
Submitted:
02 January 2026
Posted:
05 January 2026
You are already at the latest version
Abstract
Spinal cord regeneration requires a transformative strategy capable of rewriting inhibitory genetic programs while orchestrating real-time electrical communication with regenerating neural tissues. Recent advancements in precision CRISPR genome editing effectively silence or activate crucial molecular gatekeepers such as PTEN, SOCS3, and various epigenetic repressors, thereby reactivating dormant intrinsic regenerative pathways and enabling robust axonal growth. Concurrently, cutting-edge bioelectronic technologies utilizing piezoelectric, triboelectric, and magnetoelectric scaffolds have emerged, adeptly harnessing the body's inherent biomechanical energy. These innovative materials convert subtle physiological micromotions into finely tuned electrical stimuli, precisely guiding neuronal regeneration without external power sources, addressing limitations associated with traditional implants such as infection risks and mechanical incompatibility.Integrating these genetic modifications with bioelectric innovations creates a potent synergy. Genome-level reprogramming amplifies neuronal responsiveness to bioelectrical signals, markedly enhancing axonal regeneration. Simultaneously, autonomous electrical stimulation sustains and stabilizes cellular, metabolic, and synaptic improvements induced by genomic interventions, forming a closed-loop, self-sustaining therapeutic platform. This advanced system significantly transcends conventional transient recovery approaches, moving toward durable, personalized outcomes. Such convergence of advanced genetic engineering and intelligent biomaterial design represents a groundbreaking shift in regenerative neurology.Despite promising preclinical outcomes, significant translational challenges remain. Critical hurdles include ensuring precise delivery of CRISPR tools, mitigating off-target genomic effects, enhancing biocompatibility and scaffold stability, and navigating rigorous regulatory pathways. Addressing these challenges necessitates integrating next-generation gene-editing technologies, comprehensive genomic surveillance, advanced biomaterial sciences, and meticulous preclinical evaluations. Future directions in spinal cord injury research encompass multiplex genome editing, AI-driven scaffold optimization via digital twins, and tailored immune-evasive biomaterials. Collectively, this innovative approach has the potential to redefine regenerative medicine's boundaries, offering unprecedented hope for sustained, personalized recovery and dramatically improving quality of life for individuals affected by spinal cord injuries.
Keywords:
CRISPR genome editing
; self-powered bioelectronics
; spinal cord injury
; neural regeneration
; bioelectric scaffolds
1. Introduction
1.1. Global SCI Burden and Unmet Needs
Spinal-cord injury (SCI) is a lifelong condition whose absolute burden continues to increase. The Global Burden of Disease 2019 data show approximately 0.9 million incident cases and 20.6 million people living with SCI in 2019, together accounting for more than six million years lived with disability (YLDs) [1,2,3,4,5,6,7,8,9]. Compared with 1990, the prevalence has increased by approximately 82%, a growth driven not only by increased survival but also by aging populations that now contribute to an expanding share of new injuries [9]. Males and young adults remain the most affected, yet steady increases among older age groups portend added healthcare strain in the coming decades [5,9]. Despite gains in acute management, the 2019 burden analysis indicates that most survivors continue to live with substantial long-term disability, underscoring how current surgical and rehabilitative approaches rarely restore full function [9]. These realities highlight an urgent need for therapies able to overcome both the intrinsic limits of axonal regrowth and the extrinsic barriers that silence disrupted circuits.
1.2. Convergence Rationale: Genetic Rewiring Plus Self-Powered Stimulation
Genome engineering strategies are advancing beyond trophic factor supplementation toward precise circuit rewiring. In a zebrafish model, CRISPR-mediated deletion of dusp2 accelerated early axonal regeneration by increasing JNK phosphorylation [3]. A systematic meta-analysis of 71 preclinical studies confirmed that gene therapy vectors, including viral, nonviral and CRISPR-based vectors, significantly enhance locomotor recovery and axonal density after SCI, although optimal molecular targets remain to be standardized [8].
Concurrently, soft bioelectronic platforms harness applied motions to achieve neuromodulation without external power sources. Compared with conventional pulse generators, hybrid tribo-piezo nanogenerators implanted subcutaneously convert compression into therapeutic current and, via epidural electrodes, elicit coordinated hind-limb locomotion at substantially reduced stimulation amplitudes [1]. Electrospun triboelectric membranes provide conformable stimulators that harvest muscle or skin deformation for on-demand nerve activation [4], whereas piezoelectric polymer conduits guide Schwann-cell alignment and neurite extension under cyclic strain [6]. Reviews of triboelectric nanogenerator (TENG) implants further document devices that scavenge respiration, heartbeat and visceral pulsations to power chronic stimulation or sensing modules [7]. By eliminating bulky batteries and percutaneous leads, these energy-autonomous implants mitigate infection risk and improve long-term biocompatibility [7].
Bringing CRISPR editing and self-powered neuromodulation together offers a complementary dual-modality strategy: gene circuits can lift molecular brakes on regeneration, whereas locally harvested bioelectric fields retrain neuronal networks through activity-dependent plasticity. Although the two approaches have yet to be combined within a single experimental system, their mechanistic orthogonality suggests a powerful synergy that warrants systematic exploration in future studies.
This review interrogates the emerging intersection of CRISPR precision engineering and self-powered bioelectronics as a transformative paradigm for SCI care. First, we contextualize the clinical need by analyzing contemporary epidemiology and burden metrics. Next, we dissect advances in next-generation editors, base, prime and multiplex CRISPR systems, focusing on how they neutralize axonal growth inhibitors, modulate immune checkpoints and enable lineage-specific reprogramming. We then examine triboelectric, piezoelectric and magnetoelectric generators that harvest in vivo biomechanical energy to achieve closed-loop neuromodulation, emphasizing design principles that optimize charge output, mechanical compliance and biological integration. Finally, we outline convergent pathways where genetic rewiring and autonomous stimulation may act synergistically; map translational hurdles in immunogenicity, device longevity and regulatory classification; and propose metrics for preclinical validation and first-in-human trials. By integrating these technological frontiers, the review aims to chart a coherent roadmap from a molecular blueprint to bedside implementation, ultimately striving for restorative therapies that are durable, patient-tailored and globally accessible.
2. Mechanistic Findings
2.1. CRISPR Editing Targets that Unlock Axon Regrowth
Regeneration in the adult spinal cord is stifled by a double bind: external inhibitors and an intrinsic shutdown of growth genes. Elegant genetic experiments first loosened that internal brake. Deleting the phosphatase PTEN reawakens mTOR signaling, fattens growth cones and sends hundreds of corticospinal axons across millimeter-scale thoracic gaps [10,11,12,13,14,15,16,17,18,19,20,21]. The sprint, however, soon falters. A second edit, removing the cytokine repressor SOCS3, rearms STAT3 transcription and keeps axons advancing far beyond the reach of either single manipulation, at least in retinal ganglion neurons that face comparable CNS roadblocks [16].
The power of CRISPR lies in extending such lessons beyond conventional knockouts. A catalytically dead Cas9 fused to a KRAB domain has been programmed to epigenetically silence PTEN; the repression is transient yet potent and purposely crafted to avoid systemic mTOR hyperactivation, a safety angle still awaiting in vivo confirmation (Figure 1) [20]. Zooming out from single loci, chromatin itself emerges as a master gatekeeper: ATAC-seq mapping reveals that progrowth enhancers become hermetically sealed during maturation, forming an epigenetic wall that transcription factors cannot climb [17].
Here, chromatin modifiers were used to redraw the map. Environmental enrichment and pharmacological potentiators recruit the histone acetyl-transferase CBP to those shuttered genes, reopening their chromatin and triggering parallel sprouting of sensory and motor tracts in injured rodents [18]. These mechanistic strategies are operationalized in acute and chronic in vivo PTEN-KO cohorts with BDA and fluorogold tracers and biweekly behavioral assessments (Figure 2) [19]. In effect, PTEN and SOCS3 increase metabolic and cytokine checkpoints, whereas CBP-mediated acetylation opens the genomic atlas of regeneration. CRISPR now supplies fine-tuned chisels to sculpt each layer of this hierarchical barrier.
2.2. Tribo-, Piezo- and Magnetoelectric Scaffolds: Biophysical Cues for Edited Neurons
Genetically primed axons still crave directional and metabolic guidance. Self-powered bioelectronic scaffolds, materials that harvest ambient energy and translate it into neurotrophic cues, have been developed. One standout design embeds Fe₃O₄@BaTiO₃ nanoparticles in a gelatin-methacrylate matrix (Figure 3). With a simple alternating magnetic field, the hydrogel produces local electric potentials, releases HUMSC exosomes, dampens inflammation and accelerates axon extension in rat contusion models (Figure 4a) [6,10]. Upon scaling down, magnetoelectric microrobots cloaked in human neural progenitors steer through tissue and, after pulsed stimulation, reinstate almost-normal swimming in injured zebrafish larvae by three days [11].
Even “bare” magnetic nanoparticles matter: pico-Newton forces transmitted under remote fields tug on membrane receptors and cytoskeletal strands, subtly biasing neurite trajectories. This phenomenon has been cataloged across multiple neural engineering studies [12].
Mechanical energy offers a complementary route. Piezoelectric materials convert strain into voltages that mimic endogenous bioelectricity (Figure 4b). A sweeping review charts their tunability and highlights conceptual hybrids with conductive hydrogels and triboelectric nanogenerators that can tap muscle or respiratory micromotions for on-demand stimulation [13]. Theory is already progressing into practice: a K₀.₅Na₀.₅NbO₃-loaded decellularized matrix yields >500 mV under ultrasound, spikes intracellular Ca²⁺, activates the CaMK2β/PGC-1α axis, boosts ATP and restores both locomotion and bladder control in vivo [21]. Such platforms turn passive matrices into living electronics capable of sensing, harvesting and delivering energy precisely where regenerating circuits need it.
2.3. Electrical–Genetic Cross-Talk: Amplifying the Repair Program
Electrical stimulation has long nudged neurons toward regrowth; genome editing hands them the molecular tools to respond. Functional electrical stimulation (FES) alone activates cAMP- and STAT3-centered gene networks and improves both peripheral and central axon extension [14]. When PTEN- or PTEN/SOCS3-edited neurons enter the electrical microenvironments generated by magnetoelectric or piezoelectric devices, multilayer dialog occurs.
Metabolic lift. Ultrasound-driven piezoelectric currents in the KNN hydrogel increase cytosolic Ca²⁺, trigger Camk2β, upregulate PGC-1α and promote mitochondrial biogenesis, which serves as fuel for long-range axon advancement [21]. The Ca²⁺ surge may further amplify mTOR signals unchained by PTEN loss, suggesting a beneficial metabolic loop.
Immune and niche modulation. The magnetoelectric hydrogel calms early inflammation and accelerates neuronal differentiation [10], complementing CBP-mediated chromatin opening, which primes growth genes [18]. In parallel, FES-level electrical fields phosphorylate STAT3 [14]; in neurons bereft of SOCS3, those fields could plausibly extend STAT3-driven transcription for days.
Guidance precision. Magnetoelectric microrobots focus on subcellular fields that are hypothesized to bias cytoskeletal polymerization, knitting regenerating axons into target circuits faster than gene editing alone permits [11]. Conceptual tribo/piezo hybrids aim to convert gait or breathing motions into spike trains, rhythmically entraining edited neurons during rehabilitation [13].
Epigenetic locking. CBP’s acetyltransferase activity consumes acetyl-CoA. The ATP surge provoked by piezoelectric stimulation plausibly enlarges the acetyl-CoA pool, cementing the chromatin state in which CBP initiates [18,21].
Together, these intertwined layers, membrane excitability, metabolic charge, immune moderation and epigenetic stability, transform isolated gene edits into a coherent, systems-level regeneration programme. The convergence of CRISPR precision and self-powered bioelectronics thus sketches a roadmap toward durable spinal-cord repair, where neurons not only regrow but also reintegrate into functional circuits guided by a symphony of genetic and biophysical cues.
3. Design & Engineering Principles
3.1. Biomaterial Selection, Porosity and Mechanical Matching
The first requirement for any spinal scaffold is to “speak the mechanical dialect” of neural tissue. Ultrasound-responsive piezoelectric hydrogels formulated with a decellularized spinal cord matrix and K₀.₅Na₀.₅NbO₃ nanoparticles reach an elastic modulus of approximately 1 kPa, a value that nests neatly within the 0.1–3 kPa window measured in healthy rat cords and preserves stem-cell neurogenesis [22,23,24,25,26,27,28,29,30]. However, bulk softness alone cannot stop scarring. Very high interconnected porosities, often above 85 to 90%, minimize foreign-body pressure while keeping the construct as compliant as the native cord [25]. Techniques such as freeze-drying, salt leaching and gas foaming are preferred because their ice or gas templates carve tortuous, channel-like pores that resemble endoneurial fascicles, allowing oxygen, cytokines and axons to move freely [30].
Chemistry provides another axis of control. Natural polymers such as collagen or fibrin carry cell-binding motifs but lose shape under compression; synthetic aliphatic polyesters such as PLGA or PCL resist deformation but lack bioactivity. Hybrid systems, such as collagen-gelatin, collagen-chitosan or PLGA microcapsules within GelMA, allow engineers to degrade from a few weeks to several months while offering biochemical cues and mechanical strength in one package [27]. The alignment matters as well. Electrospun fibers that run parallel to the cord draw nascent axons along the lesion axis and bias infiltrating macrophages toward an anti-inflammatory M2 state, thereby uniting structural guidance with immunomodulation [25]. When millimeter-scale precision is needed, computer-aided 3D printing can sculpt microchannels that preserve more than half of the scaffold volume as open space, creating highways through which thousands of regenerating axons can advance in parallel [25].
In addition to providing passive support, modern scaffolds act as metabolic partners. Cerium oxide nanozyme fibers embedded within a porous matrix quench reactive oxygen species, polarize macrophages toward the M2 phenotype and sustain neuronal ATP production, thereby linking redox control to energy homeostasis [26]. A practical design rule therefore emerges: start with a compliance-matched backbone, sculpt highly connected pores that balance vascular ingrowth with gentle prestrain on regenerating axons, and then layer biochemical or electroactive modules that converse fluently with immune and metabolic pathways. Such multiscale orchestration converts an inert implant into a neuroinstructive platform ready to host CRISPR-guided gene programs.
3.2. Energy Harvesting and Charge Control from Spinal Micromotion
Noninvasive ultrasound pulses repeatedly compress the piezoelectric hydrogel by micrometres, providing a controllable electromechanical energy source. In this ultrasound-driven piezoelectric hydrogel, these deformations are converted into open-circuit voltages peaking at approximately 0.96 Vpp under 1 MHz ultrasound stimulation. The resulting electric fields activate Piezo1 channels, trigger CaMKII and CREB phosphorylation and, when combined with neural-stem-cell delivery, accelerate axonal regeneration in vivo [22]. A complementary strategy embeds biodegradable K₀.₅Na₀.₅NbO₃ crystals in a decellularized matrix. The pico- to nanocoulomb pulses increase ATP synthesis through a Ca²⁺/CamK2β/PGC-1α cascade, effectively converting mechanical “noise” into metabolic “fuel” for injured neurons [29].
Triboelectric nanogenerators extend the toolkit by directly translating shear and compressive forces into biphasic stimulation waveforms. The four canonical operating modes, vertical contact, in-plane sliding, single-electrode and free-standing, offer designers the flexibility to match diverse spinal kinematics without adding batteries or wires [24]. Computational models further show that micropatterning of the triboelectric surface amplifies local electric fields and sharply tunes neuronal firing thresholds simply by varying the contact-release velocity, providing engineers with a clean, chemistry-free knob to adjust charge delivery [28].
Stable routing of the harvested energy is essential. Galvanic body-coupled links in simulation report more than 20% power-transfer efficiency at 1.25 GHz to a 2-mm-deep implant via differential ring electrodes, eliminating bulky receiver coils and delivering a matched load impedance for implanted neurostimulators [23]. When integrated with scaffold-embedded piezo- and tribo-units, this link may allow energy to be buffered in microsupercapacitors and released in principle for low-duty-cycle CRISPR electroporation bursts, maintaining genomic precision while preventing Joule heating.
By weaving piezoelectric, triboelectric and galvanic modalities into soft, compliant scaffolds, current research is transforming spinal micromotion from a mechanical nuisance into an endogenous power grid. The resulting autonomous neuro-electronic hybrids can sense gene edits and stimulate a perfectly choreographed rhythm with the cord itself, suggesting fully self-sustaining therapies for spinal cord repair.
3.3. Integrated Gene-Delivery Modules (AAV, LNP, Electroporation)
Delivering CRISPR reagents to a traumatized spinal cord demands vectors able to cross the blood-spinal barrier, thread a dense extracellular matrix, and evade vigilant immune sentinels. Three complementary platforms, recombinant adeno-associated virus (AAV), lipid nanoparticles (LNPs), and in vivo electroporation of ribonucleoprotein (RNP) complexes, now anchor most experimental strategies, each balancing efficiency, payload capacity, and spatiotemporal precision in distinct ways [31,32,33,34,35,36,37].
3.3.1. AAV Vectors
Recombinant AAVs readily transduce postmitotic neurons and astroglia while provoking only mild inflammation, resulting in prolonged expression from a compact viral genome [36]. Their ≈4.7 kb packaging window accommodates streamlined or split-Cas nucleases plus single- or dual-guide cassettes driven by cell type-specific promoters, enabling one-pot editing constructs for spinal repair [31,36]. High systemic doses, however, can elicit anti-capsid immunity and off-target hepatic transduction [36]. Capsid reengineering mitigates these liabilities: the chimeric serotype AAV-DJ yields markedly higher vector-genome loads in the mouse brain and spinal cord and lower liver uptake than AAV9 after intrathecal dosing, but not after intracerebroventricular dosing [35]. This enhanced tropism permits therapeutic editing at substantially lower vector doses than many early AAV9 studies, narrowing the immunological risk envelope [35]. The remaining hurdles, finite genome capacity and the theoretical risk of insertional mutagenesis, drive ongoing work on split-vector and hybrid delivery schemes [31,36].
3.3.2. LNP Systems
Fully synthetic LNPs furnish a nonviral alternative with little preexisting immunity. State-of-the-art formulations blend ionizable cationic lipids (pKa ≈ 6.4), cholesterol, DSPC, and PEG-lipids to promote endosomal escape, structural integrity, and a tunable circulation half-life [33,37]. Surface functionalization with antibodies, peptides, or small molecules steers particles across central-nervous-system barriers, although robust parenchymal deposition remains challenging [37]. In rodent and nonhuman primates liver, LNP-delivered CRISPR mRNA or RNP routinely yields on-target editing efficiencies exceeding 60% [31,33]. Spinal-cord efficiencies are lower but improve through receptor-targeted chemistry and adjunct physical modalities such as focused ultrasound [37]. Because LNP cargo is cleared within hours to days, nuclease or editor exposure is transient, limiting cumulative off-target activity and dampening innate immune activation [32,33].
3.3.3. In Vivo Electroporation
Direct electroporation of preassembled Cas9–gRNA RNPs provides unmatched spatial and temporal control. Fei and colleagues microinjected RNPs into the axolotl spinal cord lumen and, after millisecond electric pulses, achieved > 90% knockout of multiple loci in neural stem cells without compromising tissue viability [34]. Modern electrode arrays and pulse width algorithms confine gene transfer to millimeter-scale regions, allowing multiplex editing during a single surgical session. Because RNPs carry no coding DNA, nuclease activity occurs in a brief “pulse” that ends as the protein degrades, sharply reducing prolonged Cas exposure and limiting adaptive immune recognition [32]. Challenges include the need for invasive access and the difficulty of scaling to larger vertebrates, although coupling with biodegradable scaffolds is under active investigation [31,34].
No single vector yet satisfies every requirement of spinal cord gene editing. AAVs exhibit durable expression at clinically tolerable doses but remain constrained by limited cargo space and capsid immunogenicity. LNPs enable modular, short-lived editing with a strong safety record but still wrestle with central nervous system penetration. RNP electroporation yields millimeter-scale precision but requires direct surgical access. By combining these strengths, piezoelectric hydrogels that eliminate AAV, self-powered scaffolds stocked with LNP reservoirs, or microelectronic chips that dispense RNP bursts on demand, engineers are forging integrated platforms poised to deliver safe, efficient, and autonomous CRISPR therapies for spinal-cord repair.
4. Preclinical Evidence
4.1. In Vitro Neural and Organoid Data
Spinal-cord organoid technology has progressed from rudimentary neuroepithelial spheres to patterned three-dimensional tissues that mirror the dorsal–ventral architecture of the developing cord. Protocols developed by Zhou et al. guide human pluripotent stem cells through dual-SMAD inhibition, retinoic acid exposure and graded caudalization, yielding caudal neural progenitors that express floor plate markers and generate spontaneous action potentials within four weeks [38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59]. When these spinal organoids are fused with region-matched brain or peripheral-ganglion organoids, they form “ascending-pathway” assembloids that relay tactile signals from peripheral sensory neurons to thalamic targets, establishing an entirely human platform for CRISPR guide-RNA screening of synaptic-rewiring strategies [44].
Electrical cues are emerging as potent maturation drivers. A systematic synthesis of next-generation organoid studies revealed that weak direct-current or low-frequency fields delivered through conductive polymer matrices accelerate neurite extension and synchronize network bursting without the need for exogenous neurotrophic factors; although most primary data originate from cerebral organoids, similar principles are now being applied to spinal constructs [52]. Relative to Matrigel controls, embedding organoids in hydrogels containing decellularized neonatal spinal extracellular matrix accelerates neuronal diversification and supports longer-range axonal projections [57]. Disease-centered models are likewise advancing; a risdiplam analog has corrected SMN2 splicing and restored motor–neuron output in stem-cell-derived spinal organoids, enabling time-resolved dissection of drug–gene interactions [45]. Collectively, these in vitro systems provide tractable, human-relevant testbeds for multiplex CRISPR editing, biomaterial optimization and bioelectronic stimulation before animal experimentation.
4.2. Rodent SCI Models: Functional and Histological Outcomes
In rodents, convergent use of gene-editing vectors, self-powered stimulators and electroactive scaffolds has translated into substantial functional recovery. Stewart et al. injected a retrogradely transported AAV-Cre two millimeters rostral to a severe thoracic crush, excising Pten from corticospinal neurons. Treated mice regained hind-limb weight support in both acute and three-month chronic cohorts; the benefit waned by six months in parallel with declining reporter expression, highlighting the need for durable promoters or episomal stabilization [38].
Daily neuromodulation markedly augments these molecular interventions. A flexible polyimide thin-film electrode placed subdurally over the lesion delivered charge-balanced 2 Hz biphasic pulses for one hour per day. Stimulation elevated BBB locomotor scores from week 4 through week 12, improved tactile-withdrawal thresholds and preserved white matter volume at the lesion margins without provoking microglial activation [43]. Other teams have closed the biofeedback loop by harvesting biomechanical energy in situ: a bionic Z-structured triboelectric nanogenerator mounted to hind-limb joints converts limb motion into synchronous sensory–motor stimulation, restoring ladder-rung accuracy and increasing serotonin-positive axon density in transected rats [48]. Complementary work has shown that triboelectric nanogenerator-powered electroacupuncture at governor vessel points enhances BBB recovery and dampens astrocyte activation within four weeks [59].
Electroactive scaffolds can simultaneously guide axons and provide endogenous currents. A biodegradable poly-L-lactic acid scaffold doped with potassium-sodium niobate (KNN) crystals and coated with polydopamine generated microcurrents under physiological strain, aligned regenerating axons, reduced cavitation by one-third and increased BBB scores relative to those of inert PLLA bridges [50]. Ultrasound-responsive hydrogels embedded with barium-titanate nanoparticles offer on-demand stimulation: upon insonation, they synchronized cotransplanted Schwann and neural-stem cells, accelerated remyelination and improved gait kinematics in mice [58]. Magnetic alignment can further refine architecture; an injectable β-sheet-forming peptide hydrogel exposed to an external 100 G field produced twofold longer neurofilament-positive projections across contusion gaps and reduced glial scar density by 35% [56].
Overall, rodent data demonstrate that (i) transient yet potent gene-editing pulses unlock intrinsic growth programmes, (ii) low-frequency electric fields retune spared circuits, and (iii) piezo- and triboelectric biomaterials supply both physical conduits and regenerative microcurrents. Together, these elements fulfill the dual imperatives of structural repair and electrophysiological reactivation.
4.3. Early Large-Animal Insights and Translational Gaps
Scaling these innovations to human-sized cords introduces biomechanical, immunological and surgical challenges that small animals cannot recapitulate. In a gyrencephalic mini-pig model, intravenous autoinfusion of autologous leucoconcentrate engineered to express VEGF165, GDNF and NCAM1 preserved gray matter area, reduced cleaved caspase-3 immunoreactivity and increased βIII-tubulin-positive axonal density compared with those of intact leucoconcentrate controls, demonstrating the feasibility of systemic cell-based gene delivery [39]. To streamline vector optimization, a newly created swine reporter line expresses luciferase upon successful gene transfer, enabling nonterminal, longitudinal bioluminescence imaging of vector tropism and dose–response kinetics [49].
Large-animal biomechanics reveal device-specific pitfalls. In pigs subjected to contusion–compression injury followed by dorsal myelotomy, the implantation of fibrin–carbon–microfiber bundles guided aligned axon sprouts but enlarged cystic cavities and failed to curb fibrotic encapsulation, underscoring the need for antiscar adjuncts before clinical deployment [54]. A separate thoracic reconstruction paradigm using sural nerve autografts combined with polyethylene glycol fusogen allowed partial weight bearing in three of eight swine over 12 weeks, yet MRI revealed heterogeneous bridge continuity, highlighting surgical reproducibility as a translational barrier [55].
Porcine models, with a vascular caliber, white matter proportion and innate immunity resembling those of humans, are widely regarded as the most stringent gateway for biomaterial and electroceutical evaluation [51]. Nevertheless, standardized injury severities, harmonized behavioral batteries and validated electrophysiology end points remain scarce, and the cost of good laboratory-practice facilities limits cohort sizes. Addressing these gaps through multicenter consortia will be critical for advancing CRISPR-powered, self-energizing implants toward first-in-human testing.
5. Safety & Immunogenicity
5.1. CRISPR Off-Target Mitigation and Genome Surveillance
Restoring neural circuitry in the traumatized spinal cord leaves no room for genotoxic collateral damage. Every nuclease molecule delivered with a self-powered bioelectronic scaffold must act with surgical precision, and every unintended scar must be found before first-in-human trials. Over the past decade, the field has built an increasingly sophisticated safety net that combines high-fidelity enzyme engineering with increasingly sensitive genome-wide surveillance assays, creating a virtuous cycle of mitigation and measurement [60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76].
5.1.1. From Tag Capture to Single-Molecule CHROMATIN footprints
The first unbiased safety lens was GUIDE-seq, which ligates protected oligonucleotide tags into double-strand breaks (DSBs) inside living cells; systematic use has shown that apparently similar single-guide RNAs (sgRNAs) can differ by two orders of magnitude in their off-target counts and that shortening the sgRNA spacer sharply trims collateral cuts [67]. However, GUIDE-seq requires efficient transfection and struggles in primary neurons. In vitro biochemical approaches removed cellular bottlenecks: CIRCLE-seq circularizes purified genomic DNA and virtually eliminates background reads, identifying off-targets missed by GUIDE-seq and doing so with a reference genome. This is a crucial advantage for diverse patient haplotypes [69]. Its tagmentation-based successor, CHANGE-seq, scales to hundreds of guides in parallel and shows that the chromatin context plus human single-nucleotide variation modulates approximately fifteen percent of all detected off-sites [71].
5.1.2. True Translational Relevance Comes from Capturing Editing Events Inside Intact Tissue.
DISCOVER-Seq repurposes chromatin immunoprecipitation to follow the DNA-repair sensor MRE11 at nucleotide resolution; it pinpoints off-targets during adenoviral editing of the mouse liver and patient-derived iPSCs without any sequence bias [60]. Pharmacological stabilization of MRE11 residence recently increased sensitivity even further: DISCOVER-Seq+ revealed up to fivefold more sites across human T cells and murine hepatocytes by transiently inhibiting DNA-PKcs [74]. Complementing protein-based read-outs, GUIDE-tag tethers a biotinylated donor to Cas9, enriching cleavage junctions in vivo to a detection floor of 0.2% and simultaneously mapping large deletions and translocations, which are particularly problematic for long axons [73].
5.1.3. Learning to Predict Before We Cut.
Large curated datasets are fuelling machine-learning models that flag risk in silico. A 2024 study generated 1.2 million bulged off-target alignments and showed that allowing single-base bulges dramatically reshaped specificity landscapes, urging updated scoring rules for neural targets rich in repetitive motifs [66]. CHANGE-seq data have already seeded gradient-boosting predictors that account for chromatin marks, allowing designers to triage guides for spinal injury genes such as PTEN or SOCS3 long before any wet-lab work [71].
5.1.4. Engineering Nucleases That Refuse to Miscut
Surveillance alone is inadequate without enzymes built for accuracy. Structure-guided mutagenesis produced SpCas9-HF1, in which neutralizing four backbone contacts decreased the number of GUIDE-seq-detectable off-targets to background while preserving on-target editing for more than eighty-five percent of the guides [68]. Parallel directed evolution campaigns yielded Sniper-Cas9, which survives dual positive–negative bacterial selection and maintains wild-type activity yet resists mismatched substrates in human cells [72]. Hyperaccurate HypaCas9 tightened the REC3 allosteric switch and was capable of allele-specific editing in mouse zygotes, an attractive feature for dominant spinal mutations [75]. When combined with truncated or chemically modified sgRNAs, both strategies have been shown to lower off-site cleavage without sacrificing efficiency [63,64]; these variants routinely push indels at the top off-sites below 0.1%.
5.1.5. Next-Generation Editors Remove the DSB Trigger Altogether
Base and prime editors swap single nucleotides or short stretches without cutting both DNA strands, intrinsically lowering the chance of chromosomal rearrangements. CHANGE-seq-BE profiled adenine and cytosine deaminase fusions and provided a roadmap for separating desired activity from RNA or off-target DNA deamination [61]. Prime editing further minimizes risk: whole-genome sequencing of human pluripotent stem cells revealed no increase in spontaneous single-nucleotide variants or indels after prolonged PE2 expression compared with controls [65]. The original prime-editing report also revealed fourfold lower modification at 16 canonical Cas9 off-sites while installing therapeutically relevant alleles such as HBB E6V [62].
5.1.6. Integrating Mitigation with Surveillance for Spinal Applications
In the envisioned self-powered implant, a high-fidelity or DSB-free editor would be paired upstream with a patient-specific in vitro CIRCLE-seq screen, followed by in vivo DISCOVER-Seq+ verification in large-animal spinal models. Candidate guides that survive this gate may then enter toxicology studies with a GUIDE tag to capture rare translocations under chronic electrical stimulation. Computational bulge-aware filters continuously refine the guide choice, as additional human haplotypes are sequenced.
Taken together, these advances demonstrate that precision editing now relies on a three-layer safety stack: first, bulge-aware in silico triage informed by CHANGE-seq and million-site training data; second, patient-haplotype-matched CIRCLE-seq followed by DISCOVER-Seq+ or GUIDE-tag in the relevant spinal tissue; and third, delivery of a DSB-free or high-fidelity editor whose residual off-site scarring sits below the spontaneous mutation rate of spinal neurons. By turning off-targets from an unpredictable hazard into a design variable, this workflow grants developers a quantifiable margin of genetic safety. It also shortens the translational runway because every guide that clears the screen arrives with a documented, lesion-by-lesion risk register. In practice, such diligence is what will allow CRISPR-powered, self-powered neuroelectronic implants to drive axon regrowth on a genome left unscarred.
5.2. Scaffold Biocompatibility and Foreign-Body Response
Successful translation hinges on a scaffold that slips past immune surveillance while remaining mechanically and electrically competent. The very first checkpoint is the glial front: after only four hours of platinum–iridium stimulation, microglia exhibit an amplitude-dependent rise in necrotic death, confirming that even brief electrical exposure can provoke a sharp innate response [77,78,79,80,81,82,83,84,85,86,87,88]. Therefore, any material introduced into the injured cord must dampen, rather than amplify, this early warning.
Electroconductive hydrogels convert that challenge into therapeutic leverage. A wireless piezoelectric gelatin–BaTiO₃ matrix driven by focused ultrasound produces nanoampere currents that open Piezo1 channels, polarize macrophages toward an M2 phenotype and, in rats, diminish cystic cavities and fibrotic scarring when combined with neural and umbilical-cord mesenchymal stem cells [78]. Complementary survey work has shown that hydrogels tuned for high hydration, ionic conductivity and self-repair accurately match extracellular-matrix softness, bridge millimeter-scale cord gaps and sequester pro-oxidant species that otherwise derail regeneration [79].
Conductive formulations push the dialog further. Networks laden with polypyrrole or graphene propagate endogenous action-potential-level fields, increase calcium-dependent neurogenic signaling and extend axons, effects that can be amplified by well-timed external pulses [81]. Chemistry matters as much as conductivity: a macroporous cryogel reinforced with Ti₃C₂ MXene nanosheets lowers the level of intracellular reactive oxygen species in PC-12 neurons, elevates β-III-tubulin expression and triggers robust neuritogenesis without detectable cytotoxicity, combining mechanical resilience with electrical percolation and redox buffering [82].
Energy-harvesting designs close the loop between biomechanics and cell metabolism. A biomimetic piezoelectric hydrogel that converts spinal micromotion into millivolt outputs rewires glycolytic flux, restores mitochondrial membrane potential and sharply reduces lipid-peroxidation markers in a rodent hemisection model, all without the rigid lead–tissue boundary that usually invites fibrotic encapsulation [88].
Collectively, next-generation scaffolds are evolving from passive placeholders to immunoadaptive mechano-electrical transducers. By blending soft hydrated backbones, intrinsic conductivity and active detoxification chemistry, they thin the foreign-body capsule, stabilize the lesion microenvironment and extend the therapeutic window for downstream CRISPR interventions.
5.3. Electrical, Thermal and Long-Term Degradation Limits
Delivering currents large enough to reanimate circuitry yet small enough to avoid collateral injury remains the central engineering puzzle. The foundational Shannon model links safe stimulation to charge density and charge per phase, but it was derived with large planar electrodes and narrow pulse widths that no longer represent modern interfaces [87]. A contemporary universal model now folds in electrode geometry, intercontact spacing and impedance, predicting electrochemical breakdown with submillimeter resolution and showing that microscale contacts can tolerate higher current densities when pulse width and material impedance are cooptimized [80].
These short-term culture data reinforce these predictions. When mixed glial networks are exposed to rectangular pulses of increasing amplitude, both cell death and impedance rise steeply, underscoring the dominant role of charge delivery in setting biological limits [77]. Beyond electrochemistry, Joule heating looms large. A 10 kHz spinal-cord stimulation system increases the phantom-cord temperature by approximately 0.64 °C under clinically typical settings and up to 1.7 °C near compliance ceilings, values that approach thresholds for excitability modulation yet remain below protein denaturation limits [84]. Bioheat modeling further shows that a supralinear temperature increases with the root-mean-square current until the device voltage caps impose an effective thermal fuse.
Materials must survive these stresses for years. The diamond-in-polyimide insulation wrapped around the PEDOT:PSS-coated TiN microelectrodes preserves the impedance and charge-storage capacity after accelerated saline aging and in vivo auditory-cortex use, pairing the corrosion resistance of the diamond with the flexural compliance of the polyimide [85]. A broader review of flexible deep-brain interfaces highlights two longevity strategies: passive invisibility through modulus matching and active drug-eluting layers that dispense dexamethasone or reactive-oxygen-species scavengers on demand, together extending signal-to-noise lifetimes into the multiyear range [86].
Safe-charge ceilings remain anchored in classical damage threshold data. Detailed cortical analyses have shown that surpassing material-specific charge-injection capacities causes irreversible histological damage, guiding parameter selection for both acute and chronic devices [83]. The rise of self-polarizing piezoelectric hydrogels finally invites a fresh look at dose metrics: because these materials convert intrinsic motion into charge, long-term safety may hinge as much on metabolically coupled work units as on conventional charge density [88].
Taken together, modern spinal implants navigate a multidimensional safety landscape. Electrochemical ladders are charted by predictive equations, thermal boundaries are set by pulse bandwidth and hardware compliance, and degradation curves are flattened by inert, flexible or self-healing substrates. Balancing these axes enables stimulation regimens potent enough to synergize with gene editing yet disciplined enough to preserve the tissue they aim to heal.
6. Regulatory & Ethical Pathways
6.1. FDA, EMA and NMPA Combination–Product Frameworks
When a self-powered bioelectronic scaffold is coupled with a CRISPR editing cassette, it transforms into a true combination product that blends the risk profiles of a biologic, a gene therapy and an active implantable medical device. The United States Food and Drug Administration addresses these hybrids through the Office of Combination Products, whose continuously updated guidance portfolio spells out how sponsors determine lead-center assignment, satisfy user-fee classes, incorporate human factor testing and apply unique device identifiers. By consulting this “living library” at the concept stage, developers can draft a single regulatory roadmap that anticipates which modules will be reviewed by CDER, CBER or CDRH and which cross-disciplinary questions will arise during presubmission meetings [89,90,91,92,93,94,95,96].
Across the Atlantic, the European Medicines Agency offers a parallel pathway in the 2025 Guideline for Investigational Advanced Therapy Medicinal Products. Gene or cell therapies that include an energy-harvesting scaffold qualify as combined ATMPs, instantly triggering extraquality sections for component interplay, more stringent nonclinical packages and a mandatory long-term follow-up plan [93]. Section 7 of that document expressly requires sponsors to integrate relevant ISO and IEC device standards into the clinical dossier, ensuring that electrical safety data can be evaluated alongside vector potency assays.
Global convergence is reinforced by the effects of ICH S12 on nonclinical biodistribution. Coauthored by regulators from the FDA, EMA and China’s NMPA, S12 standardizes the study design, sampling matrices and analytic sensitivity, thereby enabling one biodistribution dataset to support Investigational New Drug, Clinical Trial Application or Investigational Device Exemption submissions in multiple jurisdictions [89]. While dossier formats still differ, the three agencies share core expectations: full traceability of every component, a transparent rationale for how the device and gene therapy act together, and a lifecycle safety plan that stretches from first-in-human dosing to postmarket corrective actions.
6.2. Preclinical Package: Biodistribution and IEC/ISO Electrical Testing
Before a CRISPR-enabled scaffold may enter the clinic, regulators insist on two complementary evidence streams: biodistribution of the editing complex and bench verification that the self-powered implant behaves safely under physiological and magnetic resonance conditions. ICH S12 frames biodistribution as a staged endeavour. A single, carefully designed study in a pharmacologically relevant species can capture organ tropism, persistence and potential germ-line exposure, provided that highly sensitive assays such as quantitative PCR or in situ hybridization push detection limits as low as technically feasible [89]. The guidelines allow sponsors to minimize animal numbers through judicious sampling schedules, as long as peak and clearance phases remain well characterized.
Electrical and mechanical safety are benchmarked against ISO 14708-3, the foundational standard for implantable neurostimulators. The causes of leakage current, charge density, electromagnetic compatibility and accelerated aging define the worst-case test conditions, whereas Annex DA sets a strict 2 °C ceiling for MRI-induced temperature rise [94]. Meeting these thresholds often dictates material choices. For example, Doguet and colleagues replaced conventional copper leads with optical fibers and high-impedance traces in an optoelectronic vagus nerve stimulator, achieving full MRI compatibility at 1.5 T and 3 T without sacrificing biphasic pulse fidelity [96]. These results illustrate how rigorous device engineering can preempt regulatory concerns and enable seamless integration with gene-delivery payloads.
Because thermal injury or stray currents can alter tissue permeability and, by extension, vector shedding, regulators look for integrated risk analyses that combine GLP pathology, ISO bench data and real-time telemetry from prototype implants. The risk-based approach of EMAs even permits well-validated bench or in silico models to be used as substitutes for animal studies when safety margins are demonstrated convincingly [93]. A cohesive preclinical package therefore tells one coherent story: the gene editor remains confined to intended tissues, the scaffold performs within electrical and mechanical limits, and neither technology amplifies the other’s hazards.
6.3. Clinical Trial Design and Postmarket Duties
Once preclinical gates are cleared, first-in-human evaluation must balance scientific ambition with patient safety. The FDA’s 2015 guidance for early-phase cellular and gene therapy products recommends staggered dosing, sentinel cohorts and extended follow-up capable of catching late genotoxic events [90]. These principles align naturally with adaptive, seamless phase 1/2 study designs that have begun to dominate next-generation spinal-cord-injury trials. Scheuren and Kramer described protocols in which interim analyses resize dose-expansion cohorts and recycle participants, accelerating the evaluation of neuromodulatory interventions while maintaining robust safety oversight [95]. The same framework can be applied to CRISPR–bioelectronic studies, provided that data and safety monitoring boards receive live telemetry on pulse fidelity alongside molecular readouts of editing efficacy.
Real-world evidence is now a recognized regulatory currency. The FDA defines curated electronic health records, claims databases and disease registries as valid sources that can supplement effectiveness or even replace certain postapproval trials when data quality is high [92]. Embedding a condition-specific registry directly into the pivotal study makes the transition to postmarket surveillance almost frictionless. Device serial numbers, vector lot IDs and patient-reported outcomes flow into one longitudinal dataset that can flag emerging signals far earlier than periodic safety update reports alone. EMAs echo this expectation by requiring detailed long-term follow-up plans that integrate manufacturing provenances, immunogenicity profiles and functional outcomes over the years [93].
Lifecycle obligations do not end at marketing authorization. ISO 14708-3 obliges manufacturers to maintain field-action procedures for firmware updates, battery replacements and recalls, with all changes traceable to individual implants [94]. Sponsors therefore commit, in their marketing application, to remote diagnostics that quantify pulse amplitude and duty cycle, data that can be cross-referenced with registry outcomes to detect performance drift. By uniting adaptive trial design, registry-based evidence and ISO-anchored vigilance, developers create a cradle-to-grave safety net that satisfies FDA, EMA and NMPA expectations while offering patients a transparently managed path toward spinal-cord injury recovery through precision genome editing and self-powered neurostimulation.
7. Future Horizons and Open Questions
7.1. Next-Generation Editors and Multiplex Stimulation
Lipid-nanoparticle (LNP) delivery of ribonucleoprotein editors is redefining CRISPR therapeutics. In an optimized formulation built around the ionizable lipids SM102 and DMG-PEG 2000, adenine base and prime editor RNPs achieved more than a 300-fold gain in in vivo editing efficiency over naked protein while eliminating measurable off-target activity [97,98,99,100,101,102,103,104,105,106,107,108,109,110]. Such chemistry-defined precision allows single-dose correction of point mutations that halt axonal regrowth after trauma or drive complex genetic pathologies [100]. Beyond single loci, base editors have rewritten pathogenic CAG and GAA repeats in Huntington disease and Friedreich ataxia models, converting 66–82% of the target bases in patient fibroblasts and durably suppressing somatic expansions in mice [108].
Neuromodulation is evolving just as quickly. Differential-target multiplex spinal-cord stimulation (DTM-SCS) delivers interleaved pulse trains that sustain an 86 percent responder rate and approximately 75 percent mean pain reduction at 12 months in patients with refractory upper-limb neuropathy [98]. Combining such circuit-level control with gene editors that reset intrinsic growth programs could enable closed-loop “edit-and-electrify” strategies that turn a short postinjury window into a chronic phase of active repair.
7.2. AI-Guided Scaffold Digital Twins
The next generation of bioelectronic scaffolds will be fabricated twice: first as a high-fidelity digital twin and then on the printer bed. A proof-of-concept workflow streams sensor data from a 3D bioplotter to a machine-learning model that retunes extrusion pressure and speed in real time, producing poly-ε-caprolactone scaffolds with a 16.8% improvement in strut fidelity and a rougher topography expected to support cell attachment [99]. A musculoskeletal review extends the paradigm, suggesting that patient-specific twins can forecast load paths and long-term degeneration before any implant is produced [104]. Orthopedic-trauma case studies already use finite-element twins to predict stress distribution after revision surgery and to guide implant selection [109]. Coupled with generative AI optimizers, these twins can sift through thousands of virtual designs overnight and forward only the highest-scoring candidates for fabrication and testing, a predictive paradigm mirroring the increasing utility of in silico docking for optimizing bioactive material interfaces [101].
7.3. Immune-Stealth Materials and Patient-Specific Manufacturing
Foreign-body inflammation remains the chief obstacle to long-term bioelectronic integration. Ultrathin zwitterionic polysulfobetaine/polydopamine coatings reduce protein adsorption by 89%, reduce fibroblast adhesion by 86%, limit astrocyte activation to a 70 µm zone, and decrease blood–brain barrier leakage by 34% around neural probes [105]. This aligns with broader findings in precision nanomedicine, where zwitterionic surface modifications routinely minimize nonspecific binding and extend in vivo circulation [102]. In parallel, gelatin-methacryloyl hydrogels combine self-healing mechanics with electrical conductivity, anti-inflammatory cues, on-demand degradability, and photoprintability into patient-specific geometries [110].
Immune evasion can also be programmed biologically. CRISPR-engineered mesenchymal stromal cells and the extracellular vesicles they secrete modulate cytokine profiles, increase antiapoptotic Bcl-2, and act as low-immunogenicity carriers for genome editors [97]. When these cellular stealth tactics are combined with digital-twin-assisted 3D bioplotting [99], each patient can receive a construct whose geometry, mechanical gradient, and surface biochemistry are tuned to minimize host reactivity while maximizing regeneration.
7.4. Collaborative Roadmaps and Funding Landscapes
A recent audit of United States federal portfolios revealed that while the NIH, CDMRP, and VA support a diverse spinal-cord-injury pipeline, critical areas such as bowel, cardiovascular, and mental-health complications remain underfunded. With the impending sunset of the CDMRP Spinal Cord Injury Research Program and projected National Institutes of Health (NIH) budget cuts, total federal investment could decrease by approximately 50 percent by 2026 [103]. The authors propose a public, living funding dashboard that would allow agencies, industry, foundations, and patient groups to align priorities in real time.
This transparency is vital for capital-intensive domains, ribonucleoprotein editors [107], digital-twin manufacturing [99], and immune-stealth biomaterials [105,110], which are moving toward clinical translation. For example, coordinated road-mapping could synchronize multiplex-stimulation trials with gene-editing delivery studies, accelerating combined-modality approvals. Embedding open digital-twin models and standardized immune compatibility metrics into grant calls would promote data sharing and reproducibility. Finally, cross-agency seed funds earmarked for AI-driven scaffold twins and editor-enabled cell therapies would help promising concepts cross the “valley of death” between discovery and bedside impact.
8. Conclusion
The convergence of precision CRISPR genome editing and self-powered bioelectronic scaffolds represents a transformative approach poised to redefine SCI treatment paradigms. By targeting intrinsic molecular barriers such as PTEN and SOCS3 through precise genomic interventions, researchers have effectively reactivated latent regenerative pathways previously considered inaccessible. Concurrently, bioelectronic scaffolds exploiting piezoelectric, triboelectric, and magnetoelectric phenomena capitalize on physiological micromotions, converting them into finely tuned electrical stimuli capable of guiding neuronal regeneration without reliance on external power sources. This strategic integration not only enhances the regenerative potential of neurons but also stabilizes functional gains at the cellular and synaptic levels, achieving robust and enduring therapeutic outcomes.
Emerging evidence underscores the significant functional recovery facilitated by this synergistic approach, demonstrating remarkable advances in preclinical models. The sophisticated orchestration of genetic and electrical cues promotes extensive axonal regeneration, mitigates inflammatory responses, and encourages meaningful circuit reconstruction. Moreover, these bioengineered platforms inherently address traditional device limitations, such as infection risk, immunogenicity, and mechanical incompatibility, by seamlessly merging with native tissue dynamics and delivering therapeutic stimulation autonomously.
However, several translational hurdles persist, notably concerning precise gene-editing delivery mechanisms, off-target genomic effects, biocompatibility, long-term scaffold stability, and rigorous regulatory pathways. Addressing these challenges demands a multifaceted approach, integrating next-generation gene-editing tools with advanced biomaterial science, comprehensive genomic surveillance methods, and meticulous preclinical evaluation frameworks. Regulatory strategies, harmonized globally through guidelines from the FDA, EMA, and NMPA, provide structured pathways toward clinical translation, emphasizing the importance of stringent preclinical validation and patient safety.
The future of spinal cord regeneration research holds immense promise, particularly with innovations such as multiplex gene editing, AI-driven scaffold design via digital twins, and immune-evasive biomaterials tailored through patient-specific manufacturing. Collaborative efforts and synchronized funding landscapes are crucial to overcome the inherent complexities of merging biological editing with bioelectronic systems, ultimately translating this groundbreaking approach into widely accessible clinical therapies.
Collectively, the integration of precision genome editing and intelligent biomaterials represents more than an incremental advancement; it signifies a revolutionary leap forward, offering new hope to millions of individuals worldwide affected by spinal cord injuries. By harnessing the synergy of genetic and electrical modalities, we are not merely restoring function; we are redefining the very boundaries of regenerative medicine, setting the stage for sustained, personalized recovery and significantly enhancing quality of life.
Author Contributions
Abdullah Ayad confirms sole responsibility for the study conception, literature review, analysis, and manuscript preparation.
Funding
Not Applicable.
Ethics approval and consent to participate
Not Applicable.
Consent for publication
Not Applicable.
Availability of data and material
Not Applicable.
Competing interests
Not Applicable.
Declaration Regarding the Use of AI-Assisted Readability Enhancement
I hereby affirm that the utilization of AI-assisted tools in the refinement of the manuscript was strictly limited to enhancing its readability. At no point were AI technologies employed to supplant essential authorial responsibilities, including the generation of scientific, pedagogic, or medical insights; the formulation of scientific conclusions; or the issuance of clinical recommendations. The implementation of AI for readability enhancement was rigorously supervised under the discerning eye of human oversight and control.
Author information
Abdullah Ayad: A distinguished molecular biologist and geneticist at the forefront of research and development in molecular diagnostics and genetics. Currently serving as a staff scientist at the Al Bayan National Laboratory for advanced medical diagnostics, Abdullah’s work spans several critical areas, including molecular biology, genetics, oncology, and biotechnology. His expertise is particularly notable in the fields of next-generation sequencing (NGS) and CRISPR/Cas9 gene editing technologies.Abdullah’s academic journey in dentistry and his subsequent pivot to molecular genetics illustrate his commitment to integrating diverse fields of science to address complex health issues. He is actively involved in pioneering research on genetic diseases, such as cancer and Sandhoff syndrome, utilizing cutting-edge technologies to explore novel therapeutic strategies and diagnostics. With a strong focus on collaboration and knowledge dissemination, Abdullah has established significant partnerships both locally and internationally, engaging in collaborative projects and conferences that highlight his commitment to advancing the field of molecular genetics. His work with the Ministry of Health in Iraq and his efforts to introduce new technologies such as the Oxford Nanopore Technologies’ MinION devices are testament to his role as a key player in expanding the capabilities of genetic research and diagnostics in the region As an academic and researcher, Abdullah has contributed to numerous scholarly articles and has been an active participant in peer reviews, further enriching the scientific community. His vision for the future includes leading a diagnostics and R&D center that not only focuses on research but also serves as a hub for innovation in genetic diagnostics and therapy in Iraq.
References
- Li, C., Shan, Y., Zheng, S., Tan, P., Bai, Y., Wang, E., ... & Feng, H. (2025). Nanogenerator Neuromodulation to Enable Locomotion Rehabilitation for Spinal Cord Injury via Epidural Electrical Stimulation. Advanced Science, 2501425.
- Ding, W., Hu, S., Wang, P., Kang, H., Peng, R., Dong, Y., & Li, F. (2022). Spinal cord injury: the global incidence, prevalence, and disability from the global burden of disease study 2019. Spine, 47(21), 1532-1540.
- Shao, G. J., Wang, X. L., Wei, M. L., Ren, D. L., & Hu, B. (2023). DUSP2 deletion with CRISPR/Cas9 promotes Mauthner cell axonal regeneration at the early stage of zebrafish. Neural Regeneration Research, 18(3), 577-581.
- Xu, C., Zeng, F., Wu, D., Wang, P., Yin, X., & Jia, B. (2022). Nerve stimulation by triboelectric nanogenerator based on nanofibrous membrane for spinal cord injury. Frontiers in Chemistry, 10, 941065.
- Liu, Y., Yang, X., He, Z., Li, J., Li, Y., Wu, Y., ... & Xiang, H. (2023). Spinal cord injury: global burden from 1990 to 2019 and projections up to 2030 using Bayesian age-period-cohort analysis. Frontiers in neurology, 14, 1304153.
- Shlapakova, L. E., Surmeneva, M. A., Kholkin, A. L., & Surmenev, R. A. (2024). Revealing an important role of piezoelectric polymers in nervous-tissue regeneration: a review. Materials Today, Bio, 25, 100950.
- Wang, C., Shi, Q., & Lee, C. (2022). Advanced implantable biomedical devices enabled by triboelectric nanogenerators. Nanomaterials, 12(8), 1366.
- Cunningham, C. J., Viskontas, M., Janowicz, K., Sani, Y., Håkansson, M. E., Heidari, A., ... & Bo, X. (2023). The potential of gene therapies for spinal cord injury repair: a systematic review and meta-analysis of preclinical studies. Neural Regeneration Research, 18(2), 299-305.
- Safdarian, M., Trinka, E., Rahimi-Movaghar, V., Thomschewski, A., Aali, A., Abady, G. G., ... & Shetty, P. H. (2023). Global, regional, and national burden of spinal cord injury, 1990–2019: a systematic analysis for the Global Burden of Disease Study 2019. The Lancet Neurology, 22(11), 1026-1047.
- Liu, W., Liu, Q., Li, Z., Zhang, C., Li, Z., Ke, H., ... & Li, J. (2025). Multifunctional magneto-electric and exosome-loaded hydrogel enhances neuronal differentiation and immunoregulation through remote noninvasive electrical stimulation for neurological recovery after spinal cord injury. Bioactive Materials, 48, 510-528.
- Ye, H., Zang, J., Zhu, J., Arx, D. V., Pustovalov, V., Mao, M., ... & Pané, S. (2024). Magnetoelectric Microrobots for Spinal Cord Injury Regeneration. bioRxiv, 2024-08.
- Goya, G. F., & Raffa, V. (2018). Magnetic nanoparticles for neural engineering. Clinical Applications of Magnetic Nanoparticles, 395-410.
- Wang, X., Stefanello, S. T., Shahin, V., & Qian, Y. (2025). From Mechanoelectric Conversion to Tissue Regeneration: Translational Progress in Piezoelectric Materials. Advanced Materials, 2417564.
- Jara, J. S., Agger, S., & Hollis, E. R. (2020). Functional electrical stimulation and the modulation of the axon regeneration program. Frontiers in Cell and Developmental Biology, 8, 736.
- Liu, K., Lu, Y., Lee, J. K., Samara, R., Willenberg, R., Sears-Kraxberger, I., ... & He, Z. (2010). PTEN deletion enhances the regenerative ability of adult corticospinal neurons. Nature neuroscience, 13(9), 1075-1081.
- Sun, F., Park, K. K., Belin, S., Wang, D., Lu, T., Chen, G., ... & He, Z. (2011). Sustained axon regeneration induced by codeletion of PTEN and SOCS3. Nature, 480(7377), 372-375.
- Venkatesh, I., Mehra, V., Wang, Z., Califf, B., & Blackmore, M. G. (2018). D evelopmental C hromatin R estriction of P ro- G rowth G ene N etworks A cts as an E pigenetic B arrier to A xon R egeneration in C ortical N eurons. Developmental Neurobiology, 78(10), 960–977.
- Hutson, T. H., Kathe, C., Palmisano, I., Bartholdi, K., Hervera, A., De Virgiliis, F., ... & Di Giovanni, S. (2019). Cbp-dependent histone acetylation mediates axon regeneration induced by environmental enrichment in rodent spinal cord injury models. Science translational medicine, 11(487), eaaw2064.
- Stewart, A. N., Kumari, R., Bailey, W. M., Glaser, E. P., Bosse-Joseph, C. C., Park, K. A., ... & Gensel, J. C. (2023). PTEN knockout using retrogradely transported AAVs transiently restores locomotor abilities in both acute and chronic spinal cord injury. Experimental neurology, 368, 114502.
- Moses, C., Hodgetts, S. I., Nugent, F., Ben-Ary, G., Park, K. K., Blancafort, P., & Harvey, A. R. (2020). Transcriptional repression of PTEN in neural cells using CRISPR/dCas9 epigenetic editing. Scientific reports, 10(1), 11393.
- Shi, G., Su, T., Li, J., Wang, A., Gao, G., Tao, B., ... & Zhao, Y. (2025). Biomimetic piezoelectric hydrogel system for energy metabolism reprogramming in spinal cord injury repair. Theranostics, 15(11), 4955.
- Zhong, H., Zhou, M., Guo, J., Chen, D., Xing, C., Liu, S., ... & Ning, G. (2025). Ultrasound-Driven Wireless Piezoelectric Hydrogel Synergizes with Cotransplantation of NSCs–hUCMSCs for Structural and Functional Recovery in Spinal Cord Injury. Materials Today, Bio, 101805.
- Omi, A. I., Farina, E., Jiang, A., Khalifa, A., Srinivasan, S., & Chatterjee, B. (2024). Galvanic Body-Coupled Powering for Wireless Implanted Neurostimulators. arXiv preprint arXiv:2412.03488.
- Xu, S., Manshaii, F., Xiao, X., Yin, J., & Chen, J. (2024). Triboelectric nanogenerators for self-powered neurostimulation. Nano Research, 17(10), 8926-8941.
- Kaplan, B., & Levenberg, S. (2022). The role of biomaterials in peripheral nerve and spinal cord injury: a review. International journal of molecular sciences, 23(3), 1244.
- Zheng, G., Yu, W., Xu, Z., Yang, C., Wang, Y., Yue, Z., ... & Chen, H. (2024). Neuroimmune modulating and energy supporting nanozyme-mimic scaffold synergistically promotes axon regeneration after spinal cord injury. Journal of Nanobiotechnology, 22(1), 399.
- Chen, K., Yu, W., Zheng, G., Xu, Z., Yang, C., Wang, Y., ... & Chen, H. (2024). Biomaterial-based regenerative therapeutic strategies for spinal cord injury. NPG Asia Materials, 16(1), 5.
- Nazari-Vanani, R., Mohammadpour, R., Asadian, E., Rafii-Tabar, H., & Sasanpour, P. (2022). A computational modeling study of excitation of neuronal cells with triboelectric nanogenerators. Scientific Reports, 12(1), 13411.
- Shi, G., Su, T., Li, J., Wang, A., Gao, G., Tao, B., ... & Zhao, Y. (2025). Biomimetic piezoelectric hydrogel system for energy metabolism reprogramming in spinal cord injury repair. Theranostics, 15(11), 4955.
- Kang, N. U., Lee, S. J., & Gwak, S. J. (2022). Fabrication techniques of nerve guidance conduits for nerve regeneration. Yonsei medical journal, 63(2), 114.
- Taha, E. A., Lee, J., & Hotta, A. (2022). Delivery of CRISPR‒Cas tools for in vivo genome editing therapy: Trends and challenges. Journal of Controlled Release, 342, 345-361.
- Yip, B. H. (2020). Recent advances in CRISPR/Cas9 delivery strategies. Biomolecules, 10(6), 839.
- Kazemian, P., Yu, S. Y., Thomson, S. B., Birkenshaw, A., Leavitt, B. R., & Ross, C. J. (2022). Lipid-nanoparticle-based delivery of CRISPR/Cas9 genome-editing components. Molecular pharmaceutics, 19(6), 1669-1686.
- Fei, J. F., Knapp, D., Schuez, M., Murawala, P., Zou, Y., Pal Singh, S., ... & Tanaka, E. M. (2016). Tissue-and time-directed electroporation of CAS9 protein–gRNA complexes in vivo yields efficient multigene knockout for studying gene function in regeneration. NPJ Regenerative medicine, 1(1), 1-9.
- Chauhan, M., Daugherty, A. L., Khadir, F., Duzenli, O. F., Hoffman, A., Tinklenberg, J. A., ... & Pacak, C. A. (2024). AAV-DJ is superior to AAV9 for targeting brain and spinal cord, and detargeting liver across multiple delivery routes in mice. Journal of Translational Medicine, 22(1), 824.
- Wang, J. H., Gessler, D. J., Zhan, W., Gallagher, T. L., & Gao, G. (2024). Adeno-associated virus as a delivery vector for gene therapy of human diseases. Signal transduction and targeted therapy, 9(1), 78.
- Mendes, B. B., Conniot, J., Avital, A., Yao, D., Jiang, X., Zhou, X., ... & Conde, J. (2022). Nanodelivery of nucleic acids. Nature reviews Methods primers, 2(1), 24.
- Stewart, A. N., Kumari, R., Bailey, W. M., Glaser, E. P., Bosse-Joseph, C. C., Park, K. A., ... & Gensel, J. C. (2023). PTEN knockout using retrogradely transported AAVs transiently restores locomotor abilities in both acute and chronic spinal cord injury. Experimental neurology, 368, 114502.
- Davleeva, M. A., Garifulin, R. R., Bashirov, F. V., Izmailov, A. A., Nurullin, L. F., Salafutdinov, I. I., ... & Islamov, R. R. (2023). Molecular and cellular changes in the posttraumatic spinal cord remodeling after autoinfusion of a genetically enriched leucoconcentrate in a mini-pig model. Neural Regeneration Research, 18(7), 1505-1511.
- Zhou, G., Pang, S., Li, Y., & Gao, J. (2024). Progress in the generation of spinal cord organoids over the past decade and future perspectives. Neural Regeneration Research, 19(5), 1013-1019.
- Maisumu, G., Willerth, S., Nestor, M., Waldau, B., Schülke, S., Nardi, F. V., ... & Yakoub, A. M. (2025). Brain organoids: building higher-order complexity and neural circuitry models. Trends in Biotechnology.
- Saraswathy, V. M., Zhou, L., & Mokalled, M. H. (2024). Single-cell analysis of innate spinal cord regeneration identifies intersecting modes of neuronal repair. Nature communications, 15(1), 6808.
- Harland, B., Matter, L., Lopez, S., Fackelmeier, B., Hazelgrove, B., Meissner, S., ... & Svirskis, D. (2025). Daily electric field treatment improves functional outcomes after thoracic contusion spinal cord injury in rats. Nature Communications, 16(1), 1-16.
- Kim, J. I., Imaizumi, K., Jurjuț, O., Kelley, K. W., Wang, D., Thete, M. V., ... & Pașca, S. P. (2025). Human assembloid model of the ascending neural sensory pathway. Nature, 1-11.
- Corti, S., D’Angelo, A., Beatrice, F., Cordiglieri, C., Miotto, M., Lodato, S., ... & Ottoboni, L. (2025, April). Stem Cell-Derived Spinal Cord Organoid Model: Temporal and Cell-Specific Analysis of Molecular Effects of a Risdiplam Analog (N2. 002). In Neurology (Vol. 104, No. 7_Supplement_1, p. 256). Hagerstown, MD: Lippincott Williams & Wilkins.
- Hsu, M. N., Liao, H. T., Truong, V. A., Huang, K. L., Yu, F. J., Chen, H. H., ... & Hu, Y. C. (2019). CRISPR-based activation of endogenous neurotrophic genes in adipose stem cell sheets to stimulate peripheral nerve regeneration. Theranostics, 9(21), 6099.
- Shi, G., Su, T., Li, J., Wang, A., Gao, G., Tao, B., ... & Zhao, Y. (2025). Biomimetic piezoelectric hydrogel system for energy metabolism reprogramming in spinal cord injury repair. Theranostics, 15(11), 4955.
- Lu, B., Jia, S., Wang, Z., Wu, W., Yan, L., Zhu, L., & Hao, D. (2023). Sensory-motor coupling electrical stimulation driven by a bionic Z-structured triboelectric nanogenerator improves functional recovery from spinal cord injury. Nano Energy, 107, 108133.
- Campbell, J., Korpela, D., Han, H., Zhao, S., Webster, D., Nguyen, Y. A., ... & Carlson, D. (2025). Swine reporter model for preclinical evaluation and characterization of gene delivery vectors. bioRxiv, 2025-06.
- Zhang, J., Wang, Q., Tang, X., Chai, M., Liu, N., Jiang, Z., ... & Chen, P. (2024). A biodegradable piezoelectric scaffold promotes spinal cord injury nerve regeneration. Nano Energy, 132, 110382.
- Wathen, C. A., Ghenbot, Y. G., Ozturk, A. K., Cullen, D. K., O’Donnell, J. C., & Petrov, D. (2023). Porcine Models of Spinal Cord Injury. Biomedicines, 11(8), 2202.
- O’Hara-Wright, M., Mobini, S., & Gonzalez-Cordero, A. (2022). Bioelectric potential in next-generation organoids: electrical stimulation to enhance 3D structures of the central nervous system. Frontiers in Cell and Developmental Biology, 10, 901652.
- Islamov, R., Bashirov, F., Fadeev, F., Shevchenko, R., Izmailov, A., Markosyan, V., ... & Lavrov, I. (2020). Epidural stimulation combined with triple gene therapy for spinal cord injury treatment. International Journal of Molecular Sciences, 21(23), 8896.
- Alves-Sampaio, A., Del-Cerro, P., & Collazos-Castro, J. E. (2023). Composite fibrin/carbon microfiber implants for bridging spinal cord injury: a translational approach in pigs. International Journal of Molecular Sciences, 24(13), 11102.
- Nourbakhsh, A., Takawira, C., Barras, E., Hampton, C., Carossino, M., Nguyen, K., ... & Lopez, M. J. (2024). A novel reconstruction model for thoracic spinal cord injury in swine. PLoS One, 19(9), e0308637.
- Tran, K. A., Jin, Y., Bouyer, J., DeOre, B. J., Suprewicz, Ł., Figel, A., Walens, H., Fischer, I., & Galie, P. A. (2022). Magnetic alignment of injectable hydrogel scaffolds for spinal cord injury repair. Biomaterials Science, 10(9), 2237–2247.
- Sun, Z., Chen, Z., Yin, M., Wu, X., Guo, B., Cheng, X., ... & Zhao, Y. (2024). Harnessing developmental dynamics of spinal cord extracellular matrix improves regenerative potential of spinal cord organoids. Cell Stem Cell, 31(5), 772-787.
- Wang, H., Zhang, W., Ren, Y., Lu, J., Liu, S., Liu, L., ... & Chen, L. (2025). Ultrasound-driven piezoelectric hydrogel enhances Schwann/neural stem cells Cotransplantation for spinal cord injury repair. Materials & Design, 253, 113842.
- Wei, X., Wang, Y., Tan, B., Zhang, E., Wang, B., Su, H., ... & Wu, Z. (2022). Triboelectric nanogenerators stimulated electroacupuncture (EA) treatment for promoting the functional recovery after spinal cord injury. Materials Today, 60, 41-51.
- Wienert, B., Wyman, S. K., Richardson, C. D., Yeh, C. D., Akcakaya, P., Porritt, M. J., ... & Corn, J. E. (2019). Unbiased detection of CRISPR off-targets in vivo using DISCOVER-Seq. Science, 364(6437), 286-289.
- Lazzarotto, C. R., Katta, V., Li, Y., Urbina, E., Lee, G., & Tsai, S. Q. (2024). CHANGE-seq-BE enables simultaneously sensitive and unbiased in vitro profiling of base editor genome-wide activity. bioRxiv, 2024-03.
- Anzalone, A. V., Randolph, P. B., Davis, J. R., Sousa, A. A., Koblan, L. W., Levy, J. M., ... & Liu, D. R. (2019). Search-and-replace genome editing without double-strand breaks or donor DNA. Nature, 576(7785), 149-157.
- Guo, C., Ma, X., Gao, F., & Guo, Y. (2023). Off-target effects in CRISPR/Cas9 gene editing. Frontiers in bioengineering and biotechnology, 11, 1143157.
- Lopes, R., & Prasad, M. K. (2024). Beyond the promise: evaluating and mitigating off-target effects in CRISPR gene editing for safer therapeutics. Frontiers in Bioengineering and Biotechnology, 11, 1339189.
- Habib, O., Habib, G., Hwang, G. H., & Bae, S. (2022). Comprehensive analysis of prime editing outcomes in human embryonic stem cells. Nucleic acids research, 50(2), 1187-1197.
- Yaish, O., & Orenstein, Y. (2024). Generating, modeling and evaluating a large-scale set of CRISPR/Cas9 off-target sites with bulges. Nucleic Acids Research, 52(12), 6777-6790.
- Tsai, S. Q., Zheng, Z., Nguyen, N. T., Liebers, M., Topkar, V. V., Thapar, V., ... & Joung, J. K. (2015). GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR‒Cas nucleases. Nature biotechnology, 33(2), 187-197.
- Kleinstiver, B. P., Pattanayak, V., Prew, M. S., Tsai, S. Q., Nguyen, N. T., Zheng, Z., & Joung, J. K. (2016). High-fidelity CRISPR–Cas9 nucleases with no detectable genome-wide off-target effects. Nature, 529(7587), 490-495.
- Tsai, S. Q., Nguyen, N. T., Malagon-Lopez, J., Topkar, V. V., Aryee, M. J., & Joung, J. K. (2017). CIRCLE-seq: a highly sensitive in vitro screen for genome-wide CRISPR–Cas9 nuclease off-targets. Nature methods, 14(6), 607-614.
- Wienert, B., Wyman, S. K., Richardson, C. D., Yeh, C. D., Akcakaya, P., Porritt, M. J., ... & Corn, J. E. (2019). Unbiased detection of CRISPR off-targets in vivo using DISCOVER-Seq. Science, 364(6437), 286-289.
- Lazzarotto, C. R., Malinin, N. L., Li, Y., Zhang, R., Yang, Y., Lee, G., ... & Tsai, S. Q. (2020). CHANGE-seq reveals genetic and epigenetic effects on CRISPR–Cas9 genome-wide activity. Nature biotechnology, 38(11), 1317-1327.
- Lee, J. K., Jeong, E., Lee, J., Jung, M., Shin, E., Kim, Y. H., ... & Kim, J. S. (2018). Directed evolution of CRISPR-Cas9 to increase its specificity. Nature communications, 9(1), 3048.
- Liang, S. Q., Liu, P., Smith, J. L., Mintzer, E., Maitland, S., Dong, X., ... & Xue, W. (2022). Genome-wide detection of CRISPR editing in vivo using GUIDE-tag. Nature communications, 13(1), 437.
- Zou, R. S., Liu, Y., Gaido, O. E. R., Konig, M. F., Mog, B. J., Shen, L. L., ... & Ha, T. (2023). Improving the sensitivity of in vivo CRISPR off-target detection with DISCOVER-Seq+. Nature Methods, 20(5), 706-713.
- Ikeda, A., Fujii, W., Sugiura, K., & Naito, K. (2019). High-fidelity endonuclease variant HypaCas9 facilitates accurate allele-specific gene modification in mouse zygotes. Communications biology, 2(1), 371.
- Tsai, S. Q., Nguyen, N. T., Malagon-Lopez, J., Topkar, V. V., Aryee, M. J., & Joung, J. K. (2017). CIRCLE-seq: a highly sensitive in vitro screen for genome-wide CRISPR–Cas9 nuclease off-targets. Nature methods, 14(6), 607-614.
- Tsui, C. T., Mirkiani, S., Roszko, D. A., Birtle, M. J., DeCorby, A. E., Churchward, M. A., ... & Todd, K. G. (2025). Electrode stimulation parameter modifications elicit differential glial cell responses in vitro over a short 4-hour timecourse. Biomaterials, 123478.
- Zhong, H., Zhou, M., Guo, J., Chen, D., Xing, C., Liu, S., ... & Ning, G. (2025). Ultrasound-Driven Wireless Piezoelectric Hydrogel Synergizes with Cotransplantation of NSCs–hUCMSCs for Structural and Functional Recovery in Spinal Cord Injury. Materials Today, Bio, 101805.
- Han, C., Jiao, J., Gong, C., Li, J., Zhao, M., & Lu, X. (2025). Multidimensional exploration of hydrogels as biological scaffolds for spinal cord regeneration: mechanisms and future perspectives. Frontiers in Bioengineering and Biotechnology, 13, 1576524.
- Vatsyayan, R., & Dayeh, S. A. (2022). A universal model of electrochemical safety limits in vivo for electrophysiological stimulation. Frontiers in neuroscience, 16, 972252.
- Qin, C., Qi, Z., Pan, S., Xia, P., Kong, W., Sun, B., ... & Yang, X. (2023). Advances in conductive hydrogel for spinal cord injury repair and regeneration. International journal of nanomedicine, 7305-7333.
- Zoughaib, M., Avdokushina, S., & Savina, I. N. (2025). MXene-Reinforced Composite Cryogel Scaffold for Neural Tissue Repair. Molecules, 30(3), 479.
- Cogan, S. F., Ludwig, K. A., Welle, C. G., & Takmakov, P. (2016). Tissue damage thresholds during therapeutic electrical stimulation. Journal of neural engineering, 13(2), 021001.
- Zannou, A. L., Khadka, N., FallahRad, M., Truong, D. Q., Kopell, B. H., & Bikson, M. (2021). Tissue temperature increases by a 10 kHz spinal cord stimulation system: phantom and bioheat model. Neuromodulation: Technology at the Neural Interface, 24(8), 1327-1335.
- Hébert, C., Carlier, S., Leão, J. M., Wilfinger, C., Lissorgues, G., Yvert, B., & Rousseau, L. (2025). Diamond-in-Polyimide Insulation of Arrays of PEDOT: PSS-Coated TiN Microelectrodes for Highly Stable Future Flexible Neural Implants. physica status solidi (a), 2400540.
- Lv, S., Xu, Z., Mo, F., Wang, Y., Duan, Y., Liu, Y., ... & Cai, X. (2025). Long-term stability strategies of deep brain flexible neural interface. NPJ Flexible Electronics, 9(1), 1-17.
- Shannon, R. V. (2002). A model of safe levels for electrical stimulation. IEEE Transactions on biomedical engineering, 39(4), 424-426.
- Shi, G., Su, T., Li, J., Wang, A., Gao, G., Tao, B., ... & Zhao, Y. (2025). Biomimetic piezoelectric hydrogel system for energy metabolism reprogramming in spinal cord injury repair. Theranostics, 15(11), 4955.
- U.S. Food and Drug Administration. (2023, May). S12 nonclinical biodistribution considerations for gene therapy products [Guidance for industry]. https://www.fda.gov/media/167605/download.
- U.S. Food and Drug Administration, Center for Biologics Evaluation and Research. (2015, June). Considerations for the design of early-phase clinical trials of cellular and gene therapy products [Guidance for industry]. https://www.fda.gov/media/106369/download.
- U.S. Food and Drug Administration, Office of Combination Products. (2025, June 26). Combination products guidance documents. https://www.fda.gov/combination-products/guidance-regulatory-information/combination-products-guidance-documents.
- U.S. Food and Drug Administration. (2025, June 9). Real-world evidence. https://www.fda.gov/science-research/science-and-research-special-topics/real-world-evidence.
- European Medicines Agency. (2025, January 20). Guideline on quality, nonclinical and clinical requirements for investigational advanced therapy medicinal products in clinical trials (EMA/CAT/22473/2025). https://www.ema.europa.eu/en/documents/scientific-guideline/guideline-quality-non-clinical-clinical-requirements-investigational-advanced-therapy-medicinal-products-clinical-trials_en.pdf.
- International Organization for Standardization. (2017, April). ISO 14708-3:2017. Implants for surgery—Active implantable medical devices—Part 3: Implantable neurostimulators. https://www.iso.org/standard/60539.html.
- Scheuren, P. S., & Kramer, J. L. K. (2024, November). Next-gen spinal cord injury clinical trials: Lessons learned and opportunities for future success. eBioMedicine, 109, Article 105381. [CrossRef]
- Doguet, P., Garnier, J., Nieuwenhuys, A., Godfraind, C., Botquin, Y., Lemaire, A., Justice, J., Nonclercq, A., El Tahry, R., Corbett, B., & Delbeke, J. (2024, May 15). An optoelectronic implantable neurostimulation platform allowing full MRI safety and optical sensing and communication. Scientific Reports, 14, Article 11110. [CrossRef]
- Hazrati, A., Malekpour, K., Soudi, S., & Hashemi, S. M. (2022). CRISPR/Cas9-engineered mesenchymal stromal/stem cells and their extracellular vesicles: A new approach to overcoming cell therapy limitations. Biomedicine & Pharmacotherapy, 156, 113943.
- White, T., Justiz, R., Fishman, M., Schultz, D., Calodney, A., Cordner, H., ... & Vallejo, R. (2024). Differential Target Multiplexed Spinal Cord Stimulation for the Treatment of Chronic Intractable Upper Limb Pain: 12-Month Results From a Prospective, Multicenter Study. Neuromodulation: Technology at the Neural Interface, 27(8), 1441-1448.
- Ahmed, N., Narayanan, L. K., & Shirwaiker, R. (2024). A framework for digital twin integration in biofabrication and a scaffold 3D bioplotting case study. Manufacturing Letters, 41, 1182-1191.
- Alsuraifi, A., Husam, Y., Mohammed, N. A. R., Mahdi, F. H., Mohammed, J., Mohammed, Z., ... & Ayad, A. (2025). State of art: a narrative review on navigating pathogenesis and therapies for genetic oral disorders. Frontiers of Oral and Maxillofacial Medicine, 7.
- Abozaid, D., Ayad, A., & Azab, A. (2025). Enhancing glass ionomer cement with Citrus aurantium L. extract: a combined in vitro and in silico investigation of antimicrobial and mechanical properties. Beni-Suef University Journal of Basic and Applied Sciences, 14(1), 1-13.
- Alsuraifi, A., Mouzan, M. M., Algzaare, A., Aqeel, Z., Al-Essa, U. I., R. Mohammed, N. A., & Ayad, A. (2025). Quantum Dots: Catalysts for a New Era of Precision Medicine and Biomedical Innovation. Journal of Fluorescence, 1-64.
- Gillespie, T., Buxton, A., Kondiles, B. R., Leal-Garcia, M., Pacheco, M. R., Tran, A. V., ... & Dulin, J. N. (2025). Systematic analysis of federal funding for spinal cord injury research. medRxiv, 2025-06.
- Diniz, P., Grimm, B., Garcia, F., Fayad, J., Ley, C., Mouton, C., ... & Seil, R. (2025). Digital twin systems for musculoskeletal applications: A current concepts review. Knee Surgery, Sports Traumatology, Arthroscopy, 33(5), 1892-1910.
- Golabchi, A., Wu, B., Cao, B., Bettinger, C. J., & Cui, X. T. (2019). Zwitterionic polymer/polydopamine coating reduce acute inflammatory tissue responses to neural implants. Biomaterials, 225, 119519.
- Golabchi, A., Wu, B., Cao, B., Bettinger, C. J., & Cui, X. T. (2019). Zwitterionic polymer/polydopamine coating reduce acute inflammatory tissue responses to neural implants. Biomaterials, 225, 119519.
- Hołubowicz, R., Du, S. W., Felgner, J., Smidak, R., Choi, E. H., Palczewska, G., ... & Palczewski, K. (2025). Safer and efficient base editing and prime editing via ribonucleoproteins delivered through optimized lipid-nanoparticle formulations. Nature biomedical engineering, 9(1), 57-78.
- Matuszek, Z., Arbab, M., Kesavan, M., Hsu, A., Roy, J. C., Zhao, J., ... & Liu, D. R. (2025). Base editing of trinucleotide repeats that cause Huntington’s disease and Friedreich’s ataxia reduces somatic repeat expansions in patient cells and in mice. Nature Genetics, 57(6), 1437-1451.
- Andres, A., Roland, M., Wickert, K., Diebels, S., Stöckl, J., Herrmann, S., ... & Braun, B. J. (2025). Advantages of digital twin technology in orthopedic trauma Surgery–Exploring different clinical use cases. Scientific Reports, 15(1), 1-13.
- Rotaru-Zavaleanu, A. D., Bica, M., Dinescu, S., Ruscu, M. A., Vasile, R. C., Zavate, A. C., & Dinescu, V. C. (2025). Bioactive hydrogels for spinal cord injury repair: Emphasis on gelatin and its derivatives. Gels, 11(7), 497.
Figure 1.
Dual strategies for modulating PTEN to promote axon growth. (A) Growth factor activation of Trk receptors drives PI3K-mediated conversion of PIP₂ to PIP₃, triggering AKT/mTOR signaling that activates S6K, relieves 4E-BP1 inhibition and suppresses GSK-3β, thereby fostering axonal extension; PTEN counteracts this cascade by dephosphorylating PIP₃ back to PIP₂. (B) A catalytically inactive Cas9-KRAB fusion guided to the PTEN locus recruits KAP1, which in turn nucleates NuRD (HDAC), SETDB1 (H3K9 methylation) and HP1 complexes to enforce local chromatin compaction and transcriptional silencing.
Figure 1.
Dual strategies for modulating PTEN to promote axon growth. (A) Growth factor activation of Trk receptors drives PI3K-mediated conversion of PIP₂ to PIP₃, triggering AKT/mTOR signaling that activates S6K, relieves 4E-BP1 inhibition and suppresses GSK-3β, thereby fostering axonal extension; PTEN counteracts this cascade by dephosphorylating PIP₃ back to PIP₂. (B) A catalytically inactive Cas9-KRAB fusion guided to the PTEN locus recruits KAP1, which in turn nucleates NuRD (HDAC), SETDB1 (H3K9 methylation) and HP1 complexes to enforce local chromatin compaction and transcriptional silencing.
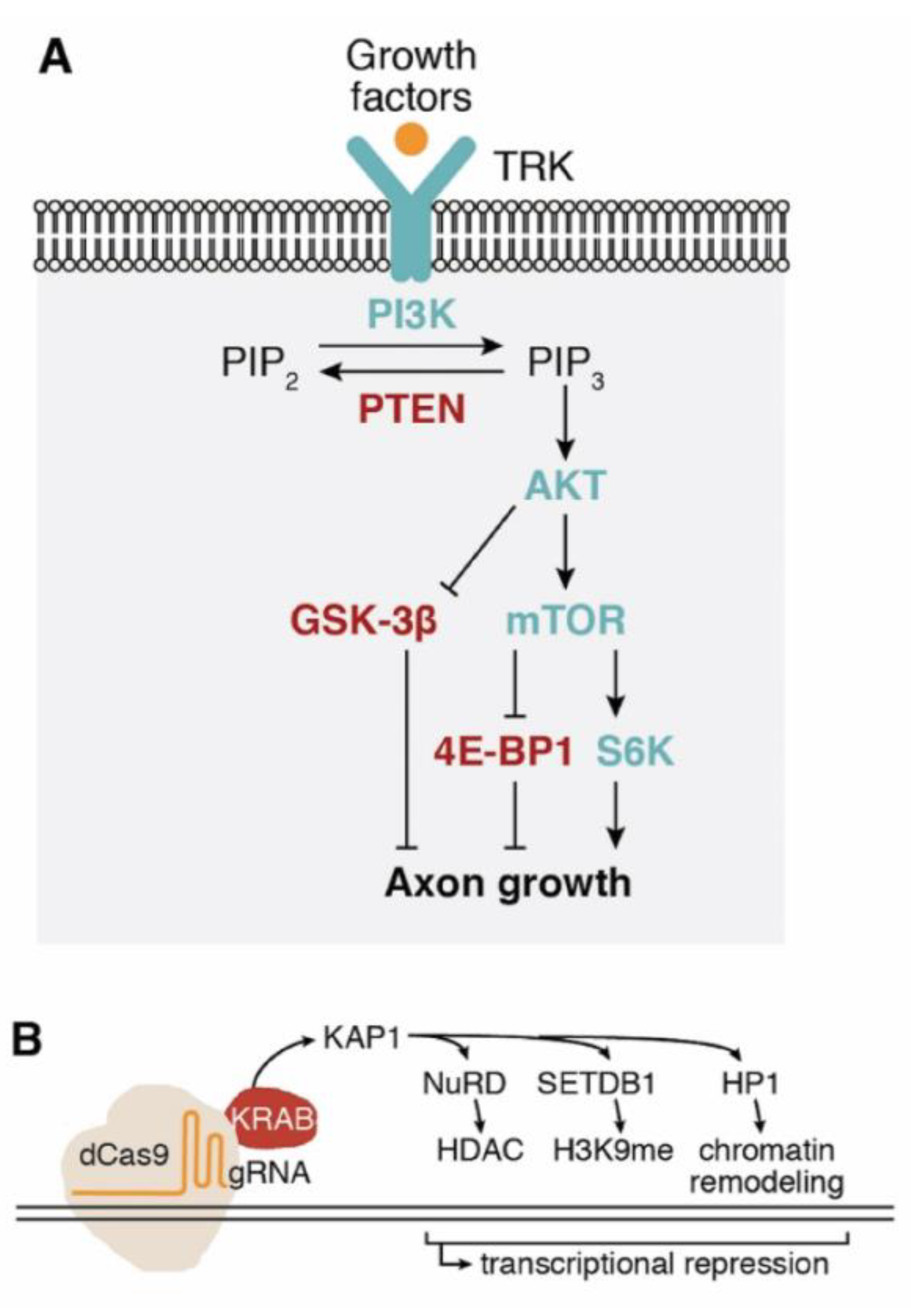
Figure 2.
Schematic of AAVrg-Cre–mediated PTEN knockout and tract-tracing workflows in a T9 compression-injury model. Retrograde AAVrg-Cre-RFP is injected 2-mm rostral to lesion at week 0 to excise PTEN in descending neurons and label their axons, followed by either (a) anterograde CST tracing via BDA injection into the motor cortex or (b) retrograde viability mapping via Fluorogold 2-mm rostral in cord. Three cohorts were studied: the acute treatment group (SCI + rostral-AAVrg at week 0, biweekly BBB scoring through week 9, harvest at week 9), the acute tract-tracing group (SCI + rostral-AAVrg at week 0, tracer at week 7, harvest at week 9), and the chronic treatment group (SCI at week 0, rostral-AAVrg at week 12, tracer at week 33, harvest at week 35) [19].
Figure 2.
Schematic of AAVrg-Cre–mediated PTEN knockout and tract-tracing workflows in a T9 compression-injury model. Retrograde AAVrg-Cre-RFP is injected 2-mm rostral to lesion at week 0 to excise PTEN in descending neurons and label their axons, followed by either (a) anterograde CST tracing via BDA injection into the motor cortex or (b) retrograde viability mapping via Fluorogold 2-mm rostral in cord. Three cohorts were studied: the acute treatment group (SCI + rostral-AAVrg at week 0, biweekly BBB scoring through week 9, harvest at week 9), the acute tract-tracing group (SCI + rostral-AAVrg at week 0, tracer at week 7, harvest at week 9), and the chronic treatment group (SCI at week 0, rostral-AAVrg at week 12, tracer at week 33, harvest at week 35) [19].
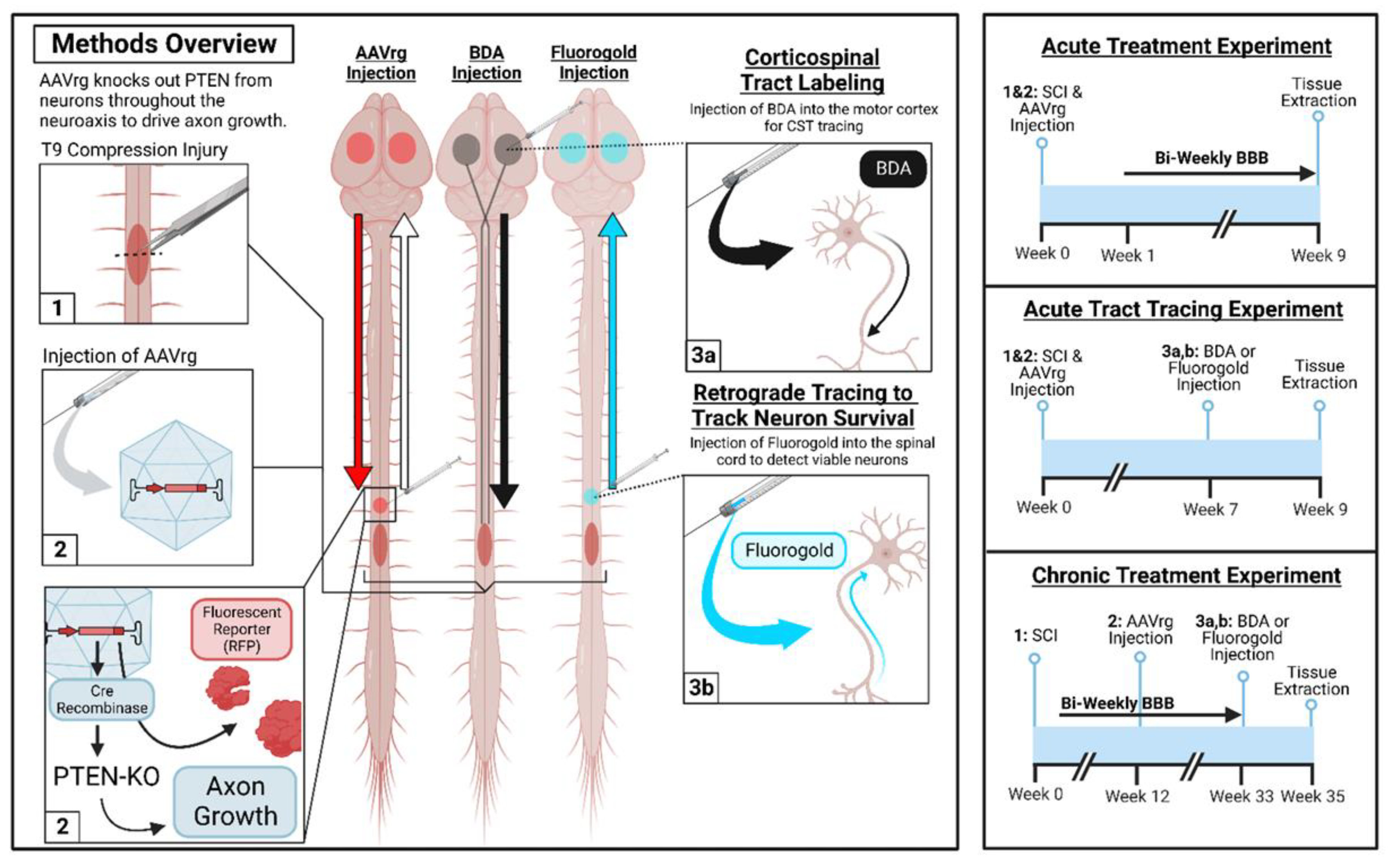
Figure 3.
Schematic of the remote-activated GMNPE hydrogel for combined neuronal regeneration and immunomodulation after SCI. Fe₃O₄@BaTiO₃ magnetoelectric nanoparticles are formed by coating Fe₃O₄ cores with a piezoelectric BaTiO₃ shell and then blending them with methacrylated gelatin (GelMA) and HUMSC-derived exosomes. When exposed to UV light, the photoinitiator crosslinks this mixture into a stable, exosome-loaded hydrogel. When implanted into the lesion cavity of a rat spinal cord hemisection model, the hydrogel remains inert until an alternating magnetic field induces strain in the Fe₃O₄@BaTiO₃ particles. This generates local electric potentials that both trigger controlled exosome release and directly stimulate resident neural cells. The resulting bioelectric and biochemical cues synergize to promote axonal outgrowth and remyelination while suppressing proinflammatory microglial activation, thereby accelerating functional repair. Adapted from Ref. [6] with permission.
Figure 3.
Schematic of the remote-activated GMNPE hydrogel for combined neuronal regeneration and immunomodulation after SCI. Fe₃O₄@BaTiO₃ magnetoelectric nanoparticles are formed by coating Fe₃O₄ cores with a piezoelectric BaTiO₃ shell and then blending them with methacrylated gelatin (GelMA) and HUMSC-derived exosomes. When exposed to UV light, the photoinitiator crosslinks this mixture into a stable, exosome-loaded hydrogel. When implanted into the lesion cavity of a rat spinal cord hemisection model, the hydrogel remains inert until an alternating magnetic field induces strain in the Fe₃O₄@BaTiO₃ particles. This generates local electric potentials that both trigger controlled exosome release and directly stimulate resident neural cells. The resulting bioelectric and biochemical cues synergize to promote axonal outgrowth and remyelination while suppressing proinflammatory microglial activation, thereby accelerating functional repair. Adapted from Ref. [6] with permission.
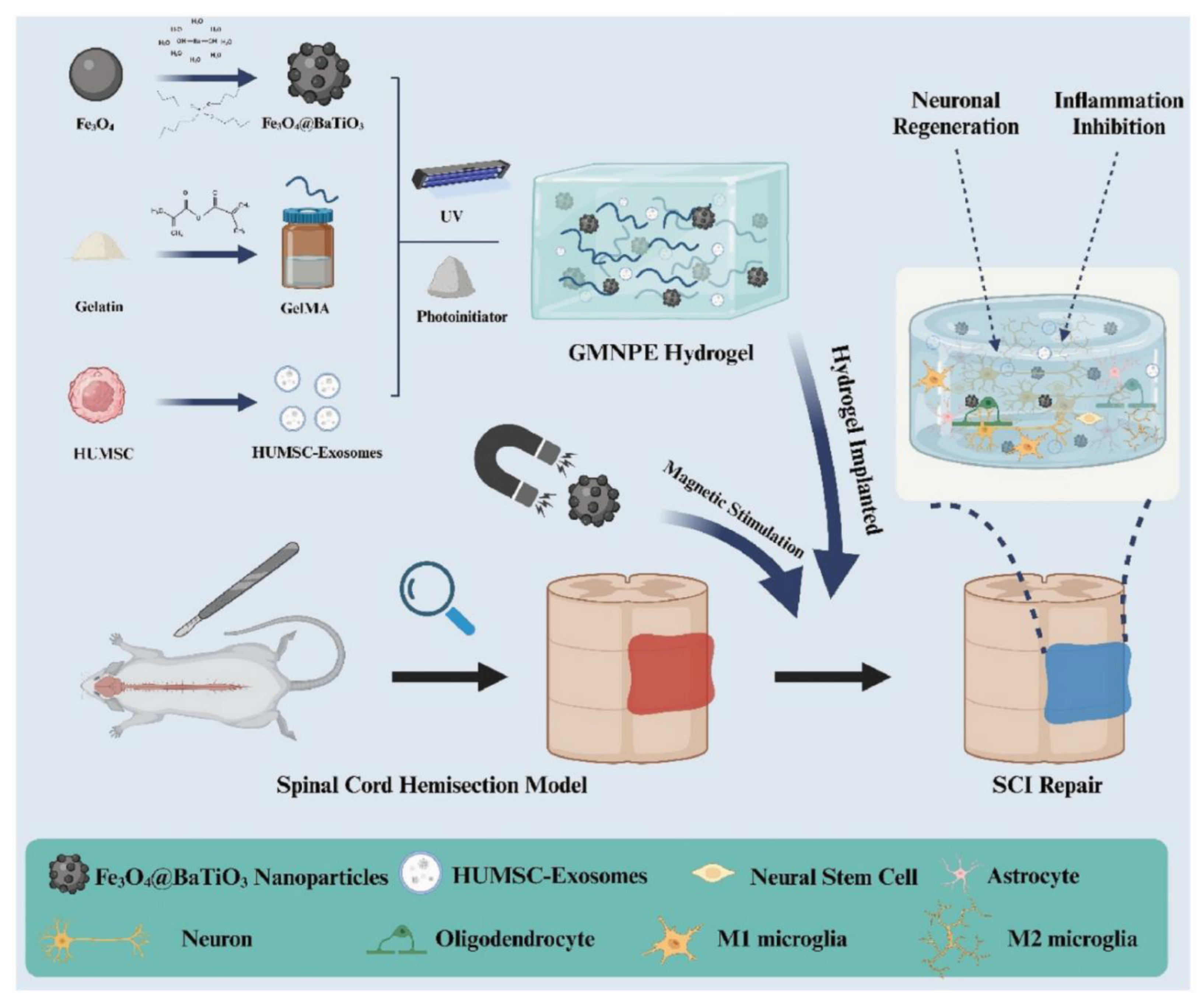
Figure 4.
Self-powered bioelectric cues from magnetoelectric nanoparticles and piezoelectric fibers. (A) Fe₃O₄@BaTiO₃ nanoparticles harness alternating magnetic fields to produce local electric potentials in a gelatin-methacrylate hydrogel (see Section 2.2). Magnetostrictive strain in the Fe₃O₄ core compresses the piezoelectric BaTiO₃ shell, generating surface polarization that depolarizes neuronal membranes. This opens L-type voltage-gated Ca²⁺ channels (L-VGCCs), triggers Ca²⁺-dependent signaling, and upregulates neurogenic proteins, ultimately driving remote neural stimulation and neurogenesis. (B) Aligned PVDF nanofibers convert cell-generated traction into a piezopotential that electrically cues stem-cell differentiation (see Section 2.2). During initial adhesion (slippage), the cells glide over unstressed fibers without deforming them, so no voltage is produced. As integrin-mediated tension increases (traction), fibers bend and create a local piezopotential that fosters neuronal outgrowth and maturation. Adapted from Ref. [6] with permission.
Figure 4.
Self-powered bioelectric cues from magnetoelectric nanoparticles and piezoelectric fibers. (A) Fe₃O₄@BaTiO₃ nanoparticles harness alternating magnetic fields to produce local electric potentials in a gelatin-methacrylate hydrogel (see Section 2.2). Magnetostrictive strain in the Fe₃O₄ core compresses the piezoelectric BaTiO₃ shell, generating surface polarization that depolarizes neuronal membranes. This opens L-type voltage-gated Ca²⁺ channels (L-VGCCs), triggers Ca²⁺-dependent signaling, and upregulates neurogenic proteins, ultimately driving remote neural stimulation and neurogenesis. (B) Aligned PVDF nanofibers convert cell-generated traction into a piezopotential that electrically cues stem-cell differentiation (see Section 2.2). During initial adhesion (slippage), the cells glide over unstressed fibers without deforming them, so no voltage is produced. As integrin-mediated tension increases (traction), fibers bend and create a local piezopotential that fosters neuronal outgrowth and maturation. Adapted from Ref. [6] with permission.
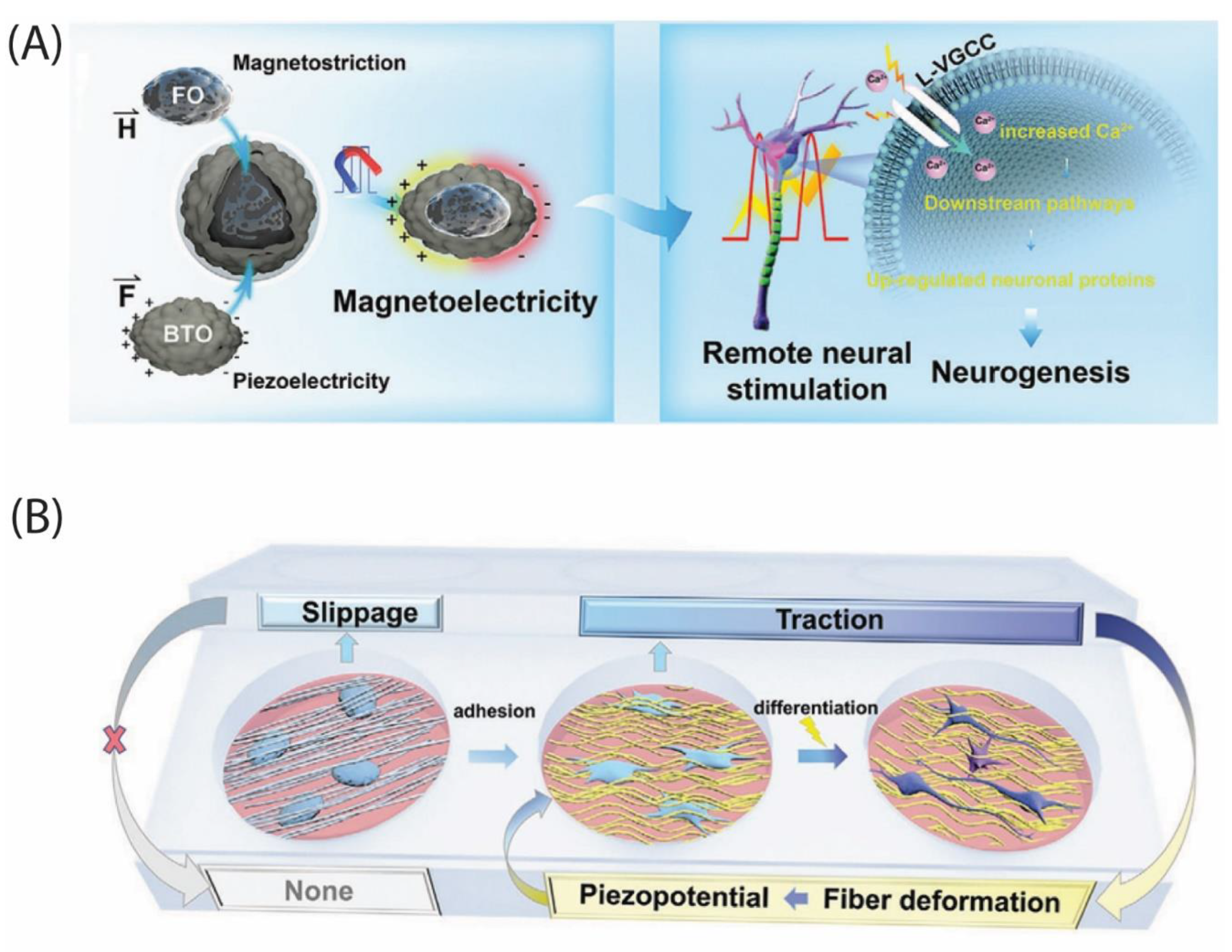
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



