
Submitted:
01 January 2026
Posted:
04 January 2026
You are already at the latest version
Abstract
Neural oscillations are fundamental to the integration of sensory, affective, and cognitive processes that contribute to pain perception. Transcranial alternating current stimulation (tACS) provides a valuable tool for investigating and modulating these oscillatory dynamics. In this review, we examine the effects of tACS on pain perception and pain-related oscillations in both healthy participants and individuals with chronic pain, highlighting methodological variability and mechanistic uncertainties that may contribute to mixed findings. We identified 14 studies, including 9 studies of experimental pain in healthy individuals and 5 of clinical pain disorders, comparing tACS to sham. Somatosensory alpha was the most frequently targeted oscillatory feature. Results varied considerably. Several studies reported reductions in pain, increases in alpha power, or changes in sensorimotor and prefrontal connectivity, but others showed no meaningful neural or behavioral effects. Out of the 14 studies, 6 demonstrated analgesic benefits and 2 showed improvements only under specific conditions or within subgroups, for a total of 8/14 studies with positive findings. Possible sources of heterogeneity include variation in stimulation duration, electrode montage, frequency alignment with individual rhythms, contextual state, and anatomical and neurophysiological differences across individuals. Pre-registered studies with sufficient power are needed to replicate effects within the most promising intervention protocols to establish a foundation in the field. We also recommend inclusion of brain imaging or electrophysiological recordings to verify whether stimulation effectively modulates the targeted neural oscillations. Finally, recent methodological advances, including phase-specific tACS, amplitude-modulated tACS, and individualized electric-field modeling, offer new opportunities to enhance mechanistic precision and clinical applicability. We argue that by integrating these approaches, future research can move beyond fixed, one-size-fits-all protocols, toward personalized, state-dependent, closed-loop tACS approaches. Exploring these frontiers will transform tACS from an exploratory tool into a reliable intervention for pain.
Keywords:
tACS
; pain perception
; neural oscillations
; alpha power
; experimental pain
; chronic pain
; brain imaging
; electrophysiological recordings
1. Introduction
Neural oscillations reflect neurophysiological mechanisms that support the integration of sensory, affective and cognitive processes involved in pain perception [1]. Human r have historically been characterized by their frequency, with major frequency bands including (from low to high) delta (0.5-4 Hz), theta (4-7 Hz), alpha (7-12 Hz), beta (12-30 Hz), and gamma (30-55 Hz for low gamma and 55-200 for high gamma). They are measured using time-frequency decomposition of scalp electroencephalography (EEG) or, less frequently, intracranial EEG (iEEG). These rhythms reflect synchrony across populations of neurons, and are thought to play a fundamental role in regulating which neurons are excitable at which times, allowing the formation of integrated networks of neuronal subpopulations across the brain [2]. Alpha rhythms are found across the brain and are associated with multiple functions, including attention [3] and sensory gating, where higher alpha power is thought to reflect stronger inhibition of incoming sensory input and lower cortical excitability – providing an important framework for interpreting alpha changes in pain [4]. Alpha 1 power (7-10 Hz), measured over temporal and somatosensory regions, has been found to negatively correlate with pain ratings both at rest and during phasic thermal stimulation [5]. Alpha amplitude is also shaped by cognitive and contextual factors, decreasing with attention to pain [6] and increasing with the expectation of pain relief [7]. A slowing of the individual peak alpha frequency (IAF) – reported in chronic pain conditions [8,9,10,11] – often occurs with increased theta and gamma activity, forming a pattern known as thalamocortical dysrhythmia, which is thought to reflect altered thalamic bursting and cortical hyperexcitability. In healthy individuals, lower IAF has been associated with increased pain sensitivity [12] and reduced nociceptive facilitation [13].
These associations offer promise for understanding pain and provide targets for neuromodulatory interventions. However, there is still uncertainty about how consistently brain oscillations relate to pain. Recent large-scale studies report no significant relationship between resting IAF and pain sensitivity, raising doubts about whether alpha slowing constitutes a stable or generalizable biomarker across pain types [14,15]. Instead, alpha dynamics appear to reflect broader cross-frequency reorganization within cortical networks. For example, Bott et al. [16] demonstrated that expectations modulate prefrontal to somatosensory connectivity at alpha frequencies, whereas gamma band coupling encodes prediction errors, consistent with predictive coding models of pain. In clinical populations, frequency-specific patterns further highlight distributed network involvement. In chronic low back pain, theta and alpha activity in the orbitofrontal cortex were linked to localized hypersensitivity, while anterior cingulate gamma and dorsolateral prefrontal theta tracked generalized hyperalgesia [17]. In neuropathic pain, higher alpha and lower delta and theta power were associated with increased pain intensity [18]. Thus, a growing body of evidence is at odds with the simple characterization of pain as a high-theta, low-alpha phenomenon. The source locations of measured rhythms, individual differences, and pain types and states may matter in complex ways. Human iEEG work further underscores this heterogeneity in oscillatory correlates of pain across individuals [19], though there may be some consistency across individuals as well. In Shirvalkar et al.’s study [19], which involved sustained iEEG recording in individuals with chronic pain across ~120 days, the most consistent correlates of pain were decreased anterior cingulate alpha/beta power for experimental heat pain and reduced orbitofrontal delta for chronic spontaneous pain.
Given the importance of brain rhythms in circuit integration and behavior, including pain,, transcranial alternating current stimulation (tACS) provides a relatively unique and important opportunity to stimulate the brain at particular frequencies [20]. TACS is a noninvasive method that delivers low-intensity sinusoidal electrical currents through scalp electrodes. When tACS is applied at a frequency close to the brain’s own ongoing rhythm, it can entrain neuronal activity by aligning the phase of neural oscillations with the stimulation waveform [21]. This effect is believed to follow the so-called Arnold tongue principle, in which the strength of entrainment increases as the stimulation frequency more closely matches the endogenous oscillatory frequency [22,23]. Another proposed mechanism involves the facilitation of synaptic plasticity via spike-timing dependent plasticity (STDP), which may allow tACS to enhance oscillatory power at the stimulated frequency [24]. The application of tACS at the IAF has been proposed to promote effective neuronal synchronization [25], to stabilize IAF against potential slowing [26], and to augment its power and gating functions. More broadly, tACS at frequencies beyond alpha may be warranted, as pain-related cortical networks show complex cross-frequency reorganizations rather than isolated alpha-band alterations [16,17,18,19].
2. Current Evidence on the Effects of tACS on Experimental and Clinical Pain
Across the growing literature on tACS, studies have examined whether rhythmic electrical stimulation can alter pain perception, modulate oscillatory activity, or reduce chronic pain symptoms. Although findings are heterogeneous, converging evidence indicates that tACS can modulate pain perception and pain-related neural activity under specific contextual and stimulation conditions. Specifically, outcome variability appears to depend on stimulation parameters (frequency, montage, intensity, duration), individual neurophysiology, and task.
Table 1 summarizes the key design features and outcomes across all published tACS experimental and clinical pain studies included in this review.
These studies exhibit substantial methodological variability. Sample sizes are typically small, and stimulation parameters including frequency, intensity, montage, electrode configuration, and duration vary widely. Many experiments also differ in whether they use individualized frequencies, stimulation focality, and whether electric field modeling or target engagement measures are reported. This heterogeneity makes it difficult to directly compare results across studies or to identify which stimulation features contribute most reliably to analgesic or neural effects. Overall, these protocol differences may underlie the inconsistencies observed in the literature. Power is also likely to be an issue. To detect a moderate effect size of Cohen’s d = 0.5 with 80% power, N = 34 participants are required for a within-person crossover and N = 130 (65 per group) are required for a between-person comparison across groups. By this standard, only two studies in Table 1 are adequately powered to detect moderate effect sizes.
2.1. Evidence from Experimental Pain Studies
Experimental pain studies show a wide range of outcomes, from clear neural modulation to limited or null effects on pain perception. Across studies, tACS altered oscillatory activity within sensorimotor and prefrontal networks, yet these neural changes did not consistently translate into behavioral analgesia.
Particularly, alpha-frequency stimulation over the somatosensory cortex has shown mixed results. Arendsen et al. [27] reported a reduction of both pain intensity and unpleasantness during α-tACS, but only under conditions of uncertain pain expectation, suggesting an interaction between oscillatory modulation and cognitive context. In contrast, May et al. [30] found no modulation of pain or autonomic responses during α- or γ-tACS over the prefrontal cortex or S1, with Bayesian analyses supporting the null hypothesis for most conditions. Some studies targeting SM1 with α-tACS also showed dissociations between neural and behavioral effects. Peng et al. [31] reported reduced pain-evoked activity and decreased SM1-network connectivity, although pain ratings did not significantly differ from sham. Fathi et al. [26], using individualized somatosensory IAF stimulation, likewise observed no overall analgesic effect, although exploratory analyses suggested reduced sensitization in women and possible stabilization of both alpha rhythms and pain perception. Li et al. [29] reported no reduction in pain, but observed disruption of laser-evoked potential habituation and changes in α₁ and θ activity during anticipation. Mediation analyses further suggested competing influences on pain, with enhanced α₁ oscillations tending toward analgesia, whereas increased θ activity was associated with hyperalgesia.
Some studies have shown more robust perceptual effects using longer stimulation durations and high-definition (HD) montages. Qi et al. [32] reported that 10 Hz HD-tACS over SM1 reduced capsaicin-induced pain, and resting α power and pain-related θ modulation predicted individual responsiveness. Sun et al. [33] found that HD-tACS over the dorsolateral prefrontal cortex (DLPFC) reduced pain and produced delayed increases in low γ power, although EEG was recorded only at rest.
Other frequency bands have also been explored. Ikarashi et al. [28] demonstrated that θ- and β-tACS over the DLPFC increased heat-pain thresholds, with θ-tACS showing an inverted U-shaped relationship between modeled electric field strength and analgesic efficacy, suggesting that both insufficient and excessive stimulation can reduce efficacy.
Finally, interpersonal and affective factors can shape tACS outcomes. Takeuchi and Terui [34] showed that synchronous dual-brain α-tACS enhanced social touch-induced analgesia in low-empathy individuals, even though overall pain ratings did not differ across conditions. These results highlight that tACS effects may depend not only on stimulation parameters but also on social and emotional context.
Together, findings on experimental pain show that tACS can alter pain-related neural activity and occasionally reduce pain perception, but results vary considerably across studies. More consistent effects may emerge when stimulation is longer, spatially focused, tailored to individual oscillatory properties, or applied in contexts that shape pain processing, such as expectation, uncertainty or social interaction.
2.2. Evidence from Chronic Pain Studies
Clinical studies generally provide more consistent indications that tACS can influence ongoing chronic pain symptoms, although sample sizes are small and protocols vary widely. Across chronic low back pain and fibromyalgia research, stimulation at individualized or symptom-related frequencies tends to be more effective than fixed-frequency protocols.
In chronic low back pain, Ahn et al. [35] found that 10 Hz bifrontal tACS reduced pain ratings relative to sham and increased somatosensory α power, with greater α enhancement predicting larger reductions in pain and disability. Prim et al. [39] also applied 10 Hz tACS over prefrontal regions and observed autonomic modulation (increased heart rate variability) and a higher proportion of clinical responders in the active condition, although group-level pain reductions did not reach statistical significance.
Fibromyalgia studies provide mixed but informative results. Bernardi et al. [37] tailored the frequency of stimulation using 4 Hz for slow-rhythm abnormalities and 30 Hz for fast-rhythm abnormalities, and observed reductions in pain intensity, improvements in cognitive and affective symptoms, and increases in α₁ power, with effects diminishing after four weeks. In contrast, Lin et al. [38] applied 50 Hz HD-tACS over M1 and found no significant differences between active and sham overall, although active stimulation reduced fibromyalgia impact scores relative to baseline.
Clinical protocols also vary widely in stimulation frequency, intensity, montage, electrode configuration, and number of sessions, ranging from single session interventions to multi-day treatment protocols. Despite this heterogeneity, several patterns emerge. Longer or repeated stimulation tends to show more consistent symptom improvement. Tailoring stimulation frequency based on patient stratification may yield stronger effects than using fixed frequencies. And finally, increases in α power or normalization of abnormal oscillatory patterns are often associated with reductions in pain.
3. Limitations, Sources of Inconsistencies, and Possible Solutions
3.1. Mechanistic Uncertainty Driven by Target and Frequency Variability
Despite the promise of tACS for both experimental [27,33] and clinical [35,36] pain modulation, a major obstacle to clinical translation remains the lack of mechanistic clarity regarding both the optimal cortical targets and stimulation frequencies. Pain processing is not localized in a single brain region, but it is highly distributed and involves contributions from sensorimotor, prefrontal, limbic, and cingulo-opercular circuits, each oscillating at multiple characteristic rhythms [1]. Moreover, individuals suffering from chronic pain can also exhibit elevated activity in limbic brain regions [40,41,42], and other brain network-level abnormalities [43,44,45,46,47], further complicating the search for analgesic targets and frequencies for clinical applications.
Although alpha activity prior to pain onset correlates with lower pain ratings in some studies [5,48,49], somatosensory alpha-tACS has not consistently modulated pain. For instance, rigorous sham-controlled studies of tonic pain failed to show robust analgesic effects for either somatosensory alpha- or prefrontal gamma-tACS, suggesting that simple one-to-one mappings between rhythm and perception may be insufficient [30]. Power is likely an issue, however, as mentioned above, and larger pre-registered studies must be done to more definitively address this issue. Complementary imaging-based work has demonstrated that alpha-tACS over M1 reduces both bilateral SM1 activity and connectivity with pain-related regions, with changes emerging most prominently after stimulation rather than during, pointing toward plasticity mechanisms that extend beyond immediate entrainment [31].
Increasing evidence also points to nonlinear dose–response relationships [28]. These nonlinearities likely arise from complex interactions between endogenous oscillatory states, network resonance frequencies, and behavioral context [1]. The mechanistic uncertainties surrounding target site selection and frequency tuning therefore represent a critical barrier: without biomarker-guided strategies, stimulation protocols risk inconsistency, with clinical trials reporting modest or null effects despite promising preclinical rationales [51]. Establishing causal links between frequency-specific entrainment, network-level modulation, and pain reduction is therefore essential for moving toward precision neuromodulation.
3.2. Sources of Inter-Individual Variability
A major challenge for effective tACS interventions is the substantial inter-individual variability in stimulation outcomes. This variability arises from a combination of anatomical, physiological, and contextual factors that shape the degree to which electrical fields engage targeted neural circuits. Structural MRI–informed modeling suggests that individual differences in head and brain anatomy lead to substantial variability in intracranial electric field strength and distribution, even when identical stimulation montages are used [28]. As a result, two individuals receiving the same nominal current may experience very different levels of cortical engagement, and behavioral efficacy may follow non-linear patterns, including inverted U relationships between field magnitude and analgesic effects. Such effects, however, remain to be replicated in well-powered studies. Variability is further amplified by the diversity of stimulation targets across studies, as sensorimotor, prefrontal, and dorsolateral prefrontal regions engage distinct pain-related networks and produce different patterns of neural modulation [27,28,29,33].
Physiological traits further contribute to variability. Baseline oscillatory dynamics, such as resting alpha power and IAF, are thought to be predictors of pain sensitivity and of responsiveness to stimulation. In controlled experiments, resting alpha power and pain-related theta reactivity jointly mediated responses to alpha HD-tACS over SM1.
Sex and gender differences may also be a factor. Complementary work has shown sex-specific effects in which women exhibited stronger modulation of heat-pain thresholds [26]. Biological rhythms, including the menstrual cycle, introduce additional variability through their effects on pain thresholds, cortical excitability, and oscillatory activity. These findings underscore the relevance of individualized dominant frequency and sex-linked neurophysiological characteristics as important covariates in stimulation research.
State-dependent psychological and contextual factors also play a significant role. In at least one study, alpha-tACS reduced pain only when stimulus intensity was uncertain and increased reactivity under certainty, demonstrating the interplay between expectancy, cortical rhythms, and sensory processing [27]. Related work has shown that alpha-tACS interacts with anticipatory alpha and theta dynamics in ways that influence pain processing [29]. Psychological traits such as empathy and, in some studies, pain-related cognitions such as catastrophizing, modulate tACS effects on social and affective dimensions of pain [52]. Autonomic differences may contribute as well, with some studies reporting increases in heart-rate variability indices after tACS and others finding no change, suggesting that systemic physiological state influences neural responsiveness [38,39]. Together, these findings show that inter-individual variability is not simply noise but a defining feature of how tACS interacts with complex biological systems. Accounting for anatomy, baseline oscillations, sex-related physiology, and state-dependent context is essential for improving reproducibility,mapping the effects of stimulation, and understanding the factors that moderate effectiveness.
To address this variability, current research is moving toward individualized and model-informed neuromodulation strategies [28]. Tailoring current intensity and electrode placement may help to achieve an optimal field distribution for each individual rather than relying on uniform stimulation parameters.
Neuroimaging provides a complementary path toward personalization. fMRI studies have shown that alpha-tACS over primary motor cortex alters local activity and connectivity with cingulate and prefrontal control regions, pointing to network-level mechanisms that can be monitored and optimized [31]. EEG biomarkers further refine this approach. Increases in low gamma power after alpha HD-tACS predict stronger analgesic responses [33], and resting alpha and theta reactivity predict SM1-specific responsiveness [32].
Individualization also extends to frequency selection. Aligning stimulation with each participant’s intrinsic alpha frequency improves entertainment and enhances modulation of pain-related processes compared with fixed-frequency approaches [37]. Reviews consistently highlight the value of combining electric-field modeling with EEG and fMRI biomarkers to guide montage design, frequency tuning, and current scaling [53]. Early clinical trials in fibromyalgia, although mixed in behavioral outcomes, demonstrate that multi-session HD-tACS can be safely implemented and provide a foundation for biomarker-guided personalization [37,38].
Altogether, these findings suggest that the most promising path forward lies in integrating electric-field modeling, individualized oscillatory biomarkers, and network-level imaging targets within adaptive, state-dependent stimulation protocols. Personalization of stimulation parameters offers a direct solution to the inter-individual variability that limits the reliability and scalability of tACS for pain relief.
3.3. Mechanistic Uncertainty Due To Lack of Simultaneous Monitoring of Oscillations During tACS
A clearer mechanistic understanding of how tACS modulates neural activity and behavior is essential for optimizing stimulation parameters such as amplitude and duration to achieve consistent analgesic effects. While many studies focus on aftereffects linked to plasticity, the online effects of tACS - particularly neuronal entrainment during stimulation - are less understood. Directly tracking neural activity during tACS would help clarify these mechanisms, but concurrent EEG recordings are technically challenging due to strong stimulation artifacts. Beyond this, current tACS protocols are typically fixed throughout stimulation, whether individualized or not, whereas optimal neuromodulation likely requires parameters that adapt dynamically to ongoing brain activity. Real-time monitoring would enable such adaptive or closed-loop frameworks by aligning stimulation parameters to moment-to-moment oscillatory dynamics.
To overcome stimulation artifacts and enable online monitoring of neuronal entrainment, Witkowski et al. [54] introduced amplitude-modulated tACS (AM-tACS), where the intended low-frequency stimulation pattern is embedded within a high-frequency carrier signal (Figure 1). Because neuronal membranes act as low-pass filters, the high-frequency carrier is too fast to modulate neural activity directly, whereas the low-frequency envelope can influence oscillations. During recording, appropriate low-pass filtering removes the carrier, allowing access to the underlying neural signal. However, nonlinearities in stimulation and recording hardware can introduce residual artifacts, and additional denoising approaches have been developed to more reliably recover the neural activity [55,56,57].
While AM-tACS has been shown to allow concurrent recordings and, with advanced denoising approaches such as stimulation artifact source separation (SASS), to reveal phase-dependent modulation of oscillations and behavior [55,56], other studies have questioned its efficacy. For example, phosphene-based evaluations indicated that AM-tACS effects rely primarily on the carrier frequency rather than the modulation frequency, and suggested that substantially higher stimulation intensities might be required for amplitude modulation to exert reliable neuromodulatory effects [58]. Therefore, while AM-tACS provides a valuable tool for artifact-free recording, its efficacy as a neuromodulatory technique relative to conventional tACS requires further validation.
3.4. Phase-Dependent and State-Dependent Modulation in tACS
A review of Table 1 indicates that parameter settings have to this date concentrated mainly on modifications of stimulation amplitude and frequency. An important, underappreciated consideration is the phase of stimulation, which has recently been shown to play a critical role, particularly in domains such as visual processing [55] and working memory [56]. When we refer to the phase of stimulation in tACS, this can denote two distinct concepts: (1) the phase of the stimulation relative to the phase of ongoing brain oscillations, and (2) the phase difference between electrode pairs in multi-electrode montages.
We discuss each of these below and highlight how they may critically influence tACS effectiveness and contribute to heterogeneous findings in human pain studies.
3.4.1. tACS Interaction with Ongoing Brain Oscillations
While tACS is often assumed to entrain brain oscillations at their endogenous frequency, recent evidence indicates that it interacts with ongoing neural activity rather than merely amplifying it. Using electrophysiological recordings in awake rhesus monkeys, Krause et al. [59] demonstrated that tACS does not simply impose rhythmicity on neurons but instead competes with intrinsic oscillations for control of spike timing. These recordings revealed that neurons with weak baseline entrainment became more rhythmic during tACS, whereas strongly entrained neurons often lost synchrony. The findings explain variability in human tACS/EEG studies and suggest opportunities for both strengthening and desynchronizing neural activity.
In humans, Fiene et al. [60] provided complementary evidence by pairing visual flicker with occipital tACS at different phase shifts. They showed that steady-state visual responses were modulated in a phase-dependent manner, and that the optimal phase shift correlated with each participant’s cortical response delay. These findings highlight that the effectiveness of tACS depends critically on the timing of stimulation relative to ongoing brain rhythms. Haslacher et al. [55,56] provided evidence that the impact of tACS critically depends on its phase relationship to ongoing oscillations. They showed that stimulation at specific phases could enhance or suppress both neural activity and perceptual dominance, and later demonstrated that working memory performance was maximized at distinct phase lags that aligned with changes in frontoparietal alpha synchrony. Another example showing the importance of timing is provided by Schreglmann et al. [61], who demonstrated that non-invasive cerebellar tACS phase-locked to the tremor rhythm in essential tremor patients suppressed tremor amplitude by disrupting the temporal coherence of pathological oscillations, with the effective phase lag varying across individuals.
Together, these studies demonstrate that the efficacy and even the direction of tACS effects (enhancement or suppression) are not fixed but depend on phase alignment with ongoing neural activity and brain state, varying not only across individuals but also across time within the same individual. For pain modulation, this implies that the analgesic effect of, for example, alpha-tACS may depend on applying the stimulation at the correct phase of the individual’s ongoing sensorimotor alpha rhythm. Current protocols do not account for this, potentially explaining why some participants respond and others do not. At the same time, state-dependent suppression can be therapeutically exploited. For instance, in neuropathic pain conditions characterized by thalamocortical dysrhythmia, tACS could be tuned to selectively suppress aberrant low-frequency theta or delta oscillations.
3.4.2. Phase Configuration of Multi-Electrode tACS Montages
In pain studies employing multi-electrode tACS montages, in-phase stimulation has often been assumed to maximize the electric field strength at the target site. However, as demonstrated by Alekseichuk et al. [62], the magnitude and distribution of intracranial electric fields are not linearly determined by electrode currents. Their invasive recordings in nonhuman primates showed that the strongest electric fields do not necessarily occur during in-phase stimulation and can instead emerge at different phase offsets between electrodes. This challenges the common assumption that in-phase configurations always produce the strongest and most focal fields, highlighting the need to treat stimulation phase as a critical parameter in tACS protocols. Moreover, they demonstrated that specific phase configurations can generate a novel phenomenon, called “traveling waves”, in which the peak electric field shifts dynamically across the brain during the stimulation cycle.
Other studies suggest that phase-shifted tACS can modulate neuronal oscillations in a spatiotemporal manner and facilitate wave propagation (beyond local entrainment) [63,64]. This pattern introduces a new dimension to how multi-electrode tACS can modulate brain activity and raises the possibility of mimicking or deliberately disrupting the natural propagation of oscillatory activity across pain-related regions (e.g., from sensory to affective regions), offering a new tool to investigate network dynamics.
3.5. Limited Access to Deep-Regions as a Constraint for Pain Modulation
In general, noninvasive brain stimulation techniques lack the capability to target deep brain regions effectively without affecting surrounding areas. As pain encompasses sensory, affective, and cognitive dimensions, effective modulation may require to selectively target brain regions involved in these processes. One such region is the insula, a key hub for integrating sensori-discriminative and affective-cognitive aspects of pain [65,66,67]. The posterior insula is more strongly associated with processing nociceptive input [68], whereas the anterior insula is most closely associated with affective and evaluative aspects [69]. Dysregulation of insular activity is often observed in chronic pain [66], and lesions in the posterior insula can abolish the unpleasantness of pain while preserving sensory detection [70]. Furthermore, iEEG evidence suggests that nociceptive pain preferentially modulates theta and alpha oscillations in the dorsal posterior insula [71], indicating that this subregion could be a promising stimulation target.
Targeting insula for pain treatment has been investigated in several studies. A recent rodent study showed that repetitive insular cortex stimulation alleviated neuropathic pain by normalizing synaptic plasticity, specifically reducing phosphorylated CRMP2, AMPAR, and NR2B expression, and dampening insular hyperactivity [72]. A pilot intracranial study in humans showed that anterior insula stimulation raised heat pain thresholds without adverse effects, supporting its role in the affective–cognitive modulation of pain [73]. Together, these converging lines of evidence underscore the insula as a promising therapeutic hub for targeting both sensory and affective dimensions of chronic pain.
However, invasive insular stimulation for pain remains limited in scope, and its clinical application is constrained by surgical risks and outcome variability. A noninvasive method capable of selectively modulating the insula without affecting the surrounding cortex would therefore offer a safer and more accessible therapeutic option while also enabling deeper investigation into the insula’s mechanistic role in pain. The same principle applies to other regions important in pain construction.
Temporal interference stimulation (TIS) builds on the principle of amplitude-modulated tACS, where two electrode pairs deliver high-frequency currents with slightly different frequencies, creating a low-frequency envelope at their intersections that can modulate neural activity (Figure 2). Grossman et al. demonstrated in mice that such deep activation is possible without engaging in the overlying cortex, and that by adjusting current ratios, the stimulation locus can be shifted [74], opening the possibility to selectively target subregions such as specific insular subregions. This steerability and focality make TIS a particularly attractive approach for noninvasively modulating deep structures involved in pain. A systematic review of human TIS studies [75] suggests that although preliminary findings indicate frequency- and target-specific modulation of cortical and subcortical activity, the evidence remains limited by small sample sizes and methodological heterogeneity. Larger, well-controlled trials are needed to establish clinical efficacy.
3.6. Methodological Limitations, Blinding Challenges, and Recommendations for Future Studies
Across the current literature, several methodological limitations contribute to the inconsistent findings regarding the effects of tACS on pain. A prominent source of variability lies in the wide range of stimulation parameters used across studies. Frequencies span from theta to high gamma, with some studies using fixed frequencies and others applying individualized alpha stimulation. Intensities span from subthreshold values to multi-milliamp protocols, and electrode configurations range from large sponge pads to high definition arrays. Because electric fields depend strongly on montage geometry and individual anatomy, the absence of standardized reporting of modeled electric fields makes it difficult to determine whether null or positive findings reflect genuine physiological effects or simply inadequate dosing. This issue is further complicated by the lack of systematic investigation into dose–response relationships. Very few studies examine how variations in frequency, current amplitude, or field strength influence neural or behavioral outcomes, leaving many assumptions about optimal stimulation parameters untested.
Stimulation duration also shows considerable heterogeneity and appears to interact with efficacy. Short 10-minute protocols often fail to produce measurable changes in pain or oscillatory activity, whereas longer sessions of 30-40 minutes tend to yield more consistent neural modulation and, in some clinical cases, reductions in pain. Some experiments report neural effects only after stimulation has ended, suggesting that aftereffects and plasticity mechanisms may be more relevant than online entertainment. Other studies observe only immediate changes during stimulation with no lasting impact. Because few studies include systematic follow-up assessments, the temporal dynamics of tACS effects remain poorly understood. Without controlled comparisons of short versus long protocols, or repeated session designs, it is unclear whether inconsistencies arise from insufficient stimulation duration or from more fundamental limitations in how tACS interacts with pain-related networks.
Blinding quality is another major concern, as sensory experiences during stimulation can differ substantially between active and sham conditions. Sham protocols vary widely, from brief ramp periods to one minute of stimulation, and only a minority of studies evaluate whether participants can reliably distinguish between conditions. Because expectation strongly modulates pain perception, inconsistent sham procedures introduce an important confound that may obscure or exaggerate stimulation effects.
Variability in outcome measures and pain models further limits comparability across studies. Experimental paradigms include tonic heat, laser stimulation, electrical pain, pressure pain, and capsaicin sensitization, each engaging partially distinct sensory and cognitive processes. Chronic pain studies involve different diagnoses, symptom profiles, and comorbid conditions. Neural readouts also vary widely, with some studies relying solely on behavioral ratings, others measuring resting EEG before and after stimulation, and still others collecting fMRI activity during pain. Differences in the timing of measurements relative to stimulation further add to inconsistency. Together, these factors make it difficult to identify stimulation features that consistently influence pain or pain related neural activity.
Individual differences also play a meaningful role in shaping responsiveness to tACS. Skull thickness, cortical folding, and cerebrospinal fluid distribution produce large inter-individual differences in electric field strength even when identical parameters are used. Baseline oscillatory properties, including resting alpha power, IAF, and pain related theta reactivity, predict whether stimulation modulates pain or neural activity. Sex related factors and hormonal variability contribute additional sources of variability. Psychological variables such as attention, expectancy, uncertainty, empathy, and catastrophizing further modulate responses to stimulation. These observations indicate that variability is not random noise but reflects the complex and dynamic nature of pain networks and their interaction with external rhythmic stimulation.
Taken together, inconsistent findings likely arise from a combination of heterogeneous stimulation parameters, variable sham procedures, insufficient exploration of dose–response and duration effects, limited verification of neural target engagement, diverse pain models, and substantial inter-individual differences. Future research would benefit from systematic testing of stimulation duration and intensity, clearer reporting of modeled intracranial electric fields, standardized sham protocols, larger and adequately powered samples, and direct measures of neural engagement during or immediately after stimulation. Incorporating individualized oscillatory biomarkers, anatomical modeling, and repeated session designs will also help clarify the conditions under which tACS reliably influences pain networks. Addressing these methodological challenges in a coordinated, evidence-based manner is essential to determine whether tACS can provide consistent and meaningful pain modulation Emerging tACS techniques are increasingly designed to target complex oscillatory dynamics and interactions [76]. Beyond the approaches discussed in this study, other strategies may help advance our understanding of pain and the role of oscillations in its processing. One example is cross-frequency coupled tACS (CFC-tACS) [77], which, rather than applying a single frequency, targets coupled oscillations to modulate their interactions. Another example is broadband tACS, which uses non-sinusoidal patterns that deliver a range of frequencies instead of a narrow sinusoid, thereby mimicking the spectral complexity of natural neural activity. Applying these techniques to pain requires systematic investigation to evaluate their potential contributions to pain modulation mechanisms.
4. Conclusions
Across experimental and clinical studies, tACS has demonstrated the capacity to modulate oscillatory activity within pain-related networks, yet its behavioral effects remain inconsistent. Evidence reviewed here shows that stimulation efficacy depends critically on several interacting factors: the choice of stimulation parameters, individual neuroanatomical and physiological characteristics, and the cognitive or emotional context in which stimulation is delivered. Studies employing longer stimulation durations, high-definition electrode montages, and frequencies tuned to each participant’s intrinsic oscillatory rhythms tend to show more reliable effects on both neural oscillations and pain perception [27,32,33]. These findings highlight that tACS does not exert uniform effects but instead interacts dynamically with ongoing network states.
A consistent observation across the literature is that tACS can alter cortical rhythms implicated in nociceptive processing, particularly within the alpha, theta, and gamma bands. Alpha-frequency stimulation over sensorimotor or prefrontal regions can modulate oscillatory power and functional connectivity within networks that integrate sensory, emotional, and cognitive aspects of pain [31,32]. These results agree with broader evidence linking alpha oscillations to top-down inhibitory control and expectancy mechanisms [1,5,16]. However, alpha enhancement does not consistently translate into analgesia, some studies show that alpha-tACS reduces pain under uncertain conditions [27], while others report no perceptual effect or even an increase in pain sensitivity when stimulation alters activity in the theta range [29]. These inconsistencies support the view that oscillatory modulations are inherently network-level phenomena rather than single-frequency biomarkers of pain inhibition. The simultaneous involvement of theta and gamma rhythms in pain modulation [17] indicates that tACS may influence cross-frequency coordination that supports sensory precision and emotional regulation.
Inter-individual variability represents another major source of inconsistency in tACS outcomes. Anatomical factors such as skull thickness and cortical folding can change how electric fields propagate through the brain, leading to large differences in the effective dose across participants. Stimulation effects also appear to follow a nonlinear pattern, where both insufficient and excessive field strengths can fail to produce modulation [28]. Physiological characteristics further contribute to this variability. Resting alpha power, IAF, and sex-related hormonal shifts all affect responsiveness to tACS [26,32]. Psychological factors, including attention, expectancy, and uncertainty additionally modulate behavioral effects, underscoring the state-dependent nature of neuromodulation [1,16]. These sources of variability are not simple noise, but reflect the complex, distributed and dynamic nature of pain.
Methodologically, several gaps currently limit mechanistic insight. Most studies rely on pre-post measures and therefore cannot distinguish transient entrainment from plasticity-driven aftereffects [24,59]. Recent advances in artifact-resistant recordings, including AM-tACS combined with denoising techniques such as stimulation artifact source separation, show that online monitoring is feasible and that neural responses depend critically on the precise timing of stimulation relative to ongoing brain rhythms [55,56]. The phase relationship between multiple electrodes is another factor that can determine the distribution of electric fields across the cortex [62]. Specific phase configurations can create traveling wave-like effects, offering new ways to manipulate the spatial and temporal characteristics of cortical activity. At the same time, standard tACS protocols have limited ability to engage deeper structures such as the insular cortex and the thalamus, which are central to pain processing [65,66,69]. Techniques such as TIS [74] and cross-frequency-coupled tACS offer promising avenues for accessing deeper or more complex oscillatory interactions, though empirical evidence remains preliminary [63,64].
As methodological rigor increases and individualized approaches become more common, future studies will be better positioned to determine the conditions under which tACS can consistently engage pain-related networks and contribute to effective neuromodulation. Combining tACS with behavioral or cognitive interventions, such as expectancy training or pain distraction tasks, may also amplify neuromodulatory effects through synergistic learning and plasticity mechanisms [35,37]. For the field to progress, it will be essential to adopt standardized protocols, report electric field strengths in addition to current amplitude, preregister experimental designs, and conduct adequately powered studies. Only through such methodological rigor will it be possible to translate tACS from an experimental technique into a dependable clinical tool for pain modulation.
Overall tACS offers a promising but still inconsistent approach for modulating neural oscillations involved in pain. Across studies, variability in stimulation parameters, individual anatomy and physiology, psychological state, and methodological rigor contributes to mixed behavioral outcomes. Evidence indicates that tACS can reliably influence pain-related rhythms under specific conditions – particularly when stimulation is longer, more focal, and tailored to individual oscillatory profiles. Progress toward clinical translation will require standardized protocols, individualized electric field modeling, real-time monitoring of neural engagement, and systematic evaluation of dose, duration, frequency, and phase. With these advances, tACS may ultimately develop into a precise and noninvasive tool for modulating pain and probing the neural mechanisms underlying its perception.
Author Contributions
Y.F. and A.D. writing of original manuscript draft. D.M.G. writing, review and editing of the final manuscript draft. G.L and T.D.W supervision, review and editing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ploner, M.; Sorg, C.; Gross, J. Brain Rhythms of Pain. Trends Cogn. Sci. 2017, 21, 100–110. [Google Scholar] [CrossRef]
- Fries, P. Rhythms for Cognition: Communication through Coherence. Neuron 2015, 88, 220–235. [Google Scholar] [CrossRef]
- Foxe, J.J.; Snyder, A.C. The Role of Alpha-Band Brain Oscillations as a Sensory Suppression Mechanism during Selective Attention. Front Psychol 2011, 2, 154. [Google Scholar] [CrossRef]
- Jensen, O.; Mazaheri, A. Shaping Functional Architecture by Oscillatory Alpha Activity: Gating by Inhibition. Front. Hum. Neurosci. 2010, 4, 186. [Google Scholar] [CrossRef]
- Nir, R.-R.; Sinai, A.; Moont, R.; Harari, E.; Yarnitsky, D. Tonic Pain and Continuous EEG: Prediction of Subjective Pain Perception by Alpha-1 Power during Stimulation and at Rest. Clin. Neurophysiol. 2012, 123, 605–612. [Google Scholar] [CrossRef]
- May, E.S.; Butz, M.; Kahlbrock, N.; Hoogenboom, N.; Brenner, M.; Schnitzler, A. Pre- and Post-Stimulus Alpha Activity Shows Differential Modulation with Spatial Attention during the Processing of Pain. Neuroimage 2012, 62, 1965–1974. [Google Scholar] [CrossRef]
- Huneke, N.T.M.; Brown, C.A.; Burford, E.; Watson, A.; Trujillo-Barreto, N.J.; El-Deredy, W.; Jones, A.K.P. Experimental Placebo Analgesia Changes Resting-State Alpha Oscillations. PLoS One 2013, 8, e78278. [Google Scholar] [CrossRef] [PubMed]
- Walton, K.D.; Dubois, M.; Llinás, R.R. Abnormal Thalamocortical Activity in Patients with Complex Regional Pain Syndrome (CRPS) Type I. Pain 2010, 150, 41–51. [Google Scholar] [CrossRef]
- Sarnthein, J.; Stern, J.; Aufenberg, C.; Rousson, V.; Jeanmonod, D. Increased EEG Power and Slowed Dominant Frequency in Patients with Neurogenic Pain. Brain 2006, 129, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Furman, A.J.; Thapa, T.; Summers, S.J.; Cavaleri, R.; Fogarty, J.S.; Steiner, G.Z.; Schabrun, S.M.; Seminowicz, D.A. Cerebral Peak Alpha Frequency Reflects Average Pain Severity in a Human Model of Sustained, Musculoskeletal Pain. J. Neurophysiol. 2019, 122, 1784–1793. [Google Scholar] [CrossRef] [PubMed]
- de Vries, M.; Wilder-Smith, O.H.; Jongsma, M.L.A.; van den Broeke, E.N.; Arns, M.; van Goor, H.; van Rijn, C.M. Altered Resting State EEG in Chronic Pancreatitis Patients: Toward a Marker for Chronic Pain. J. Pain Res. 2013, 6, 815–824. [Google Scholar] [CrossRef]
- Furman, A.J.; Meeker, T.J.; Rietschel, J.C.; Yoo, S.; Muthulingam, J.; Prokhorenko, M.; Keaser, M.L.; Goodman, R.N.; Mazaheri, A.; Seminowicz, D.A. Cerebral Peak Alpha Frequency Predicts Individual Differences in Pain Sensitivity. Neuroimage 2018, 167, 203–210. [Google Scholar] [CrossRef]
- Huber, F.A.; Kell, P.A.; Shadlow, J.O.; Rhudy, J.L. Cerebral Peak Alpha Frequency: Associations with Chronic Pain Onset and Pain Modulation. Neurobiol. Pain 2025, 18, 100180. [Google Scholar] [CrossRef]
- May, E.S.; Tiemann, L.; Gil Ávila, C.; Bott, F.S.; Hohn, V.D.; Gross, J.; Ploner, M. Assessing the Predictive Value of Peak Alpha Frequency for the Sensitivity to Pain. Pain 2025, 166, 2076–2090. [Google Scholar] [CrossRef] [PubMed]
- Bott, F.S.; Zebhauser, P.T.; Hohn, V.D.; Turgut, Ö.; May, E.S.; Tiemann, L.; Gil Ávila, C.; Heitmann, H.; Nickel, M.M.; Day, M.A.; et al. Exploring Electroencephalographic Chronic Pain Biomarkers: A Mega-Analysis. EBioMedicine 2025, 120, 105955. [Google Scholar] [CrossRef]
- Bott, F.S.; Nickel, M.M.; Hohn, V.D.; May, E.S.; Gil Ávila, C.; Tiemann, L.; Gross, J.; Ploner, M. Local Brain Oscillations and Interregional Connectivity Differentially Serve Sensory and Expectation Effects on Pain. Sci. Adv. 2023, 9, eadd7572. [Google Scholar] [CrossRef] [PubMed]
- Kenefati, G.; Rockholt, M.M.; Ok, D.; McCartin, M.; Zhang, Q.; Sun, G.; Maslinski, J.; Wang, A.; Chen, B.; Voigt, E.P.; et al. Changes in Alpha, Theta, and Gamma Oscillations in Distinct Cortical Areas Are Associated with Altered Acute Pain Responses in Chronic Low Back Pain Patients. Front. Neurosci. 2023, 17, 1278183. [Google Scholar] [CrossRef]
- Barbosa, S.P.; Junqueira, Y.N.; Akamatsu, M.A.; Marques, L.M.; Teixeira, A.; Lobo, M.; Mahmoud, M.H.; Omer, W.E.; Pacheco-Barrios, K.; Fregni, F. Resting-State Electroencephalography Delta and Theta Bands as Compensatory Oscillations in Chronic Neuropathic Pain: A Secondary Data Analysis. Brain Netw. Modul. 2024, 3, 52–60. [Google Scholar] [CrossRef]
- Shirvalkar, P.; Prosky, J.; Chin, G.; Ahmadipour, P.; Sani, O.G.; Desai, M.; Schmitgen, A.; Dawes, H.; Shanechi, M.M.; Starr, P.A.; et al. First-in-Human Prediction of Chronic Pain State Using Intracranial Neural Biomarkers. Nat. Neurosci. 2023, 26, 1090–1099. [Google Scholar] [CrossRef]
- Bikson, M.; Esmaeilpour, Z.; Adair, D.; Kronberg, G.; Tyler, W.J.; Antal, A.; Datta, A.; Sabel, B.A.; Nitsche, M.A.; Loo, C.; et al. Transcranial Electrical Stimulation Nomenclature. Brain Stimul 2019, 12, 1349–1366. [Google Scholar] [CrossRef] [PubMed]
- Helfrich, R.F.; Schneider, T.R.; Rach, S.; Trautmann-Lengsfeld, S.A.; Engel, A.K.; Herrmann, C.S. Entrainment of Brain Oscillations by Transcranial Alternating Current Stimulation. Curr. Biol. 2014, 24, 333–339. [Google Scholar] [CrossRef]
- Ali, M.M.; Sellers, K.K.; Fröhlich, F. Transcranial Alternating Current Stimulation Modulates Large-Scale Cortical Network Activity by Network Resonance. J. Neurosci. 2013, 33, 11262–11275. [Google Scholar] [CrossRef]
- Huang, W.A.; Stitt, I.M.; Negahbani, E.; Passey, D.J.; Ahn, S.; Davey, M.; Dannhauer, M.; Doan, T.T.; Hoover, A.C.; Peterchev, A.V.; et al. Transcranial Alternating Current Stimulation Entrains Alpha Oscillations by Preferential Phase Synchronization of Fast-Spiking Cortical Neurons to Stimulation Waveform. Nat. Commun. 2021, 12, 3151. [Google Scholar] [CrossRef] [PubMed]
- Vossen, A.; Gross, J.; Thut, G. Alpha Power Increase after Transcranial Alternating Current Stimulation at Alpha Frequency (α-tACS) Reflects Plastic Changes rather than Entrainment. Brain Stimul. 2015, 8, 499–508. [Google Scholar] [CrossRef]
- Riddle, J.; Frohlich, F. Targeting Neural Oscillations with Transcranial Alternating Current Stimulation. Brain Res. 2021, 1765, 147491. [Google Scholar] [CrossRef] [PubMed]
- Fathi, Y.; Dissassuca, F.; Ricci, G.; Liberati, G. Individualised Alpha-tACS for Modulating Pain Perception and Neural Oscillations: A Sham-Controlled Study in Healthy Participants. Eur. J. Pain 2025, 29, e70086. [Google Scholar] [CrossRef]
- Arendsen, L.J.; Hugh-Jones, S.; Lloyd, D.M. Transcranial Alternating Current Stimulation at Alpha Frequency Reduces Pain When the Intensity of Pain Is Uncertain. J. Pain 2018, 19, 807–818. [Google Scholar] [CrossRef]
- Ikarashi, H.; Otsuru, N.; Gomez-Tames, J.; Hirata, A.; Nagasaka, K.; Miyaguchi, S.; Sakurai, N.; Ohno, K.; Kodama, N.; Onishi, H. Modulation of Pain Perception through Transcranial Alternating Current Stimulation and Its Nonlinear Relationship with the Simulated Electric Field Magnitude. Eur. J. Pain 2024, 28, 1018–1028. [Google Scholar] [CrossRef]
- Li, X.; Jin, R.; Lu, X.; Zhan, Y.; Jiang, N.; Peng, W. Alpha Transcranial Alternating Current Stimulation Modulates Pain Anticipation and Perception in a Context-Dependent Manner. Pain 2025, 166, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- May, E.S.; Hohn, V.D.; Nickel, M.M.; Tiemann, L.; Gil Ávila, C.; Heitmann, H.; Sauseng, P.; Ploner, M. Modulating Brain Rhythms of Pain Using Transcranial Alternating Current Stimulation (tACS) - A Sham-Controlled Study in Healthy Human Participants. J. Pain 2021, 22, 1256–1272. [Google Scholar] [CrossRef]
- Peng, W.; Zhan, Y.; Jin, R.; Lou, W.; Li, X. Aftereffects of Alpha Transcranial Alternating Current Stimulation over the Primary Sensorimotor Cortex on Cortical Processing of Pain. Pain 2023, 164, 1280–1290. [Google Scholar] [CrossRef]
- Qi, X.; Jia, T.; Sun, B.; Xia, J.; Wang, C.; Hong, Z.; Zhang, Y.; Yang, H.; Zhang, C.; Liu, J. Individual Differences in Resting Alpha Band Power and Changes in Theta Band Power during Sustained Pain Are Correlated with the Pain-Relieving Efficacy of Alpha HD-tACS on SM1. Neuroimage 2025, 312, 121237. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Zhang, C.; Zhang, Q.; Xu, X.; Liu, J.; Yang, H. Analgesic Aftereffects of Alpha High-Definition Transcranial Alternating Current Stimulation over the DLPFC during Experimental Pain. Neuroimage 2025, 317, 121332. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, N.; Terui, Y. Synchronal Dual Brain Stimulation over the Somatosensory Cortex Modulated Social Touch-Induced Analgesia Depending on Empathy. J. Pain 2025, 105483. [Google Scholar] [CrossRef]
- Ahn, S.; Prim, J.H.; Alexander, M.L.; McCulloch, K.L.; Fröhlich, F. Identifying and Engaging Neuronal Oscillations by Transcranial Alternating Current Stimulation in Patients with Chronic Low Back Pain: A Randomized, Crossover, Double-Blind, Sham-Controlled Pilot Study. J. Pain 2019, 20, 277.e1–e277.e11. [Google Scholar] [CrossRef]
- Antal, A.; Bischoff, R.; Stephani, C.; Czesnik, D.; Klinker, F.; Timäus, C.; Chaieb, L.; Paulus, W. Low Intensity, Transcranial, Alternating Current Stimulation Reduces Migraine Attack Burden in a Home Application Set-up: A Double-Blinded, Randomized Feasibility Study. Brain Sci. 2020, 10, 888. [Google Scholar] [CrossRef]
- Bernardi, L.; Bertuccelli, M.; Formaggio, E.; Rubega, M.; Bosco, G.; Tenconi, E.; Cattelan, M.; Masiero, S.; Del Felice, A. Beyond Physiotherapy and Pharmacological Treatment for Fibromyalgia Syndrome: Tailored tACS as a New Therapeutic Tool. Eur. Arch. Psychiatry Clin. Neurosci. 2021, 271, 199–210. [Google Scholar] [CrossRef]
- Lin, A.P.; Chiu, C.-C.; Chen, S.-C.; Huang, Y.-J.; Lai, C.-H.; Kang, J.-H. Using High-Definition Transcranial Alternating Current Stimulation to Treat Patients with Fibromyalgia: A Randomized Double-Blinded Controlled Study. Life (Basel) 2022, 12, 1364. [Google Scholar] [CrossRef]
- Prim, J.H.; Ahn, S.; Davila, M.I.; Alexander, M.L.; McCulloch, K.L.; Fröhlich, F. Targeting the Autonomic Nervous System Balance in Patients with Chronic Low Back Pain Using Transcranial Alternating Current Stimulation: A Randomized, Crossover, Double-Blind, Placebo-Controlled Pilot Study. J. Pain Res. 2019, 12, 3265–3277. [Google Scholar] [CrossRef] [PubMed]
- Hashmi, J.A.; Baliki, M.N.; Huang, L.; Baria, A.T.; Torbey, S.; Hermann, K.M.; Schnitzer, T.J.; Apkarian, A.V. Shape Shifting Pain: Chronification of Back Pain Shifts Brain Representation from Nociceptive to Emotional Circuits. Brain 2013, 136, 2751–2768. [Google Scholar] [CrossRef] [PubMed]
- Baliki, M.N.; Petre, B.; Torbey, S.; Herrmann, K.M.; Huang, L.; Schnitzer, T.J.; Fields, H.L.; Apkarian, A.V. Corticostriatal Functional Connectivity Predicts Transition to Chronic Back Pain. Nat. Neurosci. 2012, 15, 1117–1119. [Google Scholar] [CrossRef] [PubMed]
- Vachon-Presseau, E.; Centeno, M.V.; Ren, W.; Berger, S.E.; Tétreault, P.; Ghantous, M.; Baria, A.; Farmer, M.; Baliki, M.N.; Schnitzer, T.J.; et al. The Emotional Brain as a Predictor and Amplifier of Chronic Pain. J. Dent. Res. 2016, 95, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Baliki, M.N.; Mansour, A.R.; Baria, A.T.; Apkarian, A.V. Functional Reorganization of the Default Mode Network across Chronic Pain Conditions. PLoS One 2014, 9, e106133. [Google Scholar] [CrossRef]
- Kaplan, C.M.; Kelleher, E.; Irani, A.; Schrepf, A.; Clauw, D.J.; Harte, S.E. Deciphering Nociplastic Pain: Clinical Features, Risk Factors and Potential Mechanisms. Nat. Rev. Neurol. 2024, 20, 347–363. [Google Scholar] [CrossRef]
- Napadow, V.; LaCount, L.; Park, K.; As-Sanie, S.; Clauw, D.J.; Harris, R.E. Intrinsic Brain Connectivity in Fibromyalgia Is Associated with Chronic Pain Intensity. Arthritis Rheum. 2010, 62, 2545–2555. [Google Scholar] [CrossRef]
- Ellingsen, D.-M.; Beissner, F.; Moher Alsady, T.; Lazaridou, A.; Paschali, M.; Berry, M.; Isaro, L.; Grahl, A.; Lee, J.; Wasan, A.D.; et al. A Picture Is Worth a Thousand Words: Linking Fibromyalgia Pain Widespreadness from Digital Pain Drawings with Pain Catastrophizing and Brain Cross-Network Connectivity: Linking Fibromyalgia Pain Widespreadness from Digital Pain Drawings with Pain Catastrophizing and Brain Cross-Network Connectivity. Pain 2021, 162, 1352–1363. [Google Scholar]
- Kuner, R.; Flor, H. Structural Plasticity and Reorganisation in Chronic Pain. Nat. Rev. Neurosci. 2016, 18, 20–30. [Google Scholar] [CrossRef]
- Babiloni, C.; Brancucci, A.; Del Percio, C.; Capotosto, P.; Arendt-Nielsen, L.; Chen, A.C.N.; Rossini, P.M. Anticipatory Electroencephalography Alpha Rhythm Predicts Subjective Perception of Pain Intensity. J. Pain 2006, 7, 709–717. [Google Scholar] [CrossRef]
- Tu, Y.; Zhang, Z.; Tan, A.; Peng, W.; Hung, Y.S.; Moayedi, M.; Iannetti, G.D.; Hu, L. Alpha and Gamma Oscillation Amplitudes Synergistically Predict the Perception of Forthcoming Nociceptive Stimuli. Hum. Brain Mapp. 2016, 37, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Hohn, V.D.; May, E.S.; Ploner, M. From Correlation towards Causality: Modulating Brain Rhythms of Pain Using Transcranial Alternating Current Stimulation. Pain Rep. 2019, 4, e723. [Google Scholar] [CrossRef]
- Chang, M.C.; Briand, M.-M.; Boudier-Revéret, M.; Yang, S. Effectiveness of Transcranial Alternating Current Stimulation for Controlling Chronic Pain: A Systematic Review. Front. Neurol. 2023, 14, 1323520. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhu, M.; Mo, S.; Li, X.; Wang, J. The Effect of Somatosensory Alpha Transcranial Alternating Current Stimulation on Pain Empathy Is Trait Empathy and Gender Dependent. CNS Neurosci. Ther. 2021, 27, 687–693. [Google Scholar] [CrossRef]
- Takeuchi, N. Pain Control Based on Oscillatory Brain Activity Using Transcranial Alternating Current Stimulation: An Integrative Review. Front. Hum. Neurosci. 2023, 17, 941979. [Google Scholar] [CrossRef]
- Witkowski, M.; Garcia-Cossio, E.; Chander, B.S.; Braun, C.; Birbaumer, N.; Robinson, S.E.; Soekadar, S.R. Mapping Entrained Brain Oscillations during Transcranial Alternating Current Stimulation (tACS). Neuroimage 2016, 140, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Haslacher, D.; Narang, A.; Sokoliuk, R.; Cavallo, A.; Reber, P.; Nasr, K.; Santarnecchi, E.; Soekadar, S.R. In Vivo Phase-Dependent Enhancement and Suppression of Human Brain Oscillations by Transcranial Alternating Current Stimulation (tACS). Neuroimage 2023, 275, 120187. [Google Scholar] [CrossRef]
- Haslacher, D.; Cavallo, A.; Reber, P.; Kattein, A.; Thiele, M.; Nasr, K.; Hashemi, K.; Sokoliuk, R.; Thut, G.; Soekadar, S.R. Working Memory Enhancement Using Real-Time Phase-Tuned Transcranial Alternating Current Stimulation. Brain Stimul. 2024, 17, 850–859. [Google Scholar] [CrossRef]
- Haslacher, D.; Nasr, K.; Robinson, S.E.; Braun, C.; Soekadar, S.R. Stimulation Artifact Source Separation (SASS) for Assessing Electric Brain Oscillations during Transcranial Alternating Current Stimulation (tACS). Neuroimage 2021, 228, 117571. [Google Scholar] [CrossRef]
- Thiele, C.; Zaehle, T.; Haghikia, A.; Ruhnau, P. Amplitude Modulated Transcranial Alternating Current Stimulation (AM-TACS) Efficacy Evaluation via Phosphene Induction. Sci. Rep. 2021, 11, 22245. [Google Scholar] [CrossRef]
- Krause, M.R.; Vieira, P.G.; Thivierge, J.-P.; Pack, C.C. Brain Stimulation Competes with Ongoing Oscillations for Control of Spike Timing in the Primate Brain. PLoS Biol. 2022, 20, e3001650. [Google Scholar] [CrossRef] [PubMed]
- Fiene, M.; Schwab, B.C.; Misselhorn, J.; Herrmann, C.S.; Schneider, T.R.; Engel, A.K. Phase-Specific Manipulation of Rhythmic Brain Activity by Transcranial Alternating Current Stimulation. Brain Stimul. 2020, 13, 1254–1262. [Google Scholar] [CrossRef]
- Schreglmann, S.R.; Wang, D.; Peach, R.L.; Li, J.; Zhang, X.; Latorre, A.; Rhodes, E.; Panella, E.; Cassara, A.M.; Boyden, E.S.; et al. Non-Invasive Suppression of Essential Tremor via Phase-Locked Disruption of Its Temporal Coherence. Nat. Commun. 2021, 12, 363. [Google Scholar] [CrossRef] [PubMed]
- Alekseichuk, I.; Falchier, A.Y.; Linn, G.; Xu, T.; Milham, M.P.; Schroeder, C.E.; Opitz, A. Electric Field Dynamics in the Brain during Multi-Electrode Transcranial Electric Stimulation. Nat. Commun. 2019, 10, 2573. [Google Scholar] [CrossRef] [PubMed]
- Aksenov, A.; Renaud-D’Ambra, M.; Volpert, V.; Beuter, A. Phase-Shifted tACS Can Modulate Cortical Alpha Waves in Human Subjects. Cogn. Neurodyn. 2024, 18, 1575–1592. [Google Scholar] [CrossRef]
- Wei, J.; Alamia, A.; Yao, Z.; Huang, G.; Li, L.; Liang, Z.; Zhang, L.; Zhou, C.; Song, Z.; Zhang, Z. State-Dependent tACS Effects Reveal the Potential Causal Role of Prestimulus Alpha Traveling Waves in Visual Contrast Detection. J. Neurosci. 2024, 44, e2023232024. [Google Scholar] [CrossRef]
- Isnard, J.; Magnin, M.; Jung, J.; Mauguière, F.; Garcia-Larrea, L. Does the Insula Tell Our Brain That We Are in Pain? Pain 2011, 152, 946–951. [Google Scholar] [CrossRef] [PubMed]
- McBenedict, B.; Petrus, D.; Pires, M.P.; Pogodina, A.; Arrey Agbor, D.B.; Ahmed, Y.A.; Castro Ceron, J.I.; Balaji, A.; Abrahão, A.; Lima Pessôa, B. The Role of the Insula in Chronic Pain and Associated Structural Changes: An Integrative Review. Cureus 2024, 16, e58511. [Google Scholar] [CrossRef]
- Kurth, F.; Zilles, K.; Fox, P.T.; Laird, A.R.; Eickhoff, S.B. A Link between the Systems: Functional Differentiation and Integration within the Human Insula Revealed by Meta-Analysis. Brain Struct. Funct. 2010, 214, 519–534. [Google Scholar] [CrossRef]
- Brooks, J.C.W.; Tracey, I. The Insula: A Multidimensional Integration Site for Pain. Pain 2007, 128, 1–2. [Google Scholar] [CrossRef]
- Bastuji, H.; Frot, M.; Perchet, C.; Hagiwara, K.; Garcia-Larrea, L. Convergence of Sensory and Limbic Noxious Input into the Anterior Insula and the Emergence of Pain from Nociception. Sci. Rep. 2018, 8, 13360. [Google Scholar] [CrossRef] [PubMed]
- Berthier, M.; Starkstein, S.; Leiguarda, R. Asymbolia for Pain: A Sensory-Limbic Disconnection Syndrome. Ann. Neurol. 1988, 24, 41–49. [Google Scholar] [CrossRef]
- Liberati, G.; Algoet, M.; Santos, S.F.; Ribeiro-Vaz, J.G.; Raftopoulos, C.; Mouraux, A. Tonic Thermonociceptive Stimulation Selectively Modulates Ongoing Neural Oscillations in the Human Posterior Insula: Evidence from Intracerebral EEG. Neuroimage 2019, 188, 70–83. [Google Scholar] [CrossRef]
- Kim, K.; Nan, G.; Bak, H.; Kim, H.Y.; Kim, J.; Cha, M.; Lee, B.H. Insular Cortex Stimulation Alleviates Neuropathic Pain through Changes in the Expression of Collapsin Response Mediator Protein 2 Involved in Synaptic Plasticity. Neurobiol. Dis. 2024, 194, 106466. [Google Scholar] [CrossRef]
- Liu, C.-C.; Moosa, S.; Quigg, M.; Elias, W.J. Anterior Insula Stimulation Increases Pain Threshold in Humans: A Pilot Study. J. Neurosurg. 2021, 135, 1487–1492. [Google Scholar] [CrossRef] [PubMed]
- Grossman, N.; Bono, D.; Dedic, N.; Kodandaramaiah, S.B.; Rudenko, A.; Suk, H.-J.; Cassara, A.M.; Neufeld, E.; Kuster, N.; Tsai, L.-H.; et al. Noninvasive Deep Brain Stimulation via Temporally Interfering Electric Fields. Cell 2017, 169, 1029–1041.e16. [Google Scholar] [CrossRef] [PubMed]
- Mansourinezhad, P.; Mestrom, R.M.C.; Klooster, D.C.W.; Sprengers, M.; Boon, P.A.J.M.; Paulides, M.M. Systematic Review of Experimental Studies in Humans on Transcranial Temporal Interference Stimulation. J. Neural Eng. 2025, 22, 051001. [Google Scholar] [CrossRef]
- Barzegar, S.; Kakies, C.F.M.; Ciupercӑ, D.; Wischnewski, M. Transcranial Alternating Current Stimulation for Investigating Complex Oscillatory Dynamics and Interactions. Int. J. Psychophysiol. 2025, 212, 112579. [Google Scholar] [CrossRef] [PubMed]
- Alekseichuk, I.; Turi, Z.; Amador de Lara, G.; Antal, A.; Paulus, W. Spatial Working Memory in Humans Depends on Theta and High Gamma Synchronization in the Prefrontal Cortex. Curr. Biol. 2016, 26, 1513–1521. [Google Scholar] [CrossRef]
Figure 1.
AM-tACS delivers a high-frequency carrier whose amplitude is modulated at a lower envelope frequency. During EEG recording, stimulation artifacts mainly cluster around the carrier frequency. Because of this spectral separation, filtering can attenuate the carrier-related artifacts while preserving the low-frequency envelope content. I1 = injected current.
Figure 1.
AM-tACS delivers a high-frequency carrier whose amplitude is modulated at a lower envelope frequency. During EEG recording, stimulation artifacts mainly cluster around the carrier frequency. Because of this spectral separation, filtering can attenuate the carrier-related artifacts while preserving the low-frequency envelope content. I1 = injected current.
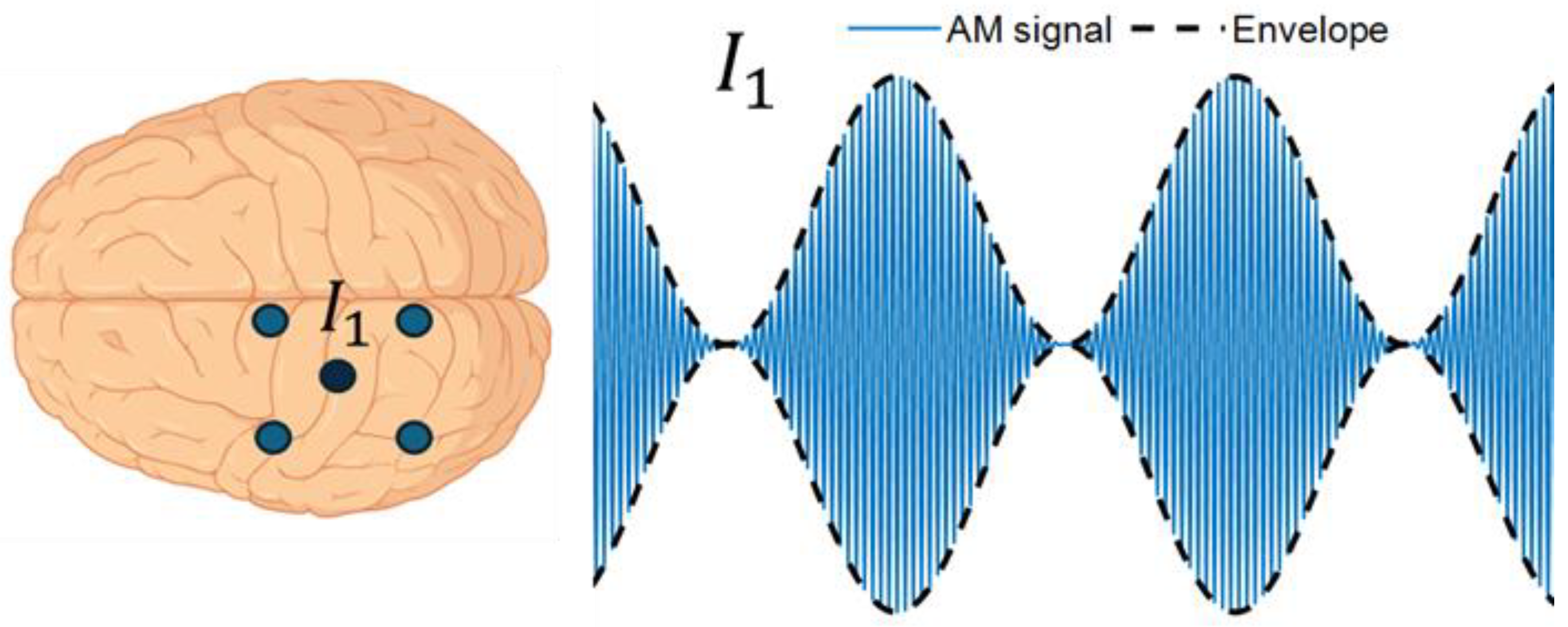
Figure 2.
TIS applies two high-frequency alternating currents with slightly different frequencies. Although each carrier frequency is too high to elicit neural firing, their interaction generates a low-frequency envelope capable of modulating neural activity selectively at a targeted focus, minimizing effects on adjacent or superficial regions and reaching deeper targets compared to conventional non-invasive brain stimulation.
Figure 2.
TIS applies two high-frequency alternating currents with slightly different frequencies. Although each carrier frequency is too high to elicit neural firing, their interaction generates a low-frequency envelope capable of modulating neural activity selectively at a targeted focus, minimizing effects on adjacent or superficial regions and reaching deeper targets compared to conventional non-invasive brain stimulation.
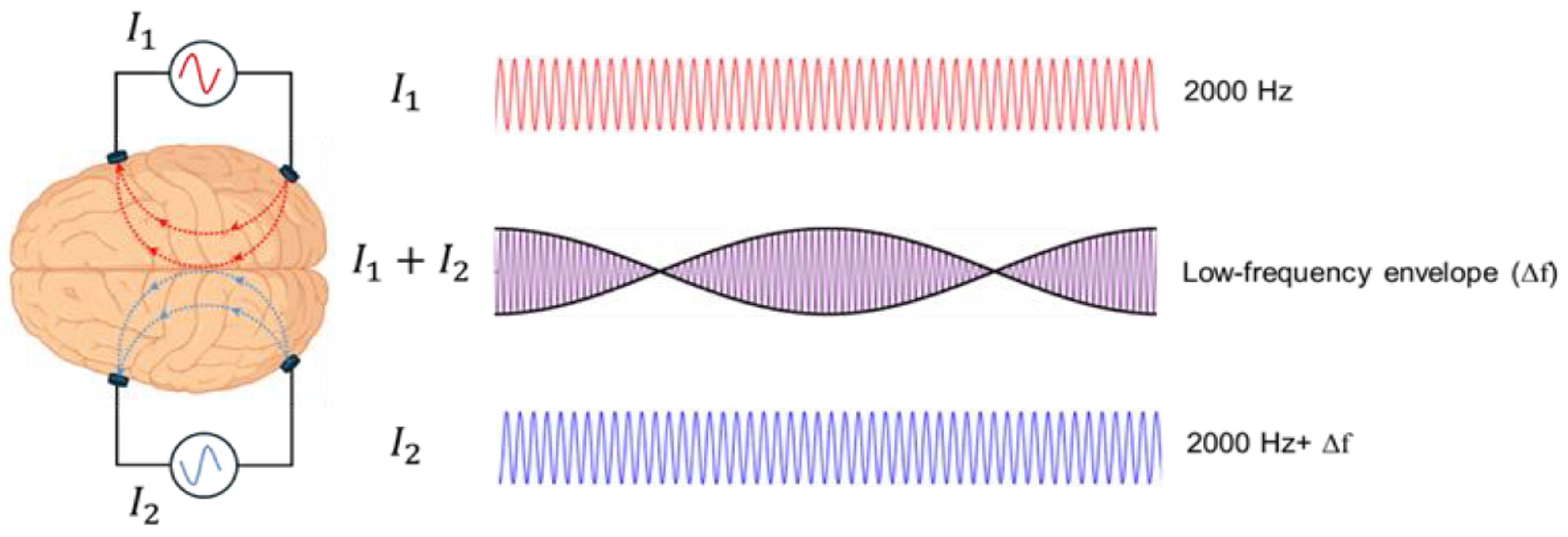

Table 1.
Summary of key characteristics of tACS studies on pain.
| Study | Participant characteristics | Study design | Stimulation montage and site | tACS parameters | Control | Outcomes | Main findings |
|---|---|---|---|---|---|---|---|
| Experimental Pain tACS Studies | |||||||
| Arendsen et al., 2018 [27] | 23 healthy, right-handed (22 female; mean age = 21.4 ± 4.7) | Within-subject, randomized crossover study | Bilateral SM1 (CP3/CP4, 5×5 cm pads) | 10 Hz, 1 mA pp, ~15–20 min (applied during pressure pain task) | Sham (tRNS 30 s ramp-up/hold/down) | Pain intensity and unpleasantness ratings (NRS) during pressure pain to the middle finger. Participants also performed a visual cue-pain task, where cues predicted the timing of pain stimuli. | Decreased pain intensity and unpleasantness during active tACS vs. sham, only when impending pain intensity was uncertain; no correlations with fear of pain or catastrophizing. |
| Fathi et al., 2025 [26] | 38 healthy participants (19 female; mean age = 28 ± 8.5) | within-subject, randomized crossover, double-blind | Contralateral SM (4×1 montage centered on C3/C4) | Individual peak alpha frequency, 2mA pp, 20 min |
Sham (1 min with ramp-up/down) | Heat pain thresholds and ratings + EEG | n.s.; trend toward reduced sensitization in women with active tACS; correlation was observed between SS-IAF and HPT during sham condition. |
| Ikarashi et al., 2024 [28] | 56 healthy, right-handed (27 female; mean age = 21.2 ± 1.0) | Between-subject, randomized, single-blind | Left DLPFC (F3–Fpz, 3×3 cm electrodes) | 6 Hz (θ) or 20 Hz (β), 1 mA, 20 min | Sham (1 min with ramp-up/down) |
Heat pain threshold (HPT) and tolerance (HPTT) using Peltier thermode (right forearm, 35 °C baseline, +0.7 °C/s); repeated measures before, during, after stimulation; | Pain reduction vs sham; θ- and β-tACS ↑ HPT during and after stimulation , no effect on HPTT; θ-tACS pain reduction followed an inverted U-shaped relation with simulated E-field in posterior DLPFC, suggesting optimal field strength for maximal pain relief |
| Li et al., 2025 [29] | 80 healthy participants (41 female; mean age = 21.8 ± 2.12) | Between-subjects, double-blinded | Right SM (4x1 montage centered on C4 and the 4 surrounding return electrodes were placed at FC2, FC6, CP6, and CP2) |
10 Hz, 20 min, 1 mA | Sham (1 min with ramp-up/down) | Ratings of pain intensity & unpleasantness in response to noxious laser stimulation, Visual cues manipulated certainty + EEG | n.s.: α-tACS produced no significant pain reduction effect on pain ratings. α-tACS disrupted normal LEP habituation, especially under certain pain expectation, with effects lasting ~30 min. It increased contralateral α₁ and midfrontal θ activity during anticipation; θ changes persisted. Mediation suggested opposing α-driven analgesic and θ-driven hyperalgesic pathways, with the latter dominating—resulting in no pain reduction. |
| May et al., 2021 [30] | 29 healthy, right-handed (13 female; mean age = 25.7 ± 4.0) | Within-subject, randomized crossover, double-blinded | PFC (F3/F4) or S1 (CP3/CP4), large pads 5×5 cm | 10 Hz (α) or 80 Hz (γ), 10 min, 1 mA pp | Sham (30 s 10 Hz with ramps) | Tonic heat pain (VAS, continuous ratings), autonomic measures (skin conductance, ECG), EEG (pre/post, 5 min) | n.s. for all conditions; no modulation of pain, autonomic responses, or oscillations. Bayesian analysis supported null, except α-S1 (inconclusive). |
| Peng et al., 2023 [31] | 53 healthy participants (26 active, 27 sham; 32 female; mean = age ~21) | Between-subject, double-blinded | Left or right SM1 (C3/C4-centered 4×1 HD montage) | 10 Hz, 20 min, 1 mA peak-to-peak | Sham (1 min with ramp-up/down) |
fMRI during noxious laser stimulation (pain-evoked activity, connectivity), pain ratings (intensity, unpleasantness, NRS) | n.s.; Active α-tACS attenuated pain-evoked activity in bilateral SM1 and left M1 vs sham. Mediation: reduced SM1 activity indirectly decreased pain ratings. Functional connectivity between SM1 and DLPFC, S1, MCC, SMA decreased with active vs sham. |
| Qi et al., 2025 [32] | 31 healthy participants (14 female; mean age = 23 ± 2.3; final after artifact exclusion) | Within-subject, randomized crossover, double-blinded | Contralateral SM1 (4×1 HD montage centered on C4) | 10 Hz, 30 min, 1.5 mA pp | Sham (1 min with ramp-up/down) | Capsaicin-induced pain (NRS), rest EEG power | Pain reduction vs sham. Resting α and pain-related θ changes at C4 predicted efficacy (α → θ → pain relief). |
| Sun et al., 2025 [33] | 45 healthy (28 female; mean age = 21 ± 2.4) | Within-subject, randomized crossover, single-blinded | ipsilateral DLPFC (F3-centered, 3×1 HD montage) | 10 Hz, 30 min, 1.5 mA pp | Sham (1 min with ramp-up/down) | Capsaicin-induced pain (NRS), rest EEG power | Pain reduction vs sham; Delayed increase of low γ at DLPFC; weak correlation with pain reduction. |
| Takeuchi & Terui, 2025 [34] | 32 healthy participants, 16 pairs (18 females, 14 males; mean age = 21.0±1.4) | Within-subject, randomized crossover | Right S1 (2 cm posterior to C4, concentric 1X1 tACS) | 10 Hz, peak-to-peak 3 mA, 5 min | Sham (1 min with 30 sec ramp-up/down) | Electrical pain intensity ratings (NRS) and empathy scores from the IRI (Interpersonal Reactivity Index). Participant pairs alternated as pain-receiver and touch-giver while receiving mild electrical pain and gentle brush stroking on the forearm to examine the effects of tACS on touch-induced analgesia. |
Though overall pain ratings did not differ across conditions, synchronous dual-brain “hyper”-tACS enhanced social touch–induced pain reduction in low-empathy receivers vs. sham. |
| Chronic Pain tACS Studies | |||||||
| Ahn et al., 2019 [35] | 20 patients with chronic low back pain (8 male, 12 female; mean pain duration ≈ 85 months). | Within-subject, randomized crossover, double-blind, sham-controlled | Bifrontal (F3/F4, 5×5 cm; return 5×7 cm at Pz) | 10 Hz, 1 mA per F3/F4 (2 mA return at Pz), 40 min | Sham (1 min with ramp-up/down) | Chronic low back pain ratings (DVPRS, ODI), rest EEG (α-power pre/post, correlation with pain relief) | Pain reduction vs sham; 10 Hz -tACS increased somatosensory α-power, and α enhancement correlated with ↓ chronic pain ratings (DVPRS) and perceived disability (ODI); exploratory Wilcoxon test showed significant ↓ DVPRS , ODI change n.s. |
| Antal et al., (2020) [36] | 25 migraine patients (Active tACS: N = 16, age = 31.1±8.9; Sham: N = 9, age = 28.1±10.5) | Between-subject, randomized parallel groups, double-blinded | Occipital cortex (Oz, 4×4 cm) with return at Cz (5×7 cm) | 140 Hz, 0.4 mA, 15 min per session up to 5 sessions total over a six week period. Patients were instructed to use tACS during onset of migraine attacks. | Sham (simulator turned off after 30 sec) | patients recorded termination of migraine attacks within 2 hours post-stimulation (defined as NAS<1), NAS pain intensity (0-10) before/after migraine attacks, analgesic medication use, and attack recurrence. |
During migraines without use of analgesic medication, a significantly greater number of migraine attacks terminated following active tACS compared to sham. Significant ↓ NAS pain severity for active tACS compared to sham at 2-4 hours post-stimulation. |
| Bernardi et al., (2021) [37] | 15 individuals diagnosed with fibromyalgia syndrome | Within-subject, randomized crossover,double blind 10 sessions (30 min/day, 5 days/week × 2 weeks) + physiotherapy (60 min/session) with 4-week washout between conditions |
Anode positioned individually over scalp region with greatest EEG abnormality (e.g., F3, C3, CP5, C4/Cp2, or similar); cathode over ipsilateral mastoid. large sponge pads (5 × 7 cm) |
1–2 mA alternating current Participants with increased slow rhythms (theta, delta, α₁) received β-tACS at 30 Hz Participants with increased fast rhythms (β, α₂) received θ-tACS at 4 Hz |
Random noise stimulation with random amplitude and frequency, respectively, in the intervals (1–2) mA and (0–100) Hz |
Resting EEG (α₁, θ, δ, β power); Visual Analog Scale (VAS) for pain; neuropsychological and self-report assessments (MoCA, TMT-A/B, HVLT-R, Rey Complex Figure, PROCOG-P, EMQ-R, BDI-II, BSI, STAI); SF-36 for health status. | Pain reduction vs sham; tailored tACS (4 Hz or 30 Hz, 1–2 mA) increased resting α₁ power and reduced pain (VAS ↓ at T1) vs random noise stimulation (RNS). Improved cognitive and neuropsychological performance (MoCA, TMT-B, PROCOG-P, EMQ-R) and reduced depressive symptoms (BDI-II, BSI). EEG normalization mainly over sensorimotor/M1 areas; pain relief effects faded by 4 weeks |
| Lin et al., 2022 [38] | 38 patients with fibromyalgia (30F; mean age 48.6 ± 12.9); 35 completed (active = 18, sham = 17) | Between-subject, randomized, double-blind, sham-controlled | Left M1 (anode = C3; 4 cathodes = Cz, F3, T7, P3; radius ≈ 7.5 cm, 4×1 HD montage) | 50 Hz, 20 min/session, 10 sessions over 2 weeks | Sham (10 s ramp-up, 19 min 40 s no current, 10 s ramp-down) | NRS (pain intensity), FIQ (quality of life), BAI (anxiety), BDI-II (depression), PSQI (sleep quality), PPT (pressure pain threshold), plasma Tau and Aβ₁₋₄₂ | Both groups showed within-group symptom improvement; no significant differences between active and sham in NRS, FIQ, or secondary measures. Active HD-tACS significantly reduced FIQ vs baseline but not vs sham. One suicide attempt occurred (likely unrelated). HD-tACS was otherwise well tolerated. |
| Prim et al., 2019 [39] | 20 patients with chronic low back pain (8M, 12F; mean age 43 ± 13; mean pain duration ≈7 years) | Within-subject, randomized, crossover, double-blind, sham-controlled | Bilateral prefrontal cortex (F3/F4, 5×5 cm) with return at Pz (5×7 cm) | 10 Hz, 1 mA (zero-to-peak), 40 min; in-phase stimulation; ≥7 days washout | Sham (1 min 10 Hz with ramp-up/down) | ECG for HRV (RSA, LF, LF/HF, SDNN, RMSSD); pain (DVPRS), disability (ODI), and pressure pain threshold (PPT) pre/post | No significant change in RSA or LF/HF. SDNN increased after 10 Hz-tACS vs sham, indicating modulation of overall autonomic balance. Pain ratings and HRV changes were uncorrelated. Twice as many participants were clinical responders (≥2 pt pain reduction) in active vs sham, though not statistically significant. Blinding was successful and no adverse effects were reported. |
| * n.s. = no significant analgesic effect on pain ratings. | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



