
Submitted:
24 December 2025
Posted:
25 December 2025
You are already at the latest version
Abstract
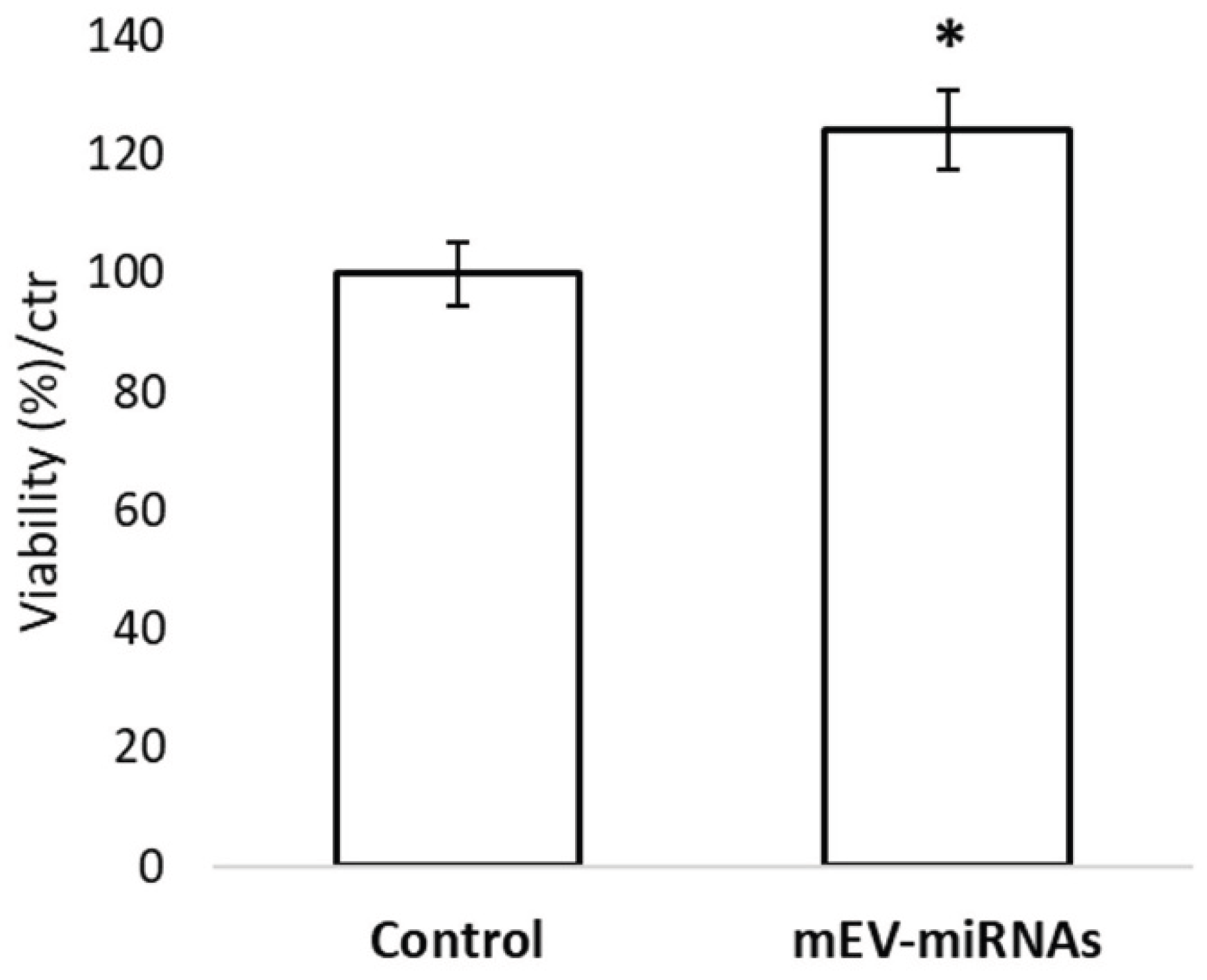
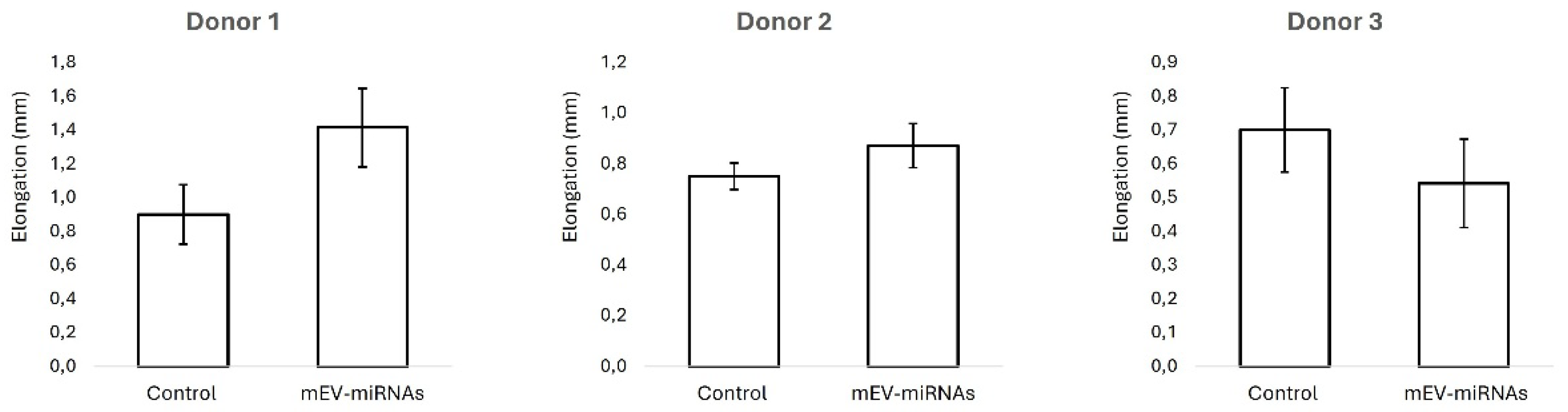
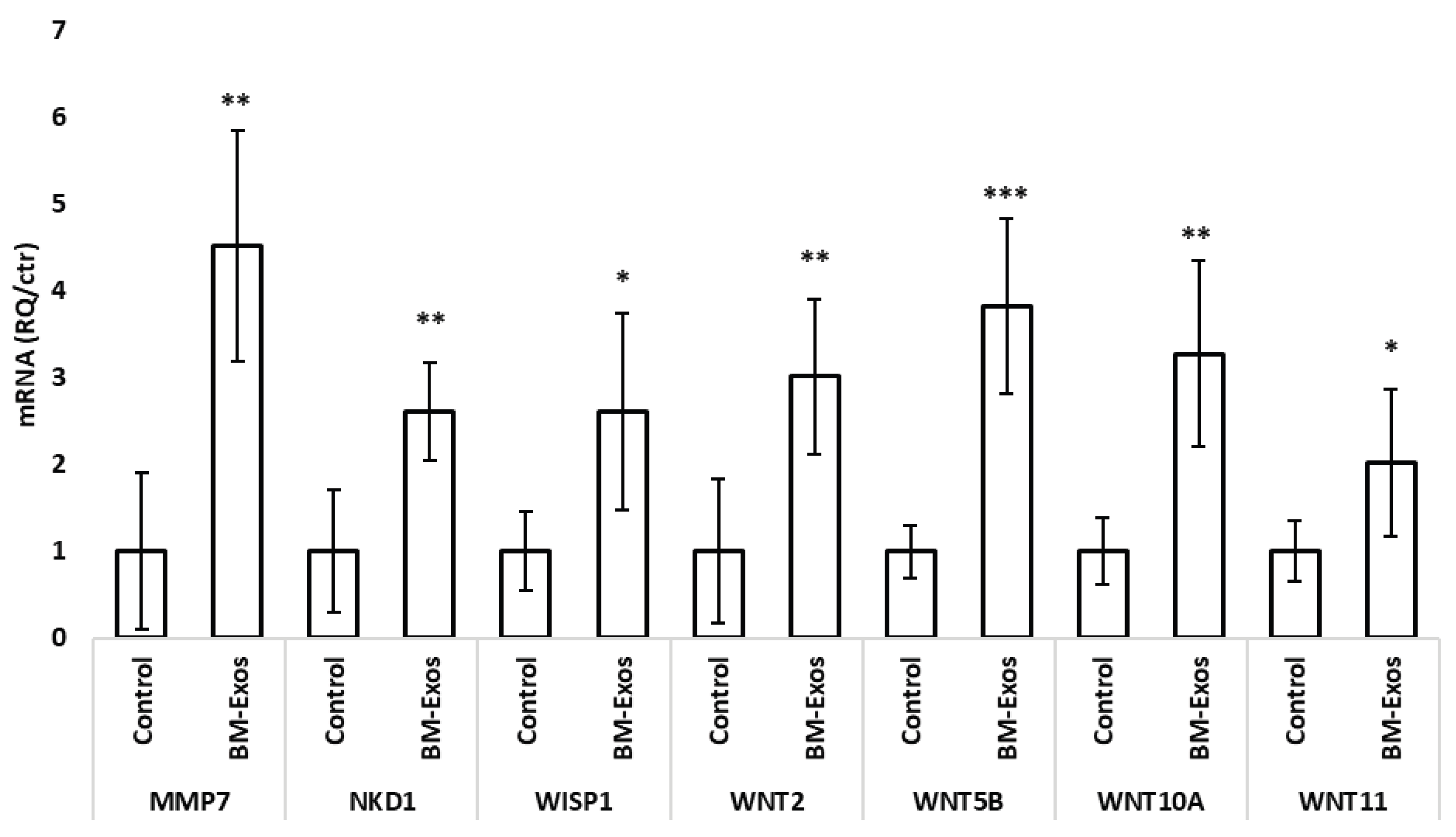
Androgenetic alopecia (AGA) and telogen effluvium (TE) are common hair loss disorders characterized by dysregulated hair follicle cycling and impaired dermal papilla cell function. Emerging evidences propose exosomes as key mediators of intercellular communication, largely via their microRNA (miRNA) cargo. Milk-derived exosomes (Mi-Exos) represent an accessible and biologically active source of regulatory miRNAs with therapeutic potential. This study evaluated the in vitro effects of bovine milk–derived exosomes (MEV-miRNAs) on human hair follicles. MEV-miRNAs were enriched in miRNA families (Let-7, miR-21, miR-30, miR-200, and miR-148/152) previously implicated in hair follicle regulation. Proliferation of hair follicle dermal papilla (HFDP) cell was assessed, and human hair follicles were cultured ex vivo to measure shaft elongation, and modulation of the WNT signaling pathway by qRT-PCR. MEV-miRNAs significantly increased HFDP cell viability after 24 hours compared with controls. Human hair follicles showed a non-significant trend toward increased elongation following treatment. Gene expression analysis revealed significant up-regulation of key WNT pathway components, including WNT2, WNT5B, WNT10A, WNT11, MMP7, WISP1, and NKD1, indicating activation of pro-regenerative signaling. Overall, MEV-miRNAs exhibit pro-proliferative and signaling-modulatory effects, supporting their potential as a novel therapeutic strategy for AGA and TE.
Keywords:
1. Introduction
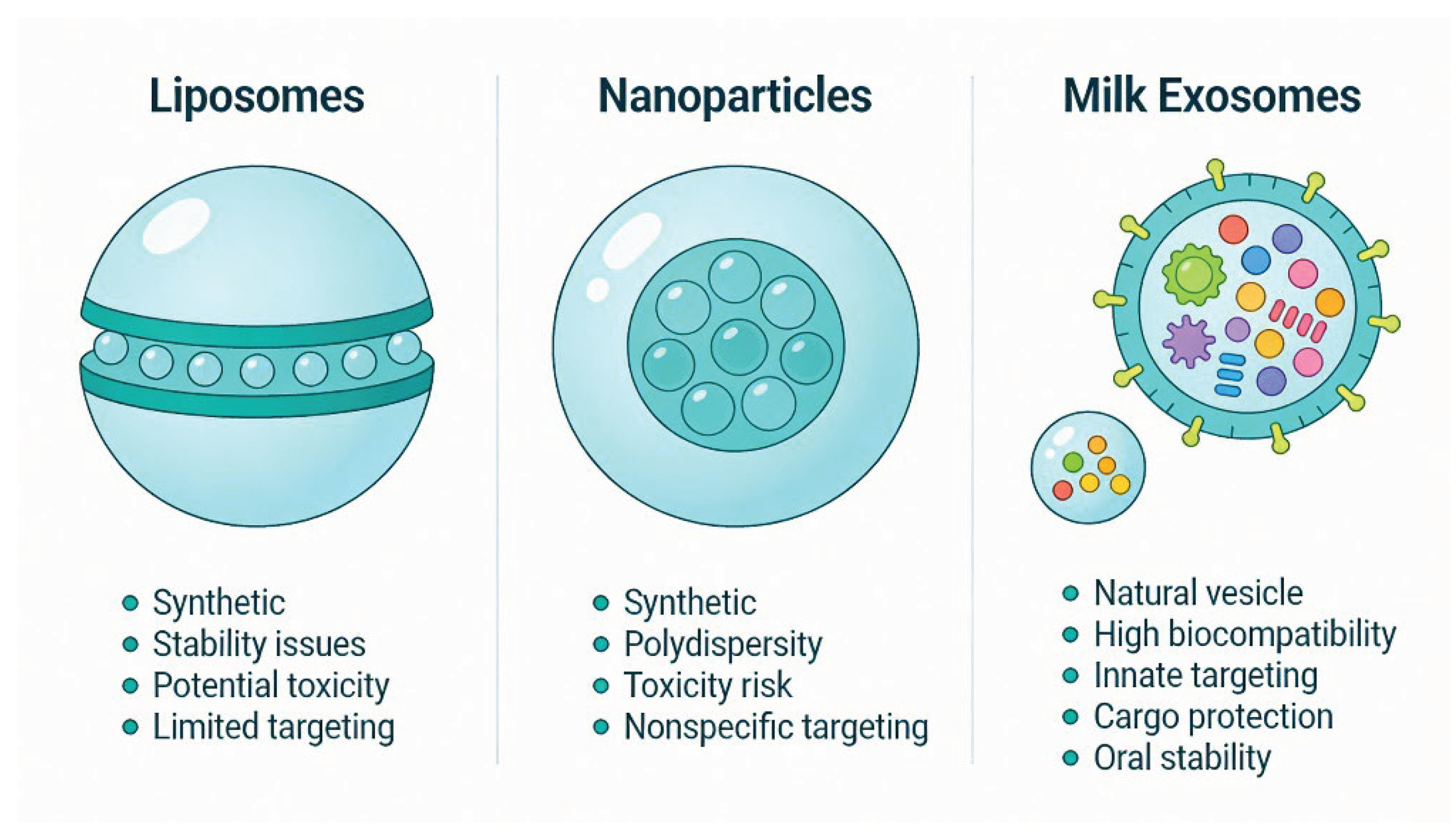
2. Materials and Methods
2.1. Exosomes Isolation and miRNA Characterisation
2.2. RNA Extraction and Sequencing
2.3. Bioinformatic Analysis of Small RNA-seq Data
2.4. Hair Follicle Dermal Papilla Cells Culture and Proliferation Assay (MTT Assay)
2.5. Human Hair Follicles Cultivation and Elongation Measurement
2.6. Hair Follicles’ Taqman Array Analysis of WNT Signaling Pathway
2.7. Statistical Analysis
3. Results
3.1. Identifying the miRNA fingerprint of mEV-miRNAs
3.2. Proliferative Activity on HFDP Cells
3.3. Elongation of Human Hair Follicles
3.4. WNT Signaling Pathway Differentially Expressed Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kost, Y.; Muskat, A.; Mhaimeed, N.; Nazarian, R.S.; Kobets, K. Exosome therapy in hair regeneration: A literature review of the evidence, challenges, and future opportunities. J Cosmet Dermatol 2022, 21(8), 3226–31. [Google Scholar] [CrossRef]
- Tang, X.; Cao, C.; Liang, Y.; Han, L.; Tu, B.; Yu, M.; Wan, M. Adipose-Derived Stem Cell Exosomes Antagonize the Inhibitory Effect of Dihydrotestosterone on Hair Follicle Growth by Activating Wnt/β-Catenin Pathway. Stem Cells Int. 2023, 2023, 5548112. [Google Scholar] [CrossRef]
- Chen, X.; Liu, B.; Li, Y.; Han, L.; Tang, X.; Deng, W.; Lai, W.; Wan, M. Dihydrotestosterone Regulates Hair Growth Through the Wnt/β-Catenin Pathway in C57BL/6 Mice and In Vitro Organ Culture. Front Pharmacol 2020, 10. [Google Scholar] [CrossRef]
- Zhang, Y; Huang, J; Fu, D; Liu, Z; Wang, H; Wang, J; Qu, Q; Li, K; Fan, Z; Hu, Z; Miao, Y. Transcriptome Analysis Reveals an Inhibitory Effect of Dihydrotestosterone-Treated 2D- and 3D-Cultured Dermal Papilla Cells on Hair Follicle Growth. Front Cell Dev Biol. 2021, 9, 724310. [Google Scholar] [CrossRef]
- Cedirian, S; Bruni, F; Quadrelli, F; Caro, G; Fortuna, M; Rossi, A; Piraccini, BM; Starace, M. Clinical study on the efficacy and tolerability of a topical regenerative treatment in patients with telogen effluvium and mild androgenetic alopecia. J Cosmet Dermatol 2023, 22(12), 3347–51. [Google Scholar] [CrossRef]
- Rudnicka, L; Kaczorowska, A. Treatment of diseases associated with non-cicatricial alopecia. Dermatology 2021, 108, 504–16. [Google Scholar]
- Kim, H; Jang, Y; Kim, EH; Jang, H; Cho, H; Han, G; Song, HK; Kim, SH; Yang, Y. Potential of Colostrum-Derived Exosomes for Promoting Hair Regeneration Through the Transition From Telogen to Anagen Phase. Front Cell Dev Biol 2022, 10. [Google Scholar] [CrossRef]
- Xia, Y; Wei, K; Hu, LQ; Zhou, CR; Lu, ZB; Zhan, GS; Pan, XL; Pan, CF; Wang, J; Wen, W; Xu, J; He, ZC; Huang, CJ; Chen, L. Exosome-mediated transfer of miR-1260b promotes cell invasion through Wnt/β-catenin signaling pathway in lung adenocarcinoma. J Cell Physiol. 2020, 235(10), 6843–53. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J; Li, S; Li, L; Li, M; Guo, C; Yao, J; Mi, S. Exosome and exosomal microRNA: trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015, 13(1), 17–24. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. Journal of Cell Biology 2013, 200(4), 373–83. [Google Scholar] [CrossRef]
- Ge, Q; Zhou, Y; Lu, J; Bai, Y; Xie, X; Lu, Z. miRNA in plasma exosome is stable under different storage conditions. Molecules 2014, 19(2), 1568–75. [Google Scholar] [CrossRef]
- Kalluri, R; LeBleu, VS. The biology , function , and biomedical applications of exosomes. Science 2020, 367(6478), eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Torregrosa Paredes, P; Gutzeit, C; Johansson, S; Admyre, C; Stenius, F; Alm, J; Scheynius, A; Gabrielsson, S. Differences in exosome populations in human breast milk in relation to allergic sensitization and lifestyle. Allergy 2014, 69(4), 463–71. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D; Giuliani, G; Mondadori, G; Rinaldi, F. Exosomes in Dermatological Research: Unveiling Their Multifaceted Role in Cellular Communication, Healing, and Disease Modulation. Cosmetics 2025, 12(1), 16. [Google Scholar] [CrossRef]
- Wang, J; Ma, Y; Li, T; Li, J; Yang, X; Hua, G; Cai, G; Zhang, H; Liu, Z; Wu, K; Deng, X. MiR-199a-3p Regulates the PTPRF/β-Catenin Axis in Hair Follicle Development: Insights into the Pathogenic Mechanism of Alopecia Areata. Int J Mol Sci 2023, 24(24), 17632. [Google Scholar] [CrossRef]
- de Rivero Vaccari, JP; Sawaya, ME; Brand, F; Nusbaum, BP; Bauman, AJ; Bramlett, HM; Dietrich, WD; Keane, RW. Caspase-1 level is higher in the scalp in androgenetic alopecia. Dermatol Surg. 2012, 38 7 Pt 1, 1033–9. [Google Scholar] [CrossRef] [PubMed]
- Hu, S; Li, Z; Lutz, H; Huang, K; Su, T; Cores, J; Dinh, PUC; Cheng, K. Dermal exosomes containing miR-218-5p promote hair regeneration by regulating β-catenin signaling. Sci Adv. 2020, 6(30), eaba1685. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 1983, 65(1–2), 55–63. [Google Scholar] [CrossRef]
- Chomczynski, P; Mackey, K. Short technical reports. Modification of the TRI reagent procedure for isolation of RNA from polysaccharide- and proteoglycan-rich sources. Biotechniques 1995, 19(6), 942–5. [Google Scholar]
- Vigetti, D; Viola, M; Karousou, E; Rizzi, M; Moretto, P; Genasetti, A; Clerici, M; Hascall, VC; De Luca, G; Passi, A. Hyaluronan-CD44-ERK1/2 regulate human aortic smooth muscle cell motility during aging. J Biol Chem. 2008, 283(7), 4448–58. [Google Scholar] [CrossRef]
- Yoon, YC; Ahn, BH; Min, JW; Lee, KR; Park, SH; Kang, HC. Stimulatory Effects of Extracellular Vesicles Derived from Leuconostoc holzapfelii That Exists in Human Scalp on Hair Growth in Human Follicle Dermal Papilla Cells. Current Issues in Molecular Biology 2022, 44(2), 845–66. [Google Scholar] [CrossRef]
- Ferruggia, G; Zimbone, M; Brundo, MV. New technologies for hair follicle growth and regeneration: comparison between different commercial products contain exosomes. bioRxiv 2024, 2024.11.17.623990. [Google Scholar]
- Andl, T; Botchkareva, NV. MicroRNAs (miRNAs) in the control of HF development and cycling: the next frontiers in hair research. Exp Dermatol 2015, 24(11), 821–6. [Google Scholar] [CrossRef]
- Paul, S; Licona-Vázquez, I; Serrano-Cano, FI; Frías-Reid, N; Pacheco-Dorantes, C; Pathak, S; Chakraborty, S; Srivastava, A. Current insight into the functions of microRNAs in common human hair loss disorders: a mini review. Human Cell 2021, 34(4), 1040–50. [Google Scholar] [CrossRef]
- Rajar, UDM; Fahim, A; Qazi, S; Kazi, N; Maheshwary, N; Rph, AA. Platelet-Rich Plasma Therapy for Telogen Effluvium: A Comprehensive Evaluation of Efficacy and Safety. Dermis 2024, 4(1), 1–9. [Google Scholar] [CrossRef]
- Yi, R; O’Carroll, D; Pasolli, HA; Zhang, Z; Dietrich, FS; Tarakhovsky, A; Fuchs, E. Morphogenesis in skin is governed by discrete sets of differentially expressed microRNAs. Nat Genet 2006, 38(3), 356–62. [Google Scholar] [CrossRef]
- Xiang, B; Li, Yumei; Li, Jianping; Zhang, Baoyu; Li, Jianyu; Jiang, HuaiZhi; Zhang, Q. MiR-21 regulated hair follicle cycle development in Cashmere goats by targeting FGF18 and SMAD7. Animal Biotechnology 2023, 34(9), 4695–702. [Google Scholar] [CrossRef]
- Xiong, J; Wu, B; Hou, Q; Huang, X; Jia, L; Li, Y; Jiang, H. Comprehensive Analysis of LncRNA AC010789.1 Delays Androgenic Alopecia Progression by Targeting MicroRNA-21 and the Wnt/β-Catenin Signaling Pathway in Hair Follicle Stem Cells. Front Genet. 2022, 13, 782750. [Google Scholar] [CrossRef]
- Ma, T; Li, J; Jiang, Q; Wu, S; Jiang, H; Zhang, Q. Differential expression of miR-let7a in hair follicle cycle of Liaoning cashmere goats and identification of its targets. Funct Integr Genomics 2018, 18(6), 701–7. [Google Scholar] [CrossRef]
- Yan, S; Yu, Z; Ning, L; Hai-dong, W; Jian-Shan, X; Shu-yuan, G; Jia-qi, C; Xiu-ju, Y; Ting, W; Chang-sheng, D; Xiao-yan, H. Let-7b promotes alpaca hair growth via transcriptional repression of TGFβR I. Gene 2016, 577(1), 32–6. [Google Scholar] [CrossRef] [PubMed]
- Liu, N; Niu, S; Cao, X; Cheng, J; Gao, S; Yu, X; Wang, H; Dong, C; He, X. Let-7b regulates alpaca hair growth by downregulating ectodysplasin A. Mol Med Report; 2018. [Google Scholar]
- Tafazzoli, A; Forstner, AJ; Broadley, D; Hofmann, A; Redler, S; Petukhova, L; Giehl, KA; Kruse, R; Blaumeiser, B; Böhm, M; Bertolini, M; Rossi, A; Garcia Bartels, N; Lutz, G; Wolff, H; Blume-Peytavi, U; Soreq, H; Christiano, AM; Botchkareva, NV; Nöthen, MM; Betz, RC. Genome-Wide MicroRNA Analysis Implicates miR-30b/d in the Etiology of Alopecia Areata. Journal of Investigative Dermatology 2018, 138(3), 549–56. [Google Scholar] [CrossRef]
- Yan, H; Gao, Y; Ding, Q; Liu, J; Li, Y; Jin, M; Xu, H; Ma, S; Wang, X; Zeng, W; Chen, Y. Exosomal Micro RNAs Derived from Dermal Papilla Cells Mediate Hair Follicle Stem Cell Proliferation and Differentiation. Int J Biol Sci. 2019, 15(7), 1368–82. [Google Scholar] [CrossRef]
- Liu, J; Liu, B; Mu, Q; Liu, J; Li, Y; Gong, W; Chahaer, T; Song, Y; Hai, E; Wang, H; Zhang, Y; Zhao, Y. Melatonin promotes the proliferation of dermal papilla cells in cashmere goats via activation of chi-let-7d-5p/WNT2 axis. Genomics 2024, 116(6), 110961. [Google Scholar] [CrossRef]
- Benard, EL; Hammerschmidt, M. The fundamentals of WNT10A. Differentiation 2025, 142, 100838. [Google Scholar] [CrossRef]
- Hawkshaw, NJ; Hardman, JA; Alam, M; Jimenez, F; Paus, R. Deciphering the molecular morphology of the human hair cycle: Wnt signalling during the telogen-anagen transformation. Br J Dermatol 2020, 182(5), 1184–93. [Google Scholar] [CrossRef] [PubMed]
- Cesarato, N; Schwieger-Briel, A; Gossmann, Y; Henne, SK; Hillmann, K; Frommherz, LH; Wehner, M; Xiong, X; Thiele, H; Oji, V; Milani, D; Tantcheva-Poor, I; Giehl, K; Fölster-Holst, R; Teichler, A; Braeckmans, D; Hoeger, PH; Jones, G; Frank, J; Weibel, L; Blume-Peytavi, U; Hamm, H; Nöthen, MM; Geyer, M; Heilmann-Heimbach, S; Basmanav, FB; Betz, RC. Short anagen hair syndrome: association with mono- and biallelic variants in WNT10A and a genetic overlap with male pattern hair loss. Br J Dermatol. 2023, 189(6), 741–9. [Google Scholar] [CrossRef] [PubMed]
- Suthon, S; Perkins, RS; Bryja, V; Miranda-Carboni, GA; Krum, SA. WNT5B in Physiology and Disease. In Front Cell Dev Biol; 2021. [Google Scholar]
- Zheng, M; Choi, N; Jang, Y; Kwak, DE; Kim, Y; Kim, WS; Oh, SH; Sung, JH. Hair growth promotion by Necrostatin-1s. Sci Rep. 2020, 10(1), 17622. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S; Andl, T; Bagasra, A; Lu, MM; Epstein, DJ; Morrisey, EE; Millar, SE. Characterization of Wnt gene expression in developing and postnatal hair follicles and identification of Wnt5a as a target of Sonic hedgehog in hair follicle morphogenesis. Mechanisms of Development. 2001, 107(1), 69–82. [Google Scholar] [CrossRef]
- Gong, G; Fan, Y; Yan, X; Li, W; Yan, X; Liu, H; Zhang, L; Su, Y; Zhang, J; Jiang, W; Liu, Z; Wang, Z; Wang, R; Zhang, Y; Lv, Q; Li, J; Su, R. Identification of Genes Related to Hair Follicle Cycle Development in Inner Mongolia Cashmere Goat by WGCNA. Front Vet Sci 2022. [Google Scholar] [CrossRef]
- Crawford, HC; Fingleton, B; Gustavson, MD; Kurpios, N; Wagenaar, RA; Hassell, JA; Matrisian, LM. The PEA3 Subfamily of Ets Transcription Factors Synergizes with β-Catenin–LEF-1 To Activate Matrilysin Transcription in Intestinal Tumors. Molecular and Cellular Biology 2001, 21(4), 1370–83. [Google Scholar] [CrossRef]
- Botchkarev, VA; Sharov, AA. BMP signaling in the control of skin development and hair follicle growth. Differentiation 2004, 72(9–10), 512–26. [Google Scholar] [CrossRef] [PubMed]
- Deng, W; Fernandez, A; McLaughlin, SL; Klinke, DJ. WNT1-inducible signaling pathway protein 1 (WISP1/CCN4) stimulates melanoma invasion and metastasis by promoting the epithelial–mesenchymal transition. Journal of Biological Chemistry. 2019, 294(14), 5261–80. [Google Scholar] [CrossRef] [PubMed]
- Angonin, D; Raay, TJV. Nkd1 Functions as a Passive Antagonist of Wnt Signaling. PLOS ONE 2013, 8(8), e74666. [Google Scholar] [CrossRef] [PubMed]

| # | miRNA |
| 1 | Bta.Let.7.P2b2_5p |
| 2 | Bta.Let.7.P1d_5p |
| 3 | Bta.Mir.8.P1b_3p |
| 4 | Bta.Mir.8.P2b_3p |
| 5 | Bta.Mir.15.P2a_5p |
| 6 | Bta.Mir.30.P1b_5p |
| 7 | Bta.Mir.28.P2_5p |
| 8 | Bta.Let.7.P2a3_5p |
| 9 | Bta.Let.7.P2a1_5p |
| 10 | Bta.Let.7.P2a2_5p |
| 11 | Bta.Let.7.P2c3_5p |
| 12 | Bta.Mir.26.P4_5p |
| 13 | Bta.Mir.26.P1_5p |
| 14 | Bta.Mir.320.P1b_3p |
| 15 | Bta.Let.7.P1c_5p |
| 16 | Bta.Mir.26.P2_5p |
| 17 | Bta.Mir.191_5p |
| 18 | Bta.Mir.423_5p |
| 19 | Bta.Let.7.P2b1_5p |
| 20 | Bta.Mir.30.P1a_5p |
| 21 | Bta.Mir.21_5p |
| 22 | Bta.Mir.320.P1a_3p |
| 23 | Bta.Mir.29.P2d2_3p |
| 24 | Bta.Mir.29.P2b_3p |
| 25 | Bta.Let.7.P2c1_5p |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



