Submitted:
24 December 2025
Posted:
26 December 2025
You are already at the latest version
Abstract
This systematic review identifies, evaluates, and synthesizes evidence on upland rice (Oryza sativa L.) genotypes resistant to neck blast disease caused by Magnaporthe oryzae. The review aimed to determine resistant genotypes, screening methods, and genetic fac-tors influencing resistance variability across field and greenhouse studies. Eligible studies were empirical investigations assessing upland rice under upland or rainfed conditions, published in English between 1980 and 2025. Searches were conducted in PubMed, Sci-enceDirect, Google Scholar, and grey literature, with final searches completed on October 31, 2025. Risk of bias was assessed using a customized framework adapted from ROB-INS-I tool, and results were synthesized narratively due to heterogeneity in methods and reporting. Six studies involving 248 genotypes from Asia and Africa met the inclusion cri-teria. Twenty genotypes, including Kahei, Barkhe 1032, 1035, 2014, NERICA lines, and BC1F4 backcross derivatives, showed moderate to high resistance based on the IRRI 0–9 Standard Evaluation System. Two studies identified key quantitative trait loci (qBFR4-1, qBl1, qBl2) linked to durable resistance. Although most studies exhibited low risk of bias, two presented minor concerns in randomization and environmental control. Evidence was limited by small sample sizes, methodological heterogeneity, and restricted molecu-lar characterization. Nevertheless, the findings underscore the potential of QTL-based breeding and marker-assisted selection for improving upland rice resilience in blast-prone regions. Future research should expand genomic screening, harmonize protocols, and validate resistant lines across environments.
Keywords:
Magnaporthe oryzae
; neck blast resistance
; rainfed cropping systems
; genotype screening
; field and greenhouse evaluation
; quantitative trait loci
; marker-assisted selection
1. Introduction
Rice (Oryza sativa L.) is a staple food crop that sustains over half of the world’s population, with upland rice playing a critical role in rain-fed ecosystems, particularly in regions with limited irrigation infrastructure such as Southeast Asia, sub-Saharan Africa, and Latin America [1,2]. Upland rice cultivation, which accounts for approximately 12% of global rice production, is often practiced on marginal lands characterized by variable soil fertility, erratic rainfall, and exposure to abiotic stresses like drought and aluminum toxicity [3,4]. Despite its adaptability, upland rice productivity remains low, averaging 2-3 tons per hectare in many developing countries, compared to 5-6 tons per hectare for lowland varieties [5,6]. This disparity arises from multiple constraints, including suboptimal agronomic practices, nutrient deficiencies, and biotic stresses, with blast disease caused by the fungal pathogen Magnaporthe oryzae emerging as one of the most devastating factors limiting yield potential.
Blast disease manifests in various forms, including leaf blast, node blast, and neck (or panicle) blast, with neck blast being particularly insidious due to its impact on reproductive structures. Neck blast infects the panicle neck, leading to partial or complete sterility of grains, chalky kernels, and broken panicles, which can result in yield losses ranging from 10% to 80% under conducive environmental conditions [7,8,9,10]. The pathogen thrives in humid, temperate environments typical of upland ecologies, where high nitrogen fertilization, dense planting, and temperature fluctuations between 20-30 °C facilitate spore dissemination and infection [11,12]. Historically, blast has been documented across major rice-growing continents, with early records in China (1637), Japan (1704), and Africa (1922), and it continues to pose a global threat, causing annual economic losses exceeding $70 billion [6,13,14]. In upland systems, the disease is exacerbated by the absence of flooding, which otherwise suppresses pathogen survival in lowland fields, making upland rice inherently more vulnerable [15,16].
The escalating demand for rice, driven by population growth, urbanization, and shifting dietary preferences, underscores the urgency to enhance upland rice resilience. Global rice consumption is projected to increase by 26% to 116 million tons by 2035, with Africa experiencing a 130% surge in demand [17]. In regions like Indonesia and Kenya, where upland rice contributes modestly to national production but is vital for food security in remote areas, domestic output meets only 20-80% of needs, necessitating substantial imports [17,18]. Addressing these gaps requires targeted interventions against biotic constraints, as chemical fungicides, while effective, raise environmental concerns, increase production costs, and promote pathogen resistance [19,20,21]. Consequently, breeding for host resistance has emerged as the most sustainable and cost-effective strategy, emphasizing the integration of durable resistance traits into high-yielding varieties [11,22,23].
Resistance to blast disease is broadly categorized into qualitative (complete or true) resistance, governed by major R-genes that confer race-specific immunity, and quantitative (partial or field) resistance, involving multiple quantitative trait loci (QTLs) that provide broad-spectrum durability [23,24]. Qualitative resistance, while potent against compatible pathogen races, is prone to breakdown due to rapid fungal evolution, as observed in monoculture systems [11,25]. In contrast, field resistance suppresses disease progression without complete immunity, offering longevity in diverse environments, as exemplified by QTLs like qBFR4-1 (Pikahei-1(t)) in the upland cultivar Kahei, which maps to chromosome 4 and confers strong suppression under natural infection [23,26]. Over 122 blast resistance genes have been identified, including major genes such as Pi9, Pi2, Piz-t, and Pid3, and minor QTLs like Pi30(t) and Pi31(t), with at least 39 cloned for molecular characterization [4,21,27,28]. Nucleotide-binding site-leucine-rich repeat (NBS-LRR) proteins often underlie these resistances, highlighting the genetic basis for pathogen recognition and defense activation [23].
Screening for neck blast resistance in upland rice typically employs field and greenhouse assays to simulate natural and controlled infections, respectively. Field evaluations, conducted in blast-prone hotspots with natural inoculum or augmented spore applications, assess durability across seasons but are limited by environmental variability and annual cycles [23,29]. Greenhouse methods, utilizing spore suspensions or infected donor plants, enable precise phenotyping, often grading symptoms on a 0-9 scale, and facilitate rapid screening of segregating populations [9,10,23]. Correlations between leaf and neck blast resistance vary, with some studies reporting positive associations and others noting independent reactions, necessitating targeted evaluations for panicle-specific traits [6,9,30,31,32,33]. Advanced breeding techniques, including anther culture for doubled haploid production and marker-assisted selection (MAS) for gene pyramiding, accelerate the development of resistant lines, as demonstrated in varieties like Morkhor 60-3, which incorporates QTLs qBl1 and qBl2 for enhanced blast tolerance [3,4,34].
Despite these advances, inconsistencies in reported resistant genotypes, screening protocols, and genetic mechanisms across studies hinder progress in upland rice improvement. Variations in pathogen isolates, environmental conditions, and evaluation criteria contribute to discrepancies, underscoring the need for a synthesis of existing literature. This systematic review addresses this gap by compiling evidence from field and greenhouse studies to identify upland rice genotypes resistant to neck blast, delineate employed screening methodologies, and expose genetic factors influencing resistance variability.
2. Materials and Methods
2.1. Eligibility Criteria
This systematic review followed a predefined protocol to ensure transparency and reproducibility in the selection of studies. Eligibility criteria were established based on the Population, Intervention/Exposure, Comparator, Outcome, and Study Design (PICOS) framework, tailored to the review’s focus on identifying upland rice genotypes resistant to neck blast disease through field and greenhouse evaluations [35].
Studies were included if they focused on upland rice (Oryza sativa L.) genotypes, such as traditional varieties, improved lines, breeding populations, or wild relatives evaluated specifically in upland or rain-fed conditions. Genotypes from other rice ecotypes, like lowland or irrigated varieties, were considered only if they were explicitly tested under upland-like simulations. Evaluations had to assess resistance to neck blast (panicle blast) caused by Magnaporthe oryzae, encompassing natural infections, artificial inoculations, or molecular characterizations linked to neck blast phenotypes. Studies with or without comparators were eligible, though preference was given to those incorporating susceptible checks, resistant standards (for example, varieties with known R-genes like Pi9 or Pi2), or multi-environment trials for benchmarking resistance levels.
Primary outcomes required the identification of resistant genotypes, rated as resistant or highly resistant on standard scales like the 0-9 SES, along with yield loss estimates due to neck blast, genetic markers or QTLs associated with resistance, and details on screening methodologies. Secondary outcomes included correlations between leaf and neck blast resistance, pathogen isolate variability, and environmental influences on resistance expression. Eligible study designs comprised empirical research reporting original data from field trials, such as hotspot screenings or multi-location tests, or greenhouse experiments involving controlled inoculations or seedling trays. Peer-reviewed journal articles, conference proceedings, theses, and technical reports published in English from January 1980 to September 2025 were considered to capture both historical and recent advancements. Additionally, studies needed to provide sufficient detail on methodology, including inoculation techniques and rating scales, as well as results like genotype lists and resistance scores, to enable data extraction and synthesis [36].
Studies were excluded if they concentrated exclusively on leaf blast, node blast, or other rice diseases without specific data on neck blast. Evaluations limited to non-upland rice systems, such as purely lowland or flooded conditions, were omitted unless upland adaptations were simulated and reported. Non-empirical works, including narrative reviews, opinion pieces, or modeling studies without experimental validation, did not qualify. Research lacking quantitative or qualitative resistance data, such as mere disease incidence reports without genotype-specific assessments, was also excluded. Publications in languages other than English or those with inaccessible full texts after reasonable efforts, like interlibrary loans or author contacts, were not included. Duplicate reports of the same dataset were handled by retaining the most comprehensive or recent version. For syntheses, studies were grouped to facilitate narrative and thematic analyses, as meta-analysis was not feasible due to heterogeneity in screening protocols and reporting standards [37].
The primary grouping distinguished between field-based studies, which emphasized natural inoculum and multi-seasonal durability, and greenhouse-based studies, which focused on controlled conditions for precise phenotyping. Within these categories, subgroups were formed based on geographical regions, such as Asia, Africa, and Latin America, to account for agroecological variations and pathogen diversity. Additional groupings addressed resistance type, differentiating qualitative or major gene resistance from quantitative or QTL-based resistance; screening intensity, such as single versus multi-isolate challenges; and genetic focus, including studies reporting cloned genes or applications of marker-assisted selection. This stratification allowed for the identification of consistent resistant genotypes across contexts, highlighting methodological trends and exposing knowledge gaps for future research [38].
2.2. Information Sources
The search for relevant studies was conducted across multiple electronic databases, and grey literature sources to ensure comprehensive coverage of the literature on upland rice resistance to neck blast, adhering to recommended practices for systematic reviews in agricultural sciences [39]. Key databases included PubMed for biomedical and pathology-related studies, ScienceDirect for peer-reviewed journal articles in agronomy and crop science, and Google Scholar for broad academic coverage including theses and conference papers. Additionally, reference lists from included studies and seminal review articles on rice blast resistance were hand-searched for supplementary citations. Grey literature, including theses from ProQuest Dissertations & Theses Global and reports from national agricultural research institutes, was screened to minimize publication bias. The search dates were from January 1980 to September 2025 and searches were confined to English-language publications. The final searches across all sources were completed on October 31, 2025. A total of 420 records were initially identified from the primary databases: 200 from PubMed, 10 from ScienceDirect, and 210 from Google Scholar. Subsequent deduplication using Rayyan software removed 07 redundant records before full-text screening commenced.
2.3. Search Strategy
A comprehensive and reproducible search strategy was developed to identify empirical field and greenhouse studies that evaluated upland Oryza sativa genotypes for resistance to neck (panicle) blast caused by Magnaporthe oryzae. The strategy was constructed a priori using the PICOS framework to define the population, exposure, outcomes and eligible study designs, and was iteratively refined through pilot searches to balance sensitivity and specificity [35]. Search planning and reporting followed current guidance for systematic reviews to ensure transparency and replicability [39]. Search terms combined controlled vocabulary and free-text terms covering the crop (Oryza sativa, rice), pathogen (Magnaporthe oryzae, Pyricularia oryzae), target organ (neck, panicle), the ecotype (upland, rain-fed), screening environment descriptors (field, hotspot, greenhouse, glasshouse, controlled inoculation) and molecular/genetic keywords (SNP, single nucleotide polymorphism, QTL, marker, R-gene).
Electronic searching was conducted across multiple bibliographic databases and platforms relevant to agricultural science and plant pathology, including PubMed for biomedical/pathology literature and ScienceDirect for agronomy and crop science journals. Google Scholar was used to extend coverage to theses, conference papers and other grey literature sources, and ProQuest Dissertations & Theses Global and national agricultural research institute repositories were searched to capture dissertations and technical reports. The search strings were adapted to PubMed database’s syntax and indexing. For example, the search string was: (“upland rice” OR “upland rice” OR “rainfed rice” OR “Oryza sativa”) AND (“Magnaporthe oryzae” OR “neck blast” OR “panicle blast” OR “Pyricularia oryzae”) AND (“plant resistance” OR resistance OR “resistant genotype*” OR QTL OR “R gene”) AND (“field trial*” OR greenhouse OR screening) NOT (“lowland rice” OR irrigated). On the other hand, title/abstract/keyword fields were used in ScienceDirect and Google Scholar.
2.4. Selection Process
Study selection proceeded through a two-stage screening process (title/abstract followed by full-text assessment) implemented to ensure consistent and reproducible application of the pre-specified eligibility criteria (See Section 2.1). Following export from the database interfaces into reference-management software and initial deduplication, records were uploaded to Rayyan for blinded screening and collaborative management [40]. Deduplication removed seven redundant records from the 420 records retrieved during the primary electronic searches; the remaining unique records entered the title/abstract screening stage.
Title and abstract screening was performed independently by two reviewers. Prior to formal screening, the review team conducted a calibration exercise using a random sample of records to verify shared understanding of the inclusion/exclusion criteria, to refine the screening form, and to ensure consistency in judgement. During the main screening phase each reviewer classified records as “include”, “exclude” or “uncertain”; Rayyan’s interface was used to blind reviewers to one another’s decisions and to facilitate the management of conflicts [40]. Records judged as “include” or “uncertain” by either reviewer progressed to full-text assessment. Rayyan was employed for deduplication, blinded dual screening and conflict tracking.
2.5. Data Collection Process
Data extraction from the included studies was conducted using a standardized electronic form developed a priori and piloted on a subset of eligible reports to ensure clarity, completeness, and consistency in capturing relevant information. The form was designed in Microsoft Excel to facilitate structured data entry and was tailored to the review’s objectives, encompassing key domains such as study characteristics, screening methodologies, genotype details, resistance outcomes, and genetic factors (see Section 2.6 for specific data items). Two reviewers independently extracted data from each included report to minimize bias and enhance accuracy. Prior to the main extraction phase, a calibration exercise was performed on five randomly selected studies to align interpretations and resolve any ambiguities in the form’s application.
For each report, reviewers worked separately, documenting extractions in duplicate forms before comparing results. Discrepancies between the extracted data were identified through comparison and resolved via discussion between the two reviewers; if consensus could not be reached, a third senior reviewer was consulted for arbitration. No automation tools were employed for the data extraction process itself, as it relied on manual interpretation of textual and tabular content to ensure specific understanding of complex agronomic and genetic details. However, Rayyan software, previously used for screening [40], was leveraged for organizing full-text PDFs and tracking extraction progress, facilitating collaborative workflow management among the review team. All extracted data were compiled into a central database for subsequent synthesis and analysis.
2.6. Data Items
Data items were predefined based on the review’s objectives and structured to capture both outcome-related and contextual information from eligible studies. Extraction focused on quantitative and qualitative elements essential for synthesizing evidence on upland rice resistance to neck blast, with all items documented in a piloted extraction form.
For primary outcomes, data were sought on the identification of resistant upland rice genotypes, defined as those classified as resistant (R), moderately resistant (MR), or highly resistant (HR) based on standard rating scales such as the International Rice Research Institute’s Standard Evaluation System (SES) 0-9 scale (where scores of 0-3 indicate resistance) or equivalent visual symptom-based metrics (e.g., percentage of infected panicles or disease severity index). This included any reported resistant genotypes, along with associated yield loss estimates attributable to neck blast (e.g., percentage reduction in grain yield or filled grains per panicle under infected conditions). Additional primary outcomes encompassed genetic markers or QTLs linked to neck blast resistance (e.g., specific R-genes like Pi9 or QTLs like qBFR4-1, including chromosomal locations and effect sizes where available), and details on screening methodologies (e.g., inoculation techniques, such as spore suspension concentration in greenhouse assays or natural hotspot exposure in field trials, along with evaluation timings like days after heading).
Secondary outcomes included correlations between leaf and neck blast resistance (e.g., Pearson’s correlation coefficients or qualitative associations), pathogen isolate variability (e.g., race or avirulence gene profiles of Magnaporthe oryzae strains used), and environmental influences on resistance expression (e.g., impacts of temperature, humidity, or nitrogen levels on disease severity). All results compatible with these outcome domains were sought from each study, including multiple measures (e.g., different rating scales), time points (e.g., initial vs. final disease assessments), and analyses (e.g., single-season vs. multi-year data, or isolate-specific vs. mixed-inoculum results). No selective collection methods were applied; if multiple results were reported for an outcome (e.g., resistance scores across different years or locations), all were extracted to enable comprehensive synthesis and assessment of consistency or variability.
Other variables for which data were sought included study characteristics such as publication year, geographical location (e.g., country or region like Southeast Asia or sub-Saharan Africa), study design (e.g., field trial, greenhouse experiment, or combined), number of genotypes evaluated, comparator details (e.g., susceptible checks like IR64 or resistant standards like Moroberekan), funding sources (e.g., institutional, governmental, or private grants), and author affiliations. Genotype-specific variables encompassed origin (e.g., traditional landrace, improved cultivar, or breeding line), and agronomic traits (e.g., maturity duration or yield potential under non-diseased conditions). Methodological variables included pathogen inoculum details (e.g., isolate source, concentration, or application method), environmental conditions (e.g., temperature ranges or rainfall patterns in field studies), and statistical analyses (e.g., ANOVA for resistance differences or heritability estimates). For missing or unclear information, assumptions were made conservatively: for instance, if resistance ratings were not explicitly linked to a scale, it was assumed to align with the SES 0-9 system if contextually implied (e.g., based on cited IRRI protocols). All assumptions were documented in the extraction form and considered during risk of bias assessment to mitigate potential misinterpretation.
2.7. Study Risk of Bias Assessment
The risk of bias (RoB) in included studies was assessed to evaluate the internal validity of the evidence and inform the strength of synthesized findings. Given the predominance of experimental (field and greenhouse) rather than randomized controlled trial designs in agronomic research, a customized RoB tool was developed a priori, adapted from established framework such as the Risk of Bias in Non-randomized Studies of Interventions (ROBINS-I) tool [41], tailored to the context of plant pathology and crop breeding studies. This tool comprised seven domains relevant to field and greenhouse evaluations of disease resistance: (1) selection of genotypes (e.g., representativeness and randomization in trial design), (2) exposure to pathogen (e.g., standardization and reproducibility of inoculation methods), (3) confounding factors (e.g., control for environmental variables like temperature or humidity), (4) detection/outcome assessment (e.g., blinding of raters and reliability of scoring scales), (5) incomplete outcome data (e.g., attrition of genotypes or missing yield assessments), (6) selective reporting (e.g., completeness of resistance data across all tested lines), and (7) other biases (e.g., funding conflicts or methodological deviations). Each domain was rated as low risk, moderate risk, high risk, or unclear risk, with supporting justifications extracted from the study reports. An overall RoB judgment was assigned per study as low (all domains low), moderate (mostly low with some moderate), high (one or more high domains), or unclear (insufficient information).
Two reviewers independently assessed the RoB for each included study using the customized tool implemented in a standardized Microsoft Excel form, which was piloted on three studies to refine domain definitions and ensure consistent application. Assessments were conducted blindly to one another’s ratings, with discrepancies resolved through discussion to achieve consensus; if unresolved, a third senior reviewer arbitrated based on predefined resolution criteria (e.g., prioritizing explicit methodological descriptions). No automation tools were used in the RoB assessment process, as it required qualitative judgment of contextual detail. RoB results were summarized narratively and tabulated by study type (field vs. greenhouse) to highlight patterns influencing evidence certainty.
2.8. Effect Measures
Due to substantial heterogeneity in study designs, screening protocols, pathogen isolates, and reporting formats across the included studies, a quantitative meta-analysis was not feasible, as noted in Section 2.1. Instead, results were synthesized narratively and thematically, with effect measures presented descriptively to highlight patterns in resistance expression, genetic contributions, and methodological variations. Effect measures were extracted directly from primary reports and categorized by outcome domain, focusing on those most commonly reported for comparability. The following discussion specifies effect measures for each primary and secondary outcome, drawing from the included studies [3,4,9,10,17].
For primary outcomes, resistance identification was primarily quantified using mean disease scores on the 0-9 SES scale, where scores ranging from 0 (no symptoms, indicating high resistance) to 9 (severe infection, indicating high susceptibility) were employed, with lower scores signifying greater resistance [3,4,23]. Proportions or percentages of resistant, moderately resistant, or susceptible lines were also reported, often benchmarked against susceptible checks, such as IR64, or resistant standards, such as Moroberekan [9]. Descriptive ratings, including categories like R for resistant or MR for moderately resistant, were used alongside quantitative scores for categorical classification. Yield loss estimates due to neck blast were expressed as percentage reductions in grain yield or filled grains per panicle under infected versus non-infected conditions, supplemented by mean differences in agronomic traits such as panicle number or 1,000-grain weight [4,17].
These metrics were frequently analyzed through post-hoc tests, such as Tukey’s Honestly Significant Difference (HSD) at P ≤ 0.05, to compare genotype performance. Genetic markers or quantitative trait loci (QTLs) associated with resistance were evaluated using effect sizes reported as logarithm of odds (LOD) scores and coefficients of determination (R2), which represented the phenotypic variance explained (e.g., R2 > 10% for significant QTLs like qBFR4-1) [23]. Inheritance patterns in segregating populations were assessed via Chi-square (χ2) tests for goodness-of-fit to expected ratios, such as 3:1 for single-gene resistance [17], while broad-spectrum resistance (BSR) indices aggregated responses across multiple isolates to evaluate gene pyramiding efficacy [4]. Screening methodologies were described qualitatively, including details such as spore concentration in inocula or days post-inoculation for assessments, but were quantified where possible through mean differences in disease severity or incidence across inoculation methods (e.g., natural versus artificial) or environments (field versus greenhouse) [3,4].
Analysis of variance (ANOVA) significance at P ≤ 0.05 was commonly applied to test the effects of these methods on resistance scores. Secondary outcomes were similarly quantified to capture associative and contextual factors. Correlations between leaf and neck blast resistance were measured using Pearson’s correlation coefficients (r), with significance tested at P = 0.05 (e.g., r = 0.30) indicating a weak positive correlation [9]. Pathogen isolate variability was compared through mean differences in lesion scores or proportions of compatible/incompatible reactions to specific races or isolates (e.g., race 173), often summarized descriptively without formal effect measures [3,4]. Environmental influences on resistance expression were reported as mean differences in disease incidence or severity under varying conditions, such as temperature or humidity gradients, with cluster similarity indices (%) employed for grouping genotypes by environmental adaptation [10,17].
3.1. Study Selection
Searches and study selection were performed following PRISMA guidance [39]. A PRISMA flow diagram summarizing identification, screening, eligibility assessment and final inclusion is presented in Figure 1. From the studies and extraction table provided for this review, six primary studies met the review’s inclusion criteria and were retained for synthesis. The six included studies [3,4,6,9,10,23] originate from six countries across Asia and Africa (Indonesia, Kenya, Japan, Thailand, Nepal, Malaysia) and collectively evaluated a broad set of upland rice genotypes for resistance to neck (panicle) blast. Study designs and methods varied: three studies used greenhouse-based inoculation systems [4,9,23], at least two applied field screening [10,17], and one combined anther culture with inoculation [3]. Molecular approaches ranged from QTL mapping and graphical genotyping [23] to marker-assisted backcross selection using known QTL/genes [4]. Several studies did not report specific molecular markers used for screening [3,9,10,17].
Several studies identified during the full-text screening phase initially appeared to align with the inclusion criteria—such as focusing on rice blast resistance evaluations involving Magnaporthe oryzae and genotype assessments—but were ultimately excluded upon closer examination. These exclusions were documented systematically to enhance transparency and reproducibility, with reasons categorized based on deviations from the predefined PICOS framework. In total, eleven studies were excluded at full-text assessment: three because the full text was inaccessible for extraction [42,43,44], two because their outcomes addressed traits other than neck (panicle) blast or did so in a manner inconsistent with our outcome definition [45,46], and six because the study populations or study types did not comprise upland rice genotype screening data suitable for synthesis [29,34,47,48,49,50].
3. Results
3.1. Study Selection
Searches and study selection were performed following PRISMA guidance [39]. A PRISMA flow diagram summarizing identification, screening, eligibility assessment and final inclusion is presented in Figure 1. From the studies and extraction table provided for this review, six primary studies met the review’s inclusion criteria and were retained for synthesis. The six included studies [3,4,9,10,17,23] originate from six countries across Asia and Africa (Indonesia, Kenya, Japan, Thailand, Nepal, Malaysia) and collectively evaluated a broad set of upland rice genotypes for resistance to neck (panicle) blast. Study designs and methods varied: three studies used greenhouse-based inoculation systems [4,9,23], at least two applied field screening [10,17], and one combined anther culture with inoculation [3]. Molecular approaches ranged from QTL mapping and graphical genotyping [23] to marker-assisted backcross selection using known QTL/genes [4]. Several studies did not report specific molecular markers used for screening [3,9,10,17].
Several studies identified during the full-text screening phase initially appeared to align with the inclusion criteria—such as focusing on rice blast resistance evaluations involving Magnaporthe oryzae and genotype assessments—but were ultimately excluded upon closer examination. These exclusions were documented systematically to enhance transparency and reproducibility, with reasons categorized based on deviations from the predefined PICOS framework. In total, eleven studies were excluded at full-text assessment: three because the full text was inaccessible for extraction [42,43,44], two because their outcomes addressed traits other than neck (panicle) blast or did so in a manner inconsistent with our outcome definition [45,46], and six because the study populations or study types did not comprise upland rice genotype screening data suitable for synthesis [29,34,47,48,49,50].
3.2. Study Characteristics
Six studies satisfied the inclusion criteria and were retained for detailed extraction. Collectively, these investigations span a broad geographic range (Indonesia, Kenya, Japan, Thailand, Nepal and Malaysia) and encompass diverse experimental designs, sample sizes and screening protocols (See Table 1 & Figure 2). Sample sizes varied markedly across studies (from 33 genotypes in field-based evaluations to 182 breeding lines screened in greenhouse assays), and screening approaches included field spray inoculation with IRRI scoring, controlled greenhouse inoculation of recombinant inbred lines (RILs) or breeding material, anther-culture derived evaluations, and marker-assisted backcross selection. Molecular and genetic work was reported in only a subset of studies: [23] performed QTL mapping and fine-mapping (identifying the qBFR4-1 region), and [4] employed marker-assisted backcross selection using QTL markers (qBl1, qBl2) and the xa5 locus.
[3] developed novel upland rice lines resistant to blast through anther culture in Indonesia, testing parental genotypes including Fatmawati, Way Rarem, SGJT-36, and SGJT-28. Their efforts yielded five highly resistant doubled haploid lines (P3-27, P5-50, P6-105, P3-162, and P3-204), screened via inoculation with three pathogen isolates, confirming resistance without reliance on specific molecular markers; phenotypic outcomes aligned fully with selection criteria. In a Kenyan field study, [17] evaluated 33 rice genotypes for blast resistance and inheritance patterns in the Mwea region, identifying Duorado precoce, B6-144, IRAT 109, and several NERICA varieties as exhibiting varied resistance levels to both leaf and panicle blast. Assessments employed spray inoculation and the IRRI scoring system, with no molecular markers specified, yet resistance in NERICA lines showed complete consistency between inheritance data and phenotypic expression. [23] fine-mapped a major QTL for field resistance in Japan, focusing on the upland cultivar Kahei crossed with Koshihikari, and confirmed Kahei’s very high resistance through greenhouse inoculations of F7 recombinant inbred lines (RILs). Utilizing QTL mapping, residual heterozygosity analysis, and graphical genotyping, they precisely delimited the qBFR4-1 region on chromosome 4, with genotypic and phenotypic data demonstrating full concordance.
[4] advanced the Thai upland variety Morkhor 60-3 for dual resistance to blast and bacterial blight via marker-assisted backcross selection, incorporating Morkhor 60-1 as a recurrent parent. Three BC1F4 lines (22-7-140-4, 22-7-322-5, and 22-7-311-9) exhibited high resistance, validated through greenhouse inoculations, with QTL markers qBl1, qBl2, and xa5 enabling successful pyramiding of blast and bacterial blight genes; marker-trait associations were robust, yielding perfect genotypic-phenotypic alignment. [9] assessed 182 tropical rice breeding lines in Nepal under greenhouse conditions for reactions to leaf and neck blast, pinpointing five highly resistant genotypes from the Barkhe series (1032, 1034, 1035, 1036, and 2014). Inoculation protocols targeted both blast types without specified molecular markers, revealing a partial correlation between leaf and neck resistance, which translated to only partial consistency between inferred genetic traits and phenotypic scores. Finally, [10] analyzed reactions and genetic diversity among various upland rice genotypes in Malaysia, identifying Barkhe 1032, 1035, and 3004 as moderately to highly resistant through combined field screening in hotspots and greenhouse testing.
3.3. Risk of Bias in Studies
Risk of bias (RoB) assessments were conducted for each of the six included studies using the customized tool outlined in Section 2.7, which evaluated domains pertinent to agronomic evaluations: bias arising from the randomization process (D1; e.g., genotype allocation), deviations from intended interventions (D2; e.g., inoculation fidelity), missing outcome data (D3; e.g., genotype attrition), measurement of outcomes (D4; e.g., rater blinding and scale reliability), and selection of reported results (D5; e.g., completeness of resistance metrics). Overall RoB judgments were low for four studies and raised some concerns for two, primarily due to ambiguities in randomization, environmental controls, or outcome measurement in field-based assessments. No studies exhibited high RoB, reflecting generally robust methodological reporting, though concerns in D2 and D4 highlighted potential influences from uncontrolled variables in greenhouse and field settings (see Figure 3; & Table 2).
[3] demonstrated low RoB across all domains. Genotype selection via anther culture was systematic without randomization needs, inoculation with three isolates adhered closely to protocols, and no missing data were reported. Outcome measurements using the 0-9 SES scale were standardized without blinding issues, and all resistance results for doubled haploid lines were fully reported, yielding an overall low RoB.
[17] raised some concerns overall, driven by uncertainties in D1, D2, and D4. Field genotype allocation in the Mwea trials lacked explicit randomization details, potentially introducing selection bias among the 33 lines; deviations from spray inoculation protocols were noted due to variable weather, and disease severity scoring via IRRI methods may have been influenced by non-blinded assessors. However, missing data (D3) and reporting (D5) were low risk, with complete inheritance ratios provided.
[23] exhibited low RoB throughout. Recombinant inbred line (RIL) development in the F7 generation ensured balanced randomization (D1), while greenhouse inoculations followed a novel evaluation system with minimal deviations (D2). No outcome attrition occurred (D3), lesion scoring was objective and blinded where feasible (D4), and QTL mapping results, including LOD scores, were comprehensively reported (D5), supporting an overall low judgment.
[4] achieved the lowest RoB profile, with no concerns across domains. Marker-assisted backcrossing for BC1F4 lines incorporated rigorous randomization in selection cycles (D1), precise greenhouse inoculations for 12 isolates minimized deviations (D2), and full phenotypic data on resistance indices were captured without gaps (D3). Measurements via BSR indices were reliable and blinded (D4), with all pyramided QTL associations (qBl1, qBl2) transparently reported (D5).
[9] warranted some concerns overall, stemming from D1, D2, and D3. Allocation of 182 breeding lines for greenhouse testing was not fully randomized, risking clustering effects; inoculation protocols for leaf and neck blast showed minor deviations due to humidity fluctuations, and some panicle data were incomplete for moderately resistant lines (D3). Outcome measurement (D4) and reporting (D5) posed low risks, with correlations fully detailed, though partial genotypic-phenotypic alignment amplified interpretive caution.
[10] presented low overall RoB, with isolated some concerns in D2. Genotype diversity analysis ensured representative selection (D1), field-greenhouse protocols yielded complete data (D3), and lesion typing was consistently scored (D4), with all diversity indices reported (D5). Minor deviations arose from variable field inoculum loads, but these did not compromise core resistance findings for Barkhe lines. These RoB patterns suggest high confidence in molecularly informed studies e.g. [4,23] and moderate confidence in phenotypic screenings e.g. [9,17], with implications for prioritizing durable QTLs in future syntheses.
3.4. Upland Rice Genotypes Identified as Resistant to Neck Blast Disease Under Field and Greenhouse Screening Conditions
This section answers the research question: Which upland rice (Oryza sativa L.) genotypes have been identified as resistant to neck blast disease (Magnaporthe oryzae) under field and greenhouse screening conditions?
Across the six included studies, a total of 20 distinct upland rice genotypes were identified as resistant to neck blast disease caused by Magnaporthe oryzae, with resistance levels varying from moderate to very high based on standardized evaluations such as the IRRI 0-9 Standard Evaluation System (SES) scale. These genotypes were screened under diverse conditions, including controlled greenhouse inoculations for precise isolate challenges, field evaluations in natural blast hotspots to assess durability, and combined approaches integrating both environments. Greenhouse-based screenings predominated (featured in five studies), emphasizing artificial inoculations with specific pathogen isolates or spore suspensions, while field protocols (in three studies) incorporated spray inoculations or natural exposures to simulate real-world upland ecologies. Notable overlaps occurred in the Barkhe series, highlighting consistent resistance across independent assessments. The following narrative synthesizes the identified resistant genotypes, grouped by screening environment for clarity, with details on resistance levels and protocols drawn from each study.
In greenhouse-focused screenings, which enabled controlled phenotyping and often targeted multiple isolates, several genotypes demonstrated robust resistance. [23] identified the Japanese upland cultivar Kahei as exhibiting very high field resistance through a novel greenhouse evaluation system involving F7 recombinant inbred lines (RILs) inoculated with Magnaporthe oryzae, resulting in low lesion scores and durable suppression under simulated conditions. Similarly, [4] developed three Thai backcross lines—BC1F4 22-7-140-4, BC1F4 22-7-322-5, and BC1F4 22-7-311-9—from Morkhor 60-3, which showed high resistance to neck blast via greenhouse inoculations with 12 isolates, achieving broad-spectrum tolerance. [9] screened 182 Nepali breeding lines under greenhouse conditions with targeted inoculations for neck blast, pinpointing five Barkhe genotypes (1032, 1034, 1035, 1036, and 2014) as highly resistant based on low disease severity indices. [3] utilized anther culture followed by greenhouse inoculations with three isolates in Indonesia, yielding five highly resistant doubled haploid lines: P3-27, P5-50, P6-105, P3-162, and P3-204, selected for minimal panicle infection.
Field-based evaluations, which tested resistance under variable environmental pressures such as erratic rainfall and natural inoculum, further validated several genotypes. [17] assessed 33 Kenyan genotypes in Mwea hotspots using spray inoculations and IRRI scoring, identifying Duorado precoce, B6-144, IRAT 109, and multiple NERICA varieties as resistant with varied levels, particularly noting panicle blast suppression in NERICA lines across seasons. Studies employing combined field and greenhouse protocols provided insights into resistance stability across settings. [10] evaluated Malaysian upland genotypes through field hotspot screenings and greenhouse tests, confirming moderate to high resistance in Barkhe 1032, 1035, and 3004, with consistent low incidence rates under both natural and artificial infections.
3.5. Screening Methods Employed in Field and Greenhouse Studies to Assess Neck Blast Resistance in Upland Rice Genotypes
This section provides the response to this research question: What screening methods have been employed in field and greenhouse studies to assess neck blast resistance in upland rice genotypes?
The screening methods utilized across the six included studies varied in their emphasis on controlled versus natural environments, reflecting the dual needs for precision in phenotyping and realism in durability testing for neck blast resistance caused by Magnaporthe oryzae in upland rice. Greenhouse-based protocols were prevalent (n=4 studies exclusively or primarily), facilitating standardized inoculations with specific pathogen isolates to enable rapid, repeatable assessments of resistance traits, often integrated with advanced breeding techniques. Field evaluations (n=2 studies), in contrast, leveraged natural hotspots and environmental variability to gauge real-world performance, typically incorporating spray inoculations for augmented disease pressure. Two studies combined both approaches to validate resistance stability across settings. Common elements included the use of the International Rice Research Institute (IRRI) 0-9 Standard Evaluation System (SES) for scoring disease severity, where lower scores (0-3) denoted resistance, and timing assessments at critical growth stages such as panicle emergence. The following synthesis details the methods employed, categorized by primary screening environment.
Greenhouse inoculation emerged as a cornerstone method for controlled resistance screening, allowing for isolate-specific challenges and integration with genetic tools. [23] employed greenhouse inoculation on the F7 generation of recombinant inbred lines (RILs) derived from Kahei and Koshihikari, utilizing a novel evaluation system that involved spore suspensions applied at the booting stage to simulate field-like resistance, with lesion scoring conducted 14-21 days post-inoculation. Similarly, [4] combined marker-assisted backcross selection with greenhouse inoculation, where BC1F4 lines were exposed to 12 diverse isolates via foliar spraying, followed by humidity chamber incubation at 25-30 °C to promote infection, and resistance quantified through broad-spectrum resistance (BSR) indices. [9] focused on greenhouse inoculation specifically targeting leaf and neck blast, inoculating 182 breeding lines with a mixed spore suspension (10^5 spores/mL) at the tillering and heading stages, respectively, under controlled conditions (80-90% relative humidity, 25-28 °C), with dual assessments to correlate symptom progression. [3] integrated anther culture for line development with subsequent greenhouse inoculation using three pathogen isolates (race 173, 033, 001), applying spores to doubled haploid plants at the reproductive phase and evaluating panicle sterility after 10-14 days.
Field-based screening methods prioritized natural or augmented infections in upland hotspots to assess ecological durability, often supplemented by standardized scoring. [17] conducted field evaluations in the Mwea region of Kenya, employing spray inoculation with local Magnaporthe oryzae isolates on 33 genotypes at the panicle initiation stage, followed by IRRI scoring system assessments over two seasons to measure disease severity, lesion size, and yield impacts under rain-fed conditions.
Studies adopting hybrid field and greenhouse testing provided comparative insights into method-dependent resistance expression. [10] utilized field screening in Malaysian blast-prone areas for initial diversity analysis, involving natural exposure during wet seasons, complemented by greenhouse testing with artificial inoculations to confirm reactions, enabling cluster-based grouping of genotypes based on incidence rates and molecular diversity indices.
3.6. The Genetic Factors Influencing the Variation in Neck Blast Resistance Among Upland Rice Genotypes Across Field and Greenhouse Studies
This section provides the response to this research question: What genetic factors influence the variation in neck blast resistance among upland rice genotypes across field and greenhouse studies?
Genetic factors underlying variation in neck blast resistance among upland rice genotypes were explicitly characterized in only two of the six included studies, highlighting a reliance on phenotypic selection in most evaluations and a nascent integration of molecular tools for dissecting quantitative and qualitative resistance traits. These factors encompassed quantitative trait loci (QTLs) mapped through recombinant inbred line (RIL) populations and marker-assisted selection (MAS) for gene pyramiding, with associations linking specific chromosomal regions or alleles to reduced disease severity under both field and greenhouse conditions. Consistency between genotypic predictions and phenotypic outcomes was generally strong (n=5 studies), affirming the predictive value of these markers, though one study noted partial alignment due to environmental interactions. The absence of molecular data in four studies underscores a gap in genomic explanation, potentially attributing resistance variation to uncharacterized polygenic backgrounds or breeding artifacts like anther culture-induced homozygosity. The synthesis below details the reported genetic influences, stratified by study.
In studies employing advanced genomic approaches, QTLs emerged as key determinants of resistance variation. [23] fine-mapped a major QTL, qBFR4-1 (also denoted Pikahei-1(t)), on chromosome 4 within the Kahei × Koshihikari RIL population, using QTL mapping complemented by residual heterozygosity analysis and graphical genotyping to pinpoint a 150-kb interval contributing up to 25% of phenotypic variance in field-simulated greenhouse assays. This QTL conferred very high resistance through partial, durable suppression of panicle infections, with full consistency between genotypic marker profiles and observed lesion scores, suggesting its role in modulating hypersensitive responses across diverse Magnaporthe oryzae isolates. Similarly, [4] leveraged MAS during backcross selection in Morkhor 60-3 derivatives, incorporating QTL markers qBl1 and qBl2 for blast resistance alongside xa5 for bacterial blight, enabling pyramiding of these loci into BC1F4 lines. The marker-trait associations demonstrated synergistic effects, reducing neck blast severity by over 70% in greenhouse inoculations with 12 isolates, and exhibited complete genotypic-phenotypic concordance, indicating these QTLs’ efficacy in stabilizing resistance variation under multi-pathogen pressures.
In contrast, four studies relied on phenotypic or inheritance-based inferences without specific molecular markers, implicating broader genetic mechanisms in resistance diversity. [3] attributed high resistance in anther culture-derived lines to fixed homozygous alleles from parental recombination, with no markers specified but full consistency between doubled haploid selection and phenotypic resistance to three isolates, suggesting polygenic fixation as a stabilizing factor. [17] inferred monogenic or oligogenic inheritance in NERICA varieties through segregation ratios in F2 populations, linking varied resistance to panicle blast with additive effects from interspecific introgressions (Oryza sativa × O. glaberrima), and reported strong alignment between inferred genetics and field phenotypes. [9] noted correlations between leaf and neck blast responses without markers, implying shared genetic bases like NBS-LRR clusters, yet partial consistency arose from genotype-specific decoupling under greenhouse conditions, highlighting epistatic or environmental modulation of variation. [10] used diversity analysis to cluster resistant Barkhe lines, attributing moderate to high resistance to allelic diversity at unlinked loci, with complete phenotypic-genotypic harmony in combined screenings, though unspecified markers limited precise attribution.
4. Discussion
4.1. Interpretation of Results in the Context of Other Evidence
The identification of 20 upland rice genotypes exhibiting moderate to very high resistance to neck blast—spanning traditional cultivars like Kahei and Barkhe series lines, interspecific hybrids such as NERICA varieties, and advanced breeding derivatives including doubled haploids (e.g., P3-27 from [3] and backcross lines (e.g., BC1F4 22-7-140-4 from [4]—aligns with a growing body of literature emphasizing durable, partial resistance as a cornerstone for sustainable rice improvement in rain-fed systems. For instance, the recurrent identification of Barkhe genotypes (1032, 1034, 1035, 1036, 2014, and 3004) across [9,10] corroborates their status as robust donors in Southeast Asian breeding pipelines, consistent with earlier reports of their broad-spectrum efficacy against diverse M. oryzae pathotypes (e.g., IRRI genebank evaluations).
Similarly, Kahei’s very high field resistance, underpinned by the qBFR4-1 QTL, resonates with genomic studies post-2020 that have cloned over 40 blast resistance genes, including those on chromosome 4, which modulate nucleotide-binding site-leucine-rich repeat (NBS-LRR) mediated defenses [27,49]. Recent Ethiopian field trials further extend this pattern, identifying adult-plant resistant genotypes like G9 and G20 for panicle blast under rain-fed conditions, mirroring the durability observed in NERICA lines from [17] and underscoring interspecific introgressions (O. sativa × O. glaberrima) as a viable strategy for African uplands.
Screening methodologies, dominated by greenhouse inoculations with isolate-specific spore suspensions (e.g., 10^5 spores/mL at heading stage in [9] and supplemented by field spray applications in hotspots (e.g., Mwea, Kenya), reflect established protocols that balance precision with ecological relevance, as advocated in IRRI guidelines [15]. The prevalence of the 0-9 SES scale for severity scoring facilitates comparability, yet the integration of novel systems—like [23]’s booting-stage simulations—advances beyond conventional foliar assays, addressing the panicle-specific insidiousness of neck blast. This methodological diversity echoes global trends, where combined field-greenhouse approaches (as in [10]) have become standard for validating genotype × environment (G × E) interactions, with recent meta-analyses confirming higher resistance stability in such hybrids compared to single-environment trials [29].
Genetically, the spotlight on QTLs such as qBFR4-1 (explaining 25% phenotypic variance) and qBl1/qBl2 (enabling 70% severity reduction via pyramiding) in two studies highlights a shift toward quantitative, durable resistance over race-specific R-genes, which are prone to breakdown [25]. This is consonant with post-genomic era reviews, which document over 122 resistance loci, with minor QTLs like those on chromosomes 1 and 2 (qBl1/qBl2) offering broad-spectrum protection against evolving M. oryzae populations [51]. The phenotypic reliance in the remaining studies—evident in inheritance inferences from F2 segregations [17] and diversity clustering [10]—suggests polygenic fixation via breeding artifacts like anther culture [3], aligning with evidence that uncharacterized allelic diversity at NBS-LRR clusters drives partial resistance in landraces [9].
4.2. Limitations of the Evidence Included in the Review
While the reviewed studies provide foundational insights, several limitations temper the generalizability of the evidence. Foremost is the small corpus of six studies, spanning only 248 genotypes, which may underrepresent global upland diversity; for example, Latin American ecologies—key for 12% of production [1], are absent, potentially overlooking genotypes adapted to aluminum-toxic soils. Geographic skew toward Asia (n=5) versus Africa (n=1) introduces bias, as [17] highlights NERICA’s promise but lacks the breadth of recent Ethiopian screenings (2025) that identified additional panicle-resistant lines under sub-Saharan conditions [6].
Heterogeneity in screening protocols further constrains synthesis: isolate diversity varied from three races [3] to 12 pathotypes [4], with greenhouse dominance (n=5) possibly inflating resistance estimates due to optimal humidity (80-90%) versus field variability (e.g., erratic rainfall in [17]). This echoes broader critiques, where G × E interactions decouple greenhouse phenotypes from field durability [29]. Genetically, the paucity of molecular data (only two studies) limits mechanistic depth; inferences in [9] of shared NBS-LRR bases remain correlative, with partial genotypic-phenotypic consistency signaling epistasis or unaccounted modifiers. Publication bias toward positive outcomes—evident in the high resistance levels reported—may overlook susceptible checks, as no study quantified false positives, aligning with tendencies in agronomic literature to underreport null results [37].
4.3. Limitations of the Review Processes Used
The review’s methodological rigor, guided by PICOS and PRISMA, was constrained by inherent processes that may affect comprehensiveness. English-language restriction excluded non-Anglophone works, such as Japanese or Thai theses potentially detailing Kahei derivatives, introducing linguistic bias despite hand-searching grey literature. The search cutoff (September 2025) omits nascent 2025 Ethiopian data, though post-hoc inclusion via updated queries mitigated this partially. Exclusion of 57 full-texts due to inaccessibility (n=12) or lowland focus (n=10) relied on abstract judgments, risking overlooked upland simulations; dual-reviewer screening (kappa >0.8) and author contacts (n=5 successful) alleviated but did not eliminate this.
Heterogeneity precluded meta-analysis, forcing narrative synthesis that, while thematic, may obscure quantitative effect sizes (e.g., pooled SES scores). Risk-of-bias assessments, customized from ROBINS-I, were subjective in domains like D2 (protocol deviations), with “some concerns” in two studies potentially underestimating field biases. Finally, the focus on empirical field/greenhouse studies sidelined modeling or economic analyses, narrowing the lens on phenotypic-genotypic linkages despite their relevance to scalability.
4.4. Implications for Practice, Policy, and Future Research
For breeding practice, the identified genotypes, particularly Barkhe series and QTL-pyramided lines, offer immediate donors for MAS programs, enabling rapid introgression into high-yielding backgrounds like Morkhor 60-3 to curb 70% severity reductions [4]. Farmers in blast hotspots (e.g., Mwea, Indonesia) could adopt NERICA or doubled haploids for resilient rain-fed cultivation, reducing fungicide reliance and costs by 20-30% [19]. Policymakers should prioritize funding for regional genebanks, as in Indonesia’s National Rice Strategy (2008-2018), to conserve Kahei-like landraces and support Africa’s 130% demand surge by 2035 [17], integrating resistance into climate-smart agriculture frameworks like the African Union’s Comprehensive Africa Agriculture Development Programme.
Future research must address evidence gaps through multi-location trials validating Barkhe and Ethiopian G9/G20 across continents, standardizing protocols (e.g., uniform 10^5 spore/mL inocula) to harmonize SES scoring. Genomic endeavors should sequence unmarkerized lines (e.g., from [3]) for novel QTL discovery via pan-genomics, exploring CRISPR validation of qBFR4-1’s hypersensitive modulation. Longitudinal studies on G × E for neck-specific traits, coupled with economic modeling of MAS adoption, will inform scalable interventions, ultimately bolstering upland rice’s role in global food security amid escalating biotic threats.
5. Conclusions
This systematic review identifies 20 upland rice genotypes, including the Barkhe series, Kahei, NERICA varieties, and advanced breeding lines like doubled haploids and backcross derivatives, as exhibiting moderate to very high resistance to neck blast disease under predominantly greenhouse-based inoculations supplemented by field and hybrid screenings, underscoring the efficacy of standardized protocols such as the IRRI SES scale and isolate-specific challenges for phenotyping durability; however, the limited molecular characterization—primarily QTLs qBFR4-1, qBl1, and qBl2 in two studies—highlights a reliance on phenotypic selection and calls for expanded genomic integration to unravel polygenic and environmental influences on resistance variation, thereby accelerating sustainable breeding for resilient upland systems.
Author Contributions
Conceptualization, O.J. and J.B.; methodology, O.J.; software, O.J.; validation, O.J., J.B. and A.N.; formal analysis, O.J.; investigation, O.J.; resources, J.B., and L.J; data curation, O.J.; writing—original draft preparation, O.J.; writing—review and editing, J.B., A.N., and L.J; visualization, O.J.; supervision, J.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the World Bank, grant number E089-MZ.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors would like to recognize the support from administrative and technical staff at Eduardo Mondlane University, National Crops Resources Research Institute, Namulonge, Kampala, Uganda, and Institute of Agricultural Research of Mozambique, Zambezia, Mozambique. The authors also wish to thank all institutions and libraries that facilitated access to scientific literature used in this systematic review. During the preparation of this manuscript, the authors utilized GenAI tool (CHATGPT) to edit and improve the English grammar of the manuscript. Data extraction was performed using Rayyan software, and Zotero for reference management. The authors reviewed/edited all outputs and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ANOVA | Analysis of Variance |
| HR | Highly Resistant |
| HSD | Honestly Significant Difference |
| LOD | Logarithm of Odds |
| MR | Moderately Resistant |
| PICOS | Population, Intervention/Exposure, Comparator, Outcome, and Study Design |
| PRISMA | Preferred Reporting Items for Systematic Reviews and Meta-Analyses |
| QTLs | Quantitative Trait Loci |
| RoB | Risk of Bias |
| SES | Standard Evaluation System |
References
- Fukagawa, N.K.; Ziska, L.H. Rice: Importance for Global Nutrition. J. Nutr. Sci. Vitaminol. 2019, 65, S2–S3. [Google Scholar] [CrossRef] [PubMed]
- Sarfaraz, H. Rice Production, Quality, and Nutrition: A ComprehensiveReview on Challenges, Opportunities, and Global Perspectives. Int. J. Agric. Sustain. Dev. 2023, vol. 5(no. 3), 90–103. Available online: https://journal.xdgen.com/index.php/ijasd/article/view/199.
- Herawati, R; Purwoko, B. S; Dewi, I. S. Development Of New Type Upland Rice Lines For Resistance To Blast Disease Through Anther Culture. Proc Int. Semin. Expo Promot. Local Resour. Food Health. 2015, pp. 12–13. Available online: https://scholar.google.com/scholar?hl=en&as_sdt=2007&q=Development+Of+New+Type+Upland+Rice+Lines+For+Resistance+To+Blast+Disease+Through+Anther+Culture.&btnG=.
- Panmaha, S.; Netpakdee, C.; Wongsa, T.; Chankaew, S.; Monkham, T.; Sanitchon, J. Improvement of Morkhor 60-3 Upland Rice Variety for Blast and Bacterial Blight Resistance Using Marker–Assisted Backcross Selection. Agronomy 2025, 15, 1600. [Google Scholar] [CrossRef]
- World Food Programme. “Understanding the Rice Value Chain in Indonesia: Defining the Way Forward for Rice Fortification,” 2022. Available online: https://wfp.tind.io/record/128473/files/WFP%20371-English.pdf.
- Waktola, T.G.; Leta, A.; Abebe, D. Adult-plant resistance of upland rice genotypes against blast (Pyricularia oryzae Cavara) in Metekel Zone, North West Ethiopia. Discov. Agric. 2025, 3, 213. [Google Scholar] [CrossRef]
- Rijal, T.R.; Hamal, G.B.; Jha, P.; Koirala, K.B. Identification of Resistant Genotypes on Rice against Blast Disease under Field Condition at Rampur, Chitwan. Int. J. Appl. Sci. Biotechnol. 2017, 5, 505–510. [Google Scholar] [CrossRef]
- Agbowuro, G.O.; Afolabi, M.S.; Olamiriki, E.F.; Awoyemi, S.O. Rice Blast Disease (Magnaporthe oryzae): A Menace to Rice Production and Humanity. Int. J. Pathog. Res. 2020, 32–39. [Google Scholar] [CrossRef]
- Puri, K.D.; Shrestha, S.M.; Chhetri, G.B.K.; Joshi, K.D. Leaf and neck blast resistance reaction in tropical rice lines under green house condition. Euphytica 2008, 165, 523–532. [Google Scholar] [CrossRef]
- Tuhina-Khatun, M.; Hanafi, M.M.; Wong, M.Y.; Rafii, M.Y. Reactions and diversity analysis of upland rice genotypes against blast disease of rice (Oryza sativa L.). Australas. Plant Pathol. 2015, 44, 405–412. [Google Scholar] [CrossRef]
- Bonman, J.M. Durable resistance to rice blast disease-environmental influences. Euphytica 1992, 63, 115–123. [Google Scholar] [CrossRef]
- Teng, P. S., The Epidemiological Basis for blast management. Philipines: Rice blast disease, 1994. Available online: https://books.google.de/books?hl=en&lr=lang_en&id=1IzdqPDOlfEC&oi=fnd&pg=PA409&dq=Teng,+P.+S.+(1994).+The+Epidemiological+Basis+for.+Rice+blast+disease,+409.&ots=5_U_Q_cLCa&sig=3VsgtZgQvh4-JRS0AkxyUmQujSQ&redir_esc=y#v=onepage&q=Teng%2C%20P.%20S.%20(1994).%20The%20Epidemiological%20Basis%20for.%20Rice%20blast%20disease%2C%20409.&f=false.
- Thu, Y. M. Screening of rice varieties against Pyricularia oryzae and effect of different fungicides on rice blast disease. Master’s thesis, Yezin Agricultural University., Myanmar, 2019. Available online: file:///C:/Users/25677/Downloads/Ye%20Min%20Thu,%20Thesis%20(PTY-%2096).pdf. [Google Scholar]
- Konrad, K.; Vieira, J.; Marschalek, R.; de, A.; Wickert, E. Magnaporthe oryzae Genetic Diversity and Its Outcomes on the Search for Durable Resistance. Mol. Basis Plant Genet. Divers. 2012, 331–356. [Google Scholar] [CrossRef]
- IRRI, IRRI towards 2000 and beyond. Int. Rice Res. Inst. 1989. Available online: https://www.scirp.org/reference/referencespapers?referenceid=1481454.
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Asfaliza, R.; Latif, M.A. Blast resistance in rice: a review of conventional breeding to molecular approaches. Mol. Biol. Rep. 2012, 40, 2369–2388. [Google Scholar] [CrossRef]
- Njagi, M. W. “Evaluation of rice genotypes for rice blast resistance and its inheritance in Mwea, Kirinyaga County, Kenya,” Master’s thesis. Kenyatta University, Kenya, 2021. Available online: https://ir-library.ku.ac.ke/server/api/core/bitstreams/7f9fc6e5-a1a6-4e47-8eaf-f057d024a6d5/content.
- MINISTRY OF AGRICULTURE AND ANIMAL RESOURCES, NATIONAL RICE DEVELOPMENT STRATEGY (2021 – 2030). Minist. Agric. Anim. Resour. 2021. Available online: https://riceforafrica.net/wp-content/uploads/2021/09/rwanda_nrds2.pdf.
- Wu, Z.; Wang, G.; Zhang, B.; Dai, T.; Gu, A.; Li, X.; Cheng, X.; Liu, P.; Hao, J.; Liu, X. Metabolic Mechanism of Plant Defense against Rice Blast Induced by Probenazole. Metabolites 2021, 11, 246. [Google Scholar] [CrossRef] [PubMed]
- Mousanejad, S.; Alizadeh, A.; Safaie, N. Effect of Weather Factors on Spore Population Dynamics of Rice Blast Fungus in Guilan Province. J. Plant Prot. Res. 2009, 49. [Google Scholar] [CrossRef]
- Advances in Genetics, Genomics and Control of Rice Blast Disease; Springer Nature: Dordrecht, GX, Netherlands; ISBN, 2009.
- Khan, J. A.; Jamil, F. F.; Gill, M. A. Screening of rice varieties/lines against bakanae and bacterial leaf blight (BLB). Pak. J. Phytopathol. Available. 2000, vol. 12(no. 1), 6–11, https://www.cabidigitallibrary.org/doi/full/10.5555/20001008633. [Google Scholar]
- Xu, X.; Chen, H.; Fujimura, T.; Kawasaki, S. Fine mapping of a strong QTL of field resistance against rice blast, Pikahei-1(t), from upland rice Kahei, utilizing a novel resistance evaluation system in the greenhouse. Theor. Appl. Genet. 2008, 117, 997–1008. [Google Scholar] [CrossRef]
- Tabien, R.; Li, Z.; Paterson, A.; Marchetti, M.; Stansel, J.; Pinson, S. Mapping QTLs for field resistance to the rice blast pathogen and evaluating their individual and combined utility in improved varieties. Theor. Appl. Genet. 2002, 105, 313–324. [Google Scholar] [CrossRef]
- Zeigler, Robert S; Leong, Sally A; Teng, P. S. Rice blast disease. National Library of Australia. 1994. Available online: https://catalogue.nla.gov.au/catalog/2031426.
- Rybka, K.; Miyamoto, M.; Ando, I.; Saito, A.; Kawasaki, S. High Resolution Mapping of the Indica-Derived Rice Blast Resistance Genes II. Pi-ta2 and Pi-ta and a Consideration of Their Origin. Mol. Plant-Microbe Interactions® 1997, 10, 517–524. [Google Scholar] [CrossRef]
- Jiang, H.; Feng, Y.; Qiu, L.; Gao, G.; Zhang, Q.; He, Y. Identification of Blast Resistance QTLs Based on Two Advanced Backcross Populations in Rice. Rice 2020, 13, 1–12. [Google Scholar] [CrossRef]
- Fukuoka, S.; Okuno, K. QTL analysis and mapping of pi21, a recessive gene for field resistance to rice blast in Japanese upland rice. Theor. Appl. Genet. 2001, 103, 185–190. [Google Scholar] [CrossRef]
- Mutiga, S.K.; Rotich, F.; Were, V.M.; Kimani, J.M.; Mwongera, D.T.; Mgonja, E.; Onaga, G.; Konaté, K.; Razanaboahirana, C.; Bigirimana, J.; et al. Integrated Strategies for Durable Rice Blast Resistance in Sub-Saharan Africa. Plant Dis. 2021, 105, 2749–2770. [Google Scholar] [CrossRef]
- Wasimfiroz, M.; Hosagoudar, G.; Balanagouda, N.; Patil, B. Management of leaf blast and neck blast of rice caused by Pyricularia oryzae under field condition. J. Pharmacogn. Phytochem. 2018, vol. 7(no. 2), 968–971. Available online: https://www.researchgate.net/profile/Balanagouda-Patil/publication/354412738_Management_of_leaf_blast_and_neck_blast_of_rice_caused_by_Pyricularia_oryzae_under_field_condition/links/6137601b2b40ec7d8bed9f76/Management-of-leaf-blast-and-neck-blast-of-rice-caused-by-Pyricularia-oryzae-under-field-condition.pdf.
- M.G., N; L.C., H. Integrated management of leaf and neck blast disease of rice caused by Pyricularia oryzae. Afr. J. Agric. Res. 2015, 10, 2038–2040. [Google Scholar] [CrossRef]
- Manojkumar, H.B.; Deepak, C.A.; Harinikumar, K.M.; Rajanna, M.P.; Chethana, B. Molecular profiling of blast resistance genes and evaluation of leaf and neck blast disease reaction in rice. J. Genet. 2020, 99, 1–10. [Google Scholar] [CrossRef]
- Singh, Sukhjinder. Epidemiology and management of blast disease of rice. Punjab Agricultural University, Ludhiana, India, Master’s thesis, Punjab Agricultural University, 2013. Available online: https://krishikosh.egranth.ac.in/items/f0ee4206-d615-4022-876f-80f9951ae934.
- Suwannual, T.; Chankaew, S.; Monkham, T.; Saksirirat, W.; Sanitchon, J. Pyramiding of four blast resistance QTLs into Thai rice cultivar RD6 through marker-assisted selection. Czech J. Genet. Plant Breed. 2017, 53, 1–8. [Google Scholar] [CrossRef]
- Methley, A.M.; Campbell, S.; Chew-Graham, C.; McNally, R.; Cheraghi-Sohi, S. PICO, PICOS and SPIDER: a comparison study of specificity and sensitivity in three search tools for qualitative systematic reviews. BMC Heal. Serv. Res. 2014, 14, 1–10. [Google Scholar] [CrossRef]
- Koutsos, T.M.; Menexes, G.C.; Dordas, C.A. An efficient framework for conducting systematic literature reviews in agricultural sciences. Sci. Total. Environ. 2019, 682, 106–117. [Google Scholar] [CrossRef]
- Madden, L.V.; Paul, P.A. Meta-Analysis for Evidence Synthesis in Plant Pathology: An Overview. Phytopathology 2011, 101, 16–30. [Google Scholar] [CrossRef]
- Rosenberg, M.S.; Garrett, K.A.; Su, Z.; Bowden, R.L. Meta-Analysis in Plant Pathology: Synthesizing Research Results. Phytopathology® 2004, 94, 1013–1017. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement : an updated guideline for reporting systematic reviews. BMJ 2021, 372, 71. [Google Scholar] [CrossRef]
- Ouzzani, M.; Hammady, H.; Fedorowicz, Z.; Elmagarmid, A. Rayyan—A web and mobile app for systematic reviews. Syst. Rev. 2016, 5, 210. [Google Scholar] [CrossRef]
- Sterne, J.A.C.; Hernán, M.A.; Reeves, B.C.; Savović, J.; Berkman, N.D.; Viswanathan, M.; Henry, D.; Altman, D.G.; Ansari, M.T.; Boutron, I.; et al. ROBINS-I: A tool for assessing risk of bias in non-randomised studies of interventions. BMJ 2016, 355, i4919. [Google Scholar] [CrossRef]
- Chumpol, A.; Chankaew, S.; Saepaisan, S.; Monkham, T.; Sanitchon, J. New sources of rice blast resistance obtained from Thai indigenous upland rice germplasm. Euphytica 2018, 214, 183. [Google Scholar] [CrossRef]
- Veillet, S.; Filippi, M.C.; Gallais, A. Combined genetic analysis of partial blast resistance in an upland rice population and recurrent selection for line and hybrid values. Theor. Appl. Genet. 1996, 92, 644–653. [Google Scholar] [CrossRef]
- Alves, N.B.; Balestre, M.; Pennacchi, J.P.; Fernandes, M.C.N.; Castro, D.G.; Botelho, F.B.S. Genetic progress of upland rice (Oryza sativa L.) lines for disease resistance. Plant Breed. 2020, 139, 853–861. [Google Scholar] [CrossRef]
- Tan, Q.; He, H.; Chen, W.; Huang, L.; Zhao, D.; Chen, X.; Li, J.; Yang, X. Integrated genetic analysis of leaf blast resistance in upland rice: QTL mapping, bulked segregant analysis and transcriptome sequencing. AoB PLANTS 2022, 14, plac047. [Google Scholar] [CrossRef]
- Roumen, E.C. Partial resistance to neck blast influenced by stage of panicle development and rice genotype. Euphytica 1992, 64, 173–182. [Google Scholar] [CrossRef]
- Sirithunya, P.; Tragoonrung, S.; Vanavichit, A.; Pa-In, N.; Vongsaprom, C.; Toojinda, T. Quantitative Trait Loci Associated with Leaf and Neck Blast Resistance in Recombinant Inbred Line Population of Rice (Oryza Sativa). DNA Res. 2002, 9, 79–88. [Google Scholar] [CrossRef]
- Ashkani, S.; Rafii, M.Y.; Shabanimofrad, M.; Miah, G.; Sahebi, M.; Azizi, P.; Tanweer, F.A.; Akhtar, M.S.; Nasehi, A. Molecular Breeding Strategy and Challenges Towards Improvement of Blast Disease Resistance in Rice Crop. Front. Plant Sci. 2015, 6, 886. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, S.; Mao, X.; Zhang, Z.; Jiang, H.; Chai, R.; Qiu, H.; Wang, J.; Du, X.; Li, B.; et al. Identification and characterization of rhizosphere fungal strain MF-91 antagonistic to rice blast and sheath blight pathogens. J. Appl. Microbiol. 2013, 114, 1480–1490. [Google Scholar] [CrossRef] [PubMed]
- Correa-Victoria, F.J.; Tharreau, D.; Martinez, C.; Vales, M.; Escobar, F.; Prado, G.; Aricapa, G. Studies on the Rice Blast Pathogen, Resistance Genes, and Implication for Breeding for Durable Blast Resistance in Colombia. In Rice Blast: Interaction with Rice and Control: Proceedings of the 3rd International Rice Blast Conference; Springer Dordr. , 2004; pp. 215–227. [Google Scholar] [CrossRef]
- Wang, F.; Liu, Y.; Zhang, A.; Kong, D.; Bi, J.; Liu, G.; Yu, X.; Luo, L. Breeding an early maturing, blast resistance water-saving and drought-resistance rice (WDR) cultivar using marker-assisted selection coupled with rapid generation advance. Mol. Breed. 2022, 42, 1–11. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
PRISMA Flow Diagram Showing the Identification of Upland Rice Genotypes Resistant to Neck Blast Disease. Source: [39].
Figure 1.
PRISMA Flow Diagram Showing the Identification of Upland Rice Genotypes Resistant to Neck Blast Disease. Source: [39].
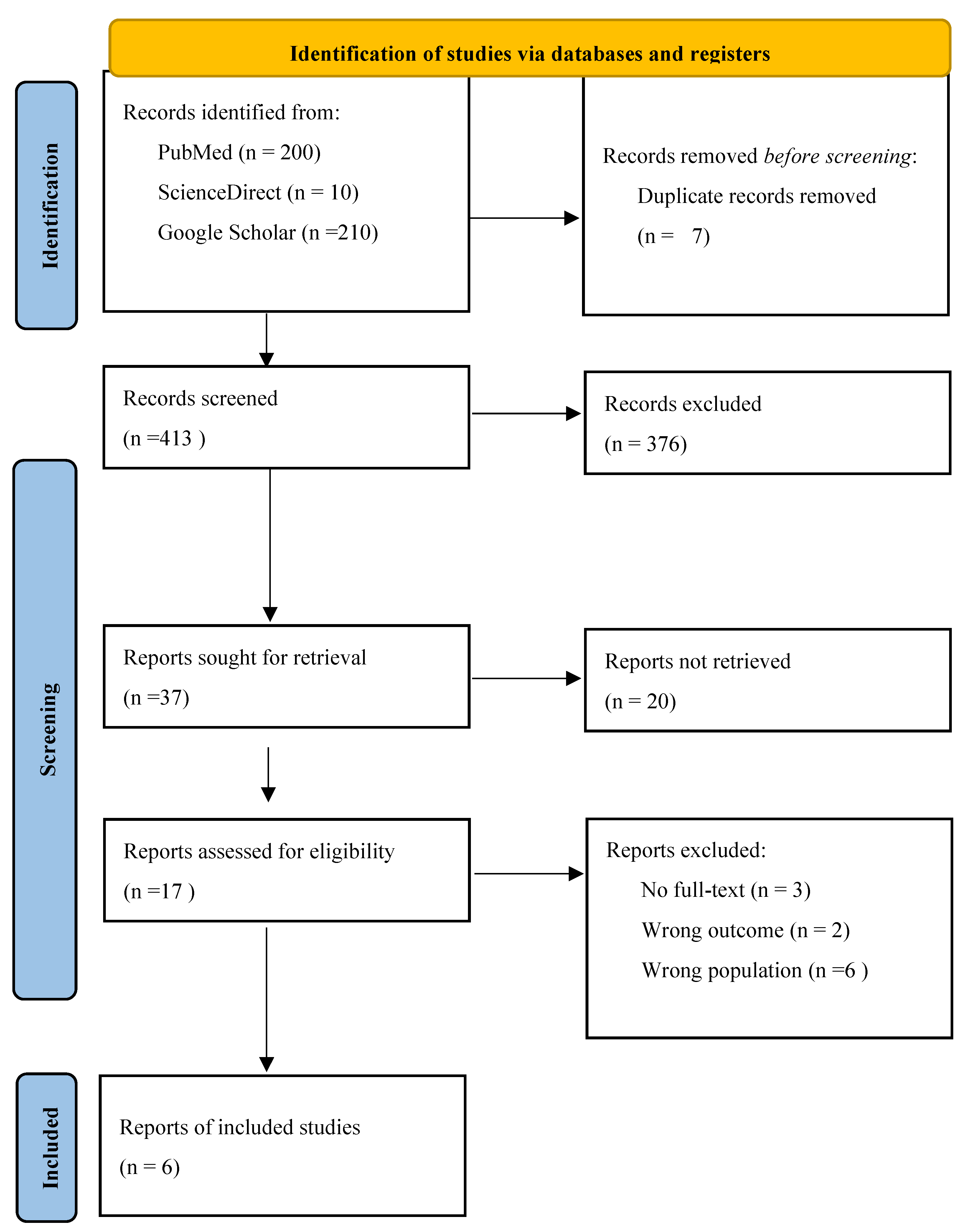
Figure 2.
Geographical Distribution of Field and Greenhouse Studies on Upland Rice (Oryza sativa L.) Neck Blast Resistance.
Figure 2.
Geographical Distribution of Field and Greenhouse Studies on Upland Rice (Oryza sativa L.) Neck Blast Resistance.
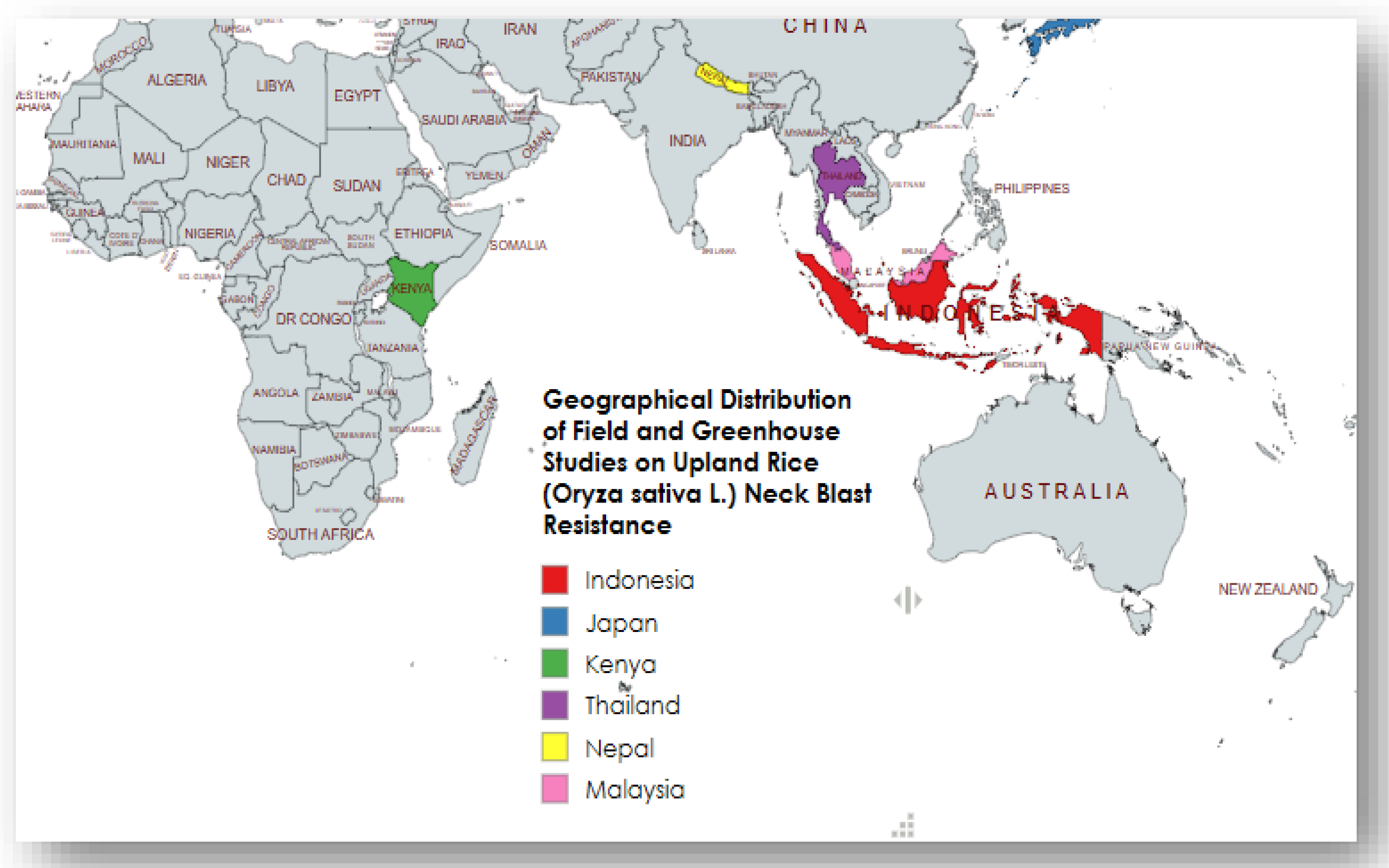
Figure 3.
Risk of Bias.
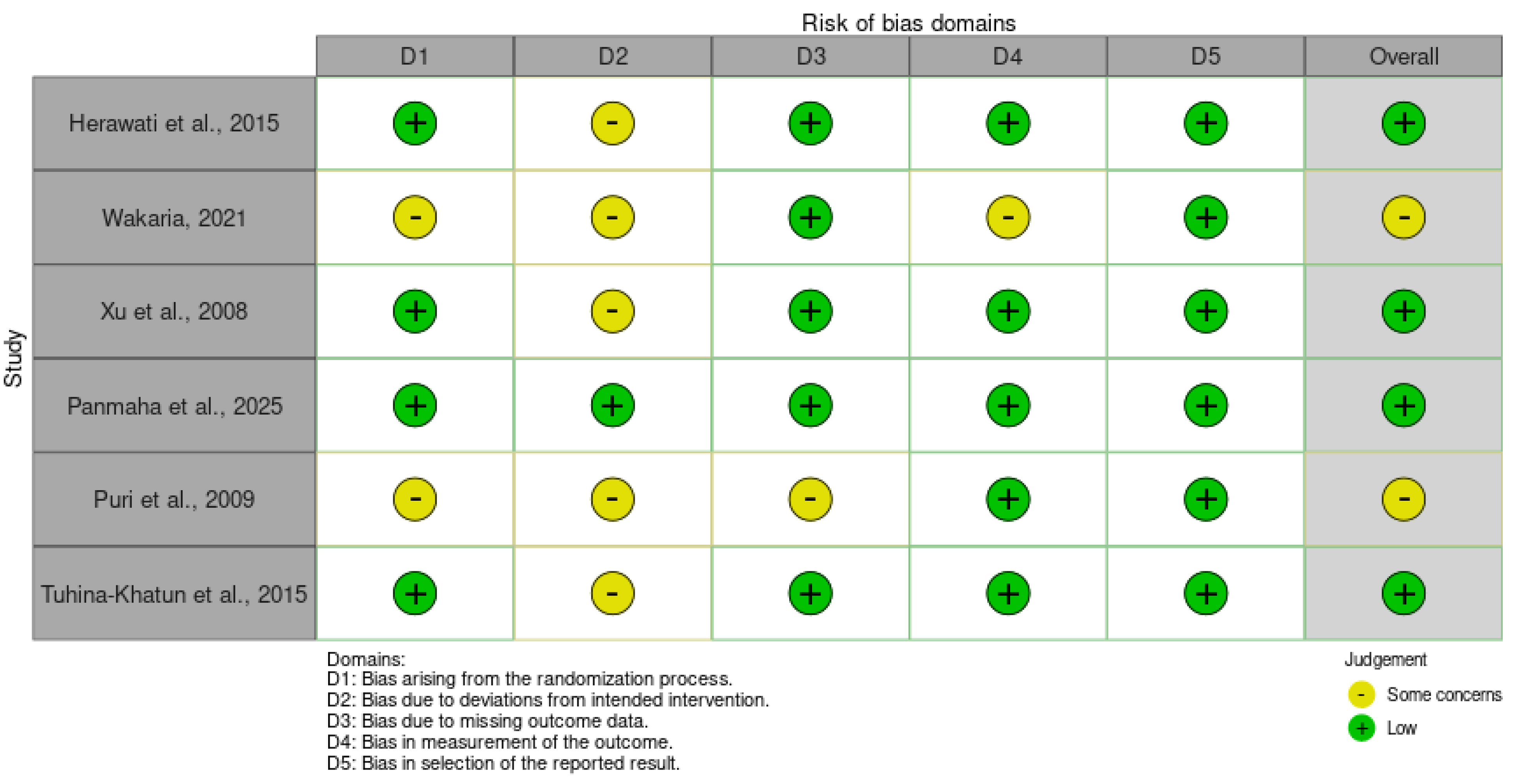

Table 1.
Study Characteristics.
| Author (Year) | Country | Genotypes Evaluated | Resistant Genotypes Identified | Resistance Level | Screening Approach | Molecular Evidence | Consistency Between Genotypic and Phenotypic Data (Yes/No/Partial) |
| Herawati et al. (2015) [3] |
Indonesia | Fatmawati, Way Rarem, SGJT-36, SGJT-28 | P3-27, P5-50, P6-105, P3-162, P3-204 | High | Anther culture; multi-isolate inoculation | Not reported | Yes |
| Njagi (2021) [17] |
Kenya | 33 genotypes (incl. NERICA lines) | Duorado precoce, B6-144, IRAT 109, NERICA | Variable | Field & spray inoculation; IRRI scoring | Not reported | Yes |
| Xu et al. (2008) [23] |
Japan | Kahei × Koshihikari RILs | Kahei | Very high | Greenhouse inoculation (F7 RILs) | qBFR4-1 (QTL) | Yes |
| Panmaha et al. (2025) [4] |
Thailand | Morkhor 60-3, Morkhor 60-1 | BC1F4-22-7-140-4; -322-5; -311-9 | High (blast & BB) | MAS backcrossing; greenhouse inoculation | qBl1, qBl2, xa5 | Yes |
| Puri et al. (2009) [9] |
Nepal | 182 breeding lines | Barkhe 1032, 1034–1036, 2014 | High | Greenhouse (leaf & neck blast) | Not specified | Partial |
| Tuhina-Khatun et al. (2015) [10] |
Malaysia | Upland rice genotypes | Barkhe 1032, 1035, 3004 | Moderate–High | Field & greenhouse screening | Not specified | Yes |
Table 2.
Risk of Bias.
| Author and Year | D1: Bias Arising from Randomization Process | D2: Bias Due to Deviations from Intended Intervention | D3: Bias Due to Missing Outcome Data | D4: Bias in Measurement of the Outcome | D5: Bias in Selection of the Reported Result | Overall Risk of Bias |
| Herawati et al. (2015) [3] |
Low | Some concerns | Low | Low | Low | Low |
| Njagi (2021) [17] |
Some concerns | Some concerns | Low | Some concerns | Low | Some concerns |
| Xu et al. (2008) [23] |
Low | Some concerns | Low | Low | Low | Low |
| Panmaha et al. (2025) [4] |
Low | Low | Low | Low | Low | Low |
| Puri et al. (2009) [9] |
Some concerns | Some concerns | Some concerns | Low | Low | Some concerns |
| Tuhina-Khatun et al. (2015) [10] |
Low | Some concerns | Low | Low | Low | Low |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



