
Submitted:
29 December 2025
Posted:
30 December 2025
You are already at the latest version
Abstract
Damage to the chorda tympani (CT) nerve through trauma or experimental nerve axotomy results in the degeneration of anterior taste buds and taste loss. Our previous work demonstrated that Il1r signaling is required for taste bud regeneration and the recovery of taste function. However, the effects of experimental axotomy on immune responses in the absence of Il1r signaling remain unclear. To test this, we performed unilateral CT sectioning in Il1r KO or wild-type mice as previously described. We found that CD45+ immune cells, CD68+ and CD206+ M2-like macrophages are significantly increased near anterior taste buds at day two post-injury in wild-type but not Il1r KO mice. By day 5, these macrophage responses were slightly elevated in wild-type mice but remained at baseline levels in KO mice, indicating that immune responses to injury were suppressed rather than delayed in the absence of Il1r signaling. However, taste buds degenerated at similar time points in both strains. These results suggest that delayed taste bud degeneration in Il1r KO mice is not the primary reason for later functional deficits, though suppressed immune response may have other consequences in the injured peripheral taste system.
Keywords:
chorda tympani nerve
; axotomy
; degeneration
; Interleukin-1
; gustatory
; macrophages
; regeneration
1. Introduction
Taste buds are heterogenous chemosensory organs that transmit information about chemical stimuli from the oral cavity to the gustatory cortex. Similar to epithelial cells, taste bud cells are not permanent and have a turnover rate of 7-30 days with each taste bud containing 40-100 specialized taste cells. Taste buds are composed of four main taste receptor cell types. Type I cells are “glial-like” with processes that encompass other cell types in the taste bud. Type II and type III cells are directly responsible for the transduction of taste information, with type II responding to bitter, sweet or umami stimuli and type III responding to sour and salty tastants. Post-mitotic type IV cells at the base of the taste bud differentiate into the other three cell types [4,5].
Taste buds are innervated by and receive trophic support from sensory afferent nerve fibers. Anterior taste buds in fungiform papillae are innervated ipsilaterally by one of two chorda tympani (CT) nerves on each side of the tongue. Unlike neurons in the central nervous system, peripheral gustatory axons have the ability to regenerate after injury. In rodents, experimental axotomy of the chorda tympani nerve is followed by degeneration of the distal axons associated with taste buds. However, within four weeks in mice, CT fibers and taste buds regenerate and normal responses to tastants are restored [6,7,8]. The sense of taste is vulnerable to damage by chemotherapeutic treatment, infection, trauma, or through direct damage to the CT nerve during surgical or dental procedures [9,10,11,12,13,14]. Lasting deficits in taste perception remain in some patients, highlighting the need for a better understanding of mechanisms mediating the recovery of gustatory function [15]. The plasticity of the peripheral taste system and bilateral innervation of anterior taste buds make rodent CT injury model ideal for elucidating mechanisms of recovery [14].
Gustatory nerve injury and taste bud degeneration elicits immune responses and activation of inflammatory pathways [16,17,18,19,20,21]. After CT axotomy in rodents there is a rapid influx of neutrophils in the degenerating taste bud field within 6 hours followed by macrophage responses over the next 48 hours [22,23,24,25,26,27]. Nerve injury also upregulates chemokine, adhesion molecules, and cytokine responses in the anterior taste field [16,17,18,19,20,21]. The IL-1 receptor (IL-1R)1 and ligands IL-1α and IL-1β are expressed in taste buds, axons, and immune cells responding to axotomy [20,28]. IL-1α and IL-1β bind to the IL-1R to trigger intracellular signaling and activate inflammatory responses. In the absence of Il1r, these responses are typically blunted in other tissues especially in the context of infection or injury [29,30,31,32]. However, lingual mRNA levels of the proinflammatory cytokine, Il1b, are upregulated at day 56-60 in Il1r KO compared to wild-type, C57BL/6J mice, which could induce inflammation at this late period [33]. This dysregulation, combined with the pleiotropism and redundancy of cytokine pathways, highlights the need to investigate immune responses in the taste bud target field after CT nerve injury.
Il1r KO mice receiving CT axotomy exhibit altered cytokine response dynamics in the injured taste bud field, dysregulated taste progenitor cell proliferation, and incomplete taste bud regeneration. These changes result in absent or minimal neural responses to taste stimuli even at >14 weeks post-injury, compared to the recovery of normal taste function at 4 weeks in wild-type mice [6]. Together, these results support the requirement for IL-1R signaling in taste bud regeneration and functional recovery. Here, we hypothesize that immune responses to CT axotomy are altered in the absence of IL-1R signaling. We performed surgical CT axotomy on wild-type (wild-type) and Il1r KO mice as in our previous work [28] then measured innate immune cell populations at different time points post-injury. We also tested whether dysregulated immune responses in Il1r KO mice delay taste bud degeneration, potentially preventing later, efficient taste cell regeneration. Identifying the dynamics of specific immune responses will potentially allow for more targeted therapeutic treatments in the restoration of gustatory function after nerve injury, chemotherapy, or infection.
2. Results
2.1. CD45+ Immune Cells Rapidly Infiltrate Axotomized Taste Papillae in Wild-Type But not Il1r KO Mice
CD45+, a pan-leukocyte cell marker, labeled cells in fungiform papillae, the lamina propria immediately beneath the lingual epithelium, and throughout the deeper muscle layer of the anterior tongue. We are interested in immune cells that potentially affect taste bud integrity and function, so we spatially limited our analyses to fungiform papillae and nearby lamina propria. CD45+ immune cells were rarely observed within the lingual epithelium or inside taste bud borders in sham-sectioned mice. In response to nerve sectioning, immune cells infiltrated taste papillae and took on an activated morphology with projections extending from the cell (Figure 1A, white arrows). In contrast, CD45+ cells observed in the lingual muscle typically had a more rounded morphology (Figure S1B). NTPdase2, which labels the numerous type I glial-like taste cells, was used to identify taste buds. We note that NTPdase2 expression is also present in lingual nerve fibers as previously reported [34](Figure 1A, open arrows).
We analyzed immune responses at day 2, day 5, and day 60 post-injury. The timing was based on previous studies in rat demonstrating elevated macrophage responses during the first week after CT nerve sectioning. At day 60 post-axotomy, wild-type, C57BL/6J mice have recovered normal taste function for over 5 weeks, while Il1r KO mice still exhibit minimal taste responsiveness. We hypothesized that inappropriate immune responses at this later period could underlie the absence of taste bud regeneration and functional recovery in KO mice. We analyzed results with two-way ANOVAs followed by Bonferroni post-tests on selected comparisons: (1) sham-sectioned wild-type vs. Il1r KO; (2) CT-sectioned wild-type vs. Il1r KO to detect strain-related differences in baseline and injury-induced immune responses, respectively; and (3) sham vs. CT sectioned groups within each strain to detect the effects of surgical treatment. Non-significant statistics are reported in Appendix A.1.
Given the wispy processes of CD45+ immune cells, we thresholded immunopositive pixels in standard-sized regions of interest as in previous studies [6,16,28]. These analyses revealed early, dramatic differences in CD45+ immune responses to axotomy in wild-type and Il1r KO groups (Figure 1C). At day 2 post-injury, there were significant main effects of surgical treatment [F(1,12) = 20.83; P = 0.0006), mouse strain [F(1,12) = 7.326; P = 0.0191], and their interaction [F(1,12) = 20.87; P = 0.0006] on immune cell density in taste cell papillae and lamina propria. Specifically, lingual immune cells were significantly elevated in CT sectioned compared to sham-sectioned control mice (P = 0.0001). Notably, CD45+ cell density increased in wild-type compared to Il1r KO axotomized mice (Figure 1 A, B; P = 0.0010), as immune responses remained at baseline, sham levels in Il1r KO mice (Table A.1). As shown in Figure 1D at day 5 post-axotomy, there was a significant main effect of mouse strain [F(1,8) = 5.484; P = 0.0473], but no difference between groups in post-tests. At day 60 post-injury (Figure 1 A & D), despite variability in the sham, Il1r KO group, there was a significant effect of surgical treatment on CD45+ cell density [F(1,9) = 8.185; P = 0.0188] that was not reflected in post-tests. These results demonstrated that CD45+ leukocyte responses are elevated in wild-type but not Il1r KO at day 2 post-nerve injury.
2.2. CD68+ Macrophage Density Increases in Wild-Type Compared to Il1r KO Taste Papillae
In previous and pilot studies we observed robust macrophage responses to CT nerve sectioning in rodents, but few lingual T cells in contrast to human fungiform papillae [16,35,36]. Neutrophil responses were rapid, ending by day 2 after nerve injury [21]. Thus, to identify the subtype of CD45+ leukocytes infiltrating taste tissue we used the standard marker CD68 to quantify macrophage responses. CD68 is expressed in all macrophages isolated from tongue except for epithelial Langerhans cells [37]. Based on the results of the CD45 analysis, we focused on day 2 post-injury since immune responses returned to baseline across all mice strains and treatments at day 5 and 60 post-injury. Identified macrophages were morphologically similar to immune cells identified using the pan-immune marker CD45, and also found in similar locations after axotomy in wild-type mice (Figure 2). Macrophages were sporadically present within (Figure 2A, inset) and often near taste buds and nerve fibers of nerve-sectioned wild-type mice. Few CD68+ macrophages were observed in Il1r KO mice (Figure 2B).
At day 2 (Figure 2C), macrophage density significantly incrPeased in CT axotomized compared to sham wild-type mice [F(1,13) = 5.068; P = 0.0423]. There were also significant main effects of mouse strain [F(1,13) = 5.068; P = 0.0423], and the interaction between strain and surgical treatment [F(1,3) = 16.16; P = 0.0015]. Post-tests showed that increased CD68+ macrophage responses to injury in wild-type mice (P = 0.0013) were absent in Il1r KO mice (P = 0.0013). In conjunction with the CD45 analyses, these results demonstrate that early macrophage responses to taste bud denervation are suppressed in the absence of IL-1R signaling.
2.3. Immune Responses to Axotomy Are Primarily Composed of M2-Like Macrophages
Macrophages exhibit polarization into M1 and M2 subtypes, though little is known about their phenotype in normal or denervated taste papillae or lingual epithelium. While macrophage responses are more complex in vivo than in vitro [38,39], further phenotyping has provided insights to immune responses to axotomy and sterile tissue injury [40,41]. We refer to these macrophages as “M2-like” and “M1-like” (abbreviated as M2 and M1, respectively) because of this caveat. M2 macrophages are associated with anti-inflammatory responses and tissue repair [42,43,44,45]. Since macrophage responses were only elevated in wild-type mice in which taste buds later regenerated, we tested whether M2 responses to axotomy are prevalent in this group. We used CD206 and CD68 to identify M2, double-positive macrophages (denoted as CD206++) in taste papillae and lamina propria based on findings in the tongue and other injured tissues [46,47,48,49]. Taste buds were not labeled with NTPdase2 in these experiments because of antibody compatibility, but were easily identified morphologically by DAPI (Figure 3A, B; white dotted lines). In sham-sectioned wild-type or Il1r KO mice, few macrophages were present in fungiform papillae and lamina propriae (Figure 3A, B).
At day 2 after axotomy, CD206++ macrophages were numerous in wild-type mice but sparse in Il1r KO animals (Figure 3A, B). A majority of CD68+ macrophages (56.6%) expressed the M2 marker, CD206. There were significant effects of surgical treatment [F(1,8) = 40.30; P = 0.0002], mouse strain [F(1,8) = 36.09; P = 0.0003], and their interaction [F(1,8) = 32.23; P = 0.0005](Figure 3D). Double-positive macrophages were significantly elevated in nerve-injured vs. sham-sectioned wild-type mice (P = 0.0001). In contrast, there was a significant difference between strains in nerve-injured groups (P = 0.0001). Specifically, CD206++ macrophages remained at baseline, sham levels in Il1r KO mice.
Since CD206++ macrophage responses to injury were minimal at day 2, we also analyzed this population at day 5 to check for delayed responses in KO mice. M2-like macrophage responses followed a similar pattern at day 5 post-injury, although at lower levels in wild-type mice compared to day 2. However, there were significant effects of surgical treatment [F(1,8) = 15.44; P = 0.0044], mouse strain [F(1,8) = 20.96; P = 0.0018], and their interaction [F(1,8) = 15.44; P = 0.0044]). As at day 2, responses were elevated in the wild-type group after axotomy (P = 0.002), and significantly reduced in Il1r KO vs wild-type groups receiving CT nerve sectioning ( = 0.0013). In sum, M2-like macrophage responses were robust in control-strain mice, particularly at day 2, but negligible in the system-wide absence of IL-1R signaling.
2.4. M1-Like Macrophages Were Absent at Day 2 and 5 After CT Nerve Injury
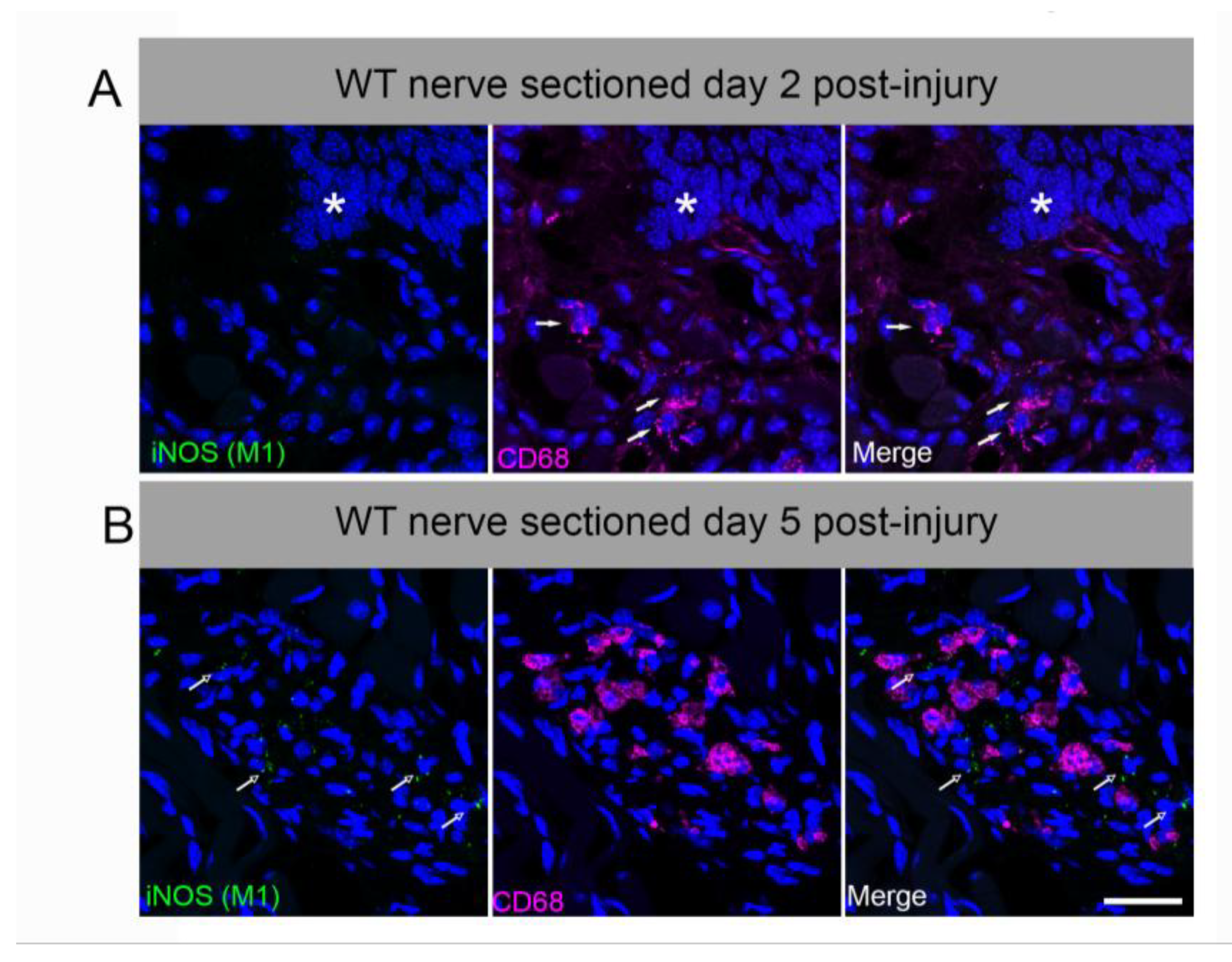
We tested whether a subset of lingual CD68+ macrophages express the proinflammatory M1-like marker, inducible nitric oxide synthase (iNOS) [45]. iNOS is not constitutively expressed, but upregulated by inflammatory stimuli including cytokines, LPS, and peripheral nerve injury [50]. Systemic LPS also induced iNOS in circumvallate taste buds and nerve fibers in previous reports [51]. We observed very limited iNOS expression in CD68+ macrophages deep in the lingual muscle and in ventral glands at day 2 or 5 after CT sectioning in wild-type mice (Figure A1). Macrophages in taste papillae and nearby lingual mucosa remained iNOS-negative, though positive control staining was observed in LPS-treated tongue and inflamed colon (not shown).
2.5. Taste Buds Degenerate in Both Wild-Type and Il1r KO Mice
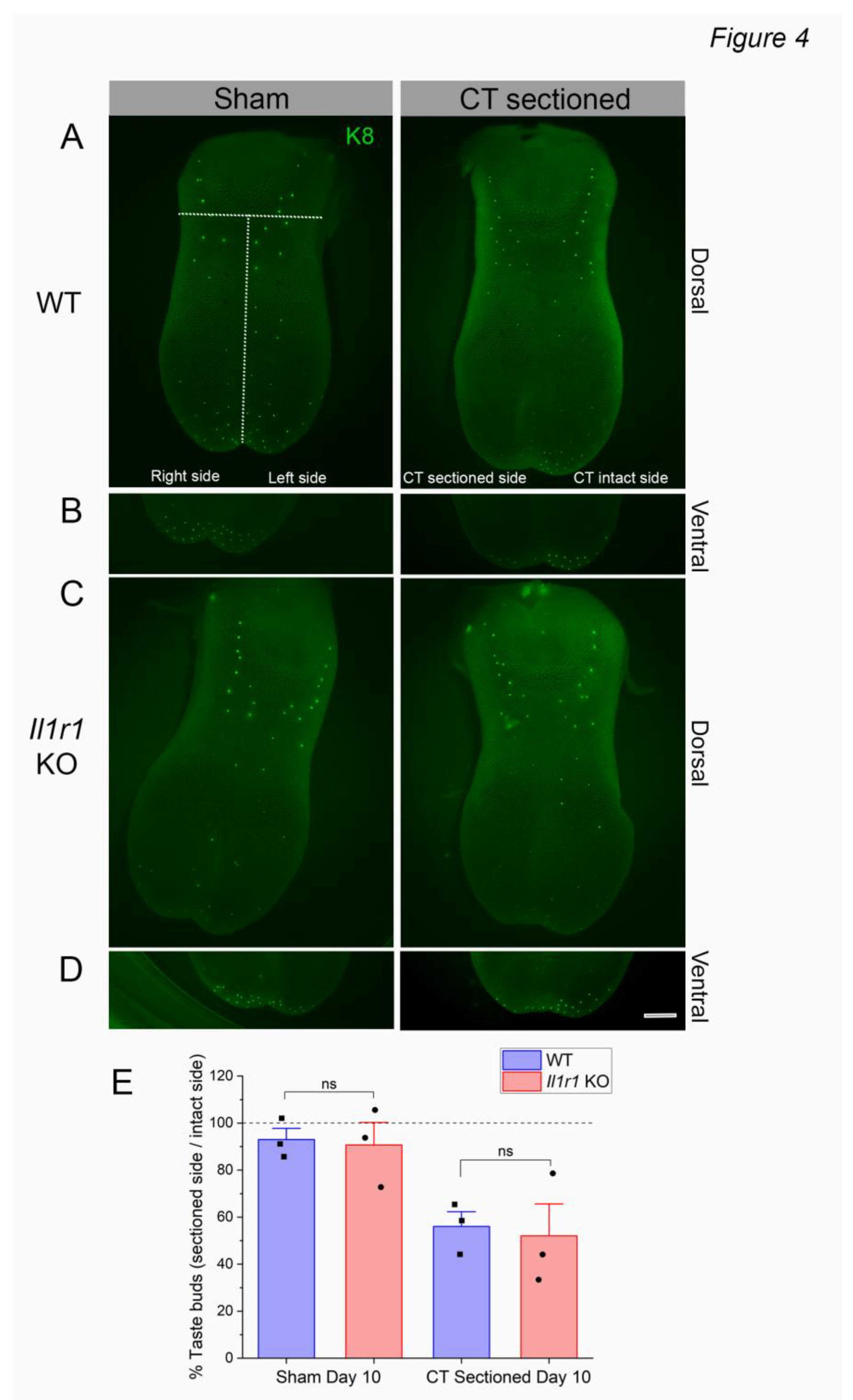
We previously demonstrated similar numbers of taste buds in wild-type and Il1r KO mice at day 5 following CT nerve axotomy [6]. However, given the reduced immune responses to injury in Il1r KO papillae (Figure 1, Figure 2 and Figure 3), taste bud degeneration could be delayed as a substrate for later deficits in regeneration and recovery in this strain. To test this, we counted taste buds expressing the marker Keratin (K)8 on the denervated and intact sides of lingual epithelial whole mounts at day 10 post-injury as shown in Figure 4A-D. We then analyzed the percentage of remaining taste buds (number taste buds on right or sectioned side / number on left or intact side X 100). As shown in Figure 4E, taste bud degeneration was similar in wild-type and Il1r KO mice. There was a significant main effect of surgical treatment [F (1, 8) = 16.85; P = 0.0034] but not strain [F (1, 8) = 0.1167; P = 7415]. There were no significant differences in post-tests, although sham vs. sectioned values approached significance for both wild-type (P = 0.0877) and Il1r KO (P = 0.0718) strains. This is not unexpected in wild-type mice, since CT nerve responses to taste stimuli appear in a subset of animals at day 11-17 post-injury indicating that taste buds are partially regenerated at this period [6]. Thus, taste buds degenerate at the same rate in the absence of IL-1R signaling despite suppressed macrophage responses to injury in these mice.
3. Discussion
Immune responses to peripheral nerve axotomy, and the consequences for subsequent axonal regeneration, have been studied in depth [52,53,54,55]. Sterile inflammation in denervated target tissue, in contrast, is not well-understood particularly in the taste system [14]. We previously demonstrated that the IL-1R is required for normal taste bud regeneration and the restoration of gustatory function [33]. In the absence of signaling through this pleiotropic cytokine pathway, taste loss persists after experimental axotomy. Here, we report that leukocytes rapidly respond to CT nerve sectioning in control-strain mice that later recover normal taste function. The immune response is dominated by CD206+ M2-like macrophages which infiltrate the denervated anterior taste tissue within days. At the same early post-injury period (i.e., days 2 and 5), CD45+ leukocytes, CD68+ macrophages, and CD206++ M2 macrophages remain at sham-like control levels in the taste tissue of axotomized Il1r KO mice. Even 8 weeks after nerve sectioning, immune responses remain minimal in KO mice indicating their absence rather than a delay.
We tested whether taste receptor cell degeneration is delayed in Il1r KO mice, but that is not the case. Instead, taste bud numbers are similar at day 5 [33] and 10 post-injury across mouse strains, despite the absence of CD45+ leukocyte responses in KO mice. Neurophysiological responses to taste stimuli return at day 11-17 after CT axotomy in a subset of wild-type mice, indicating that nerve fibers and new taste receptor cells are reconnecting at this period. Partial regeneration at this period likely explains why the reduced number of taste buds in axotomized vs. sham mice approaches but does not reach significance. It is unlikely that Il1r KO delays taste bud degeneration beyond day 10, since the CT nerve remains unresponsive to taste stimuli from day 11-60 post-injury in these mice. Our prediction was based on the likelihood that CD45+ immune cells, and particularly macrophages, clear apoptotic taste receptor cells. CD45 is expressed on all cells of hematopoietic lineage, including lymphocytes (e.g., T cells and B cells), innate immune cells, dendritic cells, and at least some fibroblasts.
Our finding that taste bud degeneration proceeds in the absence of infiltrating leukocytes highlights the uncertainty over the fate of denervated, apoptotic taste receptor cells [56]. Suzuki and colleagues reported that macrophages phagocytose nerve fibers while fibroblasts engulf taste receptor cells in posterior mouse taste buds after glossopharyngeal nerve sectioning [57]. This is consistent with our results if CD45-negative fibroblasts phagocytose anterior taste receptor cells while M2-like macrophages clear taste fiber debris. A small subpopulation of CD45+ cells isolated from murine tongue was comprised of fibroblasts in a recent study using single-cell RNA sequencing [37]. However, the sizable population of vimentin-positive stromal cells in other reports suggests that many fibroblasts are CD45-negative [58,59]. Fibroblasts play a role in wound-healing in the tongue, and are suggested to recruit macrophages to the injury site [60]. Taste cells may also be cleared by type I, glial-like taste cells [61,62]. Desquamation through the taste pore has been suggested as a mechanism for taste receptor cell clearance, though it is unclear that this mechanism could accommodate widescale taste cell death after axotomy [63,64].
Two novel, major subpopulations of CD45+ macrophages in uninjured mouse tongue were recently reported. Cx3cr1+ macrophages occupy the taste papillae and lamina propria, while Fol2+ cells reside in the lamina propria and deeper lingual muscle [37]. Both of these resident macrophage populations express CD68 [37]. The spatial segregation (i.e., taste papillae and lamina propria vs. muscle) and morphological differences in the two CD68+ macrophage subsets in the current study may correspond to these populations, though additional immune responses are likely induced by nerve sectioning. The authors suggest that Cx3Cr1+ macrophages may communicate with nearby nerve fibers during homeostasis and after injury as recently reported in skin [65]. Thus, even though taste buds degenerate at the same rate in wild-type and Il1r KO mice, we cannot rule out interactions between macrophages in papillae, lamina propria and muscle and CT nerve fibers. Since anterior taste buds are dependent on trophic support from the CT nerve, major changes in the rate that gustatory fibers degenerate are unlikely [14]. In a previous study, we also found that the subset of taste that regenerate (~40%) in Il1r KO mice are innervated. In fact, the percent of the taste bud occupied by CT fibers is slightly elevated in the absence of IL-1R signaling at day 56-60 post-injury [33].
IL-1 is a master regulatory cytokine. Knocking out its receptor will inhibit many inflammatory signals that stimulate immune responses to axotomy and subsequent nerve and taste receptor cell degeneration [66,67]. While resident CD45+ populations are similar during homeostasis (e.g. sham wild-type vs. KO groups), injury unmasks the critical role of IL-1R cytokine signaling in taste regeneration and recovery [33]. For example, macrophage chemoattractant protein (MCP)-1, intracellular adhesion molecule (ICAM)-1 and vascular adhesion molecule (VCAM)-1 are upregulated at 24 and 48 hours after CT nerve axotomy in rats [18,19]. IL-1 signaling is a major inducer of these recruitment signals, which likely results in the suppressed immune responses to nerve injury reported here [66,67,68]. One of the challenges in identifying molecular mechanisms by which IL-1 signaling contributes to taste regeneration is the huge number of biological processes that it controls [66,67,68]. The current study is a step in that direction since we report that M2-like macrophages respond to CT sectioning in control but not Il1r KO mice. Importantly, taste buds degenerate at the same rate in both strains. Even though this is a negative result, it shifts the focus of future studies to later regenerative processes.
We suggest that IL-1 signaling may have a growth factor-like role in the injured taste system. This cytokine could target taste progenitor cell proliferation, differentiation, or their ability to attract regenerating CT nerve fibers. One example of a similar non-canonical role for IL-1 occurs in skin wound healing. Keratinocyte-derived IL-1 stimulates growth factor release, including keratinocyte growth factor (KGF)-1/fibroblast growth factor (KGF)-7 from fibroblasts regulating skin wound repair [69]. In the gut, IL-1 and other cytokines induce intestinal stem cell proliferation fostering regeneration and repair [70]. The olfactory epithelium also upregulates Il1b and other proinflammatory cytokines in a tumor necrosis factor (Tnf)-overexpression model of chronic inflammation. Horizontal basal stem cells recruit macrophages and other leukocytes through an NFkB-dependent mechanism that is acutely pro-regenerative but chronically detrimental to olfactory sensory neuron regeneration and functional recovery [71,72]. The requirement for tightly regulated inflammatory responses might be similar for taste bud regeneration. Importantly, better understanding of neuroimmune mechanisms leading to taste recovery could suggest new strategies to treat the significant clinical problem of taste dysfunction from chemotherapy, injury, and long COVID-19 [73,74].
4. Materials and Methods
4.1. Animals
Adult male and female mice were 8-16 weeks old during all experimental procedures. Wild-type C57BL/6J (WT) and B6.129S7-Il1r1tm1lmx/J were obtained from the Jackson Laboratory (000664; #003245, respectively) and bred in-house. Sex differences were not observed in previous studies focused on CT sectioning in rodents [6,16,17,19,22,26]. Genotypes were confirmed using Jackson Laboratory protocols on site or outsourced (Transnetyx). Specified pathogen-free mice were kept on a 12:12 h light: dark cycle with ad libitum access to rodent chow (Envigo Tekland) and filtered tap water.
4.2. Chorda Tympani (CT) Nerve Sectioning
A ketamine (50 mg/kg) and xylazine (10 mg/kg) cocktail was administered intraperitoneally to anesthetize mice. Once a surgical plane of anesthesia was achieved, mice were moved to a water-circulating heating pad to maintain body temperature. Mice were given ketoprofen analgesic (5 mg/kg bw) prior to the start of surgery. CT nerve sectioning was done as previously described [33]. Access to the CT nerve was gained ventrally through the neck and transected after bifurcation from the lingual nerve with severed ends being left in place. Sham-sectioned mice underwent the same procedure, however, the CT nerve remained intact.
4.3. Tissue Collection and Immunofluorescence
Two, five, or 60 days after CT nerve sectioning, mice were sacrificed, and tongues were removed with the mandible still attached. Samples were fixed in paraformaldehyde for 5-7 h then tongues dissected free and placed in cryoprotectant overnight [6,75]. Tongues were frozen in O.C.T (Fisher) and 8 µm sagittal sections were obtained for immunostaining. Samples were rinsed with PBS (7.5 pH) and placed in 5% blocking solution for 45 min for CD45 analysis and 30 minutes for CD68 and CD206 at room temperature. Staining for iNOS followed a similar procedure as described, however, we included a 30-minute permeabilization step in 0.5% triton-x in PBS before placing samples in blocking solution (Sigma-Aldrich 9036-19-5). NTPdase2 (1:1000, Labome, 501943785, rabbit anti-mouse) was used stain Type I taste cells to identify taste buds. We used NTPdase2 instead of Keratin 8 since antigen retrieval interfered with immune cell staining. CD45 (1:100; R&D Systems, AF114, goat anti-mouse) is a pan-immune cell marker while CD68 (1:100; Abcam, ab53444, rat anti-mouse) identifies macrophages. To observe M1 and M2 macrophage subtypes, iNOS (1:100; Abcam, ab3523, rabbit anti-mouse) and CD206 (1:1000; Abcam, ab64693, rabbit anti-mouse) were used, respectively. Tongue sections were incubated in primary antibodies in a humidified box at 4°C overnight. Samples were rinsed with PBS and incubated in donkey anti-goat 594 (1:500; Abcam, ab150136), donkey anti-rabbit 488 (1:1000; Jackson ImmunoResearch, 711-545-152), goat anti-rabbit 488 (1:1000; Jackson ImmunoResearch, 111-545-144) and goat anti-rat 594 (1:500; Abcam, ab53444) in a humidified box at room temperature for 1 hr. Nuclei were stained with DAPI(#D1306; Invitrogen/Fisher Scientific) and slides coverslipped with Fluoromount-G (#0100-01: Southern Biotechnology).
4.4. Whole Mount Lingual Epithelium Analysis
Il1r KO or wild-type mice received CT nerve axotomy then euthanized with isofluorane followed by bilateral thoracotomy 10 days later and whole tongues were dissected from the jaw. We selected this period since we previously found no difference in the number of taste buds remaining at day 5 post-injury in wild-type or Il1r KO mice [33]. Tongues were washed in PBS and injected with ~1.5 mL collagenase A (1 mg/mL, catalog no. 10103578001; Roche) and dispase II (2.5 mg/mL, D4693-1G; Sigma). After injection, tongues were incubated in 300mL of PBS with an oxygen bubbler for 45 minutes at room temperature, then placed on a shaker and fixed in 4% PFA for 1 hour. Intact whole epithelial were dissected from the underlying muscle and washed in PBS. Lingual epithelia were placed in blocking solution for 1 hour then incubated in primary antibody K8 TROMA (1:500; DSHB) for 2 days to identify tastebuds. Epithelia were washed with PBS, incubated in secondary antibody (donkey α rat 488, 1:500, 712-545-150; Jackson ImmunoResearch) for 2 days, and counterstained with DAPI in ddH2O.
We counted taste buds on the right / denervated or left / intact side of the tongue (in sham- or chorda tympani nerve-sectioned mice) on images taken with a Leica M165 FC stereoscope. Separate images were taken of the dorsal epithelium and the ventral tip which has a high density of fungiform taste buds. We expressed the number of taste buds on the right or injured side of the tongue relative to the number on the intact side to obtain the percent degeneration. Brightness and contrast were adjusted to assist in counting taste buds.
4.5. Image Analysis
Non-overlapping images of either taste buds, papilla or in the absence of either, lamina propria, were taken at 40x magnification. CD45+ and CD68+ pixels were thresholded and calculated as % thresholded area relative to the region of interest (ROI 581.5065 mm2)[27,33,76,77]. The experimenter was blinded to surgical treatment and strain for most analyses. Serial sagittal sections were analyzed using a BX51 microscope with epifluorescence (Olympus), a digital monochrome camera (Cool Snap, Roper Scientific), and MetaMorph software (MDS Analytical Technologies). Quantification of iNOS and CD206 positive cells were done using the same epifluorescence microscope as previously described. Individual cells were considered double positive for CD68 and iNOS or CD206 if colocalization was found around DAPI positive nuclei and could be distinguished from background. For presentation purposes, z-stack images with a step-size of 1 µm were also captured with a Nikon A1R confocal/multiphoton microscope. Images of taste buds were captured at a magnification of 60x with a 3x zoom setting. We then converted images from NIS Elements (version 4.30.01) to maximum Z projections using ImageJ. Images for Figure S1 were captured with a Leica Stellaris confocal system at 40x (2.5x zoom) and a 1 µm step-size, and LAX .lif files converted to maximum Z projections in ImageJ. Images were minimally adjusted for brightness and contrast in Adobe Photoshop with the same settings applied equally to all panels.
4.6. Statistical Analysis
Two-way ANOVAs were performed in GraphPad Prism (version 10.6.0), using mouse strain (wild-type or Il1r KO) and surgical treatment (sham or CT sectioned) as main factors. Bonferroni posttests were used for multiple comparisons. P values ≤ 0.05 were considered significant. F values, degrees of freedom, and P values are reported in the text when significant, and in Appendix A.1 when non-significant.
Author Contributions
Conceptualization, JB and LPM; Methodology, JB, YB, and LPM.; Formal Analysis, JB and LPM; Investigation, JB, YB, TA, EH, OO, TH, and LPM.; Writing – Original Draft Preparation, JB and LPM; Writing – Review & Editing, JB and LM; Visualization, JB, EH, LPM; Supervision, LPM.; Project Administration, LM.; Funding Acquisition, LPM. JB used Biorender to create the graphical abstract.
Funding
This research was funded by the National Institutes of Deafness and Communication Disorders (NIDCD) R01 DC016668 to LPM.
Institutional Review Board Statement
Animal procedures followed National Institutes of Health guidelines and were approved by the Augusta University Institutional Animal Care and Use Committee.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
Appendix A.1

Table A1.
Table showing nonsignificant two-way ANOVA statistics.
| Two-way ANOVA Comparison | Main Factor | F-statistic | df | P | |
| Figure 1. CD45+ immune response | Day 5 | Surgical treatment | 0.7301 | 1, 8 |
0.4177 |
| Interaction | 4.102 | 0.0774 | |||
| Day 60 | Mouse strain | 1.575 | 1, 9 | 0.2411 | |
| Interaction | 0.4634 | 0.5132 | |||
| Figure 4. Taste bud degeneration | Day 10 | Mouse strain | 0.1167 | 1, 8 | 0.7415 |
| Interaction | 0.008495 | 0.9288 | |||
Figure A1.
Lingual M1-like macrophages expressing iNOS are rare or absent after axotomy. Macrophages were identified with CD68 (magenta, arrows). (A) Macrophages in proximity to taste buds were increased at day 2 post-injury, as in Fig. 2. However, iNOS was not co-expressed with CD68 in taste papillae or lamina propria. White asterisk marks the base of a fungiform papilla. (B) At day 5 post-axotomy, CD68+ macrophages are most prominent in the deep lingual muscle. Punctate iNOS was observed nearby, but rarely co-localized with macrophages (green, open arrows). Scale bar in B = 25 µm.
Figure A1.
Lingual M1-like macrophages expressing iNOS are rare or absent after axotomy. Macrophages were identified with CD68 (magenta, arrows). (A) Macrophages in proximity to taste buds were increased at day 2 post-injury, as in Fig. 2. However, iNOS was not co-expressed with CD68 in taste papillae or lamina propria. White asterisk marks the base of a fungiform papilla. (B) At day 5 post-axotomy, CD68+ macrophages are most prominent in the deep lingual muscle. Punctate iNOS was observed nearby, but rarely co-localized with macrophages (green, open arrows). Scale bar in B = 25 µm.
References
- Beidler, L.M.S., R.L. Renewal of cells within taste buds. J Cell Biol 1965, 27, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Finger, T.E.B.; Barlow, L.A. Cellular diversity and regeneration in taste buds. Current Opinion in Physiology 2021, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Perea-Martinez, I.; Nagai, T.; Chaudhari, N. Functional Cell Types in Taste Buds Have Distinct Longevities. PLoS One 2013, 8, e53399. [Google Scholar] [CrossRef]
- Finger, T.E.; Barlow, L.A. Cellular diversity and regeneration in taste buds. Current Opinion in Physiology 2021, 20, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Roper, S.D.; Chaudhari, N. Taste buds: cells, signals and synapses. Nat. Rev. Neurosci. 2017, 18, 485–497. [Google Scholar] [CrossRef]
- Dong, G.; Kogan, S.; Venugopal, N.; Chang, E.; He, L.; Faal, F.; Shi, Y.; McCluskey, L.P. Interleukin (IL)-1 receptor signaling is required for complete taste bud regeneration and the recovery of neural taste responses following axotomy. J Neurosci 2023. [Google Scholar] [CrossRef]
- Guagliardo, N.A.; Hill, D.L. Fungiform taste bud degeneration in C57BL/6J mice following chorda-lingual nerve transection. J. Comp. Neurol. 2007, 504, 206–216. [Google Scholar] [CrossRef]
- Cheal, M.; Dickey, W.P.; Jones, L.B.; Oakley, B. Taste fiber responses during reinnervation of fungiform papillae. J. Comp. Neurol. 1977, 172, 627–646. [Google Scholar] [CrossRef]
- Cooper, K.W.; Brann, D.H.; Farruggia, M.C.; Bhutani, S.; Pellegrino, R.; Tsukahara, T.; Weinreb, C.; Joseph, P.V.; Larson, E.D.; Parma, V.; Albers, M.W.; Barlow, L.A.; Datta, S.R.; Di Pizio, A. COVID-19 and the Chemical Senses: Supporting Players Take Center Stage. Neuron 2020, 219–233. [Google Scholar] [CrossRef]
- Doty, R.L. Systemic diseases and disorders. Handb Clin Neurol 2019, 361–387. [Google Scholar] [CrossRef]
- McManus, L.J.; Stringer, M.D.; Dawes, P.J. Iatrogenic injury of the chorda tympani: a systematic review. J Laryngol Otol 2012, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Snyder, D.J.B.; Bartoshuk, L.M. Oral sensory nerve damage: Causes and consequences. Rev Endocr Metab Disord 2016, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhou, M.; Brand, J.; Huang, L. Inflammation and taste disorders: mechanisms in taste buds. Ann. N. Y. Acad. Sci. 2009, 1170, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, H.G.; Miller, E.; White-Canale, A.; McCluskey, L.P. Immune responses in the injured olfactory and gustatory systems: a role in olfactory receptor neuron and taste bud regeneration? Chem. Senses 2022, 47. [Google Scholar] [CrossRef]
- Liu, G.; Zong, G.; Doty, R.L.; Sun, Q. Prevalence and risk factors of taste and smell impairment in a nationwide representative sample of the US population: a cross-sectional study. BMJ Open 2016, 6, e013246. [Google Scholar] [CrossRef]
- McCluskey, L.P. Up-regulation of activated macrophages in response to degeneration in the taste system: effects of dietary sodium restriction. J Comp Neurol 2004, 43–55. [Google Scholar] [CrossRef]
- Steen, P.W.; Shi, L.; He, L.; McCluskey, L.P. Neutrophil responses to injury or inflammation impair peripheral gustatory. Neuroscience 2010, 894–908. [Google Scholar] [CrossRef]
- Cavallin, M.A.; McCluskey, L.P. Upregulation of the chemokine monocyte chemoattractant protein-1 following unilateral nerve injury in the peripheral taste system. Neurosci. Lett. 2007, 413, 187–190. [Google Scholar] [CrossRef]
- Cavallin, M.A.; McCluskey, L.P. Upregulation of intracellular adhesion molecule (ICAM)-1 and vascular cell adhesion molecule (VCAM)-1 after unilateral nerve injury in the peripheral taste system. J. Neurosci. Res. 2007, 85, 364–372. [Google Scholar] [CrossRef]
- Shi, L.; He, L.; Sarvepalli, P.; McCluskey, L.P. Functional role for interleukin-1 in the injured peripheral taste system. J. Neurosci. Res. 2012, 90, 816–830. [Google Scholar] [CrossRef]
- Steen, P.W.; Shi, L.; He, L.; McCluskey, L.P. Neutrophil responses to injury or inflammation impair peripheral gustatory function. Neuroscience 2010, 167, 894–908. [Google Scholar] [CrossRef] [PubMed]
- Cavallin, M.A.M.; McCluskey, L.P. Lipopolysaccharide-induced up-regulation of activated macrophages in the degenerating taste system. J Neurosci Res 2005, 75–84. [Google Scholar] [CrossRef] [PubMed]
- DeFrancesco-Lisowitz, A.; Lindborg, J.A.; Niemi, J.P.; Zigmond, R.E. The neuroimmunology of degeneration and regeneration in the peripheral nervous system. Neuroscience 2015, 174–203. [Google Scholar] [CrossRef] [PubMed]
- Dubovy, P. Wallerian degeneration and peripheral nerve conditions for both axonal regeneration and neuropathic pain induction. Ann Ant 2011, 267–275. [Google Scholar] [CrossRef]
- Guagliardo, N.A.; West, K.N.; McCluskey, L.P.; Hill, D.L. Attenuation of peripheral salt taste responses and local immune function contralateral to gustatory nerve injury: effects of aldosterone. Am J Physiol Regul Integr Comp Physiol 2009, R1103–1110. [Google Scholar] [CrossRef]
- He, L.; Yadgarov, A.; Sharif, S.; McCluskey, L.P. Aging profoundly delays functional recovery from gustatory nerve injury. Neuroscience 2012, 208–218. [Google Scholar] [CrossRef]
- McCluskey, L.P. Up-regulation of activated macrophages in response to degeneration in the taste system: effects of dietary sodium restriction. J. Comp. Neurol. 2004, 479, 43–55. [Google Scholar] [CrossRef]
- Dong, G.; Boothe, K.; He, L.; Shi, Y.; McCluskey, L.P. Altered peripheral taste function in a mouse model of inflammatory bowel disease. Sci. Rep. 2023, 13, 18895. [Google Scholar] [CrossRef]
- Glaccum, M.B.; Stocking, K.L.; Charrier, K.; Smith, J.L.; Willis, C.R.; Maliszewski, C.; Livingston, D.J.; Peschon, J.J.; Morrissey, P.J. Phenotypic and functional characterization of mice that lack the type I receptor for IL-1. J. Immunol. 1997, 159, 3364–3371. [Google Scholar] [CrossRef]
- Labow, M.; Shuster, D.; Zetterstrom, M.; Nunes, P.; Terry, R.; Cullinan, E.B.; Bartfai, T.; Solorzano, C.; Moldawer, L.L.; Chizzonite, R.; et al. Absence of IL-1 signaling and reduced inflammatory response in IL-1 type I receptor-deficient mice. J. Immunol. 1997, 159, 2452–2461. [Google Scholar] [CrossRef]
- Horai, R.; Asano, M.; Sudo, K.; Kanuka, H.; Suzuki, M.; Nishihara, M.; Takahashi, M.; Iwakura, Y. Production of mice deficient in genes for interleukin (IL)-1alpha, IL-1beta, IL-1alpha/beta, and IL-1 receptor antagonist shows that IL-1beta is crucial in turpentine-induced fever development and glucocorticoid secretion. J. Exp. Med. 1998, 187, 1463–1475. [Google Scholar] [PubMed]
- Nadeau, S.; Filali, M.; Zhang, J.; Kerr, B.J.; Rivest, S.; Soulet, D.; Iwakura, Y.; de Rivero Vaccari, J.P.; Keane, R.W.; Lacroix, S. Functional recovery after peripheral nerve injury is dependent on the pro-inflammatory cytokines IL-1beta and TNF: implications for neuropathic pain. J. Neurosci. 2011, 31, 12533–12542. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.; Kogan, S.; Venugopal, N.; Chang, E.; He, L.; Faal, F.; Shi, Y.; McCluskey, L.P. Interleukin (IL)-1 Receptor Signaling Is Required for Complete Taste Bud Regeneration and the Recovery of Neural Taste Responses following Axotomy. The Journal of Neuroscience 2023, 43, 3439–3455. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.L. Glial responses after chorda tympani nerve injury. J Comp Neurol 2012, 520, 2712–2729. [Google Scholar] [CrossRef]
- Feng, P.; Wang, H.; Feldman, R.S.; Pribitkin, E.A.; Breslin, P.A. The T cells in peripheral taste tissue of healthy human adults: predominant memory T cells and Th-1 cells. Chem. Senses 2010, 35, 501–509. [Google Scholar] [CrossRef]
- Feng, P.; Yee, K.K.; Rawson, N.E.; Feldman, L.M.; Feldman, R.S.; Breslin, P.A. Immune cells of the human peripheral taste system: dominant dendritic cells and CD4 T cells. Brain. Behav. Immun. 2009, 23, 760–766. [Google Scholar] [CrossRef]
- Lyras, E.M.; Zimmermann, K.; Wagner, L.K.; Dörr, D.; Klose, C.S.N.; Fischer, C.; Jung, S.; Yona, S.; Hovav, A.-H.; Stenzel, W.; et al. Tongue immune compartment analysis reveals spatial macrophage heterogeneity. eLife 2022, 11, e77490. [Google Scholar] [CrossRef]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu Rev Pathol 2020, 15, 123–147. [Google Scholar] [CrossRef]
- Sica, A.M.; Mantovani, A. Macrophage plasticity and polarization: in vivo veritas. J Clin Invest 2012, 787–795. [Google Scholar] [CrossRef]
- Chen, P.; Piao, X.; Bonaldo, P. Role of macrophages in Wallerian degeneration and axonal regeneration after peripheral nerve injury. Acta Neuropathol 2015, 130, 605–618. [Google Scholar] [CrossRef]
- Stratton, J.A.S.; Shah, P.T. Macrophage polarization in nerve injury: do Schwann cells play a role? Neural Regen Res 2016, 11, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Sinha, M.; Datta, S.; Abas, M.; Chaffee, S.; Sen, C.K.; Roy, S. Monocyte and macrophage plasticity in tissue repair and regeneration. Am. J. Pathol. 2015, 185, 2596–2606. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Peng, J.; Han, G.H.; Ding, X.; Wei, S.; Gao, G.; Huang, K.; Chang, F.; Wang, Y. Role of macrophages in peripheral nerve injury and repair. Neural Regen Res 2019, 14, 1335–1342. [Google Scholar] [CrossRef]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. 2020, 15, 123–147. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Zhao, F.; Cheng, H.; Su, M.; Wang, Y. Macrophage polarization: an important role in inflammatory diseases. Front. Immunol. 2024, 15–2024. [Google Scholar] [CrossRef]
- Kiguchi, N.; Kobayashi, Y.; Saika, F.; Sakaguchi, H.; Maeda, T.; Kishioka, S. Peripheral interleukin-4 ameliorates inflammatory macrophage-dependent neuropathic pain. Pain 2015, 156, 684–693. [Google Scholar] [CrossRef]
- Huang, T.-C.; Wu, H.-L.; Chen, S.-H.; Wang, Y.-T.; Wu, C.-C. Thrombomodulin facilitates peripheral nerve regeneration through regulating M1/M2 switching. J. Neuroinflammation 2020, 17, 240–240. [Google Scholar] [CrossRef]
- Chiloeches, M.L.; Bergonzini, A.; Frisan, T.; Martin, O.C.B. Characterization of macrophage infiltration and polarization by double fluorescence immunostaining in mouse colonic mucosa. STAR Protoc 2021, 2, 100833. [Google Scholar] [CrossRef]
- McKendrick, J.G.; Jones, G.R.; Elder, S.S.; Watson, E.; T'Jonck, W.; Mercer, E.; Magalhaes, M.S.; Rocchi, C.; Hegarty, L.M.; Johnson, A.L.; et al. CSF1R-dependent macrophages in the salivary gland are essential for epithelial regeneration after radiation-induced injury. Sci Immunol 2023, 8, eadd4374. [Google Scholar] [CrossRef]
- Kubes, P.; Jenne, C. Immune Responses in the Liver. Annu Rev Immunol 2018, 36, 247–277. [Google Scholar] [CrossRef]
- Wu, Z.; Huang, Y.; Hu, W.; Ren, L.; Jiang, P.; Margolskee, R.F.; Wang, H.; Feng, S. Lipopolysaccharide-induced inflammation increases nitric oxide production in taste buds. Brain, Behavior, and Immunity 2022, 103, 145–153. [Google Scholar] [CrossRef] [PubMed]
- DeFrancesco-Lisowitz, A.; Lindborg, J.A.; Niemi, J.P.; Zigmond, R.E. The neuroimmunology of degeneration and regeneration in the peripheral nervous system. Neuroscience 2015, 302, 174–203. [Google Scholar] [CrossRef] [PubMed]
- Zigmond, R.E.; Echevarria, F.D. Macrophage biology in the peripheral nervous system after injury. Prog. Neurobiol. 2019, 173, 102–121. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Piao, X.; Bonaldo, P. Role of macrophages in Wallerian degeneration and axonal regeneration after peripheral nerve injury. Acta Neuropathol. 2015, 130, 605–618. [Google Scholar] [CrossRef]
- Bastien, D.; Lacroix, S. Cytokine pathways regulating glial and leukocyte function after spinal cord and peripheral nerve injury. Exp. Neurol. 2014, 258, 62–77. [Google Scholar] [CrossRef]
- Takeda, M.; Suzuki, Y.; Obara, N.; Nagai, Y. Apoptosis in mouse taste buds after denervation. Cell Tissue Res. 1996, 286, 55–62. [Google Scholar] [CrossRef]
- Suzuki, Y.; Takeda, M.; Obara, N.; Nagai, Y. Phagocytic cells in the taste buds of rat circumvallate papillae after denervation. Chem. Senses 1996, 21, 467–476. [Google Scholar] [CrossRef]
- Seubert, A.C.; Krafft, M.; Bopp, S.; Helal, M.; Bhandare, P.; Wolf, E.; Alemany, A.; Riedel, A.; Kretzschmar, K. Spatial transcriptomics reveals molecular cues underlying the site specificity of the adult mouse oral mucosa and its stem cell niches. Stem Cell Reports 2024, 19, 1706–1719. [Google Scholar] [CrossRef]
- Boggs, K.; Venkatesan, N.; Mederacke, I.; Komatsu, Y.; Stice, S.; Schwabe, R.F.; Mistretta, C.M.; Mishina, Y.; Liu, H.-X. Contribution of Underlying Connective Tissue Cells to Taste Buds in Mouse Tongue and Soft Palate. PLoS One 2016, 11, e0146475. [Google Scholar] [CrossRef]
- Shimizu, K.; Arimura, Y.; Ezaki, T. Roles of podoplanin positive cells in wound healing of the mouse tongue. The FASEB Journal 2015, 29. [Google Scholar] [CrossRef]
- Farbman, A.I. Fine structure of degenerating tast buds after denervation. J. Embryol. Exp. Morphol. 1969, 22, 55–68. [Google Scholar] [PubMed]
- Wilson, C.E.; Lasher, R.S.; Salcedo, E.; Yang, R.; Dzowo, Y.; Kinnamon, J.C.; Finger, T.E. Death in the Taste Bud: Engulfment of Dying Taste Receptor Cells by Glial-Like Type I Cells. Glia 2025. [Google Scholar] [CrossRef] [PubMed]
- Guth, L. Degeneration and regeneration of taste buds. In Handbook of sensory physiology; Beidler, L., Ed.; Springer: New York, 1971; Volume IV, pp. 63–74. [Google Scholar]
- Olmsted, J.M.D. The effects of cutting the lingual nerve of the dog. Journal Comp Neurol 1921, 435, 149–154. [Google Scholar] [CrossRef]
- Kolter, J.; Feuerstein, R.; Zeis, P.; Hagemeyer, N.; Paterson, N.; d’Errico, P.; Baasch, S.; Amann, L.; Masuda, T.; Lösslein, A.; et al. A Subset of Skin Macrophages Contributes to the Surveillance and Regeneration of Local Nerves. Immunity 2019, 50, 1482–1497.e1487. [Google Scholar] [CrossRef]
- Dinarello, C.A. The IL-1 family and inflammatory diseases. Clin. Exp. Rheumatol. 2002, 20, S1–13. [Google Scholar]
- Dinarello, C.A. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): an overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Werner, S.; Krieg, T.; Smola, H. Keratinocyte-fibroblast interactions in wound healing. J. Invest. Dermatol. 2007, 127, 998–1008. [Google Scholar] [CrossRef]
- Karin, M.; Clevers, H. Reparative inflammation takes charge of tissue regeneration. Nature 2016, 529, 307–315. [Google Scholar] [CrossRef]
- Chen, M.; Reed, R.R.; Lane, A.P. Chronic Inflammation Directs an Olfactory Stem Cell Functional Switch from Neuroregeneration to Immune Defense. Cell Stem Cell 2019, 25, 501–513.e505. [Google Scholar] [CrossRef]
- Lane, A.P.; Turner, J.; May, L.; Reed, R. A genetic model of chronic rhinosinusitis-associated olfactory inflammation reveals reversible functional impairment and dramatic neuroepithelial reorganization. J. Neurosci. 2010, 30, 2324–2329. [Google Scholar] [CrossRef]
- Risso, D.; Drayna, D.; Morini, G. Alteration, Reduction and Taste Loss: Main Causes and Potential Implications on Dietary Habits. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Doty, R.L. Chapter 25 - Treatments for smell and taste disorders: A critical review. In Handb. Clin. Neurol.; Doty, R.L., Ed.; Elsevier, 2019; Volume 164, pp. 455–479. [Google Scholar]
- Kumari, A.; Yokota, Y.; Li, L.; Bradley, R.M.; Mistretta, C.M. Species generalization and differences in Hedgehog pathway regulation of fungiform and circumvallate papilla taste function and somatosensation demonstrated with sonidegib. Sci Rep 2018, 16150. [Google Scholar] [CrossRef]
- Cavallin, M.A.; McCluskey, L.P. Lipopolysaccharide-induced up-regulation of activated macrophages in the degenerating taste system. J. Neurosci. Res. 2005, 80, 75–84. [Google Scholar] [CrossRef]
- He, L.; McCluskey, L.P. Regression of Lingual Lymphatic Vessels in Sodium-restricted Mice. J. Histochem. Cytochem. 2018, 66, 377–384. [Google Scholar] [CrossRef]
Figure 1.
CD45+ immune responses are suppressed in the early post-injury period in Il1r KO compared to wild-type mice. (A) The density of CD45+ immune cell responses (magenta, white arrows) increases near denervated taste buds at day 2 post-injury in wild-type mice. Taste buds are visualized with the type I taste cell marker, NTPdase2 (green), which also labels nerve fibers in the papilla (open arrows). CD45+ immune responses return to baseline at day 5 post-injury and remain at low levels at day 60 in wild-type papillae with regenerated, functional taste buds. (B) In Il1r KO mice, few CD45+ immune cells occupy taste papillae at any post-injury period tested. (C) CD45+ pixels were thresholded, expressed as the percent / standard area, and summed at each period (n=3-4 mice/group). There was a significant increase in the mean (+SEM) CD45+ responses at day 2 post-injury compared to sham wild-type mice or axotomized Il1r KO mice. Bonferroni post-tests revealed no significant differences in CD45+ immune responses among groups at (D) day 5 or (E) day 60. ***P < 0.0001. Scale bar in B = 20 µm.
Figure 1.
CD45+ immune responses are suppressed in the early post-injury period in Il1r KO compared to wild-type mice. (A) The density of CD45+ immune cell responses (magenta, white arrows) increases near denervated taste buds at day 2 post-injury in wild-type mice. Taste buds are visualized with the type I taste cell marker, NTPdase2 (green), which also labels nerve fibers in the papilla (open arrows). CD45+ immune responses return to baseline at day 5 post-injury and remain at low levels at day 60 in wild-type papillae with regenerated, functional taste buds. (B) In Il1r KO mice, few CD45+ immune cells occupy taste papillae at any post-injury period tested. (C) CD45+ pixels were thresholded, expressed as the percent / standard area, and summed at each period (n=3-4 mice/group). There was a significant increase in the mean (+SEM) CD45+ responses at day 2 post-injury compared to sham wild-type mice or axotomized Il1r KO mice. Bonferroni post-tests revealed no significant differences in CD45+ immune responses among groups at (D) day 5 or (E) day 60. ***P < 0.0001. Scale bar in B = 20 µm.
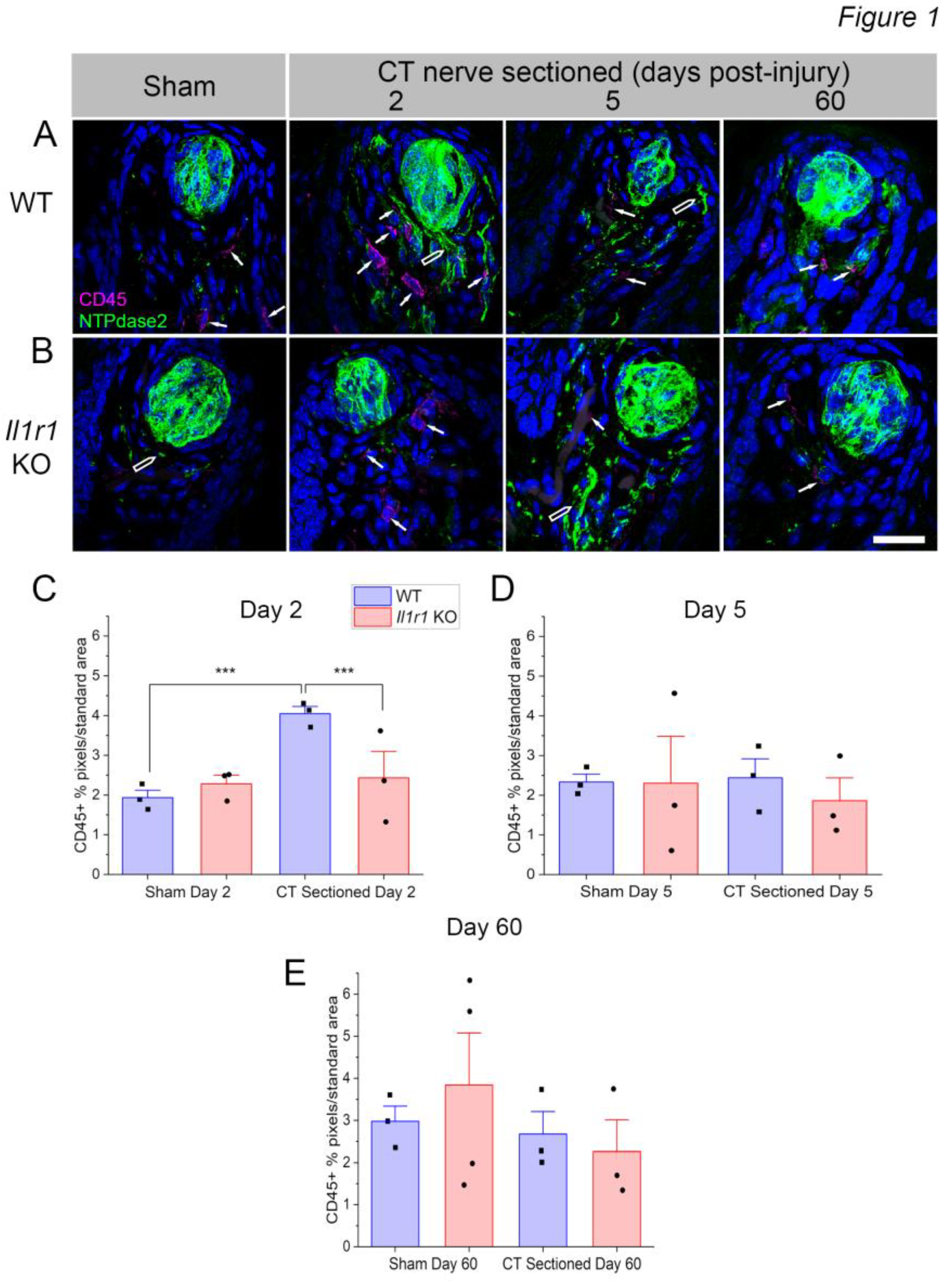
Figure 2.
CD68+ macrophage density increases at day 2 post-sectioning in wild-type but not Il1r KO mice. (A) Many CD68+ macrophages (magenta, white arrows) are present in wild-type taste papillae at day 2 after axotomy compared to sham controls. Occasionally, macrophages infiltrated denervated taste buds (inset on right). (B) Few macrophages were observed in Il1r KO papillae even after axotomy. (C) Quantification of CD68+ pixels/standard area demonstrates a significant increase in the mean (+SEM) level of macrophage responses at day 2 post-injury in wild-type mice. N = 3-4 mice / group. **P < 0.001. Scale bar in B = 20 µm.
Figure 2.
CD68+ macrophage density increases at day 2 post-sectioning in wild-type but not Il1r KO mice. (A) Many CD68+ macrophages (magenta, white arrows) are present in wild-type taste papillae at day 2 after axotomy compared to sham controls. Occasionally, macrophages infiltrated denervated taste buds (inset on right). (B) Few macrophages were observed in Il1r KO papillae even after axotomy. (C) Quantification of CD68+ pixels/standard area demonstrates a significant increase in the mean (+SEM) level of macrophage responses at day 2 post-injury in wild-type mice. N = 3-4 mice / group. **P < 0.001. Scale bar in B = 20 µm.
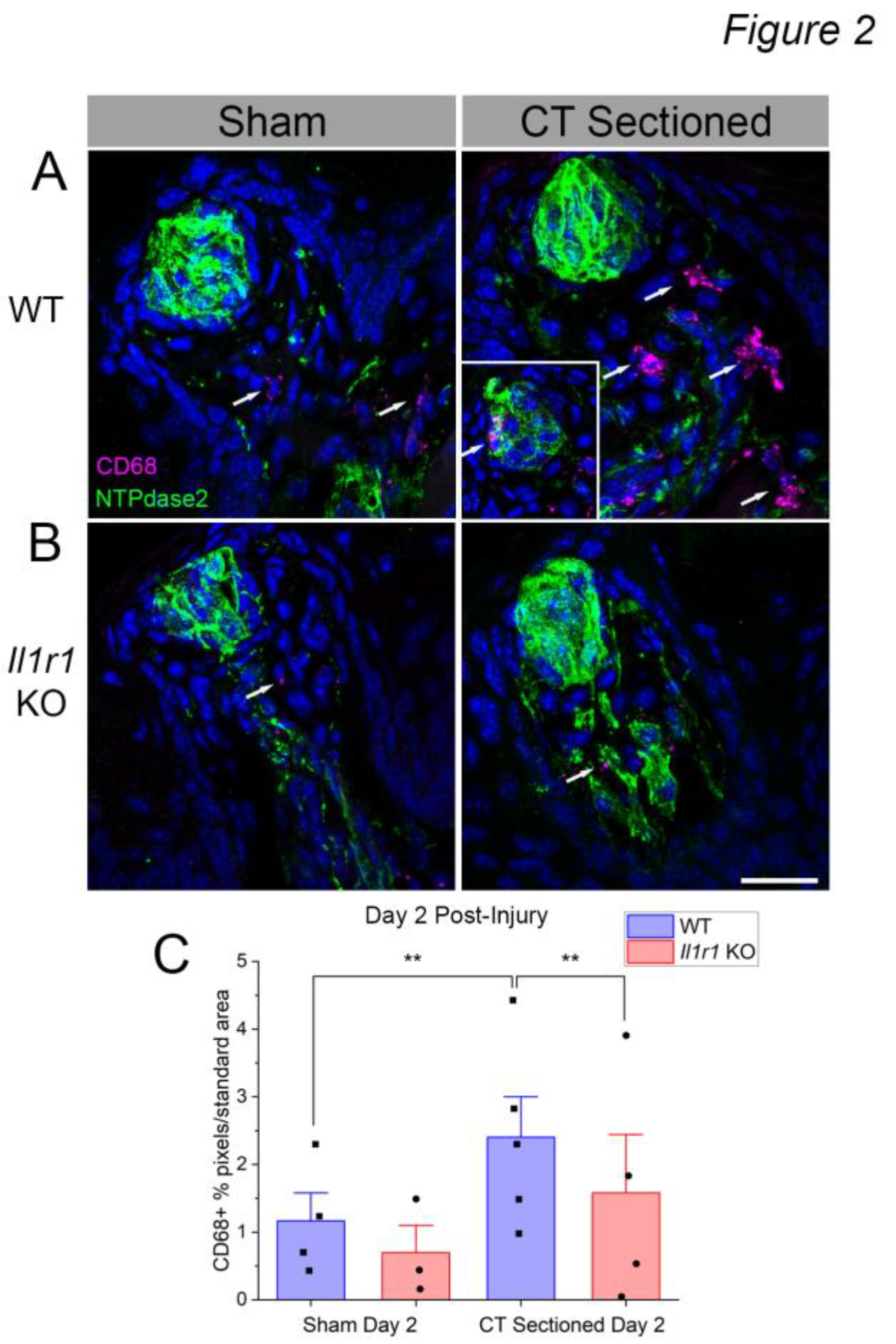
Figure 3.
CD68+CD206+ M2-like macrophage responses to nerve injury predominate in wild-type mice. In these analyses, we counted double-positive macrophages, associated with a DAPI+ nucleus, rather than quantifying immunopositive pixels as in Fig. 1-2. (A) Taste tissue , including fungiform papillae, taste buds (dotted white lines) and lamina propria, was populated by few macrophages in sham-sectioned wild-type mice. Double-positive macrophages were prominent in these regions in wild-type mice at day 2 after axotomy (white arrows). (B) At the same time, few macrophages were present in sham- or nerve-sectioned Il1r KO mice. (C) CD68+ CD206+ macrophages subsided by day 5 post-injury in wild-type mice, but remained elevated particularly at the base of papillae and in the lamina propria. The mean (+SEM) number of M2-like macrophages was significantly elevated in wild-type compared to sham-sectioned wild-type and axotomized Il1r KO mice at both (D) day 2 and (E) 5 (n=3 mice/group). **P < 0.001; ***P < 0.001. Scale bar in B = 20 µm.
Figure 3.
CD68+CD206+ M2-like macrophage responses to nerve injury predominate in wild-type mice. In these analyses, we counted double-positive macrophages, associated with a DAPI+ nucleus, rather than quantifying immunopositive pixels as in Fig. 1-2. (A) Taste tissue , including fungiform papillae, taste buds (dotted white lines) and lamina propria, was populated by few macrophages in sham-sectioned wild-type mice. Double-positive macrophages were prominent in these regions in wild-type mice at day 2 after axotomy (white arrows). (B) At the same time, few macrophages were present in sham- or nerve-sectioned Il1r KO mice. (C) CD68+ CD206+ macrophages subsided by day 5 post-injury in wild-type mice, but remained elevated particularly at the base of papillae and in the lamina propria. The mean (+SEM) number of M2-like macrophages was significantly elevated in wild-type compared to sham-sectioned wild-type and axotomized Il1r KO mice at both (D) day 2 and (E) 5 (n=3 mice/group). **P < 0.001; ***P < 0.001. Scale bar in B = 20 µm.
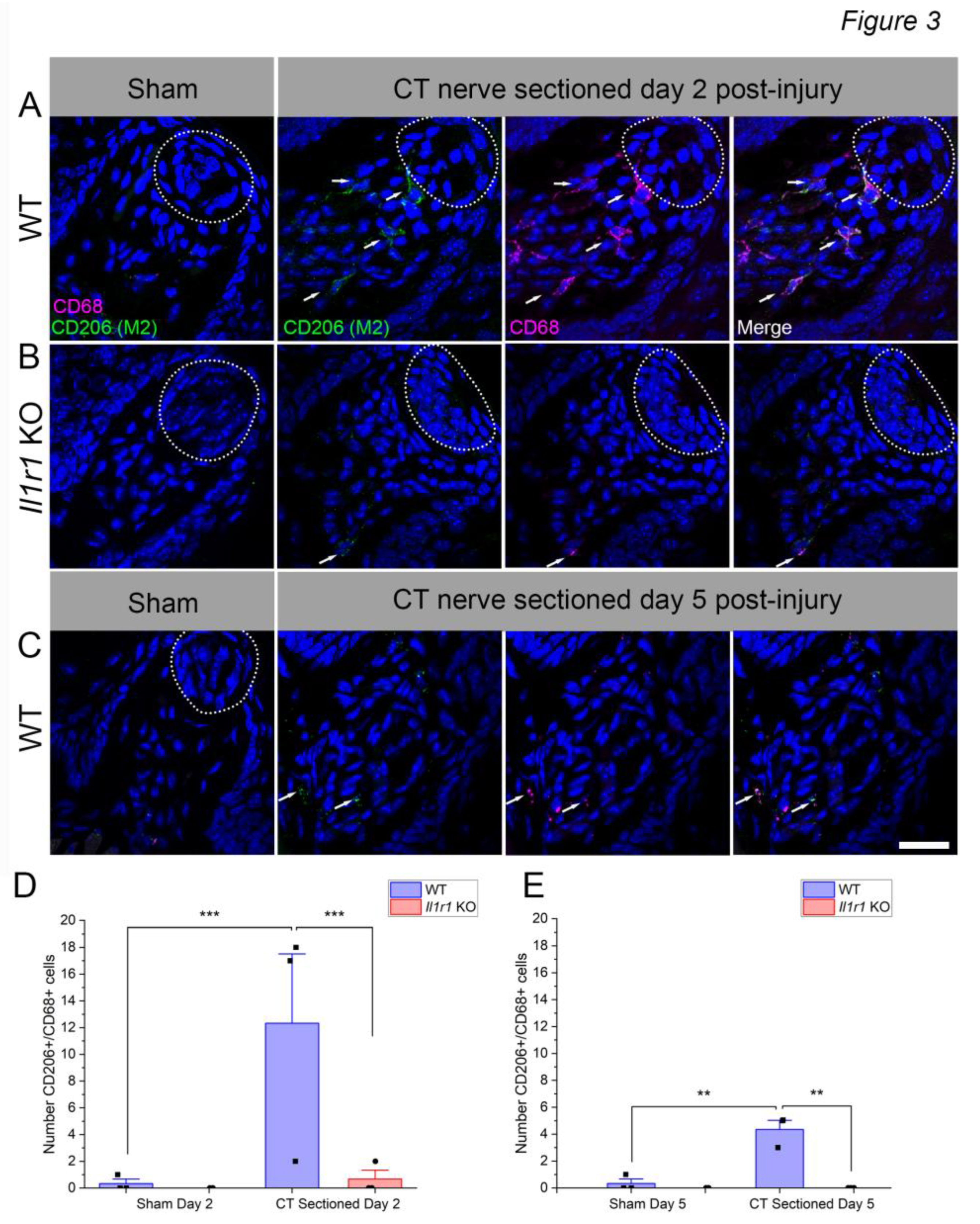
Figure 4.
Taste buds degenerate in both wild-type and Il1r KO mice at day 10 post-injury. Lingual epithelia were enzymatically and manually dissected and taste buds visualized with Keratin 8 (K8). Taste buds are shown in sham-sectioned (left) or CT nerve injured (right) lingual epithelia. Dotted lines in A, upper left, show regions in which K8+ taste buds were counted anterior to the intermolar eminence, and on the right (sectioned) and left (intact) sides of the tongue. We counted taste buds on the (A, C) dorsal epithelium and (B, D) ventral side of the tongue tip, which has a high density of taste buds. Note the reduced density of green taste buds on the right (axotomized) dorsal and ventral tongues of both strains. (E) K8+ taste buds on the right side were expressed relative to the left (intact) side to calculate percent degeneration. 100% (horizontal dotted line) represents equal numbers of taste buds on the two sides of the tongue, or no degeneration. There were no significant differences between groups, as detailed in the text. N=3 mice/group. Scale bar in D = 1 mm.
Figure 4.
Taste buds degenerate in both wild-type and Il1r KO mice at day 10 post-injury. Lingual epithelia were enzymatically and manually dissected and taste buds visualized with Keratin 8 (K8). Taste buds are shown in sham-sectioned (left) or CT nerve injured (right) lingual epithelia. Dotted lines in A, upper left, show regions in which K8+ taste buds were counted anterior to the intermolar eminence, and on the right (sectioned) and left (intact) sides of the tongue. We counted taste buds on the (A, C) dorsal epithelium and (B, D) ventral side of the tongue tip, which has a high density of taste buds. Note the reduced density of green taste buds on the right (axotomized) dorsal and ventral tongues of both strains. (E) K8+ taste buds on the right side were expressed relative to the left (intact) side to calculate percent degeneration. 100% (horizontal dotted line) represents equal numbers of taste buds on the two sides of the tongue, or no degeneration. There were no significant differences between groups, as detailed in the text. N=3 mice/group. Scale bar in D = 1 mm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



