
Submitted:
08 January 2026
Posted:
09 January 2026
You are already at the latest version
Abstract
Mangrove restoration is widely promoted as a nature-based solution to reverse coastal wetland degradation while sustaining fisheries productivity and other ecosystem services. This study evaluated whether a mangrove restoration program in Lubuk Kertang (North Sumatra, Indonesia) is associated with enhanced aquatic biodiversity and supporting water-quality conditions. Mangrove vegetation structure was assessed across restoration ages (2009–2013 plantings; 7–11 years old at the time of survey), and fish assemblages and water quality were sampled in June, August, and December 2021. We recorded 828 individual fishes representing 44 species, 27 families, and 17 orders. Fish diversity was moderate (mean Shannon–Wiener H′ = 2.15; evenness = 0.83; dominance = 0.17), with Engraulidae contributing the highest abundance. Water quality conditions were within ranges typical for estuarine mangrove habitats (DO 3.5–5.15 mg L⁻¹; pH 6.6–7.85; salinity 17.5–28.5; temperature 28.3–31.55 °C). Mangrove vegetation diversity indices were low to moderate (0.05–1.12) across restoration ages. These findings indicate that restored mangrove stands can function as aquatic habitat supporting diverse fish assemblages while maintaining basic water-quality conditions, reinforcing the role of mangrove restoration in sustainable coastal development and the Sustainable Development Goals (SDGs).
Keywords:
mangrove restoration
; fish assemblage
; aquatic biodiversity
; water quality
; estuary
; coastal resilience
; blue carbon
; sustainable development goals
; Rhizophora
; Mugilidae
1. Introduction
Mangroves are among the most productive ecosystem on Earth, providing unique habitats that support many species and offer valuable services to humans [1,2,3]. Known for their high biodiversity [4], mangroves serve as critical nurseries and food sources [5] by supplying organic matter and nutrients that sustain aquatic communities [6,7,8]. Furthermore, mangroves play an important component in the trophic chain [9] and are key sources of food for fish and invertebrates [10]. However, some of the mangrove habitats have been lost due to human activities [11,12]. In Indonesia, this decline is largely due to the expansion of aquaculture ponds [13,14] and oil palm plantations [15,16] reported that up to 80% of mangrove losses in six Southeast Asian countries were attributed to human activities, driven by regional pressures to expand aquaculture for economic development. In addition, [17] noted that Indonesia is one of the largest contributors to global carbon emissions due to mangrove conversion into ponds. Restoring degraded mangrove ecosystems is critical. According to [18], the rehabilitation and restoration of mangrove ecosystems are vital for the ecology, economy, and well-being of coastal communities. The restoration and rehabilitation program are a step to build ecosystem services to carry out community-based biodiversity conservation [19]. These initiatives must consider hydrology, mangrove vegetation, and associated fauna. While restoration aims to return habitats to their natural conditions, rehabilitation focuses on optimizing ecosystem services beyond biodiversity. Long-term success depends on many aspects [20], highlighting the importance of well-managed mangrove restoration in coastal development [21]. However, natural challenges can affect restoration efforts, as coastal areas are subject to environmental adaptations [22]. Mangroves have a profound effect on surrounding waters. For example, [23] demonstrated a positive relationship between mangrove coverage and shrimp catches in Indonesia, the Philippines, Australia, and Mexican. In addition, estuarine fish in the tropics are closely linked to mangrove habitats [24]. The restoration of mangroves is critical, as these ecosystems provide important habitats for various species [25]. In addition, mangrove litter becomes the basis of the food web, breaking down into detritus which ultimately serves as food source within the ecosystem. Given the importance of the ecological economic benefits of mangrove restoration and its role in supporting biodiversity, this study aims to determine the diversity of fish species in restored mangrove areas. The findings will help for mangrove restoration activities worldwide, especially in Indonesia.
Beyond local ecological objectives, mangrove restoration is increasingly framed as a pathway to deliver multiple Sustainable Development Goals (SDGs), including climate action (SDG 13), life below water (SDG 14), life on land (SDG 15), resilient coastal settlements (SDG 11), and livelihoods and food security (SDGs 1–2 and 8) (United Nations, 2015). Linking restoration outcomes to these development targets can strengthen the rationale for continued investment and help identify monitoring indicators that capture both ecological recovery and coastal-development benefits.
2. Materials and Methods
2.1. Study Area
The research was conducted in restored mangrove areas located in Lubuk Kertang, Pangkalan Brandan, North Sumatra (Figure 1). Sampling of aquatic fauna was carried out in June, August, and December 2021.
2.2. Procedure
- Observation of mangrove vegetation; Mangrove vegetation was observed at three locations, corresponding to restoration years: 2009, 2011, and 2013, all situated in North Sumatra. In each site, 10 plots were established using a combination of stripe and gridline methods [26] to analyze the vegetation.
- Observation of aquatic fauna; Fish collection was carried out in the waters surrounding the restored mangrove forests in Lubuk Kertang. Sampling was carried in June, August, and December 2021 at the locations shown in Figure 1. Local fishing methods were employed using boats and ‘kedera’ nets, which have a length of 150 m and a width of 1.5 m. The mesh size of the nets consists of three layers, namely 1, 1.5, and 1.6 inches. After collecting the specimens, they were identified based on [27] and [28], with species verification through FishBase.org.
- Water quality measurement; Water quality parameters were measured for each fish sampling period in June, August, and December 2021. These parameters measured included; DO (mg/L), pH, salinity (ppm), and temperature (ºC). The data was compiled and analyzed using Microsoft Excel 2016.
2.3. Data Analysis
Data analysis included the composition of fish species, diversity index (H'), evenness (J), and dominance (C). The species composition was analyzed by grouping the fish species and determining their percentage relative to the total number of fish sampled. The diversity index (H') is calculated following the Shannon and Wiener formula [29] where ni is the number of species i and N is the total number of all fish.
Evenness index (J) follows Pielou's equation [30], with the analysis that H' is the Shannon-Wiener diversity index compared to Ln number of species (S), which is presented in the formula as below:
While the analysis to obtain the dominance index (C) follows [31], C is the dominance index, ni is the number of species i and N is the total number of all fish. The equal follows the formula as below:
3. Results
3.1. Density and Diversity of Mangrove Vegetation
Mangrove restoration in Lubuk Kertang, Pangkalan Brandan, Langkat, North Sumatra has progressed through several stages, with reforestation efforts initiated in in 2009, followed by additional plantings in 2011 and 2013 at different locations within the Lubuk Kertang area. Observations at these three locations described the different densities of mangrove vegetation. The restoration in 2009 showed well progress,indicated by the presence of Rhizophora apiculata, Sonneratia alba, Xylocarpus granatum, and Ceriops tagal. The restoration area in 2011 had two species of mangroves, namely Rhizophora apiculata (planting) and Sonneratia alba. Likewise, the 3rd restoration area, namely the restoration in 2013 has 2 species, namely Rhizophora apiculata (planting) and Sonneratia alba. The restoration age of the study sites ranged from 7 to 11 years, with varying densities at different growth stages (Figure 2). Tree growth rates were the lowest compared to saplings and seedlings. Tree growth rates in the 2009 restoration area for Rhizophora apiculata was 350 ind/ha and Sonneratia alba was found with a density of 190 ind/ha. While Xylocarpus granatum and Ceriops tagal were not found at the tree level, the 2011 restoration area Rhizophora apiculata and Sonneratia alba were found with a density of 130 ind/ha. The 2013 restoration area tree level was found to have a higher density than the 2011 restoration area.
The sapling growth rate in all restoration areas had a higher density than the other tree growth rates. However, this did not occur at the level of sapling growth in the species Xylocarpus granatum and Ceriops tagal. In both species, the seedling level had a higher density than the sapling growth rate. The seedling growth rate were highest in 2013 restorations area, with densities of 4750 ind/ha (Rhizophora apiculata) and 3000 ind/ha (Sonneratia alba). The growth rate of saplings and seedlings in the 2009 and 2011 restoration areas were lower.
The mangrove vegetation diversity index was low across all restoration areas, ranging from 0.05 to 1.12. The highest diversity index was observed in the 2013 restoration (Figure 3). The average diversity indices were 0.62 (tree growth rate), 0.29 (sapling growth rate), and 0.77 (seedling growth rate). The 2013 restoration area had the highest diversity index for both saplings and seedlings.
3.2. Diversity and Composition of Fishes
A total of 828 fish individuals were sampled, representing 44 fish species from the class Actinopteri, which consists of 17 orders, 27 families, 39 genera (Table 1). The fish species composition as follows; total of all species of fish caught 4.71% were found with a percentage of less than 0.6% (20 species), 12.56% were found in the range 0.6% to 1.93% (13 species) and 79.23% were found with a percentage between 2.66% to 25.36% (11 species). Figure 4 shows the species composition based on the percentage of fish species caught during the study (percentages less than 0.60% are not shown in the figure).
The most common fish species were from the Engraulidae family (321 ind), which consisted of 3 genera, namely Setipinna, Stolephorus, and Thryssa. of the 340 ind observed about 74.76% is the genus Stolephorus, the rest are the genera Thryssa and Setipinna. During the study, the Mugilidae family found about 230 individuals consisting of 4 genera with the most genera being Crenimugil (46.52%) and Ellochelon (36.52%), the remaining 16.96% consisting of the genera Mugil and Planiliza. Another family that has a higher number than other families is Ambassidae which consists of 1 genus (3 species) with a total of 80 individuals. There were 20 species of fish with a number between 1-4 individuals during the study, the rest were found to be more than 5 individuals. Several families were found (Ephippidae, Scatophagidae, Siganidae, Channidae, Sphyraenidae, Toxotidae, Xiphiidae, Pristigasteridae, Sciaenidae, Gerreidae, Gobiidae, Chanidae, Mullidae, Cynoglossidae, Scombridae, Synbrachidae and Tentraodontidae) consisting of 1 genus and one species.
The total number of fish species present was the presence of 20 species which made up 4.71% of the total fish species found, namely Anodontostoma chacunda, Atule mate, Channa striata, Channos channos, Cynoglossus lingua, Ephippus argus, Gerres filamentosus, Hyporhamphus quoyi, Johnius tracheyephalus, Lagochepalus inermis, Lutjanus sp., Monopterus albus, Muraenesox cinereus, Periophthalmodon schlosseri, Planiliza subviridis, Scomberoides sp., Selaroides leptolepis, Sphyraena barracuda, Upeneus sulphureus, Xiphias sp. While the fish percentages with 16.06% and 79.23% are presented in Figure 4.
The fish composition of 16.06% was composed of 13 species of fish, namely: Ambassis gymnocephalus, Ambassis vachellii, Dussumieria acuta, Hemiramphus sp., Herklotsichthys dispilonotus, Lutjanus ehrenbergii, Mugil cephalus, Mugil cephalus, Rastrelliger kanagurta, Scatophagus argus, Secutor interruptus, Setipinna tenuifilis, Siganus canaliculatus, Toxotes jaculatrix. The largest composition is 79.23%, consisting of 11 species which include Ambassis nalua, Crenimugil buchanani, Crenimugil seheli, Dussumieria acuta, Eublekeeria jonesi,, Ellochelon vaigiensis, Ilisha striatula, Planiliza planiceps, Stolephorus indicus, Stolephorus waitei, Thryssa hamiltonii.
3.3. Fish diversity Index
The diversity index (H') of fish during the study period (June, August, and December 2021) ranged at a medium level (Figure 5). The average diversity index was 2.15, evenness 0.83, and dominance at 0.17. June exhibited the highest diversity, while August had the lowest. The evenness and dominance indices remained relatively stable across the months, with a slight increase in dominance in August 2021.
3.4. Water Quality
Water quality parameters namely measurements were carried out including dissolved oxygen (DO), pH, salinity, and temperature are presented in Figure 6. Dissolved oxygen concentration ranged from 3.5-5.15 mg/L, pH ranged 6.6-7.85, salinity ranged 17.5 -28.5 ppt, and temperature ranged 28.3 - 31.55°C.
4. Discussion
4.1. Density and Diversity of Mangrove Vegetation Restoration
The density of mangrove vegetation in the restoration area as presented in Figure 2A, B and C, reflects the progress of the restoration efforts that began in 2009. The dominant species, R. apiculata, has consistently been prevalent, as observed in the restoration efforts in 2011 and 2013. This condition is expected, as R. apiculata is commonly used in [32] mangrove replanting projects due to its adaptability to tidal water channels and soft, muddy soils [33,34] it in addition, its high survival rate and abundance in Southeast Asia and Australia further justify its use in restoration [35,36].
The mangrove density in 2009 was higher compared to 2011 and 2013, likely due to the earlier planting and subsequent growth. Sonneratia alba, which was less dense compared to Rhizophora apiculata, is also noted for its wide distribution in the Asian region and grown in estuarine areas [37].
The similar density in 2011 for sapling and seedling (2000 ind/ha for Rhizophora apiculata and 3000 ind/ha for Sonneratia alba) suggests comparable restoration conditions, with environmental factors influencing the outcomes [38], in which the combination of light and salinity will affect exogenous gibberellic acid (GA3).
By 2013, the tree density of Rhizophora apiculata <2009 and > 2011, indicating a natural progression in vegetation density. In contrast to Sonneratia alba, the density of trees in restoration forest was 2009>2011>2013. This is presumably because the age of vegetation in 2009 was longer so that the number of trees was more numerous and denser. Even though if we look at the diameter of the trees, the 2009 restoration forest is considered to have a lower tree diameter compared to other restoration years. Restoration forest in 2013 has a range between 10.83-16.88 cm, restoration in 2011 with a range of 10.19-18.15 cm, and restoration forest in 2009 is in the range of 10.19-16.24 cm.
4.2. Diversity and Composition of Fishes
The study identified 44 species comprise 27 families, with Stolephorus being the most abundant (74.76%), particularly Stolephorus indicus and Stolephorus waitei. The high abundance of this genus noted during the August 2021 sampling.This genus includes Engraulidae, and according to [39] and [40], they are often found in flocks in coastal areas, although some cmove into brackish waters to find food or lay eggs, whie others settle permanently. Stolephorus has a wide distribution in tropical areas, such as in the Indo-West Pacific [40]. In addition, various locations have been recorded in India [41,42], Pakistan [43], Taiwan [44], Turkey [45], and the Mediterranean Sea, including Israel [46].
The abundance of this genus at the research location is similar to the mangrove area of Segara Anakan Cilacap, Central Java [47], where it is more abundant compared to other species. These two species are thought to be found in large numbers because the mangrove waters serve as a feeding ground. This suggests that the mangrove area of the Lubuk Kertang restoration can support the the genus well and provide sufficient food. This assumption is well-founded, as crustacean groups are also found around the fishing grounds (not presented in this study). Research conducted in the Purworejo mangrove area highlights the richness of the crustacean group, which is around 19 species recorded [48]. It is suspected that one of the reasons for the presence of Stolephorus in mangrove waters is that these areas are rich in food sources.
The relationship between the presence of fish and food sources indicates that mangrove restoration forests can provide essential food for fish. This hypothesis is supported by [49] who reported that this group of species primarily feeds on crustaceans (Copepoda, Lucifer, Mysids, Acetes, and Amphipoda) and mollusks (Bivalves and gastropods), small fish, Tintinnida and Dinoflagellates. Furthermore, Stolephorus is recognized as a crustacivore fish group [50].
Another fish species often found in te study area is from the Mugilidae. commonly known as mullet. The Mugilidae caught during the study belonged to the Crenimugil, Ellochelon, Mugil, and Planiliza. The Mugilidae group is commonly found in mangrove areas as reported in the Kedungmalang Jepara mangrove region [51]. Mugilidae found in the study accounted for 27.66%, which is 11% lower than the Engraulidae. Based on the distribution patterns, the order Mugilliformes is distributed in coastal and brackish waters, with 20 genera and 75 species [52]. These results suggest that the habitat in the study is well-suited for the Mugilidae group.
It is hypothesized that the mangrove restoration area can meet the ecological needs of these fishes. This assumption is supported by [53], who stated that muddy substrate with microphytobenthos is positively correlated with fish density in this group. In addition, factors such as proximity to rivers, water depth, water clarity, and accessibility of food sources from benthic microalgae are influential
Other families (25), apart from the two families discussed above, had occurrence rates below 9%. However, the existing diversity suggests that restoring degraded mangrove ecosystems through restoration efforts can be an alternative for improving the diversity of surrounding aquatic biota. This research highlights the importance of restoration programs for the diversity of aquatic biota, particularly fish, as mangrove ecosystems provide essential food sources for surrounding species.
As reported by [54], analysis of the stomach contents of fish caught in the Lubuk Kertang mangrove ecosystem shows that these fish consume a higher proportion of plant fragments and litter, compared to other food sources. This underscores the role of mangrove restoration in enhancing aquatic productivity. In the fisheries sector, mangrove restoration is known to increase aquatic productivity. [55] and [56] also stated that fish productivity and resources are likely to increase with the expansion and productivity of mangroves areas.
Changes in mangrove ecosystem, such as deforestation, can significantly affect aquatic biota communities [57], as mangroves are an important ecosystem in coastal areas, and form the basis for food webs in both aquatic and terrestrial ecosystems [58]. Therefore, measuring the productivity of mangrove ecosystems should include assessments of fisheries productivity [59], as was done in this research.
4.3. Diversity Index of Fishes
The average results of the analysis related to the fish diversity index in the waters of the restored mangrove ecosystem indicated a medium level (2.15), reflecting the complex interactions between species in the area. Monthly variations in the diversity, evenness and dominance indices during the study were consistently in the medium range. Community stability can be described based on biodiversity [60]. The impact of land-use change at the ecosystem level is also believed to influence the distribution of exotic species, as the main factors shaping the community diversity patterns are related to trophic niches - processes like nutrition cycling or energy transfer [61].
The findings of this research further support the notion that mangrove forest restoration plays a significant role in enhancing fish species diversity. The surrounding aquatic ecosystem showed a diversity index >2, and a dominance value of 0.17. This indicates that the fish species present at the study site are diverse, with no single species dominating. This assumption is also supported by the evenness index, which has a higher value in each sampling month. These findings strengthen the idea that restoration of degraded mangrove ecosystems can help restore ecological conditions, as indicated by increased fish species diversity and improved water quality. In this context, the increase in species diversity contributes to greater functional diversity, which in turn enhances ecological stability [62].
4.4. Water Quality
The water quality around the mangrove restoration area should remain within tolerable ranges for estuarine fish communities because physicochemical conditions can directly influence fish distribution, growth, and survival. In this study, dissolved oxygen ranged from 3.5 to 5.15 mg/L, indicating generally oxygenated conditions for a tropical estuary. Nevertheless, dissolved oxygen in estuaries and mangroves can vary substantially with tidal stage, temperature, and organic matter inputs; therefore, repeated sampling across tides and seasons would strengthen inference about habitat quality.
The pH at the research site ranged from 6.6 to 7.85, which is within the tolerable range for most estuarine fishes and consistent with a slightly acidic to near-neutral mangrove environment. Mangroves can influence pH dynamics through organic matter decomposition and carbonate system processes, and they may locally buffer acidity under certain conditions [63,64]. Salinity at the study site varied from 17.5 to 28.5 ppt (as recorded during field measurements), reflecting typical estuarine mixing between freshwater inputs and tidal seawater. Such salinity variability can structure fish assemblages by favoring euryhaline taxa and influencing nursery habitat use in mangrove creeks.
Temperature at the research site ranged from 28.3 to 31.55°C, within the typical range for tropical estuarine systems. Together with the observed pH and salinity ranges, these conditions suggest that the restored mangrove area provides a physiochemically suitable habitat for aquatic biota during the study period.
4.5. Implications for Sustainable Coastal Development and the SDGs
Mangrove restoration can be understood as “green” or nature-based infrastructure that supports coastal development by maintaining ecosystem functions while reducing risk and safeguarding livelihoods. In the 2030 Agenda for Sustainable Development, such multi-benefit interventions are central to achieving the SDGs through integrated social–ecological outcomes [65].
Coastal protection is a key development co-benefit of mangroves. By attenuating waves and reducing storm-surge impacts, intact or recovering mangrove belts can lower flood damages and enhance the resilience of coastal settlements and infrastructure—supporting SDG 11 (sustainable cities and communities) and SDG 13 (climate adaptation) [66,67]. Although coastal hazard reduction was not directly measured in this study, the presence of structurally developing stands (trees, saplings, and seedlings) suggests potential for increasing protective functions as restoration sites mature.
Fisheries support and food security are also closely linked to mangrove condition. Mangroves provide nursery habitats, shelter, and detrital food pathways that can enhance coastal and offshore fisheries; prioritizing conservation or restoration near human populations can maximize returns for fisheries enhancement [68]. The moderate fish diversity observed in Lubuk Kertang (44 species; H′ ≈ 2.15) is consistent with the role of restored mangroves as functional habitat for a range of estuarine and coastal taxa, and it underpins SDG 2 (zero hunger), SDG 8 (decent work and economic growth), and SDG 14 (life below water).
Mangroves are additionally important “blue carbon” ecosystems. Protecting and restoring mangroves can contribute to climate mitigation through long-term carbon storage in biomass and especially in soils, while delivering adaptation, biodiversity, and livelihood co-benefits [69]. These attributes strengthen the case for incorporating mangrove restoration into climate and development planning (e.g., NDCs, coastal zoning, and sustainable financing mechanisms), aligning local restoration outcomes with SDG 13 and SDG 15.
To better evidence and communicate coastal-development benefits, future monitoring in Lubuk Kertang could combine ecological indicators (vegetation structure, fish assemblages, and water quality) with socio-economic and risk metrics (e.g., fisheries catch or effort, shoreline change, flood exposure, and community benefits), enabling a more direct linkage between restoration performance and SDG-relevant outcomes.
5. Conclusions
This study shows that mangrove restoration in Lubuk Kertang (7–11 years after planting) supports a moderately diverse fish assemblage (44 species; mean H′ ≈ 2.15) under estuarine water-quality conditions, alongside low-to-moderate vegetation diversity across restoration ages. These ecological outcomes suggest that restored stands can function as productive habitat for aquatic biota and can contribute to broader coastal development objectives. In particular, maintaining and restoring mangroves can help sustain fisheries resources and food security (SDGs 2, 8, 14), enhance coastal resilience to erosion and storm hazards as natural infrastructure (SDGs 11, 13), and contribute to climate-mitigation potential through blue-carbon storage (SDG 13), while reinforcing biodiversity conservation (SDG 15).
6. Recommendation
To ensure the continued success of mangrove restoration in Lubuk Kertang, long-term monitoring should be maintained for both vegetation structure and associated aquatic biota, supported by consistent water-quality measurements across seasons and tidal conditions. Monitoring could also incorporate indicators directly relevant to coastal development (e.g., fisheries effort/catch trends, shoreline change, and community benefits), and be linked with participatory management involving local communities, government agencies, NGOs, and the private sector to support SDG-aligned outcomes and sustained financing
Author Contributions
Conceptualization, O.O. and A.S.; methodology, O.O.; software, O.O.; validation, O.O.; formal analysis, O.O.; investigation, O.O. and A.S.; resources, M.A.; data curation, O.O.; writing—original draft preparation, O.O. and A.S; writing—review and editing, M.M., M.A and S.N.; visualization, O.O.; supervision, M.A. and S.N.; project administration, AS.; funding acquisition, O.O. All authors have read and agreed to the published version of the manuscript.
Funding
Ministry of Research and Technology/National Research and Innovation Agency of the Republic of Indonesia for funding this research year 2021.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors confirm the data supporting the findings are available within this paper.
Acknowledgments
The authors would like to thank the Ministry of Research and Technology/National Research and Innovation Agency of the Republic of Indonesia for funding this research year 2021 and all the field teams who assisted during the sampling in Lubuk Kertang, Langkat, North Sumatera.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Fontalo-Herazo, M.; Piou, C., Vogt, J., Saint-Paul, U.; Berger, U. Simulating harvesting scenarios towards the sustainable use of mangrove forest plantations. Wetlands Ecol Manage. 2011, 19, 397-407. [CrossRef]
- Carugati, L.; Gatto, B.; Rastelli, E.; Martire, M.L.; Coral, C.; Greco, S.; Danovaro, R. Impact of mangrove forests degradation on biodiversity and ecosystem functioning. Scientific Reports, 2018, 8:13298, 1-11. [CrossRef]
- Romanach, S.S.; DeAngelis, D.L., Koh, H.L., Li, Y., Teh, S.Y., Barizan, R.S.R., & Zhai, L. Conservation and restoration of mangroves: global status, perspectives, and prognosis. Ocean and Coastal Management, 2018, 154, 72-82. [CrossRef]
- Husain, P.; Al Idrus, A.; Ihsan, M.S. The ecosystem services of mangrove for sustainable coastal area and marine fauna in Lombok, Indonesia: A Review. Jurnal Inovasi Pendidikan dan Sains, 2020, 1(1): 1-7. [CrossRef]
- Adite, A.; Toko, I.I.; Gbankoto, A. Fish assemblages in the degraded mangrove ecosystems of the coastal zone, Benin, West Africa: implication for ecosystem restoration and resources conservation. Journal of Environmental Protection, 2013, 4, 1461-1475. [CrossRef]
- Hadwen, W.L., Russell, G.L., & Arthington, A.H. Gut content and stable isotope-derived diets of four commercially and recreationally important fish species intermittently open estuaries. Marine and Freshwater Research, 2007, 58(4). https://ui.adsabs.harvard.edu/link_gateway/2007MFRes..58..363H/doi:10.1071/MF06157.
- Zagars, M.; Ikejima, K.; Kasai, A.; Arai, N.; Tongnunui, P. Trophis characteristics of a mangrove fish community in Southwest Thailand: important mangrove contribution and intraspecies feeding variability. Estuarine Coastal and Shelf Science, 2013, 119, 145-152. [CrossRef]
- Abrantes, K.G.; Johnston, R.; Connoly, R.M.; Sheaves, M. Importance of mangrove carbon for aquatic food webs in wet-dry tropical estuaries. Estuaries and Coasts, 2015, 38, 383-399. [CrossRef]
- Muro-Torres, V.M.; Amezcua, F.; Soto-Jimenez, M.; Balart, E.F.; Serviere-Zaragoza, E.; Green, L.; Rajnohova J. Primary sources and food web structure of a tropical wetland with a high density of mangrove forest. Water, 2020, 12, 3105, 18 p. [CrossRef]
- Abrantes, K.; Sheaves, M. Incorporation of terrestrial wetland material into aquatic food webs in a tropical estuarine wetland. Estuarine. Coastal and Shelf Science, 2008. 80, (3), 401-402. [CrossRef]
- Romanach, S.S.; DeAngelis, D.L.; Koh, H.L.; Li, Y.; Teh, S.Y.; Barizan, R.S.R.; Zhai, L. Conservation and restoration of mangroves: global status, perspectives, and prognosis. Ocean and Coastal Management, 2018, 154, 72-82. [CrossRef]
- Goldberg, L.; Lagomasino, D.; Thomas, N.; Fatoyinbo, T. Global declines in human-driven mangrove loss. Glob Change Biol, 2020, 26, 5844-5855. [CrossRef]
- Onrizal, Mangrove: Important natural resources under threat. Wanamina, 2013, 3, (1), 1-9. https://onrizal.wordpress.com/2013/12/24/mangrove-sumberdaya-alam-penting-yang-terancam/ [Indonesian].
- Ilman, M.; Dargusch, P.; Dart, P.; Onrizal. A historical analysis of the drivers of loss and degradation of Indonesia’s mangroves. Land Use Policy, 2016, 54, 448-459. [CrossRef]
- Richards, D.R.; Friess, D.A. Rates and drivers of mangrove deforestation in Southeast Asia, 2000–2012. Proceedings of the National Academy of Sciences, 2016, 1132, 344-349. [CrossRef]
- Goldberg, L.; Lagomasino, D.; Thomas, N.; Fatoyinbo, T. Global declines in human-driven mangrove loss. Glob Change Biol, 2020. 26, 5844-5855. [CrossRef]
- Onrizal.; Thoha, A.S.; Ahmad, A.G.; Mansor, M. Mangrove loss drives global warming. In Proceedings of the International Conference of Science, Technology, Engineering, Environmental and Ramification Researches, 2018, 102-105. [CrossRef]
- Ellison, A.M.; Felson, A.J ; Friess, D.A. Mangrove rehabilitation and restoration as experimental adaptive management. Front. Mar. Sci, 2020, 7(327):1-19. [CrossRef]
- Andradi-Brown, D.; Howe, C.; Mace, G.; Knight, A.T. Do mangrove forest restoration or rehabilitation activities return biodiversity to pre-impact levels, Environmental Evidence 2013, 2:20. http://www.environmentalevidencejournal.org/content/2/1/20.
- Ellison, A.M.; Felson, A.J.; Friess, D.A. Mangrove rehabilitation and restoration as experimental adaptive management. Front. Mar. Sci, 2020, 7(327):1-19. [CrossRef]
- Onrizal; Thoha, A.S.; Ahmad, A.G.; Mansor, M. Mangrove loss drives global warming. Proceedings of the International Conference of Science, Technology, Engineering, Environmental and Ramification Researches. 2018, 1, 102-105, Medan, Indonesia, 30 August 2018. [CrossRef]
- Goldberg, L.; Lagomasino, D.; Thomas, N.; Fatoyinbo, T. Global declines in human-driven mangrove loss. Glob Change Biol, 2020, 26, 5844-5855. [CrossRef]
- Sukardjo, S. Fisheries associated with mangrove ecosystem in Indonesia: a view from a mangrove ecologist. Biotropia, 2004, 23, 13‐39. https://journal.biotrop.org/index.php/biotropia/article/view/201.
- Blaber, S.J.M. Mangrove and fishes: issues of diversity, dependence, and dogma. Bulletin of Marine Science Miami, 2007, 80(3), 457-472, https://www.ingentaconnect.com/content/umrsmas/bullmar/2007/00000080/00000003/art00004#.
- Negelkerken, I.; Blaber, S.J.M.; Bouillon, S.; Green, P.; Haywood, M.; Kirton, L.G.; Meynecke, J.O.; Pawlik, J.; Penrose, H.M.; Sasekumar, A.; Somerfield, P.J. The habitat function of mangrove for terrestrial and marine fauna: A review. Aquatic Botany, 2008, 89(2), 155-185. [CrossRef]
- Onrizal. Guide to the introduction and analysis of mangrove forest vegetation, 2008, https://onrizal.wordpress.com/2008/11/26/panduan-pengenalan-dan-analisa-vegetasi-hutan-mangrove/ access on 28 Dec 2025. [Indonesian].
- Kottelat, M.; AJ Whitten, S.N.; Kartikasari, S; Wirjoatmodjo. Freshwater Fishes of Western Indonesia and Sulawesi. Peripilus Edition. (HK) Ltd. And EMDI. Indonesia, 1993; 221 p.
- Allen, G. Marine fishes of South East Asia. Western Australian Museum. Perth. 1997; 147 p.
- Shannon, C.E.; Weiner, W. The Mathematical theory of communication. Urbana. IL: University of Illinois Press Urbana. 1949, 117p.
- Pielou, E.C. Species diversity and pattern diversity in the study of ecological succession. J Theore Biol, 1966, 3, 131-144. [CrossRef]
- Odum, E. P. Fundamentals of ecology (3rd Edition). WB Soundress Co., University of Minnesota, 1971; 574 p.
- Basyuni, M.; Habib, A.A.; Slamet, B.; Sulistiyono, N.; Putri, L.A.P.; Bimantara, Y.; Yusraini, E.; Lesmana, I. Assessment of one-year mangrove reforestation using Rhizophora apiculate seedling in Lubuk Kertang Village, North Sumatra. AEFS. IOP Conf. Series: Earth and Environmental Science, 2019. 260:012114. [CrossRef]
- Setyawan, A.D.; Ulumuddin, Y.I.; Ragayan, P. Review: mangrove hybrid of Rhizophora and its parental species in Indo-Malayan region. Nusantara Bioscience, 2014. 6(1), 69-81. [CrossRef]
- Mustika, D.I.; Rusdiana, O.; Sukendro, A. The development of Rhizophora apiculate at mangrove nursery of Muara Teluk Naga Village, Tangerang District, Banten. Bonorowo Wetlands, 2014, 4(2):108-116. [CrossRef]
- Giesen, W.; Wulffraat, S.; Zieren, M.; Scholten, L. Mangrove guidebook for Southeast Asia. FAO and Wetland International. Thailand, 2007; 769 p.
- Rahmat, D.; Fauziyah; Sarno. Seedling growth of Rhizophora apiculate at mangrove restoration area Sembilang National Park, South Sumatera. Maspari Journal, 2015, 7(2), 11-18.
- Qui, S.; Zhou, R.C.; Li, Y.Q.; Havanond, S.; Jaengjai, C.; Shi, S.H. Molecular evidence for natural hybridization between Sonneratia alba and S. Griffithii. Journal of Systematics and Evolution. 2008, 46(3), 391-395. [CrossRef]
- Kathiresan K & Rajendran N. Growth of mangrove (Rhizophora apiculata) seedlings as influenced by GA3, light, and salinity. Rev. Biol. Trop, 2002, 50(2), 525-530, https://pubmed.ncbi.nlm.nih.gov/12298283/.
- Whitehead, P.J.P.; Nelson, G.J.; Wongratana, T. FAO Fisheries Synopsis No. 125. Vol 7 part 2. Species Catalogue Clupeoid fishes of the world. United Nation Development Program, FAO, Rome, 1988; 176 pp.
- Dalzell, P.J.; A.D. Lewis. The fisheries biology and ecology of anchovy genera Stolephorus and Encrasicholina in the Indo-West Central Pacific Region. Fisheries Centre Research Reports 32(1). Institute for the Oceans and Fisheries, University of British Columbia, 2023, 107 p.
- Hata, H.; Lavoué, S.; Motomura, H. A New Species of the Bengal Spined Anchovy Stolephorus from the Eastern Indian Ocean and Redescription of Stolephorus dubiosus Wongratana, 1983, with Comments on the Evolution of Prepelvic Scute Numbers within Stolephorus (Clupeiformes: Engraulidae). Zoological Studies, 2022, 61, e58. [CrossRef]
- Gangan, S.; Kumar, A.P.; Jahageerdar, S.; Jaiswar, A.K. A new species of Stolephorus (Clupeidae: Engraulidae) from the Bay of Bengal, India. Zootaxa, 2020, 4743 (4): 561-574. [CrossRef]
- Musarratulain; Masood, Z.; Bibi, R.; Bibi, M.; Gul, H.; Faroog, R.Y.; Jamil, N. Growth profile of an Indian anchovy species, Stolephorus indicus (van Hasselt, 1823) of family Engraulidae from Keti Bunder, Sindh, Pakistan. Global Vetrinaria, 2015, 14(4):619-622. [CrossRef]
- Young, S.; Chiu, T.S.; Shen, S.C. A revision of the family Engraulidae (Pisces) from Taiwan. Zoological Studies, 1994. 33(3):217-227.
- Dalyan, C; Yemisken, E.; Erguden, D.; Turan, C.; Eryilmaz, L. First record of the Indian Ocean anchovy Stolephorus ensularis Hardenberg, 1933 from the northeastern Mediterranean coast of Turkey. J Appl. Ichthyol, 2014, 1-2. [CrossRef]
- Fricke, R.; Golani, D.; Appelbaum-Golani, B. First record of the Indian anchovy Stolepthorus indicus (van Hasselt, 1823) (Clupeiformes: Engraulidae in the Mediterranean Sea. BioInvasions Records, 2015, 4, 293-297. [CrossRef]
- Suprastini; Ardli, E.R.; Nuryanto, A. Diversitas dan distribusi ikan di Segara Anakan Cilacap. Scripta Biologia, 2014, 1(2), 147-15. [CrossRef]
- Rahayu, S.M.; Wiryanto; Sunarto. Biodiversity of crustacea in mangrove area, Purworejo regency. Central Java. J Sains Dasar, 2017, 6(1):57-65. [CrossRef]
- Gopal, R.V.; Ramasubramanian, V.; Santhosh, B.; Abraham, K.M. Food spectrum and dietary preferences of the Indian anchovy Stolephorus indicus (van Hasselt, 1823) from Thiruvananthapuram coast, Kerala. Indian J Fish, 2018. 65(1): 15-19. [CrossRef]
- Syafei, L.S.; Siregar, R.S.; Rahardjo, M.F.; Simajuntak, C.P.H. Diet composition and trophic niche similarities of engraulid fishes in Pabean bay, Indramayu, Indonesia. IOP Conf. Series: Earth and Environmental Science, 2020, 404, 012056. [CrossRef]
- Redjeki, S. Komposisi dan kelimpahan ikan di ekosistem mangrove di Kedungmalang Jepara. Ilmu Kelautan, 2013, 18(1): 54-60.
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the world. 5th. Ed. H,oboken (NJ): John Wiley and Sons, 2016, 752.
- Mwandy, A.W.; Mgaya, Y.D.; Ohman, M.C.; Bryceson, I.; Gullstrom, M. Distribution patterns of striped mullet Mugil cephalus in mangrove creeks, Zanzibar, Tanzinia. African Journal of Marine Science, 2010, 32(1), 85-93. [CrossRef]
- Darmarini, A. S.: Desrita, D.; Onrizal, O. Kebiasaan makanan beberapa jenis ikan di ekosistem mangrove Lubuk Kertang, Sumatera Utara. Jurnal Kelautan Tropis, 2023, 26(2), 293-300. [CrossRef]
- Hutchinson, J.; Spalding, M.D.; zu Ermgassen, P.S.E. The role mangrove in fisheries enhancement. The Nature Conservancy and Wetlands International in 2014. University of Cambridge, 2014, 54 p.
- Kalor, D.J; Indrayani, E.; Akobiarek, M.N.R. Fisheries resources of mangrove ecosystem in Demta Gulf, Jayapura, Papua, Indonesia. AACL Bioflux, 2019. 12(1): 219-229.
- Ridlo, I.A.; Kurniawan, N.; Retnaningdyah, C. Communities structure of fish in some mangrove ecosystem as a result of the restoration in Southern Beach of Malang, East Java, Indonesia. ISSMART; Journal of Physics: Conference Series, 2020, 1665, 012020. [CrossRef]
- Darmarini, A.S. ; Wardiatno, Y; Prartono T; Soewardi, K.; Samosir, A.M.; Zainuri M. Mangrove community structure in Lubuk Damar Coast, Seruway, Aceh Tamiang. Journal of Natural Resources and Environmental Management 2022, 12(1), 72-81. [CrossRef]
- Sukardjo, S. Fisheries associated with mangrove ecosystem in Indonesia: a view from a mangrove ecologist. Biotropia. 2004, 23, 13-39. [CrossRef]
- Lei, Z.; Tingting, Z.; Ziyi, Z.; Beibei, L.; Wuhui, L.; Guobao, C. Biodiversity and spatial heterogeneity of fish communities in response to geo-environmental disturbances. Estuarine, Coastal and Shelf Science, 2025, 320, 109322. [CrossRef]
- Stefani, F.; Fasola, E.; Marziali, L.; Tirozzi, P.; Schiavon, A.; Bocchi, S.; Gomarasca, S. Response of functional diversity of fish communities to habitat alterations in small lowland rivers. Biodivers Conserv, 2024, 33, 1439–1458. [CrossRef]
- Tilman, D.; Wedin, D.; Knops, J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature, 1996, 379, 718–720. [CrossRef]
- Mitra, A. Acidification of mangrove water. Mangrove Bulletin. 2018, . [CrossRef]
- Ma, Y.; Wang, W.; Gao, F.; Yu, C.; Feng, Y.; Gao, L.; Zhou, J.; Shi, H.; Liu, C., Kong, D.; Zhang, X.; Li, R.; Xie, J. Acidification and hypoxia in seawater, and pollutant enrichment in the sediments of Qi’ao Island mangrove wetlands, Pearl River Estuary, China. Ecological Indicators, 2023. 158, 111589. [CrossRef]
- United Nations. Transforming our world: the 2030 Agenda for Sustainable Development (A/RES/70/1). United Nations General Assembly, 2015, 35 p.
- Blankespoor, B.; Dasgupta, S; Lange, G.M. Mangroves as a protection from storm surges in a changing climate. Ambio, 2017, 46(4), 478–491. [CrossRef]
- IPCC. IPCC Special Report on the Ocean and Cryosphere in a Changing Climate. Cambridge University Press, 2019, 765 p.
- Hutchinson, J.; Spalding, M.D.; zu Ermgassen, P.S.E. The role mangrove in fisheries enhancement. The Nature Conservancy and Wetlands International in 2014. University of Cambridge, 2014, 54 p.
- Schindler Murray, L.; Milligan, B., et al. The blue carbon handbook: Blue carbon as a nature-based solution for climate action and sustainable development. High Level Panel for a Sustainable Ocean Economy, 2023, 84 p.
Figure 1.
Research sites in restored mangroves from 2009 (S1; blue), 2011(S2; yellow) and 2013 (S3; red) along the east coast of North Sumatra.
Figure 1.
Research sites in restored mangroves from 2009 (S1; blue), 2011(S2; yellow) and 2013 (S3; red) along the east coast of North Sumatra.
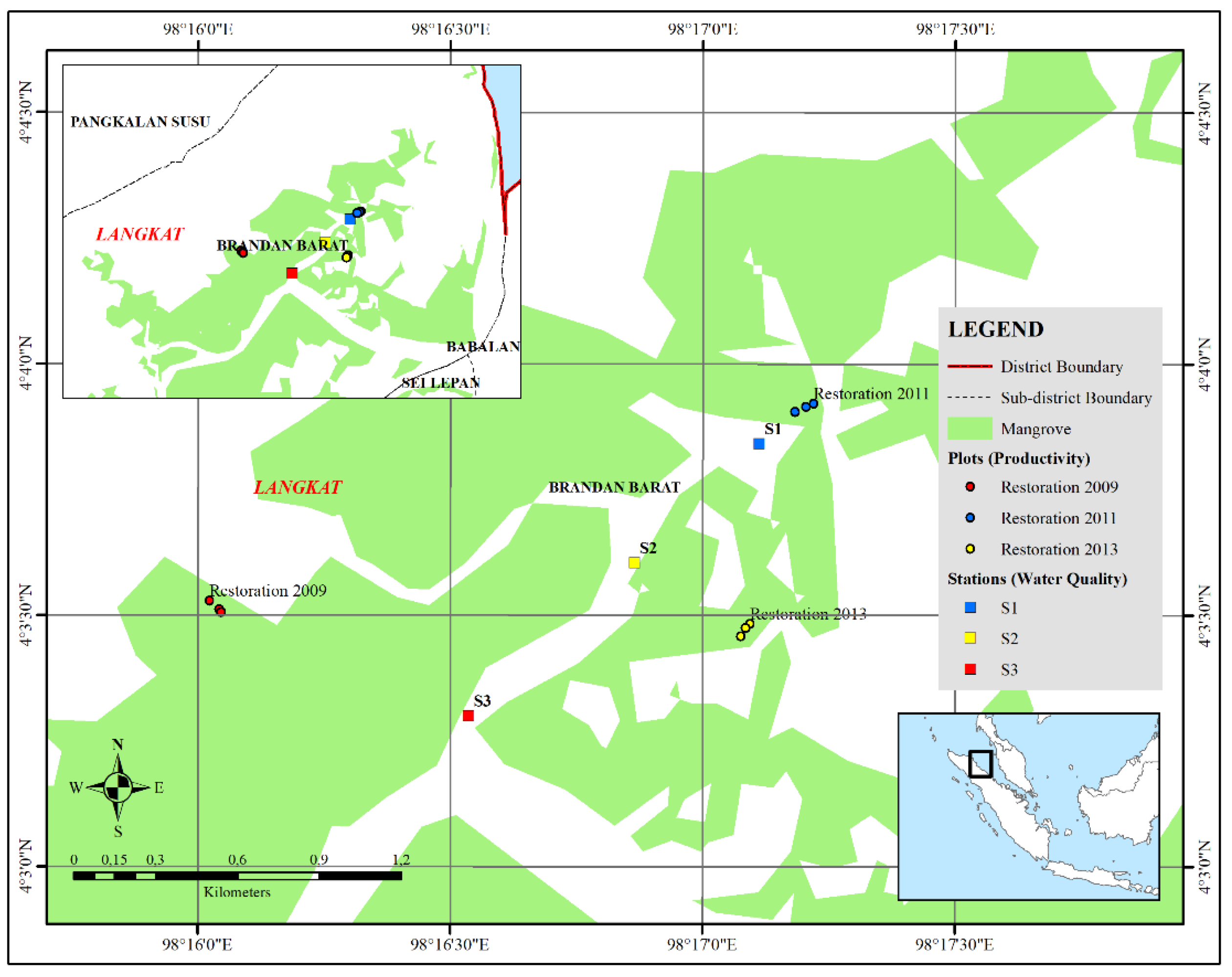
Figure 2.
Density patterns of mangrove species (ind/ha) at various growth stages in different restoration areas; A) tree growth level, B) Sapling growth level, and C) Seedling growth level.
Figure 2.
Density patterns of mangrove species (ind/ha) at various growth stages in different restoration areas; A) tree growth level, B) Sapling growth level, and C) Seedling growth level.
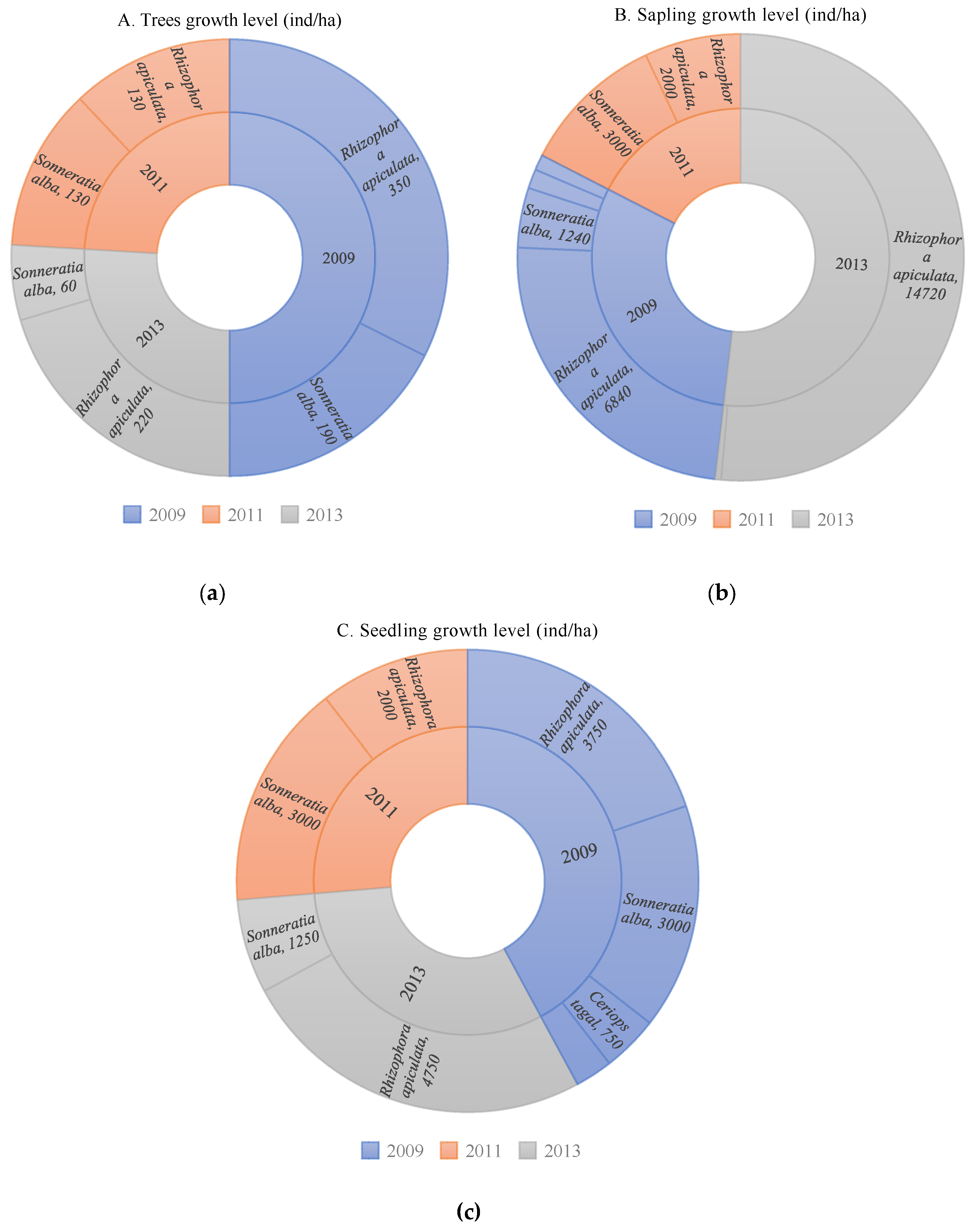
Figure 3.
Variations in mangrove vegetation diversity index based on the year of restoration.
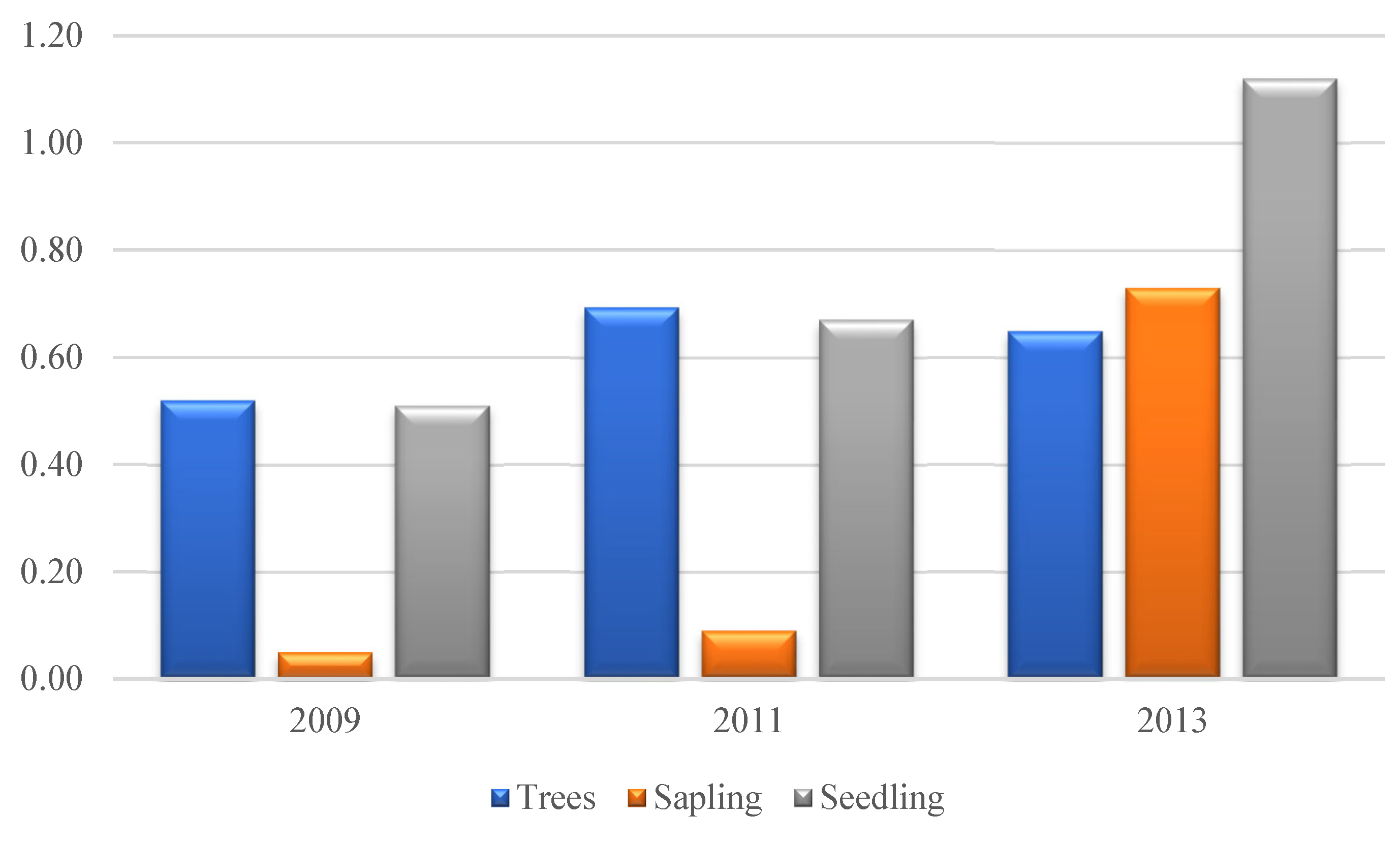
Figure 4.
Fish species composition percentages greater than 0.6% during observations in June, August, and December 2021, Ag: Ambassis gymnocephalus, An: Ambassis nalua, Av: Ambassis vachellii, Cb: Crenimugil buchanani, Crs: Crenimugil seheli, Da: Dussumieria acuta, De: Dussumieria elopsoides, Ej: Eublekeeria jonesi, Ev: Ellochelon vaigiensis, He: Hemiramphus sp., Hd: Herklotsichthys dispilonotus, Is: Ilisha striatula, Le: Lutjanus ehrenbergii, Mc: Mugil cephalus, Pp: Planiliza planiceps, Rk: Rastrelliger kanagurta, Sa: Scatophagus argus, Sc: Siganus canaliculatus, Si: Secutor interruptus, St: Setipinna tenuifilis, Sti: Stolephorus indicus, Sw: Stolephorus waitei, Tj: Toxotes jaculatrix, Th: Thryssa hamiltonii.
Figure 4.
Fish species composition percentages greater than 0.6% during observations in June, August, and December 2021, Ag: Ambassis gymnocephalus, An: Ambassis nalua, Av: Ambassis vachellii, Cb: Crenimugil buchanani, Crs: Crenimugil seheli, Da: Dussumieria acuta, De: Dussumieria elopsoides, Ej: Eublekeeria jonesi, Ev: Ellochelon vaigiensis, He: Hemiramphus sp., Hd: Herklotsichthys dispilonotus, Is: Ilisha striatula, Le: Lutjanus ehrenbergii, Mc: Mugil cephalus, Pp: Planiliza planiceps, Rk: Rastrelliger kanagurta, Sa: Scatophagus argus, Sc: Siganus canaliculatus, Si: Secutor interruptus, St: Setipinna tenuifilis, Sti: Stolephorus indicus, Sw: Stolephorus waitei, Tj: Toxotes jaculatrix, Th: Thryssa hamiltonii.
Figure 5.
Diversity index (H'), Evenness (E), and dominance (C) of fish species during the study period in June, August, and December 2021.
Figure 5.
Diversity index (H'), Evenness (E), and dominance (C) of fish species during the study period in June, August, and December 2021.
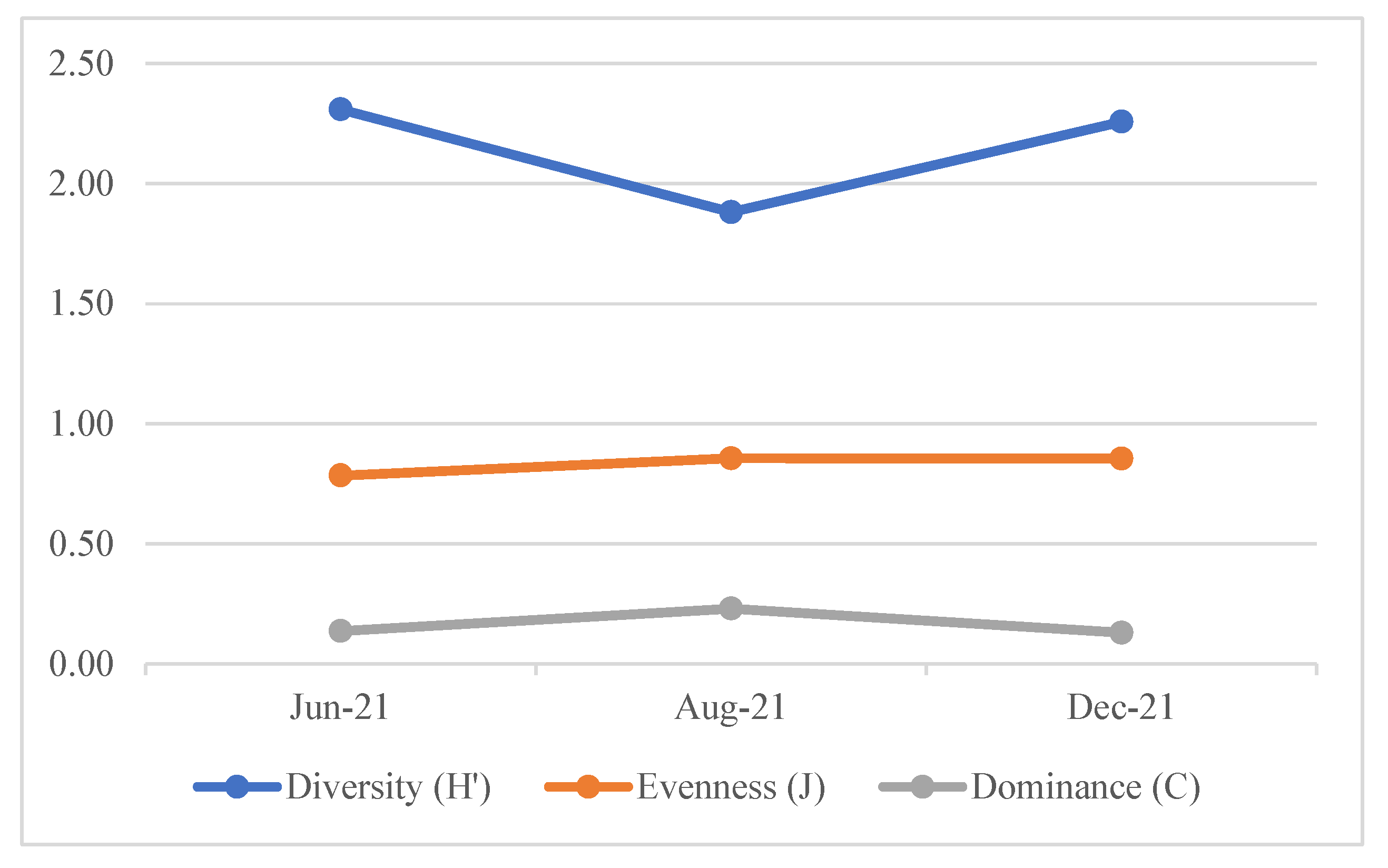
Figure 6.
Water quality parameters during the study; A) DO (mg/L), B) pH, C) salinity (ppm), D) Temperature (°C) in June, August, and December 2021.
Figure 6.
Water quality parameters during the study; A) DO (mg/L), B) pH, C) salinity (ppm), D) Temperature (°C) in June, August, and December 2021.
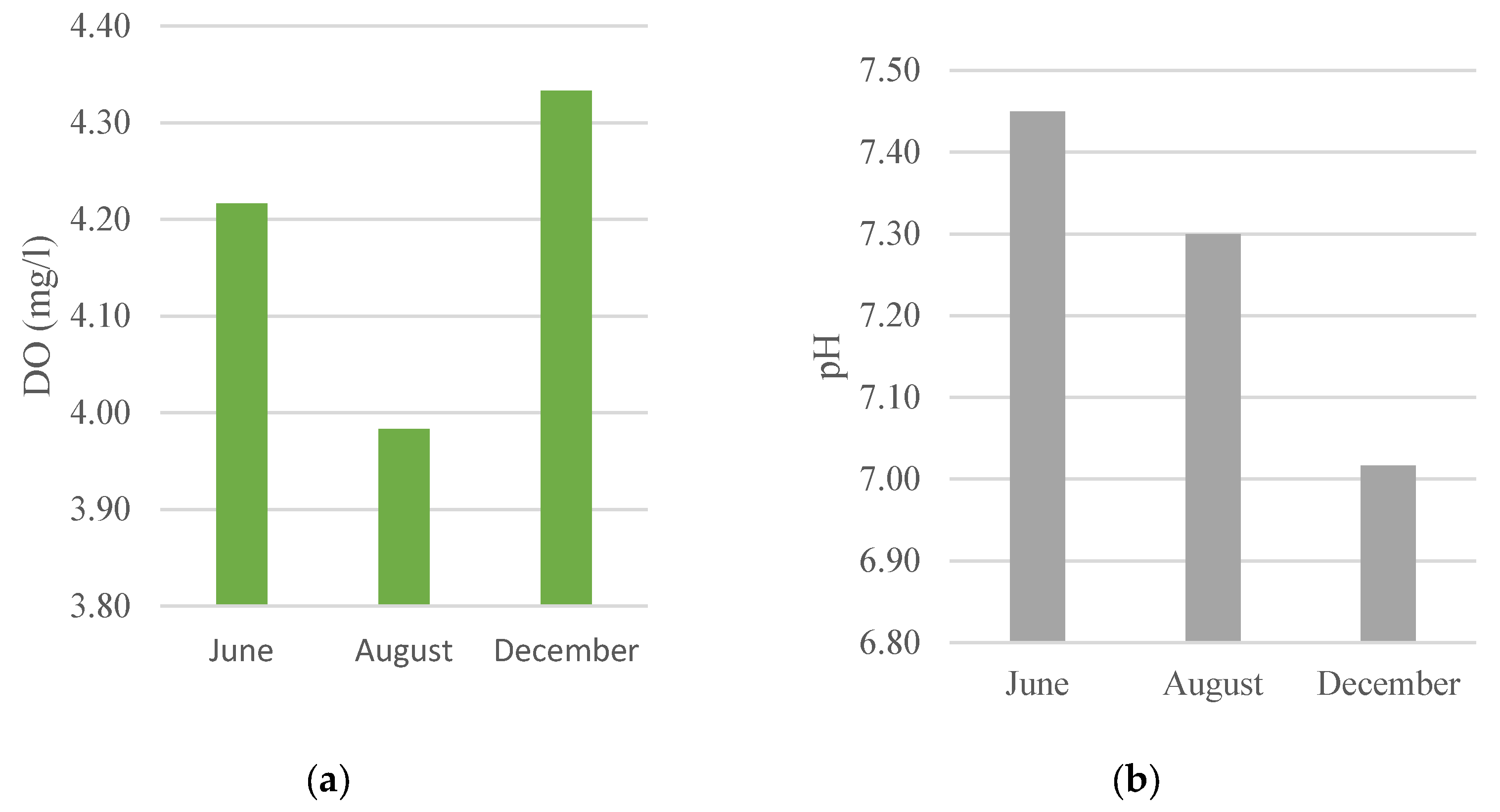
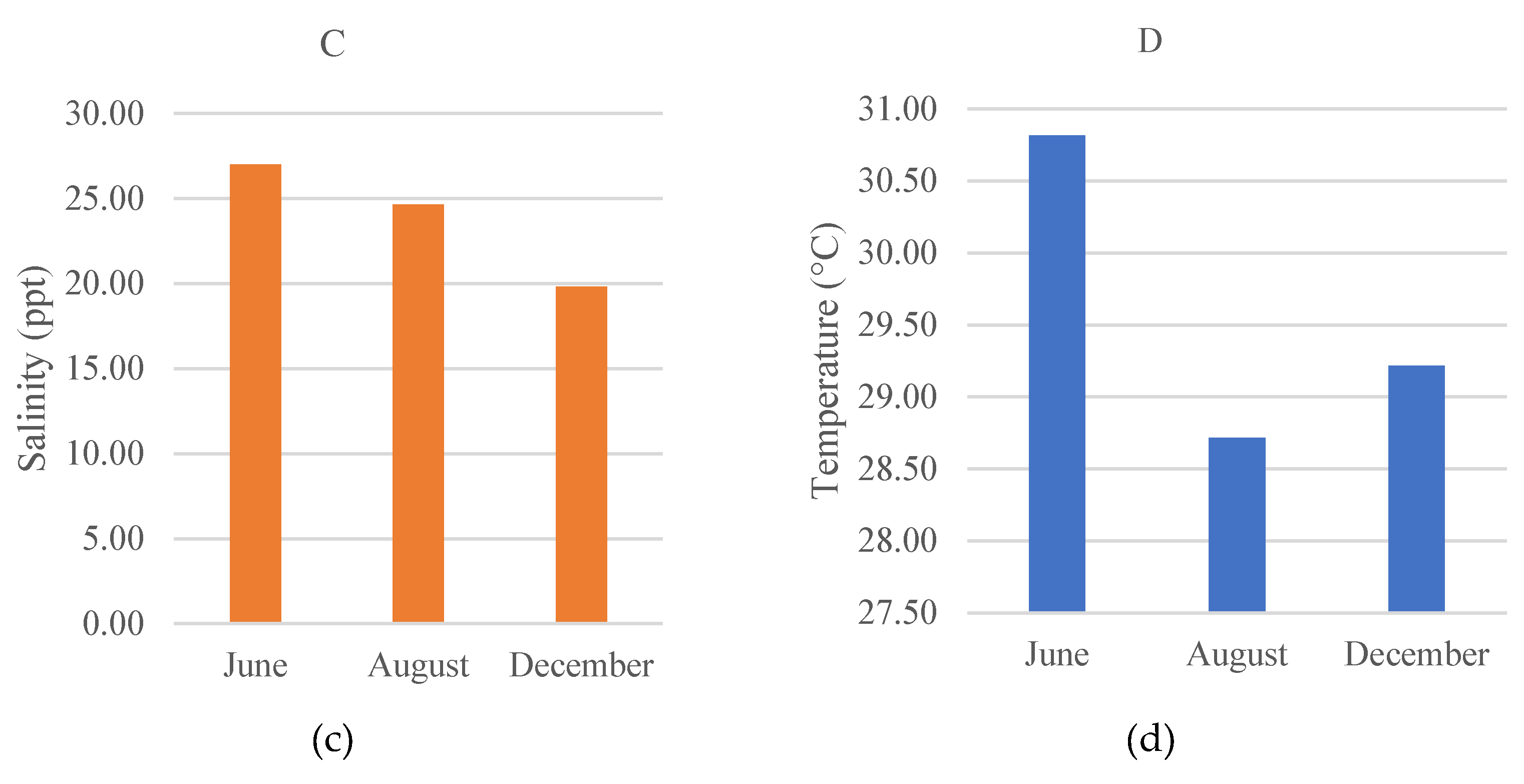

Table 2.
The diversity of fish found in the waters of the restored mangrove ecosystem is based on order, family, genus and species.
Table 2.
The diversity of fish found in the waters of the restored mangrove ecosystem is based on order, family, genus and species.
| Order | Family | Genus | Species | Relative abundance (%) |
| Acanthuriformes | Ephippidae | Ephippus | Ephippus argus | 0.36 |
| Leiognathidae | Eublekeeria | Eublekeeria jonesi | 3.26 | |
| Secutor | Secutor interruptus | 1.21 | ||
| Scatophagidae | Scatophagus | Scatophagus argus | 0.60 | |
| Siganidae | Siganus | Siganus canaliculatus | 0.60 | |
| Anabantiformes | Channidae | Channa | Channa striata | 0.12 |
| Anguilliformes | Muraenesocidae | Muraenesox | Muraenesox cinereus | 0.36 |
| Beloniformes | Hemiramphidae | Hemiramphus | Hemiramphus sp. | 1.93 |
| Hyporhamphus | Hyporhamphus quoyi | 0.12 | ||
| Carangaria | Sphyraenidae | Sphyraena | Sphyraena barracuda | 0.36 |
| Toxotidae | Toxotes | Toxotes jaculatrix | 0.97 | |
| Carangiformes | Carangidae | Scomberoides | Scomberoides sp. | 0.24 |
| Atule | Atule mate | 0.24 | ||
| Selaroides | Selaroides leptolepis | 0.24 | ||
| Xiphiidae | Xiphias | Xiphias sp. | 0.24 | |
| Clupeiformes | Clupeidae | Herklotsichthys | Herklotsichthys dispilonotus | 0.60 |
| Anodontostoma | Anodontostoma chacunda | 0.48 | ||
| Dussumieriidae | Dussumieria | Dussumieria acuta | 1.33 | |
| Dussumieria elopsoides | 3.62 | |||
| Pristigasteridae | Ilisha | Ilisha striatula | 2.54 | |
| Engraulidae | Setipinna | Setipinna tenuifilis | 1.33 | |
| Stolephorus | Stolephorus indicus | 25.36 | ||
| Stolephorus | Stolephorus waitei | 3.62 | ||
| Thryssa | Thryssa hamiltonii | 8.45 | ||
| Eupercaria | Sciaenidae | Johnius | Johnius tracheyephalus | 0.12 |
| Gerreidae | Gerres | Gerres filamentosus | 0.36 | |
| Lutjanidae | Lutjanus | Lutjanus sp. | 0.12 | |
| Lutjanus ehrenbergii | 0.72 | |||
| Gobiiformes | Gobiidae | Periophthalmodon | Periophthalmodon schlosseri | 0.12 |
| Gonorynchiformes | Chanidae | Channos | Channos channos | 0.12 |
| Mugiliformes | Mugilidae | Crenimugil | Crenimugil buchanani | 8.82 |
| Crenimugil | Crenimugil seheli | 4.11 | ||
| Ellochelon | Ellochelon vaigiensis | 10.14 | ||
| Mugil | Mugil cephalus | 1.93 | ||
| Planiliza | Planiliza planiceps | 2.66 | ||
| Planiliza subviridis | 0.12 | |||
| Mulliformes | Mullidae | Upeneus | Upeneus sulphureus | 0.36 |
| Ovalentaria | Ambassidae | Ambassis | Ambassis vachellii | 1.09 |
| Ambassis gymnocephalus | 1.93 | |||
| Ambassis nalua | 6.64 | |||
| Pleuronectiformes | Cynoglossidae | Cynoglossus | Cynoglossus lingua | 0.12 |
| Scombriformes | Scombridae | Rastrelliger | Rastrelliger kanagurta | 1.81 |
| Synbranchiformes | Synbranchidae | Monopterus | Monopterus albus | 0.24 |
| Tetraodontiformes | Tetraodontidae | Lagochepalus | Lagochepalus inermis | 0.24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



