
Submitted:
05 October 2025
Posted:
06 October 2025
You are already at the latest version
Abstract
This study investigates the osteogenic potential of rat dental pulp stem cells (DPSCs) seeded on biphasic calcium phosphate (BCP) scaffolds in a rat mandibular defect model and evaluates whether overexpression of the osteogenic transcription factor RUNX2 enhances this regenerative effect. Dental pulp stem cells (DPSCs) were isolated, characterized, and transduced with a RUNX2-expressing lentiviral vector before scaffold seeding and in vivo implantation. Bone regeneration was assessed through micro-computed tomography, histology, immunohistochemistry, and biomechanical testing. Results showed that DPSCs significantly promoted bone formation, and RUNX2 overexpression further improved bone volume, trabecular architecture, osteogenic marker expression, and biomechanical strength. Findings underscore the potential of genetically enhanced DPSC therapy for personalized, cell-based treatment of mandibular bone defects. The objectives of this study were to evaluate the DPSCs and of DDPSCs and generate bone in mandibular fractures, identify genetic markers influencing bone healing and differentiation of DPSCs, and explore the roles of genetic markers influencing bone regeneration.
Keywords:
dental pulp stem cells (DPSCs)
; RUNX2
; mandibular bone fracture
; biphasic calcium phosphate (BCP)
; scaffold
Introduction
Stem cells are remarkable for their potential to differentiate into various cell types in the body, serving as a natural repair system. Their unique abilities, including self-renewal over extended periods and differentiation into specialized cells such as muscle, blood, or bone cells, make them a cornerstone of regenerative medicine. Broadly categorized into embryonic and adult stem cells, these versatile cells have revolutionized tissue repair and regeneration [1]. A scientist successfully isolated dental pulp stem cells (DPSCs) from adult dental tissue. [2]. Since then, a variety of dental stem cells were identified, including DPSCs, Dental Follicle Stem Cells (DFSCs) [3], Periodontal Ligament Stem Cells (PDLSCs) [4], and Stem Cells from the Apical Papilla (SCAP) [5], have been identified, each contributing uniquely to dental tissue engineering.
Effective treatment of mandibular fractures is crucial due to the mandible's essential roles in mastication, speech, and facial aesthetics [6]. Improper or delayed healing can lead to malocclusion, impaired function, and aesthetic concerns, significantly affecting a patient's quality of life. Traditional treatments, such as open reduction and internal fixation, aim to restore anatomical alignment but may be associated with complications like Bacterial infections, such as Osteomyelitis, Periodontal Abscess and Salivary Gland infections, Viral infections, such as Herpes simplex virus type 1 (HSV-1), and Fungal infections, such as Oral Candidiasis, which are the least common. Therefore, enhancing bone healing processes is vital to improve outcomes and reduce recovery times [7].
Bone regeneration has been significantly advanced by the emergence of DPSCs as a promising avenue for therapeutic development. [8] Isolated from dental pulp, DPSCs are mesenchymal stem cells capable of differentiating into osteoblasts, the cells responsible for bone formation [9]. Their multipotent nature, coupled with ease of access and low morbidity at the collection site, makes them attractive for therapeutic applications [10]. Many studies in Vivo have shown that DPSCs can enhance bone healing. [11]. For instance, research indicates that DPSCs, when supported by suitable scaffolds, significantly increase bone regeneration, suggesting their potential in clinical applications [12]. Furthermore, DPSCs exhibit low immunogenicity, reducing the risk of immune rejection and potentially allowing for allogeneic transplants [11]. Their ability to interact favorably with various biomaterials further supports their use in bone tissue engineering [8]. These characteristics position DPSCs as a valuable tool in developing advanced therapies for mandibular fracture healing, aiming to improve patient outcomes and reduce the limitations associated with conventional treatments [10].
Genetic markers help play a pivotal role in bone regeneration by influencing the differentiation and activity of osteoblasts and osteoclasts, which are essential for bone formation and resorption. Variations in genes such as The Runt-related transcription factor 2 (RUNX2) and The Fibroblast growth factors (FGFs) have been linked to differences in bone healing efficiency [13]. Epigenetic mechanisms, including DNA methylation and histone modifications (H1, H2A2B), also regulate bone remodeling processes [14]. Our study is concerned with identifying these genetic factors which enhance our understanding of fracture healing and may lead to personalized therapeutic strategies [15].
Bone regeneration is a critical aspect of maxillofacial surgery, particularly for repairing mandibular fractures and defects. The human body has a natural ability to regenerate bone, yet extensive bone loss due to trauma or disease often requires additional therapeutic interventions [16]. Traditional treatments, such as autologous bone grafts, have limitations including donor site morbidity and limited availability of graft material [17]. Therefore, stem cell-based therapies, particularly those using DPSCs, have emerged as a promising alternative for bone regeneration [18].
Stem cells from the Dental Pulp were first identified by a group of scientists, led by Peter Gronthos, who characterized their multipotent nature and ability to differentiate into osteoblasts, chondrocytes, and adipocytes. [19]. Since then, numerous studies have demonstrated the osteogenic potential of DPSCs in bone tissue engineering [20]. Much research shows that DPSCs, when combined with scaffolds, significantly enhance bone healing in vivo, highlighting their potential for clinical applications [21].
Genetic factors play a crucial role in regulating bone regeneration, particularly by influencing osteogenic differentiation. Several genetic markers are integral to the differentiation of osteoblasts, with the RUNX2 gene serving as the master regulator of osteoblast differentiation. RUNX2 controls the expression of key osteogenic markers, such as collagen type I alpha 1 (COL1A1) and the secreted phosphoprotein 1, also known as Osteopontin (SPP1) [22]. Mutations in RUNX2 are linked to cleidocranial dysplasia, a condition marked by defective bone formation [23]. Osteopontin, also referred to as SPP1, plays a critical role in bone remodeling by facilitating cell adhesion, migration, and mineral deposition. Animal models lacking SPP1 exhibit delayed bone healing and impaired osteoclast function, highlighting its essential contribution to osteogenesis [24].
Odontogenesis, the process of tooth development, shares several molecular pathways with osteogenesis, making it a relevant topic in dental tissue engineering. DPSCs play a crucial role in both dentin formation and bone healing, as they originate from the neural crest and possess a high differentiation potential [25]. Many signaling pathways, such as Bone morphogenic proteins (BMPs), Wingless-type MMTV integration site family (Wnt), and (FGF), regulate both odontogenesis and osteogenesis, highlighting the overlap between these two processes [26]. This pathway enhances the proliferation, differentiation, and survival of stem cells in response to injury, making it a promising therapeutic target for enhancing mandibular fracture healing [27]. Furthermore, dentin matrix proteins, particularly Dentin Sialophosphoprotein (DSPP) play a significant role in enhancing bone regeneration. Although primarily known for its involvement in dentinogenesis, DSPP has shown to also influence bone healing and regeneration. It acts as an important matrix protein that not only supports the formation of dentin but also promotes the differentiation of osteoblasts, the cells responsible for bone formation [28].
This function makes DSPP a valuable component in tissue engineering and regenerative medicine [29]. Despite their potential, the clinical translation of DPSC-based therapies faces several challenges. The heterogeneity of DPSCs and variability in differentiation potential [30]. Furthermore, ensuring vascularization and integration of engineered bone remains a significant challenge, necessitating biomaterial innovations and scaffold modifications in mandibular fractures specifically [31].
This study builds prior work using scaffolds like Biphasic Calcium Phosphate (BCP) with stem cells [32] and aims to advance the field by analyzing genetic markers influencing DPSC-mediated repair in mandibular defects. Unlike previous research which focused mainly on calvarial models, this work addresses a key gap by applying findings to mandibular bone repair [33].
2. Materials and Methods
2.1. Isolation and Culture of DPSCs
- Collagenase I: (Sigma-Aldrich, St. Louis, MO, USA).
- Dispase: (Gibco, Thermo Fisher Scientific, Waltham, MA, USA).
- Filter (70 µm): (Falcon, Corning Inc., Corning, NY, USA).
- α-MEM (Minimal essential medium Eagle – alpha modification): (Gibco, Thermo Fisher Scientific).
- Fetal bovine serum (FBS): (Gibco, Thermo Fisher Scientific).
- Penicillin–streptomycin: (Gibco, Thermo Fisher Scientific).
2.2. Phenotypic Characterization of DPSCs
- Bovine serum albumin (BSA): (Sigma-Aldrich).
- Antibodies: Ecto-5'-nucleotidase (CD73), Thy-1 (CD90), Endoglin (CD105), CD34, CD45.
- Flow cytometer: (BD Accuri C6 flow cytometer, BD Biosciences, San Jose, CA, USA).
- Data analysis software: (FlowJo v10, FlowJo LLC, Ashland, OR, USA).
2.3. In Vivo Mandibular Fracture Model
- Isoflurane/O₂ anesthesia: (Baxter Healthcare, Deerfield, IL, USA).
- Micro-saw: (Fine Science Tools, Foster City, CA, USA).
- Biphasic calcium phosphate (BCP): (Berkeley Advanced Biomaterials Inc., Berkeley, CA, USA).
- RUNX2 lentivirus: (VectorBuilder Inc., Chicago, IL, USA).
- Resorbable collagen membrane: (Geistlich Pharma AG, Wolhusen, Switzerland).
- Vicryl Rapide™ sutures: (Ethicon, Johnson & Johnson, Somerville, NJ, USA).
- Buprenorphine (0.05 mg/kg): (Reckitt Benckiser, Slough, UK).
2.4. Genetic Marker Expression Analysis
- RNA extraction kit: (RNeasy Mini Kit, Qiagen, Hilden, Germany).
- cDNA synthesis kit: (High-Capacity cDNA Reverse Transcription Kit, Applied Biosystems, Thermo Fisher Scientific).
- SYBR Green assay kit: (PowerUp SYBR Green Master Mix, Applied Biosystems).
- SDS-PAGE: (Bio-Rad Laboratories, Hercules, CA, USA).
- PVDF membrane: (MilliporeSigma, Burlington, MA, USA).
- Chemiluminescence substrate: (SuperSignal™ Enhanced Chemiluminescence), (ECL), (Thermo Fisher Scientific).
- Paraformaldehyde (4% PFA): (Electron Microscopy Sciences, Hatfield, PA, USA).
- Triton X-100 (0.1%): (Sigma-Aldrich).
- BSA (5%): (Sigma-Aldrich).
- Primary antibodies (anti-RUNX2/OCN): (Abcam, Cambridge, UK).
- Secondary antibodies (Alexa Fluor): (Thermo Fisher Scientific).
- DAPI (4′,6-diamidino-2-phenylindole): (Sigma-Aldrich).
2.5. Histological and Immunohistochemical Evaluation
- Decalcification solution (10% EDTA): (Sigma-Aldrich).
- Hematoxylin and Eosin (H&E): (Sigma-Aldrich).
- Masson’s trichrome stain: (Polysciences, Warrington, PA, USA).
- Antigen retrieval buffer (citrate buffer pH 6): (Dako, Agilent Technologies, Santa Clara, CA, USA).
- Anti-Osteocalcin (OCN) antibody: (Abcam).
- DAB detection kit: (Dako).
- Hematoxylin counterstain: (same as above, Sigma-Aldrich implied)
- ImageJ software: (NIH, Bethesda, MD, USA).
2.6. Scaffold Preparation and Cell Seeding
- Biphasic calcium phosphate (BCP; 60/40 HA/β-TCP, 0.5–1 mm granules): (Berkeley Advanced Biomaterials Inc., Berkeley, CA, USA).
- α-MEM: (Gibco, Thermo Fisher Scientific).
2.7. Scaffold Design and Fabrication
- Type I collagen: (Corning).
- Polycaprolactone (PCL): (Sigma-Aldrich).
2.8. Methods
2.8.1. Isolation and Culture of DPSCs
Teeth were collected from healthy Male Sprague–Dawley rats (8–10 weeks, 250–300 g). Dental pulp was extirpated, minced into ~1 mm³ fragments, and digested in 3 mg/mL collagenase I (Sigma-Aldrich, St. Louis, MO, USA) and 4 mg/mL dispase (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) for 1 h at 37 °C. The resulting cell suspension was filtered (70 µm, Falcon, Corning Inc., Corning, NY, USA), centrifuged (300×g, 5 min), and plated in a (Gibco’s MEM) minimal essential medium Eagle – alpha modification (α-MEM; Gibco, Thermo Fisher Scientific) with 10% Fetal bovine serum (FBS; Gibco, Thermo Fisher Scientific) and 1% penicillin–streptomycin (Gibco, Thermo Fisher Scientific) at 1×10⁴ cells/cm². Cultures were maintained at 37 °C, 5% CO₂; passages 2–4 were used for all experiments [33].
2.8.2. Phenotypic Characterization of DPSCs
Passage 3 DPSCs were harvested, blocked with 0.5% Bovine serum albumin (BSA; Sigma-Aldrich), and stained for Ecto-5'-nucleotidase (CD73), Thy-1 (Thymocyte differentiation antigen 1) (CD90), Endoglin (CD105), Hematopoietic progenitor cell antigen (CD34), and Protein tyrosine phosphatase, receptor type C, also known as Leukocyte common antigen (CD45). After washing, cells were analyzed on a BD Accuri C6 flow cytometer (BD Biosciences, San Jose, CA, USA) and data processed with FlowJo v10 (FlowJo LLC, Ashland, OR USA) to confirm Mesenchymal stem cells (MSC) marker profile (CD73⁺, CD90⁺, CD105⁺; CD34⁻, CD45⁻) [33].
2.8.3. In Vivo Mandibular Fracture Model
Male Sprague–Dawley rats (36 Rats, 8–10 weeks, 250–300 g) were anesthetized with isoflurane/O₂ (Baxter Healthcare, Deerfield, IL, USA). A unilateral mid-body mandibular osteotomy (5 mm defect) was created with a micro-saw (Fine Science Tools, Foster City, CA, USA). Rats were randomized into three groups (n = 12 each):
- Control: defect + BCP only
- DPSC: defect + BCP + DPSCs (1×10⁶)
- RUNX2-modulated DPSC: defect + BCP + DPSCs transduced with RUNX2 lentivirus (MOI 50; VectorBuilder Inc., Chicago, IL, USA)
Defects were covered with a resorbable collagen membrane (Geistlich Pharma AG, Wolhusen, Switzerland) and closed with Vicryl Rapide™ sutures (Ethicon, Johnson & Johnson, Somerville, NJ, USA). Buprenorphine (0.05 mg/kg; Reckitt Benckiser, Slough, UK) and soft diet were provided post-operatively. Animals were sacrificed in weeks 4 and 8; mandibles harvested for analysis [31].
2.8.4. Genetic Marker Expression Analysis
- Real-Time Quantitative Reverse Transcription PCR (qRT-PCR): RNA extraction (RNeasy Mini Kit, Qiagen, Hilden, Germany), cDNA synthesis (High-Capacity cDNA Reverse Transcription Kit, Applied Biosystems, Thermo Fisher Scientific), and SYBR Green assays (PowerUp SYBR Green Master Mix, Applied Biosystems) for RUNX2, alkaline phosphatase (ALPL), Collagen type 1 alpha 1 (COL1A1), bone morphogenetic protein 2 (BMP2), and Osteocalcin (OCN); bone gamma-carboxyglutamic acid-containing protein (BGLAP) as internal control.
- Western Blot: Protein lysates separated by Sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE; Bio-Rad Laboratories, Hercules, CA, USA), transferred to Polyvinylidene fluoride (PVDF; MilliporeSigma, Burlington, MA, USA), probed for RUNX2, COL1A1, and OCN, and visualized via Thermo Fisher Scientific’s SuperSignal™ Enhanced Chemiluminescence (ECL).
- Immunocytochemistry: Cells fixed Paraformaldehyde (4% PFA; Electron Microscopy Sciences, Hatfield, PA, USA), permeabilized (0.1% Triton X-100; Sigma-Aldrich), blocked (5% BSA; Sigma-Aldrich), incubated with anti-RUNX2/OCN (Abcam, Cambridge, UK), then Alexa Fluor secondary antibodies (Thermo Fisher Scientific); nuclei counterstained with 4′,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich) [12,13].
2.8.5. Histological and Immunohistochemical Evaluation
Specimens were decalcified (10% EDTA; Sigma-Aldrich), paraffin-embedded, and sectioned (5 µm). Sections stained with Hematoxylin and Eosin (H&E; Sigma-Aldrich) and Masson’s trichrome (Polysciences, Warrington, PA, USA). For IHC, antigen retrieval (citrate buffer pH 6; Dako, Agilent Technologies, Santa Clara, CA, USA) preceded incubation with anti-OCN (Abcam) and DAB detection (Dako). Sections counterstained with hematoxylin. New bone area quantified via ImageJ (NIH, Bethesda, MD, USA) [34].
2.1.6. Scaffold Preparation and Cell Seeding
Biphasic calcium phosphate (BCP; 60/40 HA/β-TCP, 0.5–1 mm granules; Berkeley Advanced Biomaterials Inc., Berkeley, CA, USA) was pre-wetted in α-MEM overnight at 37 °C. DPSCs were resuspended at 1×10⁶ cells per 30 mg BCP and incubated in 96-well plates at 37 °C, 5% CO₂ for 12 h to allow attachment [31].
2.1.7. Scaffold Design and Fabrication
Scaffolds were designed to mimic the native extracellular matrix (ECM) architecture with high porosity (100–700 µm macropores; <100 µm micropores) to facilitate vascularization and nutrient diffusion. Fabrication techniques included salt leaching, gas forming, phase separation, freeze drying, and electrospinning. Biphasic and multiphasic constructs were produced by layering materials with distinct compositions and pore geometries. Decellularized ECM and polymeric biomaterials collagen (Type I collagen; Corning) and Polycaprolactone (PCL; Sigma-Aldrich) were functionalized with ECM proteins and growth factors [32].
2.1.8. Statistical Analysis
Data (mean ± SD) were analyzed by one-way ANOVA with Tukey’s post hoc, and repeated-measures ANOVA for time-courses; p < 0.05 considered significant. Analyses performed in GraphPad Prism 9 software [34].
3. Results
3.1. Isolation and Phenotypic Characterization Confirms the Mesenchymal Stem Cell Identity of Cultured DPSCs
This study confirms that primary cultures from rat teeth yielded an average of 1.2 × 10⁶ cells per tooth fragment by passage 2 in flow cytometry, with a typical spindle-shaped MSC morphology and >95% viability on Trypan Blue exclusion. Dental pulp stem cells (DPSCs) at P3 expressed high levels of CD73 (98.2 ± 1.1 %), CD90 (97.5 ± 1.3 %), CD105 (96.8 ± 1.5 %) and were negative for CD34 (1.2 ± 0.5 %) and CD45 (0.8 ± 0.4 %), confirming their mesenchymal identity. These phenotypic data establish the MSC profile of the cultured DPSCs and provide the baseline cell characterization for subsequent osteogenic induction and in vivo experiments (see the schematic of the in vivo model in Figure 1).
Figure 1.
Schematic representation of the in vivo mandibular defect model and experimental workflow. Male Sprague–Dawley rats underwent standardized 5 mm unilateral mid-body mandibular defects. Groups included: (1) BCP only (Control), (2) BCP + DPSCs, and (3) BCP + RUNX2-modified DPSCs. After scaffold implantation and collagen membrane coverage, wounds were closed with absorbable sutures. Post-operative care included analgesia and soft diet. Animals were sacrificed at 4 and 8 weeks for micro-CT, histology, and biomechanical testing. This schematic illustrates defect creation, treatment allocation, and analysis timeline.
Figure 1.
Schematic representation of the in vivo mandibular defect model and experimental workflow. Male Sprague–Dawley rats underwent standardized 5 mm unilateral mid-body mandibular defects. Groups included: (1) BCP only (Control), (2) BCP + DPSCs, and (3) BCP + RUNX2-modified DPSCs. After scaffold implantation and collagen membrane coverage, wounds were closed with absorbable sutures. Post-operative care included analgesia and soft diet. Animals were sacrificed at 4 and 8 weeks for micro-CT, histology, and biomechanical testing. This schematic illustrates defect creation, treatment allocation, and analysis timeline.
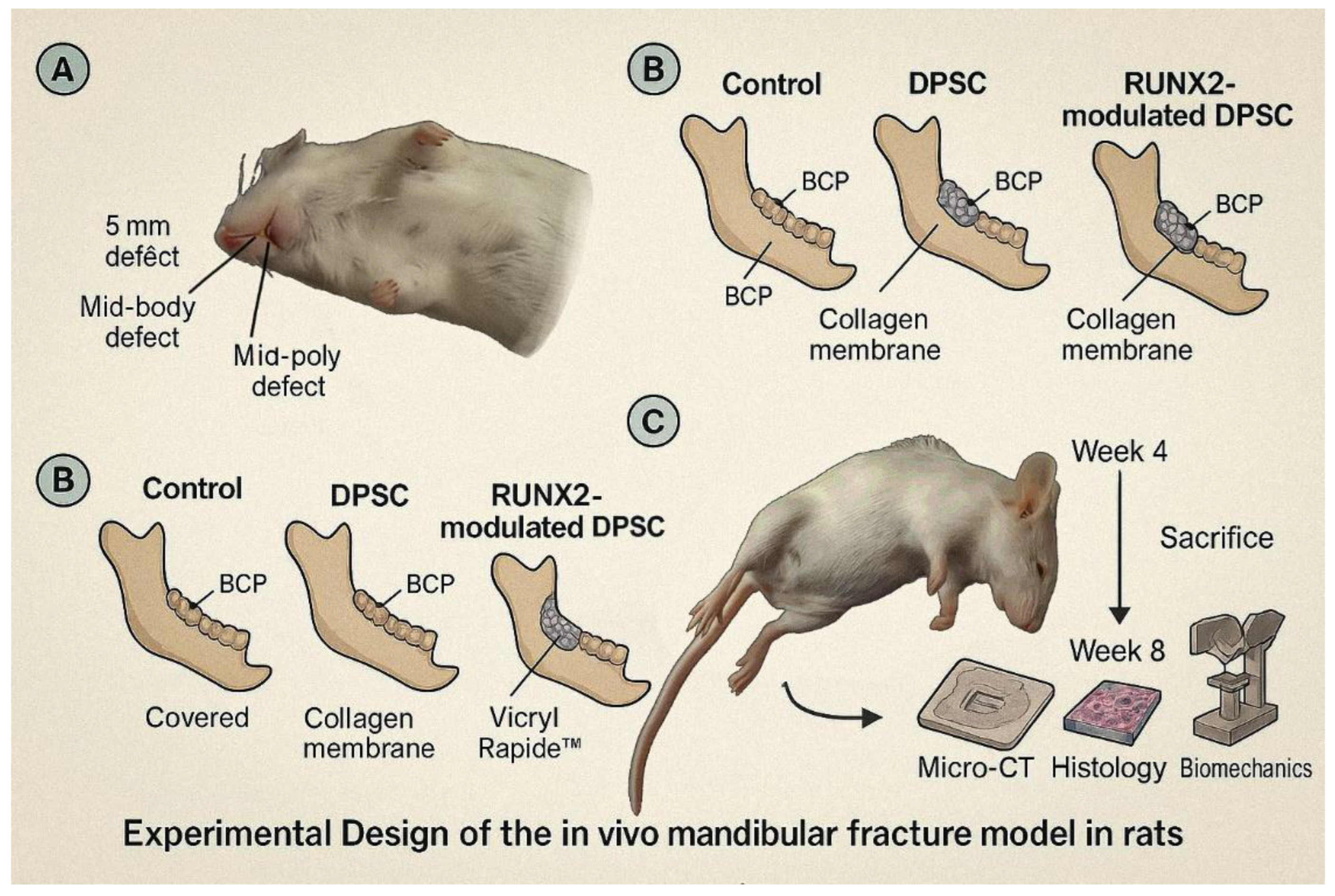
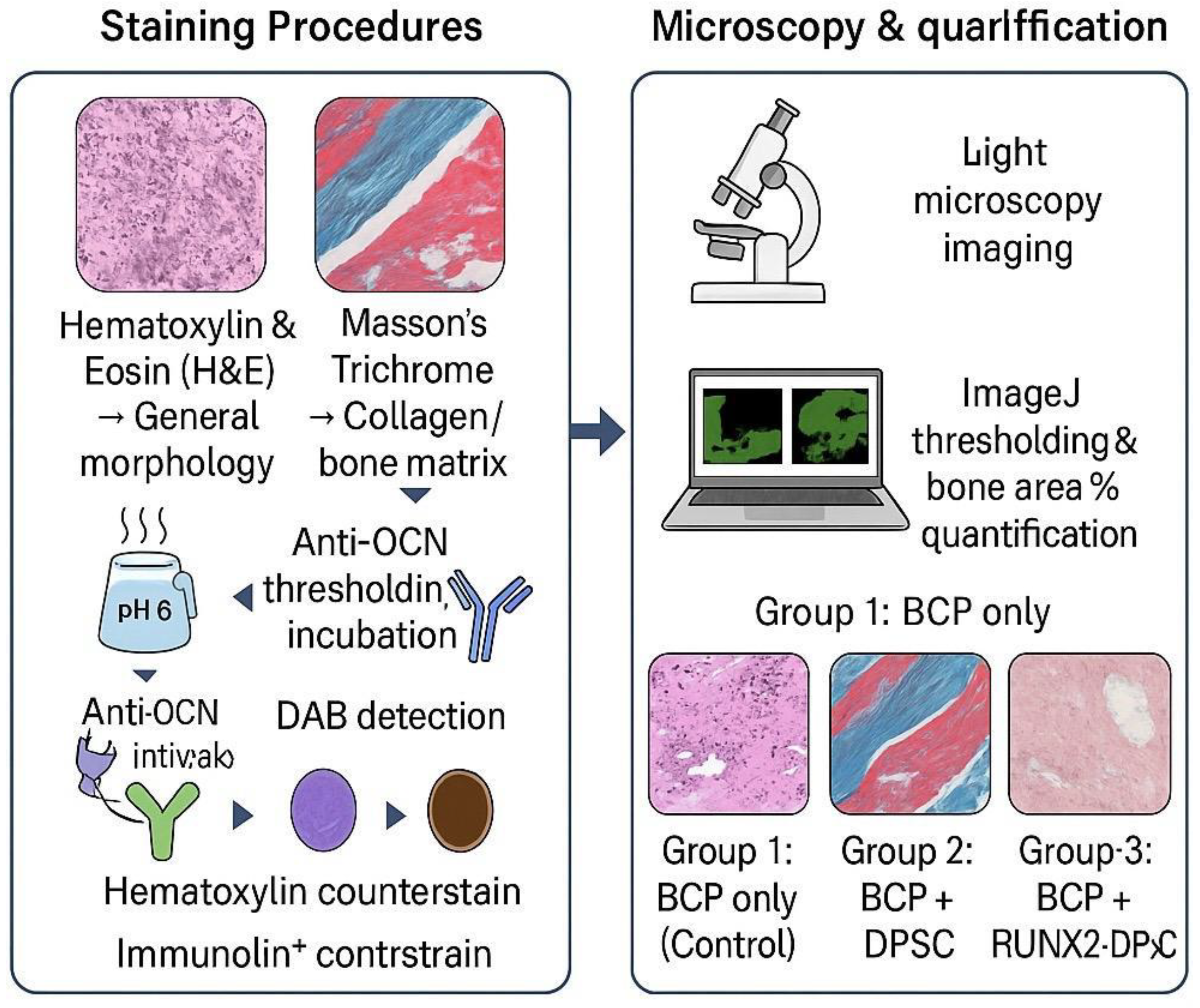
Figure 2.
Workflow of Histological and Immunohistochemical Evaluation in Mandibular Defect Healing.
3.2. Sequential and Temporal Induction of Key Osteogenic Genes Confirmed by qRT-PCR Analysis
RUNX2 (an early/mid osteogenic transcription factor) rose markedly by day 7 and peaked at day 14 before slightly declining by day 21; ALPL (ALP) and BMP2 also showed early-to-mid induction with highest fold-changes at day 14; COL1A1 increased steadily during the induction period; and OCN (BGLAP), a late mineralization marker, showed the greatest increase by day 21. My results in Table 1 showed that fold-change versus Day 0 and indicate statistical significance versus Day 0. The qRT-PCR findings provide the molecular context for the subsequent protein-level validation.
3.3. Western Blot and Immunocytochemistry Confirm Robust Elevation of RUNX2 and OCN Protein Expression
At the protein level, Western blot densitometry and immunocytochemistry confirmed the transcriptional trends: protein levels of RUNX2 and OCN increased concordantly, with densitometry showing ~4-fold (RUNX2) and ~6-fold (OCN) elevations at day 21. Immunocytochemistry confirmed robust nuclear RUNX2 and cytoplasmic OCN staining by day 14. These protein-level data validate the qRT-PCR results and demonstrate that transcriptional induction translates into increased osteogenic protein expression (see qRT-PCR quantitation in Table 1 and representative images in the Figure 4).
3.4. DPSC-BCP Scaffold Interactions: High Seeding Efficiency and Favorable Cell Adhesion
DPSC scaffold interactions were favorable: seeding efficiency onto BCP granules was 85 ± 4% after 12 h, and SEM imaging showed cells spreading over pore surfaces and establishing filopodia-mediated contacts. The attachment and spreading data document that the BCP scaffold supports efficient DPSC adhesion before implantation, which underpins the in vivo repair results presented below.
3.5. Micro-CT Morphometric Analysis Demonstrates Superior Bone Volume and Microarchitecture in DPSC-Treated Defects
Micro-CT morphometric analysis of mandibular defects showed clear, group-dependent improvements in bone metrics. Bone volume / total volume (BV/TV) at week 4 and week 8, trabecular thickness (Tb.Th) and trabecular number (Tb.N) were all improved in the DPSC + BCP group versus Control (BCP only), with further enhancement in the RUNX2-DPSC + BCP group. These quantitative micro-CT results are presented in Table 2 and indicate statistically significant differences. The micro-CT outcomes provide 3D structural evidence that DPSCs enhance bone formation and that RUNX2 overexpression further potentiates microarchitecture improvements (see Table 2 for the full morphometric dataset).
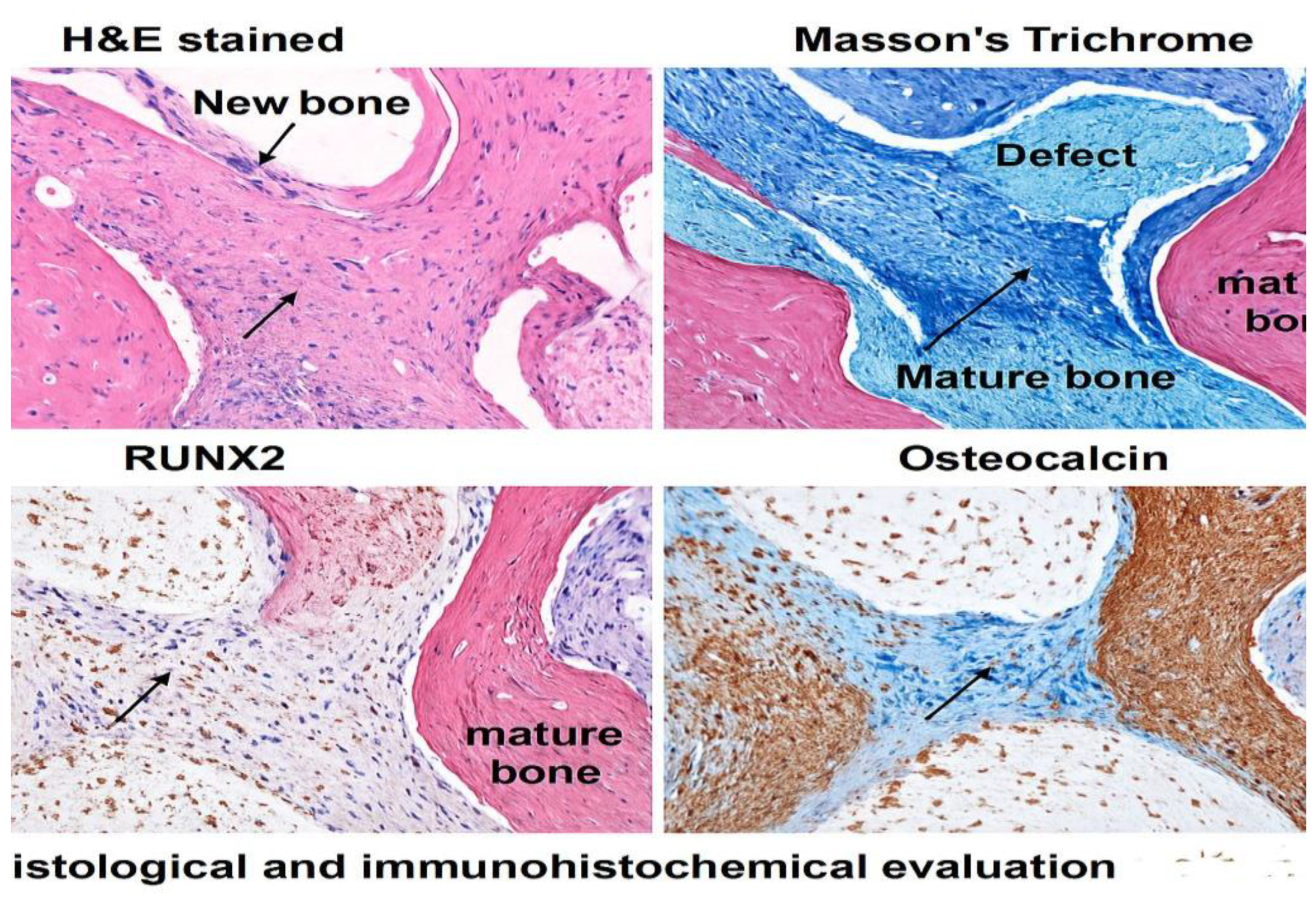
3.6. Histological and Immunohistochemical Evaluation Corroborates Enhanced Osteogenic Activity in DPSC and RUNX2-DPSC-Treated Defects
Histological and immunohistochemical evaluation corroborated imaging and molecular findings: H&E and Masson’s trichrome revealed sparse, discontinuous bone islands in Control defects, abundant woven bone bridging >60% of the gap in DPSC-treated defects by week 8, and near-complete defect closure with dense lamellar bone in the RUNX2-DPSC group. ImageJ-based area quantification of new bone confirmed these observations (see the quantification in Figure 3 and the representative histology panels in Figure 4). OCN immunostaining showed strong positive staining in DPSC and RUNX2-DPSC groups, with semi-quantitative image analysis indicating OCN-positive area fractions of 48 ± 5% (DPSC) and 62 ± 6% (RUNX2-DPSC) versus 25 ± 4% in controls, further confirming enhanced osteogenic activity in treated groups (quantitative IHC results and visual examples are provided in the figures below — as mentioned in Figure 3 and Figure 4).
Figure 3.
Error bars represent ± standard deviation (SD), indicating the variability around the mean.
Figure 3.
Error bars represent ± standard deviation (SD), indicating the variability around the mean.
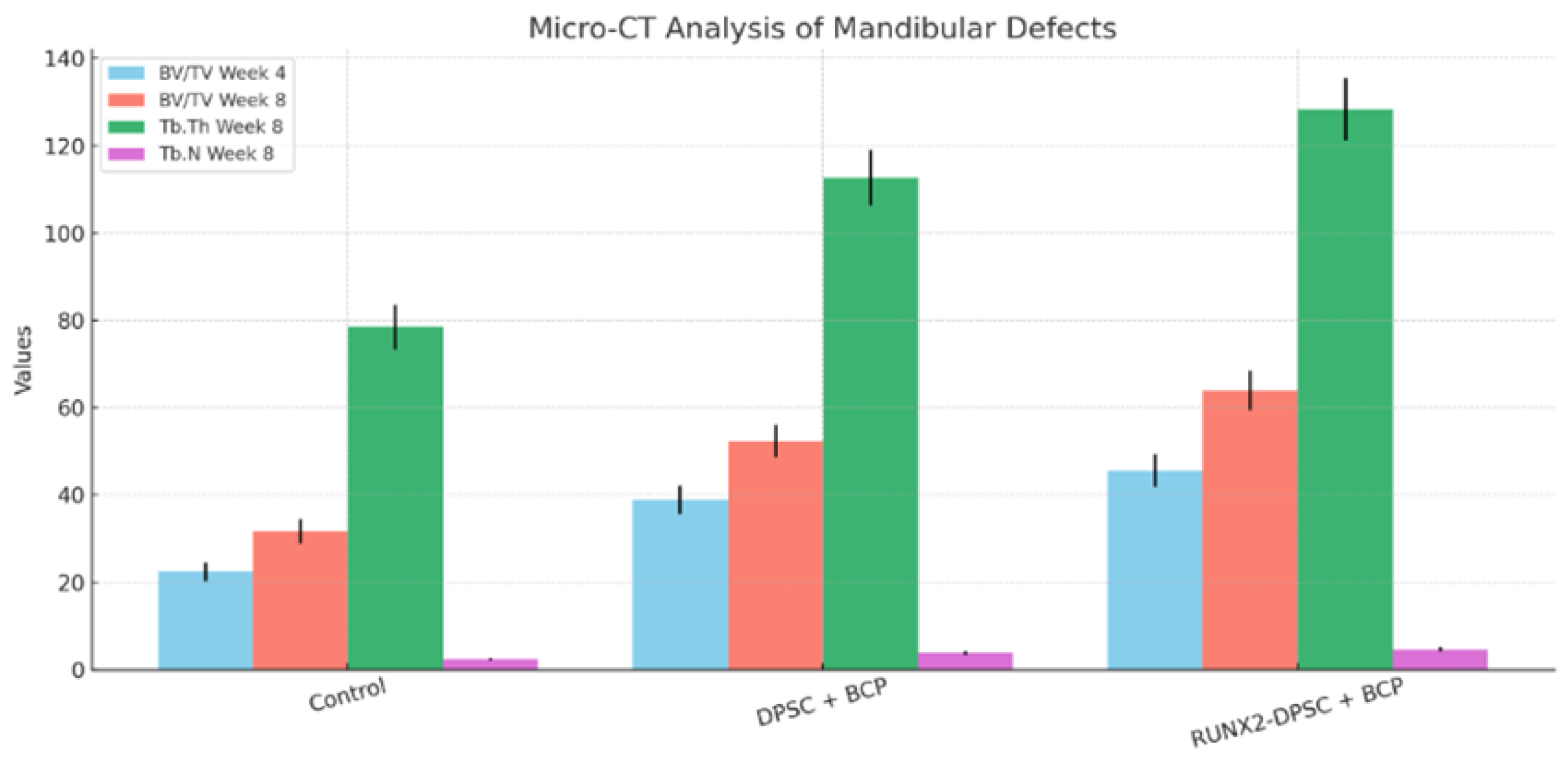
Figure 4.
Representative ImageJ-processed histological sections and quantification of new bone area in mandibular defects treated with BCP only (Control), BCP + DPSCs, and BCP + RUNX2-modified DPSCs. Binary threshold masks illustrate bone tissue (white) vs. non-bone areas (black), with corresponding bar graph showing % bone area.
Figure 4.
Representative ImageJ-processed histological sections and quantification of new bone area in mandibular defects treated with BCP only (Control), BCP + DPSCs, and BCP + RUNX2-modified DPSCs. Binary threshold masks illustrate bone tissue (white) vs. non-bone areas (black), with corresponding bar graph showing % bone area.
4. Discussion
The restoration of critical-sized mandibular defects continues to pose a considerable clinical challenge in maxillofacial surgery, frequently necessitating bone grafts that present limitations, including donor site morbidity and restricted availability. Our research presents substantial evidence that dental pulp stem cells (DPSCs) implanted onto a biphasic calcium phosphate (BCP) scaffold represent a highly promising tissue engineering approach to tackle this challenge. The primary outcomes of this study indicate that rat dental pulp stem cells (DPSCs) significantly promote bone regeneration in a rat mandibular defect model, and that the osteogenic capacity of these cells is considerably enhanced by the overexpression of the transcription factor RUNX2. A multi-faceted assessment showed that the therapeutic benefit was real. Defects treated with RUNX2-overexpressing DPSCs had not only more bone volume, but also a more mature microarchitecture and better biomechanical strength than defects treated with naive DPSCs or just scaffolds.These results strongly support the idea that DPSCs have an inherent ability to form bone, which has been well-documented in vitro [8], and successfully show that this ability works in a complex in vivo setting.
The BCP scaffold, which is known for its ability to help bones grow, seems to be the perfect three-dimensional structure that keeps cells in place, helps them grow, and eventually leads to the formation of a mineralized matrix. The substantial improvement in regenerative outcomes seen with RUNX2-overexpressing DPSCs highlights an essential principle: the mere presence of stem cells on a scaffold may not suffice for optimal repair. Instead, turning on important genetic programs that control osteoblast lineage commitment ahead of time can have a big impact on the process of regeneration. RUNX2 is very important for the differentiation of mesenchymal stem cells into pre-osteoblasts and their later maturation. It is often called the "master regulator" of osteogenesis. Our findings indicate that increasing RUNX2 expression in DPSCs before implantation effectively primes the cells, expediting their differentiation process and improving their functional performance at the defect site. This resulted in the development of bone tissue exhibiting characteristics—demonstrated by micro-computed tomography (micro-CT) metrics such as bone volume/total volume (BV/TV), trabecular number (Tb.N), and connectivity density—that more closely resemble native mandibular bone.
The in-vivo results shown here are very promising, but they also make us want to learn more about the molecular mechanisms behind them. RUNX2 overexpression probably causes more bone formation by starting a chain reaction of events that happen after it. RUNX2 may not only directly enhance the expression of bone matrix proteins such as osteocalcin and bone sialoprotein but also regulate the secretion of osteotropic factors that facilitate angiogenesis and coordinate the activity of host progenitor cells, thereby establishing a more potent regenerative niche. To transcend correlation and ascertain causation, current investigations in our laboratory are utilizing transcriptomic (e.g., RNA sequencing) and epigenetic profiling [13] of collected constructs. These analyses will elucidate the extensive gene regulatory networks activated by RUNX2 in this particular context and identify potential epigenetic modifications that stabilize the osteogenic phenotype, yielding a system-level comprehension of how genetic priming enhances DPSC-mediated repair. The scaffold–cell interface is another important place to look for ways to improve things. The BCP scaffold used in this study worked well, but its surface is not very reactive, so it may not fully mimic the changing signaling environment of the native extracellular matrix (ECM). Subsequent iterations of this strategy will emphasize the advanced functionalization of biomaterials to establish a more bioactive environment [4]. This includes putting certain ECM-derived peptides (like RGD sequences) or recombinant growth factors (like BMP-2) on the surface to give cells long-lasting, localized signals that work with their genetic programming. Also, looking into multiphasic or gradient scaffold designs that control the timing and location of biochemical and biophysical signals could better mimic the transitional zones of the mandible and help create bone tissue that is anatomically correct. Ultimately, the encouraging short-term outcomes observed in a rodent model must be situated within the broader trajectory toward clinical implementation. An essential phase will involve the evaluation of long-term safety, stability, and immunogenicity in more extensive, immunocompetent preclinical models [33]. Important questions about the possibility of ectopic bone formation, the future of the implanted cells, and the immune response to both the allogeneic cells and the scaffold material over long periods of time still need to be answered. A thorough assessment involving serial imaging, histopathology, and immune cell profiling will be crucial to mitigate risks associated with this therapeutic strategy. In conclusion, this study demonstrates that RUNX2-overexpressing dental pulp stem cells (DPSCs) on a biphasic calcium phosphate (BCP) scaffold constitute a robust combination for bone regeneration in a mandibular defect model. The results show how well a supportive biomaterial platform and the strategic genetic enhancement of stem cells work together. Our current and forthcoming endeavors are focused on elucidating the mechanistic foundations, enhancing the biomaterial component, and conducting thorough safety evaluations. We expect that these coordinated efforts will not only confirm and build on the current findings but also lay the groundwork for the creation of effective and tailored DPSC-based therapies for complicated craniofacial reconstruction.
5. Conclusions
In conclusion, this study demonstrates that RUNX2-overexpressing dental pulp stem cells (DPSCs) on a biphasic calcium phosphate (BCP) scaffold constitute a robust combination for bone regeneration in a mandibular defect model. The results show how well a supportive biomaterial platform and the strategic genetic enhancement of stem cells work together. Our current and forthcoming endeavors are focused on elucidating the mechanistic foundations, enhancing the biomaterial component, and conducting thorough safety evaluations. We expect that these coordinated efforts will not only confirm and build on the current findings but also lay the groundwork for the creation of effective and tailored DPSC-based therapies for complicated craniofacial reconstruction.
Author Contributions
Conceptualization, Tarig Bilal. and Abdulaziz I A. Buwashl.; methodology, Mohammed A. Alholaibi.; software, Abdulaziz I A. Buwashl.; validation, Tarig Bilal., and Sanusi M. Bello.; formal analysis, Tarig Bilal.; investigation, Sanusi M. Bello.; resources, X.X.; data curation, X.X.; writing—original draft preparation, Abdulaziz I A. Buwashl and Mohammed A. Alholaibi .; writing—review and editing, Sanusi M. Bello.; visualization, Sanusi M. Bello.; supervision, Tarig Bilal.; project administration, Tarig Bilal. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board (or Ethics Committee) of King Faisal University (protocol code KFU-2025-ETHICS3577).” for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is available upon request.
Acknowledgments
The authors acknowledge the support given by the department of Biomedical Sciences, College of dentistry, the department of Surgery and Diagnostic Sciences, College of Dentistry, and King Faisal University for their invaluable support throughout the research. During the preparation of this manuscript/study, the author(s) used Quill Bot premium for the purposes of grammar and sentence fluency and grammar error management. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| (DPSCs): Dental Pulp Stem Cells |
| (DFSCs): Dental Follicle Stem Cells |
| (PDLSCs): Periodontal Ligament Stem Cells |
| (SCAP): Stem Cells from the Apical Papilla |
| (HSV-1) Herpes Simplex Virus type 1 |
| (MSCs) Mesenchymal Stem Cells |
| (BM-MSCs) Bone Marrow derived Stem Cells |
| (RUNX2) Runt-Related transcription factor 2 |
| (COL1A1) Collagen Type 1 Alpha 1 |
| (SPP1) Secreted Phosphoprotein. Also known as Osteopontin |
| (BMPs) Bone Morphogenic Proteins |
| (WnT) Wingless-type MMTV integration site family |
| (FGF) Fibroblast Growth Factor |
| (DSPP) Dentin Sialophosphoprotein |
| (BCP) Biphasic Calcium Phosphate |
| RUNX2 – Runt-related Transcription Factor 2 |
| FGF – Fibroblast Growth Factor |
| MSC – Mesenchymal Stem Cells |
| CD73 – Ecto-5'-nucleotidase |
| CD90 – Thy-1 (Thymocyte Differentiation Antigen 1) |
| CD105 – Endoglin |
| CD34 – Hematopoietic Progenitor Cell Antigen |
| CD45 – Protein Tyrosine Phosphatase, Receptor Type C (Leukocyte Common Antigen) |
| BCP – Biphasic Calcium Phosphate |
| MOI – Multiplicity of Infection |
| α-MEM – Alpha Modification of Minimal Essential Medium Eagle |
| FBS – Fetal Bovine Serum |
| BSA – Bovine Serum Albumin |
| SDS-PAGE – Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis |
| PVDF – Polyvinylidene Fluoride |
| ECL – Enhanced Chemiluminescence |
| PFA – Paraformaldehyde |
| DAPI – 4′,6-diamidino-2-phenylindole |
| EDTA – Ethylenediaminetetraacetic Acid |
| H&E – Hematoxylin and Eosin |
| IHC – Immunohistochemistry |
| HA – Hydroxyapatite |
| β-TCP – Beta-Tricalcium Phosphate |
| PCL – Polycaprolactone |
| qRT-PCR – Quantitative Reverse Transcription Polymerase Chain Reaction |
| ALPL – Alkaline Phosphatase |
| COL1A1 – Collagen Type 1 Alpha 1 |
| BMP2 – Bone Morphogenetic Protein 2 |
| OCN – Osteocalcin |
| BV/TV – Bone Volume / Total Volume |
| Tb.Th – Trabecular Thickness |
| Tb.N – Trabecular Number |
References
- Weissman, I. L. (2000). Stem cells: Units of development, units of regeneration, and units in evolution. Cell, 100(1), 157-168. [CrossRef]
- Gronthos, S., Mankani, M., Brahim, J., Robey, P. G., & Shi, S. (n.d.). Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proceedings of the National Academy of Sciences U.S.A. [CrossRef]
- Zhang, J., Ding, H., Liu, X., Sheng, Y., Liu, X., & Jiang, C. (2019). Dental Follicle Stem Cells: Tissue engineering and immunomodulation. Stem Cells and Development, 28(15), 986-994. [CrossRef]
- Zhu, W., & Liang, M. (2015). Periodontal ligament stem cells: Current status, concerns, and future prospects. Stem Cells International, 2015, 972313. [CrossRef]
- Liu, Q., Gao, Y., & He, J. (2023). Stem Cells from the Apical Papilla (SCAPs): Past, present, prospects, and challenges. Biomedicines, 11(7), 2047. [CrossRef]
- Thapliyal, G. K. (2006). Peterson's Principles of Oral & Maxillofacial Surgery. Med J Armed Forces India, 62(1), 89. [CrossRef]
- Panesar, K., & Susarla, S. M. (2021). Mandibular fractures: Diagnosis and management. Seminars in Plastic Surgery, 35(4), 238-249. [CrossRef]
- Frontiers in Medicine. (2024). Gene and Cell Therapy (Vol. 11). [CrossRef]
- Namjoynik, A., Islam, M. A., & Islam, M. (2023). Evaluating the efficacy of human dental pulp stem cells and scaffold combination for bone regeneration in animal models: A systematic review and meta-analysis. Stem Cell Research & Therapy, 14, 132. [CrossRef]
- Kwack, K. H., & Lee, H. W. (2022). Clinical potential of dental pulp stem cells in pulp regeneration: Current endodontic progress and future perspectives. Frontiers in Cell and Developmental Biology, 10, 857066. [CrossRef]
- Min, Q., Yang, L., Tian, H., Tang, L., Xiao, Z., & Shen, J. (2023). Immunomodulatory mechanism and potential application of dental pulp-derived stem cells in immune-mediated diseases. International Journal of Molecular Sciences, 24(9), 8068. [CrossRef]
- Fujii, Y., Kawase-Koga, Y., Hojo, H., et al. (2018). Bone regeneration by human dental pulp stem cells using a helioxanthin derivative and cell-sheet technology. Stem Cell Research & Therapy, 9, 24. [CrossRef]
- Charoenlarp, P., Rajendran, A. K., & Iseki, S. (2017). Role of fibroblast growth factors in bone regeneration. Inflammation and Regeneration, 37, 10. [CrossRef]
- Oton-Gonzalez, L., Mazziotta, C., Iaquinta, M. R., Mazzoni, E., Nocini, R., Trevisiol, L., D'Agostino, A., Tognon, M., Rotondo, J. C., Martini, F. (2022). Genetics and epigenetics of bone remodeling and metabolic bone diseases. International Journal of Molecular Sciences, 23(3), 1500. [CrossRef]
- Dimitriou, R., & Giannoudis, P. V. (2013). The genetic profile of bone repair. Clinical Cases in Mineral and Bone Metabolism, 10(1), 19-21. [CrossRef]
- Wu, V., Helder, M. N., Bravenboer, N., Ten Bruggenkate, C. M., Jin, J., Klein-Nulend, J., & Schulten, E. A. J. M. (2019). Bone tissue regeneration in the oral and maxillofacial region: A review on the application of stem cells and new strategies to improve vascularization. Stem Cells International, 2019, 6279721. [CrossRef]
- Gómez-Barrena, E., Rosset, P., Müller, I., Giordano, R., Bunu, C., Layrolle, P., Konttinen, Y. T., & Luyten, F. P. (2011). Bone regeneration: Stem cell therapies and clinical studies in orthopaedics and traumatology. Journal of Cellular and Molecular Medicine, 15(6), 1266-1286. [CrossRef]
- Ferro, F., Spelat, R., & Baheney, C. S. (2014). Dental pulp stem cell (DPSC) isolation, characterization, and differentiation. Methods in Molecular Biology, 1210, 91–115. [CrossRef]
- Polezhaev, L. V. (1972). Restoration of lost regenerative capacity of dental tissues. In Loss and Restoration of Regenerative Capacity in Tissues and Organs of Animals (pp. 141–152). Keterpress.
- Bai, X., Cao, R., Wu, D., Zhang, H., Yang, F., & Wang, L. (2023). Dental pulp stem cells for bone tissue engineering: A literature review. Stem Cells International, 2023, 7357179. [CrossRef]
- Ahmed, B., Ragab, M. H., Galhom, R. A., & Hassan, H. Y. (2023). Evaluation of dental pulp stem cells behavior after odontogenic differentiation induction by three different bioactive materials on two different scaffolds. BMC Oral Health, 23(1), 252. [CrossRef]
- Liu, T. M., & Lee, E. H. (2013). Transcriptional regulatory cascades in Runx2-dependent bone development. Tissue Engineering. Part B, Reviews, 19(3), 254–263. [CrossRef]
- Ryoo, H. M., Kang, H. Y., Lee, S. K., Lee, K. E., & Kim, J. W. (2010). RUNX2 mutations in cleidocranial dysplasia patients. Oral Diseases, 16(1), 55–60. [CrossRef]
- Sodek, J., Ganss, B., & McKee, M. D. (2000). Osteopontin. Critical Reviews in Oral Biology and Medicine, 11(3), 279–303. [CrossRef]
- Brunello, G., Zanotti, F., Scortecci, G., Sapoznikov, L., Sivolella, S., & Zavan, B. (2022). Dentin particulate for bone regeneration: An in vitro study. International Journal of Molecular Sciences, 23(16), 9283. [CrossRef]
- Parsegian, K. (2023). The BMP and FGF pathways reciprocally regulate odontoblast differentiation. Connective Tissue Research, 64(1), 53–63. [CrossRef]
- Nusse, R. (2008). Wnt signaling and stem cell control. Cell Research, 18(5), 523–527. [CrossRef]
- Choi, Y. S., Lee, J. Y., Suh, J. S., Lee, G., Chung, C. P., & Park, Y. J. (2013). The mineralization inducing peptide derived from dentin sialophosphoprotein for bone regeneration. Journal of Biomedical Materials Research. Part A, 101(2), 590–598. [CrossRef]
- Cox, R. F., & Morgan, M. P. (2013). Microcalcifications in breast cancer: Lessons from physiological mineralization. Bone, 53(2), 437-450. [CrossRef]
- Mastrolia, I., Foppiani, E. M., Murgia, A., Candini, O., Samarelli, A. V., Grisendi, G., Veronesi, E., Horwitz, E. M., & Dominici, M. (2019). Challenges in clinical development of mesenchymal stromal/stem cells: Concise review. Stem Cells Translational Medicine, 8(11), 1135–1148. [CrossRef]
- Rana, N., Suliman, S., Mohamed-Ahmed, S., Gavasso, S., Gjertsen, B. T., & Mustafa, K. (2022). Systemic and local innate immune responses to surgical co-transplantation of mesenchymal stromal cells and biphasic calcium phosphate for bone regeneration. Acta Biomaterialia, 141, 440–453. [CrossRef]
- Yamada, S., Shanbhag, S., & Mustafa, K. (2022). Scaffolds in periodontal regenerative treatment. Dental Clinics of North America, 66(1), 111-130. [CrossRef]
- Yamada, S., Yassin, M. A., Schwarz, T., Hansmann, J., & Mustafa, K. (2021). Induction of osteogenic differentiation of bone marrow stromal cells on 3D polyester-based scaffolds solely by subphysiological fluidic stimulation in a laminar flow bioreactor. Journal of tissue engineering, 12, 20417314211019375. [CrossRef]
- Shanbhag, S., Suliman, S., Pandis, N., Stavropoulos, A., Sanz, M., & Mustafa, K. (2019). Cell therapy for orofacial bone regeneration: A systematic review and meta-analysis. Journal of clinical periodontology, 46 Suppl 21, 162–182. [CrossRef]

Table 1.
qRT-PCR fold-change (vs Day 0) for osteogenic genes during DPSC osteogenic induction (mean ± SD).
Table 1.
qRT-PCR fold-change (vs Day 0) for osteogenic genes during DPSC osteogenic induction (mean ± SD).
| Gene | Day 0 | Day 7 (fold ± SD) | Day 14 (fold ± SD) | Day 21 (fold ± SD) | Notes |
| RUNX2 | 3.9 ± 0.4 | 4.2 ± 0.5 | 6.8 ± 0.6 | 5.9 ± 0.7 | Transcription factor — early/mid osteogenic marker. |
| ALPL (ALP) | 3.1 ± 0.4 | 3.5 ± 0.4 | 7.1 ± 0.8 | 4.5 ± 0.6 | Enzymatic marker of osteoblastic activity. |
| COL1A1 | 2.7 ± 0.3 | 2.8 ± 0.3 | 5.2 ± 0.5 | 3.9 ± 0.4 | Major bone matrix collagen. |
| BMP2 | 2.9 ± 0.4 | 3.1 ± 0.4 | 6.4 ± 0.7 | 4.2 ± 0.5 | Osteoinductive growth factor. |
| OCN (BGLAP) | 1.3 ± 0.2 | 1.5 ± 0.2 | 4.7 ± 0.6 | 7.8 ± 0.9 | Late osteogenic/mineralization marker. |
All comparisons vs Day 0.
Table 2.
Micro-CT morphometric parameters for mandibular defects treated with BCP only (Control), BCP + DPSCs, and BCP + RUNX2-DPSCs at weeks 4 and 8 (mean ± SD, n = 6 per group per timepoint). BV/TV = bone volume / total volume (%); Tb.Th = trabecular thickness (µm); Tb.N = trabecular number (1/mm). Statistical analysis: one-way ANOVA with Tukey’s post hoc test.
Table 2.
Micro-CT morphometric parameters for mandibular defects treated with BCP only (Control), BCP + DPSCs, and BCP + RUNX2-DPSCs at weeks 4 and 8 (mean ± SD, n = 6 per group per timepoint). BV/TV = bone volume / total volume (%); Tb.Th = trabecular thickness (µm); Tb.N = trabecular number (1/mm). Statistical analysis: one-way ANOVA with Tukey’s post hoc test.
| Group | BV/TV (%) at Week 4 | BV/TV (%) at Week 8 | Tb.Th (µm) at Week 8 | Tb.N (1/mm) at Week 8 |
|---|---|---|---|---|
| Control (BCP only) | 22.4 ± 2.1 | 31.7 ± 2.8 | 78.5 ± 5.1 | 2.4 ± 0.3 |
| DPSC + BCP | 38.9 ± 3.2* | 52.3 ± 3.7* | 112.7 ± 6.4* | 3.8 ± 0.4* |
| RUNX2-DPSC + BCP | 45.6 ± 3.8*,† | 63.9 ± 4.5*,† | 128.3 ± 7.1*,† | 4.6 ± 0.5*,† |
* p < 0.01 vs. Control; † p < 0.05 vs. DPSC.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



