1. Introduction
Teleost fish represent important vertebrate models for elucidating the developmental and regulatory mechanisms underlying skeletal formation, providing insights into both evolutionary biology and applied fields such as aquaculture and biomedicine (Lakra et al., 2011). In vitro cell lines are indispensable experimental tools and have been extensively applied in studies of fish immunology, physiology, genetics, and related disciplines (Zhang et al., 2024; Jing et al., 2023). Fish cell culture technology originated in the 1960s, marked by the establishment of the first fish cell line, RTG-22, derived from rainbow trout (Oncorhynchus mykiss) gonadal by Wolf and Quimby (Wolf & Quimby, 1962). Since then, continuous methodological advances have rendered fish cell culture systems fundamental to modern fish biology research. Currently, fish cell lines have been reported from various tissues and organs, including the liver, kidney, brain, fin, skin, and gonads (Hongli et al., 2023; Fu et al., 2015; Jing et al., 2016; Wang et al., 2017; Wei et al., 2018; Zhang et al., 2020).
Despite these advances, bone tissue cell lines, especially those exhibiting stable osteogenic properties, remain extremely limited (Marques et al., 2007; Rafael et al., 2010; Vijayakumar et al., 2013). The fish bone matrix is highly mineralized and collagen-rich, which complicates tissue digestion and cell isolation because efficient dissociation must be balanced against preservation of cellular integrity (Witten et al., 2017). In addition, osteogenesis-related growth factors and signaling pathways display pronounced species specificity in fish, necessitating careful optimization of culture conditions and medium composition (Witten & Huysseune, 2009). Maintaining osteoblastic differentiation potential and functional stability during long-term in vitro culture therefore remains a major technical challenge. To date, osteoblast culture systems have been reported for only a limited number of fish species, such as blunt snout bream (Megalobrama amblycephala) (Guan et al., 2018) and zebrafish (Danio rerio) (Yan et al., 2024). However, cell lines derived from local strains or ecotypes remain particularly scarce, making it difficult to meet the demands of in-depth studies on the molecular mechanisms of osteogenesis.
Bone cells primarily include osteoblasts and osteocytes (Fu et al., 2004). Among these cell types, osteoblasts are the principal effector cells responsible for bone matrix synthesis and mineralization, thereby playing a central role in bone formation, remodeling, and metabolic homeostasis (Long, 2012; Parfitt, 2001). Osteogenesis is tightly regulated by a complex network of transcription factors and signaling pathways and is closely associated with the cellular microenvironment (Komori, 2006). Previous studies have demonstrated that bone tissue is generally maintained in a relatively hypoxic microenvironment, in which hypoxia inducible factor-1α (hif-1α) plays a crucial role in regulating osteoblast metabolism, survival, and differentiation (Chachami et al., 2013). These findings underscore the importance of systematically investigating osteoblast cell responses to hypoxic conditions using in vitro models.
Crucian carp (Carassius auratus), a member of the family Cyprinidae within the order Cypriniformes, is one of the most economically important aquaculture species in China (Du et al., 2021; Olsén & Bonow, 2023). This species is highly valued because of its rapid growth rate, strong environmental adaptability, and relatively stable genetic background. The Chongming crucian carp, a distinctive strain originating from the lower Yangtze River region, possesses considerable value for genetic resource conservation and applied biological research. However, to date, no studies have documented the establishment of a cell line from Chongming crucian carp. To address this knowledge gap, the present study aimed to establish a stable osteoblast cell line from Chongming crucian carp using an improved tissue explant culture method. The resulting cell line, designated Carassius auratus osteoblast cell (COBC), was systematically characterized in terms of morphology, chromosomal karyotype, biological characteristics, and the expression of osteogenesis-related genes. Furthermore, hypoxia treatment experiments were conducted to evaluate the applicability of COBC as an in vitro model for environmental stress studies. Overall, this study establishes a novel in vitro osteoblast cell model and provides a robust experimental platform for investigating skeletal development and osteogenic regulatory mechanisms in fish.
2. Materials and Methods
2.1. Ethics Statement
All animal experiments were approved by the Institutional Animal Care and Use Ethics Committee of Shanghai Ocean University (SHOU-DW-2024-054). All efforts were made to minimize the suffering of the animals. All experiments were performed in accor dance with relevant guidelines and regulations.
2.2. Primary Culture and Subculture
The Chongming crucian carp (18-20 g) used in this study were obtained from Chongming District, Shanghai. Osteoblast cells were cultured using tissue culture. After 48 h of acclimation, 4-month-old fish were disinfected by immersion in 0.01% potassium permanganate for 20 min. The fish were subsequently anesthetized with MS-222 for 5 min and transferred to a sterile operating platform, where the body surface was further disinfected with 95% ethanol. Using a sterile scalpel, the caudal fin was removed, and the tail was excised posterior to the cloacal region. The skin, muscle, and associated soft tissues were carefully removed with sterile forceps, retaining only the vertebral column and adjacent connective tissues. After washing the tissue blocks 3-5 times with PBS, the samples were minced into approximately 2 mm3 fragments using ophthalmic scissors. The minced tissues were moistened with fetal bovine serum (FBS, Gibco, USA) and transferred into 25 cm2 culture bottle (Thermo Scientific, USA), with roughly 20 fragments placed per flask. Cell information, including tissue source, date, and passage number, was labeled on the side of each flask. The flasks were initially placed upside down in a 28 ℃, 5% CO2 incubator for 3-4 h to facilitate tissue attachment and were then gently inverted. One milliliter of Leibovitz’s L-15 (L-15, Gibco, USA) complete medium (containing 1% penicillin-streptomycin, 20% FBS, 25 ng/mL EGF, and 25 ng/mL bFGF) was added to support continued culture. Cell adhesion and proliferation were monitored daily. The medium was refreshed every 2-3 days by replacing one-third of the volume, and cells were designated as the P0 generation once they reached approximately 80% confluence.
Once the primary cells had migrated from the explants and reached ≥80% confluence, subculture procedures were initiated. The cultures were gently rinsed twice with 2 mL of PBS to remove residual medium. The cells were digested with 0.5 mL of 0.25% trypsin (Thermo Scientific, USA) for approximately 5 min, and trypsinization was terminated by adding an equal volume of fresh medium, followed by centrifugation at 1200 rpm for 5 min. The resulting cell pellet was resuspended and seeded into two new culture flasks to generate the P1 cell population. Cell morphology was assessed daily, and the medium was renewed every other day. Upon formation of a confluent monolayer, cellular morphology was documented using an inverted fluorescence microscope (DMi3000, Leica).
2.3. Cryopreservation and Recovery of Cells
After selecting actively proliferating cells, they were resuspended following trypsin digestion, and an appropriate volume of serum-free cryoprotective medium was added. The resulting suspension was gently pipetted to achieve homogeneity, transferred into pre-labeled cryovials, and promptly sealed. The cryovials were first cooled at -80 ℃ overnight and subsequently transferred to liquid nitrogen for long-term preservation.
For recovery, the cryovial was removed from liquid nitrogen and rapidly thawed in a 37 ℃ water bath with gentle agitation to facilitate uniform thawing. The thawed cell suspension was immediately transferred into L-15 complete medium and gently mixed. The cells were centrifuged at 1200 rpm for 5 min, after which the pellet was resuspended in 2 mL of fresh complete medium. Medium replacement was performed after 24 h once cell adhesion was confirmed.
2.4. Investigation of Optimal Growth Conditions for COBCs
To determine the optimal culture conditions for COBCs, a CCK-8 assay (Sigma) was employed to assess the effects of different medium components on the proliferation of P10 cells. In 96-well plates, 2 × 103 cells were seeded per well, with three replicate wells included for each treatment group. The effects of 5%, 10%, 15%, and 20% FBS supplementation in M-199 and L-15 complete media were evaluated. Cell proliferation was monitored at 24, 48, 72, and 96 h to identify the optimal FBS concentration.
2.5. Chromosome Analysis of COBCs
COBCs at the 15th passage were cultured for 2 days following subculture. Subsequently, 2 mL of fresh medium containing 1 μg/mL colchicine was added, and the cultures were incubated for up to 8-12 h to arrest cells in metaphase. After trypsin digestion and resuspension, the cells were processed for chromosome preparation. Chromosomes were examined under a high-power microscope, and chromosome numbers were counted and documented.
2.6. Biological Characterization of COBCs
At passage 20 (P20), COBCs were seeded into 12-well plates and, upon reaching approximately 80% confluence, were stained and imaged using alkaline phosphatase (ALP) staining (Beyotime, Shanghai, China), Alizarin Red S staining (Sigma), Von Kossa mineralization assay (Beyotime), and Giemsa staining (Solarbio, Beijing, China). To quantify osteocalcin levels in P16 cells, a Fish osteocalcin (BGP/OCN) ELISA kit (Shanghai win-win Biotechnology Co., Ltd., China) was employed. Following color development of the antibody-antigen-enzyme complex, absorbance at 450 nm was measured using a microplate reader, and the BGP content were determined based on a standard curve.
2.7. Hypoxic Stress in COBCs
Stable P20 COBCs were seeded into culture dishes and exposed to hypoxic conditions for subsequent analyses. Based on preliminary experiments and previous reports, cells were incubated under hypoxic conditions (1% O2 and 99% N2) for 0, 12, and 24 h, followed by a 24 h reoxygenation period (r24 h) (Wang et al., 2023; Xu et al., 2024). All experiments were performed in triplicate. Samples were collected for morphological observation, gene expression profiling, and enzyme activity determination. Samples designated for gene expression and enzyme activity analyses were immediately snap-frozen in liquid nitrogen and stored at -80 ℃ until use.
2.8. Antioxidant Enzyme Activity
Collected cell samples were homogenized in physiological saline at 60 Hz for 60-120 s. The homogenates were centrifuged at 320 × g for 5 min, and the resulting supernatants from each experimental group were collected for subsequent enzyme activity assays. Superoxide dismutase (SOD) and catalase (CAT) activities, glutathione (GSH) levels, and lactate dehydrogenase (LDH) content were quantified using commercial assay kits ((Nanjing Jiancheng Biochemical Company, Nanjing, China) and measured with a microplate reader.
2.9. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR)
Total RNA was extracted using TRIzol reagent (Invitrogen, USA), and RNA concentration and purity were determined with a NanoDrop 2000 (Thermo Fisher Scientific). The cDNA was synthesized using the PrimeScript™ RT Reagent Kit with gDNA Eraser (TaKaRa, Japan), with
β-actin serving as the internal reference gene. The primers were designed based on crucian carp cDNA sequences using Primer Premier 5.0 (Premier Biosoft, USA) and synthesized by Sangon Biotech (Shanghai) Co., Ltd. (
Table 1).
The qRT-PCR was performed using SYBR Premix Ex Taq™ II (TaKaRa, Japan) in a total reaction volume of 20 μL, containing 10 μL SYBR Premix, 0.8 μL each of forward and reverse primers, 1 μL cDNA template, and 7.4 μL nuclease-free water. The amplification protocol consisted of an initial denaturation at 94 ℃ for 30 s, followed by 40 cycles of denaturation at 94 ℃ for 10 s, annealing for 30 s, and extension at 72 ℃ for 30 s. All reactions were conducted using a CFX96 Touch real-time PCR fluorescence quantification instrument (Bio-Rad, USA). Relative gene expression levels were calculated using the 2-ΔΔCT method following normalization to β-actin.
2.10. Statistical Analysis
The results are reported as mean ± standard deviation (Mean ± SD). In addition, SPSS 25.0 was used for one-way ANOVA of the experimental data and Duncan’s post-hoc test, with p < 0.05 indicating statistically significant differences. Graphics were created using GraphPad Prism 9.5 (GraphPad Software, USA).
3. Results
3.1. Primary Culture and Subculture Observation of COBCs
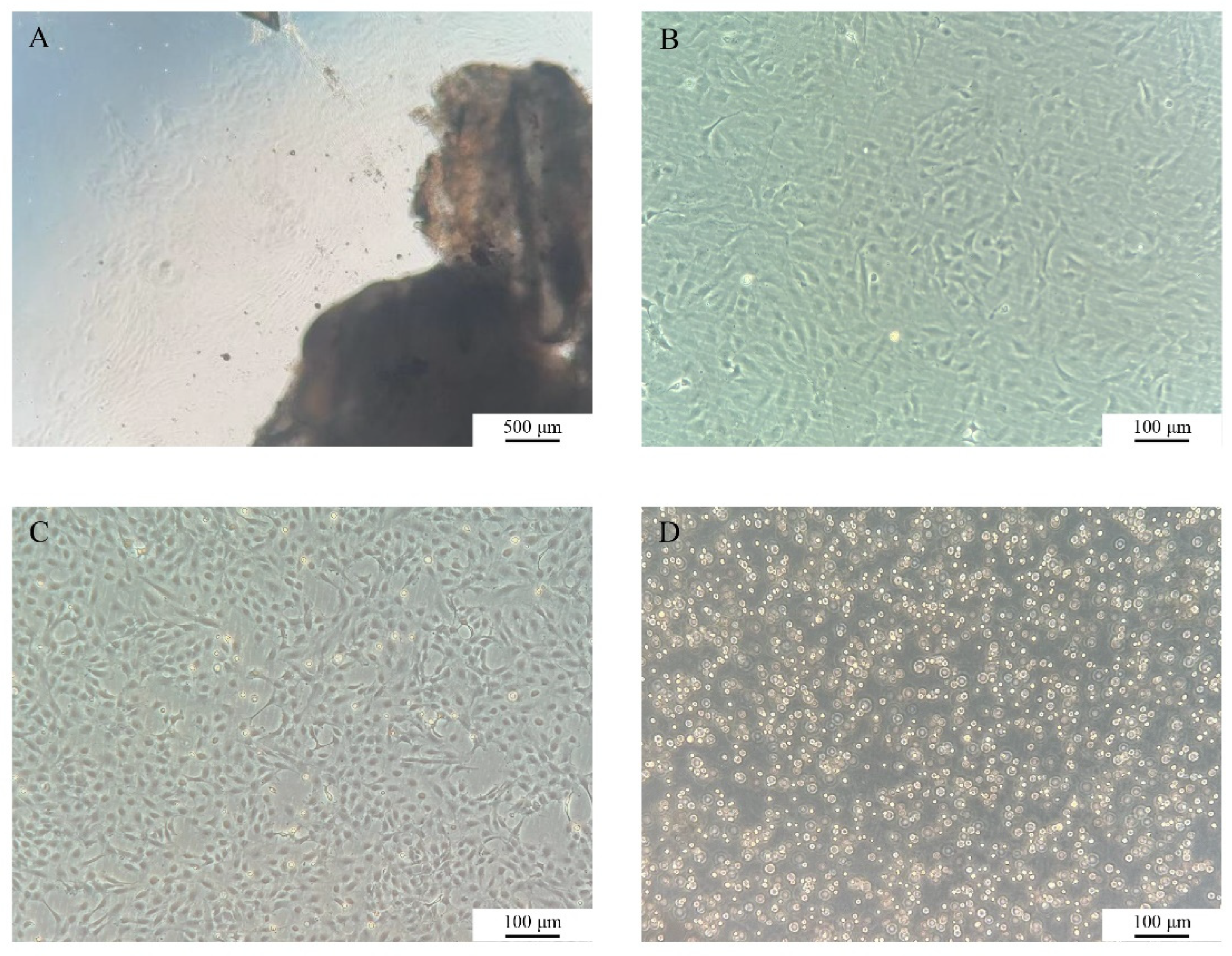
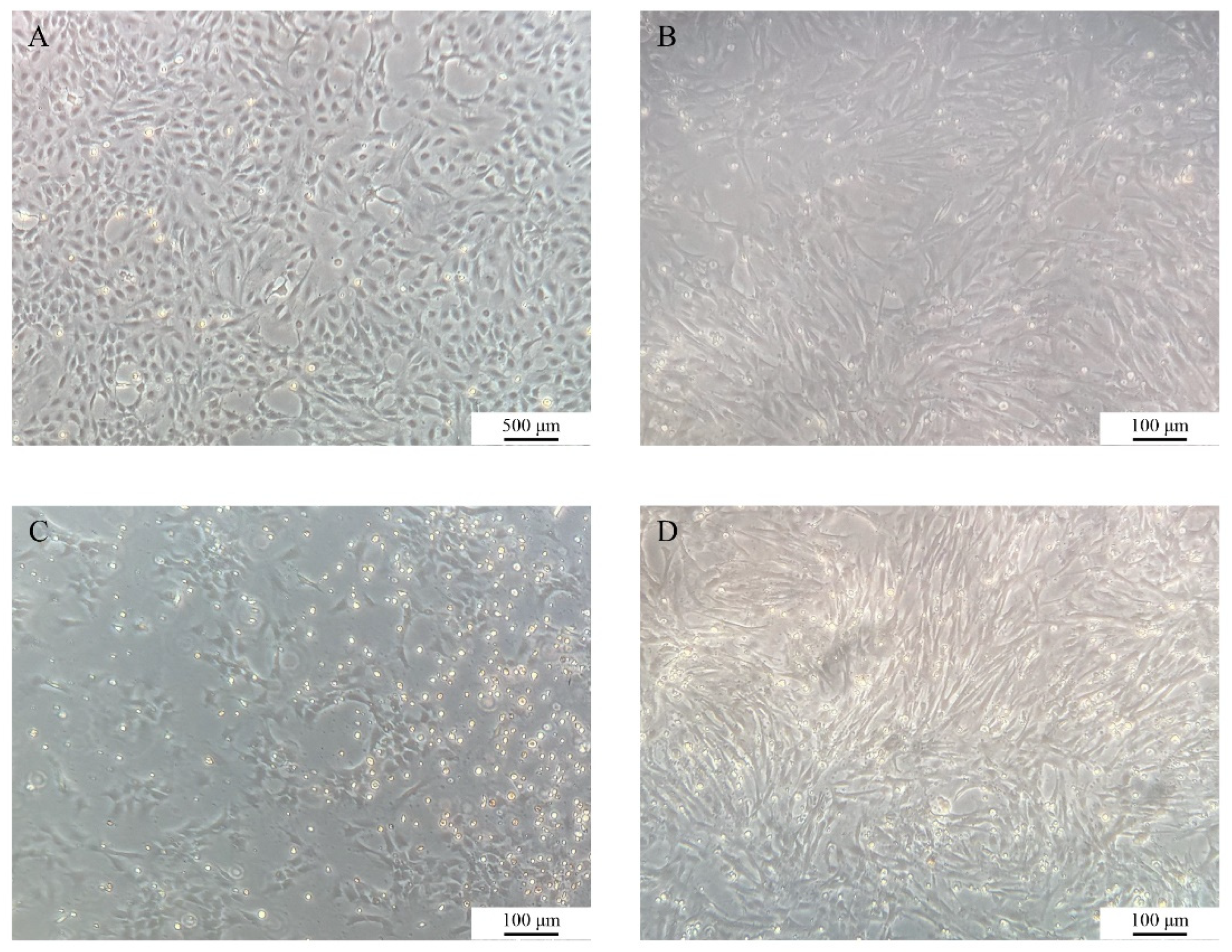
The growth dynamics of COBCs were systematically monitored under an inverted fluorescence microscope (
Figure 1). At approximately 3 days after primary culture, cells began to migrate outward from the margins of the tissue, displaying predominantly spindle-shaped or polygonal morphologies. By day 7, a distinct proliferative halo had developed around the tissues (
Figure 1A). Following subculture, the cells exhibited an accelerated proliferation rate while maintaining a stable morphology (
Figure 1B). Subculturing was performed every 2-3 days, and the COBC has since been stably propagated to the P37 (
Figure 1C). After cryopreservation and subsequent thawing, the cells exhibited good post-thaw recovery and retained normal morphology and proliferative capacity (
Figure 1D).
3.2. Effects of Culture Medium Components on the Growth of COBCs
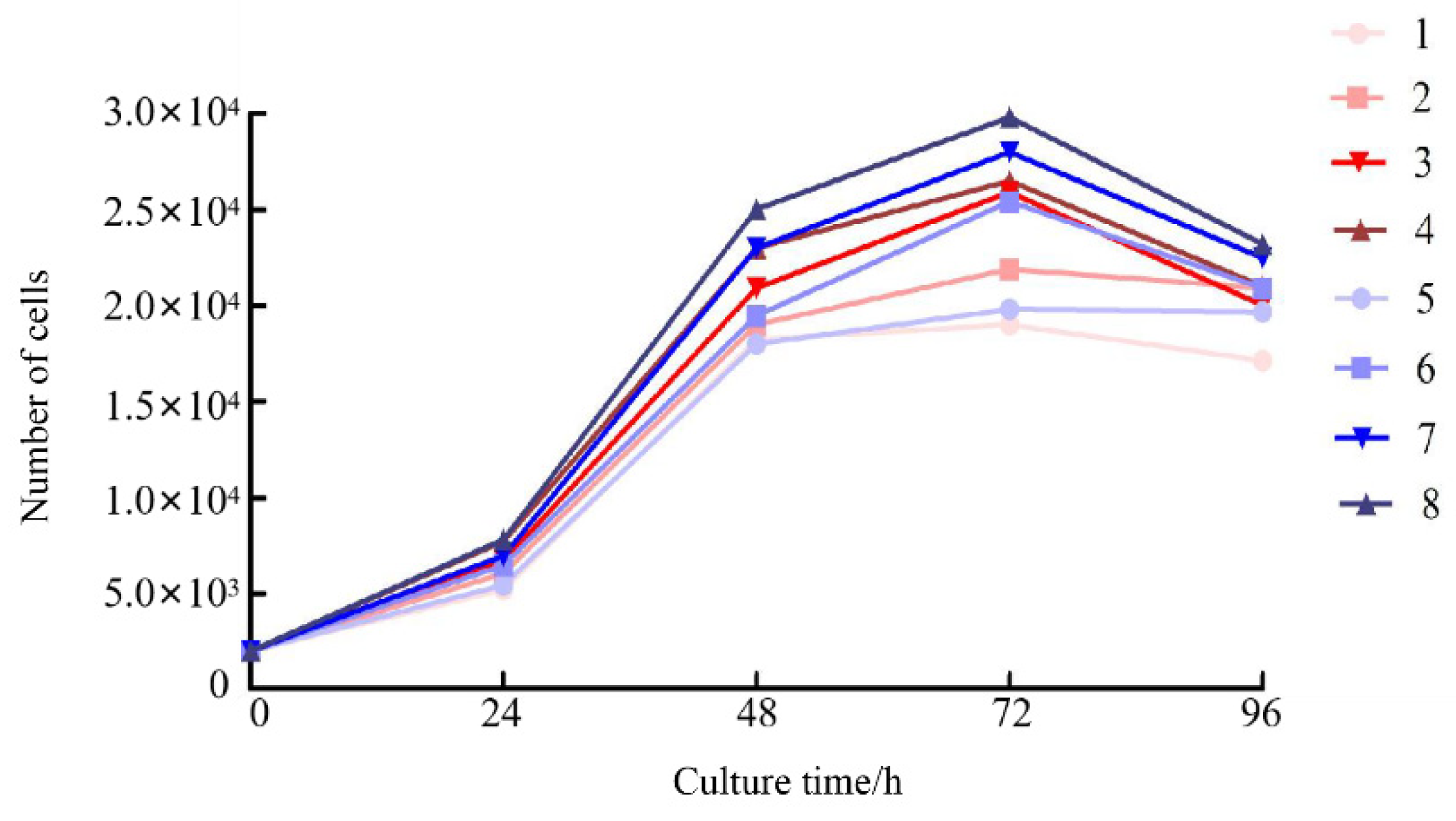
Results from the CCK-8 assay revealed marked differences in cell proliferation across FBS concentrations (20%, 15%, 10%, and 5%) in both M-199 and L-15 media (
Figure 2). The highest proliferative activity occurred in cultures supplemented with 20% FBS, whereas 5% FBS resulted in the lowest proliferation. Overall, cell abundance was consistently greater in L-15 medium than in M-199 medium under equivalent serum concentrations. In both media, the growth kinetics of COBCs were comparable, with cell numbers peaking at 48 h and subsequently declining slightly, yet remaining above baseline levels (
Figure 2). These results indicate that L-15 medium supplemented with 20% FBS provides the most favorable conditions for the proliferation of COBCs.
3.3. Chromosome Analysis
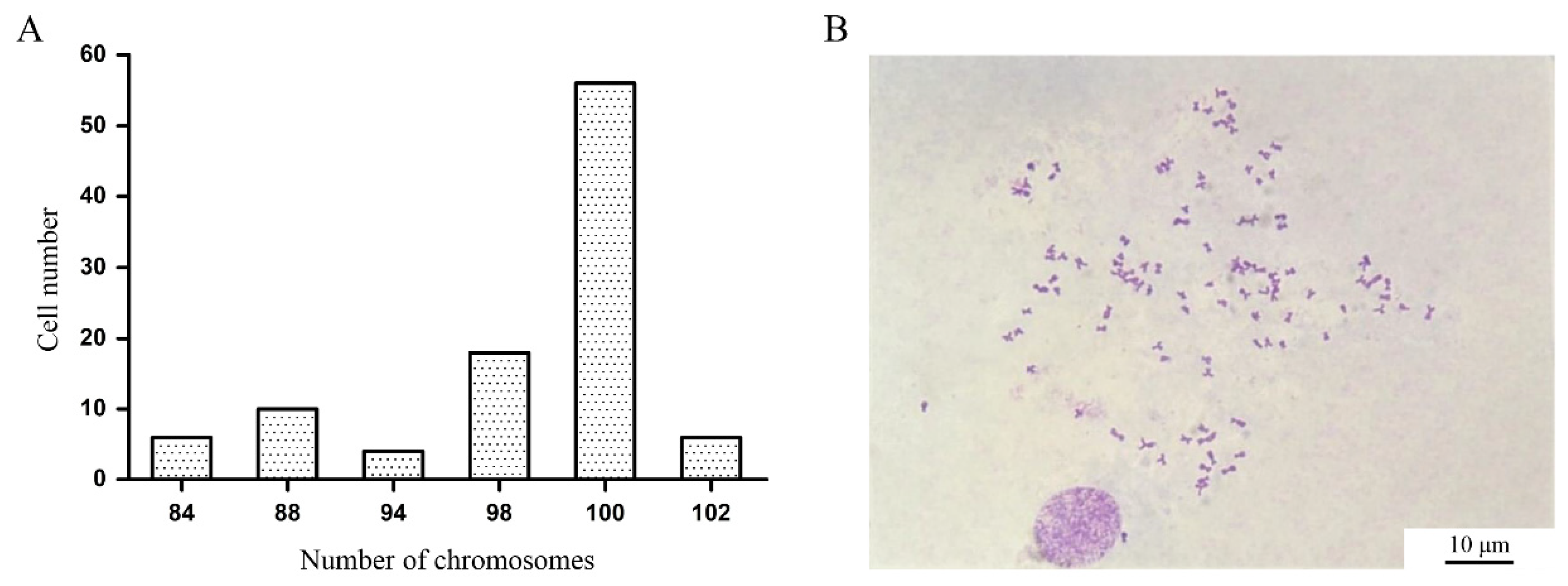
After colchicine treatment, chromosome spreads were prepared from P10 COBCs and examined. Among the 100 metaphase cells analyzed, chromosome numbers ranged from 92 to 102 (
Figure 3A). Notably, 56% of the metaphase cells exhibited 100 chromosomes, indicating that the COBCs originates from diploid Chongming crucian carp and possesses a modal chromosome number of 100 (
Figure 3B).
3.4. Biological Characterization of the COBCs
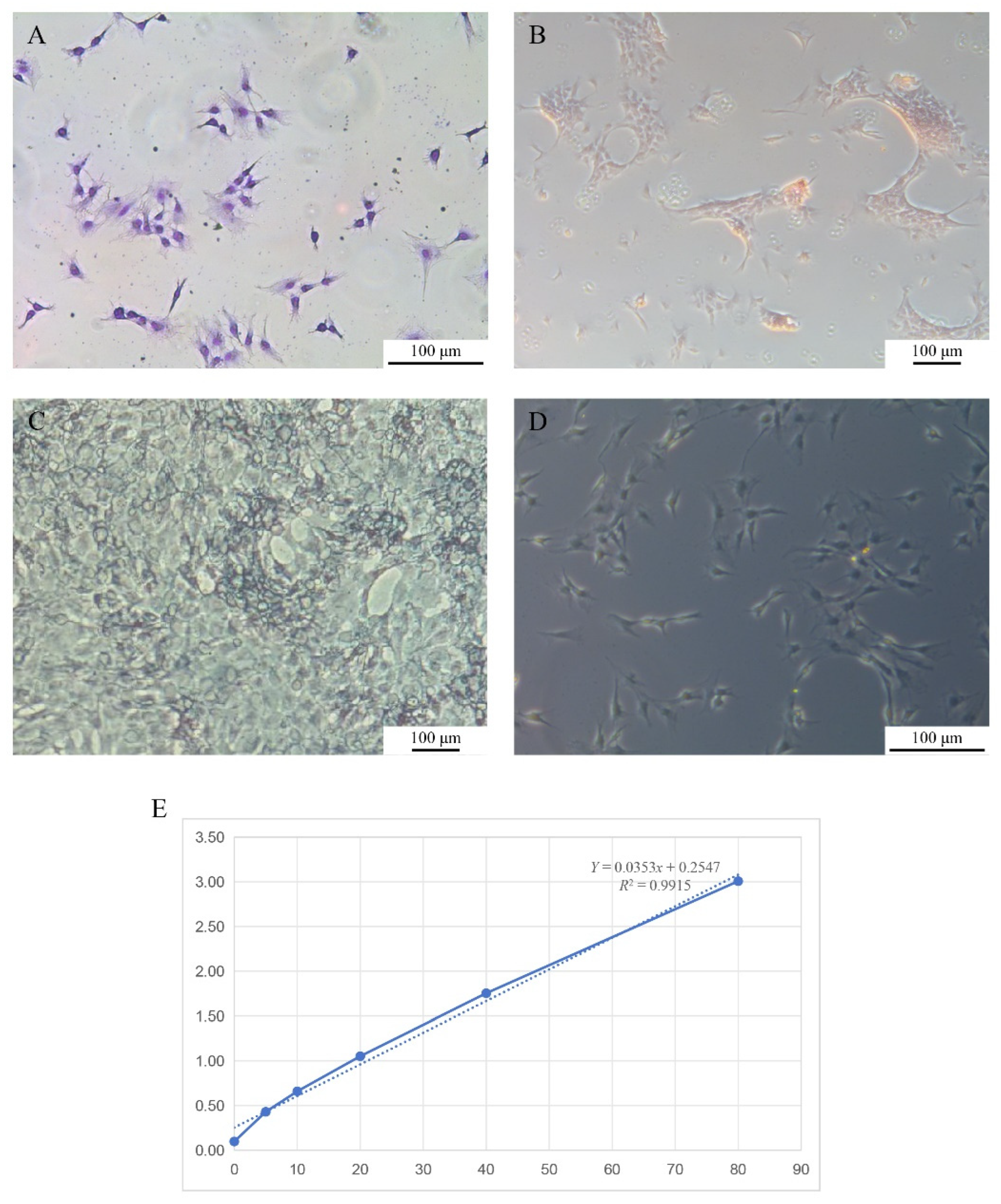
Giemsa staining revealed that the nuclei were dark blue with distinct nucleoli, which were predominantly centrally positioned within the cells (
Figure 4A). Alizarin Red S staining demonstrated that the long-term cultured COBCs formed focal aggregates and produced characteristic red-stained calcified nodules (
Figure 4B). In ALP staining, positive cells exhibited prominent blue-violet cytoplasmic precipitates (
Figure 4C). Von Kossa staining further showed that the cells were predominantly spindle-shaped or polygonal and generated abundant black mineralized nodules (
Figure 4D).
Using a fish BGP ELISA, absorbance at 450 nm was recorded and used to construct a standard curve (
Figure 4E). The average BGP content calculated from the standard curve was 36,884 ng/L.
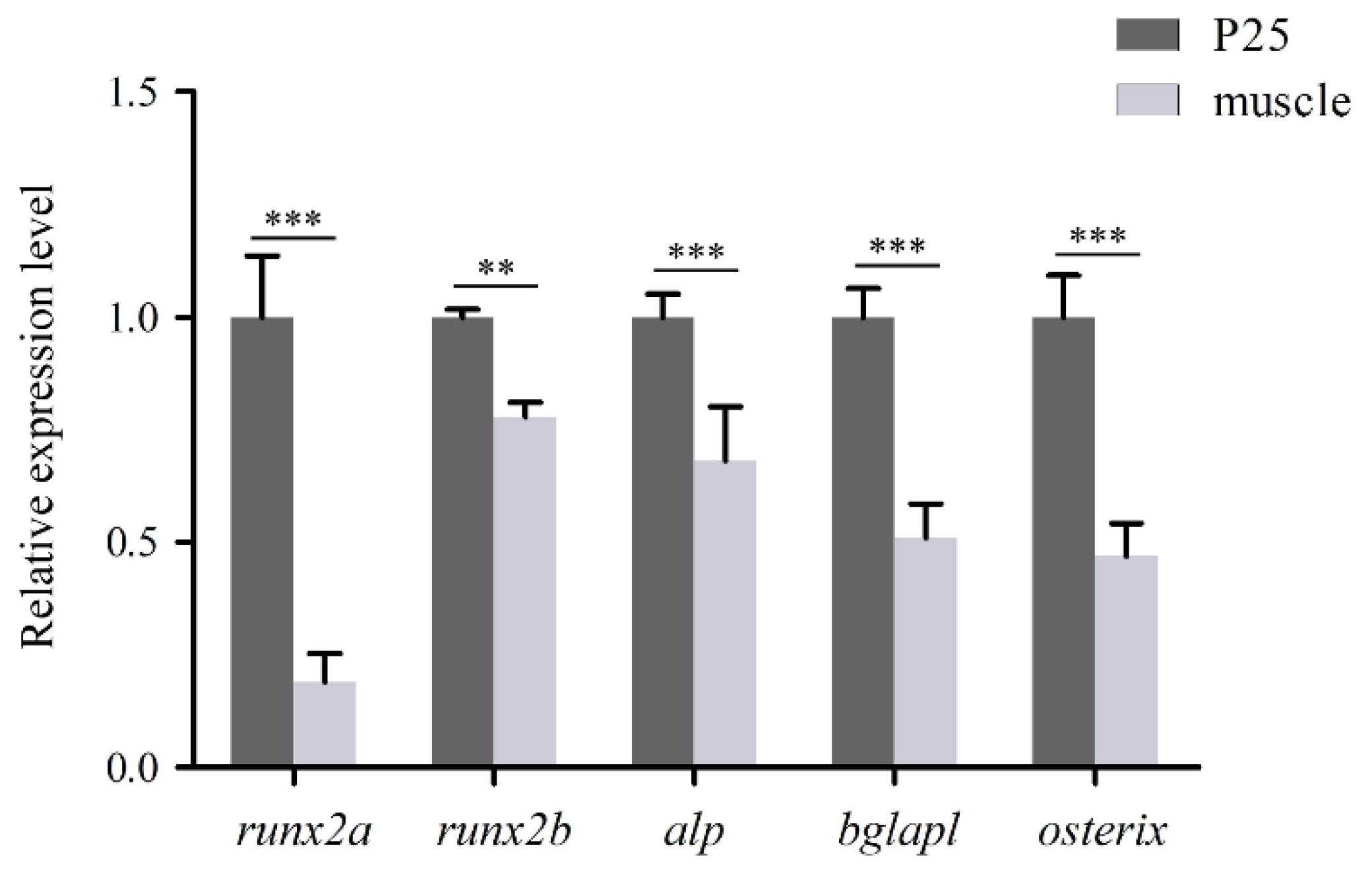
3.5. Gene Expression Analysis
The qPCR analysis demonstrated that all five genes-
runx2a,
runx2b,
alp,
bglapl, and
osterix-were detectable in COBCs. Among these genes,
runx2a showed the highest relative expression, followed by
bglapl and
osterix. The expression of all five genes in COBCs was significantly higher than in in vivo muscle tissue (
p < 0.01,
Figure 5).
3.6. Effects of Hypoxic Stress on COBCs
Compared with the normoxic control (0 h) (
Figure 6A), COBCs exhibited evident apoptotic features following 12 h of hypoxic exposure, and the extent of apoptosis increased markedly after 24 h of hypoxia (
Figure 6B, C). Following 24 h of reoxygenation, cell morphology and cell density were partially restored compared with the hypoxia groups, with fibroblast cells emerging as the dominant cell type (
Figure 6D).
3.7. Gene Expression Responses of COBCs Under Hypoxia and Reoxygenation
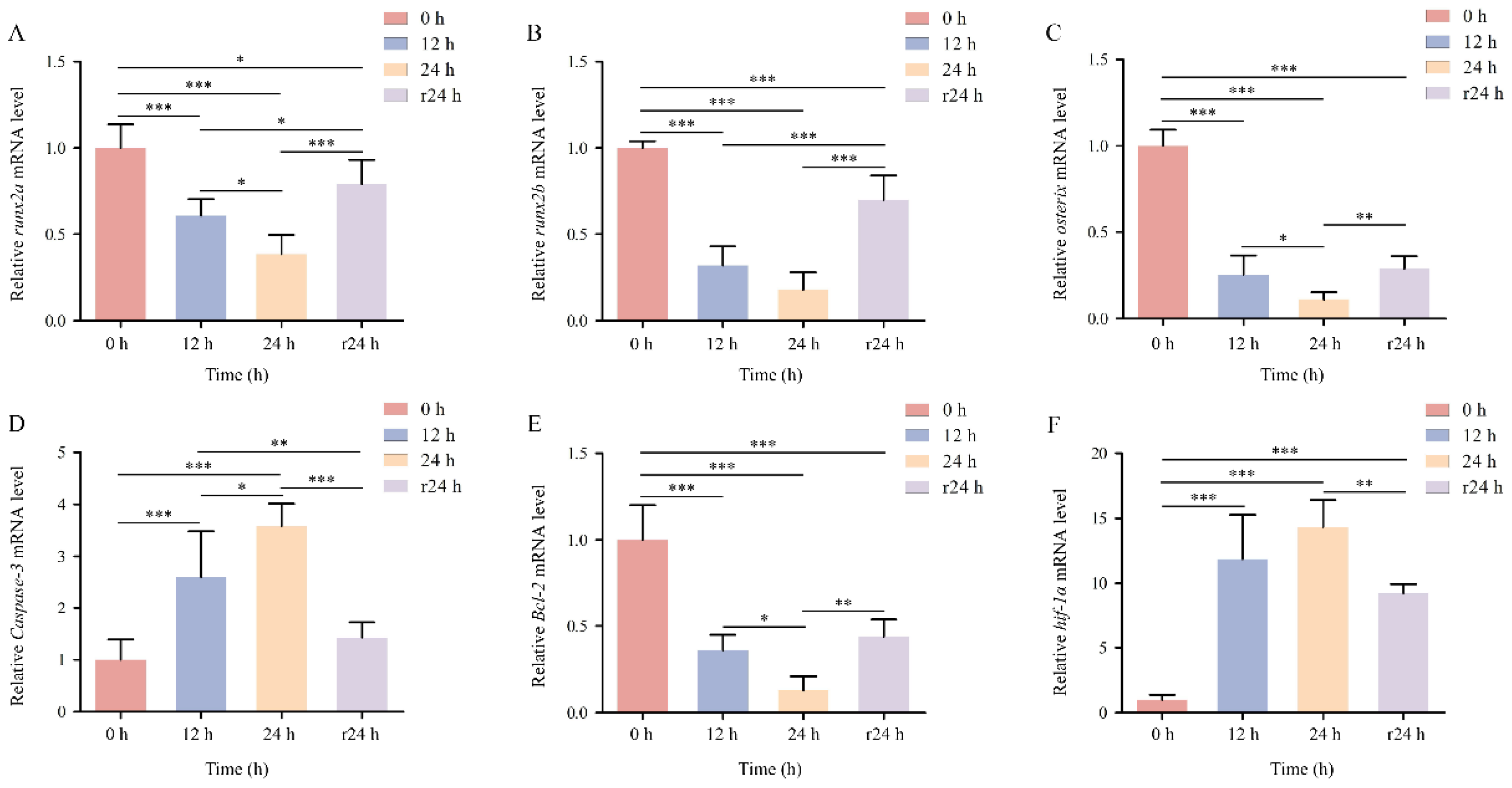
The mRNA expression levels of osteogenesis-related genes
runx2a,
runx2b, and
osterix in COBCs were significantly downregulated under acute hypoxic conditions compared with the normoxic control at all time points. The lowest expression levels were observed after 24 h of hypoxia (
p < 0.001). After 24 h of reoxygenation, the expression of these genes showed partial recovery but remained significantly lower than control levels (
p < 0.05,
p < 0.01;
Figure 7A-C).
The hypoxia-related gene
Bcl-2 displayed a similar expression pattern, characterized by suppression under hypoxia and partial recovery following reoxygenation (
Figure 7E). In contrast, the expression levels of
Caspase 3 and
hif-1α increased progressively with prolonged hypoxic exposure (
p < 0.001;
Figure 7D, F). After 24 h of reoxygenation,
Caspase 3 expression returned to baseline levels comparable to the normoxic control (
p > 0.05;
Figure 7D), whereas
hif-1α expression remained significantly elevated (
Figure 7F).
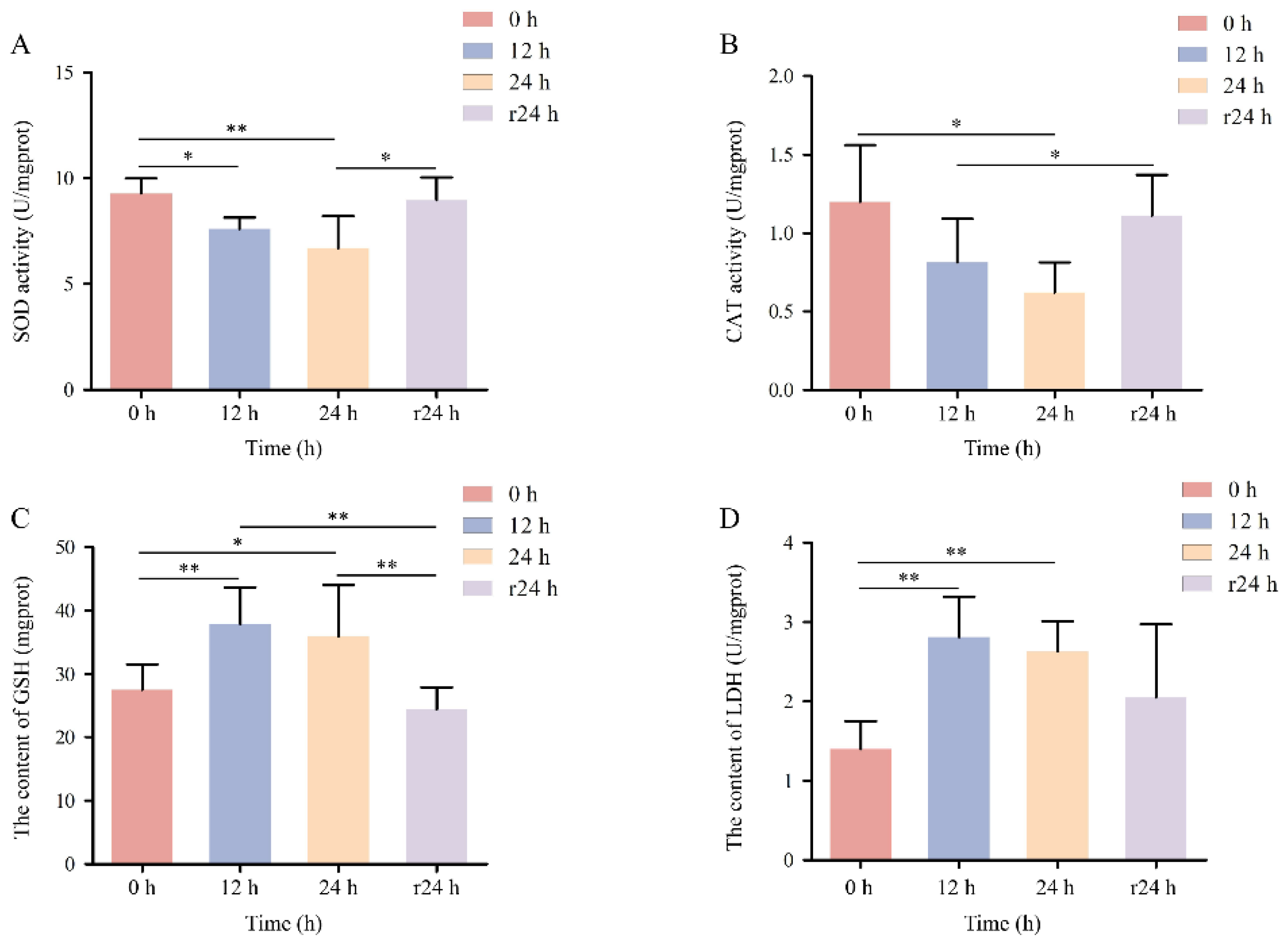
3.8. Enzyme Activity of COBCs
Enzyme activity assays demonstrated that the activities of SOD and CAT in COBCs decreased progressively with increasing hypoxia duration compared with the normoxic control (
p < 0.05). Following 24 h of reoxygenation, SOD and CAT activities recovered to levels that were not significantly different from those of the normoxic group (
p > 0.05;
Figure 8A, B). The content of GSH and LDH reached their highest levels after 12 h of hypoxic exposure and subsequently declined after 24 h of reoxygenation, ultimately returning to values comparable to those observed under normoxic conditions (
Figure 8C, D).
4. Discussion
The establishment of osteoblast cell lines in teleost fish is essential for dissecting the cellular and molecular mechanisms underlying skeletal development, metabolic regulation, and environmental adaptation (Farquharson, 2015). In contrast to mammals, in vitro osteogenic cell models in fish remain poorly developed, with particularly few reports describing cell lines derived from region-specific strains or ecotypes (Jing et al., 2023). This limitation has hindered comprehensive investigations into the evolutionary conservation of skeletal regulatory mechanisms and adaptive responses to environmental stressors. Here, we established and systematically characterized COBC, thereby providing an in vitro platform for exploring osteogenic cell biology and environmental responsiveness in teleosts. COBC displayed stable morphology, chromosomal integrity, and sustained expression of osteogenesis-related markers, demonstrating the maintenance of a differentiated osteoblastic phenotype during long-term in vitro culture. These characteristics are consistent with those reported for previously established osteoblast cell models in teleost fish (Guan et al., 2018; Yan et al., 2024). In this study, COBC was comprehensively characterized in terms of cellular morphology, proliferative behavior, osteogenic marker expression, and responses to hypoxia-reoxygenation stress. An tissue culture method was applied to establish COBC. This method creates a relatively stable microenvironment for cell migration and adhesion, thereby promoting efficient monolayer formation and improving both the success rate and efficiency of primary culture (Xiao et al., 2012). Morphological examination under an inverted microscope demonstrated that COBCs predominantly exhibited spindle-shaped and polygonal morphologies, which are characteristic features of osteoblast cells reported in various teleost species (Guan et al., 2018; Yan et al., 2024). These observations further confirm the osteogenic identity of the COBC.
Previous studies have demonstrated that long-term in vitro culture of fish cells may lead to chromosomal instability, particularly aneuploidy, as a consequence of culture stress and environmental influences (Bols et al., 1994). The COBCs established in this study maintained the same chromosome number as reported for crucian carp somatic cells even after multiple passages (Wang et al., 1982). These findings support the reliability and suitability of COBC as a stable long-term in vitro model for osteoblast studies. The composition of the culture medium and the concentration of serum are widely recognized as critical determinants of fish cell proliferation and growth in vitro (Gong et al., 2023). In this study, COBCs cultured in L-15 medium supplemented with 20% fetal bovine serum showed a significantly enhanced proliferative capacity compared with those maintained in M-199 medium. This difference may be attributed to the optimized osmotic balance and nutrient composition of L-15 medium, which is considered particularly suitable for cells derived from poikilothermic vertebrates (Rosa et al., 2010).
During the early phase of osteoblast differentiation, ALP is progressively upregulated and plays a critical role in extracellular matrix maturation. With further osteogenic maturation, matrix mineralization is initiated (Nelson et al., 2002). During the late differentiation stage, ALP activity gradually declines, whereas the expression of mineralization-associated markers, including BGP and osteopontin, reaches peak levels (Han, 2008). Consequently, ALP activity, matrix calcification, and mineralized nodule formation are widely recognized as key indicators of osteoblast functional status and differentiation stage (Huang et al., 2025). In the present study, COBCs showed robust positive staining for ALP activity, Alizarin Red, and von Kossa mineralization, demonstrating typical osteoblastic functionality and mineralization capacity. The concordant results obtained from multiple histochemical assays provide strong evidence that COBC exhibits the functional and phenotypic characteristics of mature osteoblasts (Kuchimaru et al., 2014). BGP, a well-recognized marker of late-stage osteoblast differentiation and bone matrix maturation, is selectively secreted by osteoblasts and plays a critical role in regulating bone metabolism (Jiang et al., 2006). The ELISA results demonstrated that the BGP content in COBCs was within the range reported for osteoblasts of other teleost species. This finding indicates that, under in vitro culture conditions, COBCs not only initiates osteogenic differentiation but also supports relatively complete bone matrix synthesis (Zeng et al., 2014). These findings are consistent with previous reports on fish osteoblast models, further supporting COBC as a reliable and stable in vitro osteoblast model (Guan et al., 2018; Yan et al., 2024).
Runx2 is widely regarded as the master transcriptional regulator of osteoblast differentiation, and in teleost fish, it exists as two isoforms, runx2a and runx2b. These isoforms exert distinct and stage-specific regulatory roles across different tissues and developmental stages (Li et al., 2009). Runx2 directly activates the transcription of multiple osteogenic genes by binding to osteoblast-specific cis-acting elements (OSEs), including osteocalcin (OC), osteopontin (OPN), bone sialoprotein (BSP), and type I collagen (Ducy et al., 1999; Ducy et al., 1997). Acting downstream of runx2, osterix is an indispensable transcription factor required for osteoblast lineage commitment, and loss of its function results in a complete failure of bone formation (Nakashima et al., 2002). Compared with muscle tissue, all examined osteogenic genes were significantly upregulated in COBCs. This upregulation may result from the synergistic regulation of osteogenic signaling pathways, growth factors, and hormonal cues under in vitro conditions (Marie, 2012). In addition, the use of juvenile Chongming crucian carp during a rapid growth phase may have contributed to higher bone cell purity, thereby facilitating elevated expression of osteogenesis-related genes (Lall & Lewis-McCrea, 2007).
Oxygen deprivation profoundly alters cellular gene expression profiles and energy metabolism (Semenza, 2010; Zhou et al., 2024). In the present study, COBC exhibited evident apoptotic features after 12 h of hypoxic exposure, with apoptotic severity increasing as hypoxia was prolonged, indicating that COBCs are highly sensitive to oxygen availability. Following 24 h of reoxygenation, cell morphology and cell density were partially restored, suggesting that COBC retains a certain capacity for damage reversal and cellular recovery (Xu et al., 2024). At the molecular level, hypoxic exposure markedly suppressed the expression of osteogenesis-related genes, including runx2a, runx2b, and osterix, with the lowest expression levels observed after 24 h of hypoxia. These findings suggest that hypoxic conditions exert a pronounced inhibitory effect on osteogenic differentiation (Li et al., 2025). Furthermore, the expression levels of hif-1a and Caspase-3 were significantly upregulated in a hypoxia duration-dependent manner, indicating that both the canonical HIF-1α–mediated hypoxic response pathway and apoptosis signaling pathways are concurrently activated under low-oxygen conditions (Lu et al., 2015; Rissanen et al., 2006). Comparable hypoxia-induced apoptotic responses have been reported in multiple fish cell types, including osteoclasts (Zhu et al., 2019) and gill cells (Xu et al., 2024). These findings further support the reliability of the present results.
With respect to oxidative stress responses, prolonged hypoxic exposure led to a gradual decline in SOD and CAT activities, whereas intracellular GSH and LDH content reached their maximum at 12 h of hypoxia. These changes suggest that COBCs activate compensatory antioxidant defense mechanisms in response to ROS accumulation and increased membrane permeability under hypoxic stress (Deme & Telekes, 2017; Fraternale et al., 2009; Yasui & Baba, 2006; Zhu et al., 2008). Following reoxygenation, all measured indicators progressively returned to normoxic levels, indicating that COBCs possess a measurable capacity for oxidative stress tolerance and functional recovery (Lushchak, 2011).
In summary, the present study successfully established and systematically characterized COBC. The results demonstrate that this cell line represents a reliable in vitro model for investigating osteogenic function, chromosomal stability, and responses to environmental stressors. This model provides a valuable experimental platform for studies of teleost skeletal development, physiological regulation, and environmental stress effects, and offers technical support for aquaculture skeletal deformity control, environmental risk assessment, and molecular breeding research.
5. Conclusion
This study successfully established and comprehensively characterized COBC. Under in vitro culture conditions, COBC exhibited stable morphology, robust proliferative capacity, and high genetic stability. Histochemical staining, biochemical assays, and molecular analyses collectively confirmed that COBC possesses typical and stable osteoblastic characteristics. Under hypoxia-reoxygenation stress, COBC exhibited pronounced responses to oxygen fluctuations, indicating a certain capacity for environmental stress adaptation and damage recovery. Collectively, COBC fills a critical gap in in vitro bone cell models for this species. This cell line provides a valuable in vitro model for investigating regulatory mechanisms of fish bone development and the effects of dissolved oxygen fluctuations, and offers essential experimental material and technical support for aquaculture and fish developmental biology research.
Author Contributions
Zaozao Guo: Writing-original draft, Validation, Software, Formal analysis, Methodology, Data curation. Jiamin Liu: Visualization, Software. Songlin Chen: Supervision, Resources. Guodong Zheng: Writing-review & editing, Conceptualization, Project administration, Funding acquisition. Shuming Zou: Supervision, Conceptualization, Project administration, Funding acquisition.:.
Funding
This research was funded by the National Key Research and Development Program of China (2023YFD2400300), the National Key Research and Development Program (2023YDF2401602) and Shanghai Ocean University Fish New Germplasm Breeding and Promotion Project (D-8006-25-0049).
Informed Consent Statement
Not applicable.
Institutional Review Board Statement
All animal experiments were approved by the Institutional Animal Care and Use Ethics Committee of Shanghai Ocean University (SHOU-DW-2024-054).
Data Availability
All data presented in this study are available on request from the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Chachami, G., Kalousi, A., Papatheodorou, L., Lyberopoulou, A., Nasikas, V., Tanimoto, K., Simos, G., Malizos, K. N., & Georgatsou, E. An association study between hypoxia inducible factor-1alpha (HIF-1α) polymorphisms and osteonecrosis. Plos One 2013, 8(11), e79647. [CrossRef]
- Deme, D., & Telekes, A. Prognostic importance of lactate dehydrogenase (LDH) in oncology. Orvosi Hetilap 2017, 158(50), 1977-1988.
- Du, H. Y., Xiong, S. B., Lv, H., Zhao, S. M., & Manyande, A. Comprehensive analysis of transcriptomics and metabolomics to understand the flesh quality regulation of crucian carp (Carassius auratus) treated with short term micro-flowing water system. Food Research International 2021, 147, 13, 110519. [CrossRef]
- Ducy, P., Starbuck, M., Priemel, M., Shen, J., Pinero, G., Geoffroy, V., Amling, M., & Karsenty, G. A Cbfa1-dependent genetic pathway controls bone formation beyond embryonic development. Genes & development. 1999, 13(8), 1025-1036. [CrossRef]
- Ducy, P., Zhang, R., Geoffroy, V., Ridall, A. L., & Karsenty, G. Osf2/Cbfa1: a transcriptional activator of osteoblast differentiation. Cell 1997, 89(5), 747-754. [CrossRef]
- Farquharson, C. Bones and Cartilage: Developmental and Evolutionary Skeletal Biology. In: Taylor & Francis. 2015. [CrossRef]
- Fraternale, A., Paoletti, M. F., Casabianca, A., Nencioni, L., Garaci, E., Palamara, A. T., & Magnani, M. GSH and analogs in antiviral therapy. Molecular aspects of medicine 2009, 30(1-2), 99-110. [CrossRef]
- Fu, S., Cai, Y. Regulatory effects of osteoblasts on os teoclastic bone resorption. Shaanxi Medical Journal 2004, 33(11): 1040-1041.
- Fu, X., Li, N., Lai, Y., Luo, X., Wang, Y., Shi, C., Huang, Z., Wu, S., & Su, J. A novel fish cell line derived from the brain of Chinese perch Siniperca chuatsi: development and characterization. Journal of Fish Biology 2015, 86(1), 32-45. [CrossRef]
- Gong, Z., Zhang, Q., Liu, J., Hu, G., Chen, S., & Wang, N. Establishment, characterization and application in germplasm conservation and disease resistance: An embryonic cell line from Yangtze sturgeon (Acipenser dabryanus). Aquaculture 2023, 575, 739807. [CrossRef]
- Guan, N., Lan, T., Chen, Y., Liu, L., Zhou, F., Gao. Z. Establishment and characterization of bone cell line for blunt snout bream (Megalobrama amblycephala). Journal of fisheries of china 2018, 42(4), 513-521.
- Han, Y. Isolated culture and identification about bionomics in rat osteoblast. Fuzhou: Fujian Agricultural and Forestry University. 2008.
- Huang, Y., Wang, S., Hu, D., Zhang, L., & Shi, S. Knockdown of GCNT2 promoted osteoblast differentiation by activating PI3K/AKT/mTOR pathway in osteoblasts. Scientific Reports 2025, 15(1), 42251. [CrossRef]
- Jing, H., Gao, L., Zhang, M., Wang, N., Lin, X., Zhang, L., & Wu, S. Establishment from the snout and kidney of goldfish, Carassius auratus, of two new cell lines and their susceptibility to infectious pancreatic necrosis virus. Fish Physiology and Biochemistry 2016, 42(1), 303-311. [CrossRef]
- Jing, H., Kong, F., Yuan, X., Zhang, M., Wang, N., Chen, D., Wu, S,. A review: The research development of fish cell lines. Chinese Journal of Fisheries 2023, 36(3), 120.
- Jiang, L., Zhang, J., Han, C., OuYang, Q. Clinical significance of changes of serum osteocalcin (BGP) levels in subjects of different age-groups. Journal of Radioimmunology 2006, 19.
- Komori, T. Regulation of osteoblast differentiation by transcription factors. Journal of cellular biochemistry 2006, 99(5), 1233-1239. [CrossRef]
- Kuchimaru, T., Hoshino, T., Aikawa, T., Yasuda, H., Kobayashi, T., Kadonosono, T., & Kizaka-Kondoh, S. Bone resorption facilitates osteoblastic bone metastasis by insulin-like growth factor and hypoxia. Von Kossa staining showing aberrant bone formation due to bone metastasis of murine osteosarcoma LM8 cells. Cancer science 2014, 105(5), Maycover. [CrossRef]
- Lakra, W., Swaminathan, T. R., & Joy, K. Development, characterization, conservation and storage of fish cell lines: a review. Fish Physiology and Biochemistry 2011, 37(1), 1-20. [CrossRef]
- Lall, S. P., & Lewis-McCrea, L. M. Role of nutrients in skeletal metabolism and pathology in fish—An overview. Aquaculture 2007, 267(1-4), 3-19. [CrossRef]
- Li, N., Felber, K., Elks, P., Croucher, P., & Roehl, H. H. Tracking gene expression during zebrafish osteoblast differentiation. Developmental dynamics 2009, 238(2), 459-466.
- Li, X., Ding, L., Feng, N., Sha, H., Zou, G., & Liang, H. Elucidating the Molecular Network Underpinning Hypoxia Adaptation in the Liver of Silver Carp (Hypophthalmichthys molitrix) via Transcriptome Analysis. Animals 2025, 15(24), 3577. [CrossRef]
- Long, F. Building strong bones: molecular regulation of the osteoblast lineage. Nature reviews Molecular cell biology 2012, 13(1), 27-38. [CrossRef]
- Lu, K. W., Wang, H., Xiao, G., Wang, L., Yi, X., Chu, X., & Huang, L. Primary cultured and establishment of hypoxic/reoxygenation model of human dermal microvascular endothelial cells. J. Cardiovasc. Pulmonary Dis. 2015, 34, 132-136.
- Lushchak, V. I. Environmentally induced oxidative stress in aquatic animals. Aquatic toxicology 2011, 101(1), 13-30. [CrossRef]
- Marie, P. J. Signaling pathways affecting skeletal health. Current osteoporosis reports 2012, 10(3), 190-198. [CrossRef]
- Marques, C. L., Rafael, M. S., Cancela, M. L., & Laizé, V. Establishment of primary cell cultures from fish calcified tissues. Cytotechnology 2007, 55(1), 9-13. [CrossRef]
- Nakashima, K., Zhou, X., Kunkel, G., Zhang, Z., Deng, J. M., Behringer, R. R., & De Crombrugghe, B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108(1), 17-29. [CrossRef]
- Nelson, R. L., Turyk, M., Kim, J., & Persky, V. Bone mineral density and the subsequent risk of cancer in the NHANES I follow-up cohort. BMC cancer. 2002, 2(1), 22. [CrossRef]
- Olsén, K. H., & Bonow, M. Crucian carp (Carassius carassius), an anonymous fish with great skills. Ichthyological Research 2003, 70(3), 313-331. [CrossRef]
- Parfitt, A. M. Skeletal heterogeneity and the purposes of bone remodeling: implications for the understanding of osteoporosis. In Osteoporosis. 2001, 433-447.
- Rafael, M. S., Marques, C., Parameswaran, V., Cancela, M., & Laizé, V. Fish bone-derived cell lines: an alternative in vitro cell system to study bone biology. Journal of Applied Ichthyology 2010, 26(2), 230-234. [CrossRef]
- Rissanen, E., Tranberg, H. K., Sollid, J., Nilsson, G. r. E., & Nikinmaa, M. Temperature regulates hypoxia-inducible factor-1 (HIF-1) in a poikilothermic vertebrate, crucian carp (Carassius carassius). Journal of Experimental Biology 2006, 209(6), 994-1003. [CrossRef]
- Rosa, J., Tiago, D., Dias, J., Cancela, M., & Laizé, V. Serum-specific stimulation of proliferation and mineralization of fish bone-derived cells. Journal of Applied Ichthyology 2010, 26(2), 251-256. [CrossRef]
- Semenza, G. L. Oxygen homeostasis. Wiley Interdisciplinary Reviews: Systems Biology and Medicine 2010, 2(3), 336-361.
- Vijayakumar, P., Laizé, V., Cardeira, J., Trindade, M., & Cancela, M. L. Development of an in vitro cell system from zebrafish suitable to study bone cell differentiation and extracellular matrix mineralization. Zebrafish 2013, 10(4), 500-509. [CrossRef]
- Wang, C., Li, Y. Studies on the Karyotype of Goldfish (Carassius auratus) Ⅰ. A Comparative Study of Chromosomes in Crucian and Red Dragon-eye Goldfish. Acta Genetica Sinica. 1982, (3), 238-242+249-250.
- Wang, J., Gao, X., Wang, L., Du, C., Hou, C., Liu, F., Xie, Q., Lou, B., Jin, S., & Zhu, J. Establishment of the first cell line from the small yellow croaker (Larimichthys polyactis) and its application in unraveling the mechanism of ROS-induced apoptosis under hypoxia. Aquaculture 2023, 563, 738900. [CrossRef]
- Wang, R., Zhang, N., Wang, R., Wang, S., & Wang, N. Two skin cell lines from wild-type and albino Japanese flounder (Paralichthys olivaceus): Establishment, characterization, virus susceptibility, efficient transfection, and application to albinism study. Fish Physiology and Biochemistry 2017, 43(6), 1477-1486. [CrossRef]
- Wei, S., Yu, Y., & Qin, Q. Establishment of a new fish cell line from the caudal fin of golden pompano Trachinotus ovatus and its susceptibility to iridovirus. Journal of Fish Biology 2018, 92(6), 1675-1686. [CrossRef]
- Witten, P. E., Harris, M., Huysseune, A., & Winkler, C. Small teleost fish provide new insights into human skeletal diseases. Methods in cell biology 2017, 138, 321-346.
- Witten, P. E., & Huysseune, A. A comparative view on mechanisms and functions of skeletal remodelling in teleost fish, with special emphasis on osteoclasts and their function. Biological Reviews 2009, 84(2), 315-346. [CrossRef]
- Wolf, K., & Quimby, M. Established eurythermic line of fish cells in vitro. Science,1962, 135(3508), 1065-1066. [CrossRef]
- Xie, Y., Chen, K., Ma, X. Isolation and identification and comparative study of calvarial osteoblasts and osteocytes of rats. Medical & Pharmaceutical Journal of Chinese People’s Liberation Army 2015, 27(3), 1-5.
- Xiao, Y., Zeng, L., Xu, J., & Zhou, Y. Establishment and characterization of a cell line derived from fin of Cryprinus carpiod. Chinese Journal of Cell Biology 2012, 34(8), 767-774.
- Xu, W., Feng, Y., Chen, S., Wang, H., Wen, J., Zheng, G., Wang, G., & Zou, S. Establishment and identification of the gill cell line from the blunt snout bream (Megalobrama amblycephala) and its application in studying gill remodeling under hypoxia. Fish Physiology and Biochemistry 2024, 50(6), 2475-2488. [CrossRef]
- Yan, T., Liu, T., Tong, G., Xu, H., Zhang, T., Kuang, Y. Establishment of a Zebrafish Danio rerio Osteoblast Cell Line from Vertebrae. Chinese journal of fisheries 2024, 37(6), 45-52.
- Yasui, K., & Baba, A. Therapeutic potential of superoxide dismutase (SOD) for resolution of inflammation. Inflammation Research 2006, 55(9), 359-363. [CrossRef]
- Zeng, G., Zong, S., Zou, B., Li. K. Effect of polygonatum polysaccharides on the expression of ALP and BGP in the differentiation of the osteoblast by mouse bone marrow derived mesenchymal stem cells. Chinese Journal of Osteoporosis 2014, 20(7).
- Zhang, H., Li, K., Wang, N., Zhai, J., & Chen, S. Establishment and characterization of a liver cell line from spotted knifejaw (Oplegnathus punctatus). Journal of Agricultural Biotechnology 2020, 28(2), 360-368.
- Zhang, W., Chen, H., Liu, W., Jia, K., & Yi, M. Characterizing Marine Medaka (Oryzias melastigma) Haploid Embryonic Stem Cells: A Valuable Tool for Marine Fish Genetic Research. Animals: an Open Access Journal from MDPI. 2024, 14(18), 2739. [CrossRef]
- Zhou, M., Wang, J., Cao, R., Zhang, F., Luo, X., Liao, Y., Chen, W., Ding, H., Tan, X., & Qiao, Z. Hypoxia-induced differences in the expression of pyruvate dehydrogenase kinase 1-related factors in the renal tissues and renal interstitial fibroblast-like cells of Yak (Bos Grunniens). Animals 2024. 14(21), 3110. [CrossRef]
- Zhu, H., Jia, Z., Misra, B. R., Zhang, L., Cao, Z., Yamamoto, M., Trush, M. A., Misra, H. P., & Li, Y. Nuclear factor E2-related factor 2-dependent myocardiac cytoprotection against oxidative and electrophilic stress. Cardiovascular Toxicology 2008, 8(2), 71-85. [CrossRef]
- Zhu, J., Tang, Y., Wu, Q., Ji, Y. C., & Kang, F. W. Mechanism of participation of osteocytes in the formation of osteoclasts under hypoxia. Hua xi kou qiang yi xue za zhi. 2019, 37(5), 463-468. [CrossRef]
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).