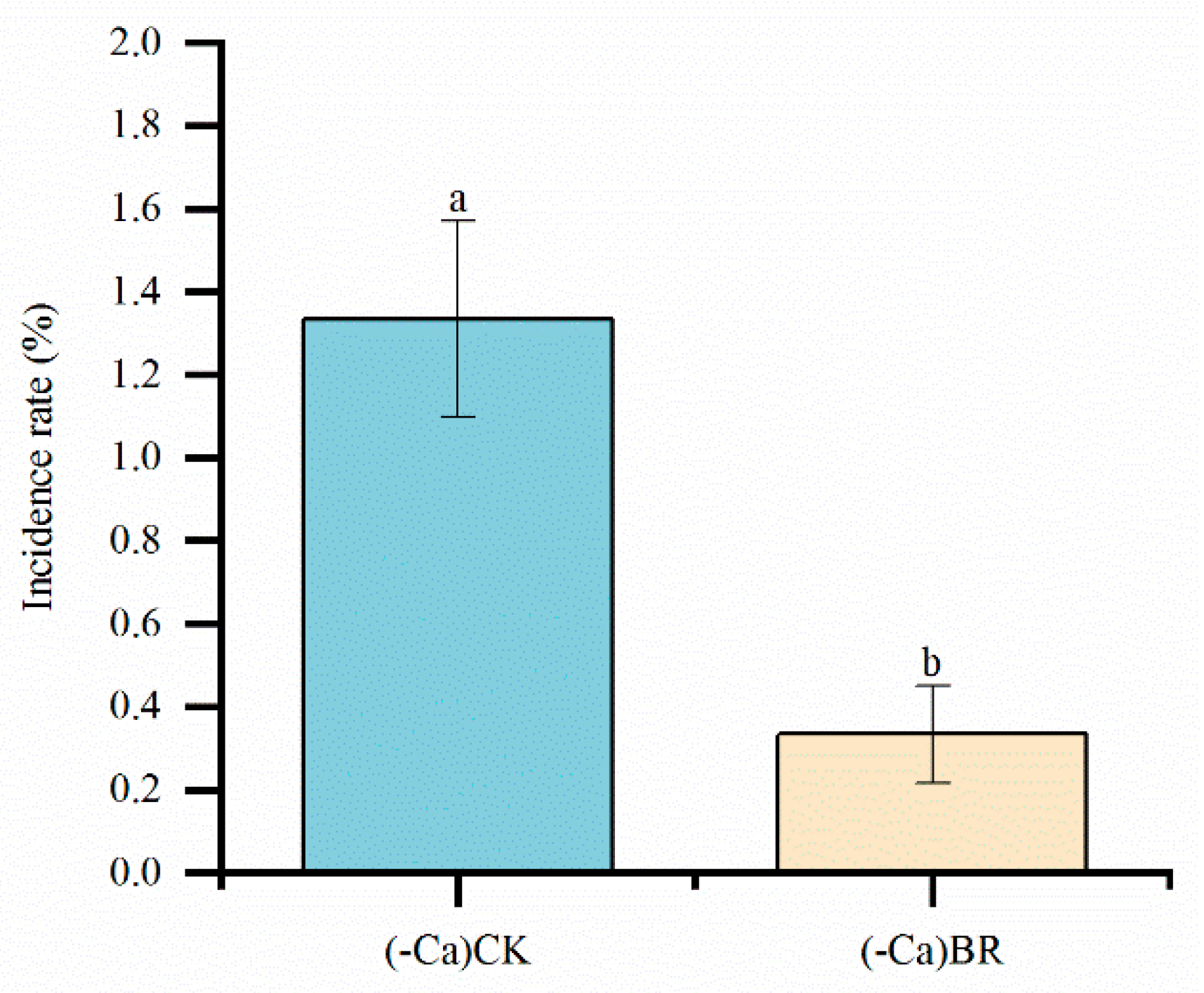
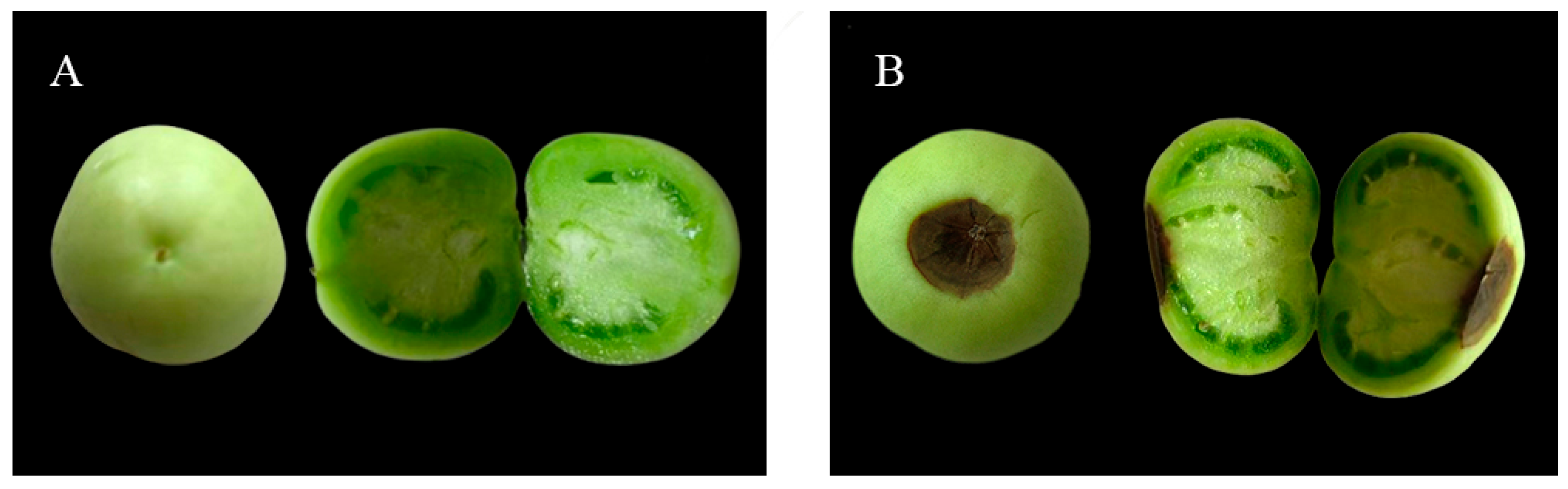
Blossom-end rot (BER) in tomatoes is a physiological disorder primarily caused by disrupted calcium absorption and transport. This study aims to investigate the physiological and molecular mechanisms by which exogenous brassinolide (BR) reduces the incidence of tomato BER under calcium-deficient conditions. The results showed that under calcium deficiency, foliar spraying of BR significantly reduced the BER incidence (from 26.67% to 6.67%) and effectively increased calcium ion content in the leaves, stems, roots, and other parts of the plant. At the physiological level, BR treatment significantly enhanced the activities of CAT, POD, and SOD in the leaves (by 105.70%, 117.12%, and 82.77%, respectively), while reducing the contents of H₂O₂ (by 36.90%) and MDA (by 16.38%). This indicates that BR alleviates membrane lipid peroxidation damage by enhancing the antioxidant defense system. Transcriptome analysis further revealed that BR treatment identified 4807, 2807, and 2554 differentially expressed genes (DEGs) in the leaves, stems, and roots, respectively, with the most significant response observed in the leaves. GO functional enrichment analysis showed that these genes are mainly involved in biological processes such as secondary metabolic processes, response to oxygen-containing compounds, and cell wall organization. KEGG pathway analysis further indicated significant enrichment in pathways such as phenylpropanoid biosynthesis, plant hormone signal transduction, and plant-pathogen interaction. Additionally, several key genes, such as the cytochrome C oxidase gene (Solyc03g013460.1), exhibited a gradient up-regulation pattern of "root > stem > leaf" in the oxidative phosphorylation pathway, while photosynthesis-related genes (e.g., PsbA, PsaA, and ND family genes) were also generally up-regulated. In summary, BR likely enhances tomato tolerance to calcium deficiency stress and effectively reduces the occurrence of BER through multiple pathways, including regulating calcium absorption and distribution, activating the antioxidant system, modulating hormone signaling pathways, and enhancing energy metabolism. This provides a theoretical basis for the application of BR in agricultural production.
Keywords:
1. Introduction
2. Results
2.1. Plant Growth Status under Calcium-Deficient Nutrient Solution + Distilled Water Spray and Calcium-Deficient Nutrient Solution + BR Spray
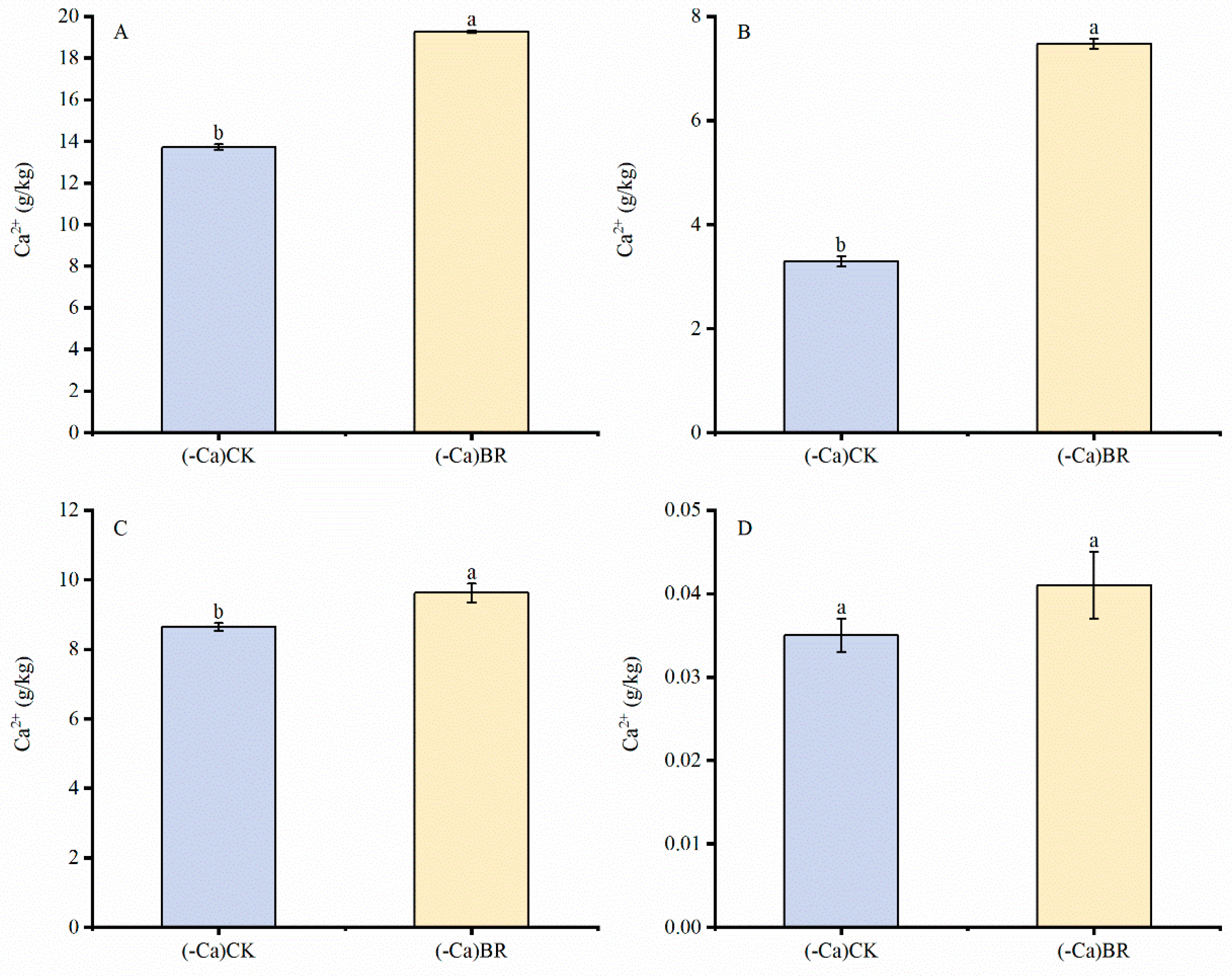
2.2. Effect of BR on Calcium Ion Content in Different Tomato Parts
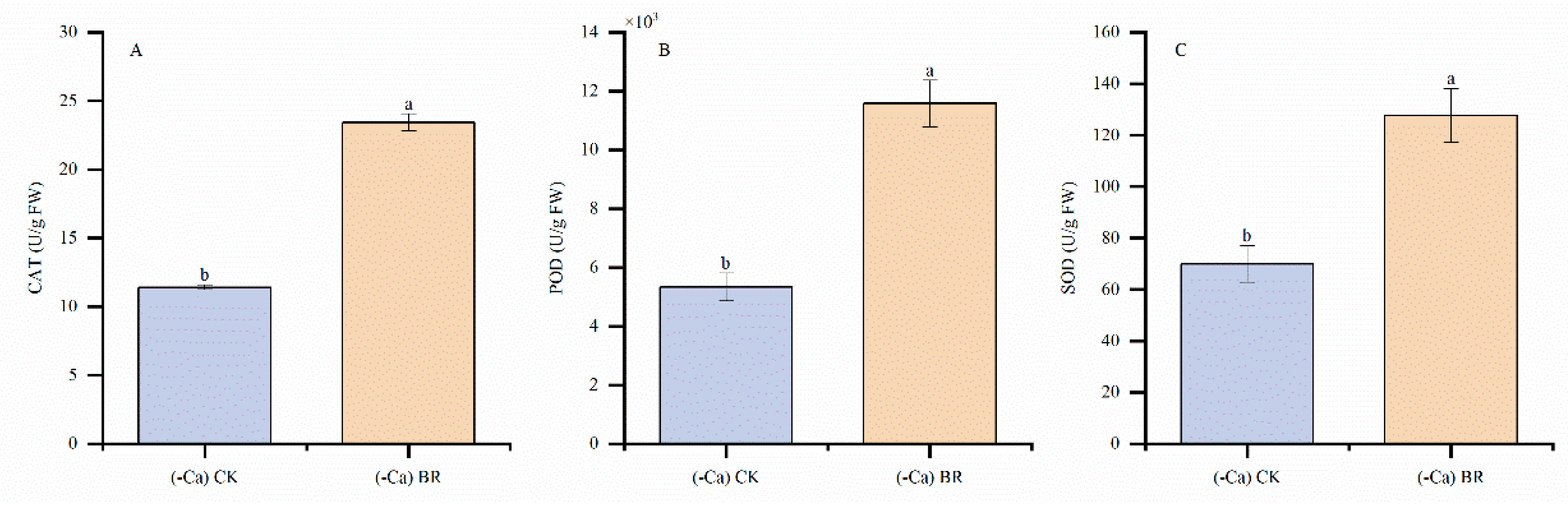
2.3. Effect of BR on Antioxidant Enzymes in Tomato Leaves
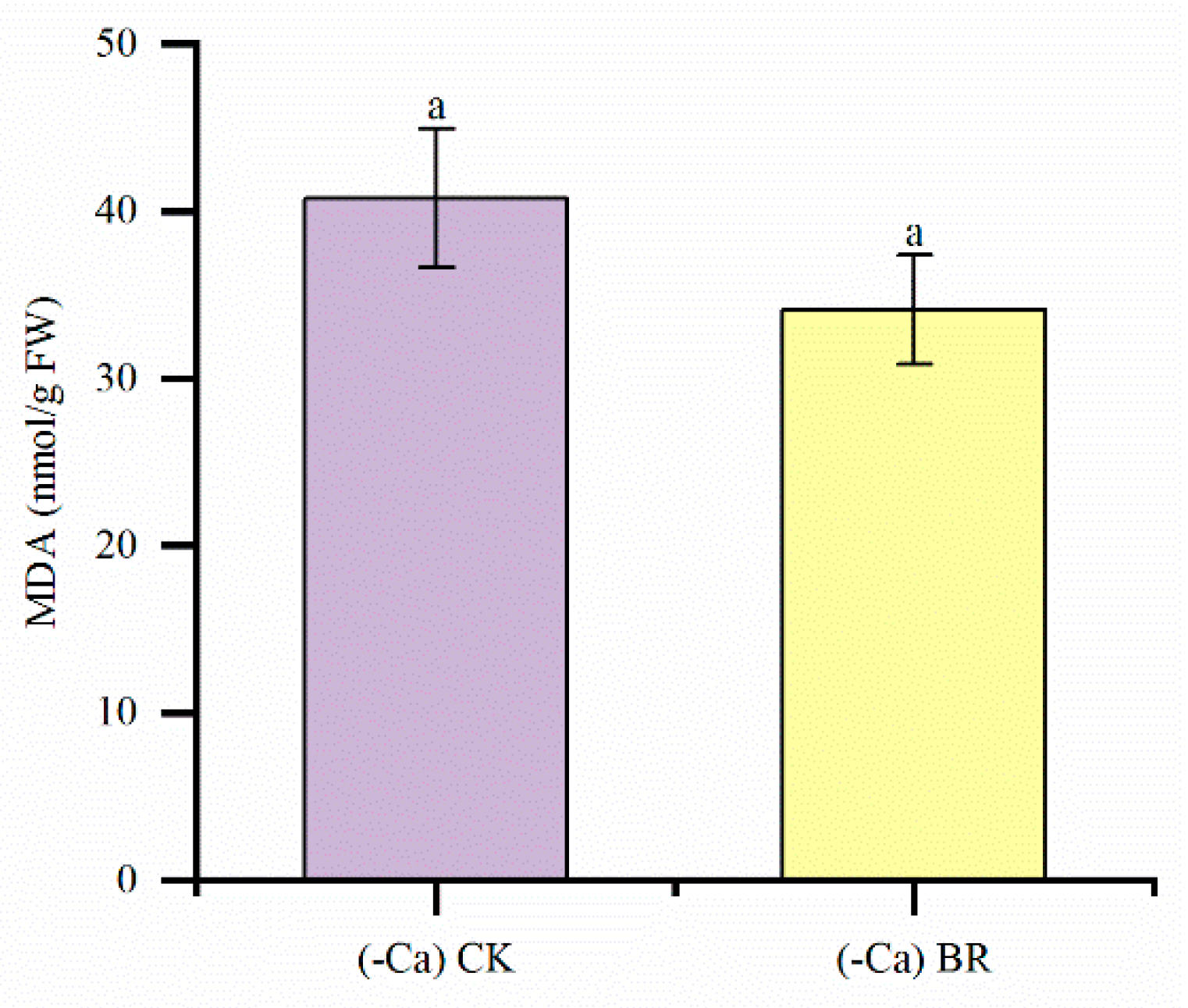
2.4. Effect of BR on MDA in Tomato Leaves
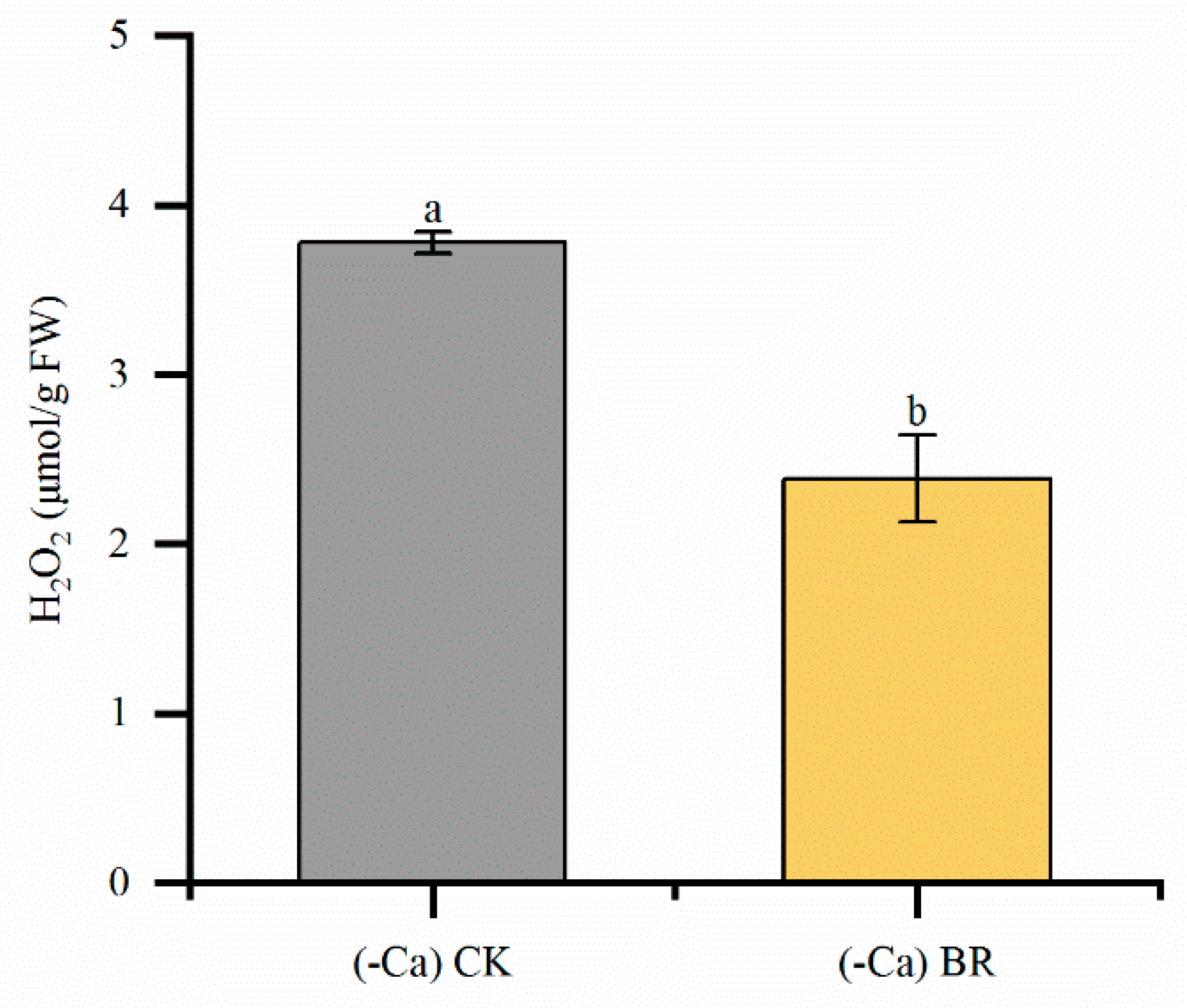
2.5. Effect of BR on H₂O₂ in Tomato Leaves
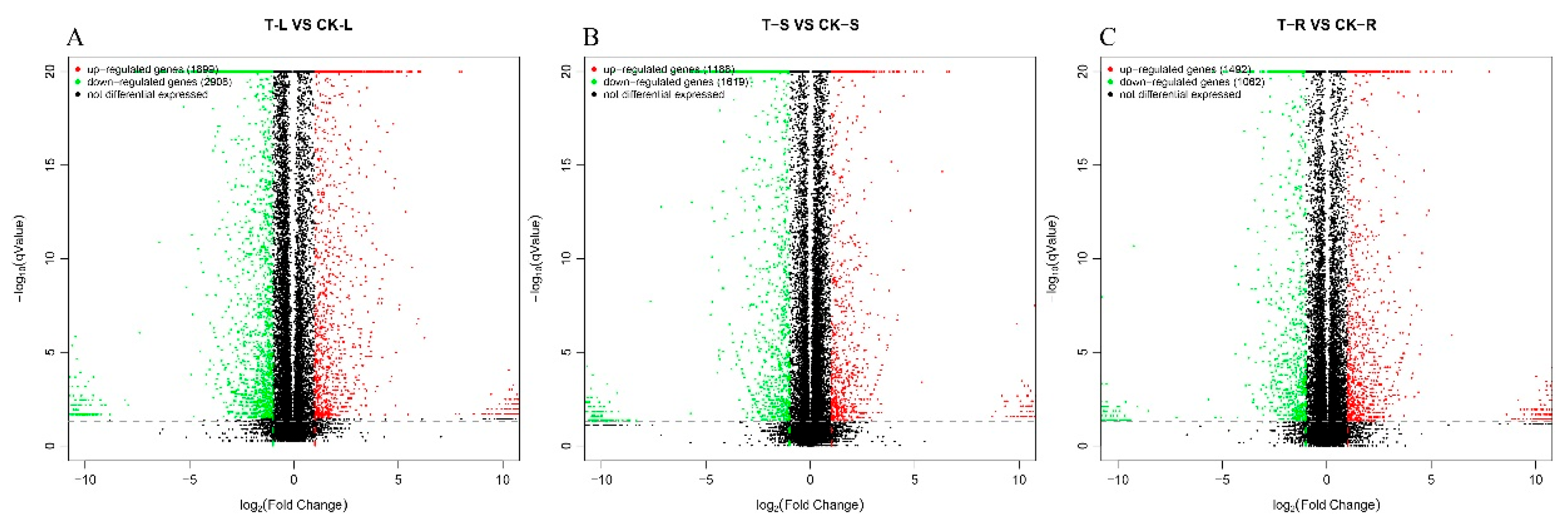
2.6. Differential Gene Screening

| Up-Regulated Gene Annotation Group | Number of Genes |
|---|---|
| Calmodulin(cam) | 1 |
| MADS-box transcription factor | 1 |
| MYB transcription factors | 3 |
| Cold shock protein | 1 |
| Glycine-rich protein | 6 |
| Glutathione S-transferase | 4 |
| Pectinesterase | 4 |
| Cytochrome c oxidase | 4 |
| Fatty acyl-coa | 2 |
| ABC transporter | 3 |
| ABA deficient protein | 1 |
| Sugar transport proteins | 3 |
| Auxin-responsive protein SAUR | 5 |
| Ribosomal proteins | 6 |
| Auxin efflux facilitator slpin | 1 |
| Atpase | 8 |
| Cytochrome P450 | 5 |
| Other | 414 |
| Down-regulated gene annotation group | |
| Heat shock protein | 3 |
| MADS-box protein | 1 |
| MYB transcription factors | 2 |
| Glycine-rich protein | 2 |
| Cytochrome P450 | 13 |
| Jasmonic acid-amido | 1 |
| Glucan endonuclease | 6 |
| Glutathione S-transferase | 1 |
| Pathogenesis-related leaf protein | 4 |
| Sugar transport proteins | 2 |
| Ethylene-responsive transcription factors | 8 |
| WRKY transcription factors | 6 |
| Serine/threonine protein kinases | 8 |
| Della protein | 1 |
| Other | 459 |
| Up-Regulated Gene Annotation Group | Number of Genes |
|---|---|
| Glutathione S-Transferase | 1 |
| Cytochrome P450 | 2 |
| Cytochrome C Oxidase | 1 |
| Pectinesterase | 1 |
| Calcium-Binding Protein | 1 |
| Sugar Transport Proteins | 3 |
| Ribosomal Protein | 1 |
| Auxin | 3 |
| Stress-Associated Protein | 3 |
| Ethylene-Responsive Transcription Factors | 3 |
| Other | 222 |
| Down-regulated gene annotation group | |
| Calcium-binding protein | 3 |
| MYB transcription factors | 9 |
| WRKY transcription factors | 8 |
| Cytochrome P450 | 7 |
| Auxin | 25 |
| Glutathione S-transferase | 2 |
| Pectinesterase | 1 |
| Serine/threonine protein kinases | 7 |
| Ethylene-responsive transcription factors | 19 |
| Jasmonic acid-amido | 1 |
| Sugar transport proteins | 2 |
| B3 domain-containing proteins | 2 |
| Bhlh transcription factors | 3 |
| Other | 344 |
| Up-Regulated Gene Annotation Group | Number of Genes |
|---|---|
| WRKY transcription factors | 2 |
| Calcium-binding protein | 1 |
| MYB transcription factors | 5 |
| ABC transporter | 1 |
| Ribosomal protein | 1 |
| Sugar transport proteins | 8 |
| Bhlh transcription factors | 4 |
| Glycine-rich proteins | 2 |
| Pectinesterase | 4 |
| Della proteins | 2 |
| Other | 436 |
| Down-regulated gene annotation group | |
| Sugar transport proteins | 2 |
| MYB transcription factor | 1 |
| Auxin-responsive proteins | 3 |
| ABC transporter | 3 |
| Bhlh transcription factor | 1 |
| Pectinesterase | 4 |
| Ethylene-responsive transcription factor | 1 |
| Other | 279 |
2.7. GO Enrichment Analysis of Differentially Expressed Genes
| Category | DEGs | GO enrichment | |||
| biological process | cellular component | molecular function | total | ||
| T-L_vs_CK-L | 4807 | 1654 | 1574 | 1422 | 4650 |
| T-S_vs_CK-S | 2807 | 986 | 914 | 864 | 2764 |
| T-R_vs_CK-R | 2554 | 853 | 846 | 759 | 2458 |
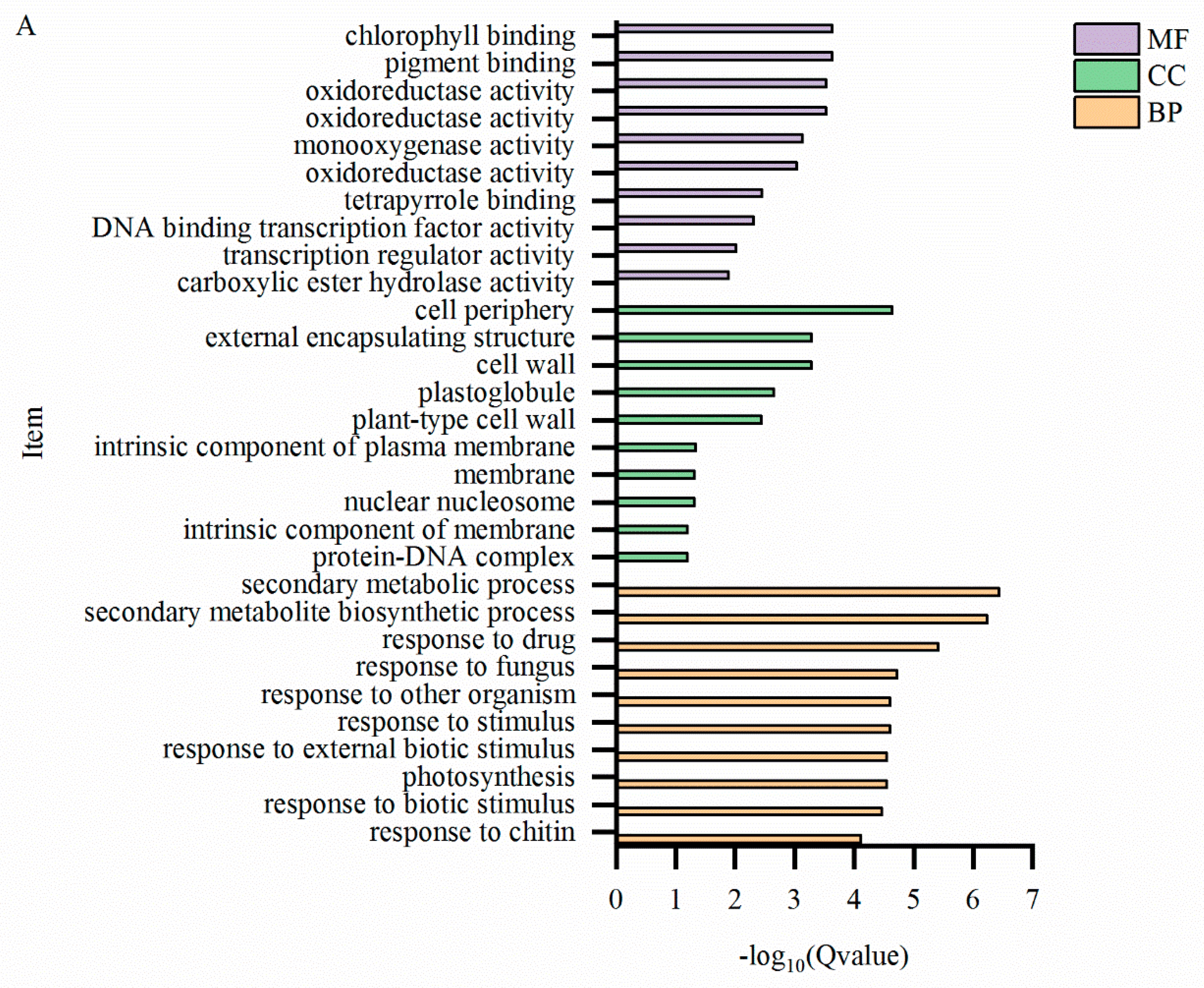
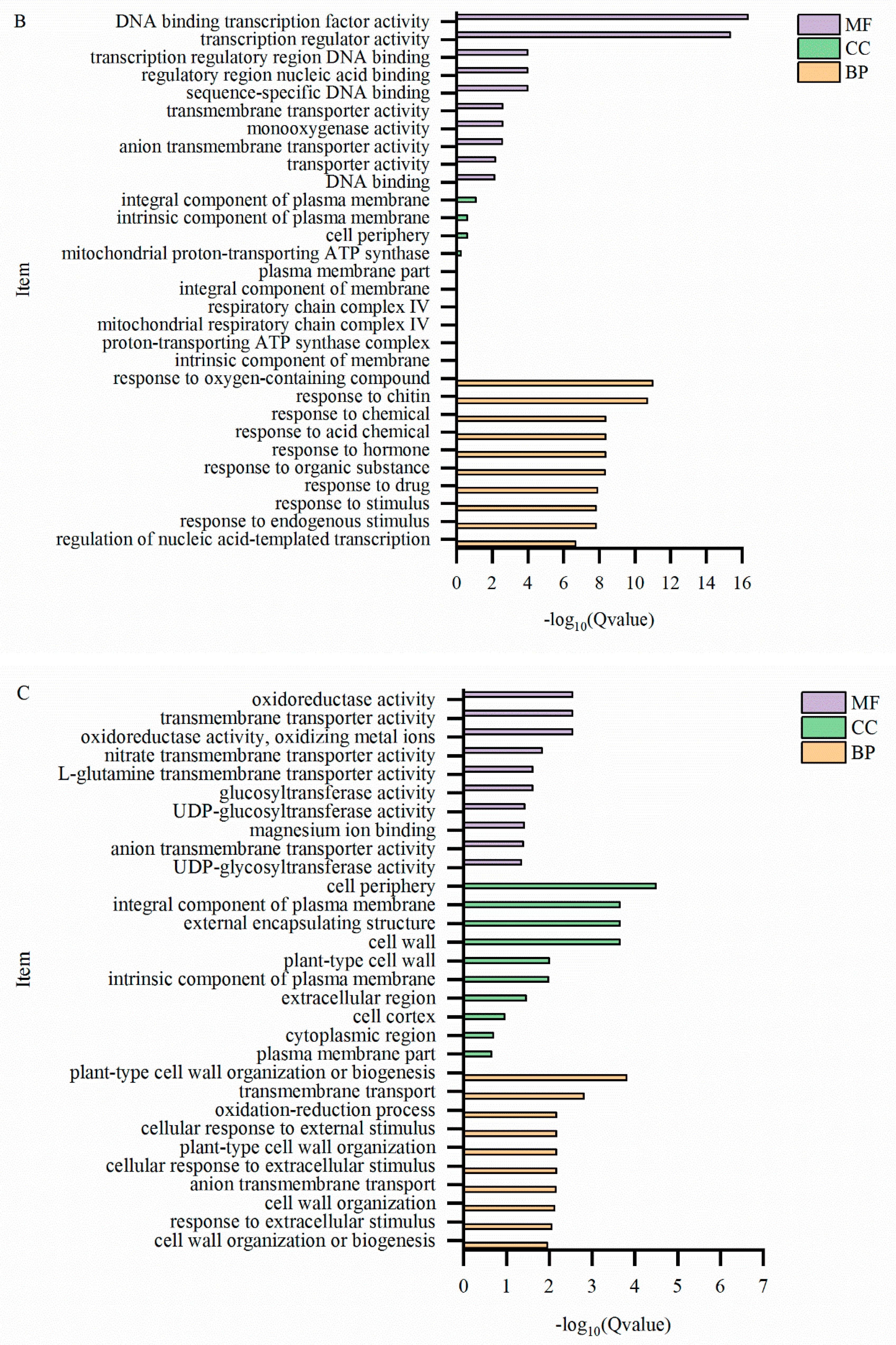
2.8. KEGG Metabolic Pathway Enrichment Analysis of Differentially Expressed Genes
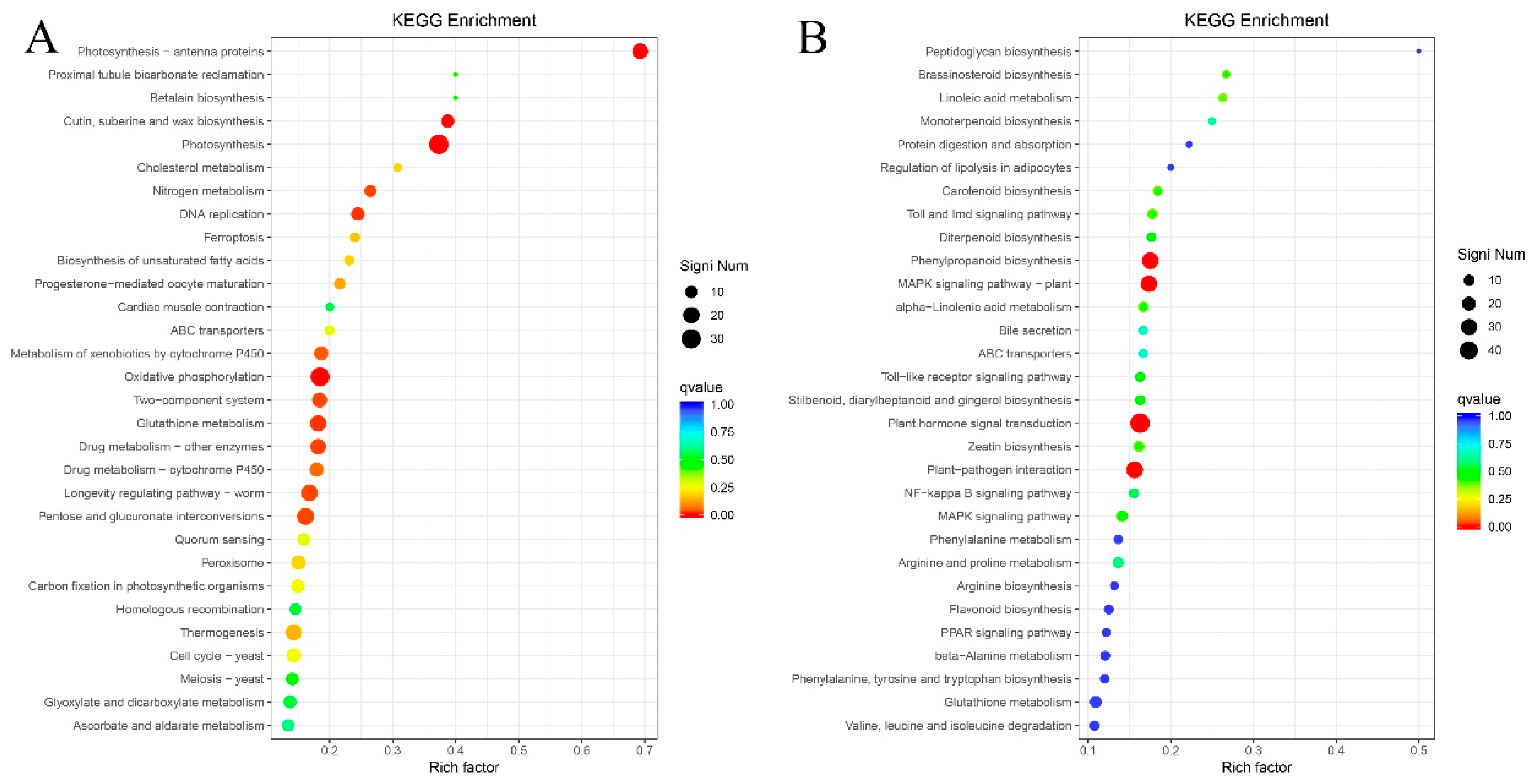
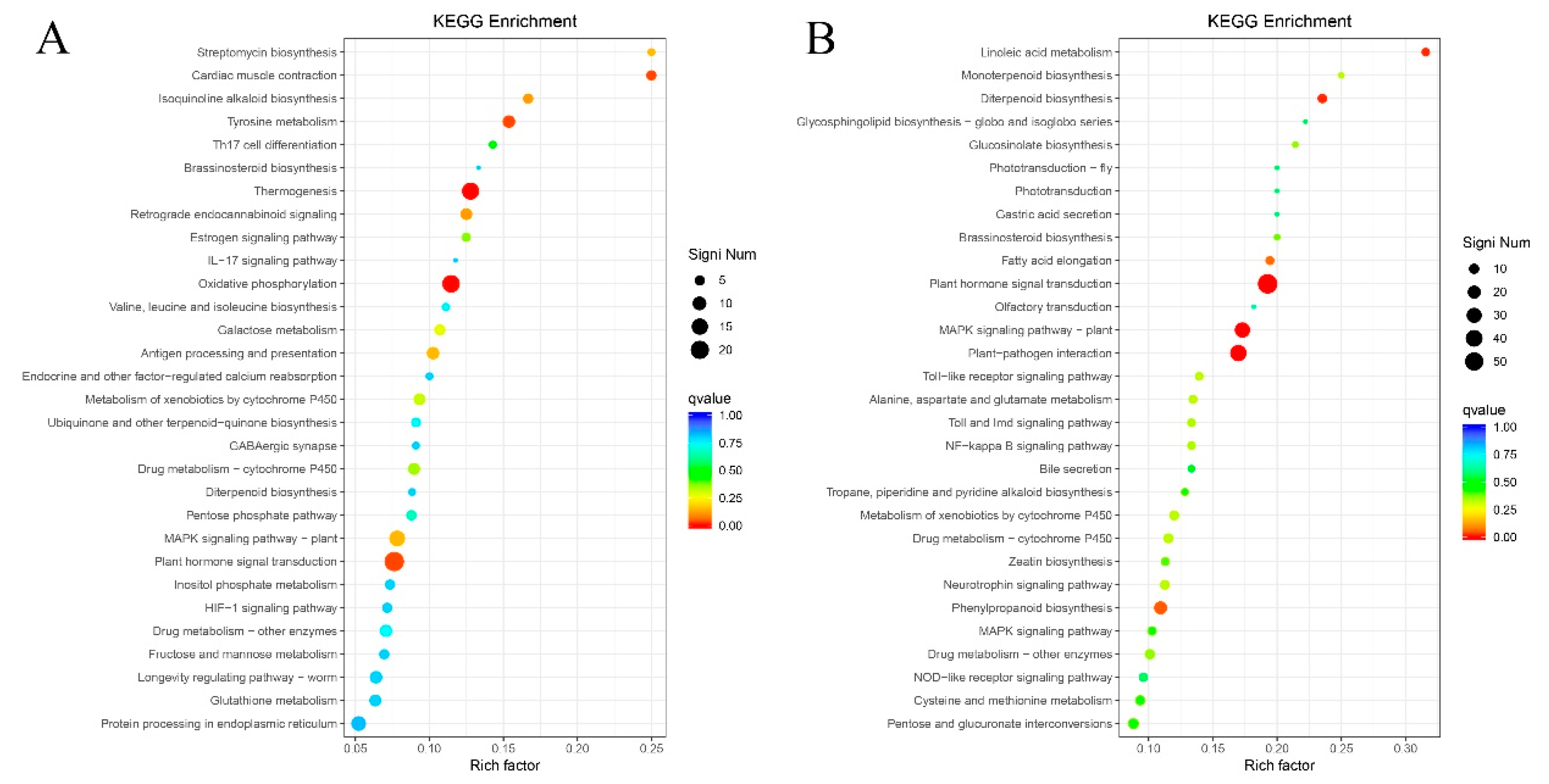
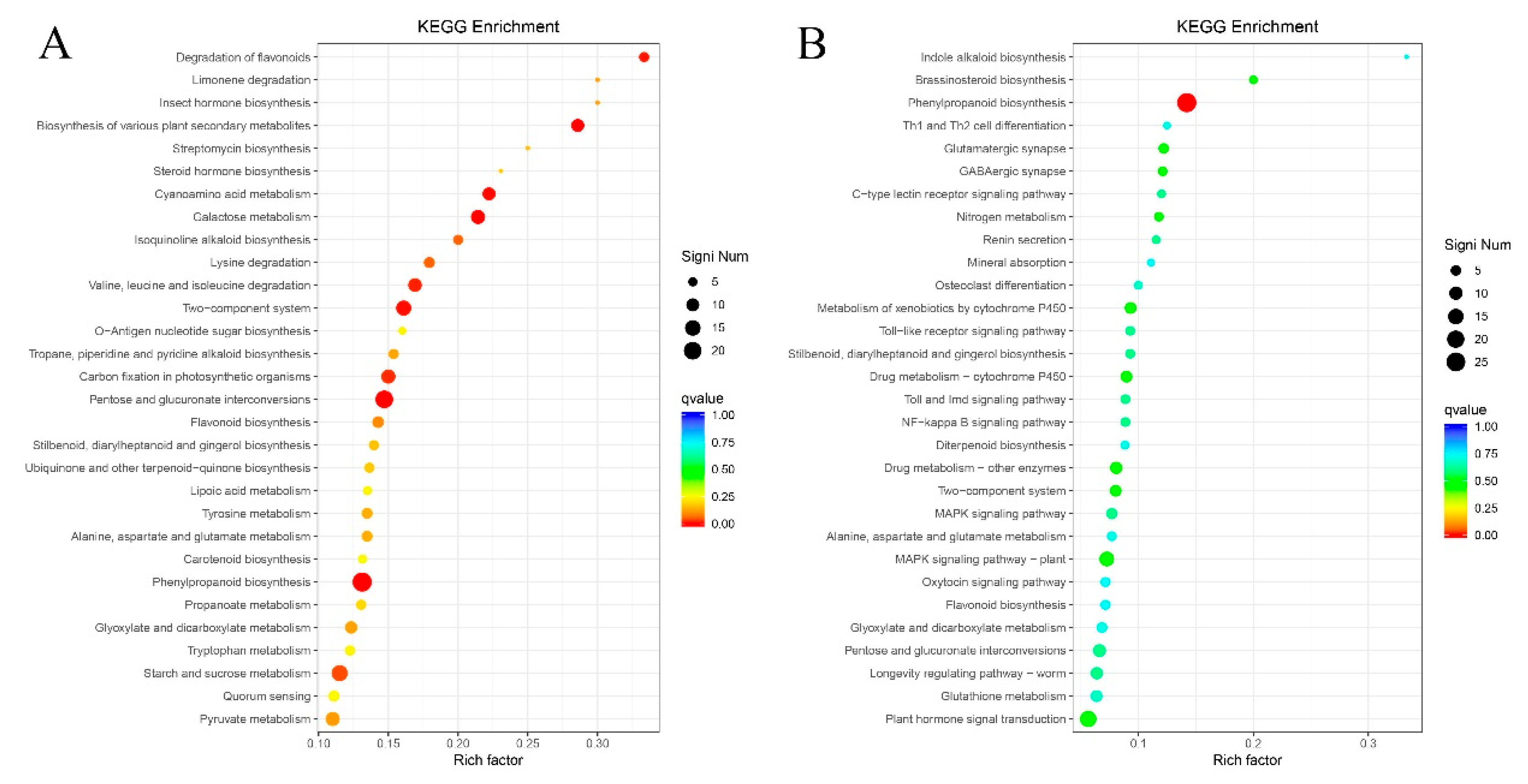
2.9. Metabolic Pathways
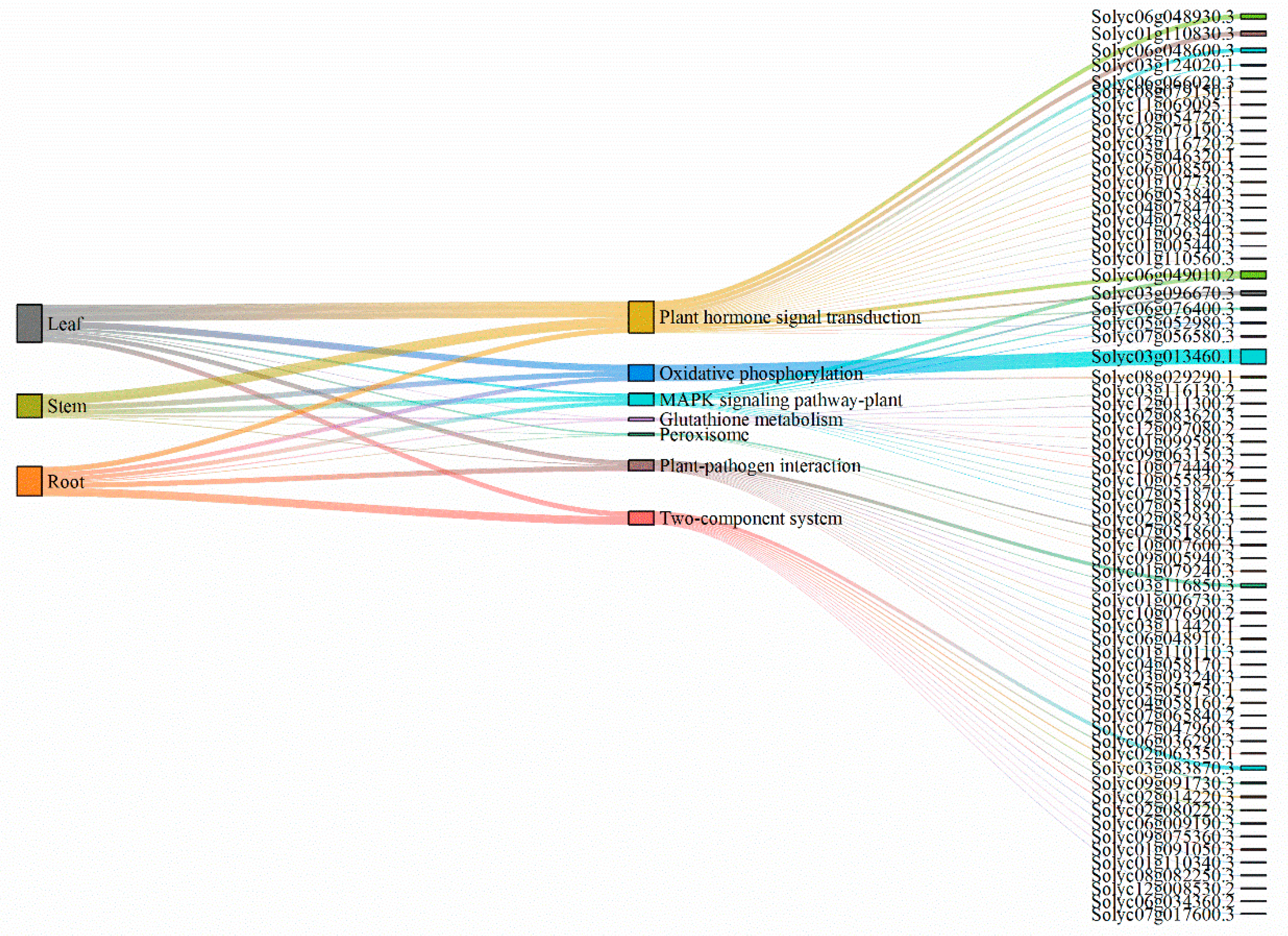
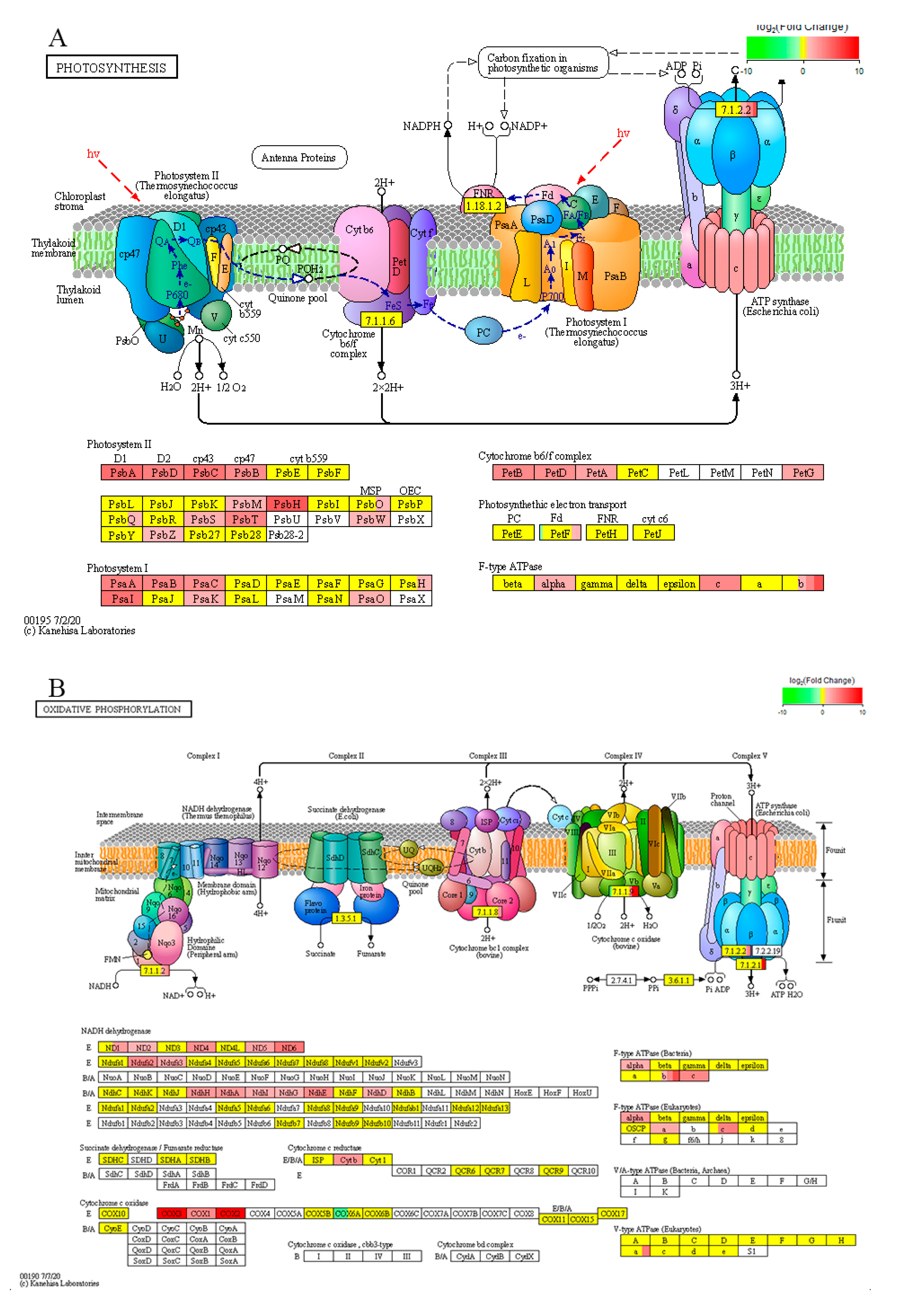
3. Discussion
3.1. BR Reduces BER Incidence by Promoting Calcium Absorption and Distribution
3.2. BR Alleviates Oxidative Damage by Activating the Antioxidant Defense System
3.3. Multi-Omics Analysis Reveals the Molecular Network Regulated by BR
3.4. BR Enhances Energy Metabolism by Regulating Photosynthesis-Related Genes
3.5. BR Enhances Disease Resistance-Related Pathways to Compensate for Immune Deficiencies Caused by Calcium Deficiency
4. Materials and Methods
4.1. Plant Material
4.2. Experimental Design
4.3. Determination of Antioxidant Enzyme Activities
4.4. Tomato Sample Total RNA Extraction, Library Construction, and Transcriptome Sequencing
4.5. Differential Gene Screening
4.6. Data Analysis
5. Conclusion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, D.; Zhang, Q.; Li, Y.; Qi, L.; Niu, R.; Zhao, X.; Shi, J.; Tian, G. Regularity of absorption and distribution of nitrogen, phosphorus, potassium, calcium, and magnesium in growth bag tomatoes in solar greenhouse. Journal of Hebei Agricultural University 2021, 44, 33–40. [Google Scholar] [CrossRef]
- Diaz-Perez, J.C.; Hook, J.E. Plastic-mulched bell pepper (Capsicum annuum L.) plant growth and fruit yield and quality as influenced by irrigation rate and calcium fertilization. Hortscience 2017, 52, 774–781. [Google Scholar] [CrossRef]
- Karlsons, A.; Osvalde, A.; Cekstere, G.; Abolina, L. Effects of Ca sprays on fruit Ca content and yield of tomato variety susceptible to blossom-end rot. Plants-Basel 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Hagassou, D.; Francia, E.; Ronga, D.; Buti, M. Blossom end-rot in tomato (Solanum lycopersicum L.): A multi-disciplinary overview of inducing factors and control strategies. Scientia Horticulturae 2019, 249, 49–58. [Google Scholar] [CrossRef]
- Reitz, N.F.; Shackel, K.A.; Mitcham, E.J. Differential effects of excess calcium applied to whole plants vs. excised fruit tissue on blossom-end rot in tomato. Scientia Horticulturae 2021, 290. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, Q.; Li, Y.; Wang, D.; Geng, X.; Jia, H.; Qi, L.; Shi, J.; Zhao, L.; Tian, D. Effect of different calcium fertilizers on quality and umbilical rot of palatable tomatoes. China Cucurbits and Vegetables 2023, 36, 65–70. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, G.; Ge, M.; Chang, Y.; Tan, J.; Ge, X.; Wang, J.; Wang, B.; Zhou, G.; Fu, T. Function and application of calcium in plant growth and development. Acta Agronomica Sinica 2024, 50, 793–807. [Google Scholar] [CrossRef]
- Hou, X. The mechanisms underlying CAMTA5-suppressed blossom-end rot occurrence in tomato. Doctor, Gansu Agricultural University, 2025. [Google Scholar]
- Schmitz-Eiberger, M.; Haefs, R.; Noga, G. Calcium deficiency - Influence on the antioxidative defense system in tomato plants. Journal of Plant Physiology 2002, 159, 733–742. [Google Scholar] [CrossRef]
- Wu, Y.; Lin, Q.; Chen, Z.; Lin, J. Effects of calcium level on calcium absorption, fruit quality and yield of infinite growth tomato. Journal of Soil and Water Conservation 2019, 33, 185–189. [Google Scholar] [CrossRef]
- Ho, L.C.; White, P.J. A cellular hypothesis for the induction of blossom-end rot in tomato fruit. Annals of Botany 2005, 95, 571–581. [Google Scholar] [CrossRef]
- Marcelis, L.F.M.; Ho, L.C. Blossom-end rot in relation to growth rate and calcium content in fruits of sweet pepper (Capsicum annuum L.). Journal of Experimental Botany 1999, 357–363. [Google Scholar] [CrossRef]
- Aslani, L.; Gholami, M.; Mobli, M. The relationship between tomato fruit growth, incidence of blossom-end rot and phytohormone content as affected by sink/source ratio. Annals of Applied Biology 2020, 177, 211–222. [Google Scholar] [CrossRef]
- Gaion, L.A.; Júnior, J.C.M.; Barreto, R.F.; D’Amico-Damião, V.; de Mello Prado, R.; Carvalho, R.F. Amplification of gibberellins response in tomato modulates calcium metabolism and blossom end rot occurrence. Scientia Horticulturae 2019, 246, 498–505. [Google Scholar] [CrossRef]
- Balate, C.A.; Souza, D.C.d.; Resende, L.V.; Freitas, S.T.d. Effect of abscisic acid on the calcium content for controlling blossom-end rot in tomato under water stress. Pesquisa Agropecuária Tropical 2018, 48, 414–419. [Google Scholar] [CrossRef]
- Dong, S.; Sun, J.; Sun, M.; Ma, Y.; Zhou, X.; Qu, Z. Effect of brassinolide spraying on soybean yield under straw returning conditions. Journal of Shenyang Agricultural University 2025, 56, 24–32. [Google Scholar] [CrossRef]
- Duan, F.; Liu, B.; Zhang, Z.; Zheng, S.; Yang, C.; Wang, Q. Regulatory effects of 2,4-epibrassinolide, polyglutamic acid and uniconazole on tuber yield and quality in sweet potato. Jiangsu Journal of Agricultural Sciences 2025, 41, 1072–1079. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, Z.; Cao, J.; Luo, X.; Wang, Y. Effects of 2,4-epibrassinolide on the growth and physiological characteristics of Phaseolus vulgaris seedlings under saline and alkaline stresses. Acta Botanica Boreali-Occidentalia Sinica 2024, 44, 1181–1189. [Google Scholar] [CrossRef]
- Li, Y. Physiological and molecular mechanism of brassinolide regulated calcium-deficiency induced tipburn resistance in mini chinese cabbage. Doctor, Gansu Agricultural University, 2022. [Google Scholar]
- Riboldi, L.B.; Gaziola, S.A.; Azevedo, R.A.; de Freitas, S.T.; Roberto, D.C.E.C.; Paulo. 24-epibrassinolide mechanisms regulating blossom-end rot development in tomato fruit. Journal of Plant Growth Regulation 2018. [Google Scholar] [CrossRef]
- Topcu, Y.; Nambeesan, S.U.; van der Knaap, E. Blossom-end rot: a century-old problem in tomato (Solanum lycopersicum L.) and other vegetables. Molecular Horticulture 2022, 2, 1. [Google Scholar] [CrossRef]
- Sun, T. Genome-wide identification and comprehensive analysis of CIPK gene family in wheat. Huazhong University of Science and Technology, 2016.
- Emery, L.; Whelan, S.; Hirschi, K.D.; Pittman, J.K. Protein phylogenetic analysis of Ca2+/cation antiporters and insights into their evolution in plants. Frontiers in Plant Science 2012, 3. [Google Scholar] [CrossRef]
- Li, P.; Zhang, G.; Gonzales, N.; Guo, Y.; Hu, H.; Park, S.; Zhao, J. Ca2+-regulated and diurnal rhythm-regulated Na+/Ca2+exchanger AtNCL affects flowering time and auxin signalling inArabidopsis. Plant Cell and Environment 2016, 39, 377–392. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Liu, Y.; Zhang, Y.; Liu, J.; Gul, Z.; Guo, X.; Abozeid, A.; Tang, Z. Effects of exogenous calcium on adaptive growth, photosynthesis, ion homeostasis and phenolics of gleditsia sinensis Lam. plants under salt stress. 2021, 11, 1–18. [Google Scholar] [CrossRef]
- Ishu; Shumayla; Madhu; Upadhyay, S.K. Complementation with TaNCL2-A reinstates growth and abiotic stress response in atncl mutant of Arabidopsis. Plant Science 2025, 353. [Google Scholar] [CrossRef] [PubMed]
- Shumayla; Tyagi, S.; Sharma, Y.; Madhu; Sharma, A.; Pandey, A.; Singh, K.; Upadhyay, S.K. Expression of TaNCL2-A ameliorates cadmium toxicity by increasing calcium and enzymatic antioxidants activities in arabidopsis. Chemosphere 2023, - 329. [Google Scholar] [CrossRef]
- Hao, F.; Cui, Z.G.; Dong, X.; Gao, Y.; Wang, R.J.; Zhang, H.; Lin, G.L. Exogenous calcium enhances castor tolerance to saline-alkaline stress by regulating antioxidant enzyme activity and activating ca2+ and ROS signaling crosstalk. International Journal of Molecular Sciences 2024, 25. [Google Scholar] [CrossRef]
- Roussin-Leveillee, C.; St-Amand, M.; Desbiens-Fortin, P.; Perreault, R.; Pelletier, A.; Gauthier, S.; Gaudreault-Lafleur, F.; Laforest-Lapointe, I.; Moffett, P. Co-occurrence of chloroplastic ROS production and salicylic acid induction in plant immunity. New Phytologist 2025, 248, 1989–2004. [Google Scholar] [CrossRef]
- Jiang, H.; Xie, Y.; Liu, Q. Molecular biology research progress of cytochrome c oxidase. Foreign Medical Sciences • Section of Pathophysiology and Clinical Medicine 2001, 20–22. [Google Scholar] [CrossRef]
- Gao, Y.; Zhou, X.; Huang, H.; Wang, C.; Xiao, X.; Wen, J.; Wu, J.; Zhou, S.; de Dios, V.R.; Rodriguez, L.G.; et al. ORANGE proteins mediate adaptation to high light and resistance to Pseudomonas syringae in tomato by regulating chlorophylls and carotenoids accumulation. International Journal of Biological Macromolecules 2025, 306. [Google Scholar] [CrossRef]
- Dai, H.; Hu, L.; Wang, J.; Yue, Z.; Wang, J.; Chen, T.; Li, J.; Dou, T.; Yu, J.; Liu, Z. Constructing a novel disease resistance mechanism model for cruciferous crops: an example from black rot. Molecular Plant Pathology 2025, 26. [Google Scholar] [CrossRef]
- Yue, L.; Wang, L.; Neuhaeuser, B.; Zhang, S.; Herren, G.; Heuberger, M.; Jung, E.; Ludewig, U.; Zipfel, C.; Keller, B. Cytoplasmic calcium influx mediated by Lr14a regulates stomatal immunity against leaf rust in wheat. bioRxiv 2024. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, D.; Li, Y.; Wang, Z.; Wu, Z.; Zhang, Q.; Jia, H.; Dong, X.; Qi, L.; Shi, J.; et al. Gibberellin positively regulates tomato resistance to Tomato Yellow Leaf Curl Virus (TYLCV). Plants-Basel 2024, 13. [Google Scholar] [CrossRef]
- Lin, Z.; Li, S.; Lin, G.; Guo, J. The accumulation of hydrogen peroxide in senescing leaves and chloroplasts in relation to lipid peroxidation. Physiology and Molecular Biology of Plants 1988, 16–22. [Google Scholar]
- Wang, Y.; Gu, Z.; Zhang, F. Multivariate analysis of pro- and anti-oxidant properties of raspberry and blackberry from different varieties at different maturity stages. Food Science 2012, 33, 81–86. [Google Scholar]
- Zheng, F.; Zhang, X.; Wang, Y.; Han, Z. Changes of H2O2 content and activities of related enzymes in leaf among ontogenetic phases in apple (Malus ssp.). Journal of Fruit Science 2013, 30, 759–764. [Google Scholar] [CrossRef]
- Auwera, G.A.V.D.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Depristo, M.A. From FastQ data to high confidence variant calls: the Genome Analysis Toolkit best practices pipeline. Current Protocols in Bioinformatics 2013, 11, 11.10.11–11.10.33. [Google Scholar] [CrossRef]
- Damian, S.; Andrea, F.; Stefan, W.; Kristoffer, F.; Davide, H.; Jaime, H.C.; Milan, S.; Alexander, R.; Alberto, S.; Tsafou, K.P. STRING v10: protein–protein interaction networks, integrated over the tree of life. Nucleic Acids Research 2015, 43. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Cherry, J.M. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nature Genetics 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Shannon, P. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Research 2003, 13, 2498–2504. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



